")
Back to Journals » OncoTargets and Therapy » Volume 10
Predictive significance of HMGCS2 for prognosis in resected Chinese esophageal squamous cell carcinoma patients
Authors Tang H, Wu Y , Qin Y, Wang H, Jia Y, Yang S, Luo S, Wang Q
Received 17 January 2017
Accepted for publication 22 April 2017
Published 15 May 2017 Volume 2017:10 Pages 2553—2560
DOI https://doi.org/10.2147/OTT.S132543
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Samir Farghaly
Hong Tang,1,* Yufeng Wu,1,* Yanru Qin,2 Hongyan Wang,1 Yongxu Jia,2 Shujun Yang,1 Suxia Luo,1 Qiming Wang1
1Department of Internal Medicine, Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, 2Department of Clinical Oncology, the First Affiliated Hospital, Zhengzhou University, Zhengzhou, Henan, Hong Kong, China
*These authors contributed equally to this work
Abstract: Despite a series of attempts during the last decades, the prognosis of esophageal squamous cell carcinoma (ESCC) remains poor. Different responses of individual tumors encouraged us to look for valuable prognostic markers. As a key regulator controlling the anabolic ketogenic pathway, 3-hydroxy-3-methylglutaryl-CoA synthase 2 (HMGCS2) has been reported to play a crucial role in colorectal cancer and prostate cancer. However, its importance to ESCC has not been verified. Therefore, a large cohort retrospective study was planned, to investigate the relationship between HMGCS2 expression and ESCC prognosis. By adopting real-time polymerase chain reaction (PCR) and immunohistochemical (IHC) staining, HMGCS2 expression was examined in tissues of 300 ESCC patients with complete resection. Besides, the association between HMGCS2 protein expression and survival time was evaluated through chi-square test and Kaplan–Meier analysis. With the use of Cox-proportional hazards model, the prognostic impact of clinicopathologic variables and biomarker expression was evaluated. Compared with their non-tumor counterparts, HMGCS2 downregulation occurred in 65.5% and 37.6% of primary ESCCs on the mRNA and protein levels (P<0.001), respectively. On the protein level, HMGCS2 expression was associated with tumor cell differentiation (P=0.003), pT status (P=0.006), and TNM stage (P=0.010). In the down-HMGCS2 expression group, the 5-year overall survival (OS) and relapse-free survival (RFS) are poorer than those in the normal expression group (19 months vs 24 months, P=0.002; 13 months vs 17 months, P=0.007, respectively). According to the TNM stage, stratified analysis revealed that its discernibility on RFS was only pronounced in patients with advanced clinical stage (P=0.001). In addition, multivariate Cox regression analysis showed that HMGCS2 expression was an independent risk factor for RFS (P=0.032) instead of OS (P=0.099). The findings of this study provided the evidence that HMGSC2 represented a potential novel prognostic biomarker for ESCC patients.
Keywords: HMGCS2, ESCC, prognosis, down-regulation, IHC, Chinese
Introduction
As the predominant histological type of esophageal cancer, esophageal squamous cell carcinoma (ESCC) is one of the most common malignancies and associated with a poor prognosis.1,2 It should be noted that despite improvements in diagnostic techniques and multimodal therapies, 5-year survival rates of ESCC remain to be 15% on average in many countries.3 Poor survival rates highlight the need to investigate potential therapeutic targets and develop effective treatments. As a complicated and heterogeneous disease, ESCC is driven by accumulative genetic epigenetic alterations in multiple cancer-related genes. Therefore, it is imperative to search reliable biomarkers for clinical diagnosis, prognosis, and specific therapies.
In order to obtain a clear picture of genetic alteration occurring in ESCC, cDNA microarray technology was applied to simultaneously detect changes in the expression of thousands of genes. Consisting of 47,000 transcripts, Affymetrix human genome U133 Plus 2.0 GeneChip was performed in this group, so as to compare gene expression profiles between tumor tissues and matched adjacent non-tumor tissues from 10 ESCC specimens. Numerous differentially expressed genes have been identified, such as CTTN, AZGP1, 3-hydroxy-3-methylglutaryl-CoA synthase 2 (HMGCS2), and SORBS2.4 Due to its significantly downregulated expression in ESCC specimens, HMGCS2 draws the attention. HMGCS2 encodes a mitochondrial enzyme HMGCS2, which is a potential regulatory point converting acetyl-CoA to ketone bodies in the pathway.5,6 Expressed in liver and a number of extrahepatic tissues, like colon, HMGCS2 has been reported to play a crucial role in several cancers, such as rectal cancer, breast cancer, and prostate cancer.7–9 However, the relationship between HMGCS2 expression and ESCC occurrence and development has not been studied by far.
In this article, the expression level of HMGCS2 in ESCC and matched adjacent non-tumor tissues was measured, and its clinicopathological significance and prognostic value in ESCC were further explored.
Materials and methods
Statement of ethics
This research was approved by the Committees for Ethical Review of Research Involving Human Subjects at Zhengzhou University, and all patients provided written informed consent for the use of ESCC and normal tissue samples. Besides, written informed consents for the original human work producing tissue samples were obtained.
Clinical samples
Cohort 1 – a total of 55 paired primary ESCC tumor tissues and their matched adjacent non-tumor tissues were obtained from Linzhou Cancer Hospital (Henan, China) between 2012 and 2013, which were immediately put into vials and stored in liquid nitrogen after surgical removal (for RNA and DNA extraction).
Cohort 2 – along with clinicopathological summaries, a total of 300 paired primary ESCC tumor tissues and their matched non-tumor tissues were obtained from Linzhou Cancer Hospital, between 2001 and 2005, which were formalin fixed and paraffin embedded after surgical resection for tissue microarray (TMA) construction.
The inclusion criteria were listed as follows: 1) histological evidence of ESCC; 2) complete surgical resection (R0); 3) no perioperative chemotherapy and/or radiotherapy treatment; and 4) a complete follow-up period of 60 months.
cDNA synthesis and quantitative real-time PCR
By utilizing TRIzol (Thermo Fisher Scientific, Waltham, MA, USA), total RNA was extracted from frozen ESCC tissues. According to the manufacturer’ instructions, an equal amount of cDNA was synthesized through the application of Advantage RT-for-PCR kit (Clontech Laboratories, Mountain View, CA, USA). In order to analyze the expression level of the corresponding GAPDH and HMGCS2, quantitative real-time polymerase chain reaction (qRT-PCR) was performed by using a SYBR Green PCR Kit (Thermo Fisher Scientific) and ABI7900HT Fast Real-Time PCR system (Thermo Fisher Scientific). Meanwhile, GAPDH served as an internal control for HMGCS2. HMGCS2 and GAPDH primers are listed in Table 1. SDS2.3 software (Thermo Fisher Scientific) was employed to analyze the relative expression levels. By means of the Ct method, the real-time value for each sample was averaged and compared. ΔΔCt(sample) = ΔCt(sample) − ΔCt(calibrator), ΔCt(sample) = Ct(sample) of target gene − Ct(sample) of GAPDH; ΔCt(calibrator) = Ct(calibrator) of target gene − Ct(calibrator) of GAPDH; calibrator was defined as pooled samples from 55 adjacent non-tumor tissues.
 | Table 1 Primer sequences used for qPCR analyses |
ESCC tissue microarray and immunohistochemical staining
TMAs containing 300 pairs of primary ESCC tissue samples and their matched adjacent non-tumor tissues were constructed as previously described.10 TMA blocks with a thickness of 5 μm were sectioned for immunohistochemical (IHC) staining. By employing the standard streptavidin–biotin–peroxidase complex method, IHC staining was performed.10 Briefly, sections were deparaffinized and rehydrated, and the endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 10 minutes. For the sake of antigen retrieval, TMA slides were boiled in 10 mmol/L sodium citrate buffer (pH 6.0) for 15 minutes in a microwave oven. Nonspecific binding was blocked with 5% normal goat serum for 10 minutes. In a humidified chamber, slides were incubated in a 1:50 dilution of anti-HMGCS2 (Abcam, Cambridge, UK) at 4°C overnight. In the next step, the TMA section was sequentially incubated with biotinylated goat anti-rabbit immunoglobulin at a concentration of 1:100 at 37°C for 30 minutes. Using ImPress Universal kit (Vector Laboratories), primary antibody staining was visualized, during which NovaRed (Vector Laboratories, Burlingame, CA, USA) acted as a substrate. Finally, 3,3′-diaminobenzidine (DAB) Substrate Kit (Dako Ltd) was used for color development and then Mayer’s hematoxylin counterstaining. The positive localization in the cells of IHC staining is cytoplasm. The positive expression cells were quantified by ImageJ software. As previously described, an immunoreactivity score system was applied.11 HMGCS2-positive cell percentage was scored as 0, <5%, negative; 1, 5%–25%, sporadic; 2, 25%–50%, focal; 3, >50%, diffuse. In addition, HMGCS2-positive staining intensity was scored as 0, negative; 1, weak; 2, moderate; 3, strong. Both positive cell percentage and cell staining intensity were determined in a double-blinded manner. According to the following formula, the total score was determined (20× microscope field with 10 random fields per subject): staining index = intensity × positive rate. In this article, a staining index <4 was considered as downexpression and a staining index of >4 was regarded as normal expression.
Statistical analysis
SPSS standard version 16.0 was adopted to perform statistical analysis, and paired two-tailed t-test was employed to compare HMGCS2 expression in primary ESCC tumors and their paired non-tumor tissues. Moreover, the statistical significance of correlations between HMGCS2 protein expression and clinicopathologic characteristics was assessed by chi-square test or Fisher’s exact tests. The points for overall survival (OS) and relapse-free survival (RFS) were calculated from the date of surgery to the date that event developed. Survival curves were plotted by the Kaplan–Meier method, and statistical analysis was performed through log-rank test. And then, parameters demonstrating prognostic significance in the univariate test were enrolled into multivariate comparisons with the aid of the Cox-proportional hazards model. A P-value <0.05 was considered to be statistically significant.
Results
HMGCS2 is significantly downregulated in primary ESCCs
In order to investigate HMGCS2 expression in ESCC specimens, the RNA expression level of HMGCS2 was examined in 55 pairs of primary ESCC tumors and their matched non-tumor tissues obtained from Linzhou Cancer Hospital (cohort 1). With the use of qRT-PCR, HMGCS2 was significantly downregulated in ESCC samples (P<0.001), and ~36 of 55 (65.5%) ESCC specimens demonstrated lower HMGCS2 expression compared with their matched non-tumor specimens (Figure 1).
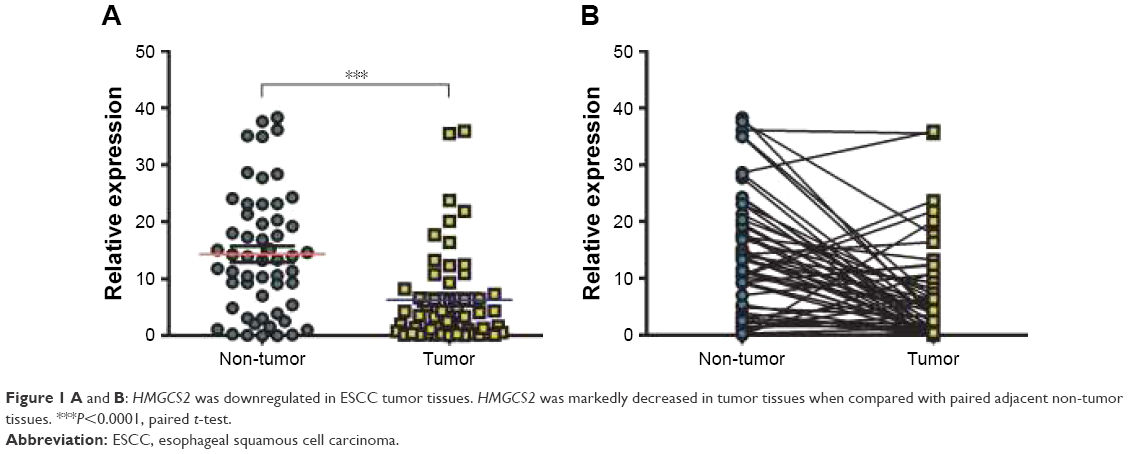 | Figure 1 A and B: HMGCS2 was downregulated in ESCC tumor tissues. HMGCS2 was markedly decreased in tumor tissues when compared with paired adjacent non-tumor tissues. ***P<0.0001, paired t-test. |
A panel of TMAs consisting of 300 matched pairs of primary ESCC and non-tumor specimens collected from Linzhou Cancer Hospital (cohort 2) were constructed, so as to confirm the findings and investigate the clinical implication of HMGCS2 during ESCC development. In 290 matched pairs of ESCC and non-tumor specimens, informative results of IHC staining were observed. Non-informative samples were not used as valid data, including lost samples, inappropriately stained samples, and samples with very few cells. By performing IHC staining, differential cytoplasmic expression of HMGCS2 was observed between primary ESCC tumor and their matched non-tumor tissues (Figure 2). Since the staining index of HMGCS2 was ≥4 in non-tumor tissues, a staining index of >4 was counted as normal. In addition, a detailed analysis of IHC data revealed that HMGCS2 was downregulated in 37.6% (109/290) of informative ESCC tumor tissues (P<0.001). In terms of HMGCS2 protein expression, there was obvious difference between ESCC and their matched non-tumor specimens.
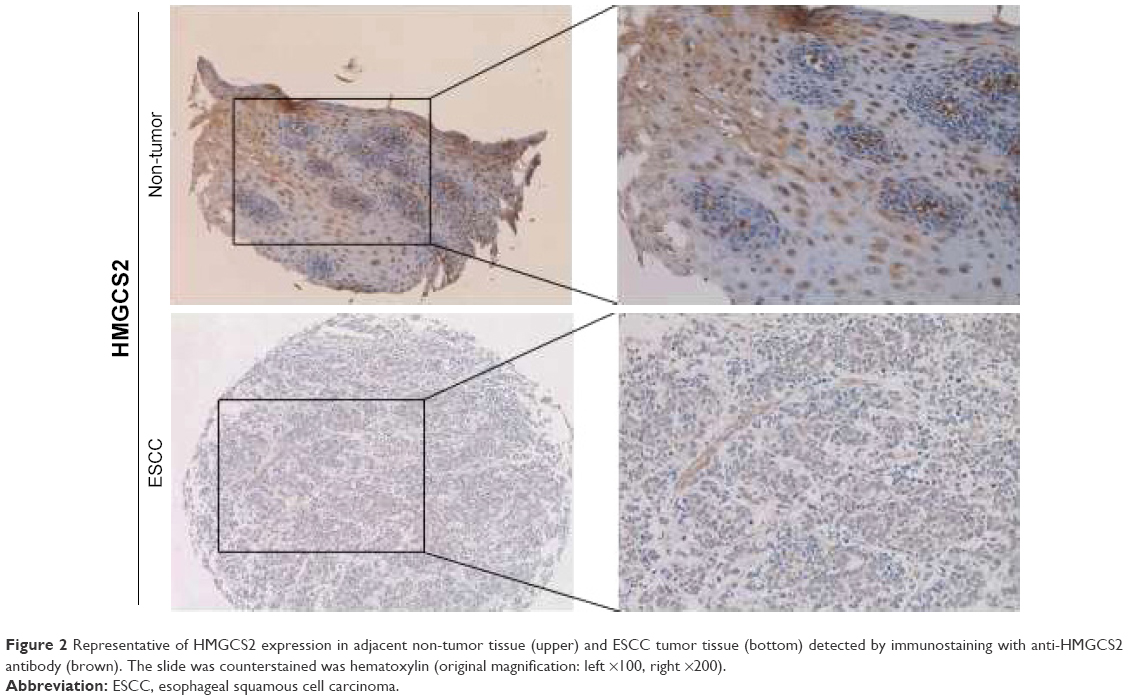 | Figure 2 Representative of HMGCS2 expression in adjacent non-tumor tissue (upper) and ESCC tumor tissue (bottom) detected by immunostaining with anti-HMGCS2 antibody (brown). The slide was counterstained was hematoxylin (original magnification: left ×100, right ×200). |
Correlation of HMGCS21 protein expression with clinicopathological variables
Subsequently, the correlation between HMGCS2 protein expression in primary ESCC and clinicopathological features of ESCC patients from Linzhou Cancer Hospital (cohort 2) are summarized in Table 2. Down-HMGCS2 expression was significantly associated with poor tumor cell differentiation (P=0.002), pT status (P=0.006), and advanced TNM stage (P=0.010). Moreover, no correlation was observed between HMGCS2 expression and other clinicopathological characteristics, such as patient’s age (P=0.275), gender (P=0.627), tumor location (P=0.306), pathologic N status (P=0.542), general classification (P=0.664), smoking status (P=0.332), and drinking status (P=1.000).
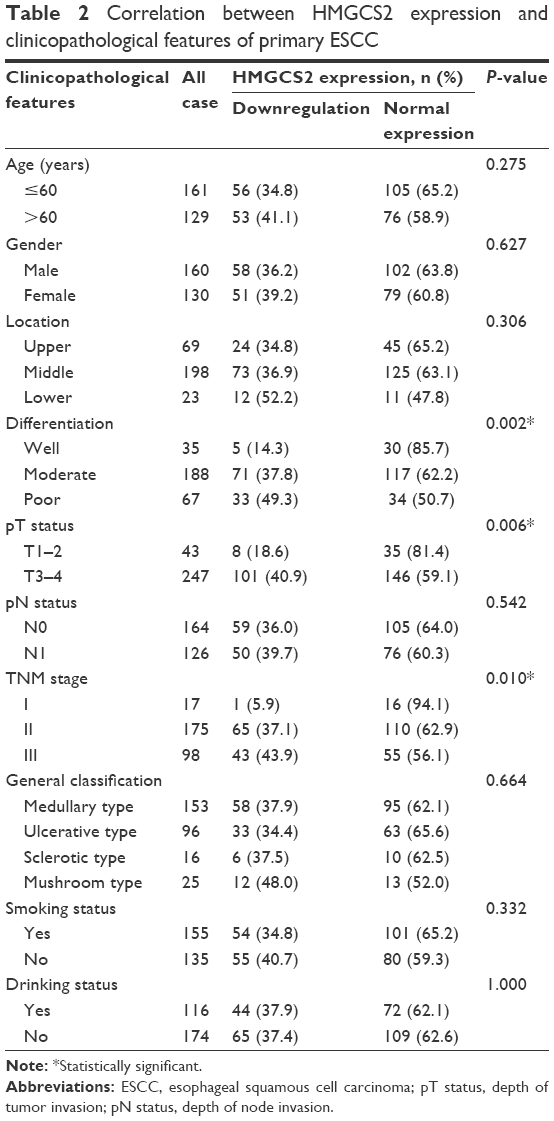 | Table 2 Correlation between HMGCS2 expression and clinicopathological features of primary ESCC |
Association between HMGCS2 expression and patient survival
Univariate analysis demonstrated that tumor cell differentiation, pathologic T and N status, TNM stage, and HMGCS2 expression were prognostic factors for OS, while pathologic T status, TNM stage, and HMGCS2 expression were prognostic factors for RFS (Table 3). In the down-HMGCS2 expression group, the median 5-year OS (19 vs 24 months; Figure 3A) and median RFS (13 vs 17 months; Figure 3B) were significantly lower than those in normal HMGCS2 expression group. Correlated with a number of clinical indexes (such as tumor cell differentiation, pathologic T or N status, and HMGCS2 expression), the TNM stage was not introduced into multivariate Cox-proportional hazard model, so as to avoid potential bias. Through the application of multivariate analysis, it was found that tumor cell differentiation and pathologic N status were independent prognostic predictors for OS, while HMGCS2 expression and pathologic T status were independent prognostic predictors for RFS (Table 4). Furthermore, no significant difference in RFS was observed in patients with TNM stages I–II (P=0.059; Figure 3C) in a stratified survival analysis according to the TNM stage. However, HMGCS2 expression could differentiate the prognosis of patients among those with TNM stage III (P=0.001; Figure 3D).
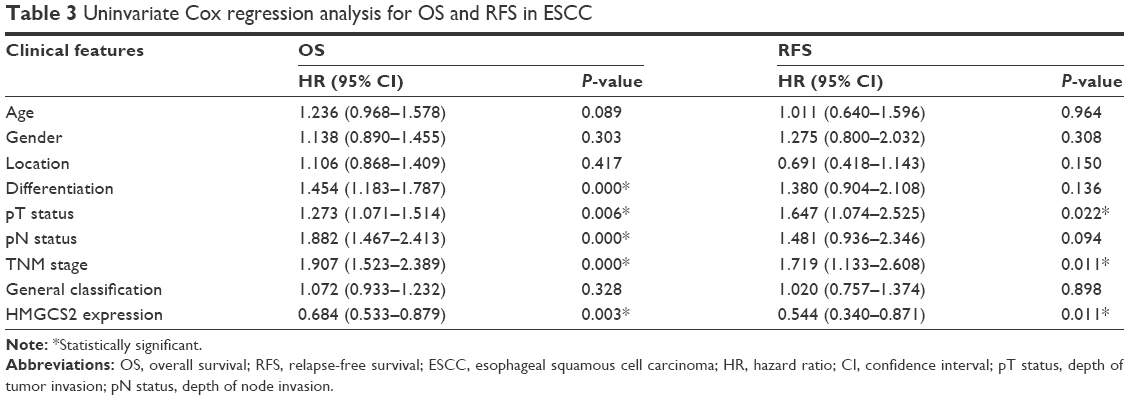 | Table 3 Uninvariate Cox regression analysis for OS and RFS in ESCC |
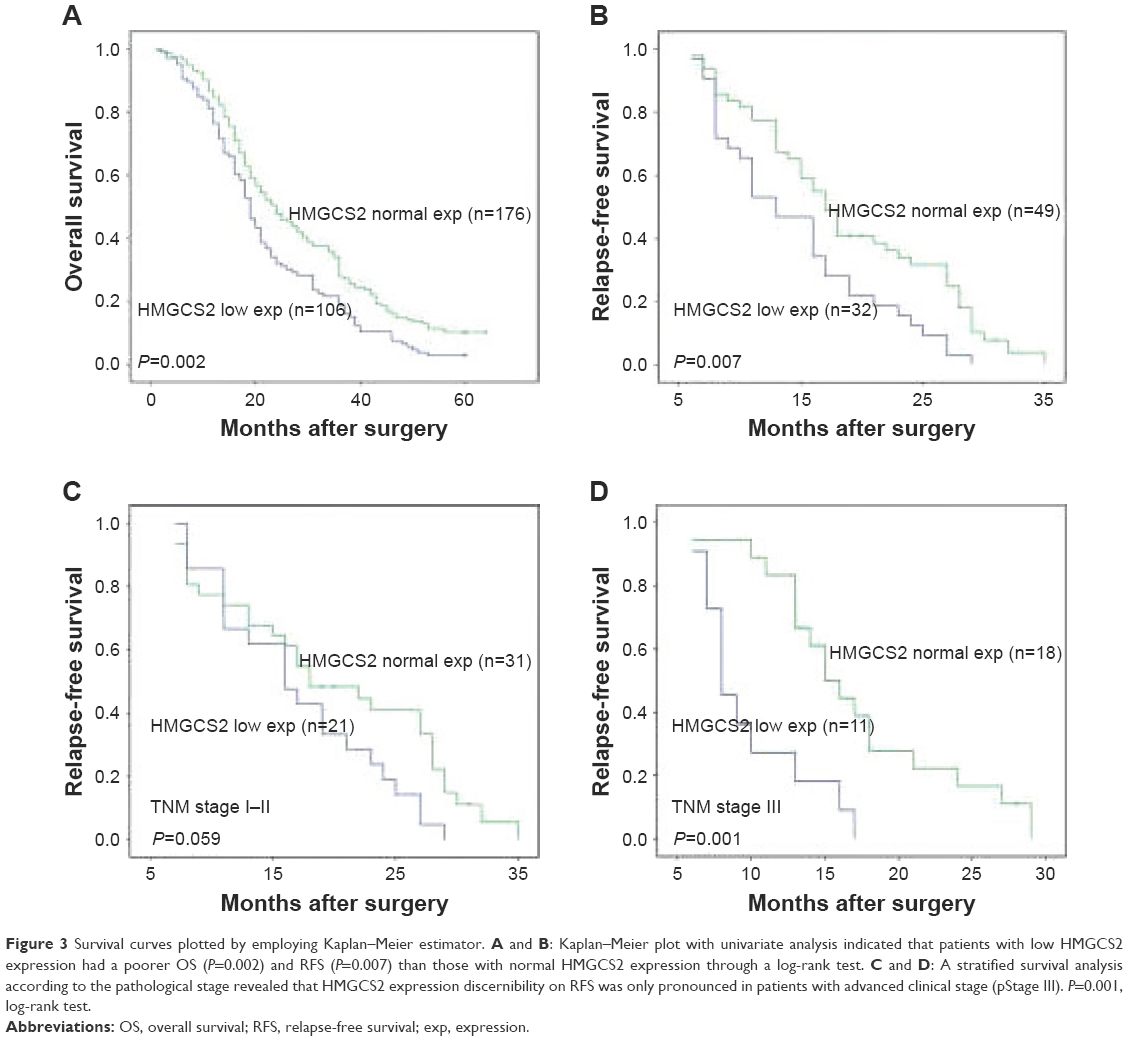 | Figure 3 Survival curves plotted by employing Kaplan–Meier estimator. A and B: Kaplan–Meier plot with univariate analysis indicated that patients with low HMGCS2 expression had a poorer OS (P=0.002) and RFS (P=0.007) than those with normal HMGCS2 expression through a log-rank test. C and D: A stratified survival analysis according to the pathological stage revealed that HMGCS2 expression discernibility on RFS was only pronounced in patients with advanced clinical stage (pStage III). P=0.001, log-rank test. |
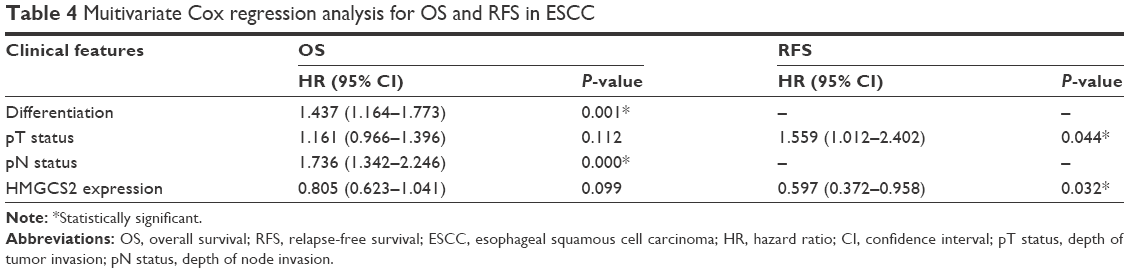 | Table 4 Muitivariate Cox regression analysis for OS and RFS in ESCC |
Discussion
Basic characteristics of cancer are closely related to major alternations in tumor cell metabolism, which is enhanced lipid biosynthesis and glycolysis to satisfy their high energy demand for conducting integral cellular processes.12,13 De novo fatty acid biosynthesis is essential for cancer cells to synthesize new cellular membranes, which are crucial in multiple membrane-based biochemical and biological processes, such as signaling cell growth receptors and protein assembly.7 Moreover, lipid metabolism is necessary for a number of transformed phenotypes in cancer cells, including cell proliferation, invasion, dissemination, and survival under oxidative or energy stress.14 By producing ketone bodies via decomposing acetyl-CoA, the ketogenic pathway is an alternative energy-producing pathway to provide energy for cancer cells in the form of ATP.15
As a mitochondrial enzyme involved in ketogenic pathway, HMGCS2 catalyzes the first reaction of ketogenesis producing HMG-CoA, so as to provide lipid-derived energy during fasting. HMGCS2 expression in humans has been reported in liver, colon, pancreas, heart, and skeletal muscle,16 and its expression is usually detected in differentiated cells.8,17 Proliferating cells do not express HMGCS2 activity.18,19 On the contrary, its expression is the characteristic of differentiated cells of colon.20 To the authors’ knowledge, no data concerning HMGCS2 expression in ESCC are available. The authors’ previous study showed that HGMCS2 was 2.2-fold downregulated in ESCC cDNA microarray, and this significant dysregulation prompted us to investigate its role in ESCC. In this study, HMGCS2 expression has been shown to be downregulated in ESCC tissues on mRNA and protein levels for the first time, when compared with their matched non-tumor tissues. It was found that ~65.5% of ESCC patients showed downregulated HMGCS2 expression on mRNA level in their tumor specimens and ~37.6% showed downregulated expression on protein level. Through chromatin immunoprecipitation experiments, HMGCS2 has been identified to be downregulated by c-Myc via a Miz-1-dependent mechanism in colon cancer.17 Since HMGCS2 is upregulated by the expression of a dominant-negative form of TCF-4 in the colon cell line, HMGCS2 may be under the control of the Wnt/APC/β-catenin signal pathway.21,22 These results give rise to the hypothesis that HMGCS2 may induce ESCC cells differentiation and/or apoptosis. However, in several other studies, high HMGCS2 expression was found in a number of tumors, such as rectal cancer and prostate cancer,7–9 which suggested that HMGCS2 could be regarded as a negative prognosis factor. Based on these studies, HMGCS2 seems to exhibit very contradicting roles depending on cellular context. Therefore, further studies in vitro and in vivo are needed to clarify the precise mechanism of HMGCS2 involved in ESCC progression.
In this study, the correlation analysis between the gene and clinicopathologic characteristics found that among patients with poor differentiation, tumor invasion, lymph node metastasis, and advanced TNM stage, downexpression level of HMGCS2 protein was observed more frequently. In addition, this study demonstrated that HMGCS2 downexpression was significantly correlated with worse survival, including OS and RFS on univariate level. And then, HMGCS2 downexpression could be considered as an independent negative prognostic factor for poor RFS in the multivariate analysis. The 5-year OS and RFS of patients with downregulated HMGCS2 expression were markedly shorter than that of those with normal expression (19 vs 24 months, 13 vs 17 months, respectively). Moreover, the prognostic value of HMGCS2 expression level was analyzed in selected patient subgroups, and it was found that in the subgroup of patients with advanced clinical stage, downregulated HMGCS2 expression was associated with a poor RFS. When compared with surgery alone, adjuvant chemotherapy or neoadjuvant chemoradiotherapy was proven to improve the clinical outcome of patients with locally advanced ESCC.23–27 Therefore, perioperative chemotherapy may be recommended for the subset of patients with down-HMGCS2 expression to improve their outcome.
Regulation of HMGCS2 expression mainly occurs on transcriptional level. Elements of HMGCS2 proximal promoter may account for its regulation by hormones, such as insulin,28 and metabolites, such as long-chain fatty acids or butyrate.20,29 Insulin-mediated HMGCS2 regulation involves forkhead transcription factors.30 Through some signals of inducing cell proliferation, HMGCS2 seems to be downregulated, such as hormonal and metabolic signals. Although HMGCS2 is a direct target of c-Myc, its expression is not related to c-Myc levels, because only some tumors were Myc dependent, such as colon rectum tumors. Therefore, HMGCS2 and c-Myc expression levels may depend on additional factors related to nutritional status or intestinal germ flora of patients.17 Associated with the ketogenic capacity, HMGCS2 expression is regulated by ketone body acetoacetate by increasing to inhibit endothelial cell growth.31,32 In another study, it is shown that HMGCS2 acts as a tumor-suppressor gene, because p53 upregulates fatty acid oxidation induced by glucose starvation. As a target gene of p53, it may activate β-oxidation to protect the cell from Warburg effect.33,34
In conclusion, the result of the present study demonstrates that downregulated HMGCS2 expression in ESCC tumor specimens indicates aggressive tumor behaviors and predicts a worse clinical outcome for the first time. These findings suggest that HMGCS2 may serve as a potential target for diagnosis and therapy of ESCC patients, especially patients with advanced clinical stage.
Acknowledgments
This work was supported by grants from the Key Science and Technology Research Foundation of Henan Province (no 152102410087 to Hong Tang; no 162102310327 to Yufeng Wu), PhD Research Foundation of Henan Cancer Hospital (no 201511004 to Yufeng Wu).
Disclosure
The authors report no conflicts of interest in this work.
References
Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ. Cancer statistics, 2009. CA Cancer J Clin. 2009;59(4):225–249. | ||
Shahbaz Sarwar CM, Luketich JD, Landreneau RJ, Abbas G. Esophageal cancer: an update. Int J Surg. 2010;8(6):417–422. | ||
Portale G, Hagen JA, Peters JH, et al. Modern 5-year survival of resectable esophageal adenocarcinoma: single institution experience with 263 patients. J Am Coll Surg. 2006;202(4):588–596; discussion 596–598. | ||
Lu P, Qiao J, He W, et al. Genome-wide gene expression profile analyses identify CTTN as a potential prognostic marker in esophageal cancer. PLoS One. 2014;9(2):e88918. | ||
Dashti N, Ontko JA. Rate-limiting function of 3-hydroxy-3-methylglutaryl-coenzyme A synthase in ketogenesis. Biochem Med. 1979;22(3):365–374. | ||
Hegardt FG. Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: a control enzyme in ketogenesis. Biochem J. 1999;338(Pt 3):569–582. | ||
Lee YE, He HL, Shiue YL, et al. The prognostic impact of lipid biosynthesis-associated markers, HSD17B2 and HMGCS2, in rectal cancer treated with neoadjuvant concurrent chemoradiotherapy. Tumour Biol. 2015;36(10):7675–7683. | ||
Gromov P, Espinoza JA, Talman ML, et al. FABP7 and HMGCS2 are novel protein markers for apocrine differentiation categorizing apocrine carcinoma of the breast. PLoS One. 2014;9(11):e112024. | ||
Saraon P, Cretu D, Musrap N, et al. Quantitative proteomics reveals that enzymes of the ketogenic pathway are associated with prostate cancer progression. Mol Cell Proteomics. 2013;12(6):1589–1601. | ||
Xie D, Sham JS, Zeng WF, et al. Heterogeneous expression and association of beta-catenin, p16 and c-myc in multistage colorectal tumorigenesis and progression detected by tissue microarray. Int J Cancer. 2003;107(6):896–902. | ||
Brown RS, Wahl RL. Overexpression of Glut-1 glucose transporter in human breast cancer. An immunohistochemical study. Cancer. 1993;72(10):2979–2985. | ||
Kroemer G, Pouyssegur J. Tumor cell metabolism: cancer’s Achilles’ heel. Cancer Cell. 2008;13(6):472–482. | ||
Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324(5930):1029–1033. | ||
Santos CR, Schulze A. Lipid metabolism in cancer. FEBS J. 2012;279(15):2610–2623. | ||
Nakamura MT, Cheon Y, Li Y, Nara TY. Mechanisms of regulation of gene expression by fatty acids. Lipids. 2004;39(11):1077–1083. | ||
Mascaro C, Buesa C, Ortiz JA, Haro D, Hegardt FG. Molecular cloning and tissue expression of human mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase. Arch Biochem Biophys. 1995;317(2):385–390. | ||
Camarero N, Mascaro C, Mayordomo C, Vilardell F, Haro D, Marrero PF. Ketogenic HMGCS2 is a c-Myc target gene expressed in differentiated cells of human colonic epithelium and down-regulated in colon cancer. Mol Cancer Res. 2006;4(9):645–653. | ||
Le May C, Pineau T, Bigot K, Kohl C, Girard J, Pegorier JP. Reduced hepatic fatty acid oxidation in fasting PPARalpha null mice is due to impaired mitochondrial hydroxymethylglutaryl-CoA synthase gene expression. FEBS Lett. 2000;475(3):163–166. | ||
Prip-Buus C, Bouthillier-Voisin AC, Kohl C, Demaugre F, Girard J, Pegorier JP. Evidence for an impaired long-chain fatty acid oxidation and ketogenesis in Fao hepatoma cells. Eur J Biochem. 1992;209(1):291–298. | ||
Camarero N, Nadal A, Barrero MJ, Haro D, Marrero PF. Histone deacetylase inhibitors stimulate mitochondrial HMG-CoA synthase gene expression via a promoter proximal Sp1 site. Nucleic Acids Res. 2003;31(6):1693–1703. | ||
van de Wetering M, Sancho E, Verweij C, et al. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell. 2002;111(2):241–250. | ||
Wanzel M, Herold S, Eilers M. Transcriptional repression by Myc. Trends Cell Biol. 2003;13(3):146–150. | ||
Lee J, Lee KE, Im YH, et al. Adjuvant chemotherapy with 5-fluorouracil and cisplatin in lymph node-positive thoracic esophageal squamous cell carcinoma. Ann Thorac Surg. 2005;80(4):1170–1175. | ||
Ando N, Iizuka T, Ide H, et al. Surgery plus chemotherapy compared with surgery alone for localized squamous cell carcinoma of the thoracic esophagus: a Japan Clinical Oncology Group Study – JCOG9204. J Clin Oncol. 2003;21(24):4592–4596. | ||
Tepper J, Krasna MJ, Niedzwiecki D, et al. Phase III trial of trimodality therapy with cisplatin, fluorouracil, radiotherapy, and surgery compared with surgery alone for esophageal cancer: CALGB 9781. J Clin Oncol. 2008;26(7):1086–1092. | ||
Sjoquist KM, Burmeister BH, Smithers BM, et al. Survival after neoadjuvant chemotherapy or chemoradiotherapy for resectable oesophageal carcinoma: an updated meta-analysis. Lancet Oncol. 2011;12(7):681–692. | ||
van Hagen P, Hulshof MC, van Lanschot JJ, et al. Preoperative chemoradiotherapy for esophageal or junctional cancer. New Engl J Med. 2012;366(22):2074–2084. | ||
Gil-Gomez G, Ayte J, Hegardt FG. The rat mitochondrial 3-hydroxy-3-methylglutaryl-coenzyme-A-synthase gene contains elements that mediate its multihormonal regulation and tissue specificity. Eur J Biochem. 1993;213(2):773–779. | ||
Rodriguez JC, Gil-Gomez G, Hegardt FG, Haro D. Peroxisome proliferator-activated receptor mediates induction of the mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase gene by fatty acids. J Biol Chem. 1994;269(29):18767–18772. | ||
Nadal A, Marrero PF, Haro D. Down-regulation of the mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase gene by insulin: the role of the forkhead transcription factor FKHRL1. Biochem J. 2002;366(Pt 1):289–297. | ||
Cherbuy C, Darcy-Vrillon B, Morel MT, Pegorier JP, Duee PH. Effect of germfree state on the capacities of isolated rat colonocytes to metabolize n-butyrate, glucose, and glutamine. Gastroenterology. 1995;109(6):1890–1899. | ||
Jain SK, Kannan K, Lim G. Ketosis (acetoacetate) can generate oxygen radicals and cause increased lipid peroxidation and growth inhibition in human endothelial cells. Free Radic Biol Med. 1998;25(9):1083–1088. | ||
Ide T, Brown-Endres L, Chu K, et al. GAMT, a p53-inducible modulator of apoptosis, is critical for the adaptive response to nutrient stress. Mol Cell. 2009;36(3):379–392. | ||
Levine AJ, Puzio-Kuter AM. The control of the metabolic switch in cancers by oncogenes and tumor suppressor genes. Science. 2010;330(6009):1340–1344. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.