Back to Journals » OncoTargets and Therapy » Volume 16
Potential Therapeutic Targets for Luminal Androgen Receptor Breast Cancer: What We Know so Far
Authors Stella S ![]() , Martorana F, Massimino M, Vitale SR
, Martorana F, Massimino M, Vitale SR ![]() , Manzella L, Vigneri P
, Manzella L, Vigneri P
Received 30 December 2022
Accepted for publication 24 March 2023
Published 7 April 2023 Volume 2023:16 Pages 235—247
DOI https://doi.org/10.2147/OTT.S379867
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Geoffrey Pietersz
Stefania Stella,1,2 Federica Martorana,1,2 Michele Massimino,1,2 Silvia Rita Vitale,1,2 Livia Manzella,1,2 Paolo Vigneri1,2
1Department of Clinical and Experimental Medicine, University of Catania, Catania, Italy; 2Center of Experimental Oncology and Hematology, A.O.U. Policlinico “G. Rodolico - San Marco”, Catania, Italy
Correspondence: Stefania Stella, University of Catania, Department of Clinical and Experimental Medicine, Center of Experimental Oncology and Hematology, A.O.U. Policlinico “G. Rodolico - San Marco”, Via S. Sofia, 78, Edificio 8D/2, Catania, Italy, Tel +39 95 378 1946, Email [email protected]; [email protected]
Abstract: Luminal Androgen Receptor Breast Cancers (LAR BCs) are characterized by a triple negative phenotype and by the expression of Androgen Receptor (AR), coupled with luminal-like genomic features. This unique BC subtype, accounting for about 10% of all triple negative BC, has raised considerable interest given its ill-defined clinical behavior and the chance to exploit AR as a therapeutic target. The complexity of AR activity in BC cells, as revealed by decades of mechanistic studies, holds promise to offer additional therapeutic options beyond mere AR inhibition. Indeed, preclinical and translational evidence showed that several pathways and mediators, including PI3K/mToR, HER2, BRCA1, cell cycle and immune modulation, can be tackled in LAR BCs. Moving from bench to bedside, several clinical trials tested anti-androgen therapies in LAR BCs, but their results are inconsistent and often disappointing. More recently, studies exploring combinations of anti-androgen agents with other targeted therapies have been designed and are currently ongoing. While the results from these trials are awaited, a concerted effort will be needed to find the biological vulnerabilities of LAR BCs which may disclose new and effective therapeutic targets, eventually improving patients’ outcomes.
Keywords: triple negative breast cancer, luminal androgen receptor breast cancer, androgen receptor, targeted therapy, clinical trials
Introduction
In the last two decades, the evolving knowledge about breast cancer (BC) biology disclosed the vast molecular heterogeneity of this disease.1 According to gene expression profile, BC can be classified into five different subtypes: luminal A, luminal B, human epidermal growth factor receptor-2 (HER2) enriched, basal-like and normal-like.2 In the clinical practice, surrogate immunohistochemistry biomarkers are commonly used to distinguish between luminal tumors, which expresses estrogen and/or progesterone hormone receptors (HR), Human Epidermal Growth Factor Receptor 2 (HER2)-enriched tumors, which overexpresses HER2, and basal-like tumors, which lacks both HR and HER2 and are therefore also referred to as triple negative breast cancer (TNBC).3 This latter category accounts for 15–20% of all BC and is usually linked with an aggressive clinical behavior, a high risk of relapse and poor prognosis.4 Further evaluations of TNBC showed a broadly heterogeneous genomic landscape compared to other BC classes.5–7 According to Lehman’s classification, TNBC entails six different molecular subgroups, namely basal-like 1, basal-like 2, immunomodulatory, mesenchymal, mesenchymal stem-like and luminal androgen receptor (LAR).5 A subsequent study by Burstein et al identified four classes of TNBC: basal-like immune-suppressed (BLIS), basal-like immune-activated (BLIA), mesenchymal and LAR.6 Despite the differences, both these classifications acknowledge LAR BC as a distinct entity accounting for about 10% of all TNBCs.8 Luminal androgen receptor BCs harbor a luminal-like gene signature and, phenotypically, an androgen receptor (AR) expression.8,9 Since its identification, the LAR BC subtype raised significant interest for its peculiar features and the potential presence of a druggable target (ie AR).8,10 However, clinical characteristics of LAR BC are still ill-defined, with evidence suggesting a favorable prognosis and others indicating poorer outcomes.11–13 Similarly, the role of AR as a predictive biomarker of response to anti-androgen therapies has not been confirmed thus far.12,14
Here, we critically review the existing literature about AR role in TNBC and also discuss the updated evidence about AR inhibition in LAR BC, starting from the pre-clinical scenario and then moving to the clinical setting.
Androgen Receptor-Mediated Signaling Pathways in AR Positive TNBC
Structurally, AR comprises three functional domains: an N-terminal domain (NTD, residues 1–555), a DNA binding domain (DBD, residues 555–623) and a carboxyl-terminal domain (CTD, residues 665–919) which includes the ligand-binding domain (LBD). The receptor’s activity depends on two transcriptional activation functions sequence: the ligand-independent AF1 (transcriptional activating function 1), sited in NTD, and the ligand-dependent AF2 (transcriptional activating function 2) located in LBD15 (Figure 1A).
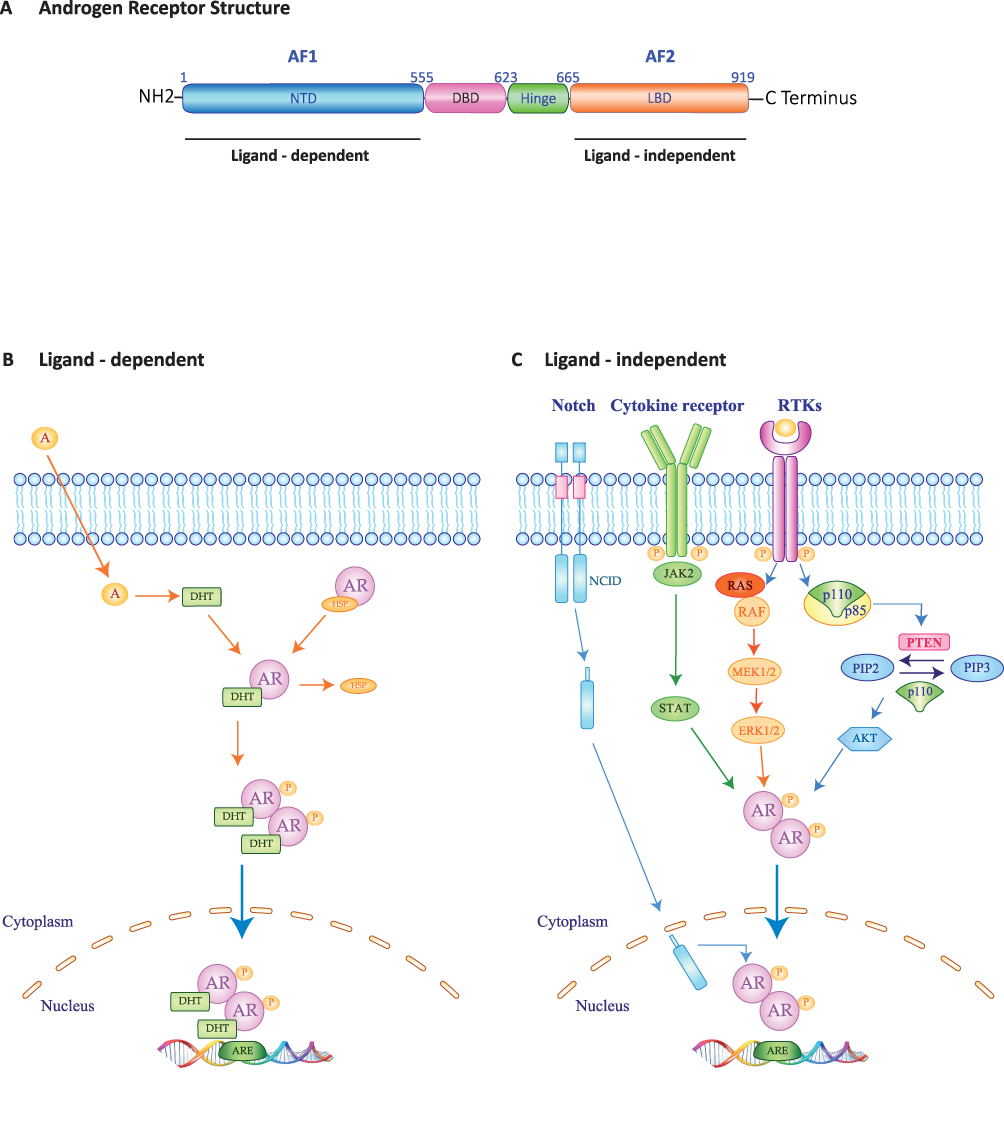 |
Figure 1 Molecular structure and schematic representation of Ligand-dependent and -independent of Androgen Receptor transactivation. (A) The androgen receptor includes three functional domains: a N-terminal domain (NTD) containing transcriptional activating functional 1 region (AF1); a DNA-binding domain (DBD); a C-terminal ligand-binding domain (LBD), including transcriptional activating functional 2 region (AF2), which is connected to the DBD by a flexible Hinge region. (B) Ligand-dependent (AR genomic pathways). AR resides in the cell cytoplasm in an active form linked to heat shock protein 90 and other chaperone proteins. In the process of ligand-dependent transactivation, AR binds the active androgens’ metabolite, such as 5a-dihydrotestosterone (DHT). AR is activated by phosphorylation (P) and dimerizes. The AR translocate into the nucleus where it binds to the androgen response elements (ARE) in the promoter regions of target genes. Depending on its interaction with other co-regulator, AR-ARE interaction leads to cell proliferation, apoptosis and survival. (C) Ligand-independent (AR genomic pathways). During this pathway, various growth factor, cytokines and receptor tyrosine kinases (RTKs) are activated and the AR is phosphorylated by multiple signalling pathways such as RAS/RAF/MAPK/ERK, PIK3/AKT/mTOR or JAK2/STAT pathways. Blue, light blue, Orange and green arrows represent signaling activation. Abbreviations: AKT, Protein kinase B; ERK, Extracellular signal-regulated kinase; JAK2, Janus kinase-signal transducer; MEK, Mitogen-activated protein; NCID, Notch intracellular domain; Notch, Neurogenic locus notch homolog protein; PIP2, phosphatidylinositol-4,5-bisphosphate; PIP3, phosphatidylinositol 3,4,5-trisphosphate; PTEN, PIP3 phosphatase and tensin homologue deleted on chromosome 10; RAF, Rapidly Accelerated Fibrosarcoma; RAS, Rat sarcoma virus; STAT3, Signal transducer and activator of transcription 3. |
Androgen Receptor has a role in several signal transduction pathways and is dynamically involved in breast carcinogenesis process. Indeed, AR displays both transcriptional/genomic and non-transcriptional/non-genomic mechanisms of action, in addition to exhibiting several cross-talks with cell-growth and survival mediators.8 The transcriptional/genomic mode can be ligand-dependent or ligand-independent. The ligand-dependent action requires a bound between androgens and their receptors, which is a key event promoting AR nuclear translocation. Into the nucleus, the AR DNA Binding Domain recognizes the androgen receptor element (ARE) (Figure 1B). This interaction determines regulation of gene expression. Androgen receptors recognize a plethora of target genes involved in cell proliferation, survival, fatty acid and lipid synthesis as well as porphyrin and androgen/estrogen metabolism.5
In the ligand-dependent mode, AR interacts and modulates the activity of several intracellular second-messengers, including mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK), Janus kinase-signal transducer and activator of transcription 3 (JAK/STAT3), neurogenic locus notch homolog protein (NOTCH)-mediated effectors and phosphatidylinositol-3-kinase/protein kinase B/mammalian target of rapamycin (PI3K/mTOR/AKT) pathways16–19 (Figure 1C). More specifically, a feedback loop between AR and ERK proteins has been proposed, which can occur either in an ERK-dependent or ERK-independent fashion.20,21 The ERK-dependent mode involves cytoplasmic AR interaction with PI3K, Src and RAS guanosine triphosphatase (GTPase), inducing ERK nuclear translocation and eventually promoting cell proliferation. In addition, following androgenic stimulation, either membrane-bound AR, G protein-coupled receptors (GPCRs) or sex hormone-binding globulin receptor (SHBGR) increase cyclic adenosine monophosphate (cAMP) expression, determining ERK pathway activation via protein kinase C, eventually converging on MAPK/ERK pathway (Figure 1C). On the other hand, the ERK-independent mode is related to the balance between mTOR and forkhead box protein O1 (FOXO1) deactivation, which induces cell proliferation. Specifically, cytoplasmic AR/PI3K interaction results in mTOR phosphorylation and activation followed by FOXO1 phosphorylation and inactivation. This latter determines apoptosis signaling downregulation via cytoplasmic AR/PI3K interaction or direct interaction between AR and FOXO1. Moreover, the modulation of intracellular Ca2+ concentration results in membrane binding of AR/GPCRs/SHBGR followed by protein kinase A (PKA) activation. Overall, the abovementioned pathways lead to increased cell proliferation.11,14,20,22,23
Evidences are also accumulating about the non-transcriptional/non-genomic mechanism of action, which does not require DNA binding and does not imply gene expression regulation.24
Androgen receptor activity is also influenced by multiple cross-talk networks, involving different signaling pathways, intracellular modulators and nuclear factors.8
For instance, the interaction between AR and Estrogen Receptor beta (ERß) can influence tumor growth in AR positive TNBC cells.25 Estrogen Receptor beta is expressed in one-third of Estrogen Receptor alpha (ERα) negative BC, including TNBC.26 Androgen receptor activation regulates the ERβ-mediated transcription by androgen responsive elements (ARE) present into the ERβ promoter. Moreover, ERβ inhibits AR nuclear translocation by stimulating the Phosphatase and tensin homolog (PTEN) protein. Activated PTEN inhibits mTOR signaling, promoting anti-proliferative effect. Given the potential ERß anti-tumor effects enhanced by AR stimulation, AR has been proposed as a favorable prognostic biomarker in ERß positive TNBC.24,27
A functional cross-talk exists between AR and HER2 signaling pathways and it seems to have a role in cell proliferation. Indeed, the simultaneous stimulation of AR and HER2 pathways in cell lines with molecular apocrine features positively modulates proliferation and exerts anti-apoptotic effects.28 Additionally, a next-generation sequencing experience on a small cohort of LAR BC patients showed a high incidence of HER2 mutations in this subset of patients.10
Androgen receptor and mitogen-activated protein kinase kinase 1 (MEK) activities also seem to be interrelated, as their co-inhibition reduces tumor growth and angiogenesis in-vivo.29
According to other evidence, BRCA1 onco-suppressor gene and its mutational status may influence AR signaling through the activation of the NH2-terminal domain (AF-1) of the receptor, especially in the presence of exogenous p160 coactivator.30 More recently, Zhang and colleagues demonstrated that BRCA1 hampers AR proliferative effects in BC cells through the activation of sirtuin (SIRT1) pathway.31 On the other hand, BRCA1 mutated cells seem to display reduced AR expression and might therefore present a reduced potency of AR-dependent signaling.30,32
Androgen receptor is also implicated in cell cycle regulation. A recent study suggests that LAR subtype may be sensitive to CDK4/6 inhibition due to a correlation between AR expression and low cyclin E1 levels.33 In addition, Michmerhuizen reported that AR and Retinoblastoma protein (Rb) expressions are positively associated, while AR has been shown to interact with other transcription factors, including MYC, which exerts a pivotal role in cell cycle control. Indeed, high AR expression is negatively correlated with MYC overexpression and promotes cyclin D-CDK4/6 activation.34
In the nucleus, FoxA1 facilitates AR chromatin access, exerting a key role in AR-driven transcription.35,36 Additionally, Fox1A is a downstream target of GATA-3 transcription factor. It was postulated that GATA-3 reduces chemotherapy responsiveness in HR negative BC, probably through AR signaling, supporting a direct correlation between AR-positivity and GATA-3 over-expression in LAR BC.37
The zinc-finger enhancer-binding protein 1 (ZEB1) is a transcription factor whose activation triggers the epithelial to mesenchymal transition (EMT), increasing tumor’s metastatic potential and predicting a poor prognosis. The AR-ZEB1 cross-talk is probably dependent on the direct binding of ZEB1 to the E-box sequence on the AR promoter. Indeed, ZEB1-knockdown cell lines present AR downregulation.38
Lastly, a role of non-coding RNAs (nc-RNAs) in LAR BC has been investigated, since these transcripts have many functional roles in regulating cancer processes, including proliferation, apoptosis, metastases and drug resistance.39,40 To this end, several studies demonstrated a cross-talk between AR and different nc-RNAs such as miR-363, let-7a, miR-30a, miR-100, miR-125, miR-328-3p, miR-204 and miR9-5p.41–47 Androgen receptor activities may be controlled in different ways by these specific nc-RNA. For instance, miR-100 and miR-125 block AR-mediated release of the metalloprotease 13 (MMP13),44 while Baldini et al reported that the miR-9-5p acts as a tumor suppressor in TNBC cell lines. The authors demonstrated that miR-9-5p resume AR-downstream signaling even in presence of AR-agonists.47 Studying the feedback loops between AR and nc-RNAs may provide hints to therapeutic targeting of AR in LAR BCs.
In conclusion, AR carries out heterogeneous functions through several mechanisms and interactions in BC. The plethora of AR intra-cellular cross-talks generate complex signaling networks, which may potentially serve as therapeutic targets in LAR BC patients.
Therapeutic Targets: Preclinical and Translational Evidences
Given the historical paucity of therapeutic options for TNBC patients, AR represented an attractive target for the treatment of LAR BC and its inhibition has been extensively studied in pre-clinical and translational models before being translated into clinical trials.8 Moreover, other biomarkers beyond AR have been identified and explored as potential therapeutic targets, but only some of them have been tested in the clinic so far. The preclinical and translational evidence about therapeutic targets in LAR BC are detailed below.
Androgen Receptor
The observation that synthetic androgens exert a tumorigenic effect in an AR positive TNBC cell line (ie MDA-MB-453 cell line) paved the way to early experiments with anti-androgen compounds in LAR BC models.48 One of the first report about the potential activity of anti-androgen bicalutamide in AR-positive TNBC dates back to Lehmann’s seminal work, in which the author described the antiproliferative effect of this compound in several LAR BC cell lines and xenograft models.5 Ensuing preclinical experiences proved the role of second generation anti-androgen enzalutamide in inhibiting the growth of LAR BC, including those with low AR expression.49,50 According to these evidences enzalutamide effect mainly relies on apoptosis stimulation.49 More recently, proxalutamide (GT0918), a newer second-generation anti-androgen, showed anti-proliferative effects in AR positive TNBC mouse xenograft models.51 All these compounds have been tested in clinical trials with inconsistent results.
PI3K/mTOR
In vitro experiments on a panel of LAR BCs cell lines showed frequent PI3K3CA activating mutations conferring sensitivity to the PI3K inhibitor dactolisib (NVP-BEZ235).5 This finding was confirmed by an ensuing study, were an enrichment of PIK3CA alterations was observed in AR positive TNBC cell line and xenograft models.52 In line with these observations, the simultaneous administration of the mTOR inhibitor rapamycin and enzalutamide exerted a potent antitumor activity in a LAR xenograft model.53 Overall, these data suggest a potential therapeutic role for the combination of AR antagonists with PI3K/mTOR inhibitors in LAR BC patients. More recently, Coussy et al investigated the efficacy of a mTOR-PIK3CA inhibitor (PF-04691502) in LAR TNBC patient-derived xenografts (PDX) models resistant to enzalutamide. They confirmed that PIK3CA and AKT1 mutations are frequent in this population and that the treatment with PF-04691502 may decrease tumor expansion in all models probably due to complete suppression of the PI3K pathway.54 According to this finding, PF-04691502 may represent a therapeutic option for LAR BC patients resistant to AR inhibitors who harbor PI3K/AKT signaling alterations.
BRCA1
It is known that BRCA1 mutated cells are sensitive to Poly[ADP-ribose] polymerase 1 (PARP-1) inhibition through a synthetic lethality mechanism.55 Based on preclinical data, the simultaneous administration of the AR inhibitor bicalutamide and of the PARP1 inhibitor veliparib (ABT-888) resulted in increased apoptosis in AR positive TNBC models and could therefore represent a viable therapeutic strategy.56
CDK4/6 and Cell Cycle
Therapeutic targeting of cyclin-dependent kinase 4/6 (CDK4/6) represented a major breakthrough in HR positive BC treatment57. According to preclinical evidence, LAR BC may also be sensitive to CDK4/6 inhibition. A single-cell analysis of in vitro and in vivo CDK2 activity in LAR BC showed a strong anticancer effect of the CDK4/6 inhibitor palbociclib. This effect was even stronger when the CDK4/6 inhibitor was combined with a PIK3CA inhibitor.33 Recently, Christenson et al investigated the effect of the dual CYP17-lyase inhibitor and AR antagonist seviteronel in combination with the CDK4/6 inhibitor abemaciclib in AR positive TNBC xenograft.58 The authors found a synergistic effect of the doublet compared to seviteronel or abemaciclib alone, due to the simultaneous inhibition of AR signaling and cell cycle pathway.58 In LAR BCs, cell cycle could also be tackled exploiting the feedback loops existing between AR and nc-RNAs. A bioinformatic analysis on four microarrays dataset identified a potential hsa-miR-3163-G2/mitotic-specific cyclin-B1 (CCNB1) regulatory axis in 44 normal breast tissues and 37 AR positive TNBC tissues.59 Previous study on prostate cancer models showed that AR is implicated in CCNB1 gene regulation through an interaction with E2F transcription factor, eventually influencing cell cycle progression.60 Accordingly, cyclin B1 may be a potential treatment target for AR positive TNBC.
BET
Bromodomain (BRD) and extra terminal domain (BET) family proteins modulate protein–protein interactions, promoting cell cycle progression, proliferation, and differentiation.61 These proteins interact with AR and may represent a therapeutic target in LAR BC.62 Indeed, using gene expression analysis and immunoblotting assays, Park et al demonstrated a potent anti-tumor effect of the BET-inhibitor JQ1 in AR positive TNBC cell lines. They also showed that JQ1 had a significant anti-tumor activity in TNBC xenograft mouse models either as monotherapy or in combination with anti-AR therapy.63
KDM1A
Preclinical studies investigated the functions of lysine (K)-specific demethylase 1A (KDM1A), a demethylase of mono- and di-methylated histone H3 lysine 4 in multiple cancer types including BC, demonstrating that high expression levels of this protein are associated with poor prognosis.64,65 KDM1A can act as a transcriptional activator or repressor to regulate target genes and its activity is critical for many cellular processes.66,67 The KDM1A inhibitor ORY-1001 has been tested in two human TNBC cell lines, where it restored apoptosis by inactivating AR expression and inhibited cells growth.68 Hence, KDM1A may represent an additional therapeutic target for AR positive TNBC.
TME
The tumor microenvironment (TME) is able to influence cancer development, progression, and treatment resistance.69 A specific TME profile has been associated to each TNBC subtype. In particular, luminal androgen receptor BCs are characterized by an immunosuppressive and pro-tumorigenic phenotype with expression of a stromal signature able to promote an immunosuppressed TME. These data suggest that LAR BC might be treated by targeting regulatory T-cells or immunosuppressive pathways including the adenosine pathway.70 For example, Kwilas et al reported that androgen deprivation with enzalutamide or abiraterone induced immunogenic modulation in AR positive TNBC cells by the expression of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and apoptosis associated genes.71 This evidence poses the basis for the combinatorial use of anti-androgen agents and immune checkpoint inhibitors (ICIs) in LAR BC.
Therapeutic Targets: Clinical Evidences
So far, AR has been considered the main therapeutic target for patients with LAR BC and it has been exploited in several clinical trials with controversial and inconsistent results (Table 1). Given these disappointing outcomes and based on the existing translational and preclinical evidence, many studies are currently ongoing in this population testing anti-androgen agents in combination with other compounds, aiming at increasing the efficacy of these therapeutic strategies (Figure 2) (Table 2). Updated evidence from clinical trials in LAR BC is summarized below.
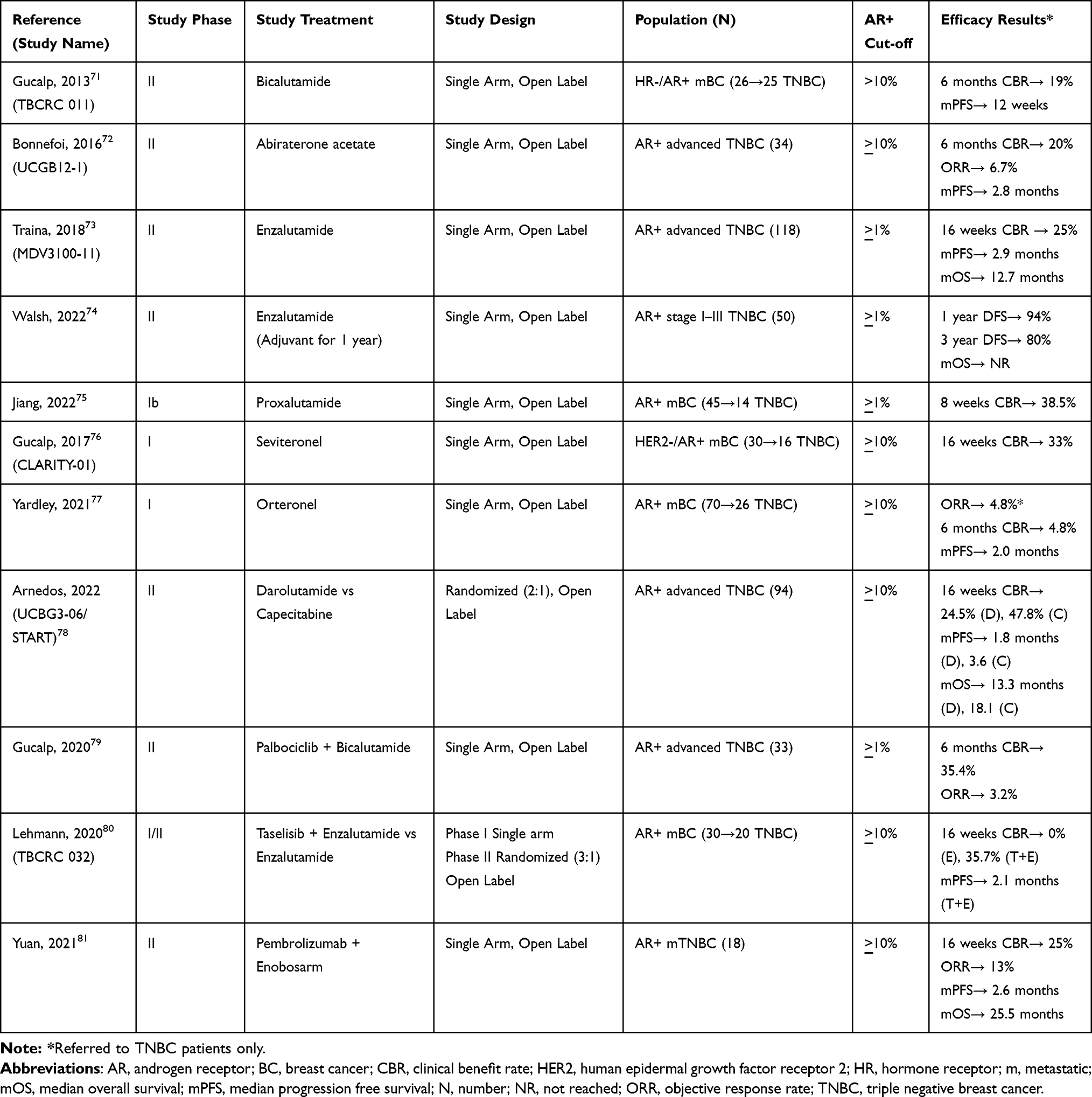 |
Table 1 Published Studies in Luminal Androgen Receptor Triple Negative Breast Cancer |
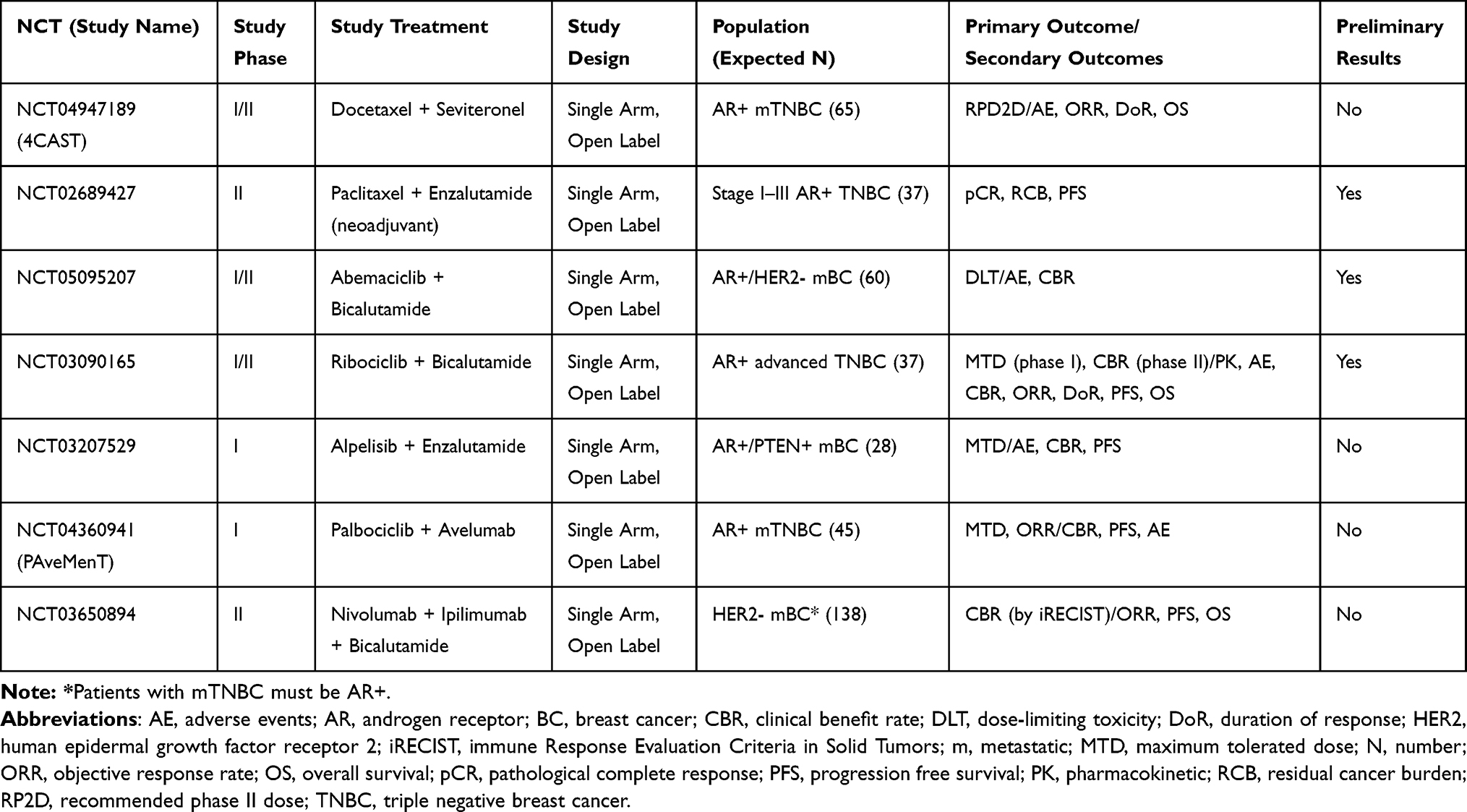 |
Table 2 Ongoing Studies in Luminal Androgen Receptor Triple Negative Breast Cancer |
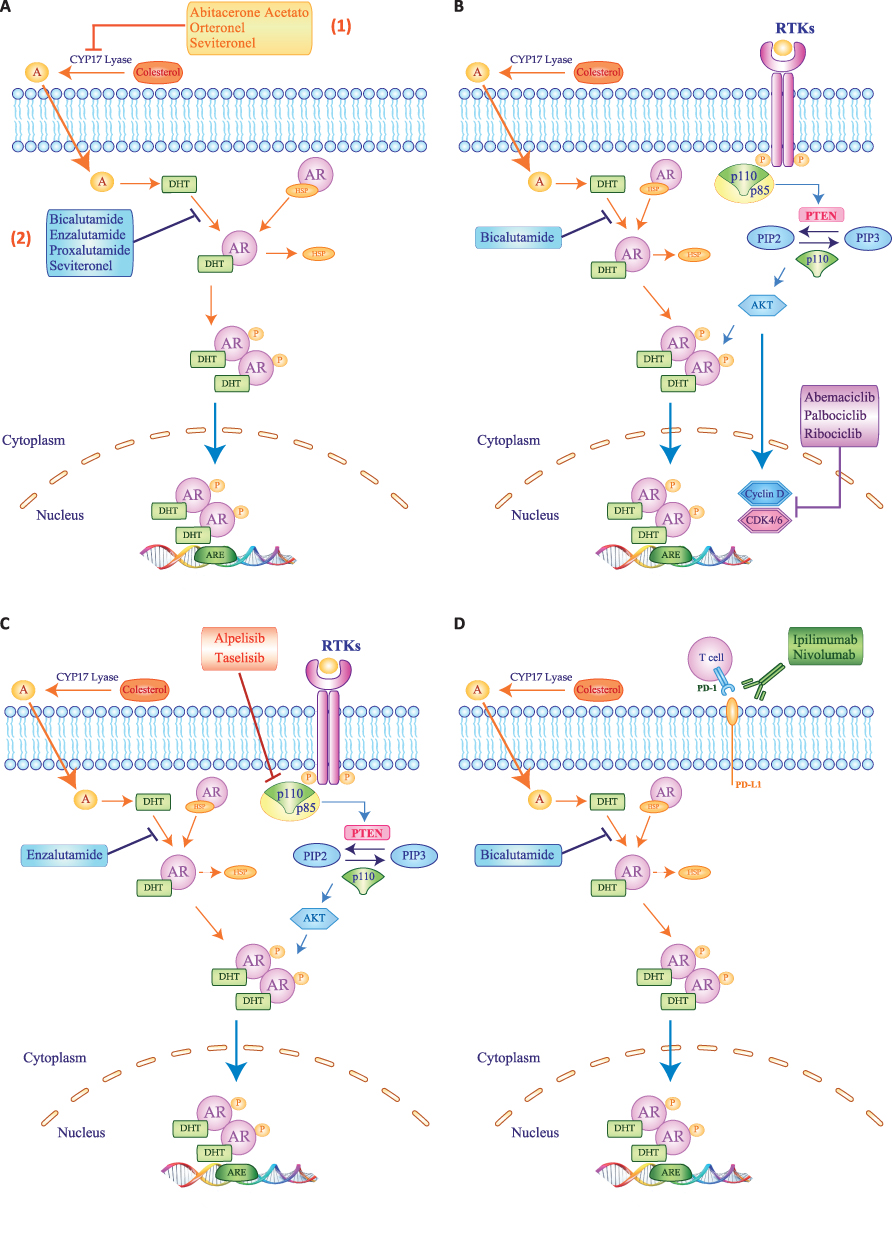 |
Figure 2 Schematic representation of therapeutic strategies used to inhibit Androgen Receptor signaling in LAR TNBC patients. (A) Androgen are produced from cholesterol and CYP17-lyase inhibitors, like Abiraterone Acetate, Orteronel and Seviteronel, inhibit the conversion of cholesterol in androgen (1). On the contrary, the antiandrogens bicalutamide, enzalutamide, proxalutamide or seviteronel block androgen binding to androgen receptor (AR) (2). Different treatment strategies currently combining antiandrogen therapy, such as bicalutamide or enzalutamide with CDK4/6 inhibitor Abemaciclib, Palbociclib or Ribociclib (B), or with PI3K inhibitors Alpelisib or Raselisib (C), or immunotherapy with Ipilimumab or Nivolumab (D). Blue, light blue and orange arrows represent signaling activation, while red, blue, purple and green bars depict inhibition of target signals. |
The anti-androgen agent bicalutamide was tested in 25 women with metastatic TNBC and AR expression >10% at immunohistochemistry (IHC). In this Phase II trial, the clinical benefit rate (CBR) at 6 months was 19% and the median progression-free survival (mPFS) was 12 weeks.72 Similar results emerged from a single-arm phase II trial of abiraterone acetate, a 17-[α]-hydroxylase/17,20-lyase (CYP17) inhibitor, in 34 patients with advanced LAR (AR>10%). In this study, 6-months CBR was 20%, with a dismal objective response rate (ORR) of 6.7% and a mPFS of 2.8 months.73 Consistently, in a larger cohort of LAR patients with AR expression >1% (n=118), monotherapy with enzalutamide provided a 25% CBR at 16 weeks while the attained mPFS was 2.9 months.74 In the adjuvant setting, enzalutamide has been administered for up to 1 year to 50 patients with stage I–III LAR, with >1% threshold for AR positivity. The 1, 2 and 3-year disease free survival (DFS) rates were 94%, 92% and 80%, respectively, while overall survival (OS) is still immature.75 Another phase Ib trial tested the anti-androgen proxalutamide in a Chinese cohort of mBC patients with AR>1%. Among these, 14 patients with LAR obtained an 8-weeks CBR of 38.5%.76 Seviteronel and orteronel are nonsteroidal inhibitors of 17, 20-lyase, which block estradiol and testosterone synthesis. In phase I studies enrolling subjects with advanced TNBC and AR>10%, seviteronel showed a 16-week CBR of 33% in 16 patients, while orteronel provided a disappointing 4.8% 6 CBR at 6 months and mPFS of 2 months in 26 patients.77,78 Recently, results of the first randomized phase II trial in patients with LAR BC have been presented. Patients with advanced TNBC and at least 10% expression of AR were assigned to receive the anti-androgen darolutamide (n=61) or capecitabine (n=33). Dorulatamide failed to show better outcomes in terms of 16 weeks CBR (24.5% in the experimental arm versus 47.8% in the control arm) and mPFS (1.8 months in the experimental arm versus 3.6 months in the control arm).79
Given the scarce activity of anti-androgen monotherapy, different combination strategies have been proposed. For instance, ongoing trials are evaluating enzalutamide plus paclitaxel in the neo-adjuvant phase (NCT02689427) and seviteronel plus docetaxel in the advanced setting (NCT04947189). Preliminary efficacy results are available for the combination of the CDK4/6 inhibitor palbociclib and bicalutamide in patients with AR positive (>1%) TNBC, showing a 33% CBR at 6 months.80 Early phase trials are also ongoing combining abemaciclib or ribociclib with bicalutamide in the same subset of patients (NCT05095207, NCT03090165). Simultaneous inhibition of androgen receptor and phosphatidyl-inositol-3-kinase (PI3K) has been addressed in a phase I/II study testing enzalutamide and taselisib. Twenty patients with metastatic LAR were included in this trial, attaining a 16 weeks CBR of 35.7% and a mPFS of 2.1 months.81 Another trial is currently evaluating enzalutamide with the alfa-subunit selective PI3K inhibitor alpelisib (NCT03207529).
Finally, the immune-checkpoint inhibitor pembrolizumab was combined with the androgen receptor modulator enobosarm in patients with >10% AR positive metastatic TNBC. The observed 16-week CBR was 25.5% with an ORR of 13%, a mPFS of 2.6 months and mOS of 25 months.82 Other ongoing trials are currently enrolling patients with LAR to test immunotherapy combination strategies, such as palbociclib plus avelumab (NCT04360941) or bicalutamide plus nivolumab and ipilimumab (NCT03650894).
Discussion and Conclusion
In last decades, improved understanding of cancer biology led to the discovery of several actionable targets and enabled the development of new treatments that ultimately paved the way to personalized medicine.83 This paradigm shift from “one size fits all” to “to each his own” involved almost every tumor type, including BC.84 However, while several targeted options have been developed for HR positive and HER2 positive tumors in the last decades, TNBC has been considered an undruggable subtype for a long time.85 The seminal works by Lehman and Burstein dissected TNBC molecular profile, unravelling its profound heterogeneity and defining a new and unique entity of LAR BC.5,6 Since then, a plethora of studies focused on this BC subtype, aiming to clarify the role of AR in disease pathogenesis, progression, prognosis and response to therapies. Overall, AR seems to exert a pleiotropic role in BC cells, as confirmed by the plethora of networks, which are able to establish with different mediators.8 Beyond the mechanistic evidence about AR functioning, many researches focused on the potential role of this receptor as a druggable target, taking also into account decades of successful AR inhibition in prostate cancer.20,86 Indeed, preclinical evidence showed encouraging signs of activity of different anti-androgen agents (bicalutamide, enzalutamide and proxalutamide) in LAR BC cell lines and xenograft models,5,14,49–51 but these results did not translate into significant clinical activity. In clinical trials enrolling advanced LAR BC patients, monotherapy with an antiandrogen agent provided a disappointing mPFS of about 2–3 months and in a head-to-head comparison capecitabine outperformed darolutamide in terms of both CBR and mPFS.72–74,79 Overall, blocking AR alone does not seem to exert a clinically relevant anti-tumor effect in LAR BC patients, and AR overexpression should probably be considered a diagnostic biomarker rather than a therapeutic target for this BC subtype.
The reasons for the discrepancy between the preclinical and clinical activity of antiandrogen agents in LAR BC are still unknown. A potential explanation may be the lack of standardization in AR detection methods, both in terms of positivity thresholds and antibodies staining.87 Additionally, the presence of AR splice variants (AR-V), such as AR-V7, may determine primary resistance to anti-androgen agents, as already shown in prostate cancer.88,89 Also, AR involvement into several intracellular networks may foster escape mechanisms, which eventually determine resistance to anti-androgen monotherapy. The pleiotropic role of AR in BC cells can be a cause of treatment resistance but may also be exploited for treatment purposes, as shown by several combinatorial approaches developed so far.33,53,71 While some combinations, such as anti-androgen agents plus PI3K/mToR inhibitors, CDK4/6 inhibitors or immunotherapeutic agents, display promising results in the preclinical and translational contexts, confirmation from ongoing clinical trials is still needed.
Several other questions about the management of LAR BC patients remain open, including whether AR should be routinely evaluated in TNBC and with which method and threshold. Additionally, it is debated whether newer anti-androgen compounds should be tested as well as which combination strategies deserve further development in clinical trials. In order to address all these controversies, further researches are needed with the ultimate goal to improve the outcomes of patients with LAR BC.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Prat A, Perou CM. Deconstructing the molecular portraits of breast cancer. Mol Oncol. 2011;5(1):5–23. doi:10.1016/j.molonc.2010.11.003
2. Perou CM, Sorlie T, Eisen MB, et al. Molecular portraits of human breast tumours. Nature. 2000;406(6797):747–752. doi:10.1038/35021093
3. Eliyatkin N, Yalcin E, Zengel B, Aktas S, Vardar E. Molecular classification of breast carcinoma: from traditional, old-fashioned way to a new age, and a new way. J Breast Health. 2015;11(2):59–66. doi:10.5152/tjbh.2015.1669
4. Foulkes WD, Smith IE, Reis-Filho JS. Triple-negative breast cancer. N Engl J Med. 2010;363(20):1938–1948. doi:10.1056/NEJMra1001389
5. Lehmann BD, Bauer JA, Chen X, et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest. 2011;121(7):2750–2767. doi:10.1172/JCI45014
6. Burstein MD, Tsimelzon A, Poage GM, et al. Comprehensive genomic analysis identifies novel subtypes and targets of triple-negative breast cancer. Clin Cancer Res. 2015;21(7):1688–1698. doi:10.1158/1078-0432.CCR-14-0432
7. Zhao S, Ma D, Xiao Y, et al. Molecular subtyping of triple-negative breast cancers by immunohistochemistry: molecular basis and clinical relevance. Oncologist. 2020;25(10):e1481–e1491. doi:10.1634/theoncologist.2019-0982
8. Gerratana L, Basile D, Buono G, et al. Androgen receptor in triple negative breast cancer: a potential target for the targetless subtype. Cancer Treat Rev. 2018;68:102–110. doi:10.1016/j.ctrv.2018.06.005
9. Garrido-Castro AC, Lin NU, Polyak K. Insights into molecular classifications of triple-negative breast cancer: improving patient selection for treatment. Cancer Discov. 2019;9(2):176–198. doi:10.1158/2159-8290.CD-18-1177
10. Stella S, Vitale SR, Massimino M, et al. Molecular analysis of luminal androgen receptor reveals activated pathways and potential therapeutic targets in breast cancer. Cancer Genomics Proteomics. 2022;19(4):464–476. doi:10.21873/cgp.20333
11. Giovannelli P, Di Donato M, Auricchio F, Castoria G, Migliaccio A. Androgens induce invasiveness of triple negative breast cancer cells through AR/Src/PI3-K complex assembly. Sci Rep. 2019;9(1):4490. doi:10.1038/s41598-019-41016-4
12. Giovannelli P, Di Donato M, Galasso G, Di Zazzo E, Bilancio A, Migliaccio A. The androgen receptor in breast cancer. Front Endocrinol. 2018;9:492. doi:10.3389/fendo.2018.00492
13. Hickey TE, Robinson JL, Carroll JS, Tilley WD. Minireview: the androgen receptor in breast tissues: growth inhibitor, tumor suppressor, oncogene? Mol Endocrinol. 2012;26(8):1252–1267. doi:10.1210/me.2012-1107
14. Barton VN, D’Amato NC, Gordon MA, Christenson JL, Elias A, Richer JK. Androgen receptor biology in triple negative breast cancer: a case for classification as AR+ or quadruple negative disease. Horm Cancer. 2015;6(5–6):206–213. doi:10.1007/s12672-015-0232-3
15. Davey RA, Grossmann M. Androgen receptor structure, function and biology: from bench to bedside. Clin Biochem Rev. 2016;37(1):3–15.
16. Cuenca-Lopez MD, Montero JC, Morales JC, Prat A, Pandiella A, Ocana A. Phospho-kinase profile of triple negative breast cancer and androgen receptor signaling. BMC Cancer. 2014;14:302. doi:10.1186/1471-2407-14-302
17. Garay JP, Karakas B, Abukhdeir AM, et al. The growth response to androgen receptor signaling in ERalpha-negative human breast cells is dependent on p21 and mediated by MAPK activation. Breast Cancer Res. 2012;14(1):R27. doi:10.1186/bcr3112
18. Tarulli GA, Butler LM, Tilley WD, Hickey TE. Bringing androgens up a NOTCH in breast cancer. Endocr Relat Cancer. 2014;21(4):T183–T202. doi:10.1530/ERC-14-0248
19. Ueda T, Bruchovsky N, Sadar MD. Activation of the androgen receptor N-terminal domain by interleukin-6 via MAPK and STAT3 signal transduction pathways. J Biol Chem. 2002;277(9):7076–7085. doi:10.1074/jbc.M108255200
20. Pietri E, Conteduca V, Andreis D, et al. Androgen receptor signaling pathways as a target for breast cancer treatment. Endocr Relat Cancer. 2016;23(10):R485–R498. doi:10.1530/ERC-16-0190
21. Chia KM, Liu J, Francis GD, Naderi A. A feedback loop between androgen receptor and ERK signaling in estrogen receptor-negative breast cancer. Neoplasia. 2011;13(2):154–166. doi:10.1593/neo.101324
22. Simoncini T, Genazzani AR. Non-genomic actions of sex steroid hormones. Eur J Endocrinol. 2003;148(3):281–292. doi:10.1530/eje.0.1480281
23. Li J, Al-Azzawi F. Mechanism of androgen receptor action. Maturitas. 2009;63(2):142–148. doi:10.1016/j.maturitas.2009.03.008
24. Anestis A, Karamouzis MV, Dalagiorgou G, Papavassiliou AG. Is androgen receptor targeting an emerging treatment strategy for triple negative breast cancer? Cancer Treat Rev. 2015;41(6):547–553. doi:10.1016/j.ctrv.2015.04.009
25. Rizza P, Barone I, Zito D, et al. Estrogen receptor beta as a novel target of androgen receptor action in breast cancer cell lines. Breast Cancer Res. 2014;16(1):R21. doi:10.1186/bcr3619
26. Dey P, Wang A, Ziegler Y, et al. Estrogen receptor beta 1: a potential therapeutic target for female triple negative breast cancer. Endocrinology. 2022;163(12):12. doi:10.1210/endocr/bqac172
27. Monaco A, Licitra F, Di Gisi M, et al. ERβ in triple-negative breast cancer: emerging concepts and therapeutic possibilities. Endocrines. 2021;2(3):356–365. doi:10.3390/endocrines2030033
28. Naderi A, Hughes-Davies L. A functionally significant cross-talk between androgen receptor and ErbB2 pathways in estrogen receptor negative breast cancer. Neoplasia. 2008;10(6):542–548. doi:10.1593/neo.08274
29. Naderi A, Chia KM, Liu J. Synergy between inhibitors of androgen receptor and MEK has therapeutic implications in estrogen receptor-negative breast cancer. Breast Cancer Res. 2011;13(2):R36. doi:10.1186/bcr2858
30. Park JJ, Irvine RA, Buchanan G, et al. Breast cancer susceptibility gene 1 (BRCAI) is a coactivator of the androgen receptor. Cancer Res. 2000;60(21):5946–5949.
31. Zhang W, Luo J, Yang F, et al. BRCA1 inhibits AR-mediated proliferation of breast cancer cells through the activation of SIRT1. Sci Rep. 2016;6:22034. doi:10.1038/srep22034
32. Yeh S, Hu YC, Rahman M, et al. Increase of androgen-induced cell death and androgen receptor transactivation by BRCA1 in prostate cancer cells. Proc Natl Acad Sci U S A. 2000;97(21):11256–11261. doi:10.1073/pnas.190353897
33. Asghar US, Barr AR, Cutts R, et al. Single-cell dynamics determines response to CDK4/6 inhibition in triple-negative breast cancer. Clin Cancer Res. 2017;23(18):5561–5572. doi:10.1158/1078-0432.CCR-17-0369
34. Michmerhuizen AR, Spratt DE, Pierce LJ, Speers CW. ARe we there yet? Understanding androgen receptor signaling in breast cancer. NPJ Breast Cancer. 2020;6:47. doi:10.1038/s41523-020-00190-9
35. Robinson JL, Macarthur S, Ross-Innes CS, et al. Androgen receptor driven transcription in molecular apocrine breast cancer is mediated by FoxA1. EMBO J. 2011;30(15):3019–3027. doi:10.1038/emboj.2011.216
36. Robinson JL, Carroll JS. FoxA1 is a key mediator of hormonal response in breast and prostate cancer. Front Endocrinol. 2012;3:68. doi:10.3389/fendo.2012.00068
37. Kim S, Moon BI, Lim W, Park S, Cho MS, Sung SH. Expression patterns of GATA3 and the androgen receptor are strongly correlated in patients with triple-negative breast cancer. Hum Pathol. 2016;55:190–195. doi:10.1016/j.humpath.2016.04.013
38. Graham TR, Yacoub R, Taliaferro-Smith L, et al. Reciprocal regulation of ZEB1 and AR in triple negative breast cancer cells. Breast Cancer Res Treat. 2010;123(1):139–147. doi:10.1007/s10549-009-0623-7
39. Garofoli M, Volpicella M, Guida M, Porcelli L, Azzariti A. The role of non-coding RNAs as prognostic factor, predictor of drug response or resistance and pharmacological targets, in the cutaneous squamous cell carcinoma. Cancers. 2020;12(9):2552. doi:10.3390/cancers12092552
40. Ravaioli S, Maltoni R, Pasculli B, et al. Androgen receptor in breast cancer: the “5W” questions. Front Endocrinol. 2022;13:977331. doi:10.3389/fendo.2022.977331
41. Nakano K, Miki Y, Hata S, et al. Identification of androgen-responsive microRNAs and androgen-related genes in breast cancer. Anticancer Res. 2013;33(11):4811–4819.
42. Zhang W, Liu X, Liu S, et al. Androgen receptor/let-7a signaling regulates breast tumor-initiating cells. Oncotarget. 2018;9(3):3690–3703. doi:10.18632/oncotarget.23196
43. Lyu S, Liu H, Liu X, et al. Interrelation of androgen receptor and miR-30a and miR-30a function in ER(-), PR(-), AR(+) MDA-MB-453 breast cancer cells. Oncol Lett. 2017;14(4):4930–4936. doi:10.3892/ol.2017.6781
44. Ahram M, Mustafa E, Zaza R, et al. Differential expression and androgen regulation of microRNAs and metalloprotease 13 in breast cancer cells. Cell Biol Int. 2017;41(12):1345–1355. doi:10.1002/cbin.10841
45. Al-Othman N, Hammad H, Ahram M. Dihydrotestosterone regulates expression of CD44 via miR-328-3p in triple-negative breast cancer cells. Gene. 2018;675:128–135. doi:10.1016/j.gene.2018.06.094
46. Yang F, Shen Y, Zhang W, et al. An androgen receptor negatively induced long non-coding RNA ARNILA binding to miR-204 promotes the invasion and metastasis of triple-negative breast cancer. Cell Death Differ. 2018;25(12):2209–2220. doi:10.1038/s41418-018-0123-6
47. Bandini E, Fanini F, Vannini I, et al. miR-9-5p as a regulator of the androgen receptor pathway in breast cancer cell lines. Front Cell Dev Biol. 2020;8:579160. doi:10.3389/fcell.2020.579160
48. Doane AS, Danso M, Lal P, et al. An estrogen receptor-negative breast cancer subset characterized by a hormonally regulated transcriptional program and response to androgen. Oncogene. 2006;25(28):3994–4008. doi:10.1038/sj.onc.1209415
49. Cochrane DR, Bernales S, Jacobsen BM, et al. Role of the androgen receptor in breast cancer and preclinical analysis of enzalutamide. Breast Cancer Res. 2014;16(1):R7. doi:10.1186/bcr3599
50. Barton VN, D’Amato NC, Gordon MA, et al. Multiple molecular subtypes of triple-negative breast cancer critically rely on androgen receptor and respond to enzalutamide in vivo. Mol Cancer Ther. 2015;14(3):769–778. doi:10.1158/1535-7163.MCT-14-0926
51. Li H, Song G, Zhou Q, et al. Activity of preclinical and phase I clinical trial of a novel androgen receptor antagonist GT0918 in metastatic breast cancer. Breast Cancer Res Treat. 2021;189(3):725–736. doi:10.1007/s10549-021-06345-x
52. Lehmann BD, Bauer JA, Schafer JM, et al. PIK3CA mutations in androgen receptor-positive triple negative breast cancer confer sensitivity to the combination of PI3K and androgen receptor inhibitors. Breast Cancer Res. 2014;16(4):406. doi:10.1186/s13058-014-0406-x
53. Robles AJ, Cai S, Cichewicz RH, Mooberry SL. Selective activity of deguelin identifies therapeutic targets for androgen receptor-positive breast cancer. Breast Cancer Res Treat. 2016;157(3):475–488. doi:10.1007/s10549-016-3841-9
54. Coussy F, Lavigne M, de Koning L, et al. Response to mTOR and PI3K inhibitors in enzalutamide-resistant luminal androgen receptor triple-negative breast cancer patient-derived xenografts. Theranostics. 2020;10(4):1531–1543. doi:10.7150/thno.36182
55. Genta S, Martorana F, Stathis A, Colombo I. Targeting the DNA damage response: PARP inhibitors and new perspectives in the landscape of cancer treatment. Crit Rev Oncol Hematol. 2021;168:103539. doi:10.1016/j.critrevonc.2021.103539
56. Luo J, Jin J, Yang F, et al. The correlation between PARP1 and BRCA1 in AR positive triple-negative breast cancer. Int J Biol Sci. 2016;12(12):1500–1510. doi:10.7150/ijbs.16176
57. Spring LM, Wander SA, Andre F, Moy B, Turner NC, Bardia A. Cyclin-dependent kinase 4 and 6 inhibitors for hormone receptor-positive breast cancer: past, present, and future. Lancet. 2020;395(10226):817–827. doi:10.1016/S0140-6736(20)30165-3
58. Christenson JL, O’Neill KI, Williams MM, et al. Activity of combined androgen receptor antagonism and cell cycle inhibition in androgen receptor positive triple negative breast cancer. Mol Cancer Ther. 2021;20(6):1062–1071. doi:10.1158/1535-7163.MCT-20-0807
59. Qiu P, Guo Q, Yao Q, Chen J, Lin J, Chang S. Hsa-mir-3163 and CCNB1 may be potential biomarkers and therapeutic targets for androgen receptor positive triple-negative breast cancer. PLoS One. 2021;16(11):e0254283. doi:10.1371/journal.pone.0254283
60. Li Y, Zhang DY, Ren Q, et al. Regulation of a novel androgen receptor target gene, the cyclin B1 gene, through androgen-dependent E2F family member switching. Mol Cell Biol. 2012;32(13):2454–2466. doi:10.1128/MCB.06663-11
61. Fu LL, Tian M, Li X, et al. Inhibition of BET bromodomains as a therapeutic strategy for cancer drug discovery. Oncotarget. 2015;6(8):5501–5516. doi:10.18632/oncotarget.3551
62. Urbanucci A, Barfeld SJ, Kytola V, et al. Androgen receptor deregulation drives bromodomain-mediated chromatin alterations in prostate cancer. Cell Rep. 2017;19(10):2045–2059. doi:10.1016/j.celrep.2017.05.049
63. Park IH, Yang HN, Jeon SY, et al. Anti-tumor activity of BET inhibitors in androgen-receptor-expressing triple-negative breast cancer. Sci Rep. 2019;9(1):13305. doi:10.1038/s41598-019-49366-9
64. Huang M, Chen C, Geng J, et al. Targeting KDM1A attenuates Wnt/beta-catenin signaling pathway to eliminate sorafenib-resistant stem-like cells in hepatocellular carcinoma. Cancer Lett. 2017;398:12–21. doi:10.1016/j.canlet.2017.03.038
65. Maiques-Diaz A, Somervaille TC. LSD1: biologic roles and therapeutic targeting. Epigenomics. 2016;8(8):1103–1116. doi:10.2217/epi-2016-0009
66. Jin Y, Ma D, Gramyk T, et al. Kdm1a promotes SCLC progression by transcriptionally silencing the tumor suppressor rest. Biochem Biophys Res Commun. 2019;515(1):214–221. doi:10.1016/j.bbrc.2019.05.118
67. Shi Y, Lan F, Matson C, et al. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell. 2004;119(7):941–953. doi:10.1016/j.cell.2004.12.012
68. Wang T, Zhang F, Sun F. ORY-1001, a KDM1A inhibitor, inhibits proliferation, and promotes apoptosis of triple negative breast cancer cells by inactivating androgen receptor. Drug Dev Res. 2022;83(1):208–216. doi:10.1002/ddr.21860
69. Baghban R, Roshangar L, Jahanban-Esfahlan R, et al. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun Signal. 2020;18(1):59. doi:10.1186/s12964-020-0530-4
70. Bareche Y, Buisseret L, Gruosso T, et al. Unraveling triple-negative breast cancer tumor microenvironment heterogeneity: towards an optimized treatment approach. J Natl Cancer Inst. 2020;112(7):708–719. doi:10.1093/jnci/djz208
71. Kwilas AR, Ardiani A, Gameiro SR, Richards J, Hall AB, Hodge JW. Androgen deprivation therapy sensitizes triple negative breast cancer cells to immune-mediated lysis through androgen receptor independent modulation of osteoprotegerin. Oncotarget. 2016;7(17):23498–23511. doi:10.18632/oncotarget.8274
72. Gucalp A, Tolaney S, Isakoff SJ, et al. Phase II trial of bicalutamide in patients with androgen receptor-positive, estrogen receptor-negative metastatic breast cancer. Clin Cancer Res. 2013;19(19):5505–5512. doi:10.1158/1078-0432.CCR-12-3327
73. Bonnefoi H, Grellety T, Tredan O, et al. A phase II trial of abiraterone acetate plus prednisone in patients with triple-negative androgen receptor positive locally advanced or metastatic breast cancer (UCBG 12-1). Ann Oncol. 2016;27(5):812–818. doi:10.1093/annonc/mdw067
74. Traina TA, Miller K, Yardley DA, et al. Enzalutamide for the treatment of androgen receptor-expressing triple-negative breast cancer. J Clin Oncol. 2018;36(9):884–890. doi:10.1200/JCO.2016.71.3495
75. Walsh EM, Gucalp A, Patil S, et al. Adjuvant enzalutamide for the treatment of early-stage androgen-receptor positive, triple-negative breast cancer: a feasibility study. Breast Cancer Res Treat. 2022;195(3):341–351. doi:10.1007/s10549-022-06669-2
76. Jiang H, Ouyang Q, Yin Y, et al. Proxalutamide in patients with AR-positive metastatic breast cancer: results from an open-label multicentre phase Ib study and biomarker analysis. Eur J Cancer. 2022;176:1–12. doi:10.1016/j.ejca.2022.08.025
77. Gucalp A, Danso MA, Elias AD, et al. Phase (Ph) 2 stage 1 clinical activity of seviteronel, a selective CYP17-lyase and androgen receptor (AR) inhibitor, in women with advanced AR+ triple-negative breast cancer (TNBC) or estrogen receptor (ER)+ BC: CLARITY-01. J Clin Oncol. 2017;35(15_suppl):1102. doi:10.1200/JCO.2017.35.15_suppl.1102
78. Yardley D, Peacock N, Young R, et al. Abstract P5-14-04: a Phase 2 study evaluating orteronel, an inhibitor of androgen biosynthesis, in patients with androgen receptor (AR)-expressing metastatic breast cancer: interim analysis. Cancer Res. 2016;76(4_Supplement):P5-14-04-P15-14–04. doi:10.1158/1538-7445.SABCS15-P5-14-04
79. Arnedos M, Goncalves A, Pulido M, et al. 213MO primary endpoint analysis of a randomized phase II of darolutamide or capecitabine in patients with triple-negative androgen receptor-positive advanced breast cancer (UCBG3-06 START trial). Ann Oncol. 2022;33:S635. doi:10.1016/j.annonc.2022.07.252
80. Gucalp A, Boyle LA, Alano T, et al. Phase II trial of bicalutamide in combination with palbociclib for the treatment of androgen receptor (+) metastatic breast cancer. J Clin Oncol. 2020;38(15_suppl):1017. doi:10.1200/JCO.2020.38.15_suppl.1017
81. Lehmann BD, Abramson VG, Sanders ME, et al. TBCRC 032 IB/II multicenter study: molecular insights to AR antagonist and PI3K inhibitor efficacy in patients with AR(+) metastatic triple-negative breast cancer. Clin Cancer Res. 2020;26(9):2111–2123. doi:10.1158/1078-0432.CCR-19-2170
82. Yuan Y, Lee JS, Yost SE, et al. A phase II clinical trial of pembrolizumab and enobosarm in patients with androgen receptor-positive metastatic triple-negative breast cancer. Oncologist. 2021;26(2):99–e217. doi:10.1002/onco.13583
83. Chang L, Ruiz P, Ito T, Sellers WR. Targeting pan-essential genes in cancer: challenges and opportunities. Cancer Cell. 2021;39(4):466–479. doi:10.1016/j.ccell.2020.12.008
84. Jacobs AT, Martinez Castaneda-Cruz D, Rose MM, Connelly L. Targeted therapy for breast cancer: an overview of drug classes and outcomes. Biochem Pharmacol. 2022;204:115209. doi:10.1016/j.bcp.2022.115209
85. Bianchini G, De Angelis C, Licata L, Gianni L. Treatment landscape of triple-negative breast cancer - expanded options, evolving needs. Nat Rev Clin Oncol. 2022;19(2):91–113. doi:10.1038/s41571-021-00565-2
86. Aurilio G, Cimadamore A, Mazzucchelli R, et al. Androgen receptor signaling pathway in prostate cancer: from genetics to clinical applications. Cells. 2020;9(12):12. doi:10.3390/cells9122653
87. You CP, Tsoi H, Man EPS, Leung MH, Khoo US. Modulating the activity of androgen receptor for treating breast cancer. Int J Mol Sci. 2022;23(23):15342. doi:10.3390/ijms232315342
88. Antonarakis ES, Lu C, Wang H, et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med. 2014;371(11):1028–1038. doi:10.1056/NEJMoa1315815
89. Ferguson DC, Mata DA, Tay TK, et al. Androgen receptor splice variant-7 in breast cancer: clinical and pathologic correlations. Mod Pathol. 2022;35(3):396–402. doi:10.1038/s41379-021-00924-5
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.