")
Back to Journals » Drug Design, Development and Therapy » Volume 13
Potential therapeutic effects of Cordyceps cicadae and Paecilomyces cicadae on adenine-induced chronic renal failure in rats and their phytochemical analysis
Authors Li L , Zhang T, Li C, Xie L , Li N, Hou T, Wang Y, Wang B
Received 17 July 2018
Accepted for publication 27 September 2018
Published 19 December 2018 Volume 2019:13 Pages 103—117
DOI https://doi.org/10.2147/DDDT.S180543
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Ling Li,1,* Tong Zhang,1,* Chunru Li,2 Lu Xie,3 Ning Li,4 Tianling Hou,1 Yuqin Wang,2 Bing Wang1
1Experiment Center for Teaching and Learning, Shanghai University of Traditional Chinese Medicine, Shanghai, China; 2Zhejiang BioAsia Pharmaceutical Co., Ltd., Pinghu, Zhejiang, China; 3Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, China; 4Institute of Chinese Materia Medica, Shanghai University of Traditional Chinese Medicine, Shanghai, China
*These authors contributed equally to this work
Background: Natural Cordyceps cicadae (C. cicadae) has been utilized extensively in traditional Chinese medicine to treat chronic renal diseases, heart palpitations, infantile convulsions, and dizziness. However, given its slow growth and immoderate exploitation, C. cicadae resources have been severely depleted. By contrast, Paecilomyces cicadae (P. cicadae), as the anamorph stage of C. cicadae, is easy to cultivate, and this kind of cultivated P. cicadae has good and controllable quality.
Purpose: This study aimed to compare the therapeutic effects of C. cicadae and P. cicadae on adenine-induced chronic renal failure (CRF) rats. In accordance with the aforementioned studies, our work subsequently analyzed the intrinsic relationships between the efficacy and pharmacodynamic substances of C. cicadae and P. cicadae to conclude whether or not P. cicadae could be used as an alternative to C. cicadae in treating CRF.
Methods: Rats were administered with C. cicadae (1.0 g/kg) or P. cicadae (1.0 g/kg) by gavage for 4 weeks. Furthermore, we applied Fourier transform infrared spectroscopy, gas chromatography–mass spectrometry, liquid chromatography–tandem mass spectrometry, and ultraviolet spectrophotometry to comprehensively detect and analyze the chemical constituent differences from ten batches each of C. cicadae and P. cicadae.
Results: This study revealed that both C. cicadae and P. cicadae exerted obvious therapeutic effects on CRF and were more consistent with their chemical compositions.
Conclusion: P. cicadae can be used as an alternative to C. cicadae for treating CRF to cater to market demands.
Keywords: Cordyceps cicadae, Paecilomyces cicadae, chronic renal failure, FTIR, GC-MS, LC-MS/MS, identification
Introduction
Natural Cordyceps cicadae (C. cicadae) of the family Clavicipitaceae and order Hypocreales is a kind of fungus that parasitizes Lepidoptera larvae.1 C. cicadae, also known as Isaria cicadae and C. cicadae Shing,2 has been used in traditional Chinese medicine (TCM) for over 1,000 years.3 The fungus contains biologically active chemical substances, such as polysaccharides,4 nucleosides,5 cyclopeptides,6 myriocin,7 inorganic elements,8 and amino acids.9 Modern pharmacological studies have shown that C. cicadae exerts active functions, including liver and kidney protection, blood fat reduction, analgesia–antipyresis, anticonvulsion, and antitumor activities.1 C. cicadae is mainly used for chronic renal diseases,10 palpitations, infantile convulsions, and dizziness in clinics.11 Given the great attention for health care and the desirable medical value of C. cicadae, the demand for C. cicadae has grown rapidly in recent years. However, C. cicadae propagates slowly and lacks resistance; thus, the supply is insufficient to meet the demand. Moreover, because of its immoderate exploitation, Cordyceps resources are severely depleted.12,13 By contrast, as an entomopathogenic genus of the family Clavicipitaceae, Paecilomyces cicadae (P. cicadae) is thought to be the anamorph of C. cicadae,14 which was obtained by artificial fermentation technologies. P. cicadae has great applications in TCM and functional foods with abundant resources.15 Ren et al successfully developed a method to enhance the yield to stabilize and further control the quality of P. cicadae.16 Chyau et al found that P. cicadae could protect renal tubular cells against Cyclosporine A-induced apoptosis and improve urea and creatinine clearance.17 Previous studies indicated that polysaccharides obtained from P. cicadae presented notable health effects and pharmacological properties such as improvement of renal function and inhibition of oxygen radicals. As shown by in vivo or in vitro experiments,18,19 it means that not only C. cicadae but also P. cicadae can protect the kidneys. Given the complexity of the chemical compositions and mechanisms of TCM, whether or not can P. cicadae replace C. cicadae remains unknown.
The present study aimed to compare the therapeutic effects between C. cicadae and P. cicadae on rats with chronic renal failure (CRF). This work also intended to determine the differences in chemical constituents between the two species, track the intrinsic relationships between the efficacy and constituent pharmacodynamic substances, and search for methods for rapidly identifying C. cicadae and P. cicadae (Figure 1). Recently, many studies have shown significant clinical effects of C. cicadae on CRF.10,20 Moreover, polysaccharides,21 cordycepin, adenosine, and inosine22 in C. cicadae are effective for renal protection, specifically reducing the inflammatory response and kidney damage. Given the abovementioned information, our experiments chose CRF models to explore whether C. cicadae and P. cicadae have the same effect or not. Uremic clearance granule (UCG) was demonstrated to protect the glomerular endothelial cells and prevent glomerulosclerosis and interstitial fibrosis, moreover, stabilize renal function and repair renal pathological changes.23,24 So, the experiments were performed on rats with adenine-induced CRF against UCG as positive control.
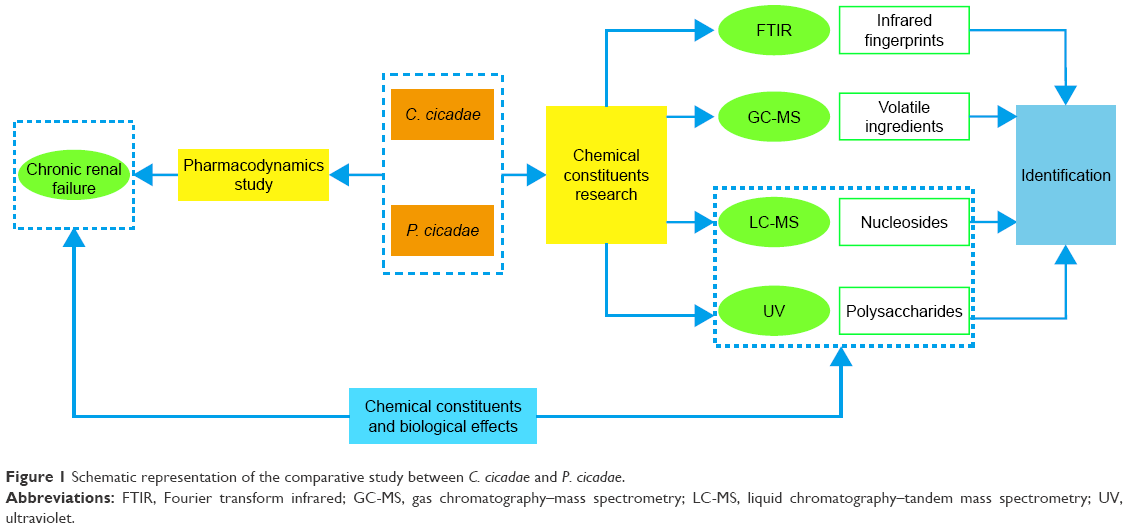 | Figure 1 Schematic representation of the comparative study between C. cicadae and P. cicadae. |
At present, many methods are available for the qualitative and quantitative analysis of C. cicadae. Examples of these techniques include phenol-sulfuric acid method combined with ultraviolet (UV) spectrophotometry for detecting polysaccharides,25 HPLC-diode array detector for simultaneously testing eight nucleosides,26 gas chromatography–mass spectrometry (GC-MS) for determining the saturated fatty acid content,27 HPLC-variable wavelength detector for obtaining the cordycepic acid content,28 capillary gas chromatography for acquiring the squalene content,29 and atomic absorption spectrometry for determining the microelemental content.9 To date, works on C. cicadae and P. cicadae have mainly focused on comparing between single components or certain chemical compositions. A comprehensive comparative analysis has not yet been conducted. GC-MS was performed to determine the volatile constituents, liquid chromatography–tandem mass spectrometry (LC-MS/MS) was used to detect 13 kinds of nucleosides, and UV spectrophotometry was conducted to quantitatively analyze polysaccharides. These methods were used to compare the differences between C. cicadae and P. cicadae. In the present study, methods were developed to authenticate C. cicadae and P. cicadae.
Materials and methods
Sample collection and chemicals
Samples of P. cicadae (Lot No 20111222) and C. cicadae (Batch No 100715) for pharmacological experiments and samples of P. cicadae (No S1–S10) and C. cicadae (No S11–S20) for chemical composition analysis experiments were provided by Zhejiang BioAsia Pharmaceutical Co., Ltd (Pinghu, Zhejiang, China). The C. cicadae samples originated from two provinces in China, ie, S11 and S12 from Anji, Zhejiang Province; S13–S15 from Dabieshan, Anhui Province; S16–S18 from Fuyang, Zhejiang Province; and S19 and S20 from Wuhu, Anhui Province. These fungal samples were authenticated by senior experimentalist Junsong Li (Experimental Center for Teaching and Learning, Shanghai University of Traditional Chinese Medicine, Shanghai, China). The P. cicadae and C. cicadae samples were preserved in the Experiment Center for Teaching and Learning, Shanghai University of Traditional Chinese Medicine. The crude drug of P. cicadae and C. cicadae was freshly dissolved in purified water and prepared into 0.1 g/mL suspension, then mixed with the vortex before use. UCG (No 20110305) was obtained from Consun Pharmaceutical Group Co., Ltd (Inner Mongolia, China) and dissolved in purified water and prepared into 0.3 g/mL solution.
Animals and treatment
A total of 70 male Sprague–Dawley rats initially weighing 160–180 g were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd., with a certification number of SCXK (Beijing) 2012-0001. The animals were fed with standard diet and given water ad libitum. All procedures involving animals and their care were approved by the Animal Care Committee of the China Academy of Chinese Medical Sciences in accordance with its regulations for animal experiments. The animals were sacrificed under ether anesthesia.
Experimental design
After 1 week of acclimation, 10 rats, as the normal control group, were given an equal volume of distilled water by gavage at around 20 mL/kg, and the remaining 60 rats were given 250 mg/kg adenine suspension (freshly dissolved in distilled water) by gavage continuously for 21 days to establish a model of renal injury.30,31 Then, the rats were evenly divided according to the values of blood urea nitrogen (BUN) and creatinine (Crea). However, some rats died as the disease progressed. The rest were used in the following experiments. Thus, the four groups included the CRF model group (n=9), C. cicadae group (1.0 g crude drug/kg, n=14), P. cicadae group (1.0 g crude drug/kg, n=11), and UCG group (3 g/kg, n=11). The animals in each group were given a dosage of 1 mL/100 g body weight of corresponding drug or water once daily for 4 weeks. During the experiment, changes in body weight, physical state, fur condition, and activities were observed.
Biochemical analysis
All serum parameters, including BUN, Crea, total protein (TP), and albumin (ALB) levels; calcium (Ca), phosphorus (P), potassium (K), sodium (Na), and chloride (Cl) contents, were analyzed using a HumaLyzer-2000 semiauto chemistry analyzer (Human, Magdeburg, Germany). An MEK-6318K automatic hematology analyzer (Nihon Kohden, Tokyo, Japan) was used to accomplish the blood routine tests, including red blood cell (RBC) count, hemoglobin (HGB), hematocrit (HCT, %), and mean corpuscular volume (MCV). Malondialdehyde (MDA) level, superoxide dismutase (SOD) activity, and glutathione (GSH) content from the blood obtained from the eye socket vein of rats were also determined. A day before the animals were sacrificed, 24-hour urine was collected from the metabolic cages and used to determine urine protein levels.
Histology
After the animals were sacrificed, the kidneys were weighed and then fixed in 10% formaldehyde overnight. The fixed organs were washed with tap water, dehydrated with gradient ethanol (Sakura automatic dehydrator, Sakura Corporation, Osaka, Japan), cleared in xylene, processed and embedded in paraffin, and then stained with hematoxylin and eosin (Sakura SH-100 automatic dyeing machine, Sakura Corporation). The number of normal glomeruli and the degree of adenine metabolite crystallization were determined in three consecutive 10×20 fields per slide under an optical microscope (Olympus BH-2 binocular microscope with automatic microphotographic system, Olympus Corporation, Tokyo, Japan) equipped with a computer-controlled digital camera.
Infrared (IR) fingerprints by FTIR spectroscopy
The structural characteristics of C. cicadae and P. cicadae were identified using a Fourier transform infrared (FTIR) spectrometer in accordance with an optimized procedure.32 In total, 4 mg of S1–S20 was mixed thoroughly with 200 mg of potassium bromide. Then, 100 mg of each mixture powder was obtained and pressed into a transparent thin pellet. All measurements were performed in the spectral range of 4,000–400 cm−1 with a resolution of 4 cm−1 using an IR spectrometer (TENSOR 27, Bruker Corporation, Karlsruhe, Germany). The IR spectra of samples were processed using Thermo Scientific OMNIC™ (version 8.2, Thermo Fisher Scientific, Waltham, MA, USA) to obtain the average IR spectra.
Volatile ingredient determination by GC-MS
Volatile components were extracted and analyzed as described by a previous study.33 A C. cicadae sample (2 g) placed in a 1,000 mL round-bottom flask for simultaneous distillation–extraction (SDE) was added to 200 mL of distilled water and boiled slightly with a heater. Then, 30 mL of ether was added into a 250 mL round-bottom flask for SDE and heated at 45°C for 4 hours. After cooling down, the ether layer was collected into a centrifuge tube with 5 g of anhydrous sodium sulfate and stored overnight. Subsequently, the mixture was filtrated, and the filtrate was placed in an evaporating dish and evaporated to dryness in a water bath under 45°C. The residue was reconstituted with 0.5 mL of ether and transferred to a centrifuge tube for centrifugation at 10,000 rpm (ST16R high-speed refrigerated centrifuge, Thermo Fisher Scientific). GC-MS analysis of the test solution was performed on a gas chromatograph (HP-7890, Agilent Technologies, Santa Clara, CA, USA) coupled with a mass selective detector (HP-5977, Agilent Technologies) equipped with an HP-5MS-fused silica column (30 m×0.25 mm, 0.25 μm). The oven temperature was 60°C, held for 1 minute and ramped at 5°C/min to 200°C, held for 5 minutes and then again to 270°C at 10°C/min and finally held for 10 minutes. The sample was injected at 230°C with an injection volume of 1.0 μL using helium as carrier gas (1.0 mL/min) and split at the ratio of 1:5. The mass spectrometer was operated in the electron ionization mode at 70 eV with a continuous scan from 35 to 500 m/z. The atlases of samples were imported from TXT format into the Similarity Evaluation System for Chromatographic Fingerprint of Traditional Chinese Medicine software (version 2012, Chinese Pharmacopoeia Commission, China).
Nucleoside determination by LC-MS/MS
Nucleoside extracts from C. cicadae were prepared as previously reported.34 C. cicadae powder (0.2 g) was screened with a 60-mesh sieve and mixed with 10 mL of 10% (v/v) methanol. The supernatant was obtained through ultrasonic ice bath (KH-400KDB Ultrasonic Cleaner, Kunshan Ultrasonic Instrument Co., Ltd., Kunshan, China) at 40 kHz for 2 hours and centrifuged (ST16R high-speed refrigerated centrifuge, Thermo Fisher Scientific) at 5,000 rpm for 15 minutes. The residue was re-extracted twice, and the resultant supernatant was centrifuged with constant stirring at 13,000 rpm for 10 minutes. Chromatographic separation was performed on a QTRAP 5500 LC-MS/MS system (AB SCIEX Co., Ltd., Boston, MA, USA). Compounds were separated by using an ultrahigh-performance liquid chromatography (UPLC) column (50×2.1 mm, 1.8 μm, Waters Acquity UPLC HSS T3, Waters Co., Ltd., USA). The mobile phase composed of water (A) and acetonitrile (B) was obtained with the following gradient elution conditions: 0.01–3 minutes (99:1–96:4), 3–5 minutes (96:4–70:30), 5–6 minutes (70:30), 6–6.01 minutes (70:30–99:1), and 6.01–7 minutes (99:1). The column temperature was 25°C, the flow rate was maintained at 0.4 mL/min, the temperature in the autosampler was set at 15°C, and the injection volume was 10 μL.
Polysaccharide determination by UV spectrophotometry
Crude polysaccharides were prepared in accordance with a slightly optimized version of the procedure.35 Precisely weighed (1 g) powdered C. cicadae or P. cicadae was extracted with a 20-fold volume of distilled water through ultrasonic (KH-400KDB Ultrasonic Cleaner, Kunshan Ultrasonic Instrument Co., Ltd., Kunshan, China) at 90°C for 90 minutes and centrifuged (ST16R high-speed refrigerated centrifuge, Thermo Fisher Scientific) at 10,000 rpm for 15 minutes (duplicated twice). After each extraction, the polymers were filtered, and the filtrate was combined and condensed to 5 mL and diluted to 10 mL. The extracts were further precipitated with ethanol until 80% and then left to stand overnight. The precipitate was then collected by vacuum filtration and added to 250 mL of distilled water under ultrasonication for 30 minutes at 70°C. An appropriate amount of sample solution was mixed with 1 mL phenol solution (5%) and 5 mL sulfuric acid and then incubated at 40°C for 30 minutes. After the sample solution was cooled down, its absorbance was measured at 490 nm by UV spectrophotometry (8454 UV–Vis, Agilent Technologies).
Statistical analysis
The statistical significance of the chemical composition data was assessed using paired t-test, whereas the hematological and biochemical results were analyzed using independent t-test. Meanwhile, the histopathological data were subjected to Ridit analysis.36,37 A value of P<0.05 was considered to be statistically significant.
Results
Animal signs
Compared with the control rats, the model rats showed growth inhibition, weight loss, polydipsia, polyuria, hair dryness and loss, chills and back arch, and poor mental state. The other group of rats improved in appearance and mental state during administration. The rats were weighed before the beginning of the treatment and weekly during the treatment period. The data of 4-week cumulative body weight gain were presented in Figure 2A. The CRF model group showed negative growth on weight gain, whereas the C. cicadae, P. cicadae, and positive control groups all exhibited accumulated increases (P<0.01). In Figure 2B, urine output in each treated group increased, especially in the C. cicadae group (P<0.05).
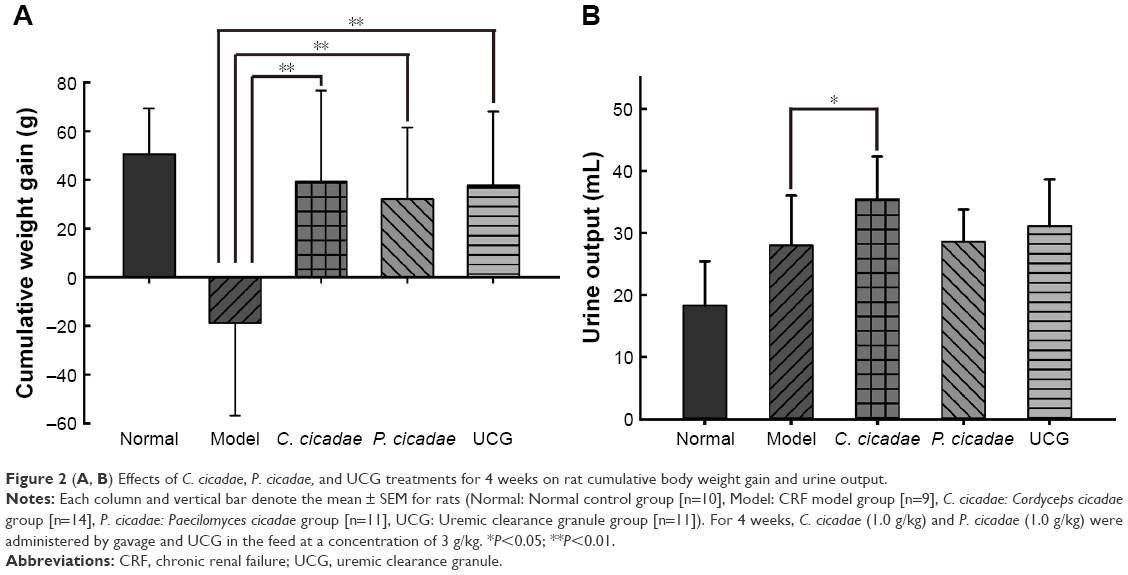 | Figure 2 (A, B) Effects of C. cicadae, P. cicadae, and UCG treatments for 4 weeks on rat cumulative body weight gain and urine output. |
BUN and Crea in serum
Three weeks after modeling, the serum BUN and Crea levels were significantly increased (P<0.01), which demonstrated the success in modeling. As shown in Figure 3A and B, the 4-week treatment of C. cicadae, P. cicadae, and UCG caused significant decreases (P<0.01) in serum BUN and Crea levels. In Figure 3C and D, every group had a slight, but statistically insignificant, reduction in urine protein, whereas the P. cicadae group showed a marked reduction in 24-hour urine protein (P<0.05).
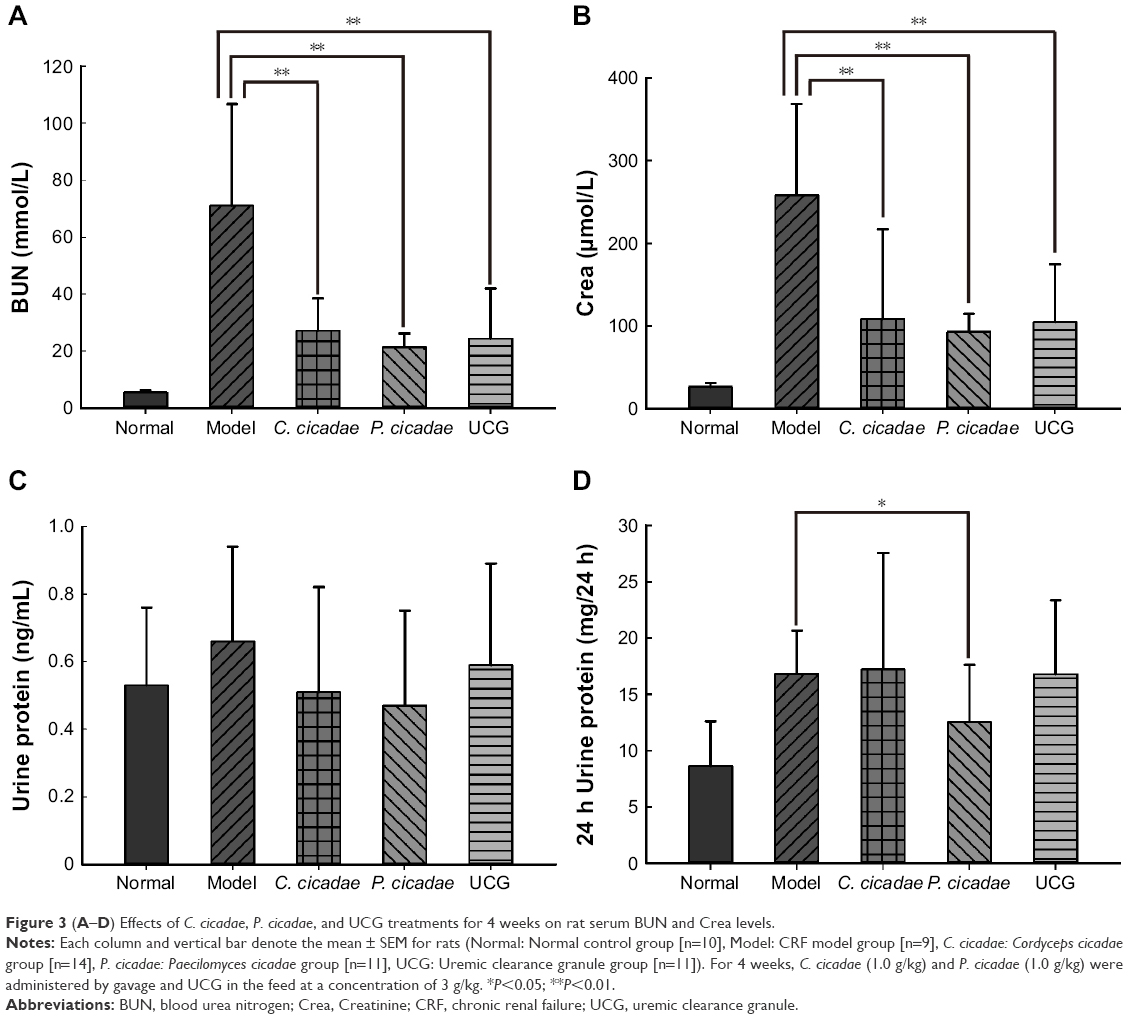 | Figure 3 (A–D) Effects of C. cicadae, P. cicadae, and UCG treatments for 4 weeks on rat serum BUN and Crea levels. |
MDA and renal antioxidant enzymes
The effects of C. cicadae and P. cicadae oxidative stress were determined by measuring the MDA levels and GSH content and analyzing the activities of oxidative-stress-associated enzymes, such as SOD. As shown in Figure 4, the C. cicadae and P. cicadae groups exhibited lower MDA levels, and the UCG group showed a slightly higher MDA level than that of the model group. SOD activity was increased in the C. cicadae, P. cicadae, and UCG groups, of which the P. cicadae and UCG groups showed significant difference compared with the model group (P<0.05). C. cicadae, P. cicadae, and UCG abated GSH levels. Compared with the model group, the C. cicadae and P. cicadae groups showed lower MDA and GSH levels and higher SOD activity, which suggested that C. cicadae and P. cicadae can inhibit oxidative stress and enhance antioxidant capacity.
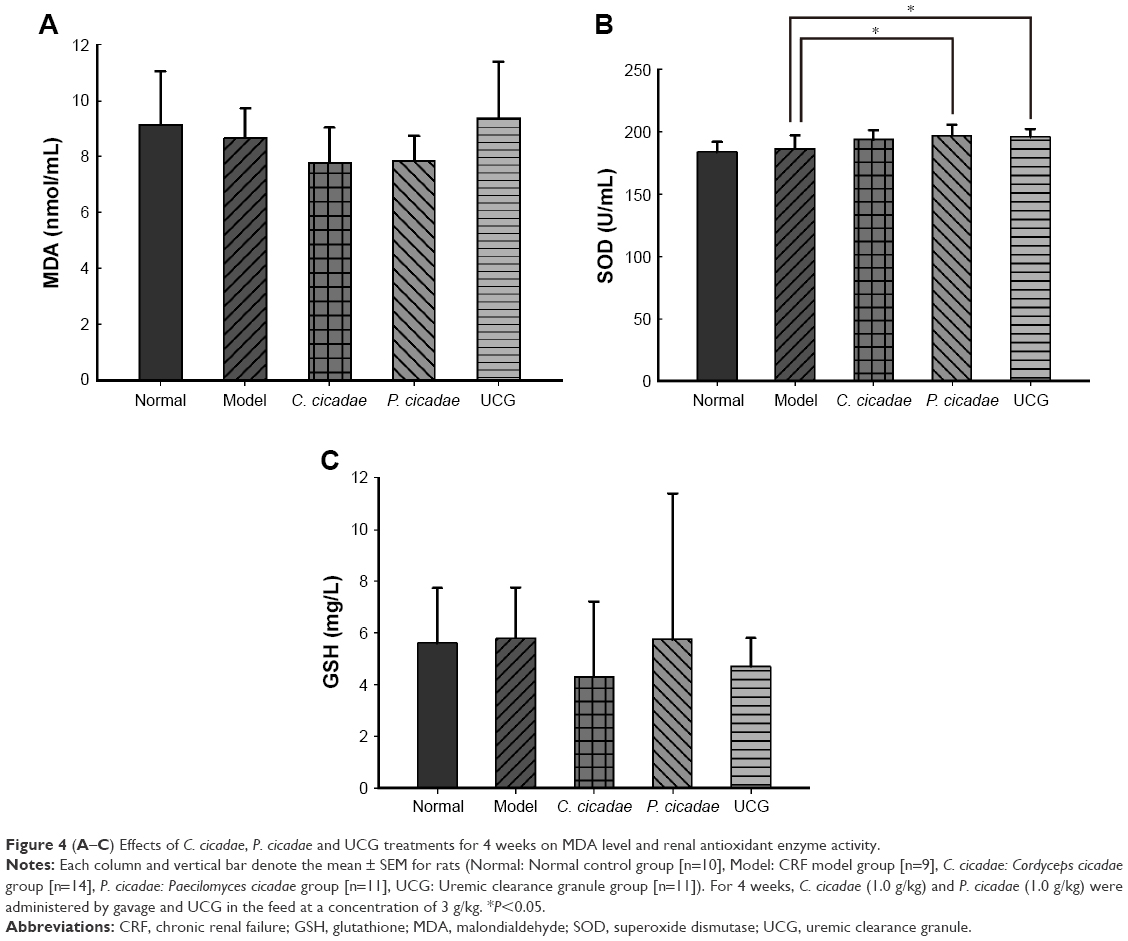 | Figure 4 (A–C) Effects of C. cicadae, P. cicadae and UCG treatments for 4 weeks on MDA level and renal antioxidant enzyme activity. |
Blood biochemical measurements
As shown in Table 1, adenine-induced model rats exhibited decreased TP and ALB levels; ion disorders of Ca, P, Na, K, and Cl; and significant reduction in RBC, HCT, and HGB with obvious appearance of anemia symptoms. After treatment, the TP and ALB levels in each group were close to normal values but higher than those in the model group, and the change was significant in the UCG group. Each treatment group presented with increased serum Ca levels that were significant in the UCG group (P<0.05) and insignificantly decreased serum P levels. The serum Na, K, and Cl levels also showed no obvious changes. Thus, C. cicadae and P. cicadae successfully ameliorated the ion disorders. Compared with the normal group, the CRF model group showed obvious anemia symptoms with significantly decreased RBC, HGB, and HCT levels. With further comparison, the RBC, HGB, and HCT in the treated groups were higher than those in the model group. C. cicadae, P. cicadae, or UCG treatments increased MCV levels, most especially the C. cicadae and UCG groups (P<0.05 and P<0.01, respectively). These results demonstrated that C. cicadae and P. cicadae can alleviate anemia symptoms to a certain extent.
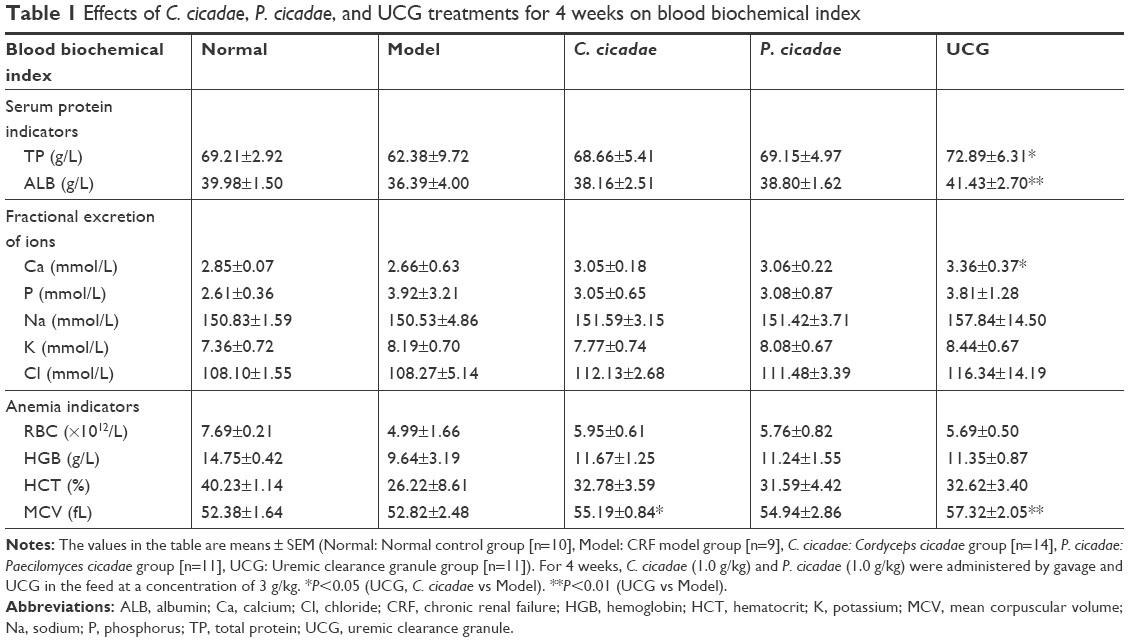 | Table 1 Effects of C. cicadae, P. cicadae, and UCG treatments for 4 weeks on blood biochemical index |
Histopathology
The kidneys in the model group were pale and enlarged significantly. Furthermore, the kidneys of rats administered with C. cicadae, P. cicadae, or UCG increased to varying degrees and appeared to improve relative to those of the model group. Given the histopathological sections, the model group showed significantly less glomeruli with purine crystals that presented as brownish sediments, crystalline deposition, interstitial inflammation in the proximal and distal convoluted tubules, and severe cell proliferation. The rats given adenine plus C. cicadae, P. cicadae, or UCG concomitantly showed improvement in histological appearance relative to that of the model group, especially in the C. cicadae and UCG groups. After modeling, the kidneys of the model group enlarged significantly. However, with the 4-week treatment both C. cicadae and P. cicadae can significantly decrease the kidney indices and increase the number of the glomerulus (Figure 5). More glomeruli (Figure 6) and less renal crystallization (Figure 7) were observed in each treated group relative to those in the CRF model group. According to Ridit analysis, when the fiducial limit >0.5, P<0.05. As shown in Table 2, C. cicadae, P. cicadae, and UCG treatments significantly abated the crystallization degree in kidneys (P<0.05).
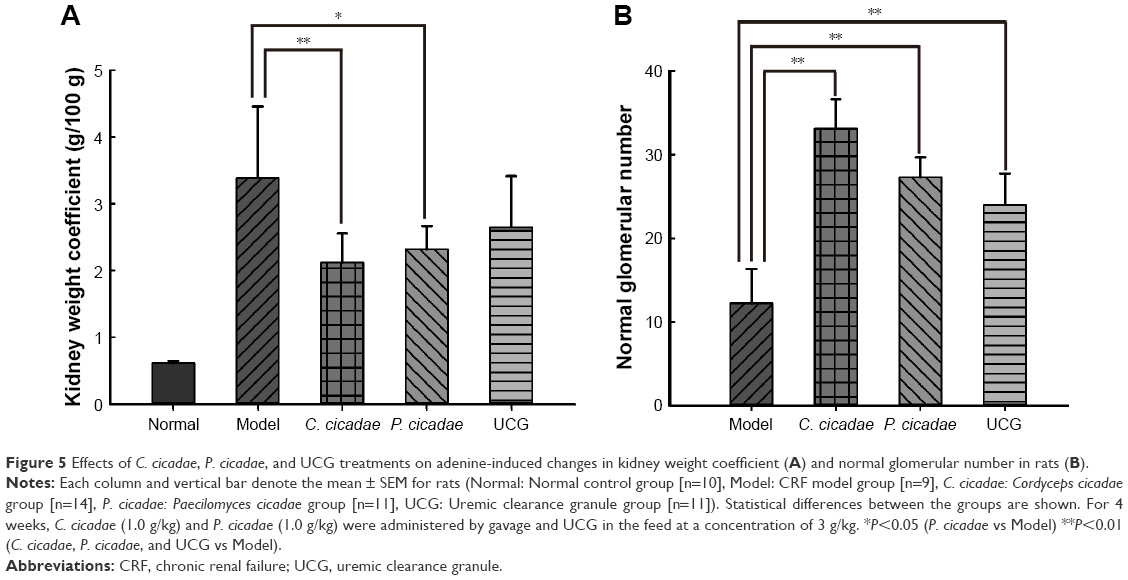 | Figure 5 Effects of C. cicadae, P. cicadae, and UCG treatments on adenine-induced changes in kidney weight coefficient (A) and normal glomerular number in rats (B). |
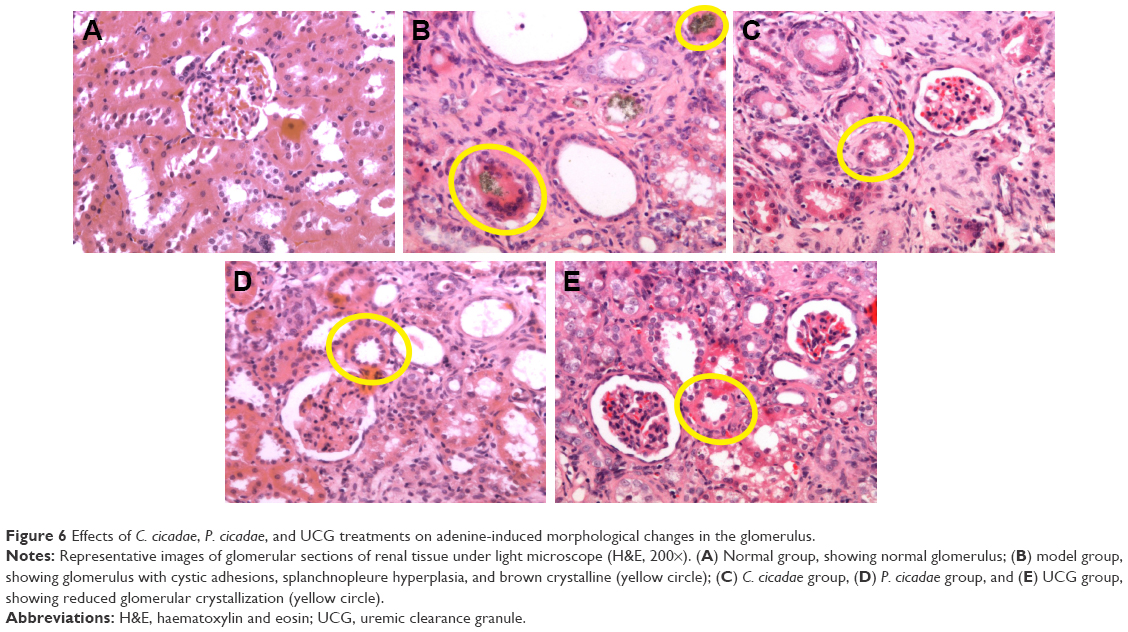 | Figure 6 Effects of C. cicadae, P. cicadae, and UCG treatments on adenine-induced morphological changes in the glomerulus. |
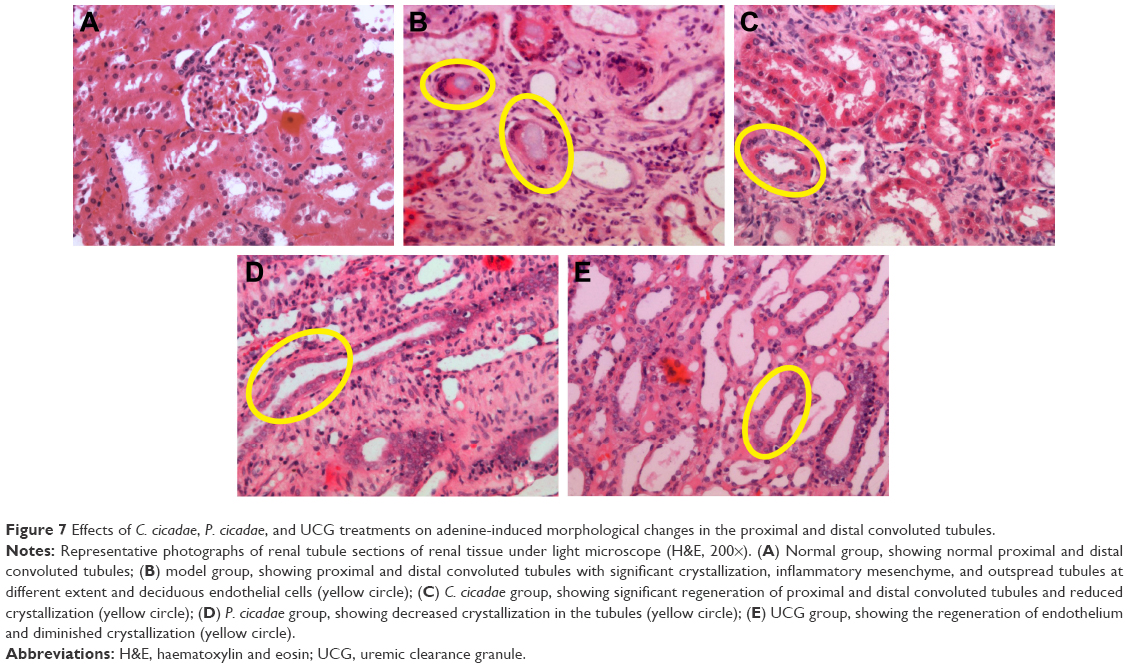 | Figure 7 Effects of C. cicadae, P. cicadae, and UCG treatments on adenine-induced morphological changes in the proximal and distal convoluted tubules. |
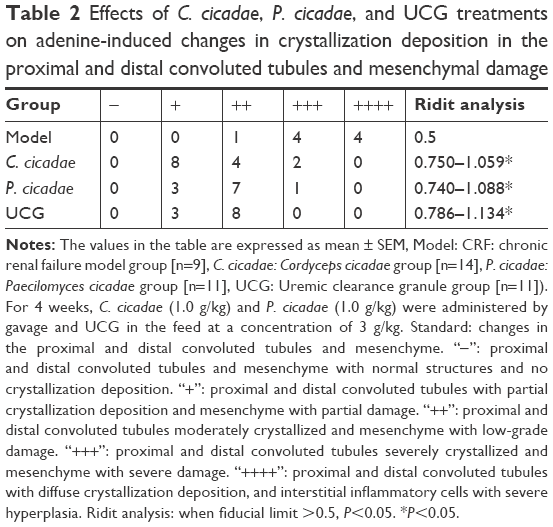 | Table 2 Effects of C. cicadae, P. cicadae, and UCG treatments on adenine-induced changes in crystallization deposition in the proximal and distal convoluted tubules and mesenchymal damage |
FTIR spectroscopy
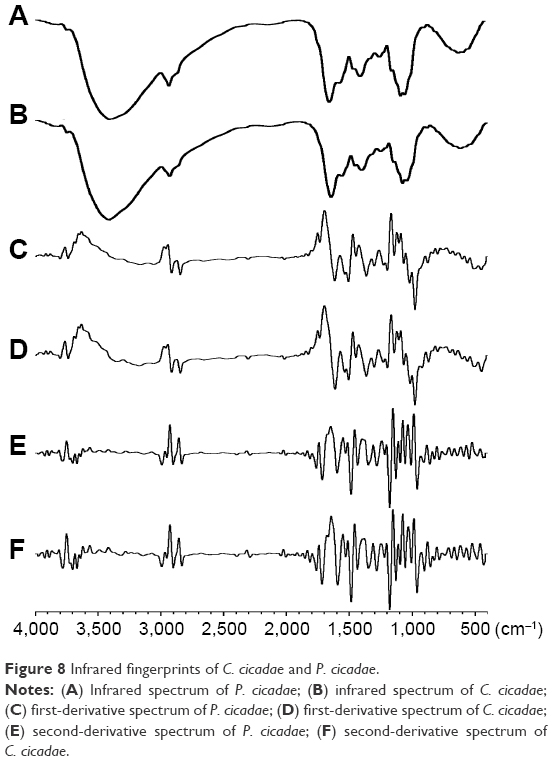
A total of ten batches each of C. cicadae and P. cicadae were used to prepare samples to be subjected to FTIR spectroscopy. Considering the overall distribution characteristics of IR spectra (Figure 8A and B), P. cicadae was basically the same as C. cicadae. The similarity analysis showed a result of over 0.90 for the ten batches of C. cicadae and P. cicadae. The abovementioned results showed that the IR fingerprints of the ten batches each of C. cicadae and P. cicadae were superior in consistency.
 | Figure 8 Infrared fingerprints of C. cicadae and P. cicadae. |
First- and second-derivative spectra can enhance IR resolution; therefore, this study used the Norris derivative filter to process reference spectra and obtain the first- and second-derivative spectra of C. cicadae and P. cicadae. As shown in Figure 8C–F, many peaks in different positions were more obvious in the high-resolution second-derivation spectra than in the first-derivative spectra. The absorption peaks of 1,680 and 1,640 cm−1 were attributed to the sheer vibration of the nitrogenous heterocycle N−H and the superposition of the C−C group stretching vibration, respectively. These results suggested the presence of purine, pyrimidine, steroids, or triterpenoids with double bonds. The absorption of 991 cm−1 was inferred to represent the bending vibration of the C−H group in the double bonds of steroids or triterpenoids. The absorption peak at ~1,516 cm−1 was attributed to the bending vibration of the aromatic or heterocyclic ring C=C skeleton. These results confirmed the presence of aromatic compounds in both C. cicadae and P. cicadae. Furthermore, the heights of the peak at ~1,516 cm−1 differed from each other and implied that C. cicadae contains more aromatic compounds than do P. cicadae. Therefore, GC-MS was used in the succeeding experiments to explore the volatile compounds in C. cicadae and P. cicadae and determine the different constituents for identifying the two species.
GC-MS
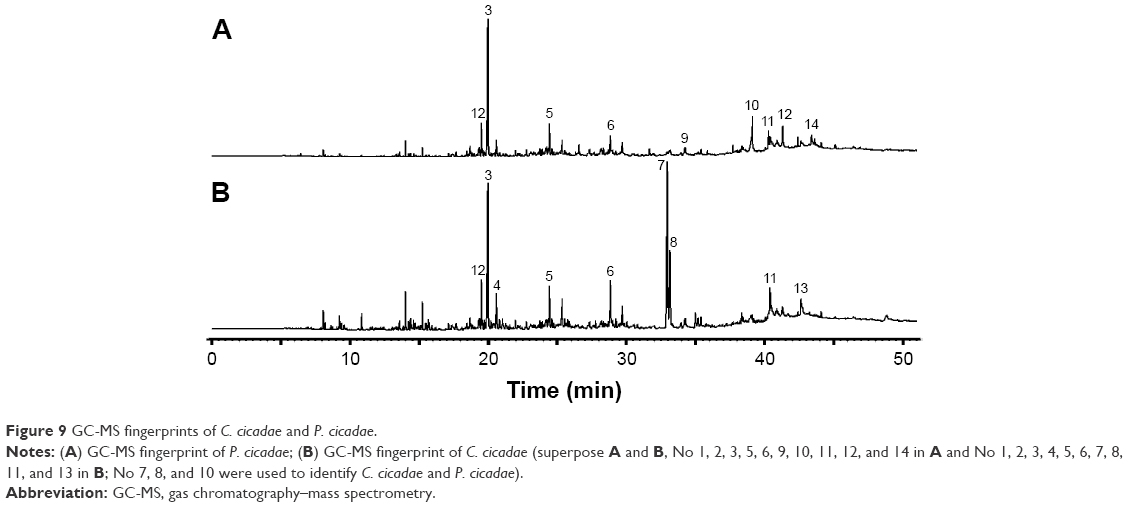
Twenty batches of samples were used to prepare sample resolutions, which were tested by GC-MS. The GC-MS spectra of each sample were then recorded. Control fingerprint spectra with the conditions of multipoint correction were generated for the retention time of the chromatographic peak, full peak match, and time width of 0.1 minute (Figure 9). The data matching results were obtained by using the data matching function of the system, where P. cicadae S1–S10 showed 20 common peaks and C. cicadae S11–S20 presented 39 common peaks. Then, 18 common peaks were noted in S1–S20, of which butylated hydroxytoluene is a commonly used antioxidant with antagonistic effects of UV-induced cellular immunosuppression and protective effects on the skin against photoaging caused by UV exposure. Meanwhile, 5,5-diphenylhydantoin, known as phenytoin, is usually used to treat epilepsy and arrhythmia. Six of the ten component compositions with the highest peak areas in C. cicadae and P. cicadae were the same. The highest peak area percentages of the top ten ingredients in C. cicadae and P. cicadae were 51.58% and 40.15%, respectively.
 | Figure 9 GC-MS fingerprints of C. cicadae and P. cicadae. |
GC-MS was adopted to detect the total volatile components of C. cicadae and P. cicadae. By searching the NIST05a database, we noted that 19 of 23 differential components were aliphatic hydrocarbons, and the rest belonged to ester compounds. Among these constituents, the esters (Z,Z)-9,12-octadecyl dioleate and 9-cis-11-octadecenoic acid methyl ester were 15.59% and 7.35%, respectively, in C. cicadae but 0.49% and 0.83%, respectively, in P. cicadae. In addition, the diisobutyl phthalate contents in C. cicadae and P. cicadae accounted for 0.38% and 0.00%, respectively, whereas the eicosane fat hydrocarbon compounds were 0.44% and 7.20%, respectively. The contents of the abovementioned four components in C. cicadae and P. cicadae significantly differed and were used as the basis for rapid identification. Notably, only the diammonium phthalate in C. cicadae presented with cytotoxicity, genetic toxicity, reproductive development toxicity, and neurotoxicity,38 which can be used as quality control standards of C. cicadae. Therefore, P. cicadae is safer than C. cicadae to some extent.
LC-MS/MS
For LC-MS/MS, ten batches each of C. cicadae and P. cicadae were used to prepare test solutions, which were subsequently measured. The typical chromatograms of the 13 analytes in multiple reaction monitoring mode are presented in Figure 10. The contents of 13 kinds of nucleosides are presented in Figure 11.
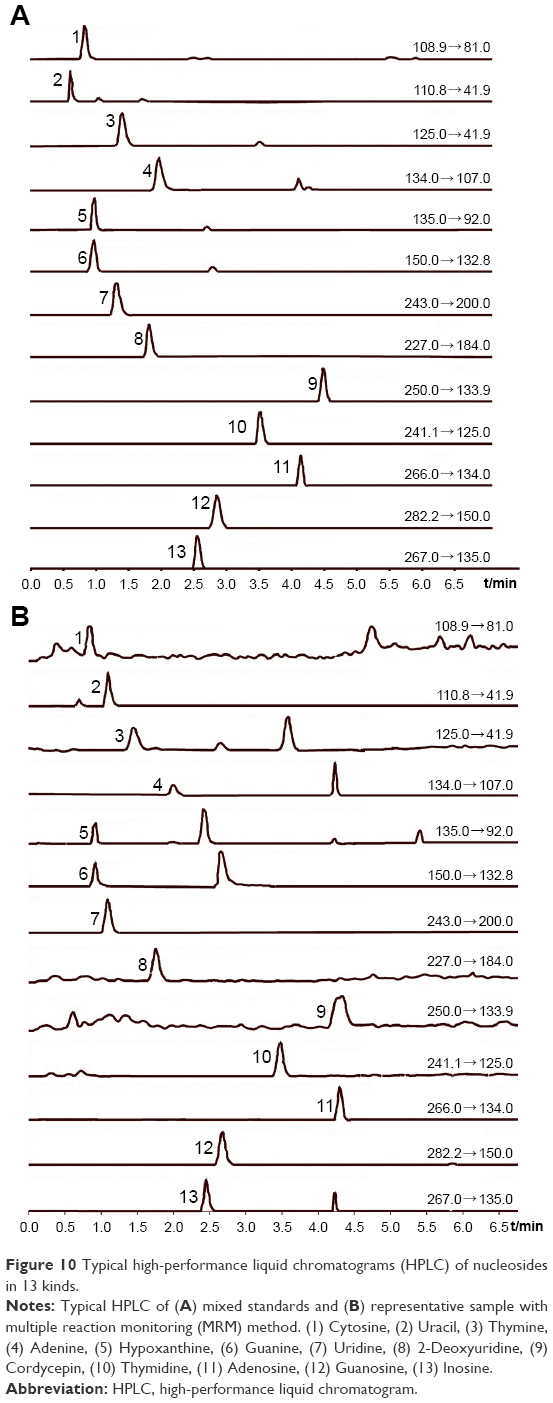 | Figure 10 Typical high-performance liquid chromatograms (HPLC) of nucleosides in 13 kinds. |
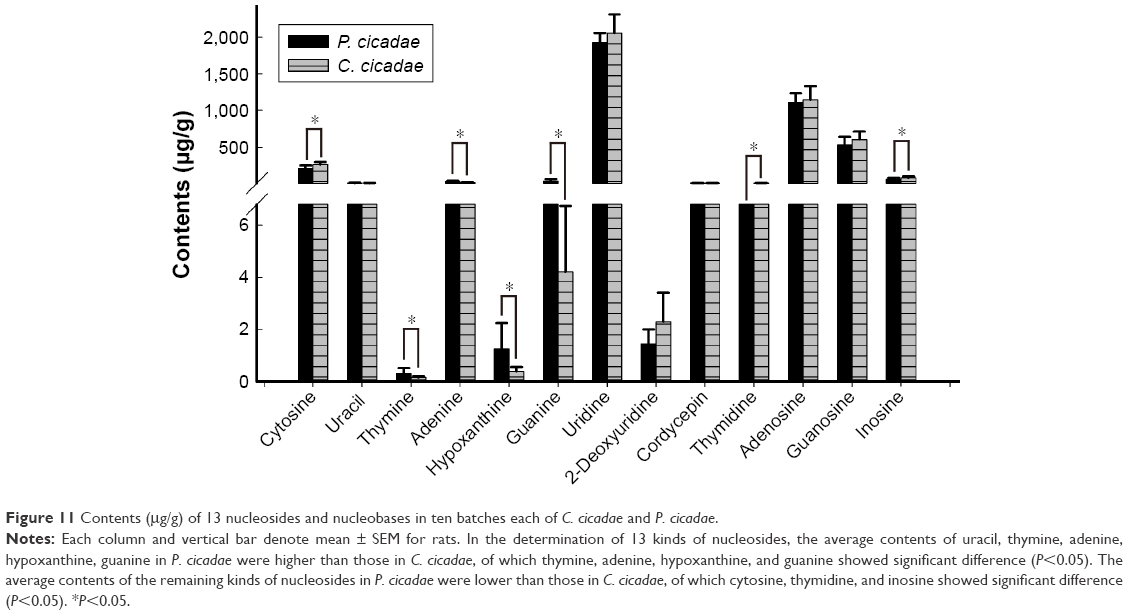 | Figure 11 Contents (μg/g) of 13 nucleosides and nucleobases in ten batches each of C. cicadae and P. cicadae. |
The range of total nucleosides in P. cicadae was 3,439.73–4,293.23 μg/g, whereas those in C. cicadae was 3,477.87–4,976.66 μg/g. Among the constituents of P. cicadae and C. cicadae, uridine had the highest concentrations (1,926.35 and 2,055.18 μg/g, respectively) and thymine had the lowest concentrations (0.30 and 0.15 μg/g, respectively). Component analysis revealed that cytosine, guanine, uridine, adenosine, and inosine mainly affected the total nucleoside contents in P. cicadae. In the analysis of ten batches of C. cicadae, the total nucleoside contents of C. cicadae from the Anji County of Zhejiang Province were higher than those from other areas, and the uridine, adenosine, and inosine contents differed significantly. As shown by the LC-MS/MS results, C. cicadae and P. cicadae contained cordycepin, adenosine, hypoxanthine, guanosine and other nucleoside substances. The results obtained by FTIR and LC-MS/MS are mutually confirmed.
During the determination of 13 kinds of nucleosides, the average contents of uracil, thymine, adenine, hypoxanthine, and guanine in P. cicadae were higher than those in C. cicadae, of which thymine, adenine, hypoxanthine, and guanine showed significant difference (P<0.05). The average contents of the remaining kinds of nucleosides in P. cicadae were lower than those in C. cicadae, of which cytosine, thymidine, and inosine showed significant difference (P<0.05).
UV spectrophotometry
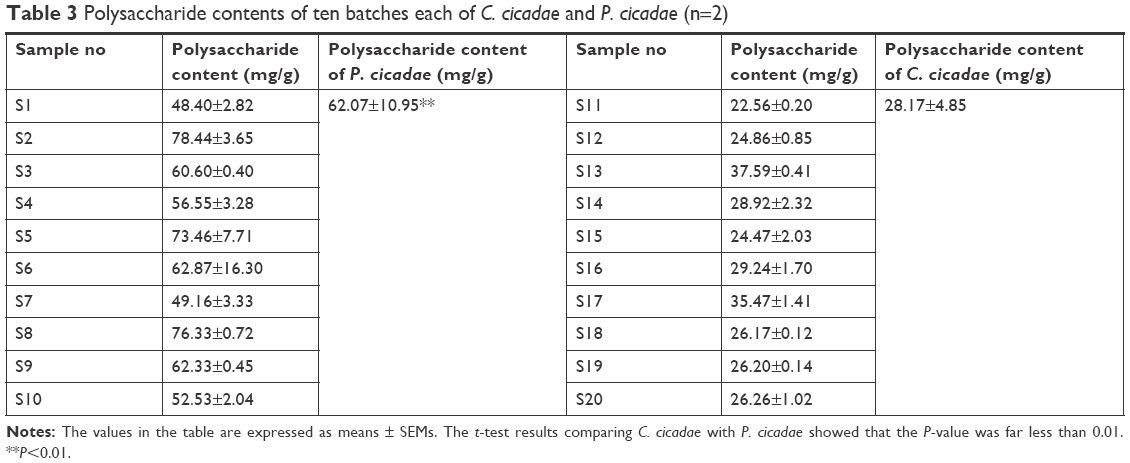
Ten batches each of C. cicadae and P. cicadae were used for sample preparation and measurement by UV spectrophotometry. Table 3 shows the polysaccharide content at 490 nm wavelength determined by UV spectrophotometry. The polysaccharide content of P. cicadae was 62.07 mg/g. The t-test results comparing C. cicadae with P. cicadae revealed that the P-value was far less than 0.01 (P=0.00002), indicating that the polysaccharide content was significantly different between C. cicadae and P. cicadae.
 | Table 3 Polysaccharide contents of ten batches each of C. cicadae and P. cicadae (n=2) |
Discussion
Natural C. cicadae has a long history of use as a traditional medicine in China. However, given its slow growth and immoderate exploitation, C. cicadae resources have been severely depleted. Naturally, because of high value in medicine, the produce of cultivated P. cicadae, which is controllable, can meet market demand and has promising prospects in the future.
Three weeks after modeling, adenine-induced rats had significant weight loss and poor mental state and expressed obvious symptoms of anemia. The serum BUN and Crea levels were significantly increased; TP, ALB, RBC, HCT, and HGB were decreased; and ion disorder of Ca, P, Na, K, and Cl was observed, suggesting the success of modeling. After 4-week administration, rats of the two groups exhibited accumulated increases in weight; improved mental state; significant decrease in serum BUN and Crea levels; reduced urinary protein concentration and 24 hours urine protein; increased TP, ALB, RBC, HCT, and HGB; and improved ion disorder and anemia. The C. cicadae group significantly decreased BUN and Crea (P<0.01) and increased MCV (P<0.05), whereas the P. cicadae group showed a marked reduction in BUN and Crea (P<0.01) and 24-hour urine protein (P<0.05) and increase in SOD (P<0.05). Comprehensive results of serum biochemical indicators demonstrated that P. cicadae had better therapeutic effects on CRF than C. cicadae did. According to histopathological observation, both C. cicadae and P. cicadae can significantly decrease the kidney indices, increase the number of the glomerulus, reduce the deposition of adenine metabolites in the renal tubules, and ease the degree of kidney disease. However, C. cicadae exhibited better effects than P. cicadae on the above aspects. The mechanism may be related to the downregulated PAI-1. Given the serum biochemical indicators and pathological tissue sections of adenine-induced CRF rats, we demonstrated that both C. cicadae and P. cicadae significantly ameliorated adenine-induced CRF in rats and had comparatively therapeutic effects.
In the present study, we established fingerprints of ten batches each of C. cicadae and P. cicadae from different time points and production places via FTIR and GC-MS, which provides the basis for authenticity identification. Based on the results of LC-MS/MS, both C. cicadae and P. cicadae had 13 kinds of nucleosides with the highest uridine content and the lowest thymine content. In addition, the results of UV spectrophotometry suggested that two species had a large amount of plant polysaccharides. Moreover, the results of FTIR showed that C. cicadae and P. cicadae were similar to the one-dimensional spectra, with both containing polysaccharides, glycosides, hydrocarbons, steroids, terpenoids, saponins, proteins, and amino acids. The results of this study revealed that both C. cicadae and P. cicadae exerted obvious therapeutic effects on CRF and were more consistent with their chemical compositions. Hence, P. cicadae can be used to treat CRF instead of C. cicadae. However, the treatment effects of P. cicadae on other diseases entail further efficacy and clinical studies.
Moreover, polysaccharides,39 cordycepin, adenosine, and inosine22 in C. cicadae are effective for renal protection, specifically reducing the inflammatory response and kidney damage. Given the abovementioned information, our study focused on whether the efficacies of C. cicadae and P. cicadae are closely related to their constituent pharmacodynamic substances. Our study demonstrated that by using LC-MS/MS, C. cicadae contains almost the same cordycepin and adenosine contents as those in P. cicadae. As the two-sample t-test results showed, the differences in total nucleoside contents between the ten batches of C. cicadae and those of P. cicadae were statistically insignificant. However, C. cicadae possesses a significantly higher inosine content than P. cicadae (P<0.05). UV spectrophotometry revealed that the polysaccharide content of P. cicadae was significantly higher (2.2 times) than that of C. cicadae (P<0.01). As can be seen in the determination of effective medicinal components, including cordycepin, adenosine, inosine, and polysaccharides, although C. cicadae and P. cicadae were significantly capable of treating rats with adenine-induced CRF, inosine and polysaccharide contents were significantly different between them. These results suggested that not only cordycepin, adenosine, inosine, and polysaccharides but also other nucleoside and fat-soluble small molecular components may play a role in protecting renal function. Future studies are needed to examine the protective effects of other components.
As reported by other studies, the identification methods of Cordyceps sinensis are mainly as follows: morphological and microscopic identification methods, HPLC-UV spectrophotometry,40 LC-MS/MS,41 depth profiling mid-IR photoacoustic spectroscopy,42 polymerase chain reaction and lateral flow rapid identification method,43 internal transcribed spacer sequence analysis, and random amplified polymorphic DNA molecular marker method.44 However, studies that verify C. cicadae have not been conducted. Based on the results of detection, this article mainly provides three methods for rapid identification to fill gaps in scientific research: (1) GC-MS, to determine the volatile components, including (Z,Z)-9,12-octadecyl dioleate, 9-cis-11-octadecenoic acid methyl ester, diisobutyl phthalate, and eicosane. As shown by the results of GC-MS, diisobutyl phthalate can only be found in C. cicadae, and C. cicadae contains more (Z,Z)-9,12-octadecyl dioleate and 9-cis-11-octadecenoic acid methyl ester and lesser eicosane than P. cicadae. (2) LC-MS/MS, to determine the hypoxanthine, guanine, and inosine contents. C. cicadae possesses lesser hypoxanthine and guanine and more inosine contents than P. cicadae. Inosine is the main identification index of natural C. sinensis, artificial aweto and Cordyceps militaris. The component is also an indicator for C. cicadae and P. cicadae. (3) UV spectrophotometry, to determine polysaccharides. C. cicadae has much lesser polysaccharides than P. cicadae. These examinations revealed significant differences between the two species.
Conclusion
Three specific methods including GC-MS, LC-MS/MS, and UV spectrophotometry developed in this study provide users with easy authentication methods and make a major contribution to the detection of C. cicadae and P. cicadae and suggested that both C. cicadae and P. cicadae have similar chemical compositions and have various polysaccharides and volatile compounds and different contents in 13 kinds of nucleosides. What is more, we demonstrated that C. cicadae and P. cicadae can protect the glomerulus and reduce the deposition of adenine metabolites in the renal tubules. The results of the abovementioned experiments for pharmacological analysis suggested that not only cordycepin, adenosine, inosine, and polysaccharides but also other nucleoside and fat-soluble small molecular components play a comprehensive role in protecting renal function. In conclusion, P. cicadae can be used as an alternative to C. cicadae for treating CRF to cater to market demands.
Acknowledgments
This research was funded by grants from Program of Shanghai Committee of Science and Technology (Grant No 17401902300) and Program of Shanghai Academic/Technology Research Leader (Grant No 18XD1403700).
Author contributions
BW and YW contributed to the conception of the paper; LL, CL, and NL contributed significantly to complete the experimental work; LL and LX prepared the manuscript; TH and TZ contributed to the constructive discussions. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Shi Z, Pan HJ, Fan LF. Advances in research of polysaccharides in Cordyceps species. Food Technol Biotechnol. 2009;47(3):304–312. | ||
Kiho T, Miyamoto I, Nagai K, Ukai S, Hara C. Minor, protein-containing galactomannans from the insect-body portion of the fungal preparation Chán huā (Cordyceps cicadae). Carbohydr Res. 1988;181(88):207–215. | ||
Li IC, Hsu J, Lin TW, Lin WH, Chen CC. Prenatal developmental toxicity study of HEA-enriched Cordyceps cicadae mycelia in Sprague-Dawley rats. SDRP J Food Sci Technol. 2017;2(1):1–7. | ||
Li SP, Zhao KJ, Ji ZN, et al. A polysaccharide isolated from Cordyceps sinensis, a traditional Chinese medicine, protects PC12 cells against hydrogen peroxide-induced injury. Life Sci. 2003;73(19):2503–2513. | ||
Yang FQ, Guan J, Li SP. Fast simultaneous determination of 14 nucleosides and nucleobases in cultured Cordyceps using ultra-performance liquid chromatography. Talanta. 2007;73(2):269–273. | ||
Kuo YC, Lin LC, Don MJ. Cyclodesipeptide and dioxomorpholine derivatives isolated from the insect-body portion of the fungus cordyceps cicadae. J Chin Med. 2002;13(4):209–219. | ||
Yu JW, Xu HJ, Mo ZH, Zhu HL, Mao XB. Determination of myriocin in natural and cultured Cordyceps cicadae using 9-fluorenylmethyl chloroformate derivatization and high-performance liquid chromatography with UV-detection. Anal Sci. 2009;25(7):855–859. | ||
Wang JH, Zhang ZL, Wang YQ. Chemical constituents from mycelia and spores of fungus Cordyceps cicadae. Chin Herb Med. 2017;9(2):188–192. | ||
Ge F, Xia CR, Li CR, et al. Analysis of the chemical compositions of Paecilomyces cicadae fermented mycelia and Cordyceps cicadae fruit body. Mycosystema. 2007;26(1):68–75. | ||
Jin ZH, Chen YP. Clinical observation on Cordyceps cicadae Shing tang in preventing the progression of chronic renal failure. Chin Arch Tradit Chin Med. 2006;24(8):1457–1459. | ||
Wang HY, Chen YP. Clinical experience of Professor Chen about Cordyceps cicadae. Chin J Inform Traditional Chin Med. 2000;7(11):71. | ||
Chiu CP, Hwang TL, Chan Y, et al. Research and development of Cordyceps in Taiwan. Food Sci Human Wellness. 2016;5(4):177–185. | ||
Liu SQ, Wen R, Xia M, Jiang N. Determination of the active ingredient produced in the artificial cultivated Cordyceps sobolifera. J Anhui Agricult Sci. 2008;36(2):429–467. | ||
Fukatsu T, Sato H, Kuriyama H. Isolation, inoculation to insect host, and molecular phylogeny of an entomogenous fungus Paecilomyces tenuipes. J Invertebr Pathol. 1997;70(3):203–208. | ||
Kim HS, Kim YJ, Lee HK, et al. Activation of macrophages by polysaccharide isolated from Paecilomyces cicadae through toll-like receptor 4[J]. Food chem toxicol. 2012;50(9):3190–3197. | ||
Ren XY, He L, Cheng JW, Chang JM. Optimization of the solid-state fermentation and properties of a polysaccharide from Paecilomyces cicadae (Miquel) Samson and its antioxidant activities in vitro. PLoS One. 2014;9(2):e87578. | ||
Chyau CC, Chen CC, Chen JC, Yang TC, Shu KH, Cheng CH. Mycelia glycoproteins from Cordyceps sobolifera ameliorate cyclosporine-induced renal tubule dysfunction in rats. J Ethnopharmacol. 2014;153(3):650–658. | ||
Jin ZH, Chen YP, Deng YY. The mechanism study of Cordyceps sobolifera mycelium preventing the progression of glomerulosclerosis. Chin J Integ Tradit West Nephrol. 2005;6(3):132–136. | ||
Zhu R, Chen YP, Deng YY, et al. Cordyceps cicadae extracts ameliorate renal malfunction in a remnant kidney model. J Zhejiang Univ Sci B. 2011;12(12):1024–1033. | ||
Wojcikowski K, Johnson DW, Gobe G. Herbs or natural substances as complementary therapies for chronic kidney disease: ideas for future studies. J Lab Clin Med. 2006;147(4):160–166. | ||
Wang Y, Yin HP, Lv XB, et al. Protection of chronic renal failure by a polysaccharide from Cordyceps sinensis[J]. Fitoterapia. 2010,81(5): 397–402. | ||
Li B, Zhang WC, Fu CX, Zheng Y. Effect of Cordyceps and adenosine on kidney oxidative stress of diabetic mice. World Latest Med Inform. 2016;42(10):770–772. | ||
Wu W, Huang YR, Wan YG, et al. Effects and mechanisms of UCG ameliorating renal interstitial fibrosis by regulating TGF-β1/SnoN/Smads signaling pathway in renal failure rats. China J Chin Mater Med. 2016;41(12):2291–2297. | ||
Tang SF, Guo SS, Hong QG. Effects of niaoduqing capsule and its ingredients on expressions of TGF-1 FN LN in renal tissue of CRF rats modeled by 5/6 nephrectomy. Chin Arch Traditional Chin Med. 2008;26(8):1656–1658. | ||
Wei CY, Li WQ, Shao SS, et al. Structure and chain conformation of a neutral intracellular heteropolysaccharide from mycelium of Paecilomyces cicadae. Carbohydr Polym. 2016;136(4):728–737. | ||
Lv HH, Yang AL, Zhang H. HPLC-DAD method for simultaneous determination of 8 nucleosides in Cordyceps cicadae. Pract Pharm Clin Remedies. 2015;18(12):1466–1469. | ||
Wei YL, Yang MF, Liu AY. Analysis of composition and content of fatty acids in Cordyceps ciecadae Shing. Sci Technol Food Industry. 2014;35(5):303–318. | ||
Tyl LW, Zhang P. Determination of active ingredients in Cordyceps cicadae. Jiangsu J Traditional Chin Med. 2006;27(1):45–46. | ||
Wei YL, Yang MF, Liu AY, Zou X. Determination of squalene in Cordyceps cicadae by capillary gas chromatography. Chin J Pharmaceutical Analysis. 2014;34(11):1975–1978. | ||
Fretellier N, Bouzian N, Parmentier N, et al. Nephrogenic systemic fibrosis-like effects of magnetic resonance imaging contrast agents in rats with adenine-induced renal failure. Toxicol Sci. 2013;131(1):259–270. | ||
Guo Y, Jiang Q, Gui D, Wang N. Chinese Herbal Formulas Si-Wu-Tang and Er-Miao-San synergistically ameliorated hyperuricemia and renal impairment in rats induced by adenine and potassium oxonate. Cell Physiol Biochem. 2015;37(4):1491–1502. | ||
Yang P, Song P, Sun SQ, Zhou Q, Feng S, Tao JX. Differentiation and quality estimation of Cordyceps with infrared spectroscopy. Spectrochim Acta A Mol Biomol Spectrosc. 2009;74(4):983–990. | ||
Ding HY, Liu Y, Yu SJ, Zhang LW, Li CR, Fan MZ. Studies on the volatile components of artificial culture of Cordyceps formosana. Food Ferment Ind. 2012;38(6):171–176. | ||
Zong SY, Han H, Wang B, et al. Fast simultaneous determination of 13 nucleosides and nucleobases in Cordyceps sinensis by UHPLC-ESI-MS/MS. Molecules. 2015;20(12):21816–21825. | ||
Wang JB, Ouyang Z, Zhao M, et al. Optimization of extraction proof polysaccharides from Cordyceps cicadae by response surface methodology. Nat Product Res Devel. 2014;26:438–443. | ||
Hashimoto H, Hirose S, Kano S, Tsunematsu T, Kabasawa K. Studies on clinical subsets and severity of systemic lupus erythematosus based on a 1987 questionnaire conducted in Japan – clinical analysis of the outcome and treatments in clinical subsets. Ryumachi. 1992;32(1):27–38. | ||
Wang M. Efficacy analysis on chronic renal failure at different stages treated with TCM enema and high-position sequential colon dialysis. World J Integr Tradit West Med. 2016;02:235–237. | ||
Ma N, Xu HB. Progress on toxicology of diisobutyl phthalate. Chin J Food Hygiene. 2010;22(5):467–470. | ||
Chiu CH, Chyau CC, Chen CC, Lin CH, Cheng CH, Mong MC. Polysaccharide extract of Cordyceps sobolifera attenuates renal injury in endotoxemic rats. Food Chem Toxicol. 2014;69:281–288. | ||
Ikeda R, Nishimura M, Sun Y, Wada M, Nakashima K. Simple HPLC-UV determination of nucleosides and its application to the authentication of Cordyceps and its allies. Biomed Chromatogr. 2008;22(6):630–636. | ||
Guo FQ, Li A, Huang LF, Liang YZ, Chen BM. Identification and determination of nucleosides in Cordyceps sinensis and its substitutes by high performance liquid chromatography with mass spectrometric detection. J Pharm Biomed Anal. 2006;40(3):623–630. | ||
Du CW, Zhou JM, Liu JF. Identification of Chinese medicinal fungus Cordyceps sinensis by depth-profiling mid-infrared photoacoustic spectroscopy. Spectrochim Acta A Mol Biomol Spectrosc. 2017;173:489–494. | ||
Wong YL, Wong KL, Shaw PC. Rapid authentication of Cordyceps by lateral flow dipstick. J Pharm Biomed Anal. 2015;111:306–310. | ||
Lam KY, Chan GK, Xin GZ, et al. Authentication of Cordyceps sinensis by DNA analyses: comparison of ITS sequence analysis and RAPD-derived molecular markers[J]. Molecules. 2015;20(12):22454–22462. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.