Back to Journals » Infection and Drug Resistance » Volume 15
Prevalence of CRISPR-Cas Systems and Their Possible Association with Antibiotic Resistance in Enterococcus faecalis and Enterococcus faecium Collected from Hospital Wastewater
Authors Alduhaidhawi AHM, AlHuchaimi SN, Al- Mayah TA, Al-Ouqaili MTS ![]() , Alkafaas SS
, Alkafaas SS ![]() , Muthupandian S, Saki M
, Muthupandian S, Saki M ![]()
Received 13 January 2022
Accepted for publication 12 March 2022
Published 19 March 2022 Volume 2022:15 Pages 1143—1154
DOI https://doi.org/10.2147/IDR.S358248
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Athraa Harjan Mohsen Alduhaidhawi,1 Sundus Nsaif AlHuchaimi,2 Thikra Abdullah Al- Mayah,2 Mushtak TS Al-Ouqaili,3 Samar Sami Alkafaas,4 Saravanan Muthupandian,5,6 Morteza Saki7
1Department of Biology, Faculty of Science, University of Kufa, Najaf, Iraq; 2Department of Parasitology, Faculty of Medicine, University of Kufa, Najaf, Iraq; 3Department of Microbiology, College of Medicine, University of Anbar, Ramadi, Al-Anbar Governorate, Iraq; 4Department of Chemistry, Division of Biochemistry, Faculty of Science, Tanta University, Tanta, Egypt; 5Department of Microbiology and Immunology, Division of Biomedical Sciences, School of Medicine, College of Health Sciences, Mekelle University, Mekelle, 1871, Ethiopia; 6Department of Pharmacology, AMR and Nanomedicine Laboratory, Center for Transdisciplinary Research, Saveetha Dental College, Saveetha Institute of Medical and Technical Sciences (SIMATS), Chennai, 60007, India; 7Department of Microbiology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Correspondence: Saravanan Muthupandian, Department of Microbiology and Immunology, Division of Biomedical Sciences, School of Medicine, College of Health Sciences, Mekelle University, Mekelle, 1871, Ethiopia, Tel +919443077097, Email [email protected] Morteza Saki, Department of Microbiology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran, Tel +989364221187, Email [email protected]
Purpose: This study aimed to evaluate the presence of CRISPR-Cas system genes and their possible association with antibiotic resistance patterns of Enterococcus faecalis and Enterococcus faecium species isolated from hospital wastewater (HWW) samples of several hospitals.
Methods: HWW samples (200 mL) were collected from wastewater discharged from different hospitals from October 2020 to March 2021. The isolation and identification of enterococci species were performed by standard bacteriology tests and polymerase chain reaction (PCR). Antibiotic resistance was determined using the disc diffusion. The presence of various CRISPR-Cas systems was investigated by PCR. The association of the occurrence of CRISPR-Cas systems with antibiotic resistance was analyzed with appropriate statistical tests.
Results: In total, 85 different enterococci species were isolated and identified using phenotypic methods. The results of PCR confirmed the prevalence of 50 (58.8%) E. faecalis and 35 (41.2%) E. faecium, respectively. In total, 54 (63.5%) of 85 isolates showed the presence of CRISPR-Cas loci. The incidence of CRISPR-Cas was more common in E. faecalis. CRISPR1, CRISPR2, and CRISPR3 were present in 35 (41.2%), 47 (55.3%), and 30 (35.3%) enterococci isolates, respectively. The CRISPR-Cas positive isolates showed significant lower resistance rates against vancomycin, ampicillin, chloramphenicol, erythromycin, rifampin, teicoplanin, tetracycline, imipenem, tigecycline, and trimethoprim-sulfamethoxazole in comparison with CRISPR-Cas negative isolates. The results showed that the presence of CRISPR-Cas genes was lower in multidrug-resistant (MDR) isolates (53.1%, n = 26/49) compared to the non-MDR enterococci isolates (77.8%, n = 28/36) (P = 0.023).
Conclusion: This study revealed the higher prevalence of E. faecalis than E. faecium in HWWs. Also, the lack of CRISPR-Cas genes was associated with more antibiotic resistance rates and multidrug resistance in E. faecalis and E. faecium isolates with HWW origin.
Keywords: antibiotic resistance, CRISPR-Cas, Enterococcus faecalis, Enterococcus faecium, hospital wastewater, HWW
Introduction
In recent years, hospitals and healthcare centers have become the main repositories for numerous multidrug-resistant (MDR) pathogenic bacteria, including Enterococcus species.1,2 One of the reasons for this phenomenon is the indiscriminate and unsupervised use of antibiotics in the treatment of infection, which creates a selective pressure to develop resistance. The spread of antibiotic-resistant bacteria (ARBs) from hospitals, which can be done through various routes including hospital wastewater (HWW), plays an important role in spreading this resistance to the community.1–3 Because of their ever-increasing resistance, enterococci have surpassed staphylococci as the second most prevalent cause of nosocomial infections.4 Two species Enterococcus faecalis and Enterococcus faecium are primarily responsible for these infections. The emergence of vancomycin-resistant enterococci (VRE) that are resistant to several antibiotics has become a major problem for healthcare systems in recent years.5,6 The rise of VRE, has also resulted in heavier use of newer antibiotics such as daptomycin and linezolid to combat them which may lead to increased resistance to these drugs.6
Enterococci are used as markers of fecal contamination and are part of the hygiene requirements for water and food products due to their preferred intestinal habitat, broad occurrence, and strength and simplicity of cultivation.7 Pathogenicity islands, plasmids, and antibiotic resistance genes comprise as much as 25% of the genomes of hospital- adapted lineages of enterococci species. According to the literature, most of both E. faecalis and E. faecium from clinical infection origin lack the clustered regularly interspaced short palindromic repeats (CRISPRs) loci and CRISPR-associated (Cas) protein-encoding genes known as CRISPR-Cas systems.8
CRISPR-Cas systems constitute a multi-step adaptive immune response that defends prokaryotes against foreign invading genetic material. A genomic locus including the CRISPR and nonrepetitive spacer sequences, as well as 6 to 20 genes producing CRISPR-Cas related proteins, make up the system. CRISPR-Cas systems consist of 2 classes and six types (types I, III, and IV belong to class 1 and types II, V and VI belong to class 2).9,10
Recent reports have shown that CRISPR-Cas systems can be used for genome engineering. Also, they impose a strong selective pressure on pathogenic bacteria to acquire virulence traits and antibiotic resistance.10–12 CRISPR1-cas, orphan CRISPR2, and CRISPR3-cas are three types of CRISPR-Cas systems found in the enterococci genome. CRISPR1 and CRISPR2 both have seven repeats of a 37-bp palindromic motif that bears no resemblance to any of the 29-bp spacer sequences. Despite this, they likely originated from pheromone-responsive type plasmids, plasmids integrated inside the E. faecalis V583 genome, and enterococcal prophage and phage, due to short spacer sequences.11,12
The available data regarding the possible association of CRISPR-Cas with antibiotic resistance in enterococci with HWW origin in Middle Eastern countries is scarce. The existence of the CRISPR-Cas system has been found to have a highly substantial negative association with the antibiotic resistance in some bacteria.13,14 However, very few studies have addressed this issue so far, and even in some of them, a positive association has been reported.9 Hence, this study aimed to evaluate the presence of CRISPR-Cas system genes and their possible association with antibiotic resistance patterns of E. faecalis and E. faecium isolated from HWW samples of several hospitals.
Materials and Methods
Ethical Approval
This study was approved by the Ethics Committee of the University of Kufa, Najaf, Iraq (ethics code: MEC-10) following the Declaration of Helsinki. No human and animal samples were used in this study.
Sample Collection
The HWW samples were collected from wastewater discharged from different hospitals during February 2020 and March 2021. Water sampling was carried out at frequent and regular intervals from the wastewater leaving the hospital. For each sample, 200 mL of hospital sewage was collected in sterile bottles and kept in ice packs and transported to the laboratory for microbiological testing.
Enterococci Isolation and Identification
To isolate and presumptive identification of Enterococcus species, 0.5 mL of the HWW samples were inoculated in 9.5 mL of tryptic soy broth (TSB) (Merck, Germany) and incubated for 24 h at 37 °C. Bacterial growths were streaked onto bile esculin azide agar (BEA) (Merck, Germany) as selective agar plates and incubated at 37 °C for 24 h. The evidence of the presence of enterococci was the formation of dark brown or black colonies. These dark brown colonies that regarded to be presumptive isolates, were further investigated by a panel of standard bacteriology tests including Gram staining, growth in NaCl 6.5% broth, and catalase test.15,16 The confirmed enterococci species were inoculated into TSB/glycerol (20%) and stocked at – 80 °C for further analysis.
Confirmation of Enterococci Isolates by Polymerase Chain Reaction (PCR)
The final confirmation of the Enterococcus species was based on the detection of the genus-specific tuf gene (product size 112 bp) using previously described primers (Table 1).17 Firstly, DNA extraction was performed by boiling method according to the previous described protocol with some modification.17 The bacterial suspension was centrifuged at 10,000 rpm for 10 min and supernatant was discarded and pellet washed with 200 μL saline and further centrifuged at 10,000 rpm for 10 min. Then the supernatant was discarded. A volume of 150 μL of lysis buffer was added and cells were lysed in a heating block at 100 °C for 10 min and then were centrifuged at 10,000 rpm for 10 minutes. Finally, the supernatant was used as a DNA template.
 |
Table 1 List of Control Strains/Genes, PCR Primers and Products Size |
The E. faecalis and E. faecium isolates were confirmed and identified using a multiplex PCR (Biorad S1000 Thermal Cycler, USA) with specific primers presented in Table 1.17 The 25 μL PCR reaction mixture was composed of 12 μL of Master Mix [Dream Taq PCR Master Mix (2X), Thermo Scientific, Pittsburgh, PA, USA], 3 μL of the DNA template, 5 μL of nuclease free water, and 0.5 μL of each primer (10 pMol). The different standard enterococci species were used as positive controls (Table 1). The PCR cycling conditions consisted of the initial denaturation 95 °C/3 min followed by 30 cycles of denaturation at 95 °C for 30s, annealing at 48 °C (E. faecium) and 52 °C (E. faecalis) for 30s, and extension at 72 °C for 1 min, followed by a final extension at 72 °C for 7 min. An agarose gel containing 2% Tris- Borate-EDTA with 2 μg ethidium bromide/mL was used to confirm the amplification of targeted genes using electrophoresis at 110 V for 40 min. The amplicons were detected by a UV light transilluminator and their size band was determined using a 100 bp DNA molecular weight marker.17
Antimicrobial Susceptibility Test (AST)
The AST of enterococci isolates were evaluated against vancomycin (30 µg), ampicillin (10 µg), chloramphenicol (30 µg), ciprofloxacin (5 µg), erythromycin (15 µg), linezolid (30 µg), rifampin (5 µg), teicoplanin (30 µg), tetracycline (30 µg), imipenem (10 µg), tigecycline (15 µg), and trimethoprim-sulfamethoxazole (1.25–23.75 µg) (MAST Co, UK) according to the Clinical and Laboratory Standards Institute (CLSI).18 For antibiotics that were not on the CLSI list, the European Committee on Antimicrobial Susceptibility Testing (EUCAST) recommendations were used.19 Disc diffusion method was performed using Muller-Hinton (MH) agar (Merck, Darmstadt, Germany) for AST. Physiological saline (0.85%) was used to prepare bacterial suspensions from overnight cultures to closely match the MacFarland standard 0.5. The suspensions were spread on MH agar plates using sterile cotton swabs. Then, the antibiotic discs were placed on the plates. The plates were incubated at 37 °C for 18 h. CLSI and EUCAST breakpoints were used to interpret the results. Quality control was performed using Staphylococcus aureus ATCC 25923 and E. faecalis ATCC 29212.16 MDR resistotypes were determined based on the resistance of isolates to three or more antibiotics from different examined classes.14 The tested antibiotics were belong to 10 different classes as follows: C1, penicillins; C2, carbapenems; C3, fluoroquinolones; C4, folate pathway antagonists; C5, phenicols; C6, glycopeptides; C7, tetracyclines; C8, ansamycins; C9, oxazolidinones; and C10, macrolides.
Detection of CRISPR-Cas Systems
The different CRISPR-Cas systems were screened using the previously published primers.11 The PCR mix for detection of various CRISPR-Cas systems contained 12.5 μL of the 2X Master Mix, and 1 μL of each primer (10 pM), 2 μL of DNA sample, and 8.5 μL of nuclease-free water. The amplification condition was carried out with the following thermal cycling program: initial denaturation at 95 °C for 5 min, 34 cycles of amplification consisting of 95 °C for 30s, 30s at 60 °C for all CRISPR-Cas systems, and 72 °C for 45s, with 72 °C for 5 min in the final polymerization. Gel electrophoresis was used to analyze PCR products in a 1% agarose gel at 100 V for 1 h in a 1X TBE buffer containing the DNA safe stain. PCR product size was correlated with a 100 based-pair DNA ladder to confirm the conjunction with their expected PCR amplicon size.
Data Analysis
Prism GraphPad, version 8.0 (GraphPad Software, Inc., CA, US) was used for all descriptive statistical analyses. Chi-square or Fisher’s exact tests were used to compare the antibiotic resistance rates between E. faecalis and E. faecium isolates and the associations between the presence of CRISPR-Cas genes and resistance to each antibiotic in both isolates. A P-value ≤ 0.05 was considered a significant association.
Results
Isolates Prevalence
In total, 50 water samples were collected from 6 hospitals, from which 85 different enterococci species were isolated and identified using culture and phenotypic methods. The results of PCR confirmed the prevalence of 50 (58.8%) E. faecalis and 35 (41.2%) E. faecium, respectively.
Antibiotic Resistotypes
The resistance rates of E. faecium and E. faecalis isolates are shown in Table 2. The highest rate of resistance was documented against ampicillin (58.8%), followed by tetracycline (55.3%) and ciprofloxacin (54.1%). The linezolid (8.2%), imipenem (8.2%), rifampin (9.4%), and tigecycline (12.9%) with the lowest resistance rates showed the strongest effects against enterococci isolates. The total resistance rate of vancomycin was 29.4%. The resistance rates to linezolid and trimethoprim-sulfamethoxazole antibiotics were significantly higher in E. faecalis than E. faecium (P-value < 0.05). In total, 49 (57.6%) isolates (30 E. faecalis and 19 E. faecium, respectively) were MDR with 10 different resistotypes (Table 3). The R7 (ampicillin-ciprofloxacin-tetracycline-erythromycin-trimethoprim-sulfamethoxazole) with the rate of 24.5% was the most prevalent resistotype.
 |
Table 2 Resistance Rates of Different Enterococci Strains Against Studied Antibiotics |
 |
Table 3 Resistotypes of 49 Multidrug-Resistant Enterococci Isolates Based on the Studied Antibiotics Classes |
Prevalence of CRISPR-Cas Systems
The prevalence of three different CRISPR-Cas elements in enterococci isolates is presented in Table 4. In total, 54 (63.5%) of 85 isolates showed positive PCR bands for the presence of CRISPR-Cas loci. CRISPR1, CRISPR2, and CRISPR3 were present separately or in combination in a total of 35 (41.2%), 47 (55.3%), and 30 (35.3%) enterococci isolates, respectively. The incidence of CRISPR-Cas was more common in E. faecalis (n = 40, 80.0%) than E. faecium (n =14, 40.0%) isolates (P = 0.0002). CRISPR1, CRISPR2, and CRISPR3 were observed in 25 (50.0%), 39 (78.0%), and 18 (36.0%) E. faecalis isolates, respectively. Also, CRISPR1, CRISPR2, and CRISPR3 were detected in 10 (28.6%), 8 (22.9%), and 12 (34.3%) E. faecium isolates, respectively. The coexistence of CRISPR/Cas loci was found in 32 (59.3%) isolates (Table 4).
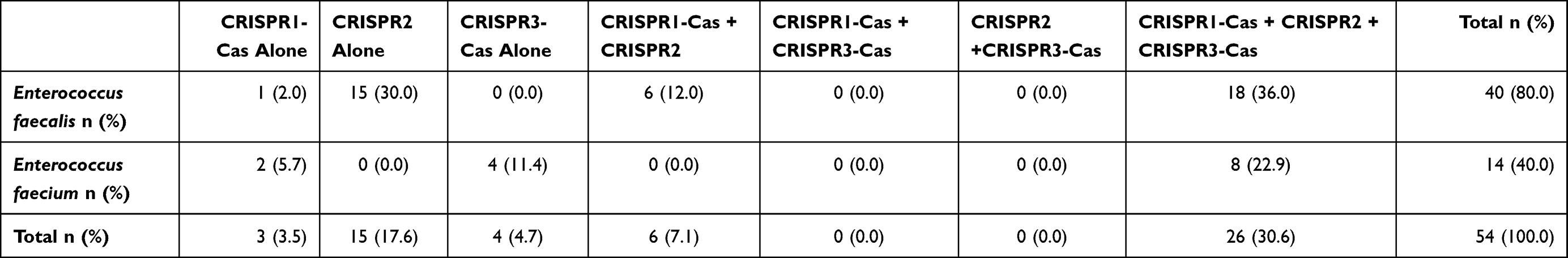 |
Table 4 The Occurrence of CRISPR/Cas Luci in 50 Enterococcus faecalis and 35 Enterococcus faecium Isolates |
Association Between the Occurrence of CRISPR-Cas Elements and Antibiotic Resistance
The association between the occurrence of CRISPR-Cas elements and antibiotic resistance of 85 enterococci isolates is summarized in Table 5. There was a negative association between the occurrence of CRISPR-Cas elements and antibiotic resistance rates in enterococci isolates. The CRISPR-Cas positive E. faecalis isolates showed significant lower resistance rates against vancomycin, ampicillin, chloramphenicol, erythromycin, rifampin, teicoplanin, tetracycline, imipenem, tigecycline, and trimethoprim-sulfamethoxazole in comparison with CRISPR-Cas negative E. faecalis strains. Also, the CRISPR-Cas positive E. faecium isolates showed significant lower resistance rates against vancomycin, ampicillin, chloramphenicol, erythromycin, teicoplanin, and tetracycline in comparison with the CRISPR-Cas negative E. faecium strains (Table 5). Finally, the results showed that the presence of CRISPR-Cas genes was lower in MDR isolates (53.1%, n = 26/49) compared to the non MDR enterococci isolates (77.8%, n = 28/36) (P = 0.023).
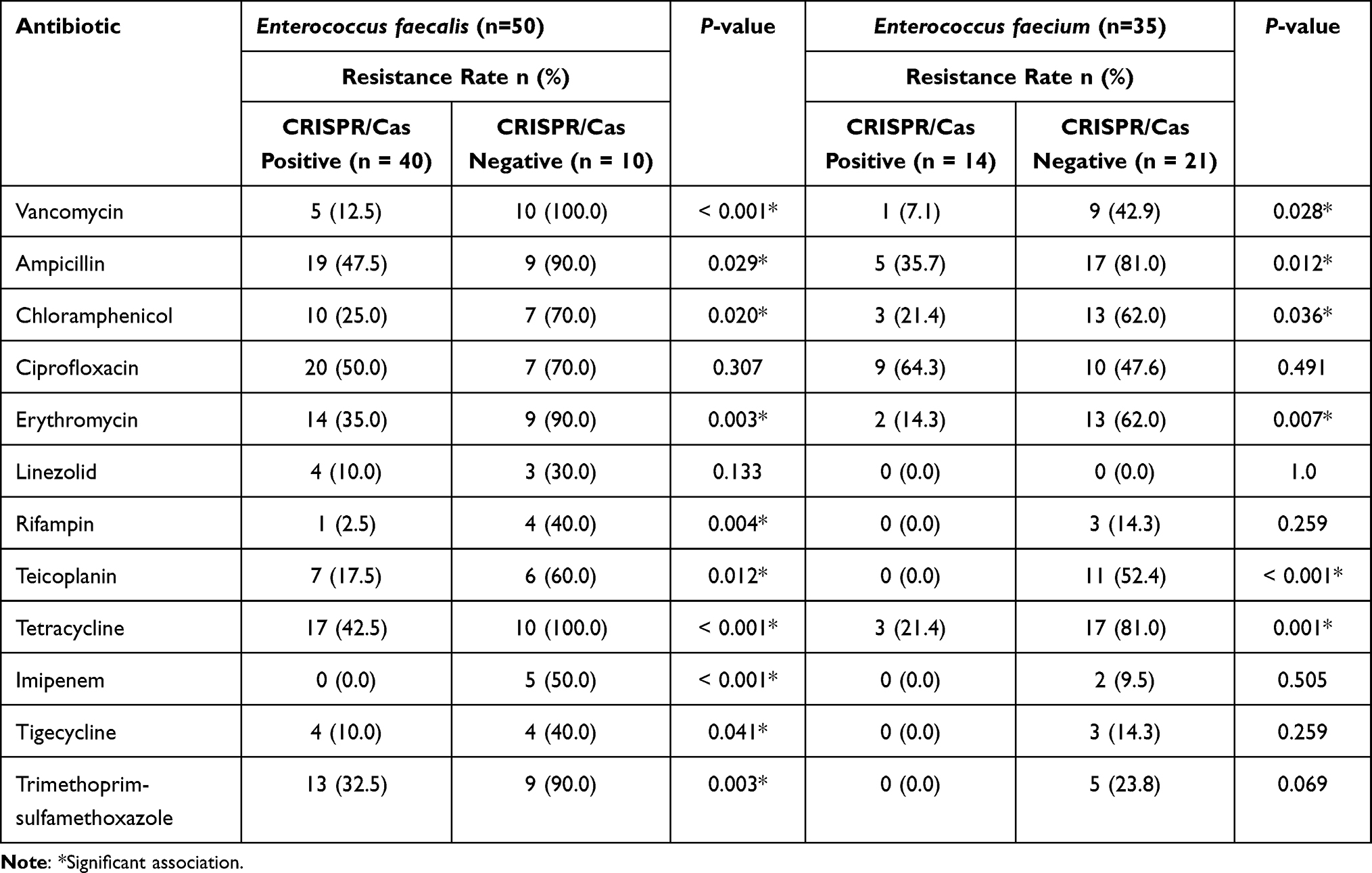 |
Table 5 Association Between the Occurrence of CRISPR/Cas Elements and Antibiotic Resistance in Enterococcus faecalis and Enterococcus faecium Isolates |
Discussion
At this time, little is known about the antibiotic resistance mechanisms of enterococci in the water environment. Also, reports of an association between the presence of CRISPR-Cas genes and antibiotic resistance in enterococci strains isolated from aquatic environments, especially HWWs, are rare. The strength of this study was to address this issue in enterococci strains isolated from HWWs samples.
In this study, E. faecalis was the most predominant (58.8%) strain followed by E. faecium (41.2%). In contrast to these results, Gotkowska-Płachta,20 reported E. faecium (42.9%) as the most prevalent isolate in HWW samples, followed by E. faecalis (31.0%). However, the predominance of the two species of E. faecium and E. faecalis compared to other strains in their study was similar to the present study. In a study by Molale-Tom et al21 from South Africa, the most prevalent Enterococcus isolate in the wastewater treatment plant samples was E. hirae (21.0%), followed by E. faecalis (21.0%) and E. faecium (19.0%). Iweriebor et al22 reported the E. faecalis as the most prevalent isolate in hospital and domestic wastewater effluents that was similar to the current study. Depending on the type of wastewater and the treatment methods used, the variety of the wastewater microbiota and their antibiotic resistance profile may be different.20 There are several survival strategies that distinct species of enterococci can engage them to persist in different environments, including starvation and the viable but non-culturable (VBNC) state. These strategies allow enterococci to survive even in chlorinated wastewater.22
This study investigated the resistance patterns of enterococci against different antibiotics. The results revealed that ampicillin and tetracycline with resistance rates of 58.8% and 55.3% were the less effective antibiotics, respectively. These results were consistent with a report by Molale-Tom et al21 from South Africa. However, they reported a low resistance rate (8.0%) against ciprofloxacin that was in contrast to the finding of the current study (54.1%). In this study, the overall resistance rate to trimethoprim-sulfamethoxazole was 31.8%, although this rate was significantly higher in E. faecalis strains than E. faecium. This result was lower than a previous report by Gotkowska-Płachta (87.5%) from Poland.20 In another study, Matlou et al23 reported a resistance rate of 12.5% for sulfamethoxazole antibiotic among enterococci isolates from surface and ground water. One of the unexpected results of this study was the lower resistance rate to vancomycin in comparison to previous reports. In this study, the resistance rates of vancomycin and teicoplanin were 29.4% and 28.2%, respectively. These lower resistance rates may be due to the low prescription of glycopeptides in treatment protocols of healthcare systems. Several previous studies by Gotkowska-Płachta et al (more than 75.0%),20 Molale-Tom et al (62.0%),21 Iweriebor et al (more than 80%),22 Matlou et al (78.6%),23 and Oravcova et al (86.0%)24 have reported high level resistance rates to vancomycin and/or teicoplanin among enterococci isolated from aquatic environments, which contradicted the results of the current study. However, low resistance rates of 2.8% and 3.3% have been reported by Karimi et al25 from Iran and Mbanga et al26 from South Africa, respectively. Even in advanced countries, medical professionals face significant challenges when dealing with VRE strains, particularly if they have multiple antibiotic resistance factors.23 VanA, B, C, D, E, G, L, M, and N are currently known operons of vancomycin resistance in enterococci species.27
The current study showed a relatively low percentage of resistance to linezolid (8.2%) and tigecycline (12.9%) among enterococci. However, these rates were higher than the previous studies conducted in Czech Republic (linezolid 0.0% and tigecycline 0.0%)24 and Iran (linezolid 3.4%).25 Also, the low resistance rates of rifampin (9.4%) and imipenem (8.2%) confirmed that these antibiotics are still among the best effective choices against enterococci. These findings were contrary to previous reports from Poland,20 Czech Republic,24 and Pakistan.28
Almost more than third of the strains showed resistance against erythromycin (44.7%) and chloramphenicol (38.8%). Molale-Tom et al21 from South Africa, reported higher and lower resistance rates against erythromycin (51.0%) and chloramphenicol (22.0%), respectively. Also, Matlou et al (30.4%)23 and Oravcova et al (0.0%)24 reported lower resistance rates against chloramphenicol than the current research. However, higher resistance proportions against erythromycin were reported in previous studies from Czech Republic24 and Iran25 in comparison to the current results.
In this work, 57.6% of enterococci isolates were MDR. This proportion was lower than previous reports from South Africa (68.0%, 80.4%, 80.8%)21,23,26 and Poland (94.0%).20 Nevertheless, the fact that more than half of the isolates tested had MDR in this study was a warning sign that regulatory measures need to be taken to prevent this phenomenon from more spreading.
In this research, 63.5% (54/85) of enterococci isolates from HWWs had CRISPR-Cas elements. To the best of our knowledge, this experiment was one of the rare studies that has dealt with this issue. A previous study by Dos Santos et al,14 showed a lower frequency rate (51.1%) of CRISPR-Cas in enterococci collected from human and food sources. Using PCR, the frequency of CRISPR2 (55.3%) was higher than CRISPR1 (41.2%) and CRISPR3 (35.3%) in this study which was in line with previous reports by Gholizadeh et al11 from Iran and Huescas et al29 from Brazil. In contrast to our results, Dos Santos et al14 reported CRISPR3 (67.4%) as the most prevalent type in enterococci isolates. In a previous study by Sanderson et al,30 orphan CRISPR (without Cas genes) was detected in 27 genomes of 39 enterococci isolates from wastewater treatment plants using bioinformatics experiments by CRISPRdb software. Another study by Lyons et al31 that focused on prevalence of CRISPR1-cas1 gene in enterococci isolates from non-clinical and environmental sources, the aforesaid gene was found in 32.7% of all isolates that was lower than the current findings. Also, they reported the occurrence of CRISPR1-cas1 gene in 23.6%, 73.1%, and 7.1% of E. faecalis, E. hirae, and E. faecium strains, respectively. In this study, the prevalence of CRISPR1 was higher in E. faecalis (50.0%) than E. faecium isolates (28.6%) that was similar to Lyons et al.31 In the current study, the frequency of CRISPR-Cas systems was significantly higher in E. faecalis (80.0%) than E. faecium (40.0%). This finding was in line with a previous report by Huescas et al29 from Brazil, who found that the CRISPR-Cas elements of E. faecalis were detected more often than those of E. faecium in non-clinical samples. However, they reported more and less prevalence of CRISPR2 and CRISPR3-cas in the E. faecalis and E. faecium isolates from food and animal fecal samples than in our study, respectively.
This study revealed the incidence of 50.0%, 78.0%, and 36.0% for CRISPR1, CRISPR2, and CRISPR3 in E. faecalis isolates, respectively. In a previous report by Gholizadeh et al11 from Iran, the attendance of CRISPR1-cas, orphan CRISPR2, and CRISPR3-cas was seen in 13%, 55.3%, and 17.4% of clinical E. faecalis, respectively. Also, in another study from Iran, the prevalence rates of 53.4%, 17.7%, and 10.4% were reported for CRISPR2, CRISPR3, and CRISPR1 in E. faecalis isolates from urinary tract infections (UTI) and dental-root canal (DRC) samples.32 Also, this research showed the prevalence of 28.6%, 22.9%, and 4.3% for CRISPR1, CRISPR2, and CRISPR3 in E. faecium isolates, respectively. In this regard, Huescas et al29 reported a higher prevalence rate of CRISPR2 (32.7%), but lower frequency of CRISPR1-cas (5.6%) and CRISPR3-cas (1.8%) genes in non-clinical E. faecium isolates. These discrepancies in the results of various studies can be due to the sample size, the source of the sample collection, and the technique used in the screening of CRISPR-Cas systems.
One of the strengths of the current study was evaluating of the association of CRISPR-Cas systems presence with resistance against a wide range of antibiotics in non-clinical enterococci isolates. Given that few studies have addressed this issue so far, the results of this study provided valuable epidemiological information. The results showed that there was a significant negative association between the occurrence of CRISPR-Cas elements and antibiotic resistance rates against vancomycin, ampicillin, chloramphenicol, erythromycin, rifampin, teicoplanin, tetracycline, imipenem, tigecycline, and trimethoprim- sulfamethoxazole in E. faecalis isolates. Also, the CRISPR-Cas positive E. faecium isolates showed significant lower resistance rates against vancomycin, ampicillin, chloramphenicol, erythromycin, teicoplanin, and tetracycline in comparison with the CRISPR-Cas negative E. faecium strains. Notably, the presence of CRISPR-Cas elements was lower in MDR isolates compared to non MDR enterococci isolates. These results were in good agreement with a recent study by Gholizadeh et al32 who showed that CRISPR-cas loci are negatively correlated with antibiotic resistance, as well as carrying antibiotic-resistant genes in E. faecalis isolates from UTI and DRC samples. Also, our results confirmed the previous findings obtained by Dos Santos et al,14 Palmer and Gilmore,33 and Burley and Sedgley34 who showed that MDR properties among enterococci isolates were associated with a lack of CRISPR-Cas systems. It is believed that CRISPR-Cas acts as a barrier to the acquisition of antibiotic resistance genes because most of these genes are commonly spread by plasmids in enterococci isolates.32 Consistent with the current study, the negative effects of the occurrence of CRISPR-Cas elements against antibiotic resistance has been reported in other bacteria including Klebsiella pneumoniae,13,35 and Escherichia coli.36 Also, Ren et al37 showed that point mutation in cas1 and cas2 genes may be contribute to multiple drug resistance in Shigella species. However, in contrast to the aforesaid studies, Shabbir et al9 claimed that CRISPR-Cas system promotes antimicrobial resistance in Campylobacter jejuni.
The present study had some limitations as follows: the lack of sequencing of the CRISPR-Cas elements and also not examining the clonal genetic linkage of the enterococci due to limited financial resources.
Conclusion
This study was among the rare studies that addressed the prevalence of CRISPR-Cas systems and their effects on antibiotic resistance in enterococci collected from HWW samples. The results showed the higher prevalence of E. faecalis than E. faecium in HWWs. Also, different abundance rates of various type II CRISPR-Cas systems with the predominance of CRISPR2 were observed among E. faecalis and E. faecium isolates from non-clinical origin. In this research, a significant negative association was seen between the presence of CRISPR-Cas systems and antibiotic resistance against penicillins, carbapenems, folate pathway antagonists, phenicols, glycopeptides, tetracyclines, ansamycins, and macrolides among enterococci isolates. Similarly, the presence of CRISPR-Cas was associated with lower multidrug resistance of enterococci isolates. This inverse relationship may provide new impetus for the fight against infection caused by MDR pathogens with the help of CRISPR-Cas elements.
Data Sharing Statement
All data generated or analyzed during this study are included here and are available from the corresponding author on reasonable request.
Ethics Approval and Informed Consent
This study was approved by the Ethics Committee of the University of Kufa, Najaf, Iraq (ethics code: MEC-10) following the Declaration of Helsinki. No human and animal samples were used in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Chia PY, Sengupta S, Kukreja A, Ponnampalavanar SS, Ng OT, Marimuthu K. The role of hospital environment in transmissions of multidrug-resistant gram-negative organisms. Antimicrob Resist Infect Control. 2020;9(1):29. doi:10.1186/s13756-020-0685-1
2. Gouliouris T, Raven KE, Moradigaravand D, et al. Detection of vancomycin-resistant Enterococcus faecium hospital-adapted lineages in municipal wastewater treatment plants indicates widespread distribution and release into the environment. Genome Res. 2019;29(4):626–634. doi:10.1101/gr.232629.117
3. Kraemer SA, Ramachandran A, Perron GG. Antibiotic pollution in the environment: from microbial ecology to public policy. Microorganisms. 2019;7(6):180. doi:10.3390/microorganisms7060180
4. Růžičková M, Vítězová M, Kushkevych I. The characterization of Enterococcus genus: resistance mechanisms and inflammatory bowel disease. Open Med. 2020;15(1):211–224. doi:10.1515/med-2020-0032
5. Remschmidt C, Schröder C, Behnke M, Gastmeier P, Geffers C, Kramer TS. Continuous increase of vancomycin resistance in enterococci causing nosocomial infections in Germany− 10 years of surveillance. Antimicrob Resist Infect Control. 2018;7(1):1–7. doi:10.1186/s13756-018-0353-x
6. Boccella M, Santella B, Pagliano P, et al. Prevalence and antimicrobial resistance of Enterococcus species: a retrospective cohort study in Italy. Antibiotics. 2021;10(12):1552. doi:10.3390/antibiotics10121552
7. Ramos S, Silva V, Dapkevicius MD, Igrejas G, Poeta P. Enterococci, from harmless bacteria to a pathogen. Microorganisms. 2020;8(8):1118. doi:10.3390/microorganisms8081118
8. García-Solache M, Rice LB. The Enterococcus: a model of adaptability to its environment. Clin Microbiol Rev. 2019;32(2):e00058–18. doi:10.1128/CMR.00058-18
9. Shabbir MA, Wu Q, Shabbir MZ, et al. The CRISPR- cas system promotes antimicrobial resistance in Campylobacter jejuni. Fut Microbiol. 2018;13(16):1757–1774. doi:10.2217/fmb-2018-0234
10. Kamruzzaman M, Iredell J. CRISPR-Cas system in antibiotic resistance plasmids in Klebsiella pneumoniae. Front Microbiol. 2020;10:2934. doi:10.3389/fmicb.2019.02934
11. Gholizadeh P, Aghazadeh M, Ghotaslou R, et al. CRISPR-cas system in the acquisition of virulence genes in dental-root canal and hospital-acquired isolates of Enterococcus faecalis. Virulence. 2020;11(1):1257–1267. doi:10.1080/21505594.2020.1809329
12. Gholizadeh P, Köse Ş, Dao S, et al. How CRISPR-Cas system could be used to combat antimicrobial resistance. Infect Drug Resist. 2020;13:1111–1121. doi:10.2147/IDR.S247271
13. Wang G, Song G, Xu Y. Association of CRISPR/Cas system with the drug resistance in Klebsiella pneumoniae. Infect Drug Resist. 2020;13:1929. doi:10.2147/IDR.S253380
14. Dos Santos BA, de Oliveira JD, Parmanhani-da-silva BM, Ribeiro RL, Teixeira LM, Neves FP. CRISPR elements and their association with antimicrobial resistance and virulence genes among vancomycin-resistant and vancomycin-susceptible enterococci recovered from human and food sources. Infect Genet Evol. 2020;80:104183. doi:10.1016/j.meegid.2020.104183
15. Ahmed ES, El-Tahan MH, Khalafall GM, Sedik MZ. Physico-chemical and microbial profile of agriculture wastewater samples from El Maryotia canal in Egypt. Plant Arch. 2021;21(1):650–657. doi:10.51470/PLANTARCHIVES.2021.v21.S1.099
16. Mariam SH. A sampling survey of enterococci within pasteurized, fermented dairy products and their virulence and antibiotic resistance properties. PLoS One. 2021;16(7):e0254390. doi:10.1371/journal.pone.0254390
17. Iweriebor BC, Obi LC, Okoh AI. Virulence and antimicrobial resistance factors of Enterococcusspp. isolated from fecal samples from piggery farms in Eastern Cape, South Africa. BMC Microbiol. 2015;15(1):136. doi:10.1186/s12866-015-0468-7
18. CLSI. Performance Standards for Antimicrobial Susceptibility Testing.
19. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters, version 8.1; 2018.
20. Gotkowska-Płachta A. The prevalence of virulent and multidrug-resistant enterococci in river water and in treated and untreated municipal and hospital wastewater. Int J Environ Res Public Health. 2021;18(2):563. doi:10.3390/ijerph18020563
21. Molale-Tom LG, Bezuidenhout CC. Prevalence, antibiotic resistance and virulence of Enterococcus spp. from wastewater treatment plant effluent and receiving waters in South Africa. J Water Health. 2020;18(5):753–765. doi:10.2166/wh.2020.086
22. Iweriebor BC, Gaqavu S, Obi LC, Nwodo UU, Okoh AI. Antibiotic susceptibilities of enterococcus species isolated from hospital and domestic wastewater effluents in Alice, eastern cape province of South Africa. Int J Environ Res Public Health. 2015;12(4):4231–4246. doi:10.3390/ijerph120404231
23. Matlou DP, Bissong ME, Tchatchouang CD, et al. Virulence profiles of vancomycin-resistant enterococci isolated from surface and ground water utilized by humans in the North West Province, South Africa: a public health perspective. Environ Sci Pollut Res. 2019;26(15):15105–15114. doi:10.1007/s11356-019-04836-5
24. Oravcova V, Mihalcin M, Zakova J, Pospisilova L, Masarikova M, Literak I. Vancomycin-resistant enterococci with vanA gene in treated municipal wastewater and their association with human hospital strains. Sci Total Environ. 2017;609:633–643. doi:10.1016/j.scitotenv.2017.07.121
25. Karimi F, Samarghandi MR, Shokoohi R, Godini K, Arabestani MR. Prevalence and removal efficiency of enterococcal species and vancomycin-resistant enterococci of a hospital wastewater treatment plant. Avicenna J Environ Health Eng. 2016;3(2):e8623. doi:10.5812/ajehe.8623
26. Mbanga J, Abia AL, Amoako DG, Essack SY. Longitudinal surveillance of antibiotic resistance in Escherichia coli and Enterococcus spp. from a wastewater treatment plant and its associated waters in KwaZulu-Natal, South Africa. Microb Drug Resist. 2021;27(7):904–918. doi:10.1089/mdr.2020.0380
27. George SK, Suseela MR, El Safi S, et al. Molecular determination of van genes among clinical isolates of enterococci at a hospital setting. Saudi J Biol Sci. 2021;28(5):2895–2899. doi:10.1016/j.sjbs.2021.02.022
28. Rehman M, Hasan KA, Bin-Asif H, et al. Differentiating Enterococcus lineages in combined sewer overflow and potable water combating to hospital acquired high-level β-lactam resistance. Environ Challenges. 2021;4:100094. doi:10.1016/j.envc.2021.100094
29. Huescas CGY, Pereira RI, Prichula J, Azevedo PA, Frazzon J, Frazzon APG. Frequency of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs) in non-clinical Enterococcus faecalis and Enterococcus faecium strains. Braz J Biol. 2019;79(3):460–465. doi:10.1590/1519-6984.183375
30. Sanderson H, Ortega-Polo R, Zaheer R, et al. Comparative genomics of multidrug-resistant Enterococcus spp. isolated from wastewater treatment plants. BMC Microbiol. 2020;20(1):1–7. doi:10.1186/s12866-019-1683-4
31. Lyons C, Raustad N, Bustos MA, Shiaris M. Incidence of type II CRISPR1-Cas systems in Enterococcus is species-dependent. PLoS One. 2015;10(11):e0143544. doi:10.1371/journal.pone.0143544
32. Gholizadeh P, Aghazadeh M, Ghotaslou R, et al. Role of CRISPR-Cas system on antibiotic resistance patterns of Enterococcus faecalis. Ann Clin Microbiol Antimicrob. 2021;20(1):49. doi:10.1186/s12941-021-00455-6
33. Palmer KL, Gilmore MS. Multidrug-resistant enterococci lack CRISPR-cas. MBio. 2010;1(4):e00227–00210. doi:10.1128/mBio.00227-10
34. Burley KM, Sedgley CM. CRISPR-Cas, a prokaryotic adaptive immune system, in endodontic, oral, and multidrug-resistant hospital-acquired Enterococcus faecalis. J Endod. 2012;38(11):1511–1515. doi:10.1016/j.joen.2012.07.004
35. Mackow NA, Shen J, Adnan M, Khan AS, Fries BC, Diago-Navarro E. CRISPR-Cas influences the acquisition of antibiotic resistance in Klebsiella pneumoniae. PLoS One. 2019;14(11):e0225131. doi:10.1371/journal.pone.0225131
36. Aydin S, Personne Y, Newire E, et al. Presence of Type IF CRISPR/Cas systems is associated with antimicrobial susceptibility in Escherichia coli. J Antimicrob Chemother. 2017;72(8):2213–2218. doi:10.1093/jac/dkx137
37. Ren L, Deng LH, Zhang RP, et al. Relationship between drug resistance and the clustered, regularly interspaced, short, palindromic repeat-associated protein genes cas1 and cas2 in Shigella from giant panda dung. Medicine. 2017;96(7):e5922. doi:10.1097/MD.0000000000005922
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.