Back to Journals » Infection and Drug Resistance » Volume 14
Prevalence and Pattern of Resistance in NS5A/NS5B in Hepatitis C Chronic Patients Genotype 3 Examined at a Public Health Laboratory in the State of São Paulo, Brazil
Authors de Torres Santos AP ![]() , Martins Silva VC, Mendes-Corrêa MC
, Martins Silva VC, Mendes-Corrêa MC ![]() , Lemos MF, de Mello Malta F
, Lemos MF, de Mello Malta F ![]() , Santana RAF
, Santana RAF ![]() , Dastoli GTF, de Castro VFD, Pinho JRR, Moreira RC
, Dastoli GTF, de Castro VFD, Pinho JRR, Moreira RC ![]()
Received 24 January 2020
Accepted for publication 12 September 2020
Published 24 February 2021 Volume 2021:14 Pages 723—730
DOI https://doi.org/10.2147/IDR.S247071
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Ana Paula de Torres Santos,1,2 Vanessa Cristina Martins Silva,1 Maria Cássia Mendes-Corrêa,3 Marcilio Figueiredo Lemos,1 Fernanda de Mello Malta,4 Rúbia Anita Ferraz Santana,5 Gregório Tadeu Fernando Dastoli,5 Vanessa Fusco Duarte de Castro,5 João Renato Rebello Pinho,2,4,5 Regina Célia Moreira1
1Laboratory of Viral Hepatitis, Virology Center, Instituto Adolfo Lutz, São Paulo, SP, Brazil; 2Divisão de Laboratório Central, Laboratório de Imunologia, Faculdade de Medicina, Hospital das Clínicas, Universidade de São Paulo, São Paulo, SP, Brazil; 3LIM-52-Institute of Tropical Medicine, Department of Infectious Diseases, School of Medicine, University of São Paulo, São Paulo, SP, Brazil; 4Laboratory of Tropical Gastroenterology and Hepatology “João de Queiroz and Castorina Bettencourt Alves”‑LIM 07‑Institute of Tropical Medicine Department of Gastroenterology, School of Medicine, University of São Paulo, São Paulo, SP, Brazil; 5Albert Einstein Medicina Diagnóstica, Hospital Israelita Albert Einstein, São Paulo, SP, Brazil
Correspondence: Regina Célia Moreira
Instituto Adolfo Lutz, Centro de Virologia, Av: Dr Arnaldo, 355 Pacaembu, SP, Cep 01246-000, Brazil
Tel +55 11 30682911
Email [email protected]
Purpose: Globally, it is estimated that 71 million people are chronically infected with hepatitis C, and 10– 20% of these will develop cirrhosis and hepatocellular carcinoma. The development of new direct-acting antiviral (DAA) drugs has contributed to sustained virological response (SVR), eliminating the infection and achieving cure of chronic hepatitis C. However, treated patients can develop HCV resistance to DAAs, which can contribute to the failure of treatment. Here, we aimed to evaluate the prevalence and specific pattern of NS5A and NS5B resistance-associated substitutions (RAS) in samples from patients chronically infected with HCV genotype 3a at a public health laboratory, Instituto Adolfo Lutz, São Paulo, Brazil.
Patients and Methods: Serum samples from the enrolled individuals were submitted to “in-house” polymerase chain reaction amplification of NS5A and NS5B non-structural protein genes, which were then sequenced by Sanger method.
Results: A total of 170 and 190 samples were amplified and analyzed for NS5A and NS5B, respectively. For NS5A, 20 (12.0%) samples showed some important RAS; 16 (9.0%) showed some type of substitution and 134 (79.0%) showed no polymorphism. No sample showed any RAS for NS5B.
Conclusion: This study found important RAS in samples from naïve chronic HCV patients in some areas from São Paulo. The most prevalent were A62S, A30K, and Y93H, which could indicate an increase in resistance to some DAAs used in HCV treatment.
Keywords: HCV, nonstructural NS5A/NS5B, resistance, polymorphism, RAS
Introduction
Hepatitis C virus (HCV) infection is one of the main causes of chronic liver disease, with approximately 71 million chronically infected individuals worldwide, many of whom are unaware of their infection. Developments in diagnostic procedures and improvements in therapy, with the discovery of new drugs, and prevention have ensured better clinical care for patients with HCV-related liver disease.1–3
HCV is classified into 7 genotypes (GT) and approximately 67 subtypes. GT1 is the most prevalent HCV genotype worldwide followed by GT3 (22%), GT2 (13%), and GT4 (13%). Studies carried out in Brazil have indicated a GT3 prevalence of 30.2%.4 Recent studies have shown that HCV GT3 is associated with more rapid disease progression and lower rates of response to treatment, in comparison with other genotypes, especially in patients with cirrhosis and those who have not showed response to previous treatment.5,6
The non-structural proteins NS3/NS4, NS5A, and NS5B, together, contribute to the HCV life cycle, including HCV RNA translation, posttranslational processing, HCV replication, and virus assembly and release.7,8 The goal of therapy of HCV infection is sustained virological response (SVR), and the development of direct antiviral agents (DAAs) has revolutionized the therapeutics for chronic hepatitis C. These include the NS3/4A protease inhibitors, NS5A inhibitors, and NS5B polymerase inhibitors, which are further subdivided into nucleoside and non-nucleoside polymerase inhibitors, with or without ribavirin, besides the combined therapies.9,10
Resistance-associated substitutions (RAS) are generated at a baseline and in chronic hepatitis C patients who fail to respond to DAA treatment. RAS in NS5A may affect the efficacy of re-treatment, as NS5A inhibitor is included in all available DAA regimens.11 A study by Hezode et al in G1-G6 HCV patients, treated with Sofosbuvir and Velpatasvir, detected 28% of RAS in NS5A at baseline, but irrespective of this, high rates of SVR were observed at 12 weeks. In addition, the study did not report any observed resistance of NS5B.6
The current study aimed to evaluate the prevalence and the specific pattern of NS5A and NS5B RAS in samples from DAA naïve patients chronically infected with HCV (GT3), whose samples were sent to a public health laboratory of Viral Hepatitis, Instituto Adolfo Lutz, São Paulo, Brazil.
Methods
Samples
This study included serum samples from DAA naïve patients chronically infected with HCV GT3a, and those were sent to a public health laboratory in São Paulo State/Brazil, from January 2015 to February 2016; the serum samples were stored at −20°C until use. The samples were obtained from different regions of São Paulo State: Vale do Paraíba, Vale do Ribeira, and São Paulo Metropolitan area (São Paulo and ABCD).
HCV RNA Extraction
HCV RNA was extracted from 200 µL of plasma using the NucliSENS easyMAG™ kit (BioMérieux, Marcy l’Etoile, France), following the manufacturer’s instructions.
Amplification of the NS5A Region
The HCV NS5A region was amplified by PCR in two overlapping parts using the same sets of primers (Table 1). Complementary DNA (cDNA) and the first round were performed using the SuperScript III One-Step RT-PCR System with the Platinum Taq DNA Polymerase kit (Invitrogen™, ThermoFisher Brand, Carlsbad, USA). The reverse transcription was performed under the following conditions: an initial denaturation step of 50°C for 30 min and 94°C for 2 min, followed by 35 cycles of 94°C for 15 s, 62°C for 30 s, and 68°C for 90 s, and a final extension at 68°C for 5 min. The second round was performed using Platinum Taq kit (Invitrogen™, ThermoFisher Brand, Carlsbad, USA) with a genotype-specific primer for GT3a, as described previously.12 After the amplification cycles, a final extension at 72°C for 7 min was added for all reactions. The amplified fragments were visualized by 2% agarose gel electrophoresis and gel red staining.
 |
Table 1 Oligonucleotides for NS5A and NS5B Gene Amplification of HCV |
Amplification of the NS5B Region
For the NS5B region, the cDNA was generated using M-MLV reverse transcriptase and random hexamers (InvitrogenTM, Carlsbad, CA, USA). After incubation at 70°C for 10 min, reverse transcription was carried out at 37°C for 60 min, followed by heat inactivation at 95°C for 15 min. The HCV NS5B region was amplified by nested PCR in two overlapping parts (primers are described in Table 1).13 The first round of PCR was performed under the following conditions: 49 cycles of 95°C for 30 s, 63°C for 30 s and 72°C for 30 s, followed by a final extension at 72°C for 10 min. The second round was performed under the following conditions: an initial denaturation step of 95°C for 5 min, 34 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 30 s, followed by a final extension at 72°C for 10 min. The amplified fragments were visualized by 2% agarose gel electrophoresis and gel red staining.
Sequence Analysis
After quantification of the amplified product, each sample was sequenced using the commercial BigDye® Terminator v3.1 Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Inc., USA). For the NS5A region, an initial purification step was performed using the ExoSAP-IT PCR Clean-up Kit (GE Healthcare, Chalfont St. Giles, UK), following the manufacturer’s instructions. The final fragments were purified using the Ethanol/EDTA method, following the instructions of the BigDye® Terminator kit. Thereafter, the sequencing reaction was analyzed on an ABI3500 automated sequencer (Applied Biosystems, Thermo Fisher Brand, Foster City, CA, USA).
Chromatograms obtained from the sense and antisense sequences were edited to obtain consensus sequences using Sequencher™ 4.7 software (Gene Codes Corporation, Michigan, USA). The file generated in the “fasta” format was submitted to Geno2Pheno site (https://hcv.geno2pheno.org/) for characterization of genotypes and identification of polymorphisms and mutations related to drug resistance.
Results
Study Population
All samples were from naïve DAA patients. For NS5A, 87 patients were from São Paulo Metropolitan area and 83 from Vale do Paraiba or Vale do Ribeira. For NS5B, 108 patients were from São Paulo Metropolitan area and 90 from Vale do Paraiba or Vale do Ribeira. Patients were waiting for new drugs, and all were being monitored to receive DAAs from public health service. RAS was analyzed in sequences obtained from chronic HCV-infected patients naïve to DAA - NS5A (N = 170; 110 males and 60 females) and NS5B-inhibitors (N = 198; 126 males and 72 females). Viral load of these patients ranged from log 2.78 – log 7.24 (median log 5.74). Patients were aged 17 to 79 years (median 54 years).
NS5A Region
A total of 170 HCV GT3a samples were amplified, sequenced, and analyzed for NS5A RAS and 134 (79.0%) samples showed no polymorphism; 16 (9.0%) samples showed natural polymorphism and 20 (12.0%) samples showed RAS at baseline. As the majority of samples were from male patients (64.7%) and aged from 40 to 59 years old, these were the groups with the greatest number of detected polymorphisms for NS5A. These data are shown in Table 2.
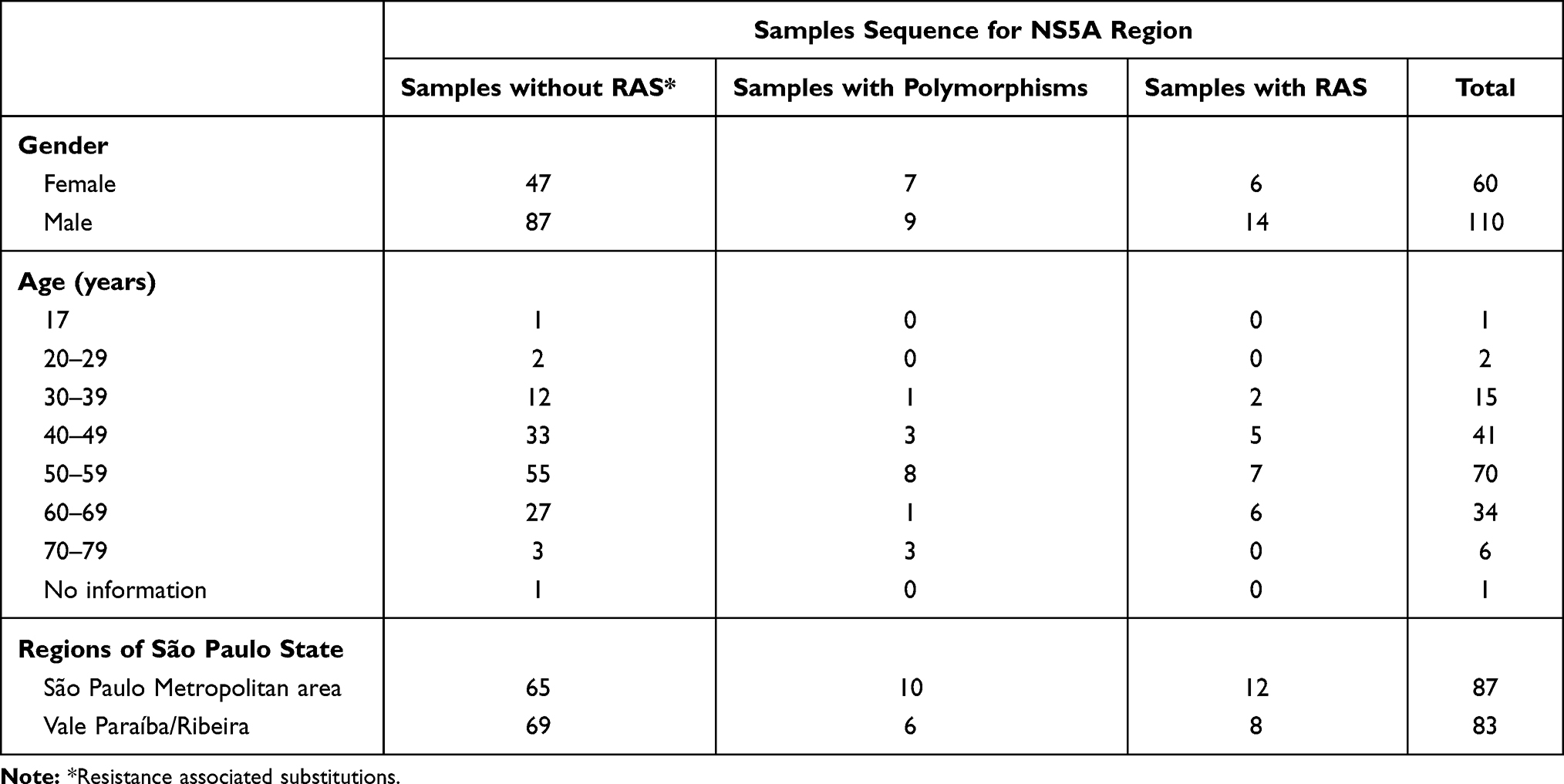 |
Table 2 Demographic Characterization of the Patients According to NS5A Region Results |
Some relevant polymorphisms were found in our analysis. From 170 samples analyzed for NS5A, we observed RAS in 20 samples (12.0%) (Table 3). Among mutations in the NS5A detected for Daclatasvir, the most frequent RAS was A30K+A62S (70.0%), followed by A30T/E/K/S (10.0%) and polymorphisms A30K+A62P, A30S+Y93H+A62M, Y93H+A62L, and A30K (with the frequency of 5.0% each). For Elbasvir and Ledispavir the most frequent RAS was A30K (80.0%), followed by A30T/E/K (10.0%), A30S+Y93H (5.0%), and Y93H (5.0%). For Velpatasvir, only 2 samples showed RAS, 1 sample with polymorphism A30S+Y93H and 1 sample with Y93H. For Pibrentasvir, no samples showed RAS. These polymorphisms are described in Table 4.
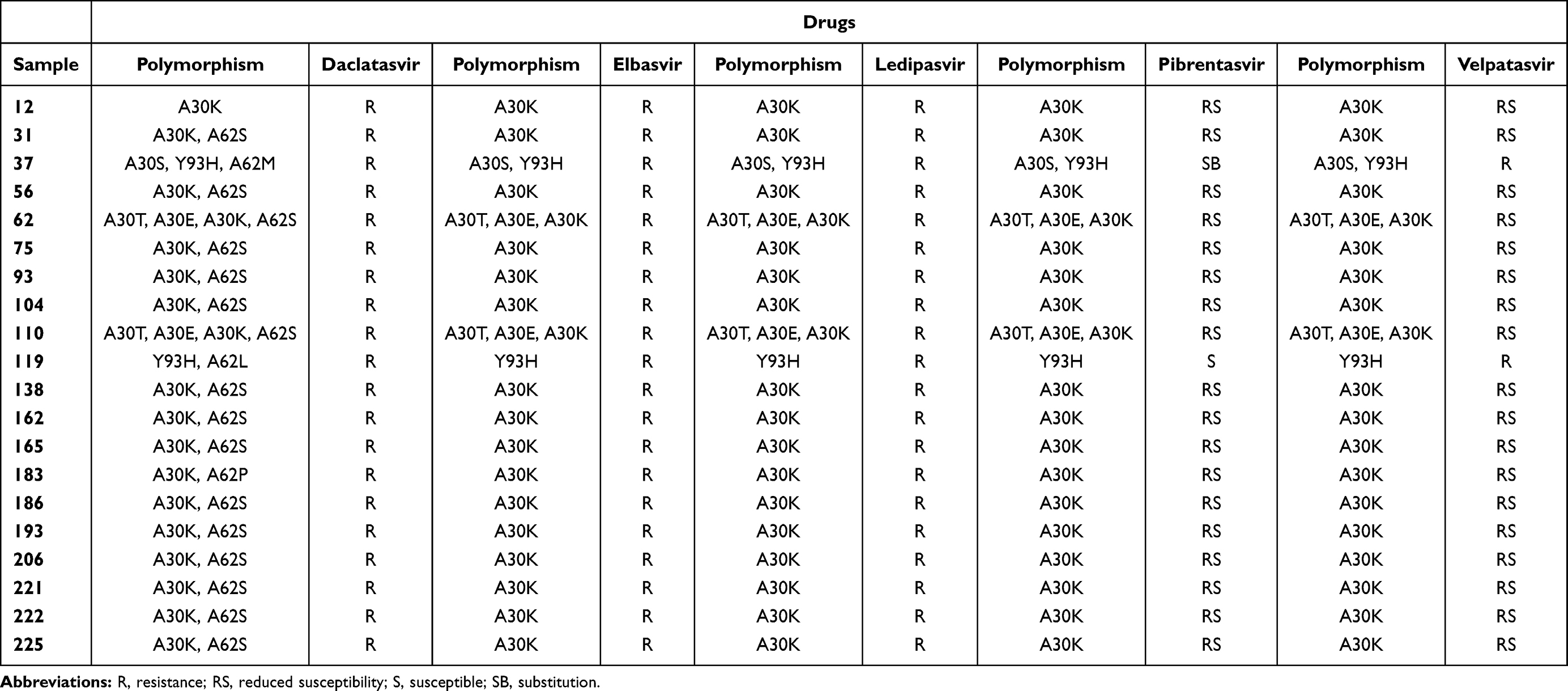 |
Table 3 Distribution of the Amino Acid Mutations in the HCV NS5A Region in DAA-Naïve Patients Infected with HCV Genotype 3a (n=20) |
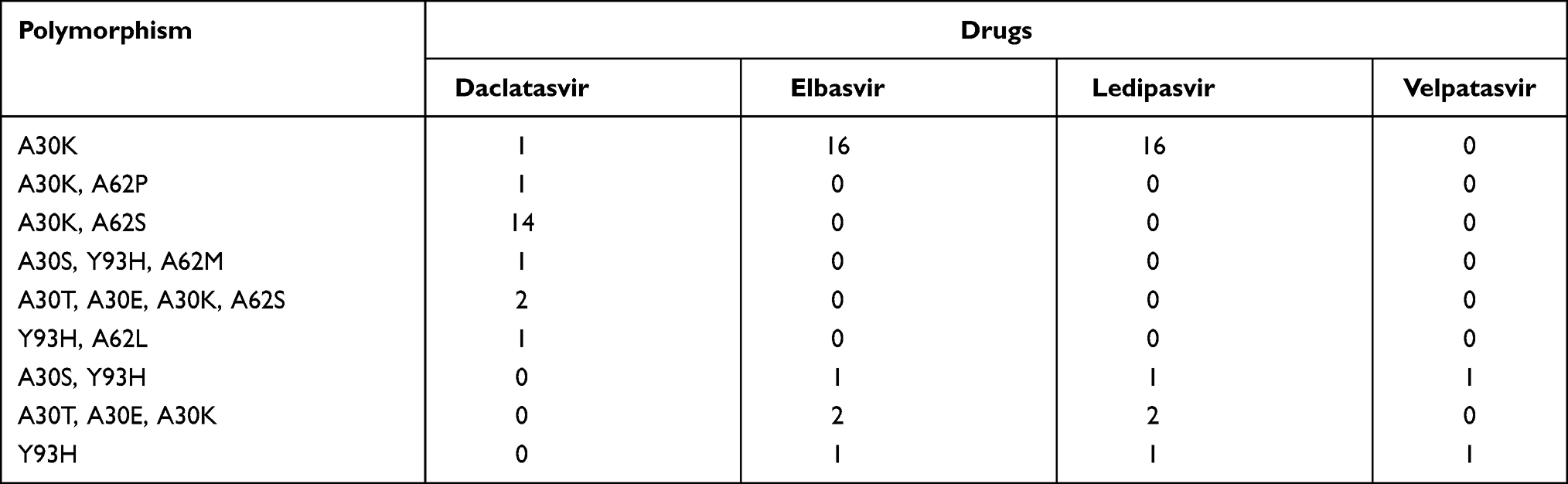 |
Table 4 Frequency of Mutations Associated with Resistance to DAAs in the NS5A Region of Genotype 3a |
NS5B Region
None of the 198 samples of HCV GT3a analyzed for NS5B region showed RAS for sofosbuvir.
Discussion
Assessment of RAS in baseline in a candidate for therapy seems to be an option to define therapeutic strategies and to predict resistance to sustained response to DAAs. In Brazil, DAAs are freely dispensed in the entire country for the treatment of chronic HCV patients. Instituto Adolfo Lutz in São Paulo, besides performing diagnostic test by viral load, also monitors the emergence of resistance strains in this population. Samples are not sent to the laboratory with medical records or any clinical information. Hence, it was not possible to follow up with the patients after drug administration; it was only possible to detect RAS in a pre-administration phase of the drugs.
Sequencing was performed by Sanger method,14 which is useful in a diagnostic laboratory and for public health. Although deep sequence is more sensitive than other methods, Sanger sequencing is easier to perform and most of the studies on HCV RAS have used this methodology to detect polymorphisms that are clinically important.15,16 Pawlotsky (2016) highlighted the importance of resistance in baseline and how Sanger method was useful for detecting RAS in diagnostic laboratory and detect strains with clinical importance.9
The population examined in the current study was heterogeneous, composed mainly by men above 50 years old, from different regions of São Paulo state and with high viral load. These characteristics represent most of the chronic HCV patients waiting for DAA treatment in Brazil and maybe these results could help other patients in the country. Besides, GT3a is the second most prevalent HCV genotype in Brazil and is associated with faster disease progression and lower rates of response to treatment, especially in patients with cirrhosis.4,17
The main polymorphisms observed in this study were A62S, A30K, and Y93H, alone or associated with another polymorphism, which confers high levels of resistance to the NS5A inhibitors Daclatasvir, Ledipasvir, Elbasvir, and Velpatasvir, but not to Pibrentasvir. The presence of NS5A polymorphisms at position 62 can reduce susceptibility to some NS5A inhibitors in other genotypes, but for GT3, their impact is unclear.18 A previous study suggested that the variations at position 62 did not have substantial impact on daclatasvir potency.19 Hernandez et al observed that A30K and Y93H mutations in NS5A reduced viral susceptibility to Daclatasvir and Ledispavir, respectively. The real impact of these resistances depends on some factors such as drug regimen, treatment adherence, and cirrhosis.20
A previous study showed that RAS Y93H and Q30K confer resistance to daclatasvir, ledipasvir, ombitasvir, elbasvir, and velpatasvir, but has low impact on the potency of pibrentasvir. Pibrentasvir is a next-generation pan-genotypic drug that remains active even with common polymorphisms that can occur for NS5A genotype 3.21 In contrast to the above study, another study demonstrated that Y93H polymorphism has clinical relevance to pibrentasvir.22
In our study, several combinations of RAS were observed, as shown in Tables 3 and 4. Some substitutions when occurring alone have been associated with weak resistance to NS5A inhibitors, but when combined with other substitutions, can decrease the viral sensitivity to DAAs.9,12 In addition, we observed some sequences that contained a polymorphism associated with resistance, although it has not yet been determined to be an important mutation associated with therapeutic failure for some drugs, such as pibrentasvir. The characterization (substitution on scored position) determined by geno2pheno software occurs based on its role in resistance to DAAs, which for pibrentasvir is possible, but not immediate.23 In G3a patients, the A30K substitution has never appeared in combination with Y93H, which could suggest a restriction for co-selection of these RAS in the same genome.24
Variations in amino acid residues can occur in all sites of action of antiviral drugs. According to Cuypers et al, the most important RASs to NS5A inhibitors are M/L28T/V, Q/L30E/H/R/S, L31M/V, H58D, and Y93C/H/N, which are capable of reducing susceptibility to NS5A inhibitors by conformational changes.25
The presence of Y93H in NS5A could facilitate the selection of resistant strains under treatment. Therefore, it is necessary to avoid the fitness of resistant variants as the predominant viral population and prevent DAA failure due to baseline resistant variants. In addition, failing DAA-based therapy should be discontinued as soon as possible to avoid an increase in the frequency of RASs, to preserve HCV re-treatment options.15,24
Daclastavir proved to be superior against resistant variant Y93H, which has a natural prevalence of >10% and displays high level of resistance to both Ledipasvir and Daclatasvir in GT1b replicon cells. Furthermore, for HCV GTs 2–4, Daclatasvir has significantly higher potency in vitro compared to Ledipasvir, with the highest fold resistance values for variant Y93H in GT3.26
Efficacy of the DAAs, depending on baseline polymorphisms, may be associated with the reduction of treatment or re-treatment. Sometimes SVR is not reached or 12 weeks of treatment is not enough. RAS is generated in chronic HCV patients and it could affect re-treatment with other available drugs.27 A recent study showed that a longer duration of treatment with Sofosbuvir+Ledipasvir and the addition of Ribavirin can reduce or even eliminate the impact of baseline NS5A RAVs. Only 50% of the patients that had two or more baseline resistance-related variants cleared the virus, with the lowest SVR rates observed with variant Y93H/N.25
Velpatasvir is a potent pan-genotypic HCV NS5A inhibitor with activity against HCV GT1 to GT6. It has better in vitro activity against GT2 and GT3 than the NS5A inhibitors Ledipasvir and Daclatasvir. A study performed by Lawitz et al observed emergent RAS at three positions in GT3 (Y93, L31, E92) and, although pre-treatment RAS was associated with a slightly low response in patients with GT3, current HCV treatment strategies are based on combined use of DAAs and should overcome any minor impact of pretreatment RASs. Combination treatment with Velpatasvir and Sofosbuvir may provide an effective treatment option for patients infected with HCV GT1 to GT6.28
No polymorphism was found for the NS5B region in the 198 sequences analyzed. Polymerase inhibitors have a high genetic barrier, so few patients show therapeutic failure in regimen containing sofosbuvir. Costa et al, in a study at Rio de Janeiro, G1/G3 HCV patients treated with daclatasvir and sofosbuvir showed high SVR. These authors also observed no NS5A RAS in 28% of HCV/HIV coinfected and 16.8% in monoinfected patients. No RAS was observed for NS5B.29 Sofosbuvir is a nucleoside NS5B polymerase inhibitor with pan-genotypic activity, and whose most frequent resistance mutations are L159F and C316N in GT1a-infected patients, S282G/T and C316N/H/F in the GT1b-infected, L159F and S282T in those with GT2 infection, L159F and V321A in the GT3-infected, and E237G, S282T and V321A, in those with GT4 infection.5 Chen et al also observed that there was a notable absence of RAS in NS5B, except in 1 patient who carried the high-resistance S282T alone in 100% of the viral genomes sequenced.24
In conclusion, this study found important RAS in samples from naïve chronic GT 3a HCV patients in some areas from São Paulo. The most prevalent were A62S, A30K, and Y93H, which could indicate an increase in resistance to some DAAs offered to treat HCV.
Ethics Statement
The procedures described in this study were initiated only after obtaining approval from the ethics committees of the participating institutions. Following the Ministry of Health guidelines30, informed consent was obtained from all patients who had detectable viral load, allowing the collection and storage of their blood samples for studying Protease Inhibitor (IP) resistance and other drugs for HCV treatment. Further, in this study, we ensured that all sensitive information remained confidential, guaranteeing patient anonymity. Therefore, the requirement for additional consent to conduct this study was waived. This study was approved by the Ethical Committee in Research at the Instituto Adolfo Lutz (CEPIAL) # 1.040.338.
Funding
This study was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP (grant number: #2017/01809-9 and #2020/12529-0).
Disclosure
All the authors who took part in this study declare that they have no conflicts of interest or disclosures with respect to the manuscript.
References
1. Heffernan A, Cooke GS, Nayagam S, Thursz M, Hallett TB. Scaling up prevention and treatment towards the elimination of hepatitis C: a global mathematical model. Lancet. 2019;393(10178):1319–1329. doi:10.1016/S0140-6736(18)32277-3
2. European Association for the Study of the Liver (EASL). Recommendations on treatment of Hepatitis C 2018. J Hepatol. 2018;69(2):
3. World Health Organization (WHO). Global hepatitis report, 2017. Geneva; 2017. Available from: https://apps.who.int/iris/bitstream/handle/10665/255016/9789241565455-eng.pdf.
4. Gower E, Estes C, Blach S, Razavi-Shearer K, Razavi H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J Hepatol. 2014;61(1suppl):S45–S57. doi:10.1016/j.jhep.2014.07.027
5. Sagnelli E, Starace M, Minichini C, et al. Resistance detection and re-treatment options in hepatitis C virus-related chronic liver diseases after DAA-treatment failure. Infection. 2018;46(6):761–783. doi:10.1007/s15010-018-1188-3
6. Hezode C, Reau N, Svarovskaia ES, et al. Resistance analysis in patients with genotype 1–6 HCV infection treated with sofosbuvir/velpatasvir in the Phase III studies. J Hepatol. 2018;68(5):895–903. doi:10.1016/j.jhep.2017.11.032
7. Kim CW, Chang KM. Hepatitis C virus: virology and life cycle. Clin Mol Hepatol. 2013;19(1):
8. Scheel TKD, Rice CM. Understanding the hepatitis C virus life cycle paves the way for highly effective therapies. Nat Med. 2013;19(7):837–849. doi:10.1038/nm.3248
9. Pawlotsky JM. Hepatitis C virus resistance to direct-acting antiviral drugs in interferon-free regimens. Gastroenterology. 2016;151(1):
10. Hussain T. Paritaprevir/ritonavir-ombitasvir and dasabuvir, the 3D regimen for the treatment of chronic hepatitis C virus infection: a concise review. Hepat Med. 2016;8:61–68. doi:10.2147/HMER.S72429
11. Vermehren J, Sarrazin C. The role of resistance in HCV treatment. Best Pract Res Clin Gastroenterol. 2012;26(4):487–503. doi:10.1016/j.bpg.2012.09.011
12. Malta FM, Medeiros-Filho JE, Azevedo RS, et al. Sequencing of E2 and NS5A regions of HCV genotype 3a in Brazilian patients with chronic hepatitis. Mem Inst Oswaldo Cruz. 2010;105(1):92–98. doi:10.1590/S0074-02762010000100014
13. Sandres-Sauné K, Deny P, Pasquier C, Thibaut V, Duverlie G, Izopet J. Determining hepatitis C genotype by analyzing the sequence of the NS5b region. J Virol Methods. 2003;109(2):187–193. doi:10.1016/S0166-0934(03)00070-3
14. Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA. 1977;74(12):5463–5467. doi:10.1073/pnas.74.12.5463
15. Sharafi H, Alavian SM. Hepatitis C resistance to NS5A inhibitors: is it going to be a problem? World J Hepatol. 2018;10(9):543–548. doi:10.4254/wjh.v10.i9.543
16. Sarrazin C. The importance of resistance to direct antiviral drugs in HCV infection in clinical practice. J Hepatol. 2016;64(2):
17. Sorbo MC, Cento V, Di Maio VC, et al. Hepatitis C virus drug resistance associated substitutions and their clinical relevance. Drug Resist Updat. 2018;37:17–39. doi:10.1016/j.drup.2018.01.004
18. Bartolini B, Giombini E, Taibi C, et al. Characterization of naturally occurring NS5A and NS5B polymorphisms in patients infected with HCV genotype 3a treated with direct-acting antiviral agents. Viruses. 2017;9(8):212. doi:10.3390/v9080212
19. Wang C, Valera L, Jia L, Kirk MJ, Gao M, Fridell RA. In vitro activity of daclatasvir on hepatitis C virus genotype 3 NS5A. Antimicrob Agents Chemother. 2013;57(1):
20. Hernandez D, Zhou N, Ueland J, Monikowski A, McPhee F. Natural prevalence of NS5A polymorphisms in subjects infected with hepatitis C virus genotype 3 and their effects on the antiviral activity of NS5A inhibitors. J Clin Virol. 2013;57(1):13–18. doi:10.1016/j.jcv.2012.12.020
21. Ng TI, Krishnan P, Pilot-Matias T, et al. In vitro antiviral activity and resistance profile of the next-generation Hepatitis C Virus NS5A inhibitor pibrentasvir. Antimicrob Agents Chemother. 2017;61(5):e02558–16. doi:10.1128/AAC.02558-16
22. Gottwein JM, Pham LV, Mikkelsen LS, et al. Efficacy of NS5A inhibitors against Hepatitis C Virus genotypes 1–7 and escape variants. Gastroenterology. 2018;154(5):
23. Kalaghatgi P, Sikorski AM, Knops E, et al. Geno2pheno [HCV] - A web-based interpretation system to support Hepatitis C treatment decisions in the era of direct-acting antiviral agents. PLoS One. 2016;11(5):e0155869. doi:10.1371/journal.pone.0155869
24. Chen Q, Perales C, Soria ME, et al. Deep-sequencing reveals broad subtype-specific HCV resistance mutations associated with treatment failure. Antiviral Res. 2020;174:104694. doi:10.1016/j.antiviral.2019.104694
25. Cuypers L, Ceccherini-Silberstein F, Van Laethem K, Li G, Vandamme AM, Rockstroh JK. Impact of HCV genotype on treatment regimens and drug resistance: a snapshot in time. Rev Med Virol. 2016;26(6):408–434. doi:10.1002/rmv.1895
26. Kjellin M, Kileng H, Akaberi D, et al. Effect of the baseline Y93H resistance-associated substitution in HCV genotype 3 for direct-acting antiviral treatment: real-life experience from multicenter study in Sweden and Norway. Scand J Gastroenterol. 2019;54(8):
27. Krishnan P, Pilot-Matias Y, Schnell G, et al. Pooled resistance analysis in patients with Hepatitis C Virus genotype 1 to 6 infection treated with Glecaprevir-Pibrentasvir in Phase 2 and 3 clinical trials. Antimicrob Agents Chemother. 2018;62(10):e01249–18. doi:10.1128/AAC.01249-18
28. Lawitz EJ, Dvory-Sobol H, Doehle BP, et al. Clinical resistance to velpatasvir (GS-5816), a novel pan-genotypic inhibitor of the Hepatitis C Virus NS5A protein. Antimicrob Agents Chemother. 2016;60(9):
29. Costa VD, Brandão-Mello CE, Nunes EP, et al. Treatment of chronic HCV infection with DAAs in Rio de Janeiro/Brazil: SVR rates and baseline resistance analyses in NS5A and NS5B genes. PLoS One. 2019;14(5):e0216327. doi:10.1371/journal.pone.0216327
30. Ministry of Health (MS). Clinical protocol and therapeutic guidelines for viral hepatitis C and coinfections. HCV genotype 1 and advanced fibrosis. Brasília – Df; 2013; Suppl 2:26. Available from: http://www.saude.gov.br/images/pdf/2014/abril/02/pcdt-hepatite-c-suplemento-2013.pdf.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.