Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 21
PPARA Gene Polymorphisms and Their Association with COPD in Tibetan and Han Populations from Gansu, China
Authors Gao R, Zhang M, Zhang Y, Wang Y, Yang Z ![]() , Lin A
, Lin A ![]() , Wu Y, Wang X
, Wu Y, Wang X
Received 19 December 2025
Accepted for publication 9 April 2026
Published 16 May 2026 Volume 2026:21 590361
DOI https://doi.org/10.2147/COPD.S590361
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jill Ohar
Renjie Gao,1,* Miao Zhang,1,* Yangjie Zhang,1 Yunchao Wang,2 Zhen Yang,1 Ao Lin,3 Yanlin Wu,4 Xinhua Wang1
1School of Public Health, Gansu University of Chinese Medicine, Lanzhou, Gansu, People’s Republic of China; 2School of Basic Medicine, Gansu University of Chinese Medicine, Lanzhou, Gansu, People’s Republic of China; 3Department of Thoracic Surgery, The First Affiliated Hospital of Guangzhou Medical University; School of Public Health, Guangzhou Medical University, Guangzhou, Guangdong, People’s Republic of China; 4Department of Pulmonary Disease, Affiliated Hospital of Gansu University of Chinese Medicine, Lanzhou, Gansu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yanlin Wu, Department of Pulmonary Disease, Affiliated Hospital of Gansu University of Chinese Medicine, Lanzhou, Gansu, 730000, People’s Republic of China, Email [email protected] Xinhua Wang, School of Public Health, Gansu University of Chinese Medicine, Lanzhou, Gansu, 730000, People’s Republic of China, Email [email protected]
Background: Chronic Obstructive Pulmonary Disease (COPD) is influenced by both genetic and environmental factors, with chronic hypoxia playing a pivotal role in its pathogenesis. Peroxisome proliferator-activated receptor alpha (PPARA), a core regulator of the hypoxia pathway, exhibits genetic differentiation in high-altitude populations. This study aimed to investigate the association between PPARA gene polymorphisms and COPD susceptibility in Tibetan and Han populations in Gansu Province.
Methods: A case-control study design was employed. A total of 1276 participants (399 Tibetans and 877 Han Chinese) were recruited from Gansu province. Using Haploview software, four tag SNPs of the PPARA gene (rs135538, rs135539, rs135549, rs4253758) were selected. Genotyping was performed using the iMLDR (Improved Multiple Ligase Detection Reaction) multiplex SNP genotyping technique.
Results: In the Tibetan population, the rs135539A>C polymorphism under a dominant model was associated with an increased risk of COPD (CC+CA vs. AA: OR = 1.779, 95% CI: 1.080– 2.903, P=0.024). In the Han population, rs135549T>C under a dominant model was associated with a reduced COPD risk (CC+CT vs. TT: OR = 0.719, 95% CI: 0.547– 0.944, P = 0.017). The rs4253758T>C SNP exhibited a dual effect in Han Chinese: under an additive model, both TC (OR = 1.722, 95% CI: 1.504– 2.813, P = 0.030) and CC (OR = 1.778, 95% CI: 1.083– 2.922, P = 0.023) genotypes increased risk compared to the TT genotype. However, under a recessive model, the CC genotype was protective compared to TT+TC (OR = 0.572, 95% CI: 0.356– 0.918, P = 0.021). No significant associations were found for other SNPs.
Conclusion: PPARA gene polymorphisms are associated with COPD susceptibility in a population-specific manner among Tibetan and Han populations in Gansu. The rs135539A>C variant increases COPD risk in Tibetans, whereas rs135549T>C and rs4253758T>C exert protective and dual effects, respectively, in the Han population.
Keywords: chronic obstructive pulmonary disease, PPARA, single nucleotide polymorphism, ethnicity
Introduction
Chronic Obstructive Pulmonary Disease (COPD) is a common respiratory disorder characterized by persistent airflow limitation and an abnormal chronic inflammatory response of the airways and lungs to noxious particles or gases.1,2 Clinically, it presents with chronic cough, sputum production, and progressive dyspnea, following an irreversible course that imposes a substantial burden on patients, families, and society.3 According to the World Health Organization, the global prevalence and mortality of COPD remain high. In China alone, the prevalence among individuals aged 40 and above has reached 13.7%, showing a significant increase over the past decade.4 Historical data indicate that approximately 400 million people were affected globally by 2011,5 and projections suggest this number will exceed 600 million by 2050.6 Mortality has also risen, from 2.4 million in 1990 to 3.23 million in 2019—a 34% increase7—with annual deaths expected to reach 4.3 million by 2040.8 COPD is now the third leading cause of death worldwide.9
COPD pathogenesis results from a complex interplay between environmental exposures and individual genetic susceptibility.10 Tobacco smoke, including active smoking and secondhand exposure, is the primary environmental risk factor.11 Other key drivers include prolonged exposure to indoor and outdoor air pollutants such as biomass fuel, fumes, particulate matter (PM2.5), and ozone.12 Occupational exposure to organic/inorganic dust, chemical irritants, and vapors is another significant independent risk factor.13 Host factors, besides rare genetic defects like alpha-1 antitrypsin deficiency, include impaired lung development in early life, frequent childhood respiratory infections, and a history of asthma, all of which reduce lung function reserve and increase susceptibility to COPD in adulthood.14 Family and genome-wide association studies (GWAS) have identified multiple genetic loci associated with COPD susceptibility, confirming its polygenic nature.15,16 Among candidate genes, peroxisome proliferator-activated receptor alpha (PPARA) has recently attracted considerable attention.
Located on human chromosome 22q13.31, PPARA encodes the PPARα protein, a member of the nuclear receptor transcription factor superfamily. PPARα plays a central role in regulating lipid metabolism, energy homeostasis, inflammation, and oxidative stress.17,18 Notably, PPARA is also recognized as a key gene for high-altitude adaptation.19 It is closely linked to the hypoxia-inducible factor (HIF) oxygen-sensing pathway. Under normoxia, HIF-1α is degraded by prolyl hydroxylases (PHDs). During hypoxia, PHD activity is inhibited, leading to the accumulation of HIF-1α and HIF-2α, which translocate to the nucleus and activate the transcription of hypoxic adaptive genes, including PPARA.20,21 PPARA cooperates with HIF-2α and PHDs to regulate erythropoiesis, angiogenesis, and cellular energy metabolism, thereby enhancing oxygen utilization and survival under hypoxic conditions.20
The pathophysiological process of COPD is closely associated with chronic hypoxia. Airway obstruction and parenchymal destruction often lead to localized or systemic oxygen deficiency, which may persistently activate the HIF pathway and disrupt normal PPARA function. Studies show that PPARα activation can suppress key pro-inflammatory signaling pathways such as NF-κB, thereby attenuating pulmonary inflammation.22 It also enhances mitochondrial function via regulators like PGC-1α, countering oxidative stress-induced cellular damage.23,24 Furthermore, as a master regulator of fatty acid oxidation, dysfunctional PPARα may lead to abnormal lipid metabolism in lung tissue, exacerbating cellular injury and death.25,26 Therefore, PPARA polymorphisms may influence its expression or activity, altering an individual’s capacity for inflammation control, oxidative stress resistance, and energy metabolism, ultimately affecting COPD risk.
Tibetans are one of the oldest high-altitude populations in the world, having inhabited the Qinghai-Tibet Plateau for thousands of generations. This prolonged exposure to chronic hypobaric hypoxia has driven unique genetic adaptations, including variants in genes involved in the hypoxia-inducible factor (HIF) pathway, such as PPARA.20,21 In contrast, the Han Chinese population predominantly resides at lower altitudes and has not experienced such sustained hypoxic selective pressure. These distinct environmental exposures and evolutionary histories provide a natural experiment to investigate gene–environment interactions in the pathogenesis of COPD. Specifically, comparing Tibetan and Han populations allows us to assess whether PPARA polymorphisms confer differential susceptibility to COPD under contrasting hypoxic conditions, thereby offering insights into the role of hypoxia adaptation genes in COPD development.
Based on this background, we hypothesize that PPARA single nucleotide polymorphisms (SNPs) are associated with COPD susceptibility in Tibetan and Han populations. To test this hypothesis, we employed a case-control design and used the iMLDR high-throughput genotyping technique to systematically analyze the distribution of four selected PPARA tag SNPs (rs135538, rs135539, rs135549, rs4253758) in Tibetan and Han populations from Gansu. We further investigated the correlation between these genetic variants and COPD risk, aiming to provide novel molecular evidence for the ethnic-specific genetic mechanisms of COPD and a theoretical basis for early screening and personalized prevention in this region.
Methods
Sample Size Estimation and Study Population
Sample Size Estimation
This study is based on the hypothesis that PPARA gene polymorphisms show no statistically significant association with COPD in Tibetan and Han populations. To test this hypothesis, sample size estimation was performed using PASS 11.0 statistical software. Estimation parameters are as follows: significance level α=0.05 (two-tailed test), statistical power (1- β)=0.8. Due to limited research on the association between PPARA and COPD, literature review18 was consulted. Referencing the exposure rate between PPARA SNPs and acute high-altitude pulmonary edema, an estimated value of 0.25 was used, with a minimum estimated OR of 1. The following formula was applied:
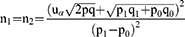
Calculations indicate that 442 cases and 442 controls are required for each of the Tibetan and Han ethnic groups, totaling 1768 cases. However, due to the challenges in conducting the baseline survey, the study ultimately enrolled only 1276 subjects.
Study Population
This case-control study aimed to investigate the association between PPARA gene polymorphisms and susceptibility to COPD in Tibetan and Han populations in Gansu Province. Han subjects were consecutively recruited from COPD patients and concurrent healthy examinees at the Department of Pulmonary Diseases, Affiliated Hospital of Gansu University of Chinese Medicine, Lanzhou City, Gansu Province. Tibetan subjects were selected from a natural population cohort in Tianzhu County, Wuwei City, Gansu Province. A total of 1276 subjects were enrolled, comprising 164 Tibetan COPD cases and 235 controls, alongside 439 Han COPD cases and 438 Han controls.
The study was conducted per the Declaration of Helsinki and approved by the Institutional Review Board of Guangzhou Medical University Center (Approval No.: 202405010). Written informed consent was obtained from all participants.
COPD Diagnosis and Pulmonary Function Testing
According to the 2023 Global Initiative for Chronic Obstructive Pulmonary Disease (GOLD), COPD is considered diagnosed when risk factors associated with dyspnea, cough, sputum production, and smoking are present, and the ratio of forced expiratory volume in one second (FEV1) to forced vital capacity (FVC) measured 30 minutes after inhaling 400 µg of salbutamol is less than 70%. Lung function testing was performed using the EasyOne spirometer (NDD Medizintechnik AG, Switzerland) according to the manufacturer’s instructions.27
Exclusion Criteria: (1) Presence of contraindications for pulmonary function testing: history of surgery, cardiovascular disease, or severe concomitant illnesses; (2) Respiratory diseases such as asthma, bronchial asthma, chronic bronchitis, or emphysema; (3) Inability to complete the survey questionnaire.
SNP Selection and Genotyping
Potential risk SNPs were identified from the dbSNP database (http://www.ncbi.nlm.nih.gov/SNP) using the following criteria: located within PPARA or its 2000 bp flanking regions, minor allele frequency (MAF) > 0.05 in the Chinese population, and low linkage disequilibrium (LD, r2 < 0.8). Four SNPs were selected for analysis: rs135538C>G, rs135539A>C, rs135549T>C, and rs4253758T>C (Table 1 and Figure 1). Genomic DNA was extracted from peripheral blood samples using the Tiangen Blood Genomic DNA Extraction Kit (DP349). Genotyping was performed using the TaqMan-MGB probe method. To ensure reliability, a negative control was included on each plate, and 10% of samples were randomly selected for repeat genotyping (Kappa ≥ 0.80).
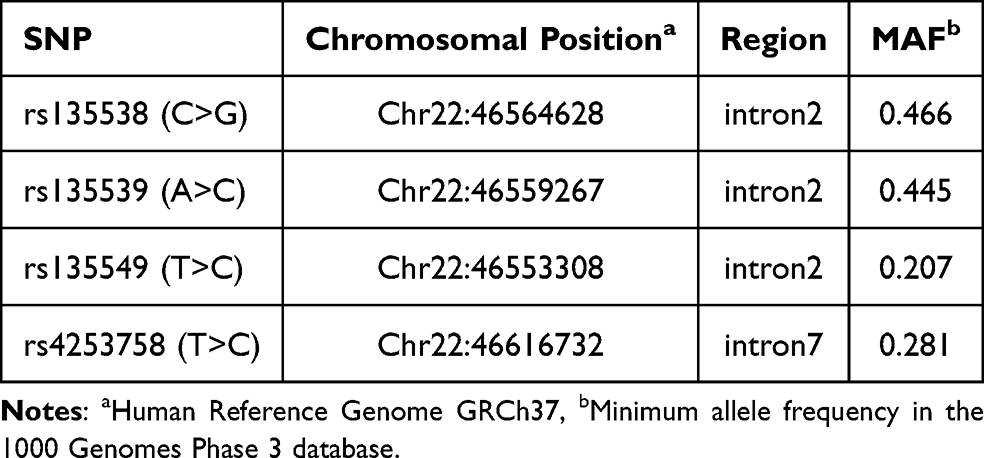 |
Table 1 Selected tag SNPs of the PPARA Gene |
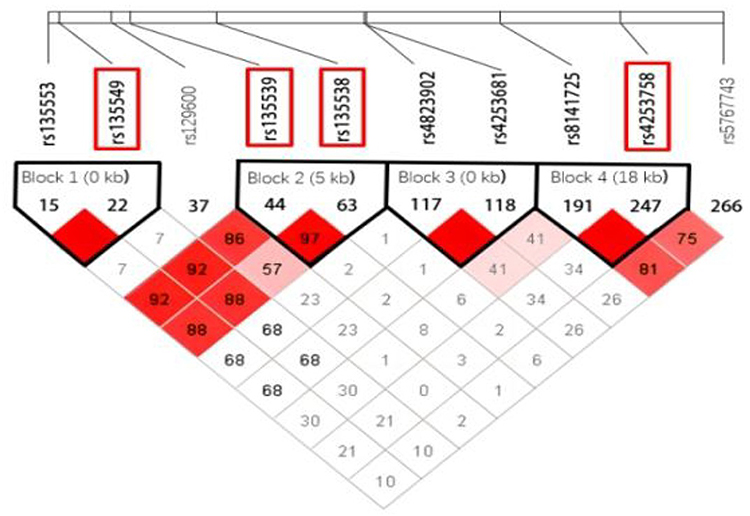 |
Figure 1 Linkage disequilibrium plot of the PPARA gene. |
Quantitative Real-Time PCR
PCR reaction mixtures were prepared using the Takara Premix Ex Taq™ (Probe qPCR) kit, vortexed for 15 seconds, and briefly centrifuged. A standard DNA concentration of 30 ng/μL was used. In each well of a 384/96-well plate, 18.5 μL of PCR reaction mix was combined with 1.5 μL of DNA template. The plate was sealed, briefly centrifuged at 2000 rpm for 2 minutes, and placed in a 7900 HT or StepOnePlus Real-Time PCR System. The thermal cycling conditions were as follows: pre-read → pre-denaturation at 95°C for 10 min → 40 cycles of denaturation at 95°C for 15s and annealing/extension at 60°C for 1 min → final extension at 60°C for 1 min → post-read.
Statistical Analysis
Baseline characteristics between cases and controls were compared using Pearson’s chi-square test. The association between genetic polymorphisms and COPD susceptibility was assessed using multivariate logistic regression, adjusting for sex, age, smoking, alcohol consumption, education level, and hypertension. The Akaike Information Criterion (AIC) was used to select the best-fitting genetic model (lowest AIC). Stratified analyses and interaction tests were performed by sex, age, BMI, education, marital status, smoking, and alcohol use. Heterogeneity across subgroups was evaluated using the Breslow-Day test, and pooled effect sizes were calculated using the Cochran-Mantel-Haenszel (CMH) test. All analyses were conducted using SPSS 27.0, with a two-sided P < 0.05 considered statistically significant.
Results
Demographic Characteristics
As shown in Table 2, significant differences were observed between Tibetan and Han groups in the distribution of sex, age, and smoking status (P < 0.05) (Supplementary Tables 1 and 2).
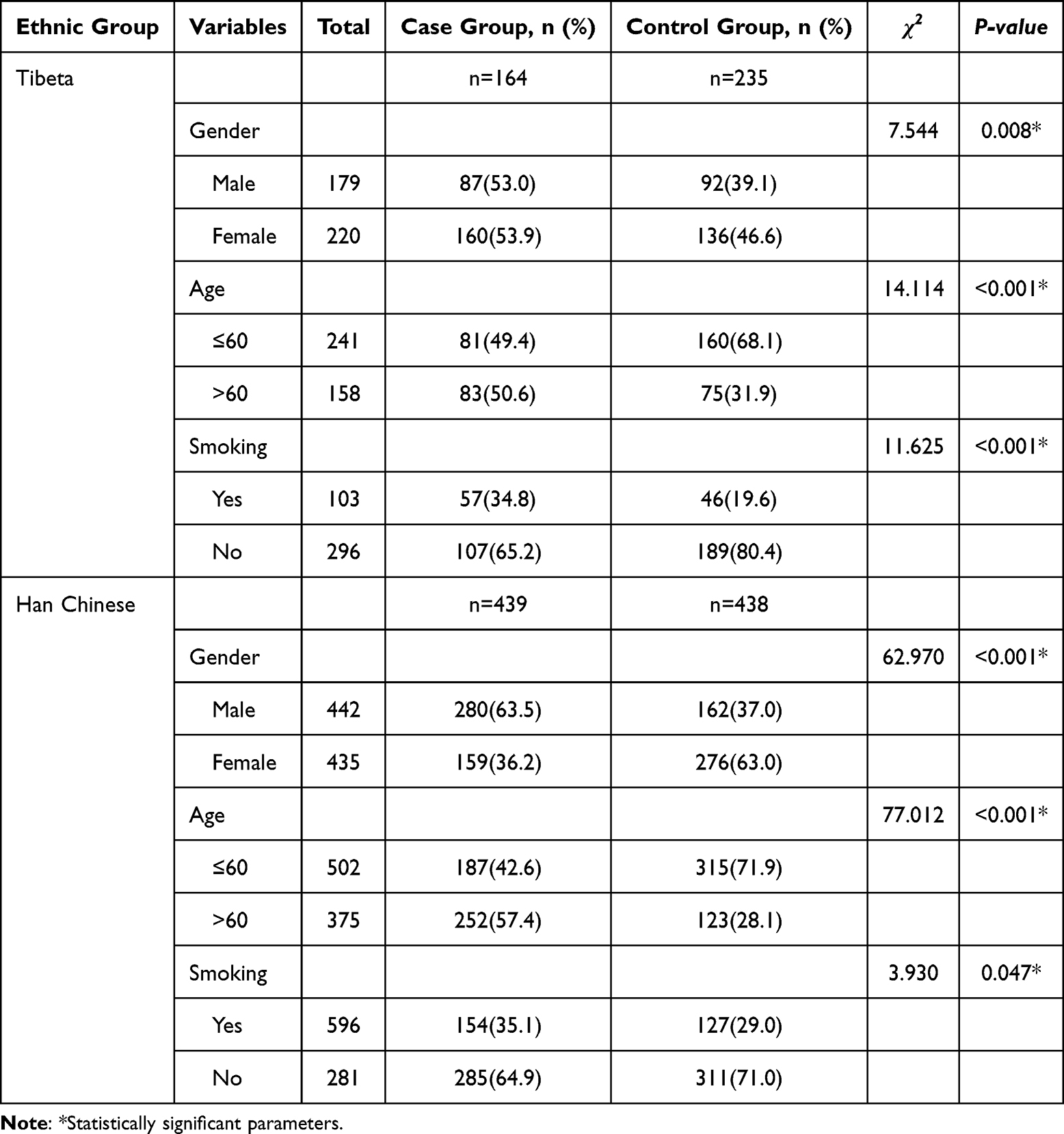 |
Table 2 Demographic Characteristics of Tibetan and Han Populations |
Association of PPARA SNPs with COPD Susceptibility
After adjusting for gender, age, smoking, alcohol consumption, education level, and hypertension, logistic regression results showed that in the Tibetan population, the rs135539A>C locus was associated with a 77.9% increased risk of COPD in the dominant model (CC+CA vs. AA: OR = 1.779, 95% CI: 1.080–2.903, P = 0.024). In the Han Chinese population, rs135549T>C was associated with a 28.1% reduction in COPD risk in the dominant model (CC+CT vs. TT: OR = 0.719, 95% CI: 0.547–0.944, P = 0.017); rs4253758T>C exhibited dual effects: In the additive model among Han Chinese, individuals with the TC genotype had a 72.2% increased risk of COPD compared to those with the wild-type TT genotype (TC vs. TT: OR = 1.722, 95% CI = 1.504–2.813, P = 0.030), while those with the CC genotype had a 77.8% increased risk (CC vs. TT: OR=1.778, 95% CI=1.083–2.922, P=0.023); In the recessive model among the Han Chinese population, individuals with the CC genotype had a 42.8% lower risk of developing COPD compared to those with the TT+TC genotype (CC vs. TT+TC: OR=0.572, 95% CI=0.356–0.918, P=0.021) (Table 3 and Supplementary Tables 3–6).
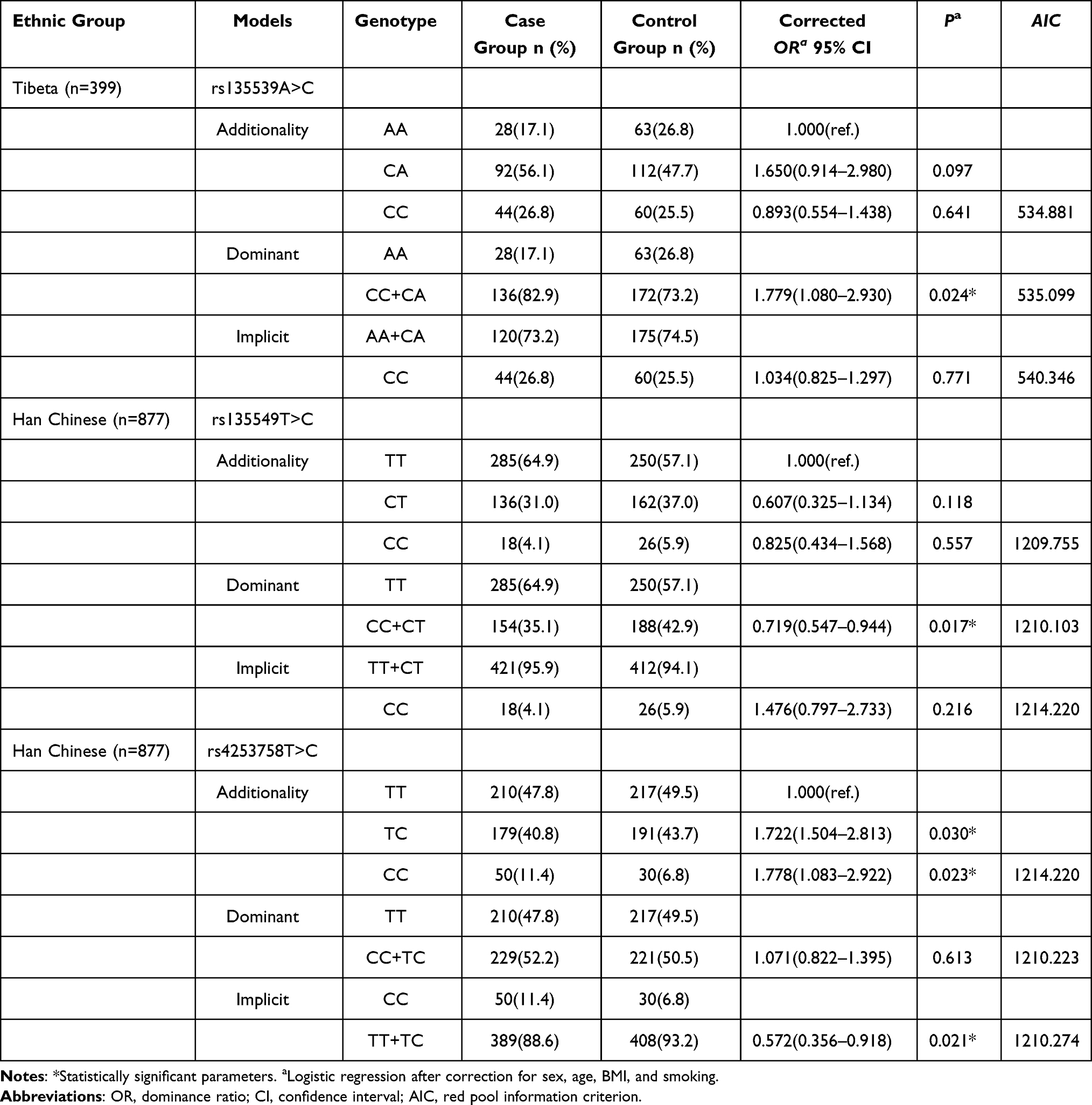 |
Table 3 Association Analysis of rs135539A>C, rs135549T>C, and rs4253758T>C with COPD Susceptibility in Tibetan and Han Populations |
Stratified and Interaction Analysis for PPARA rs135539A>C
Stratified analysis for rs135539A>C in the Tibetan population (Table 4) showed no significant heterogeneity or multiplicative interaction across subgroups (P-heterogeneity > 0.05, P-interaction > 0.05). The combined CA+CC genotype exhibited a protective effect in males (OR = 0.587, 95% CI: 0.355–0.971), individuals aged >60 (OR = 0.592, 95% CI: 0.357–0.981), and non-smokers (OR = 0.562, 95% CI: 0.339–0.932).
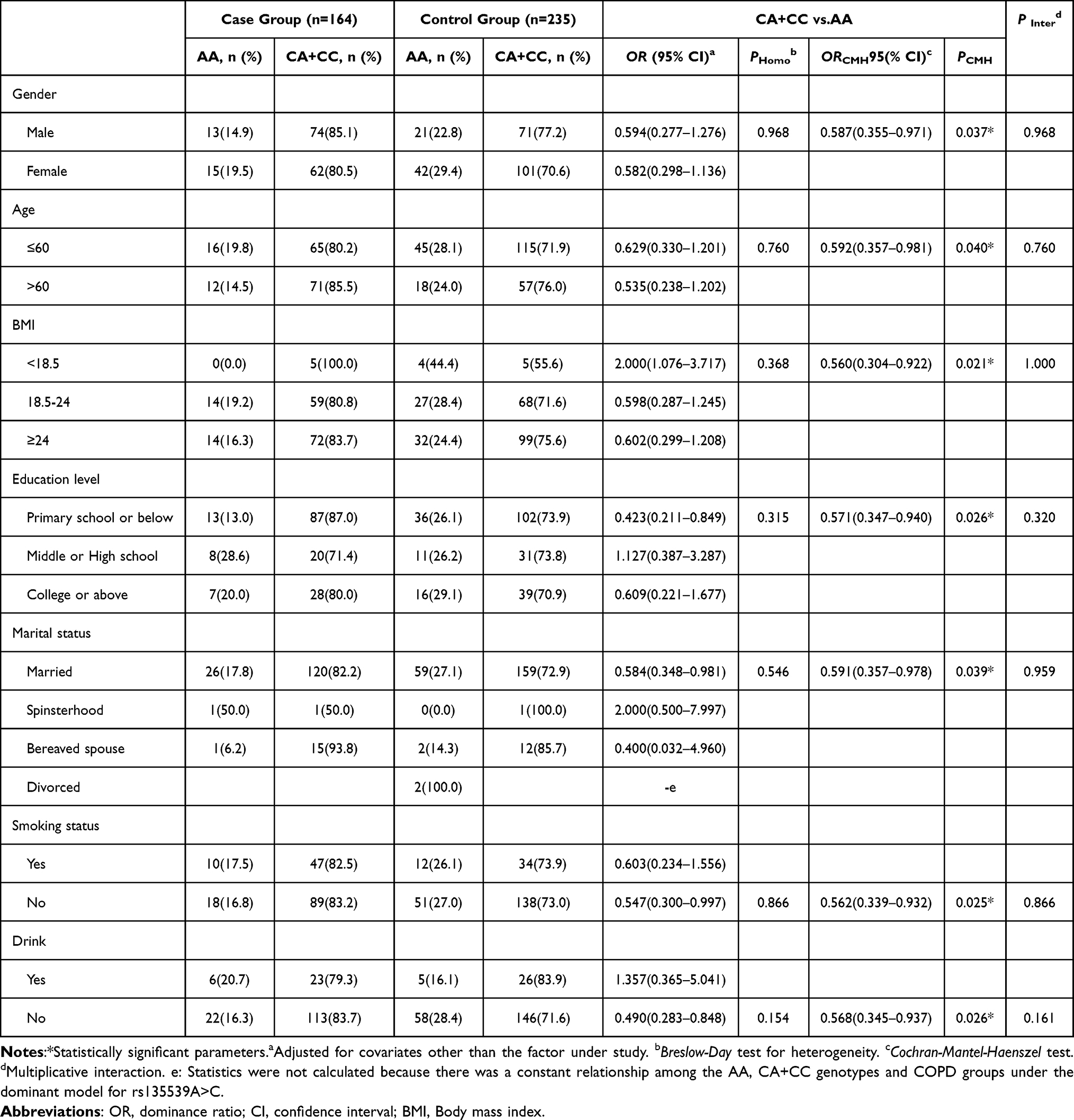 |
Table 4 Stratified Analysis and Interaction Analysis for rs135539A>C in the Tibetan Population |
Stratified and Interaction Analysis for PPARA rs135549T>C
For rs135549T>C in the Han population (Table 5), no significant heterogeneity or interaction was observed. The CT+CC genotype was associated with increased COPD risk in males (OR = 1.418, 95% CI: 1.068–1.882), those aged >60 (OR = 1.367, 95% CI: 1.030–1.815), and non-smokers (OR = 1.385, 95% CI: 1.054–1.820).
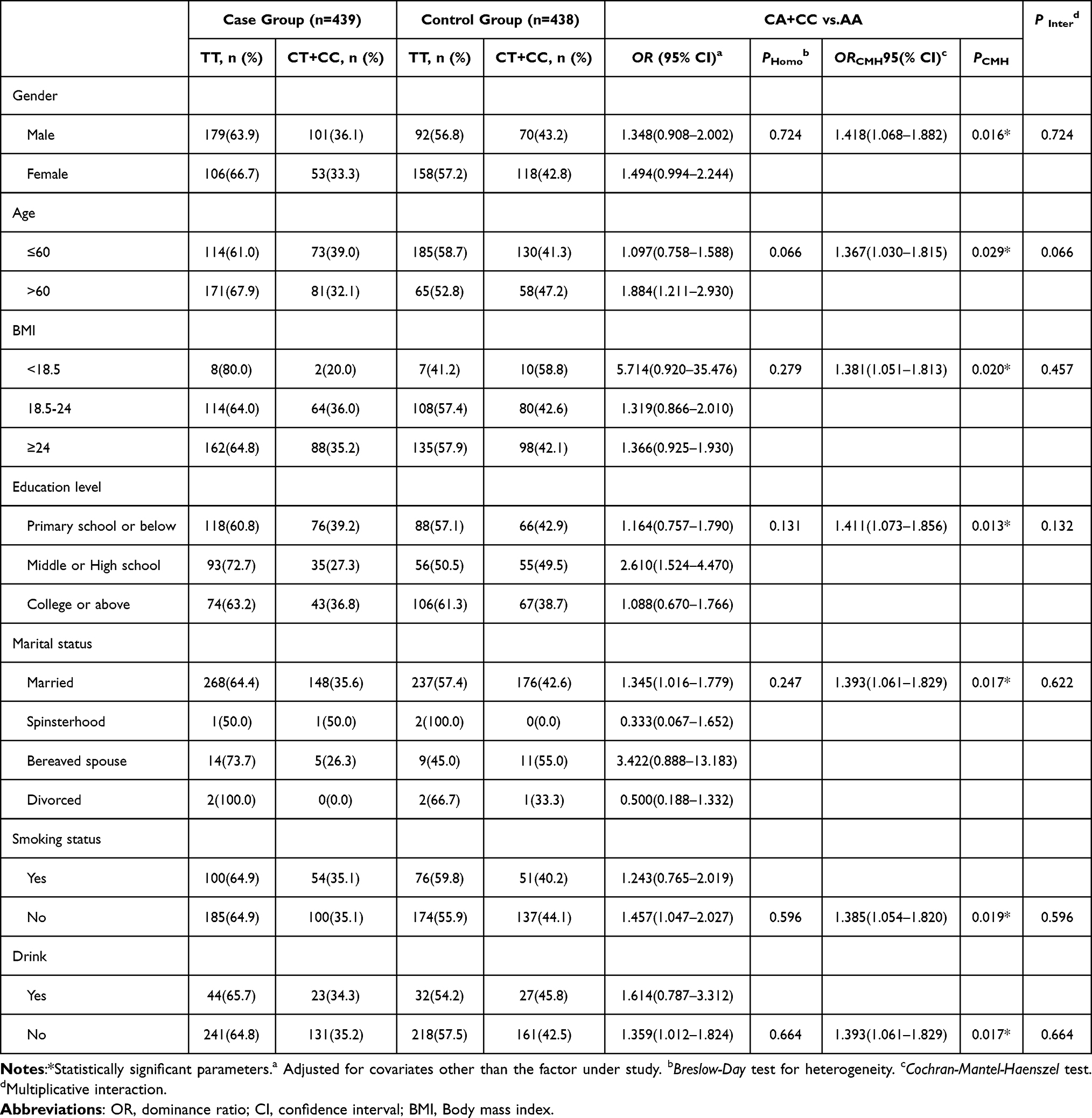 |
Table 5 Stratified Analysis and Interaction Analysis for rs135549T>C in the Han Chinese Population |
Stratified and Interaction Analysis for PPARA rs4253758T>C
For rs4253758T>C in the Han population (Table 6), no significant heterogeneity or interaction was found. The TT+TC genotype was associated with increased COPD risk in males (OR = 1.932, 95% CI: 1.171–3.187), individuals aged >60 (OR = 1.641, 95% CI: 1.006–2.677), and non-smokers (OR = 1.787, 95% CI: 1.109–2.881).
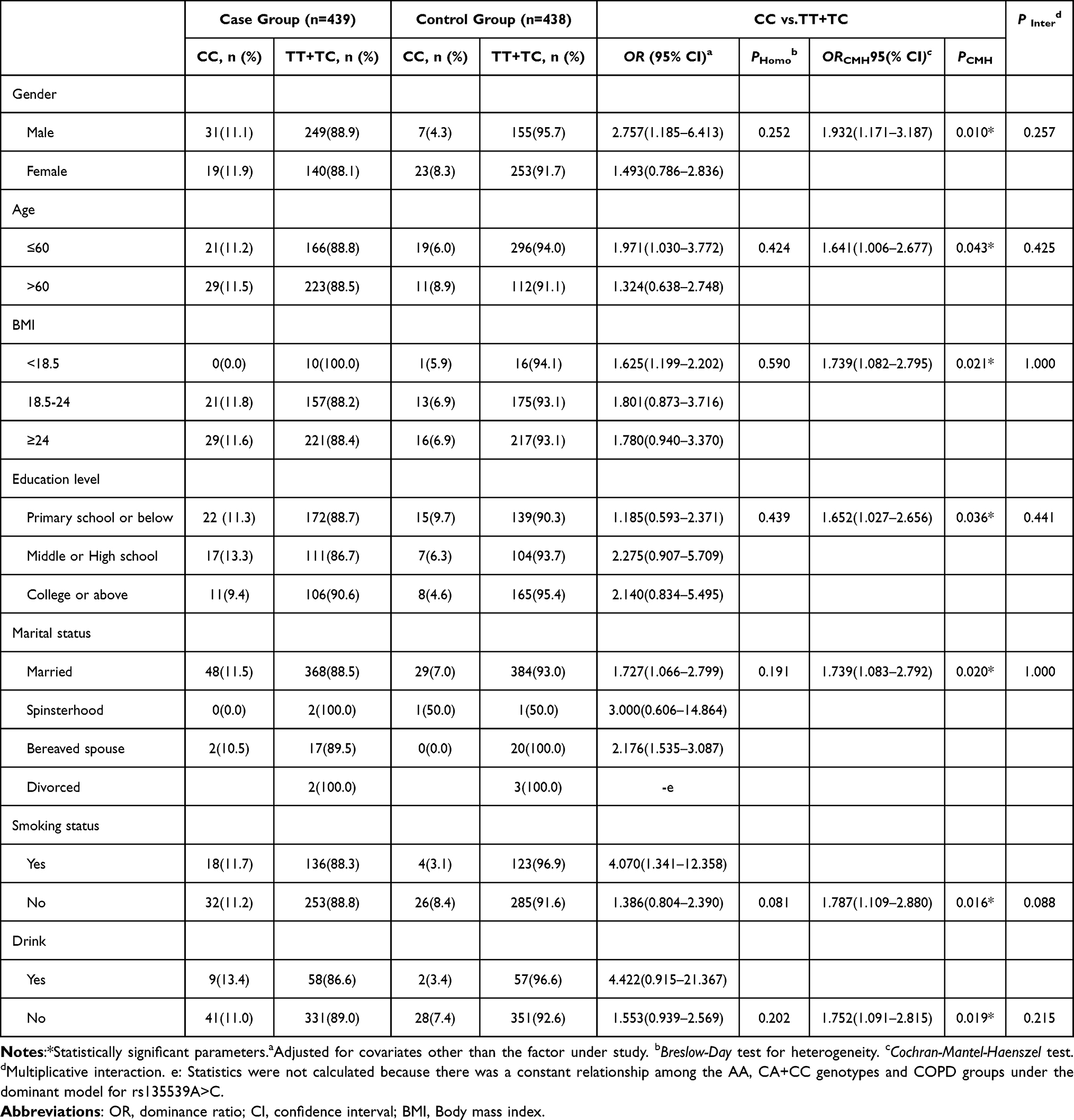 |
Table 6 Stratified Analysis and Interaction Analysis for rs4253758T>C in the Han Chinese Population |
Association Between SNP Genotypes and COPD Severity
COPD severity was graded based on post-bronchodilator FEV1% predicted: mild (≥80%), moderate (50–79%), severe (30–49%), and very severe (<30%). As shown in Table 7, no significant associations were found between rs135539A>C, rs135549T>C, or rs4253758T>C genotypes and COPD severity in their respective populations (all P > 0.05, Kruskal–Wallis H-test).
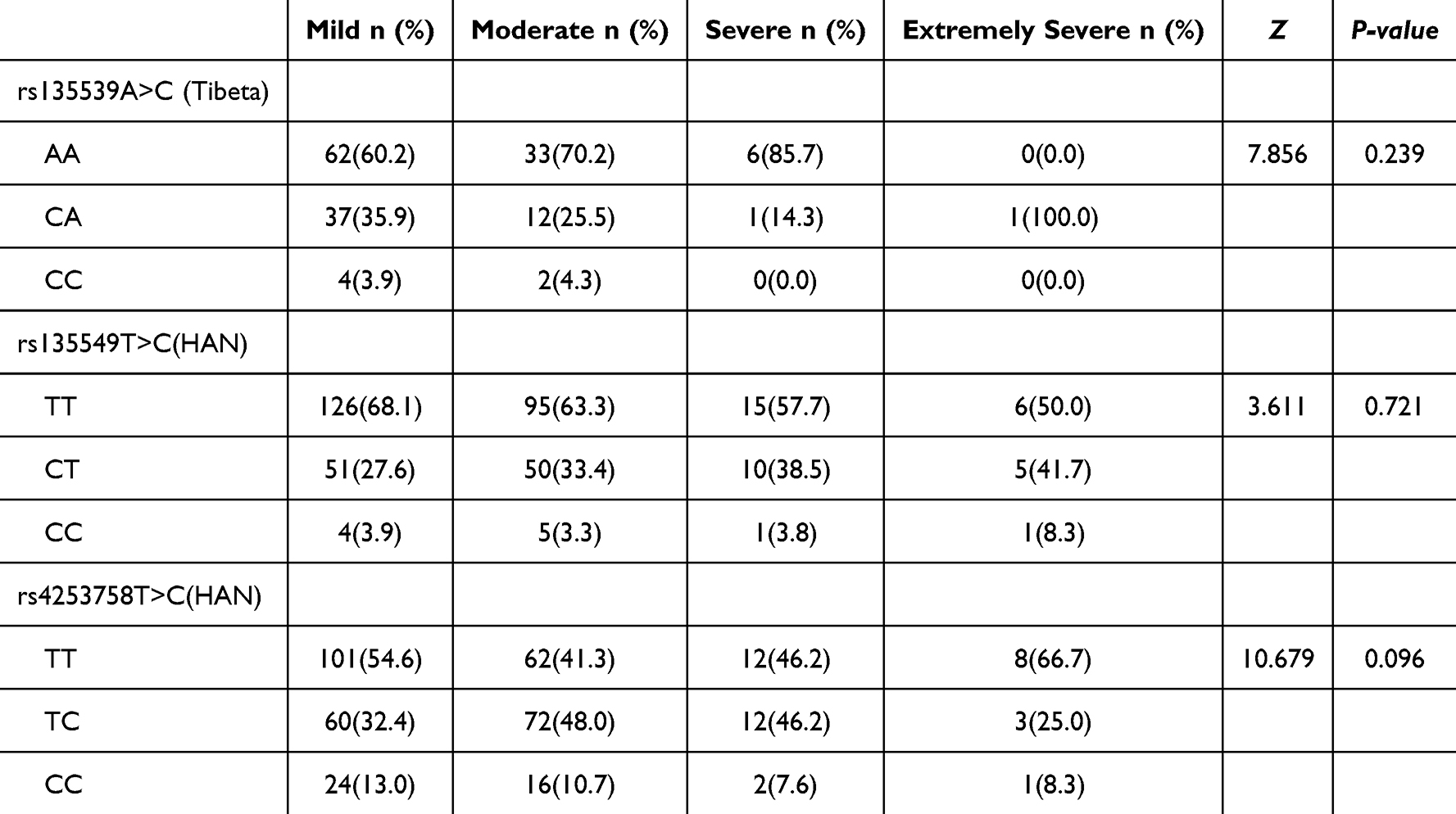 |
Table 7 Distribution of SNP Genotypes by COPD Severity [n(%)] |
Discussion
Genetic polymorphisms are considered to be closely associated with the genetic variation of COPD. Currently, there are few studies on the relationship between PPARA gene polymorphisms and COPD. To elucidate the role of PPARA polymorphisms in COPD development, we investigated the association between four PPARA gene SNPs (rs135538C>G, rs135539A>C, rs135549T>C, rs4253758T>C) and COPD risk in a Gansu Tibetan population in China. Notably, the distribution of PPARA gene polymorphisms and COPD susceptibility differed between Tibetan and Han populations in Gansu. Three SNPs (rs135539A>C, rs135549T>C, rs4253758T>C) in the PPARA gene showed higher susceptibility in individuals aged >60 years, males, and smokers (P<0.05). Among the Tibetan population, the rs135539A>C allele was associated with increased COPD risk, with CC+CA genotypes exhibiting higher susceptibility than AA (P<0.05). Among the Han population, the rs135549T>C allele was associated with reduced COPD risk, with CC+CT genotypes showing higher susceptibility than TT (P<0.05). rs4253758T>C exhibited dual effects in the Han Chinese population: increased COPD risk in the additive model (TC or CC genotypes showed higher susceptibility than TT genotype, P<0.05), and decreased COPD risk in the recessive model (CC genotype showed lower susceptibility than TT+TC genotypes, P<0.05). rs135538C>G was not associated with COPD severity.
PPARA, as a nuclear receptor transcription factor, plays a pivotal role in inflammation, oxidative stress, and lipid metabolism, contributing to the progression of COPD.28–30 COPD patients often exhibit reduced airway oxygen partial pressure, creating a chronic hypoxic environment that activates the HIF oxygen-sensitive pathway, leading to the accumulation and activation of HIF-1α and HIF-2α.31,32 The PPARA gene is a crucial regulator of the HIF pathway. Under hypoxic conditions, it interacts with HIF-2α and PHD to regulate erythropoiesis and angiogenesis, enhance energy and oxygen utilization, and mitigate hypoxic injury.33,34
In this study, we found that in the Tibetan population dominant model, individuals carrying the CC+CA genotype had a 77.9% increased risk of developing COPD compared to those with the wild-type AA genotype (CC+CA vs. AA: OR=1.779, 95% CI=1.080–2.930, P=0.024). The rs135539A>C variant is located within an intron region of the PPARA gene, a non-coding segment of DNA situated between exons.35 Early studies suggested introns lacked biological function as mere non-coding sequences; however, current research indicates introns participate in splicing processes, influencing mature mRNA formation.36,37 Regulatory elements within introns modulate splicing sites and efficiency, thereby regulating gene expression. The Fast study38 suggests introns may contain regulatory sequences such as transcription start sites, enhancers, and suppressors, directly influencing transcriptional control. Xie et al39 demonstrated that the rs135539A>C mutation may suppress PPARA expression, thereby increasing COPD susceptibility. While no direct studies have yet established an association between rs135539A>C and COPD risk, research indicates its involvement in inflammation regulation, oxidative stress, lung development, and apoptosis.40,41 Bitos et al42 demonstrated that COPD symptoms resemble those observed in populations chronically exposed to high-altitude hypoxic environments, suggesting hypoxia plays a role in COPD pathogenesis. Compared to Han Chinese populations, Tibetan populations possess unique genetic backgrounds with differences in certain genetic polymorphisms. The rs135539A>C variant is more prevalent in Tibetan populations and exhibits stronger effects. Living in high-altitude regions, the Tibetan population faces unique environmental challenges such as hypoxia and cold temperatures. These environmental factors may interact with the rs135539A>C variant, jointly affecting PPARA gene expression and function, thereby increasing COPD susceptibility. Hypoxic environments exacerbate lung injury and inflammatory responses, while the rs135539A>C variant further impairs the lungs’ adaptive capacity to these environmental stresses. The PPARα receptor encoded by the PPARA gene plays a crucial role in fatty acid oxidation and energy metabolism.43 In the Tibetan population, the rs135539A>C variant may influence COPD susceptibility by affecting PPARα function, thereby disrupting pulmonary adaptation to hypoxia and energy metabolism processes.44 Therefore, the role of rs135539A>C in the Tibetan population may differ from that in the Han Chinese population. Additionally, specific environmental factors and lifestyle characteristics in the Gansu region may also influence the relationship between this variant and COPD.
Our study found that in the dominant model among Han Chinese, individuals carrying the CC+CT genotype had a 28.1% lower risk of developing COPD compared to those with the wild-type TT genotype (CC+CT vs. TT: OR=0.719, 95% CI=0.547–0.944, P=0.017), indicating that the C allele at rs135549T>C is associated with a reduced risk of COPD in the Han Chinese population. The PPARA gene is a key regulator of fatty acid metabolism, involved in crucial biological processes such as fatty acid oxidation and inflammatory responses.45,46 PPARA also functions as a key anti-inflammatory factor by suppressing the production and release of inflammatory mediators.47 Zhang et al48 demonstrated that the C allele may downregulate PPARA gene expression, thereby affecting fatty acid metabolism and inflammatory regulation. Dubois et al49 showed that the C allele alters PPARA protein function, impacting its regulatory effects on target genes. In the Han Chinese population, this variant may reduce COPD risk by enhancing PPARA activity, thereby strengthening lung cells’ resistance to oxidative stress and inflammation. This anti-inflammatory effect may be particularly pronounced in Han Chinese, synergizing with regional environmental factors and lifestyle to maintain lung health.
We found that in the additive model among Han Chinese, individuals carrying the TC genotype at rs4253758T>C had a 72.2% increased risk of COPD compared to those with the wild-type TT genotype (TC vs. TT: OR=1.722, 95% CI=1.504–2.813, P=0.030), while those with the CC genotype had a 77.8% increased risk (CC vs. TT: OR=1.778, 95% CI=1.083–2.922, P=0.023). In the recessive model among Han Chinese, the rs4253758T>C variant was associated with a 42.8% reduced risk of COPD in CC genotype carriers compared to TT+TC genotype carriers (CC vs. TT+TC: OR=0.572, 95% CI=0.356–0.918, P=0.021). The rs4253758T>C variant exhibits dual effects in the Han Chinese population. This may arise because PPARα, encoded by the PPARA gene, regulates lipid metabolism, inflammatory responses, and cell proliferation. Variants at the rs4253758 site may enhance protein function or alter expression levels, thereby influencing lung tissue resistance to oxidative stress and inflammatory damage.50 This phenomenon highlights the importance of race-environment interactions in the role of genetic factors in disease susceptibility.51 Furthermore, this dual effect may also be associated with interactions involving multiple factors such as other genetic backgrounds, lifestyle, environmental exposures, and epigenetic modifications. Future studies need to further explore these potential mechanisms.
In summary, this study identified associations between rs135539A>C, rs135549T>C, and rs4253758T>C variants and COPD susceptibility by regulating PPARA gene expression levels, thereby influencing downstream fatty acid oxidation, inflammation suppression, and HIF pathway homeostasis. The dual effect of rs4253758T>C demonstrates the impact of PPARA gene-environment interactions on disease outcomes. This study provides a plausible biological framework for interpreting genetic association results, positioning the PPARA gene at the core of the link between hypoxic adaptation, metabolic regulation, and COPD pathogenesis. It offers important scientific evidence for identifying early diagnostic biomarkers and screening susceptible populations for COPD. However, this study has certain limitations. Our current research focused only on four loci of the PPARA gene and did not conduct a comprehensive analysis. Furthermore, large-scale cohort studies and functional studies are needed to fully understand the specific mechanisms of the PPARA gene in COPD.
Conclusion
This case-control study is the first to systematically demonstrate population-specific associations between PPARA gene polymorphisms and COPD susceptibility among Tibetan and Han populations in Gansu, China. The rs135539A>C variant increases COPD risk in Tibetans, whereas rs135549T>C confers protection and rs4253758T>C shows a dual role in Han Chinese. These findings underscore the importance of considering ethnic background in genetic studies of complex diseases like COPD and provide valuable molecular epidemiological insights for developing population-specific strategies for COPD prevention and precision medicine.
Abbreviations
COPD: Chronic Obstructive Pulmonary Disease; PPARA: Peroxisome Proliferator-Activated Receptor Alpha; HIF: Hypoxia-Inducible Factor; PHD: Prolyl Hydroxylase Domain enzyme; SNP: Single Nucleotide Polymorphism; GWAS: Genome-Wide Association Study; FEV1: Forced Expiratory Volume in 1 second; FVC: Forced Vital Capacity; MAF: Minor Allele Frequency; PCR: Polymerase Chain Reaction.
Data Sharing Statement
The data supporting this study’s findings are available from the corresponding author upon reasonable request.
Ethical Approval and Consent
The study was conducted per the Declaration of Helsinki and approved by the Institutional Review Board of Guangzhou Medical University Center (Approval No.: 202405010). Written informed consent was obtained from all participants.
Acknowledgments
We thank the School of Public Health, Gansu University of Chinese Medicine. We also acknowledge all laboratory members for their technical guidance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Major Project of the Gansu Provincial Joint Research Fund (23JRRA1520), the Lanzhou Major Project (HXLH-JBGS03), and the 2025 Open Fund Project of the Collaborative Innovation Center for Traditional Chinese Medicine Prevention and Control of Nutrition- and Environment-Related Diseases in Northwest China (ZYXT-25-9).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Sandelowsky H, Weinreich UM, Aarli BB, et al. COPD - do the right thing. BMC Fam Pract. 2021;22(1):244. doi:10.1186/s12875-021-01583-w
2. Uwagboe I, Adcock IM, Lo Bello F, Caramori G, Mumby S. New drugs under development for COPD. Minerva Med. 2022;113(3):471–14. doi:10.23736/s0026-4806.22.08024-7
3. Al Wachami N, Guennouni M, Iderdar Y, et al. Estimating the global prevalence of chronic obstructive pulmonary disease (COPD): a systematic review and meta-analysis. BMC Public Health. 2024;24(1):297. doi:10.1186/s12889-024-17686-9
4. James SL, Abate D, Abate KH, et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990-2017: a systematic analysis for the global burden of disease study 2017. Lancet. 2018;392(10159):1789–1858. doi:10.1016/s0140-6736(18)32279-7
5. Vos T, Lim SS, Abbafati C, et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990-2019: a systematic analysis for the global burden of disease study 2019. Lancet. 2020;396(10258):1204–1222. doi:10.1016/s0140-6736(20)30925-9
6. Boers E, Barrett M, Su JG, et al. Global burden of chronic obstructive pulmonary disease through 2050. JAMA Netw Open. 2023;6(12):e2346598. doi:10.1001/jamanetworkopen.2023.46598
7. Wang H, Ye X, Zhang Y, Ling S. Global, regional, and national burden of chronic obstructive pulmonary disease from 1990 to 2019. Front Physiol. 2022;13:925132. doi:10.3389/fphys.2022.925132
8. Zhang J, Xie Y, Kwong JS, et al. The efficacy and safety of revefenacin for the treatment of chronic obstructive pulmonary disease: a systematic review. Front Pharmacol. 2021;12:667027. doi:10.3389/fphar.2021.667027
9. Waeijen-Smit K, Houben-Wilke S, DiGiandomenico A, Gehrmann U, Franssen FME. Unmet needs in the management of exacerbations of chronic obstructive pulmonary disease. Intern Emerg Med. 2021;16(3):559–569. doi:10.1007/s11739-020-02612-9
10. Ji J, Zhao Q, Yuan J, Yuan Z, Gao N. Causal associations between chronic obstructive pulmonary disease and common comorbidities: evidence from comprehensive genetic methods. Int J Chron Obstruct Pulmon Dis. 2025;20:601–610. doi:10.2147/copd.S498513
11. Rey-Brandariz J, Pérez-Ríos M, Ahluwalia JS, et al. Tobacco patterns and risk of chronic obstructive pulmonary disease: results from a cross-sectional study. Arch Bronconeumol. 2023;59(11):717–724. doi:10.1016/j.arbres.2023.07.009
12. Yang IA, Jenkins CR, Salvi SS. Chronic obstructive pulmonary disease in never-smokers: risk factors, pathogenesis, and implications for prevention and treatment. Lancet Respir Med. 2022;10(5):497–511. doi:10.1016/s2213-2600(21)00506-3
13. Chen Y, Peng C, Zhang H, et al. Exposure to occupational risk factors is associated with the severity and progression of chronic obstructive pulmonary disease. Medicine. 2023;102(6):e32908. doi:10.1097/md.0000000000032908
14. Marcon A, Locatelli F, Dharmage SC, et al. The coexistence of asthma and COPD: risk factors, clinical history and lung function trajectories. Eur Respir J. 2021;58(5):2004656. doi:10.1183/13993003.04656-2020
15. Chen G, Jin Y, Chu C, et al. A cross-tissue transcriptome-wide association study reveals GRK4 as a novel susceptibility gene for COPD. Sci Rep. 2024;14(1):28438. doi:10.1038/s41598-024-80122-w
16. Wen H, Zhang R, Zhong B, Liu H, Liu C. Cross-trait genome-wide association study identifies shared genetic risk loci between COPD and Five autoimmune diseases. Int J Chron Obstruct Pulmon Dis. 2025;20:3019–3034. doi:10.2147/copd.S533401
17. Maciejewska-Skrendo A, Massidda M, Tocco F, Leźnicka K. The influence of the differentiation of genes encoding peroxisome proliferator-activated receptors and their coactivators on nutrient and energy metabolism. Nutrients. 2022;14(24):5378. doi:10.3390/nu14245378
18. Shen Y, Zhang J, Yang J, et al. Association of EPAS1 and PPARA gene polymorphisms with high-altitude headache in Chinese Han population. Biomed Res Int. 2020;2020:1593068. doi:10.1155/2020/1593068
19. Chen Z, Dong Z, Zeng R, et al. Association between single nucleotide polymorphisms in EPAS1 and PPARA genes and high altitude polycythemia in Chinese Tibetan population. Front Genet. 2025;16:1519108. doi:10.3389/fgene.2025.1519108
20. Li H, Pei W, Wang Y, Zhang Y, Yang Z, Wang X. Mechanism of action of the plateau-adapted gene PPARA in COPD. Front Biosci. 2024;29(2):68. doi:10.31083/j.fbl2902068
21. Aibara D, Takahashi S, Yagai T, et al. Gene repression through epigenetic modulation by PPARA enhances hepatocellular proliferation. iScience. 2022;25(5):104196. doi:10.1016/j.isci.2022.104196
22. Lucarelli R, Gorrochotegui-Escalante N, Taddeo J, Buttaro B, Beld J, Tam V. Eicosanoid-activated PPARα inhibits NFκB-dependent bacterial clearance during post-influenza superinfection. Front Cell Infect Microbiol. 2022;12:881462. doi:10.3389/fcimb.2022.881462
23. Wang X, Wang J, Ying C, Xing Y, Su X, Men K. Fenofibrate alleviates NAFLD by enhancing the PPARα/PGC-1α signaling pathway coupling mitochondrial function. BMC Pharmacol Toxicol. 2024;25(1):7. doi:10.1186/s40360-023-00730-6
24. Kim TS, Jin YB, Kim YS, et al. SIRT3 promotes antimycobacterial defenses by coordinating mitochondrial and autophagic functions. Autophagy. 2019;15(8):1356–1375. doi:10.1080/15548627.2019.1582743
25. Li J, Lu Y, Li N, et al. Chemerin: a potential regulator of inflammation and metabolism for chronic obstructive pulmonary disease and pulmonary rehabilitation. Biomed Res Int. 2020;2020:4574509. doi:10.1155/2020/4574509
26. Xu JJ, Xu F, Wang W, et al. Paeoniae Radix Rubra can enhance fatty acid β-oxidation and alleviate gut microbiota disorder in α-naphthyl isothiocyanate induced cholestatic model rats. Front Pharmacol. 2022;13:1002922. doi:10.3389/fphar.2022.1002922
27. Agustí A, Celli BR, Criner GJ, et al. Global initiative for chronic obstructive lung disease 2023 report: GOLD executive summary. Eur Respir J. 2023;61(4):2300239. doi:10.1183/13993003.00239-2023
28. Lee IT, Yang CC, Yang CM. Harnessing peroxisome proliferator-activated receptor γ agonists to induce Heme Oxygenase-1: a promising approach for pulmonary inflammatory disorders. Cell Commun Signal. 2024;22(1):125. doi:10.1186/s12964-024-01501-4
29. Yamamoto Y, Shirai Y, Sonehara K, et al. Dissecting cross-population polygenic heterogeneity across respiratory and cardiometabolic diseases. Nat Commun. 2025;16(1):3765. doi:10.1038/s41467-025-58149-y
30. Zhang L, Guo R, Wu H, et al. The crucial role of the PPAR signaling pathway in the diagnosis and treatment of chronic obstructive pulmonary disease: an analysis of gene expression and macrophage polarization. Int J Chron Obstruct Pulmon Dis. 2025;20:2287–2304. doi:10.2147/copd.S518592
31. Simargi Y, Turana Y, Icksan AG, et al. A multicenter study of COPD and cognitive impairment: unraveling the interplay of quantitative CT, lung function, HIF-1α, and clinical variables. Int J Chron Obstruct Pulmon Dis. 2024;19:1741–1753. doi:10.2147/copd.S466173
32. Liu Q, Zhang Y, Duan R, et al. Transcriptome reveals the landscape of alveolar macrophages exposed to combined hypoxia with cigarette smoke extract. Respir Res. 2025;26(1):241. doi:10.1186/s12931-025-03303-9
33. Bhandari S, Cavalleri GL. Population history and altitude-related adaptation in the sherpa. Front Physiol. 2019;10:1116. doi:10.3389/fphys.2019.01116
34. Wang H, Wang L, Li Y, et al. The HIF-2α/PPARα pathway is essential for liraglutide-alleviated, lipid-induced hepatic steatosis. Biomed Pharmacother. 2021;140:111778. doi:10.1016/j.biopha.2021.111778
35. Bai ZZ, Jin GE, Wu-Ren T, Ga Q, Ge RL. Energy power in mountains: difference in metabolism pattern results in different adaption traits in Tibetans. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 2012;28(6):488–493.
36. Mishra A, Mohammad G, Thinlas T, Pasha MA. EGLN1 variants influence expression and SaO2 levels to associate with high-altitude pulmonary oedema and adaptation. Clin Sci. 2013;124(7):479–489. doi:10.1042/cs20120371
37. Wang T, Lu H. Ganoderic acid A inhibits ox-LDL-induced THP-1-derived macrophage inflammation and lipid deposition via Notch1/PPARγ/CD36 signaling. Adv Clin Exp Med. 2021;30(10):1031–1041. doi:10.17219/acem/137914
38. Fast NM. Intron splicing: U12 spliceosomal introns not so ‘minor’ after all. Curr Biol. 2021;31(14):R912–r914. doi:10.1016/j.cub.2021.06.008
39. Xie HJ, Hai B, Wu M, et al. Analysis on the association between PPARα/γ polymorphisms and lipoprotein(a) in a Chinese Han population. Mol Genet Genomics. 2014;289(5):981–987. doi:10.1007/s00438-014-0866-9
40. Gu S-J, Chen D-H, Guo Z-R, Zhou Z-Y, Hu X-S, Wu M. Effect of obesity on the association between common variations in the PPAR gene and C-reactive protein level in Chinese Han population. Endocrine. 2015;48(1):195–202. doi:10.1007/s12020-014-0218-x
41. Liu M, Zhang J, Guo Z, et al. Association and interaction between 10 SNP of peroxisome proliferator-activated receptor and non-HDL-C. Zhonghua Yu Fang Yi Xue Za Zhi. 2015;49(3):259–264.
42. Bitos K, Furian M, Mayer L, et al. Effect of high-flow oxygen on exercise performance in COPD patients randomized trial. Front Med. 2020;7:595450. doi:10.3389/fmed.2020.595450
43. Li Y, Xiong Z, Yan W, et al. Branched chain amino acids exacerbate myocardial ischemia/reperfusion vulnerability via enhancing GCN2/ATF6/PPAR-α pathway-dependent fatty acid oxidation. Theranostics. 2020;10(12):5623–5640. doi:10.7150/thno.44836
44. Bougarne N, Weyers B, Desmet SJ, et al. Molecular actions of PPARα in lipid metabolism and inflammation. Endocr Rev. 2018;39(5):760–802. doi:10.1210/er.2018-00064
45. Li Q, Zhang W, Cheng N, et al. Pectolinarigenin ameliorates Acetaminophen-induced acute liver injury via attenuating oxidative stress and inflammatory response in Nrf2 and PPARa dependent manners. Phytomedicine. 2023;113:154726. doi:10.1016/j.phymed.2023.154726
46. Brocker CN, Patel DP, Velenosi TJ, et al. Extrahepatic PPARα modulates fatty acid oxidation and attenuates fasting-induced hepatosteatosis in mice. J Lipid Res. 2018;59(11):2140–2152. doi:10.1194/jlr.M088419
47. Zhou B, Zhao G, Li H, Zhao Q, Liu D. FNDC5/PPARa pathway alleviates THP-1-derived macrophage pyroptosis and its mechanism. Altern Ther Health Med. 2023;29(3):32–42.
48. Zhang C, Jiao L, Bai H, et al. Association of POR and PPARα polymorphisms with risk of anti-tuberculosis drug-induced liver injury in Western Chinese Han population. Infect Genet Evol. 2020;79:104147. doi:10.1016/j.meegid.2019.104147
49. Dubois V, Eeckhoute J, Lefebvre P, Staels B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J Clin Invest. 2017;127(4):1202–1214. doi:10.1172/jci88894
50. Wang YM, Ji R, Chen WW, et al. Paclitaxel alleviated sepsis-induced acute lung injury by activating MUC1 and suppressing TLR-4/NF-κB pathway. Drug Des Devel Ther. 2019;13:3391–3404. doi:10.2147/dddt.S222296
51. Tam VC, Suen R, Treuting PM, et al. PPARα exacerbates necroptosis, leading to increased mortality in postinfluenza bacterial superinfection. Proc Natl Acad Sci U S A. 2020;117(27):15789–15798. doi:10.1073/pnas.2006343117
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.