Back to Journals » Research and Reports in Tropical Medicine » Volume 8
Potential for sublethal insecticide exposure to impact vector competence of Aedes albopictus (Diptera: Culicidae) for dengue and Zika viruses
Authors Richards SL ![]() , White AV, Balanay JAG
, White AV, Balanay JAG ![]()
Received 28 January 2017
Accepted for publication 3 May 2017
Published 29 May 2017 Volume 2017:8 Pages 53—57
DOI https://doi.org/10.2147/RRTM.S133411
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Thomas Unnasch
Stephanie L Richards, Avian V White, Jo Anne G Balanay
Department of Health Education and Promotion, College of Health and Human Performance, East Carolina University, Greenville, NC, USA
Abstract: Chikungunya, dengue, and Zika viruses (CHIKV, family Togaviridae, genus Alphavirus; DENV and ZIKV, family Flaviviridae, genus Flavivirus) are arboviruses that cause human epidemics. Due to the lack of vaccines for many mosquito-borne diseases, there is a need for mosquito control. In the US and other regions, residual barrier insecticide sprays are applied to foliage where female mosquitoes rest and/or sugar feed between blood meals. Residual sprays are an important control method for anthropogenic day-active sylvan mosquitoes such as Aedes albopictus (vector of CHIKV, DENV, and ZIKV) that are difficult to control using ultralow-volume sprays applied only at dusk or dawn when these mosquitoes are not active. In this exploratory study, we analyzed the extent to which ingestion of a sublethal dose of the active ingredient bifenthrin affected vector competence (i.e., infection, dissemination, and transmission) of Ae. albopictus for DENV and ZIKV. Two incubation periods (IPs; 7 and 14 d) were tested at 28°C for insecticide-fed and sugar-fed mosquitoes. We show that mosquitoes that were fed bifenthrin (0.128 µg/mL) mixed with sucrose solution exhibited significantly lower DENV infection rates and body titers after a 14-d IP. During the 7-d IP, one mosquito (sugar group) transmitted ZIKV. During the 14-d IP, 100% of mosquitoes showed body and leg ZIKV infections, and one mosquito (sugar+bifenthrin group) transmitted ZIKV. This is a preliminary communication, and more studies will be required to further investigate these findings. We expect the findings of this small-scale study to spur larger-scale investigations of the impacts of insecticides on mechanisms regulating vector competence, and exposure to other active ingredients, and aid in development of new insecticides.
Keywords: mosquito, insecticide exposure, arbovirus
Introduction
Chikungunya, dengue, and Zika viruses (CHIKV, family Togaviridae, genus Alphavirus; DENV and ZIKV, family Flaviviridae, genus Flavivirus) cause human epidemics across the world. In recent years, DENV has become more important than malaria in morbidity and economic impact, and several recent ZIKV outbreaks have affected humans, with particular concern for development of microcephaly in infants.1–4 The vectors of both arboviruses are Aedes aegypti L. and Aedes albopictus Skuse.5 It has been shown that CHIKV has increased its global spread, in part, due to the high degree of vector competence in Ae. albopictus.6,7
Residual insecticide barrier sprays are increasingly used for mosquito control in conjunction with, or instead of, ultralow-volume applications with no residual effects. The effects of residual sprays can persist on foliage for up to 21 d, but degrade over time, depending on environmental conditions.8 Mechanisms for insecticide resistance (e.g., overproduction of detoxification enzymes) can lower energy reserves and reduce immune functions that may impact vector competence and fitness.9–11 Mosquito populations may develop resistance due to exposure to sublethal doses of insecticides; therefore, more information is needed to understand how insecticides may influence vector competence. While both Ae. aegypti and Ae. albopictus oviposit in natural and artificial water-holding containers within human dwellings, Ae. albopictus is found in sylvan environments, thereby making evaluation of barrier insecticides that are applied to foliage of adult resting sites important. One US study showed dichlorodiphenyltrichloroethane (DDT)-resistant populations of Ae. albopictus.12 A separate study carried out by us showed possible resistance and/or resistance in Ae. albopictus, Culex pipiens, and Culex quinquefasciatus from several US regions exposed to bifenthrin, deltamethrin, etofenprox, malathion, permethrin, and phenothrin.13
Ae. aegypti adults were exposed to malathion-impregnated papers prior to exposure to a DENV-infected blood meal.14 The same study found no difference in infection rates (whole body) between treatment and control groups at 10 d postinfection. When insecticide-resistant Anopheles gambiae infected with Plasmodium falciparum were exposed to deltamethrin-treated bednets, they showed reduced parasite loads, compared to mosquitoes exposed to untreated bednets.15 Others have shown that insecticide-resistant mosquitoes infected with P. falciparum are more susceptible to DDT than uninfected mosquitoes.10 Nutritional and larvicide (Bacillus thuringiensis israelensis [B.t.i.]) stress reduced occurrence of P. falciparum in An. gambiae, but did not decrease parasite concentration.16 The exposure of Ae. aegypti larvae to sublethal concentrations of B.t.i. did not impact adult survivorship or DENV infection or dissemination rates; however, adults from B.t.i.-fed larvae laid more eggs, compared to the control.17 The same study showed that B.t.i.-fed mosquitoes developed faster than the control mosquitoes. There is clearly a link between insecticide exposure, mosquito infection, and potentially, mosquito fitness that should be explored.
There are limited published vector competence studies for Ae. albopictus and ZIKV, especially for US populations. There is no vaccine for ZIKV, and although there is a vaccine for DENV, it is not yet at sufficient production capacity and does not protect all age groups in the population.18,19 Thus, protection from mosquito bites and targeted vector control are of utmost importance. The anthropogenic nature of the DENV and ZIKV transmission cycle, along with international travel and vector range expansion, necessitates that health agencies prepare for outbreaks. We investigate the extent to which mosquito–insecticide interactions impact vector competence for Ae. albopictus infected with DENV and ZIKV, thereby providing practical information to improve control of a primary vector.
Materials and methods
Insecticide detection in stock
A standard solution of bifenthrin was prepared by dissolving 0.01 g of technical-grade active ingredient (AI; Sigma-Aldrich, St. Louis, MO, USA) in acetone (40 mL) and used to prepare the calibration curve for quantification. The concentration of the prepared AI (bifenthrin) stock was confirmed in order to verify the bifenthrin dose being fed to mosquitoes by analyzing three to four replicate samples (1 µL) of the stock solution via capillary gas chromatography with flame ionization detector (GC-FID; Agilent GC 6850 instrument; Agilent Technologies, Santa Clara, CA, USA). The GC conditions used were adapted from our previous study.8 The linearity of the detector response was checked before conducting analysis by using the calibration curves.
Mosquitoes and insecticide treatment
We used an Ae. albopictus (F22) colony originating from Louisiana. We tested the insecticide resistance/susceptibility status of this population in accordance with the Centers for Disease Control (CDC) bottle bioassay protocol.20 Our findings showed 100% mortality in this Ae. albopictus colony after exposure to a diagnostic dose of 12.6 µg bifenthrin/bottle at a diagnostic time of 30 min.
To improve feeding rate and increase sample size, mosquitoes were deprived of sucrose and fed only water for 48 h prior to the experiments. To test the effects of insecticide ingestion on vector competence, mosquitoes in the insecticide group were fed a solution containing 0.128 µg/mL bifenthrin mixed with a 20% sucrose solution for a feeding period of 4 h. Mosquitoes were observed feeding on the sucrose solution in both treatment and control groups. The concentration of bifenthrin was verified by GC-FID using our previously established methods.8 Mosquitoes in the control group were fed a 20% sucrose solution containing no insecticide for 4 h. After this period, the sucrose solution was removed from both groups and replaced with water until infectious blood meals were offered to mosquitoes 48 h later.
Vector competence evaluation
Vector competence assays were conducted using our previously established methods.21 Briefly, the mosquitoes were assayed for DENV and ZIKV during 7- and 14-d IPs. A Southeast Asian DENV-2 isolate (16803) and a Puerto Rican ZIKV isolate (PRVABC59) were used for infecting mosquitoes. Viral cultures were propagated for use in blood meals according to standard procedures used in our laboratory.22 Freshly propagated DENV and ZIKV were mixed individually with defibrinated bovine blood (Hemostat, Dixon, CA, USA) prior to mosquito feeding. Mosquitoes were blood fed in 1-L cardboard cages with mesh screening and held at 28°C with 80% humidity in a 14:10 light:dark cycle before being transferred to incubators at 28°C for the duration of the experiments. Samples of the blood meal were stored at −80°C until further processing to determine viral titer. Mosquitoes (8- to 9-d old) were fed for 45 min on pledgets soaked with warmed (35°C) DENV- or ZIKV-infected blood at a dose of 5.4 log10 plaque-forming unit equivalents/mL. Mosquitoes were anesthetized with cold, and each fully engorged blood-fed female was transferred to a 1-L cage (according to treatment) for the duration of the experiment.
At the end of 7- and 14-d IP, 5–10 live mosquitoes from each group were anesthetized with cold, and legs and wings were removed following standard methods used in our laboratory.23 Legs were transferred to sample tubes containing 0.5 mL of RNAlater and two 5-mm glass beads and coded for each mosquito. Mosquitoes were allowed to expectorate saliva into capillary tubes filled with a 1:1 mixture of fetal bovine serum and 20% sucrose for 30 min using our methods as previously described.23 Following the salivation period, bodies and saliva samples were transferred to separate tubes categorized by mosquito code containing 0.5 mL of RNAlater and two 5-mm glass beads, and all were stored at −80°C until further processing.
Virus assay
Samples were homogenized and centrifuged as described previously.22 Nucleic acids were extracted using the QIAmp viral RNA kit (Qiagen, Valencia, CA, USA). The amount of viral RNA in each sample was determined using quantitative real-time TaqMan reverse transcriptase polymerase chain reaction (qRT-PCR) with the LightCycler® 480 Instrument (Roche, Mannheim, Germany) and the Superscript III One-Step qRT-PCR kit (Thermo Fisher Scientific, Waltham, MA, USA) programmed as follows: 48°C for 30 min, 95°C for 2 min, and 45 cycles at 95°C for 15 s, 60°C for 30 s, and finally, 40°C for 30 s. The following primers/probes were used: ZIKV 1163c: 5′-CCACTAACGTTCTTTTGCAGACAT-3′, ZIKV 1087: 5′-CCGCTGCCCAACACAAG-3′, ZIKV 1108: 5′-/56-FAM/AGCCTACCT/ZEN/TGACAAGCAGTCAGACACTCAA/3IABkfq/-3′; DENV 237: 5′-CATGGCCCTKGTGGCG-3′, DENV 305: 5′-CCCCATCTYTTCAGTATCCCTG-3′, DENV 254: 5′-/56-FAM/TCCTTCGTTTCCTAACAATCC/3BHQ_2/-3′ (IDTDNA, Coralville, IA, USA). Standard curves were based on plaque assays used to determine titer. The infection rate was the percentage of all blood-fed mosquitoes that had infected bodies. The dissemination rate was the percentage of blood-fed mosquitoes that had infected legs. The transmission rate was the percentage of blood-fed mosquitoes that had infected saliva.
Statistical analysis
The chi-square test was used to detect significant differences in rates of infection, dissemination, and transmission between groups (P<0.05). Nonparametric analysis of variance was used to evaluate differences in titers of body and leg tissues and saliva between IPs and treatments. If significant differences were observed, then a Duncan test was used to determine differences in the means.
Results
Effects of insecticide ingestion on vector competence
During the 7-d IP, we observed 80%–90% DENV infection rate and 10%–30% DENV dissemination rate in both experimental groups (Table 1). Mosquitoes that were fed bifenthrin (0.128 µg/mL) exhibited statistically equivalent DENV infection rates (X2=5.49, df=1, P=0.057) and significantly lower body titers (F=6.83, df=1, P=0.024) after a 14-d IP, as compared to mosquitoes fed only sugar. No DENV transmission was observed during the 7- or 14-d IP.
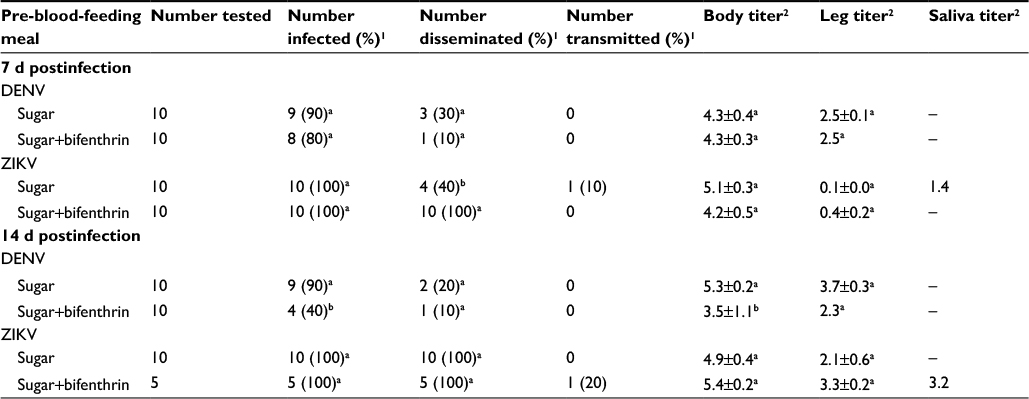 | Table 1 Mean titers (log10 plaque-forming unit equivalents [PFUeq] DENV/mL or ZIKV/mL)±standard error of rate of infection, dissemination, and transmission for Aedes albopictus stratified by pre-blood-feeding meal, virus type, and incubation period Notes: Analyses were conducted separately for each virus. 1Same letter in the same column for each virus indicates no significant difference between treatments by chi-square test. 2Same letter in the same column indicates no significant difference between treatments by means comparison. |
During both the 7- and 14-d IP, 100% of mosquitoes showed ZIKV body infections. Mosquitoes in the bifenthrin group showed 100% dissemination rates during both the 7- and 14-d IP; however, mosquitoes fed only sugar prior to the ZIKV-infected blood meal showed 40% and 100% dissemination during the 7- and 14-d IP, respectively. Two mosquitoes transmitted ZIKV (sugar group – 7-d IP; sugar+bifenthrin group – 14-d IP; Table 1).
Discussion
Research aimed at elucidating the effects of insecticide–mosquito interactions has largely been focused on vector control and development of resistance. Little attention has been given to the impacts of insecticides on vector competence. Insecticide pressure on mosquito populations is a continuing threat as mosquito control is the primary method of protecting humans from mosquito-borne diseases.
Ae. albopictus is one of the primary vectors of DENV and CHIKV, is a competent vector in the field24 and laboratory,6,7,21,25,26 and feeds on the blood of humans and other animals.27 A study in Gabon (Central Africa) showed that 137 and 45 Ae. albopictus pools were positive for ZIKV and DENV, respectively, although transmission was not tested in these cases.28 Another study showed transmission (capillary tube assay) of ZIKV by several Aedes spp., including Ae. aegypti.29 The same study did not evaluate ZIKV vector competence in Ae. albopictus.
We have shown infection, dissemination, and transmission of DENV26 and ZIKV (current study) for Ae. albopictus. After a 14-d IP, Ae. albopictus that ingested bifenthrin showed increased ZIKV transmission rate and decreased DENV infection rate. More information is needed to elucidate these differences between viruses and insecticide groups as the current exploratory study had small sample sizes. Additional experiments are planned to explore these differences at a larger scale. Sublethal doses of insecticides obtained (by contact and/or ingestion) from foliage utilized in barrier sprays may impact gut bacteria, the mosquito immune response, or affect other unknown factors that may impact subsequent vector competence.
We have previously shown that, after field application of bifenthrin, leaf surfaces contain 0–25.6 µg/mL,8 and the current study exposed Ae. albopictus to 0.128 µg/mL of bifenthrin. Future studies should assay a variety of bifenthrin doses in this range (0–25.6 µg/mL) from sublethal to lethal concentrations. Exposure of mosquitoes to bottles (as in the CDC bottle bioassay) coated with active ingredients of insecticide would be another alternative to insecticide ingestion, and we are planning to investigate this method of insecticide exposure. Our findings that ZIKV is transmitted at 7 d (one mosquito in the sugar-only group) and at 14 d (one mosquito in the sugar+bifenthrin group) should be interpreted with caution due to our sample sizes of 5–10 mosquitoes in each treatment group used in this exploratory experiment. Larger-scale studies with increased sample size should be planned to further characterize the vector competence of different populations of Ae. albopictus. This is a preliminary communication, and more studies are needed to investigate this aspect of vector competence.
Acknowledgment
A poster of this paper was presented at the East Carolina University World Environmental Health Day in September 2016, and the abstract with interim findings can be viewed at: http://thescholarship.ecu.edu/handle/10342/5963.
Disclosure
The authors report no conflicts of interest in this work.
References
Gubler DJ. The economic burden of dengue. Am J Trop Med Hyg. 2012;86(5):743–744. | ||
Halasa YA, Shepard DS, Zeng W. Economic cost of dengue in Puerto Rico. Am J Trop Med Hyg. 2012;86(5):745–752. | ||
Plourde AR, Bloch EM. A literature review of Zika virus. Emerg Infect Dis. 2016;22(7):1185–1192. | ||
Brasil P, Pereira JP Jr, Moreira ME, et al. Zika virus infection in pregnant women in Rio de Janeiro. N Engl J Med. 2016;375(24):2321–2334. | ||
Gardner LM, Chen N, Sarkar S. Global risk of Zika virus depends critically on vector status of Aedes albopictus. Lancet Infect Dis. 2016;16(5):522–523. | ||
Vega-Rua A, Zouache K, Caro V, et al. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the Southeast of France. PLoS One. 2013;8(3):e59716. | ||
Vega-Rúa A, Zouache K, Girod R, Failloux AB, Lourenço-de-Oliveria R. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of Chikungunya virus. J Virol. 2014;88(11):6294–6306. | ||
VanDusen AE, Richards SL, Balanay JA. Evaluation of bifenthrin barrier spray on foliage in a suburban eastern North Carolina neighborhood. Pest Manag Sci. 2016;72(5):1004–1012. | ||
Rivero A, Vézilier J, Weill M, Read AF, Gandon S. Insecticide control of vector-borne diseases: when is insecticide resistance a problem? PLoS Pathog. 2010;6(8):e1001000. | ||
Alout H, Ndam NT, Sandeu MM, et al. Insecticide resistance alleles affect vector competence of Anopheles gambiae s.s. for Plasmodium falciparum field isolates. PLoS One. 2013;8(5):e63849. | ||
Chan HH, Zairi J. Permethrin resistance in Aedes albopictus (Diptera: Culicidae) and associated fitness costs. J Med Entomol. 2013;50(2):362–370. | ||
Marcombe S, Farajollahi A, Healy SP, Clark GG, Fonseca DM. Insecticide resistance status of United States populations of Aedes albopictus and mechanisms involved. PLoS One. 2014;9(7):e101992. | ||
Richards SL, Balanay JAG, Fields M, Vandock K. Baseline insecticide susceptibility screening against six active ingredients for Culex and Aedes (Diptera: Culicidae) mosquitoes in the United States. J Med Entomol. 2017;54(3):682–695. | ||
Lee HL, Argubano RA, Ahmad R. Effect of sublethal dosages of malathion on the oral susceptibility of Aedes aegypti to dengue-2 virus infection. Southeast Asian J Trop Med Public Health. 1997;28(3):664–665. | ||
Kristan M, Lines J, Nuwa A, Ntege C, Meek SR, Abeku TA. Exposure to deltamethrin affects development of Plasmodium falciparum inside wild pyrethroid resistant Anopheles gambiae s.s. mosquitoes in Uganda. Parasit Vectors. 2016;9:100. | ||
Vantaux A, Ouattarra I, Lefèvre T, Dabiré KR. Effects of larvicidal and larval nutritional stresses on Anopheles gambiae development, survival, and competence for Plasmodium falciparum. Parasit Vectors. 2016;9:226. | ||
Alto BW, Lord CC. Transstadial effects of Bti on traits of Aedes aegypti and infection with dengue virus. PLoS Negl Trop Dis. 2016;10(2):e0004370. | ||
Capeding MR, Tran NH, Hadinegoro SR; CYD14 Study Group. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: a phase 3, randomized, observer-masked, placebo-controlled trial. Lancet. 2014;384(9951):1358–1365. | ||
Sáez-Llorens X, Tricou V, Yu D, et al. Safety and immunogenicity of one versus two doses of Takeda’s tetravalent dengue vaccine in children in Asia and Latin America: interim results from a phase 2, randomised, placebo-controlled study. Lancet Infect Dis. Epub 2017 Mar 29. | ||
Centers for Disease Control and Prevention. CDC bottle bioassay. 2015. Available from: http://www.cdc.gov/parasites/education_training/lab/bottlebioassay.html. Accessed May 9, 2016. | ||
Richards SL, Anderson SL, Alto BW. Vector competence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) for dengue virus in the Florida Keys. J Med Entomol. 2012;49(4):942–946. | ||
Richards SL, Pesko K, Alto BW, Mores CN. Reduced infection in mosquitoes exposed to blood meals containing previously frozen flaviviruses. Virus Res. 2007;129(2):224–227. | ||
Anderson SL, Richards SL, Smartt CT. A simple method for examining arbovirus transmission in mosquitoes. J Am Mosq Control Assoc. 2010;26(1):108–111. | ||
Effler PV, Pang L, Kitsutani P; Hawaii Dengue Outbreak Investigation Team. Dengue fever, Hawaii, 2001–2002. Emerg Infect Dis. 2005;11(5):742–749. | ||
Fernández Z, Moncayo A, Forattini OP, Weaver SC. Susceptibility of urban and rural populations of Aedes albopictus from São Paulo, Brazil, to infection by dengue-1 and -2 viruses. J Med Entomol. 2004;41(5):961–964. | ||
van Dodewaard CA, Richards SL, Harris JW. Effects of blood coagulate removal method on Aedes albopictus (Diptera: Culicidae) life table characteristics and vector competence for dengue virus. J Med Entomol. 2016;53(1):39–47. | ||
Richards SL, Ponnusamy L, Unnasch TR, Hassan HK, Apperson CS. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in relation to availability of human and domestic animals in suburban landscapes of central North Carolina. J Med Entomol. 2006;43(3):543–551. | ||
Grard G, Caron M, Mombo IM, et al. Zika virus in Gabon (Central Africa) – 2007: a new threat from Aedes albopictus? PLoS Negl Trop Dis. 2014;8(2):e2681. | ||
Diagne CT, Diallo D, Faye O, et al. Potential of selected Senegalese Aedes spp. mosquitoes (Diptera: Culicidae) to transmit Zika virus. BMC Infect Dis. 2015;15:492. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.