Back to Journals » Cancer Management and Research » Volume 10
Polymorphisms in MYCN gene and neuroblastoma risk in Chinese children: a 3-center case–control study
Authors Zhou H ![]() , Zhuo Z, Chen S, Zhao J
, Zhuo Z, Chen S, Zhao J ![]() , Mo Y
, Mo Y ![]() , Zhang J, He J, Ruan J
, Zhang J, He J, Ruan J
Received 17 March 2018
Accepted for publication 28 April 2018
Published 2 July 2018 Volume 2018:10 Pages 1807—1816
DOI https://doi.org/10.2147/CMAR.S168515
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Haixia Zhou,1,* Zhenjian Zhuo,2,* Shanshan Chen,1 Jie Zhao,1 Yixiao Mo,1 Jiao Zhang,3 Jing He,1,4 Jichen Ruan1
1Department of Hematology, The Second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, China; 2School of Chinese Medicine, Faculty of Medicine, The Chinese University of Hong Kong, Hong Kong; 3Department of Pediatric Surgery, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, China; 4Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong, China
*These authors contributed equally to this work
Introduction: Neuroblastoma is an embryonal tumor of the sympathetic nervous system. The MYCN oncogene is amplified in some neuroblastoma patients and correlated with poor prognosis. However, less is known regarding the relationship between MYCN gene single-nucleotide polymorphisms (SNPs) and neuroblastoma risk.
Patients and methods: To investigate the contribution of MYCN gene polymorphisms to neuroblastoma risk, we performed a 3-center case–control study by genotyping 4 SNPs in the MYCN gene from 429 cases and 884 controls.
Results: The results showed that only rs57961569 G>A was associated with neuroblastoma risk (GA vs GG: adjusted odds ratio =0.76, 95% confidence interval =0.60–0.98, P=0.033), while the other 3 SNPs were not (rs9653226 T>C, rs13034994 A>G, and rs60226897 G>A). Stratified analysis revealed that rs57961569 GG carriers were more likely to develop neuroblastoma in the following subgroups: children older than 18 months, tumor derived from the adrenal gland, and clinical stages III + IV. The increased neuroblastoma risk associated with the rs9653226 variant CC genotypes was more evident in the following subgroups: females, tumor derived from the adrenal gland, and clinical stages III + IV. The presence of 2–3 risk genotypes had a significant relationship with the following subgroups: tumor derived from the adrenal gland and clinical stages III + IV.
Conclusion: This study demonstrates a weak impact of MYCN gene polymorphisms on neuroblastoma risk, which should be further validated.
Keywords: neuroblastoma, susceptibility, MYCN, polymorphism
Introduction
Neuroblastoma, the most common extracranial solid tumor in childhood, develops from neural crest progenitor cells.1,2 Neuroblastoma accounts for ~7% of all childhood malignancies, yet is responsible for 15% of all pediatric oncology deaths.3–5 The prognosis of neuroblastoma patients is widely variable. Some patients may spontaneously regress without chemotherapy, while some relapse with therapy-resistant disease.6 Neuroblastoma cases are generally classified into low-, intermediate-, and high-risk groups.7 Among them, high-risk neuroblastoma patients comprise ~50% of all neuroblastoma cases.8 The 5-year survival rates of high-risk neuroblastoma cases seldom exceed 40% despite intensive, multimodal therapy.9 The difficulty in treating these high-risk neuroblastoma cases might be the distant metastasis of tumors to bone marrow.7,10
The etiology of neuroblastoma has been partly elucidated but remains obscure. Previous studies have indicated that environmental factors may influence risk of neuroblastoma; however, the theory lacks direct evidence.11,12 Growing attention has been directed to genetic and gene–environmental factors as underlying risks of neuroblastoma. Currently, numerous genetic variants have been determined to play critical roles in neuroblastoma carcinogenesis. For example, PHOX2B13,14 and ALK15,16 gene mutations were found in some neuroblastoma cases. Genome-wide association studies have also identified polymorphisms located in TP53, HACE1, BARD1, LIN28B, LMO1, and CASC15 genes associated with neuroblastoma risk.1,17,18
MYCN, a member of the MYC protooncogene family, was first identified in neuroblastoma cell lines.19 MYCN is a critical regulator of various cellular processes, including proliferation, differentiation, and apoptosis.20–23 Moreover, aberration expression of MYCN is associated with tumor initiation and progression.24–26 Amplification of MYCN is present in ~20% of neuroblastoma cases.27 MYCN is closely associated with aggressive tumor type and poor prognosis.28,29 Therefore, exploring the role of MYCN in carcinogenesis remains a research hotspot. However, few studies have been proposed to evaluate the association of MYCN gene polymorphisms and neuroblastoma risk. Given the importance of the MYCN gene in cancer initiation and development, we intend to explore whether MYCN gene polymorphisms could predispose a patient to neuroblastoma risk.
Patients and methods
Participants
A total of 429 neuroblastoma cases and 884 healthy controls from 3 centers were included in this study.30–32 All of the cases were newly confirmed and histopathologically diagnosed neuroblastoma patients without progressive disease or previous treatments. The controls were age-, gender-, and ethnicity-matched to cases and were randomly recruited from children undergoing routine medical examination at the same hospital during the same period. All of the enrolled subjects were of Chinese Han ethnicity. More specifically, 36 cases and 72 controls were enrolled from The Second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University (Table S1), 275 cases and 531 controls were enrolled from Guangzhou Women and Children’s Medical Center, and the remaining 118 cases and 281 controls were recruited from The First Affiliated Hospital of Zhengzhou University. Written informed consent was obtained from all of the participants’ parents. Details of the selection process of the included participants were provided in our previous publication.33–35 The institutional review board of the abovementioned 3 hospitals approved the study.
Single-nucleotide polymorphism (SNP) selection and genotyping
We chose potentially functional SNPs of interest from the dbSNP database (http://www.ncbi.nlm.nih.gov/) based on the following criteria: 1) minor allele frequency >5% for Chinese Han subjects; 2) potentially functional, such as affecting the binding capacity of transcription factor or microRNA binding sites, or leading to amino acid alterations. We chose 4 potentially functional SNPs in the MYCN gene (rs57961569 G>A, rs9653226 T>C, rs13034994 A>G, and rs60226897 G>A) for analysis. As predicted by SNPinfo (http://snpinfo.niehs.nih.gov/), all of them were located in the transcription factor binding sites. Moreover, they all had a minor allele frequency >5% in Chinese Han subjects. As shown in Figure S1, there was no significant linkage disequilibrium (R2<0.8) among these 4 SNPs (R2=0.220 between rs57961569 and rs9653226; R2=0.185 between rs57961569 and rs13034994; R2=0.103 between rs57961569 and rs60226897; R2=0.348 between rs9653226 and rs13034994; R2=0.161 between rs9653226 and rs60226897; R2=0.500 between rs13034994 and rs6022689). First, the genomic DNA was extracted from the peripheral blood donated by subjects. The MYCN gene polymorphisms were detected using TaqMan real-time PCR, as described elsewhere.36–39 Eight blank wells containing water as negative controls were also placed in each 384-well plate as the means of quality control. In addition, we randomly regenotyped 10% of the samples and got a concordance rate of 100%.
Statistical analysis
Testing of Hardy–Weinberg equilibrium for the selected polymorphisms in controls was performed using a goodness-of-fit χ2 test. A 2-sided χ2 test was used for comparisons of 2 groups in allele frequencies and demographic variables. We evaluated the associations between genotypes and neuroblastoma risk using odds ratios (ORs) and 95% confidence intervals (CIs) calculated from logistic regression analysis. Risk genotypes were rs57961569 GG, rs9653226 CC, rs13034994 GG, and rs60226897 GG; covariates, including age and gender, were used for adjustment. Statistical analysis was performed using SAS software version 9.4 (SAS Institute, Cary, NC, USA). P-values <0.05 were considered significant.
Results
Population characteristics
The population demographics for the Guangzhou and Zhengzhou subjects were presented in our former publication.30–32 The population demographics for the Wenzhou subjects are shown in Table S1, while those of the combined subjects are shown in Table S2. No statistically significant differences were observed between neuroblastoma cases and controls regarding age (P=0.229, P=0.484, P=0.496, P=0.119) and gender (P=0.510, P=0.196, P=1.000, P=0.840) for the Guangdong, Henan, Wenzhou, and combined subjects, respectively.
Correlation of MYCN gene polymorphisms with neuroblastoma susceptibility
The genotype frequencies of MYCN gene polymorphisms and neuroblastoma susceptibility between all cases and controls for combined subjects and divided subjects are shown in Tables 1 and S3, respectively. All of the 4 genotyped MYCN SNPs in controls conformed to the Hardy–Weinberg equilibrium for combined subjects (rs57961569 G>A, P=0.379; rs9653226 T>C, P=0.569; rs13034994 A>G, P=0.907; and rs60226897 G>A, P=0.526). In single genotype analysis, only 1 SNP rs57961569 G>A was associated with neuroblastoma risk (GA vs GG: adjusted OR [AOR)] =0.76, 95% CI =0.60–0.98, P=0.033). No statistically significant associations were found between the other 3 SNPs (rs9653226 T>C, rs13034994 A>G, and rs60226897 G>A) and neuroblastoma risk. We found that compared with subjects carrying 0–1 risk genotypes, subjects carrying 2–3 combined risk genotypes of MYCN exhibited enhanced risk of neuroblastoma, but this finding was nonsignificant (AOR =1.25; 95% CI =0.99–1.58).
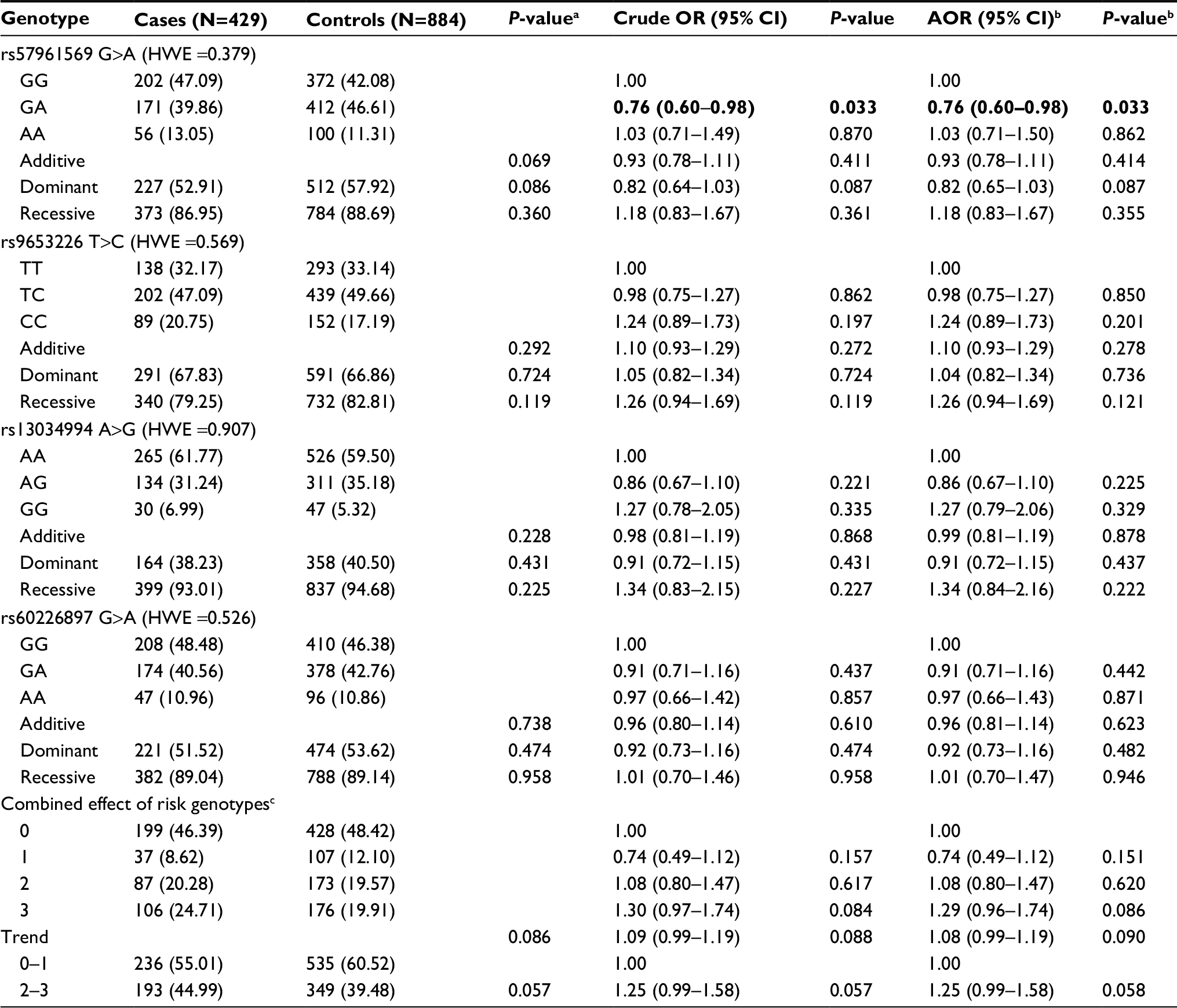 | Table 1 Genotype frequencies of MYCN gene polymorphisms and neuroblastoma susceptibility Notes: aχ2 test for genotype distributions between neuroblastoma patients and controls. bAdjusted for age and gender. cRisk genotypes were rs57961569 GG, rs9653226 CC, rs13034994 GG, and rs60226897 GG. Bold figures indicate 95% CI excluded 1 or P<0.05. Abbreviations: AOR, adjusted odds ratio; CI, confidence interval; HWE, Hardy–Weinberg equilibrium; OR, odds ratio. |
Stratification analysis
Stratification analysis was further conducted to evaluate the effects of different strata (age, gender, tumor sites of origin, and clinical stages) on the association between the selected polymorphisms and neuroblastoma risk (Table 2). Concerning the rs57961569 polymorphism, significant association was detected in the following subgroups: children older than 18 months (GG vs GA: AOR =1.38; 95% CI =1.01–1.88, P=0.040), tumor derived from the adrenal gland (GG vs GA: AOR =1.88; 95% CI =1.30–2.72, P=0.0008), and clinical stages III + IV (GG vs GA: AOR =1.40, 95% CI =1.02–1.92, P=0.037). The conferred increased neuroblastoma risk associated with the rs9653226 variant CC genotypes was more evident in the following subgroups: females (AOR =1.63, 95% CI =1.07–2.48, P=0.023), tumor derived from the adrenal gland (AOR =1.79, 95% CI =1.21–2.63, P=0.003), and clinical stages III + IV (AOR =1.63, 95% CI =1.15–2.31, P=0.006). In the stratified analysis of the cumulative effects of risk genotypes, we found that the presence of 2–3 risk genotypes had a significant relationship with the following subgroups: tumor derived from the adrenal gland (AOR =1.66, 95% CI =1.18–2.32, P=0.003) and clinical stages III + IV (AOR =1.35, 95% CI =1.004–1.81, P=0.047).
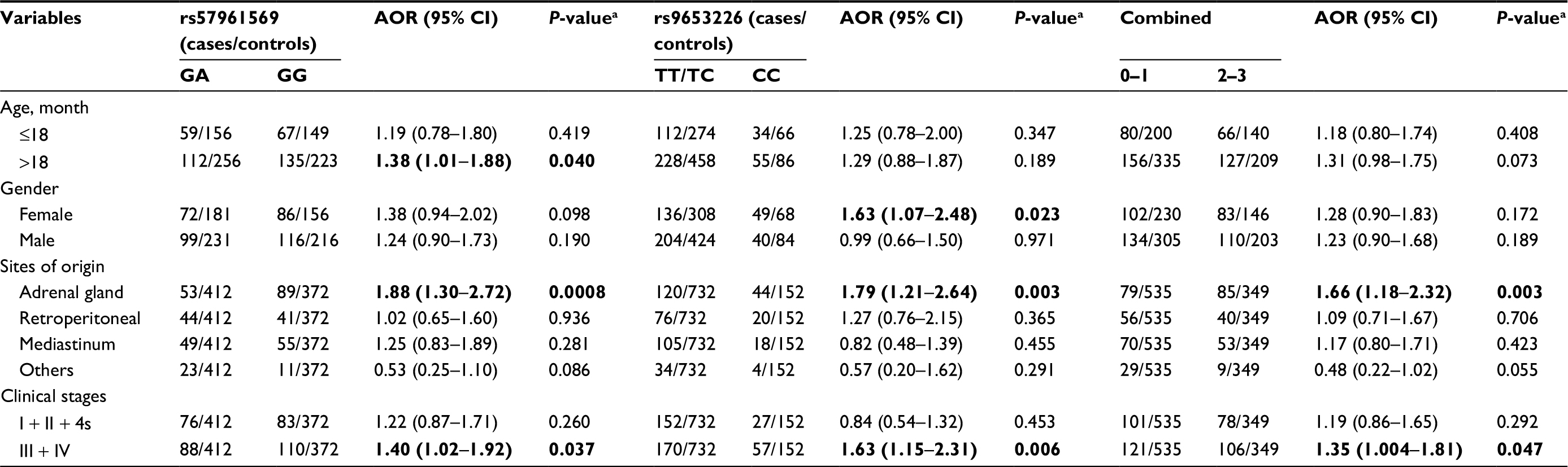 | Table 2 Stratification analysis of risk genotypes with neuroblastoma susceptibility Notes: aAdjusted for age and gender. Bold figures indicate 95% CI excluded 1 or P<0.05. Abbreviations: AOR, adjusted odds ratio; CI, confidence interval. |
Discussion
In the current study, we explored the impact of SNPs of the MYCN gene on the risk of neuroblastoma in the Chinese population. Our data revealed that the rs57961569 G>A polymorphism in the MYCN gene presented significant inverse associations with neuroblastoma risk.
MYCN is located on chromosome 2p24.3 and encodes a pleiotropic nuclear phosphoprotein. The encoding protein consists of 2 domains: a carboxy-terminal DNA-binding and protein interaction domain and an amino-terminal transcriptional activation domain. In healthy conditions in humans and mice, MYCN expression is high in certain tissues in the developing embryo, while it is low or even absent in adult tissues.40,41 Amplification of MYCN could promote proliferation and cell cycle progression. MYCN could also enhance neuroblastoma cell migration and invasion through downregulation of integrins α1 and β1.42,43 Moreover, MYCN modulates antigens on the surface of tumor cells, thus influencing immune surveillance.44 Brandetti et al45 demonstrated that MYCN functions as an immunosuppressive oncogene in neuroblastoma cells by negatively regulating the expression of ligands for DNAM-1and NKG2D NK-cell-activating receptors. Importantly, a study conducted by Dahlin et al46 was the first to investigate the relationship between MYCN gene variants and cancer risk. The study failed to establish a significant relationship between MYCN gene variant rs922 G>A and medulloblastoma risk from 243 cases and 247 controls of Swedish and Danish children. The significance of MYCN in the initiation and development of cancer is self-evident.
Herein, we investigated for the first time whether MYCN gene SNPs could affect the risk of neuroblastoma in Chinese children. Among the 4 analyzed SNPs, only rs57961569 G>A was associated with neuroblastoma risk. In the combined analysis, subjects carrying 2–3 risk genotypes tended to have increased neuroblastoma risk in comparison to those with 0–1 risk genotypes. This phenomenon was quite biologically plausible, as a single polymorphism in each gene may not be strong enough to influence the risk of cancer. Intriguingly, stratified analysis showed that individuals harboring the rs57961569 GG alleles were more likely to have a tumor in the following subgroups: children older than 18 months, tumor derived from the adrenal gland, and clinical stages III + IV. The contributing role of the rs9653226 CC genotype to increasing neuroblastoma risk was more evident in the females, tumor derived from the adrenal gland, and clinical stages III + IV subgroups. In the stratified analysis of the cumulative effects of risk genotypes, we found that the presence of 2–3 risk genotypes had a positive relationship with the following subgroups: tumor derived from the adrenal gland and clinical stages III + IV. On one hand, the same polymorphism might have a different role in cancer risk, depending on different ethnicities, regions, and cancer types. On the other hand, a conflicting role might also be the small sample size in the stratified analysis.
Several limitations accompany the merits of this study. First, the sample size was relatively small, although we gathered samples from 3 centers, especially for the stratification analysis. As a result, the strength of the statistical power may not be strong enough. Second, being a retrospective investigation, only genetic information in the MYCN gene was measured. Measuring other environmental factors, such as childhood exposure, dietary habits, and health situation, is critical in helping to further elucidate the role of MYCN polymorphisms in neuroblastoma risk. Third, only 4 SNPs in the MYCN gene were analyzed, so more polymorphisms in the MYCN gene should be investigated.
Overall, in the Chinese population studied, we provide the first evidence that polymorphisms in MYCN gene could influence neuroblastoma risk in a low-impact manner. Larger sample size studies with additional functional analysis are needed to better elucidate the role of MYCN polymorphisms in neuroblastoma risk.
Acknowledgment
This study was funded by grants from the Pearl River S&T Nova Program of Guangzhou (No: 201710010086), the Scientific Research Foundation of Wenzhou (No: 2015Y0492), the Zhejiang Provincial Medical and Health Science and Technology plan (No: 2009A148), and the Zhejiang Provincial Science and Technology Animal Experimental Platform Project (No: 016C37113).
Disclosure
The authors report no conflicts of interest in this work.
References
Matthay KK, Maris JM, Schleiermacher G, et al. Neuroblastoma. Nat Rev Dis Primers. 2016;2:16078. | ||
Capasso M, Diskin SJ. Genetics and genomics of neuroblastoma. Cancer Treat Res. 2010;155:65–84. | ||
Irwin MS, Park JR. Neuroblastoma: paradigm for precision medicine. Pediatr Clin North Am. 2015;62(1):225–256. | ||
Park JR, Bagatell R, Cohn SL, et al. Revisions to the International Neuroblastoma Response Criteria: A Consensus Statement From the National Cancer Institute Clinical Trials Planning Meeting. J Clin Oncol. 2017;35(22):2580–2587. | ||
Esposito MR, Aveic S, Seydel A, Tonini GP. Neuroblastoma treatment in the post-genomic era. J Biomed Sci. 2017;24(1):14. | ||
Maris JM. Recent advances in neuroblastoma. N Engl J Med. 2010;362(23):2202–2211. | ||
Cheung NK, Dyer MA. Neuroblastoma: developmental biology, cancer genomics and immunotherapy. Nat Rev Cancer. 2013;13(6):397–411. | ||
Schwab M, Westermann F, Hero B, Berthold F. Neuroblastoma: biology and molecular and chromosomal pathology. Lancet Oncol. 2003;4(8):472–480. | ||
Maris JM, Hogarty MD, Bagatell R, Cohn SL. Neuroblastoma. Lancet. 2007;369(9579):2106–2120. | ||
DuBois SG, Kalika Y, Lukens JN, et al. Metastatic sites in stage IV and IVS neuroblastoma correlate with age, tumor biology, and survival. J Pediatr Hematol Oncol. 1999;21(3):181–189. | ||
Cook MN, Olshan AF, Guess HA, et al. Maternal medication use and neuroblastoma in offspring. Am J Epidemiol. 2004;159(8):721–731. | ||
Menegaux F, Olshan AF, Neglia JP, Pollock BH, Bondy ML. Day care, childhood infections, and risk of neuroblastoma. Am J Epidemiol. 2004;159(9):843–851. | ||
Mosse YP, Laudenslager M, Khazi D, et al. Germline PHOX2B mutation in hereditary neuroblastoma. Am J Hum Genet. 2004;75(4):727–730. | ||
Bourdeaut F, Trochet D, Janoueix-Lerosey I, et al. Germline mutations of the paired-like homeobox 2B (PHOX2B) gene in neuroblastoma. Cancer Lett. 2005;228(1–2):51–58. | ||
Chen Y, Takita J, Choi YL, et al. Oncogenic mutations of ALK kinase in neuroblastoma. Nature. 2008;455(7215):971–974. | ||
Mosse YP, Laudenslager M, Longo L, et al. Identification of ALK as a major familial neuroblastoma predisposition gene. Nature. 2008;455(7215):930–935. | ||
Tolbert VP, Coggins GE, Maris JM. Genetic susceptibility to neuroblastoma. Curr Opin Genet Dev. 2017;42:81–90. | ||
He J, Zou Y, Wang T, et al. Genetic variations of GWAS-identified genes and neuroblastoma susceptibility: a replication study in Southern Chinese children. Transl Oncol. 2017;10(6):936–941. | ||
Schwab M, Alitalo K, Klempnauer KH, et al. Amplified DNA with limited homology to myc cellular oncogene is shared by human neuroblastoma cell lines and a neuroblastoma tumour. Nature. 1983;305(5931):245–248. | ||
Kramer M, Ribeiro D, Arsenian-Henriksson M, Deller T, Rohrer H. Proliferation and survival of embryonic sympathetic neuroblasts by MYCN and activated ALK signaling. J Neurosci. 2016;36(40):10425–10439. | ||
Guglielmi L, Cinnella C, Nardella M, et al. MYCN gene expression is required for the onset of the differentiation programme in neuroblastoma cells. Cell Death Dis. 2014;5:e1081. | ||
Westermark UK, Wilhelm M, Frenzel A, Henriksson MA. The MYCN oncogene and differentiation in neuroblastoma. Semin Cancer Biol. 2011;21(4):256–266. | ||
Petroni M, Veschi V, Prodosmo A, et al. MYCN sensitizes human neuroblastoma to apoptosis by HIPK2 activation through a DNA damage response. Mol Cancer Res. 2011;9(1):67–77. | ||
Beckers A, Van Peer G, Carter DR, et al. MYCN-driven regulatory mechanisms controlling LIN28B in neuroblastoma. Cancer Lett. 2015;366(1):123–132. | ||
Stermann A, Huebener N, Seidel D, et al. Targeting of MYCN by means of DNA vaccination is effective against neuroblastoma in mice. Cancer Immunol Immunother. 2015;64(10):1215–1227. | ||
Williams RD, Chagtai T, Alcaide-German M, et al. Multiple mechanisms of MYCN dysregulation in Wilms tumour. Oncotarget. 2015; 6(9):7232–7243. | ||
Thompson D, Vo KT, London WB, et al. Identification of patient subgroups with markedly disparate rates of MYCN amplification in neuroblastoma: a report from the International Neuroblastoma Risk Group project. Cancer. 2016;122(6):935–945. | ||
Brodeur GM, Seeger RC, Schwab M, Varmus HE, Bishop JM. Amplification of N-myc in untreated human neuroblastomas correlates with advanced disease stage. Science. 1984;224(4653):1121–1124. | ||
Seeger RC, Brodeur GM, Sather H, et al. Association of multiple copies of the N-myc oncogene with rapid progression of neuroblastomas. N Engl J Med. 1985;313(18):1111–1116. | ||
He J, Zou Y, Liu X, et al. Association of common genetic variants in pre-microRNAs and neuroblastoma susceptibility: a two-center study in Chinese children. Mol Ther Nucleic Acids. 2018;11:1–8. | ||
Zhang Z, Chang Y, Jia W, et al. LINC00673 rs11655237 C>T confers neuroblastoma susceptibility in Chinese population. Biosci Rep. 2018;38(1):BSR20171667. | ||
Zhuo ZJ, Liu W, Zhang J, et al. Functional polymorphisms at ERCC1/XPF genes confer neuroblastoma risk in Chinese children. EBioMedicine. 2018;30:113–119. | ||
He J, Zhong W, Zeng J, et al. LMO1 gene polymorphisms contribute to decreased neuroblastoma susceptibility in a Southern Chinese population. Oncotarget. 2016;7(16):22770–22778. | ||
He J, Wang F, Zhu J, et al. The TP53 gene rs1042522 C>G polymorphism and neuroblastoma risk in Chinese children. Aging (Albany NY). 2017;9(3):852–859. | ||
He J, Wang F, Zhu J, et al. Association of potentially functional variants in the XPG gene with neuroblastoma risk in a Chinese population. J Cell Mol Med. 2016;20(8):1481–1490. | ||
Gong J, Tian J, Lou J, et al. A polymorphic MYC response element in KBTBD11 influences colorectal cancer risk, especially in interaction with a MYC regulated SNP rs6983267. Ann Oncol. 2017;29(3):632–639. | ||
Li J, Zou L, Zhou Y, et al. A low-frequency variant in SMAD7 modulates TGF-beta signaling and confers risk for colorectal cancer in Chinese population. Mol Carcinog. 2017;56(7):1798–1807. | ||
Lou J, Gong J, Ke J, et al. A functional polymorphism located at transcription factor binding sites, rs6695837 near LAMC1 gene, confers risk of colorectal cancer in Chinese populations. Carcinogenesis. 2017;38(2):177–183. | ||
Zou D, Lou J, Ke J, et al. Integrative expression quantitative trait locus-based analysis of colorectal cancer identified a functional polymorphism regulating SLC22A5 expression. Eur J Cancer. 2018;93:1–9. | ||
Zimmerman KA, Yancopoulos GD, Collum RG, et al. Differential expression of myc family genes during murine development. Nature. 1986;319(6056):780–783. | ||
Grady EF, Schwab M, Rosenau W. Expression of N-myc and c-src during the development of fetal human brain. Cancer Res. 1987;47(11):2931–2936. | ||
van Golen CM, Soules ME, Grauman AR, Feldman EL. N-Myc overexpression leads to decreased beta1 integrin expression and increased apoptosis in human neuroblastoma cells. Oncogene. 2003;22(17):2664–2673. | ||
Tanaka N, Fukuzawa M. MYCN downregulates integrin α1 to promote invasion of human neuroblastoma cells. Int J Oncol. 2008;33(4):815–821. | ||
Song L, Ara T, Wu HW, et al. Oncogene MYCN regulates localization of NKT cells to the site of disease in neuroblastoma. J Clin Invest. 2007;117(9):2702–2712. | ||
Brandetti E, Veneziani I, Melaiu O, et al. MYCN is an immunosuppressive oncogene dampening the expression of ligands for NK-cell-activating receptors in human high-risk neuroblastoma. Oncoimmunology. 2017;6(6):e1316439. | ||
Dahlin AM, Hollegaard MV, Wibom C, et al. CCND2, CTNNB1, DDX3X, GLI2, SMARCA4, MYC, MYCN, PTCH1, TP53, and MLL2 gene variants and risk of childhood medulloblastoma. J Neurooncol. 2015;125(1):75–78. |
Supplementary materials
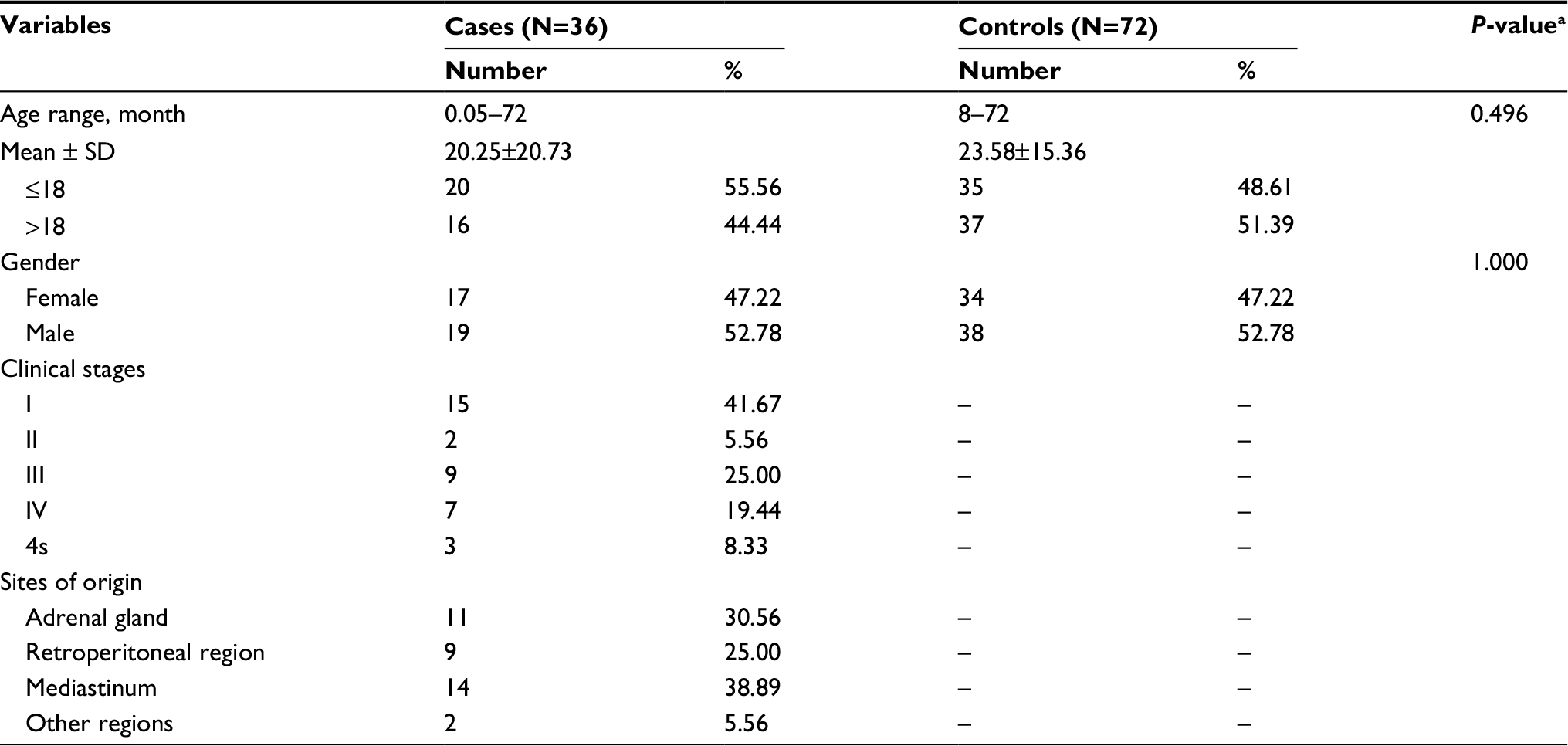 | Table S1 Frequency distribution of demographic characteristics for Wenzhou subjects Notes: aTwo-sided χ2 test for distributions between neuroblastoma patients and controls. “–” indicates no value. |
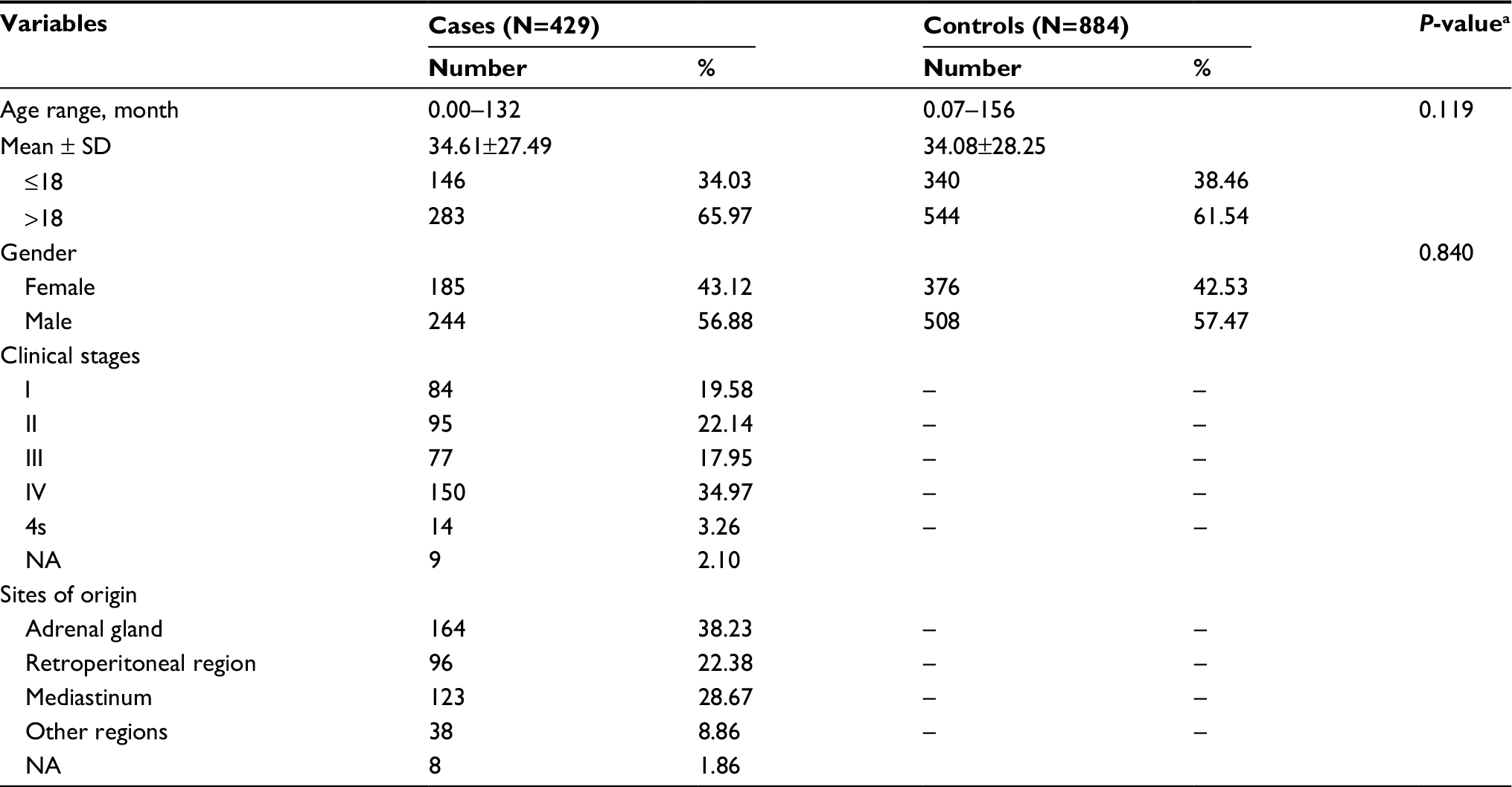 | Table S2 Frequency distribution of demographic characteristics for combined subjects Notes: aTwo-sided χ2 test for distributions between neuroblastoma patients and controls. “–” indicates no value. Abbreviation: NA, not available. |
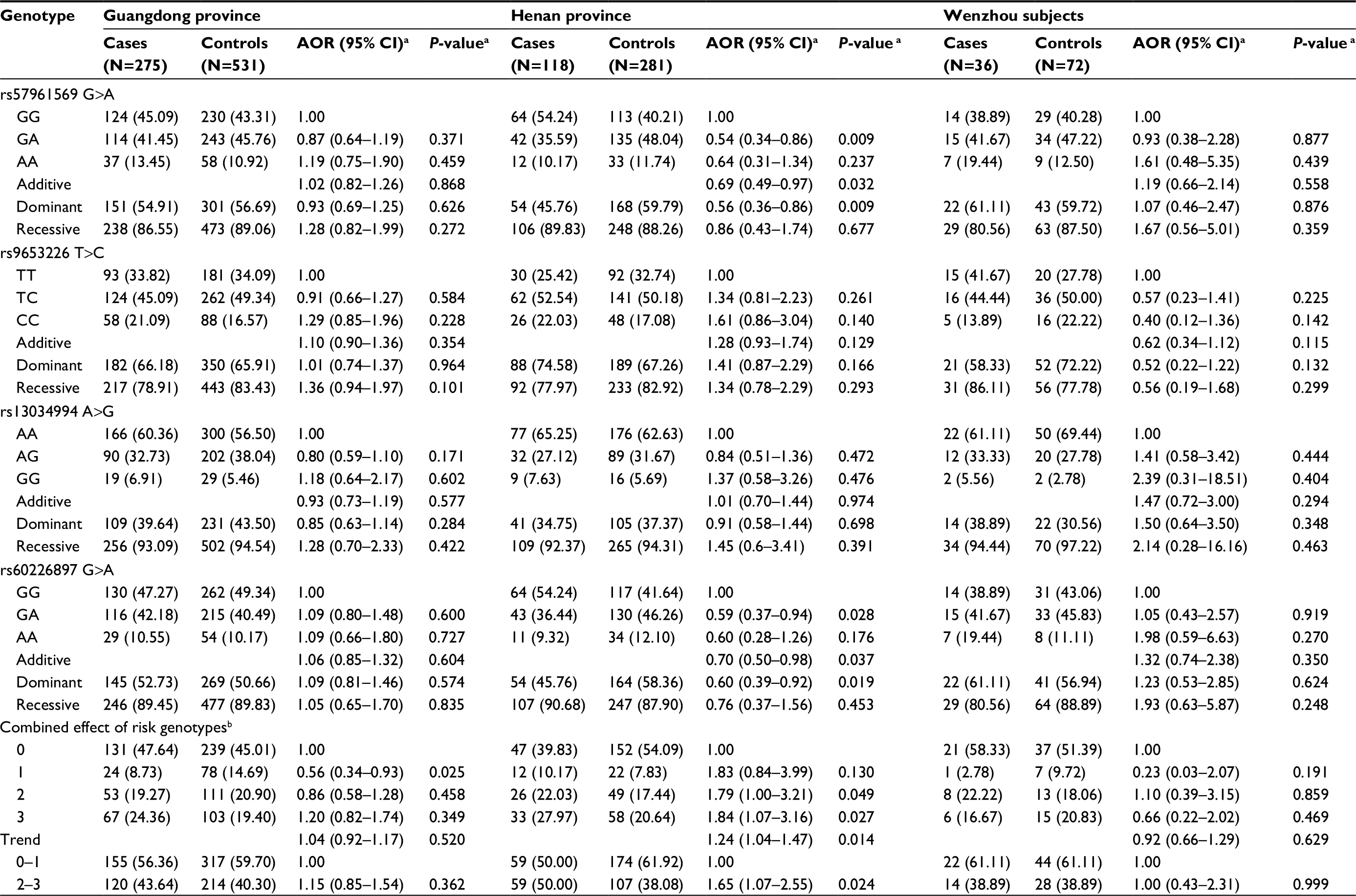 | Table S3 Genotype frequencies of MYCN gene polymorphisms and neuroblastoma susceptibility (divided subjects) Notes: aAdjusted for age and gender. bRisk genotypes were rs57961569 GG, rs9653226 CC, rs13034994 GG, and rs60226897 GG. Abbreviations: AOR, adjusted odds ratio; CI, confidence interval. |
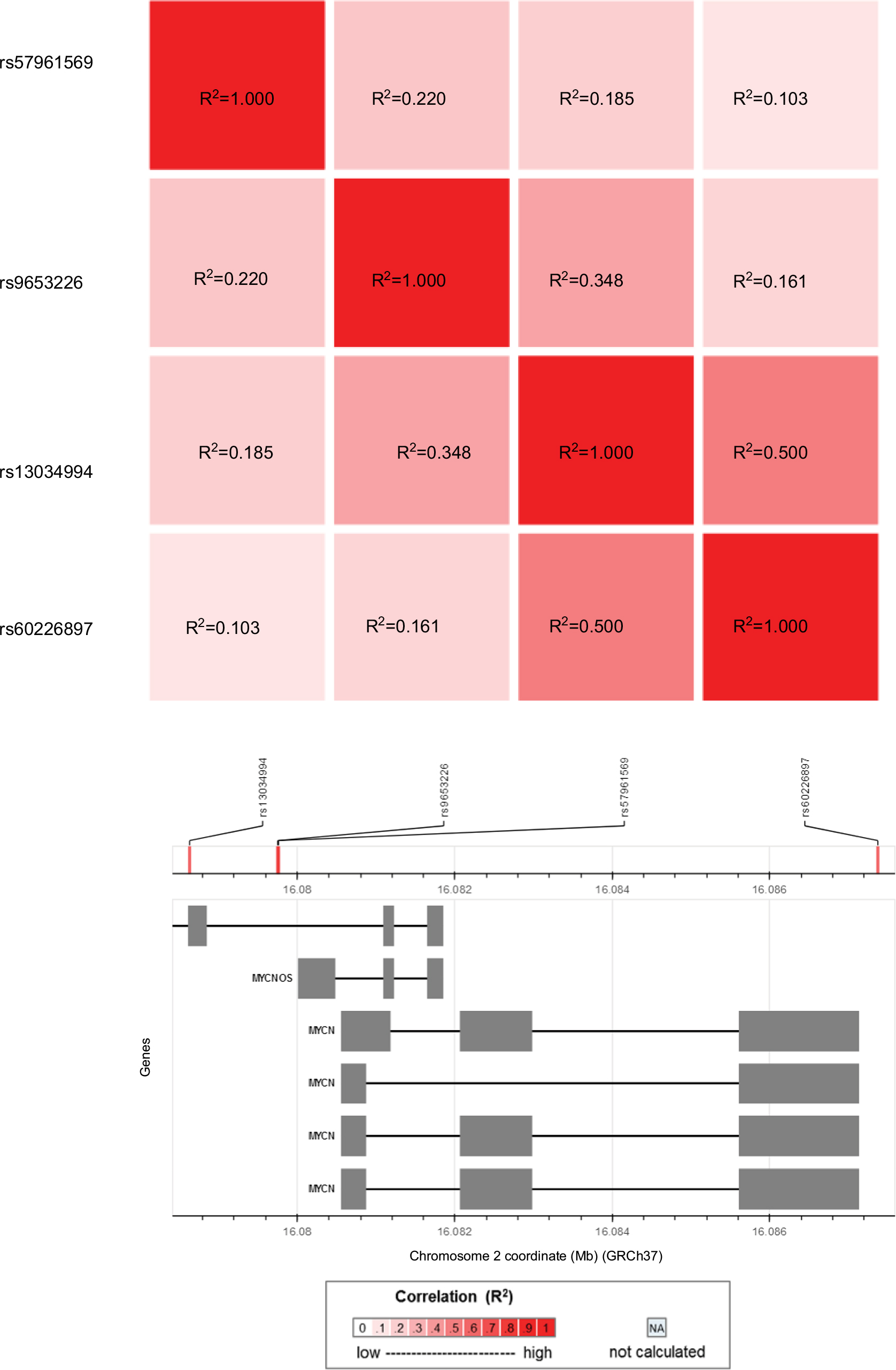 | Figure S1 Linkage disequilibrium analysis for the 4 selected polymorphisms in MYCN gene in Han Chinese population consisting of CHB (Han Chinese in Beijing, China) and CHS (Southern Han Chinese) subjects. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.