Back to Journals » The Application of Clinical Genetics » Volume 17
Polymorphisms in Immune Genes and Their Association with Tuberculosis Susceptibility: An Analysis of the African Population
Authors Wodelo W, Wampande EM ![]() , Andama A, Kateete DP, Ssekatawa K
, Andama A, Kateete DP, Ssekatawa K ![]()
Received 31 December 2023
Accepted for publication 25 March 2024
Published 29 March 2024 Volume 2024:17 Pages 33—46
DOI https://doi.org/10.2147/TACG.S457395
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Martin Maurer
Wycliff Wodelo,1 Eddie M Wampande,1,2 Alfred Andama,3 David Patrick Kateete,1 Kenneth Ssekatawa4,5
1Department of Immunology and Molecular Biology, School of Biomedical Science, College of Health Science, Makerere University, Kampala, Uganda; 2Department of Veterinary Medicine, School of Veterinary Medicine and Animal Resources, College of Veterinary Medicine, Animal Resources and Biosecurity, Makerere University, Kampala, Uganda; 3Department of Medical Microbiology, School of Medicine, College of Health Science, Makerere University, Kampala, Uganda; 4Department of Science, Technical and Vocational Education, Makerere University, Kampala, Uganda; 5Africa Center Excellence in Materials Product Development and Nanotechnology (MAPRONANO ACE), Makerere University, Kampala, Uganda
Correspondence: Eddie M Wampande, Email [email protected]
Abstract: Tuberculosis remains a global health concern, with substantial mortality rates worldwide. Genetic factors play a significant role in influencing susceptibility to tuberculosis. This review examines the current progress in studying polymorphisms within immune genes associated with tuberculosis susceptibility, focusing on African populations. The roles of various proteins, including Toll-like receptors, Dendritic Cell-Specific Intercellular Adhesion Molecule-3 Grabbing Non-Integrin, vitamin D nuclear receptor, soluble C-type lectins such as surfactant proteins A and D, C-type Lectin Domain Family 4 Member E, and mannose-binding lectin, phagocyte cytokines such as Interleukin-1, Interleukin-6, Interleukin-10, Interleukin-12, and Interleukin-18, and chemokines such as Interleukin-8, monocyte chemoattractant protein 1, Regulated upon activation, normal T-cell expressed and secreted are explored in the context of tuberculosis susceptibility. We also address the potential impact of genetic variants on protein functions, as well as how these findings align with the genetic polymorphisms not associated with tuberculosis. Functional studies in model systems provide insights into the intricate host-pathogen interactions and susceptibility mechanisms. Despite progress, gaps in knowledge remain, highlighting the need for further investigations. This review emphasizes the association of Single Nucleotide Polymorphisms with diverse aspects of tuberculosis pathogenesis, including disease detection and Mycobacterium tuberculosis infection.
Keywords: tuberculosis, polymorphisms, immune genes, African populations, genetic variants, Mycobacterium tuberculosis
Background
Tuberculosis (TB) is a pervasive bacterial infection caused by Mycobacterium tuberculosis (Mtb), contributing significantly to global mortality. Despite Mtb’s latent infection in a substantial proportion of the global population, only a fraction progresses to active TB. The disproportionate burden of TB in Sub-Saharan Africa and Southeast Asia underscores its public health significance.1,2 Mycobacterium tuberculosis’s intracellular nature and host-specific interactions highlight the genetic basis of susceptibility.
Numerous studies have explored the connection between genomic loci and TB susceptibility, encompassing candidate genes, family aggregation, twin studies, house contacts, and genome-wide analyses.3 Key players in TB susceptibility include genes encoding proteins such as major histocompatibility complex/human leukocyte antigen (MHC/HLA), tumor necrosis factors (TNFs) and their receptors, immune-related GTPases (IRGs), NRAMP1 (SLC11A1), Toll-like receptors (TLRs), vitamin D nuclear receptor (VDR), and cell surface proteins like collectins.4,5
The complex interplay between host genetics and TB susceptibility underscores the need for a nuanced understanding. While existing literature offers insights, inconsistencies arise, necessitating a deeper exploration of innate immune genes. Genetic polymorphisms associated with TB susceptibility display varying frequencies across populations, suggesting the role of environmental selection pressures.5,6
This review focuses on polymorphisms in immune genes linked to pulmonary TB susceptibility in African populations, Table 1. By elucidating the interaction between host molecules and Mtb, we aim to contribute to the design of effective strategies against tuberculosis.
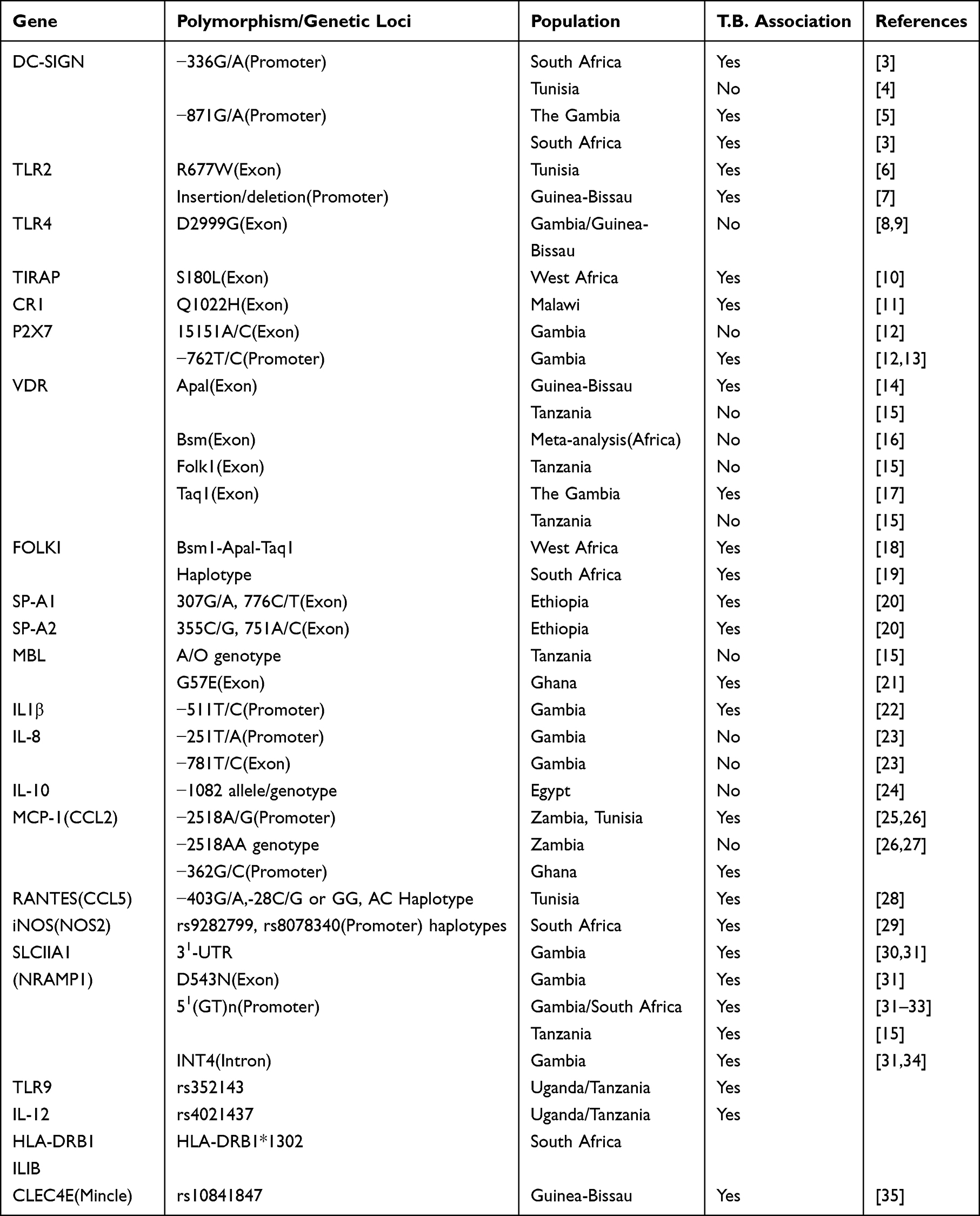 |
Table 1 Association Studies of Polymorphisms in Immune Genes and Tuberculosis |
The African Context
Understanding the genetic basis of tuberculosis (TB) susceptibility is a complex endeavor that requires careful consideration of diverse populations. Focusing on African populations in TB polymorphism studies is a strategic choice driven by various scientific, epidemiological, and ethical considerations. This justification aims to elucidate the rationale behind concentrating exclusively on African populations.
High TB Burden in Africa
Africa bears a substantial burden of TB, with high prevalence rates and a significant impact on public health.7 By concentrating on African populations, we prioritize a region where TB poses a severe threat to community well-being. Tuberculosis in Africa is characterized by a multifaceted interplay of factors, including TB-HIV co-infection, poverty, weak healthcare systems, and drug resistance. These unique challenges underscore the necessity for tailored interventions that address the specific needs of African communities.
Genetic Diversity in African Populations
Africa is characterized by unparalleled genetic diversity, owing to its rich history, diverse ethnicities, and unique population structures.8 Investigating TB polymorphisms in this context allows for a nuanced exploration of genetic variants that may be specific to African populations. This approach aligns with the growing recognition of the importance of population-specific genetic studies.
Prevalence of HIV Co-Infection
TB-HIV co-infection represents a critical aspect of TB epidemiology in Africa. HIV weakens the immune system, heightening susceptibility to TB infection and increasing the risk of TB reactivation among individuals with latent TB. Numerous studies have underscored the strong association between HIV and TB in Africa, with a significant proportion of TB cases occurring among HIV-positive individuals.1,9
Impact of Poverty and Socioeconomic Factors
Poverty and socioeconomic disparities significantly fuel the TB epidemic in Africa. Factors such as poor living conditions, overcrowding, limited healthcare access, and inadequate nutrition contribute to the transmission and persistence of TB in many African communities.10 These challenges exacerbate efforts to control TB and impede the achievement of optimal treatment outcomes.
Challenges Posed by Weak Healthcare Systems
Africa’s healthcare systems grapple with resource constraints, infrastructure deficiencies, and staffing shortages, presenting substantial barriers to effective TB control. Inadequate diagnostic capacities, limited access to quality medications, and suboptimal adherence to treatment regimens further complicate TB management in numerous African countries.11 Strengthening healthcare systems and enhancing access to TB care are paramount for reducing the burden of TB in Africa.
Emergence of Drug Resistance
The emergence of drug-resistant TB strains, including multidrug-resistant (MDR) and extensively drug-resistant (XDR) variants, poses a formidable challenge to TB control efforts across Africa. Limited access to quality diagnostic tools, inappropriate treatment regimens, and poor treatment adherence contribute to the proliferation of drug-resistant TB strains in the region.12 Addressing the menace of drug-resistant TB demands enhanced surveillance, rigorous infection control measures, and expanded access to effective treatment modalities.
Environmental and Socioeconomic Factors
The interplay between genetics and environmental factors is a crucial aspect of TB susceptibility. African populations often face unique environmental challenges and socioeconomic conditions that influence disease dynamics. Investigating TB polymorphisms in this context allows for a holistic understanding of the gene-environment interactions contributing to TB susceptibility.
Precision Medicine and Targeted Interventions
African populations deserve tailored medical interventions that consider their unique genetic makeup. Precision medicine initiatives aim to develop therapies specific to individual genetic profiles.13 By focusing on TB polymorphisms in African populations, we contribute to the development of targeted interventions that address the specific needs of this population. Therefore, the decision to study TB polymorphisms in African populations is rooted in the region’s high TB burden, genetic diversity, unique environmental challenges, and the pursuit of precision medicine. By concentrating our efforts in this context, we aim to unravel population-specific genetic factors influencing TB susceptibility, ultimately contributing to more effective and targeted strategies for TB control in Africa.
Epidemiological Data
Tuberculosis remains a pressing public health concern, particularly in African populations, where it continues to exert a significant burden on health systems and communities. The incidence rates, mortality rates, and demographic characteristics associated with TB in Africa are shown in Table 2.14
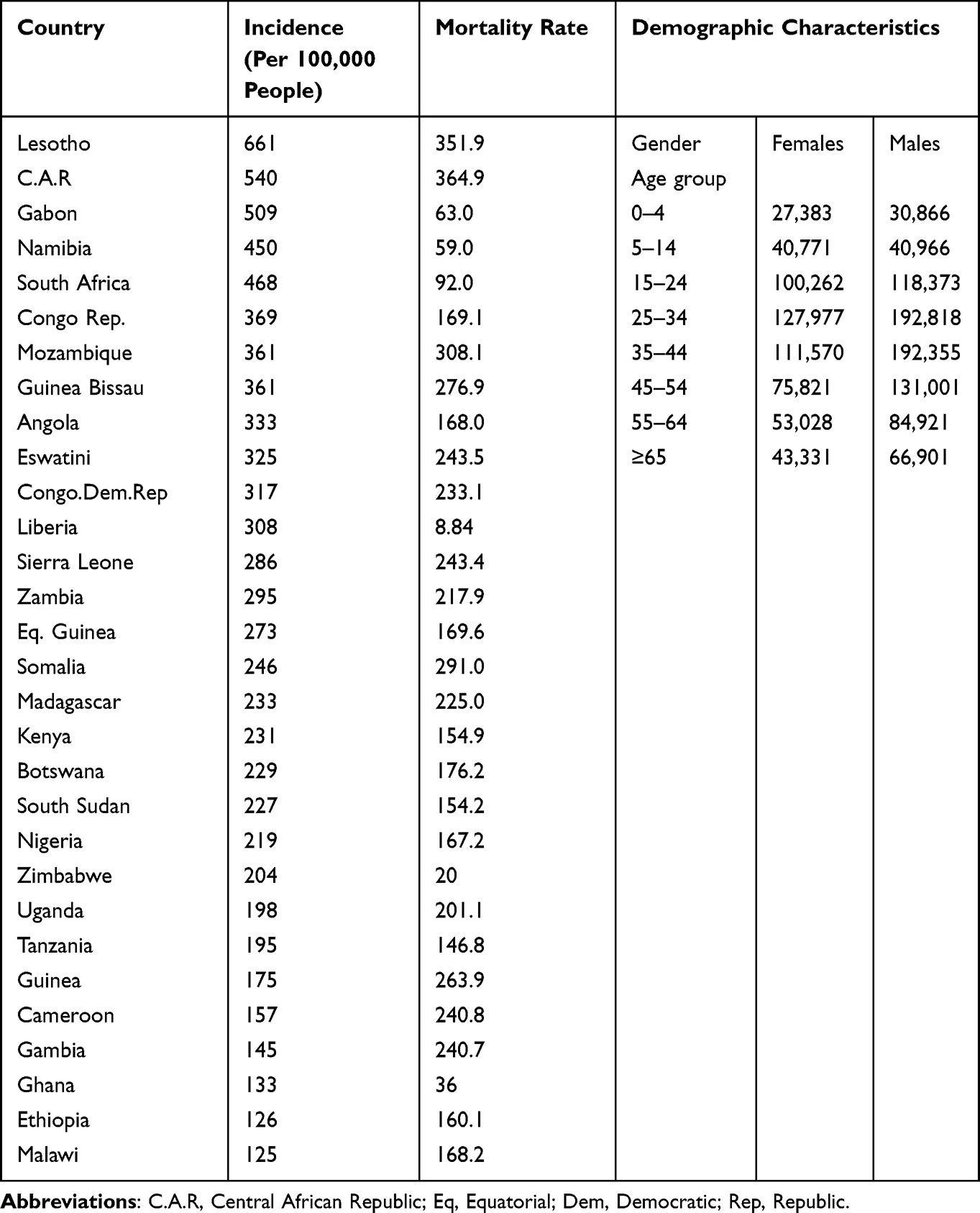 |
Table 2 The Incidence, Mortality Rates, and Demographic Characteristics Associated with TB in Africa |
TB Incidence Rates
Countries such as Lesotho, C.A.R, Gabon, South Africa have the highest Tb incident rates on the continent. However, progress has been made in reducing TB incidence in several African countries through increased access to diagnostic and treatment services, Table 2.
TB Mortality Rates
Somalia, Mozambique, Lesotho and C.A.R are among the countries with the highest TB mortality rates in Africa. TB mortality rates in Africa are closely linked to factors such as HIV co-infection, drug resistance, and healthcare access. While mortality rates have declined in recent years due to improved TB control efforts, they remain significant, particularly in countries with high HIV prevalence.
Demographic Features
Young adults, particularly those in the economically productive age group, are disproportionately affected by TB. Additionally, men are more likely to develop TB than women in many African countries, due to social and behavioral factors as seen in Table 2. Socioeconomic disparities, including poverty, overcrowding, and limited access to healthcare, contribute to the unequal distribution of TB within communities. Urban areas tend to have higher TB burdens due to population density and environmental factors, although rural areas are not exempt from the disease.
Roles of Proteins in Host – Pathogen Interaction
In the realm of tuberculosis susceptibility, extensive research is focused on elucidating the intricate roles of various proteins to decipher the underlying mechanisms governing host-pathogen interaction. These proteins, encompassing Toll-like receptors (TLRs), Dendritic Cell-Specific Intercellular Adhesion Molecule-3 Grabbing Non-Integrin (DC-SIGN), the vitamin D nuclear receptor, soluble C-type lectins such as surfactant proteins A and D, C-type Lectin Domain Family 4 Member E (CLEC4E), and mannose-binding lectin (MBL), represent integral components of the immune system’s response to Mycobacterium tuberculosis (MTB) infection.
As we embark on an in-depth exploration of the nuanced interplay between these proteins and MTB, we intent to uncover their individual and collective contributions to TB susceptibility. Farthermore, discerning the roles of phagocyte cytokines, including Interleukin-1 (IL-1), Interleukin-6 (IL-6), Interleukin-10 (IL-10), Interleukin-12 (IL-12), and Interleukin-18 (IL-18), alongside chemokines such as Interleukin-8 (IL-8), monocyte chemoattractant protein 1 (MCP-1), and Regulated upon activation, normal T-cell expressed and secreted (RANTES). This comprehensive investigation serves to enhance our understanding of the immune response to TB infection, paving the way for novel insights into TB susceptibility mechanisms and potential avenues for therapeutic intervention.
The Human Leukocyte Antigens(HLAs)
The Human Leukocyte Antigen (HLA) molecules are proteins found on the surface of cells in the human body. They play a crucial role in the immune system by presenting antigens, which are fragments of proteins from pathogens like bacteria and viruses, to T cells. This process is essential for the immune system to recognize and respond to foreign invaders while distinguishing them from the body’s own cells. HLA molecules are highly diverse and polymorphic, meaning they exist in many different forms within human populations. This diversity is important for immune system function and contributes to individual differences in immune responses to pathogens, susceptibility to diseases, and compatibility for organ and tissue transplantation.
Recent studies have revealed that specific Human Leukocyte Antigen (HLA) alleles might contribute to either increased or decreased susceptibility to tuberculosis (TB) within African communities.5 For instance, investigations have established correlations between certain HLA class I alleles—such as HLA-B39:01 and HLA-B58:01—and elevated TB susceptibility among West African populations. Conversely, alternative HLA class I alleles, like HLA-B*57:01, have demonstrated a protective association against TB within African cohorts.15,16
Furthermore, HLA class II alleles have emerged as significant factors in TB susceptibility among African populations.16 Notably, research has identified links between particular HLA-DRB1 alleles—such as HLA-DRB115:01 and HLA-DRB104:05—and heightened susceptibility to TB among East and West African groups. Conversely, contrasting HLA-DRB1 alleles, including HLA-DRB110:01 and HLA-DRB111:01, have shown protective effects against TB within African populations.
It is crucial to recognize the intricate nature of the relationship between HLAs and TB susceptibility, influenced by diverse factors such as genetic background, environmental exposures, and host-pathogen interactions. Moreover, the prevalence of specific HLA alleles may exhibit variability across different African populations, underscoring the necessity for population-specific investigations to comprehensively grasp the genetic determinants of TB susceptibility.
Soluble C-Type Lectins (Collectins)
Collectins, including MBL, SP-A, and SP-D, are calcium-dependent lectins crucial for innate immunity. Previous data on calcium-dependent (C type) lectins reveals the close relationship between the bacillus with MBL and SP-A, SP-D17 Specifically, SP-A and SP-D, located in the alveoli and respiratory epithelium, play a pivotal role in the defense against intracellular pathogens, including Mtb. SP-A and SP-D, collagen-like glycoproteins, serve as opsonins, enhancing phagocytosis by alveolar macrophages.17–19 Additionally, SP-A interacts with alveolar macrophages, modulating TLR2 and TLR4 pathways,18 showcasing the intricate immune pathways influenced by these collectins. SP-A’s impact on inflammatory cells depends on the cell state, resulting in the inflammatory paradox. While enhancing cytokine release during infection, SP-A also suppresses macrophage secretion, revealing a complex regulatory role in the immune response.
Polymorphic changes in the genes encoding SP-A and SP-D, particularly in the amino acid sequences, contribute to susceptibility to TB.20 Genetic changes in SP-A and SP-D amino acid sequences have functional consequences. Single nucleotide polymorphisms in these proteins occur when a single nucleotide (adenine, thymine, cytosine, or guanine) in a DNA sequence is altered or substituted with a different single nucleotide at a specific position in the genome. These genetic variants may either result in a synonymous or non-synonymous substitution. Since interactions of SP-A and SP-D with the same and or different pathogens occur. A polymorphism in SP-A reduces binding to Mtb, increasing susceptibility, while a genetic change in SP-D alters the sequence, affecting bacterial surface binding and increasing uptake by alveolar macrophages.
Membrane Binding Lectins (MBL)
Membrane-binding lectin, a soluble pathogen recognition receptor, plays a pivotal role in host defense against bacterial pathogens. Its hexameric structure, akin to the C1q component of the complement system, facilitates the opsonization of microbes, enhancing phagocytosis. MBL also contributes to inflammation by clearing apoptotic cells and releasing cytokines.21,22 MBL gene polymorphisms contribute to variations in blood levels and hexamer formation. MBL gene duplication results in the identification of two genes, MBL1 and MBL2, with MBL2 being the representative in humans. The MBL2 gene encompasses four coding regions (exons) separated by three non-coding regions (introns) at codon positions 52, 54, and 57, featuring variant alleles, D, B, and C, respectively. Certain alleles are linked to impaired hexamer formation and reduced blood levels.5
Mutations at specific codons resulting in low MBL levels increase susceptibility to various infections, particularly in individuals with other immune defects. Despite numerous studies exploring MBL alleles and their polymorphisms (Table 1), inconsistent results underscore the imperative to comprehensively understand the relationship between TB establishment and the polymorphic variants in the MBL gene.
Mannose-binding lectin-2 variants exhibit relatively frequent occurrences, albeit unevenly distributed across human populations.23 Notably, allele B is prevalent in Asian and native South African populations but absent in the West African population. Conversely, allele D is confined to North Africa and Caucasian populations, while allele C is predominant in Sub-Saharan Africa. These allele frequency disparities shed light on the biological advantage of MBL low levels in specific populations.
In summary, individuals with low MBL levels face increased susceptibility to various infections, particularly when coupled with other immune defects. Despite numerous studies exploring MBL alleles and their polymorphisms (Table 1), inconsistent results underscore the imperative to comprehensively understand the relationship between TB establishment and the polymorphic variants in the MBL gene. Notably, the allele frequency disparities observed, especially the prevalence of allele B in certain populations and the restriction of allele D to specific regions, shed light on the biological advantages associated with MBL low levels in distinct populations. Further investigations are warranted to unravel the precise mechanisms linking MBL polymorphisms and TB establishment, paving the way for targeted interventions and a deeper understanding of host-pathogen interactions.
CLEC4E (C-Type Lectin Domain Family 4 Member E)
CLEC4E is a calcium-dependent lectin characterized by carbohydrate recognition domains (CRDs), facilitating its involvement in pathogen recognition and cellular interactions.24 Specifically, CLEC4E encodes the receptor Mincle, situated on the outer membrane of macrophages. Mincle is responsible for recognizing trehalose-6, 6-dimycolate (TDM), a component in the mycobacterial cell wall.25 This recognition triggers signaling cascades within the innate immune system, initiating a robust response to mycobacterial infection.
A single nucleotide polymorphism (SNP) at rs10841847 within CLEC4E has emerged as a crucial determinant in TB susceptibility. The minor allele (G˃A) at this locus is associated with an increased vulnerability to pulmonary TB. This emphasizes the significance of CLEC4E in the context of TB pathogenesis.25 However, the complex landscape of TB susceptibility is likely influenced by multiple genetic factors, necessitating further investigations. While the association between CLEC4E and pulmonary TB is evident, published studies present inconsistent findings. Notably, research in admixed South African populations did not yield statistically significant results, while a study in Northern China reported a decreased predisposition to active TB associated with CLEC4E minor alleles.24,26,27 These discrepancies underscore the need for more comprehensive research to establish the causal link between CLEC4E and TB susceptibility, considering potential confounding factors.
Studies involving mice lacking the Mincle gene offer insights into its role in the immune response against Mycobacterium tuberculosis (Mtb). Depletion of Mincle resulted in the inability to recruit and adhere neutrophils to the site of invasion, impacting the formation of lung granulomas.28 Altered DNA sequences, such as those associated with the rs10841847 SNP, disrupt the normal functioning of Mincle receptors, potentially compromising the ability to bind tubercle glycolipid and increasing susceptibility to TB. Interestingly, studies with knockout mice showed a paradoxical outcome, with a higher bacterial load compared to wild-type mice, suggesting a Mincle-mediated immune response against Mtb.29 This paradox may be attributed to additional genetic factors influencing the sustained immune response, indicating the complexity of host-pathogen interactions in secondary TB.
Pathogen Recognition Receptors (PRRs)
Pathogen recognition receptors are membrane and transmembrane receptors located on the dendritic cells, macrophages, and epithelial cells (exogenous sensors) that recognize different types of Pathogen Associated Molecular Patterns (PAMPs). Macrophage PRRs and phagocytic receptors causative to mycobacterial diseases, especially T.B., have been discussed in several recent reviews.17,24,25 These include cell membrane-bound receptors, such as the mannose receptor (M.R., CD206), dendritic cell-specific ICAM-3-grabbing non-integrin (DC-SIGN) (CD209), Dectin-1, and TLRs. However, in this review, we shall focus on the DC-SIGN and TLRs, which have been associated with Tuberculosis in the African population. This is because they have been adequately studied and have been associated with tuberculosis disease susceptibility and are expressed on many immune cells that recognize numerous molecules of different pathogens including the Mycobacterium tuberculosis.
The exploration of Pathogen Recognition Receptors (PRRs) in dendritic cells, macrophages, and epithelial cells is crucial for deciphering how these receptors recognize diverse Pathogen Associated Molecular Patterns (PAMPs). The emphasis on cell membrane-bound receptors, such as mannose receptor (M.R., CD206), dendritic cell-specific ICAM-3-grabbing non-integrin (DC-SIGN), and Toll-Like Receptors (TLRs), provides a foundation for understanding the initial steps of host-pathogen recognition.30–32
The selective focus on DC-SIGN and TLRs, particularly TLR2, TLR4, and TLR9, is well-justified, considering their extensive associations with TB susceptibility in the African population.31,33,34 These receptors, expressed on various immune cells, serve as critical mediators in the immune response against Mycobacterium tuberculosis. Their well-studied roles in recognizing M. tuberculosis components make them pivotal players in the host defense against TB.35,36
The investigation of Toll-Like Receptors (TLRs) further deepens our understanding of the genetic variants associated with TB susceptibility. The elucidation of TLR1, TLR2, TLR4, and TLR9 variations across different populations provides a nuanced perspective on the global landscape of TB genetics.31 Polymorphisms in these TLRs, including TLR1 G1805T, TLR2 T597C, T1350C, TLR6 C745T, and TLR4 Asp 299Gly, Thr399Ile, reveal potential risk factors for TB susceptibility.31,33,34,37
The signaling pathways initiated by TLR2 and TLR4, leading to the activation of NF-κB and other cascades, are critical in understanding TB susceptibility.35 The identification of specific genetic changes in TLR2, TLR4, and TLR9 associated with TB susceptibility, such as the R753Q variant in TLR2, adds a layer of complexity to our understanding.38 The review delves into the influence of these variants on signaling pathways and their impact on TB progression.
Toll-Like Receptors (TLRs)
Toll-like receptors expressed on various immune cell types, serve as critical mediators in the immune response against a spectrum of pathogens, including Mtb.31,39 Recognizing a diverse array of microbial molecules, TLRs initiate transcription and proinflammatory gene processes. TLRs are categorized based on their cellular localization—surface-bound (eg, TLR2 and TLR4) and intracellular (eg, TLR8 and TLR9).35 The mycobacterial cell wall components are recognized by multiple TLRs, emphasizing their crucial role in the immune response to MTB.34,40
Genetic variants within TLRs have been identified globally, with polymorphisms in TLR1, TLR2, TLR4, TLR8, and TLR9 associated with TB susceptibility.26,31,40 Notably, TLR2 stands out with higher prevalence in genetic variants, forming a heterodimer with TLR1 or TLR6.36 The TLR2 signaling pathway, particularly involving the TIR domain-containing adaptor protein (TIRAP), plays a pivotal role in the inflammatory response to MTB.35,36
In the context of the African population, TLR-2, TLR-4, and TLR-9 emerge as key players in TB susceptibility. The R753Q variant in TLR2 influences disease progression in children.38 Contrasting reports on TLR4 variant D299G highlight its association with TB susceptibility in HIV-infected patients in Tanzania.37,41 The intricate interplay between HIV and TLR variants underscores the multifactorial nature of susceptibility in co-infected individuals. The signaling pathways initiated by TLR2 and TLR4, culminating in the activation of NF-κB, are central to TB susceptibility. Noteworthy changes in these domains correlate with variations in TB susceptibility. The nonsynonymous SNP S180L in TIRAP confers protective signal transduction within TLR2.10 Additionally, TLR7 and TLR8 genetic polymorphisms are associated with TB susceptibility, influencing phagocytosis and immune activation.42 Evidence from TLR9-deficient mice further supports the critical role of TLRs in the innate defense against MTB.26 These results provide proof that TLRs and their signaling moieties in their pathways possess susceptible genes that are associated with Tuberculosis. However, due to variations within individual genes in the different racial populations, it is very hard to make a close linkage of the genetic factors that bring about the association.
Dendritic Cell-Specific Intercellular Adhesion Molecule-3 Grabbing Non-Integrin (DC-SIGN)
DC-SIGN is characterized by a carbohydrate recognition domain (CRD) at its extracellular COOH-terminal end. Its expression on macrophages is influenced by tissue type and activation state.32,39 Notably, DC-SIGN is induced in alveolar macrophages from M. tuberculosis-infected patients, marking its relevance in the context of TB infection.35 The binding of M. tuberculosis to DC-SIGN initiates phagocytosis, allowing the pathogen to enter human monocyte-derived D.C.s. This interaction directly interferes with dendritic cell maturation, contributing to the intricate host-pathogen dynamics.33 The cytoplasmic tail of DC-SIGN, housing three conserved motifs, plays a crucial role in ligand binding, receptor signaling, phagocytosis, and intracellular trafficking of ligand particles.33 This calcium-dependent lectin, known for its affinity for mannose-rich molecules, is expressed predominantly on dendritic cells (D.C.s) and, to varying degrees, on macrophages.34 The dynamic interplay between TB and DC-SIGN involves ligand binding, phagocytosis, and modulation of dendritic cell maturation.32,39
Genetic variations in DC-SIGN have been implicated in TB susceptibility, with polymorphisms in the promoter region (eg, −871A/G and −336A/G) associated with distinct susceptibility patterns (Table 1). Notably, studies in the untranslated regions (UTRs) of CD209 and CD207R genes have revealed variations in gene frequency within African indigenous individuals and the Caucasian American population, underscoring the importance of ethnic, racial, and geographical diversity in TB susceptibility assessment.36 A genetic comparison between African and Eurasian populations has revealed notable disparities, particularly in the frequency of the polymorphic allele −871G. This allele is significantly more prevalent in Eurasians than in Africans, suggesting a genetic migration and close association leading to its introduction into the South African population.43 The absence of haplotypes −871G and −336A in most African populations, except for colored South Africans, further emphasizes the impact of genetic migration on TB susceptibility. In conclusion, such genetic variations tend to influence translation and gene polymorphisms.
Solute Carrier Family 11A, Member 1, SLC11A1 or Natural Resistance-Associated Macrophage Protein 1, or NRAMP1)
The SLC11A1 (NRAMP1) gene encodes a membrane transporter protein whose function is to regulate the cytoplasmic cation levels, especially iron. Iron is essential for the cell to generate reactive oxygen and nitrogen intermediates. It, therefore, acts as a divalent cation transporter across phagosomal membranes.38 The origin of the gene from a mouse is responsible for its resistance to infections by intracellular pathogens, especially the mycobacteria.37
SLC11A1’s function as a divalent cation transporter across phagosomal membranes is critical in modulating the concentration of metal ions within the phagosome. This regulation is paramount for macrophage defense against bacterial infections, specifically by reducing the metal ion concentration where bacterial pathogens reside. A defective NRAMP gene, however, results in increased metal ion accumulation, supporting bacterial growth and compromising the host’s ability to resist infection.
Studies across diverse populations and recent meta-analyses have highlighted the influence of SLC11A1 polymorphisms on TB susceptibility. Four key polymorphisms (3=-UTR, D543N, 5= (G.T.) and INT4 have been extensively studied globally. Although findings exhibit some contradictions in various populations, a meta-analysis in China and an association in Turkey underscore the significance of these polymorphisms in TB development.
A systematic review conducted in China further demonstrates the role of SLC11A1 polymorphisms, particularly at the D543N and INT4 loci, in response to infection rather than solely to Mycobacterium tuberculosis.42,44,45 Additionally, studies in the Gambia suggest potential links between SLC11A1 gene polymorphisms, abnormal iron levels, gene-nutrient interactions, and mortality prediction in HIV-infected patients. This raises intriguing questions about the potential risk of TB development in HIV patients with clinically abnormal iron levels and gene-nutrient interactions. Genetic interactions, such as those observed between TLR2 and polymorphism rs3731865 in African Americans, emphasize the significance of gene-gene interactions in host-pathogen genetic studies. These findings highlight the complexity of allelic heterogeneity within SLC11A1 and underscore the importance of considering various factors in understanding TB susceptibility.
Highlighted associations with SNPs in SLC11A1 are attributed to adjacent SNPs in linkage disequilibrium with reported variants. However, the 5- (G.T.) repeat in the promoter region of SLC11A1 has been shown to affect promoter activity, with allele 3 displaying greater basal and lipopolysaccharides (LPS)-stimulated activity. This intriguing gene activity, although insufficient for establishing clear evidence for in vivo activity, links high activity with resistance and low activity with susceptibility to tuberculosis.46
Chemokines (Chemotactic Cytokines)
Chemokines are a large family of structurally homologous cytokines, secreted proteins that signal through cell surface G protein-coupled heptahelical chemokine receptors that stimulate leukocyte movement and regulate the migration of leukocytes from the blood to tissues. Therefore, chemokines are proinflammatory cytokines of low molecular size, which coordinate the migration and activation of different leukocyte populations. The structurally homologous chemokines are categorized into four groups based on conserved cysteine arrangement: CXC, CC, C, and CX3C. These low molecular-size proinflammatory cytokines play a pivotal role in coordinating the migration and activation of various leukocyte populations.1
Upon activation of alveolar macrophages by intracellular M. tuberculosis, a cascade of chemokines is released. Notable among these are IL-8 (CXCL8), monocyte chemoattractant protein 1 (MCP-1, CCL2), RANTES (CCL5), and CXCL10 (IP-10). These chemokines act as molecular messengers, orchestrating the migration and activation of T cells and macrophages to sites of Mtb infection. This orchestrated response forms the characteristic tuberculous granuloma, a fortress against the dissemination of the organism.2 Systematic reviews and research articles have extensively documented the significance and genetic variations within various chemokines combating Mycobacterium tuberculosis.25,47,48 Polymorphic variations, particularly in the promoter regions of these genes, can modulate transcription and translation, influencing the overall production and activity of chemokines.3,4
The production of chemokines is indispensable for recruiting inflammatory cells to the infection site. Early recruitment of macrophages is pivotal for infection control.5 M. tuberculosis, a potent inducer of chemokines, triggers protective and immunopathogenic host responses. Studies on human macrophages infected in vitro have revealed the induction of CCL2, CCL3, CCL4, and CCL5 in response to virulent mycobacterial strains.49 Regulated upon activation, normal T-cell expressed, and secreted (RANTES) plays a critical role in T-cell activation and macrophage activation.50 Clinical studies have detected the presence of MCP-1, MIP-1a, RANTES, and IP-10 in the serum and bronchoalveolar lavage of TB patients.51 Alveolar macrophages, in particular, produce a distinct profile of chemokines, suggesting a role in the influx of cells to the infection site, and influencing granuloma formation.52 The expression of chemokines by macrophages not only influences TNF-α production but also affects chemokine receptor expression. Specifically, CCL2, CCL3, CCL4, CCL5, CXCL10, and CXCL13 expression is modulated by macrophages in response to Mtb infection.53 Furthermore, TNF-α itself can influence the chemokine network expression, indirectly impacting granuloma formation.54 This intricate interplay extends to mycobacterial cell wall components, regulating the induction of chemokine secretion by macrophages.55
Rantes (Ccl5)
RANTES (regulated upon activation, normal T-cell expressed, and secreted), a member of the C-C chemokine subfamily or beta chemokine. RANTES exhibits chemotactic properties for monocytes, memory T cells, and eosinophils. In addition, it triggers histamine release from human basophils.56,57 Notably, in a murine model, RANTES has been identified as a key player in promoting granuloma formation within M. tuberculosis-infected lungs.56 This process involves the recruitment of immune cells to the infected area, subsequently limiting the intracellular growth of mycobacteria and inducing direct antimicrobial activity, notably through the release of nitric oxide (NO) in alveolar macrophages.58 The protective immunity conferred by RANTES, as evidenced in knockout mice, underscores its significance in the defense against TB.20
Exploring the genetic landscape, the promoter region of the RANTES gene reveals two variants, −403G/A and −28C/G, associated with Tuberculosis (Table 1). These single nucleotide polymorphisms may confer either resistance or susceptibility to TB.59 A recent discovery highlights a functionally important polymorphism, In1.1T/C, located in the intron of RANTES. This polymorphism regulates gene transcription by differentially binding to alternative forms of nuclear proteins. The complexity of RANTES transcription regulation is further unraveled through haplotype analysis. Haplotypes II (A-C-T) and V (G-C-C) exhibit opposing effects on RANTES transcription, with II up-regulating and V down-regulating the process. The luciferase assay, correlating RANTES SNPs with transcription activity, supports this functional insight. Consequently, RANTES concentrations in bronchoalveolar lavage fluid of pulmonary TB patients demonstrate acute elevation during infection and subsequent decline during convalescence. This underscores the dynamic role of RANTES up-regulation in the immune response against M. tuberculosis infection.
Mcp-1 (Ccl2)
The monocyte chemoattractant protein-1, or MCP-1, is a β chemokine and a potent chemotactic factor for monocytes and macrophages. During lung infection by M. tuberculosis, the human monocytes within the body are stimulated to release MCP-1.60 During the host M. tuberculosis-specific immune response, MCP-1 participates in the inhibition of the in vivo dissemination of M. Tuberculosis regulating the secondary immune response. Therefore, MCP-1 is a vital promoter and regulator of a host’s defense against Tuberculosis infection. However, experimental studies in mice have shown that a deficiency in the chemokine MCP-1 alters the monocyte recruitment and formation of the granuloma, and yet it has the advantage of decreased M. Tuberculosis susceptibility.41 Studies on genotype and allele frequencies especially at the locus −2518 have shown contradictory results and have not yielded concrete and adequate proof for the effects of the polymorphisms in the MCP-1 gene. However, the carriers of the −2518 GG genotype were reported to produce high levels of MCP-1, which inhibits IL-12 production in response to M. Tuberculosis and promotes active pulmonary T.B.,61 whereas Thye et al62 initially found the association with the −2518G allele with T.B. resistance in Ghana, and further genotyping analysis using Microarray technology identified −362C allele as the only true protective variant of MCP-1, with a linkage disequilibrium in the region of −2518 and-362. This study is a deterrent note for the need to carry out a comprehensive genetic investigation to search for the actual cause of the polymorphism that exists in the variant and gene of interest.
Conclusion
In summary, the exploration of genetic factors influencing tuberculosis susceptibility in African populations represents a significant step forward in our understanding of the complex interplay between host genetics and infectious diseases. The comprehensive analysis presented in this article delves into the polymorphisms within immune genes and their association with TB susceptibility specifically within the African context. This study contributes valuable insights into the genetic landscape of TB susceptibility in African populations. By focusing on this region, the research sheds light on population-specific genetic variants that may play a pivotal role in determining an individual’s susceptibility to TB.
One key research gap pertains to the lack of comprehensive studies that consider the diverse genetic, environmental, and socio-economic factors influencing TB outcomes across different African populations. Existing research often fails to adequately account for the genetic diversity present within African populations, leading to inconsistent findings and limited generalizability of results. Moreover, there is a scarcity of longitudinal studies that can elucidate the temporal dynamics of TB susceptibility and progression in African populations, hindering our understanding of the disease’s trajectory and the efficacy of interventions over time.
Additionally, the reliance on small sample sizes and single-center studies limits the statistical power and reliability of findings, further exacerbating the inconsistency of outcomes observed in earlier research efforts. Furthermore, there is a scarcity of studies that integrate genetic, environmental, and socio-economic data to comprehensively assess the determinants of TB susceptibility in African populations, impeding our ability to develop targeted interventions tailored to the specific needs of these populations.
To address these research gaps and enhance our understanding of TB susceptibility in African populations, we propose several suggestions for further similar research. Firstly, there is a need for larger, multicenter studies that encompass diverse African populations to ensure the generalizability of findings and account for population-specific factors. Such studies should employ longitudinal designs to capture the temporal dynamics of TB susceptibility and progression, allowing for the identification of predictive biomarkers and the evaluation of interventions over time.
Moreover, future research efforts should prioritize the integration of genetic, environmental, and socio-economic data through interdisciplinary collaborations between geneticists, epidemiologists, clinicians, and social scientists. This holistic approach will provide a more nuanced understanding of the complex interactions underlying TB susceptibility in African populations and inform the development of targeted interventions that address the multifactorial nature of the disease.
Furthermore, leveraging emerging technologies such as genomic sequencing, big data analytics, and machine learning algorithms can facilitate the identification of novel genetic determinants and biomarkers associated with TB susceptibility in African populations. By harnessing these innovative approaches, researchers can overcome the limitations of earlier studies and advance our understanding of TB epidemiology in Africa, ultimately contributing to the development of effective strategies for TB prevention, diagnosis, and treatment tailored to the specific needs of African populations.
Abbreviations
MHC, Major histocompatibility complex; SLC11A1, Solute carrier family 11a member 1; HLA, Human leukocyte antigen; VDR, Vitamin D nuclear receptor; TNF, Tumour necrosis factor; IRGM, Immunity-related GTPase family M protein; SNP, Single Nucleotide Polymorphism; MTBC, Mycobacterium tuberculosis complex; MCP-1, Monocyte chemoattractant protein 1; MBL2, Mannose-binding lectin 2 gene; Mtb, Mycobacterium tuberculosis; NRAMP1, Natural resistance Associated Macrophage Protein 1 gene; SP-A, Surfactant protein A; SP-D, Surfactant protein D; CRD, Carbohydrate recognition domain; RANTES, Regulated upon activation, normal T-cell expressed, and secreted; TB, Tuberculosis; TLR, Toll-like receptor; DC-SIGN, Dendritic Cell-Specific Intercellular adhesion molecule-3 grabbing non-integrin; WHO, World Health Organization; PRR, Pathogen Recognition Receptor; PAMP, Pathogen Associated Molecular Pattern; CLEC4E, C-type Lectin Domain Family 4 Member E; IL, Interleukin; NO, Nitric Oxide.
Data Sharing Statement
All data generated by this manuscript has been integrated into the manuscript.
Acknowledgments
We are thankful to the Makerere University Library for the provision of electronic information resources that enabled us to complete the review article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors declare that this research project was funded by the Africa Centre of Excellence in Materials, Product Development & Nanotechnology; MAPRONANO ACE, Grant No. P151847IDA credit 5797-UG, College of Engineering Design Art and Technology, Makerere University.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Harding E. WHO global progress report on tuberculosis elimination. Lancet Respir Med. 2020;8(1):19. doi:10.1016/S2213-2600(19)30418-7
2. Dye C, Reeder JC, Terry RF. Research for universal health coverage. Sci Transl Med. 2013;5(199):199ed13. doi:10.1126/scitranslmed.3006971
3. Barreiro LB, Neyrolles O, Babb CL, et al. Promoter variation in the DC-SIGN-encoding gene CD209 is associated with tuberculosis. PLoS Med. 2006;3(2):e20. doi:10.1371/journal.pmed.0030020
4. Ben-Ali M, Barreiro LB, Chabbou A, et al. Promoter and neck region length variation of DC-SIGN is not associated with susceptibility to tuberculosis in Tunisian patients. Hum Immunol. 2007;68(11):908–912. doi:10.1016/j.humimm.2007.09.003
5. Yim JJ, Selvaraj P. Genetic susceptibility in tuberculosis. Respirology. 2010;15(2):241–256. doi:10.1111/j.1440-1843.2009.01690.x
6. Pepperell CS, Casto AM, Kitchen A, et al. The role of selection in shaping diversity of natural M. tuberculosis populations. PLoS Pathog. 2013;9(8):e1003543. doi:10.1371/journal.ppat.1003543
7. World Health Organization, Global Tuberculosis Report; 2020.
8. Campbell MC, Tishkoff SA. African genetic diversity: implications for human demographic history, modern human origins, and complex disease mapping. Annu Rev Genomics Hum Genet. 2008;9(1):403–433. doi:10.1146/annurev.genom.9.081307.164258
9. Gray JM, Cohn DL. Tuberculosis and HIV coinfection. Semin Respir Crit Care Med. 2013;34(1):32–43. doi:10.1055/s-0032-1333469
10. Harling G, Ehrlich R, Myer L. The social epidemiology of tuberculosis in South Africa: a multilevel analysis. Soc Sci Med. 2008;66(2):492–505. doi:10.1016/j.socscimed.2007.08.026
11. Oga-Omenka C, Tseja-Akinrin A, Sen P, et al. Factors influencing diagnosis and treatment initiation for multidrug-resistant/rifampicin-resistant tuberculosis in six sub-Saharan African countries: a mixed-methods systematic review. BMJ Glob Health. 2020;5(7):e002280. doi:10.1136/bmjgh-2019-002280
12. Andrews JR, Shah N, Gandhi N, et al. Multidrug-resistant and extensively drug-resistant tuberculosis: implications for the HIV epidemic and antiretroviral therapy rollout in South Africa. J Infect Dis. 2007;196(Suppl s3):S482–90. doi:10.1086/521121
13. Fullwiley D, Gibbon S. Wellcome Trust–Funded Monographs and Book Chapters Genomics in emerging and developing economies. In: Gibbon S, editor. Handbook of Genomics, Health and Society. London (UK): Routledge © 2018 selection and editorial matter Sahra Gibbon, Barbara Prainsack, Stephen Hilgartner and Janelle Lamoreaux, individual chapters, the contributors; 2018:228–237.
14. Chakaya J, Khan M, Ntoumi F, et al. Global Tuberculosis Report 2020 - Reflections on the Global TB burden, treatment and prevention efforts. Int J Infect Dis. 2021;113(Suppl 1):S7–s12. doi:10.1016/j.ijid.2021.02.107
15. Selvaraj P, Uma H, Reetha AM, et al. HLA antigen profile in pulmonary tuberculosis patients & their spouses. Indian J Med Res. 1998;107:155–158.
16. Singh SP, Mehra NK, Dingley HB, et al. Human leukocyte antigen (HLA)-linked control of susceptibility to pulmonary tuberculosis and association with HLA-DR types. J Infect Dis. 1983;148(4):676–681. doi:10.1093/infdis/148.4.676
17. Torrelles JB, Torrelles J, Azad A, et al. Role of C-type lectins in mycobacterial infections. Curr Drug Targets. 2008;9(2):102–112. doi:10.2174/138945008783502467
18. Beharka AA, Gaynor CD, Kang BK, et al. Pulmonary surfactant protein A up-regulates activity of the mannose receptor, a pattern recognition receptor expressed on human macrophages. J Immunol. 2002;169(7):3565–3573. doi:10.4049/jimmunol.169.7.3565
19. Ferguson JS, Voelker DR, McCormack FX, et al. Surfactant Protein D Binds to Mycobacterium tuberculosis Bacilli and Lipoarabinomannan via Carbohydrate-Lectin Interactions Resulting in Reduced Phagocytosis of the Bacteria by Macrophages1. J Immunol. 1999;163(1):312–321. doi:10.4049/jimmunol.163.1.312
20. Salam N, Gupta S, Sharma S, et al. Protective immunity to Mycobacterium tuberculosis infection by chemokine and cytokine conditioned CFP-10 differentiated dendritic cells. PLoS One. 2008;3(8):e2869. doi:10.1371/journal.pone.0002869
21. Vieira F, Kung JW, Bhatti F. Structure, genetics and function of the pulmonary associated surfactant proteins A and D: the extra-pulmonary role of these C type lectins. Ann Anat. 2017;211:184–201. doi:10.1016/j.aanat.2017.03.002
22. Naqvi KF, Endsley JJ. Myeloid C-type lectin receptors in tuberculosis and HIV immunity: insights into co-infection? Front Cell Infect Microbiol. 2020;10:263. doi:10.3389/fcimb.2020.00263
23. Garred P, Larsen F, Seyfarth J, et al. Mannose-binding lectin and its genetic variants. Genes Immun. 2006;7(2):85–94. doi:10.1038/sj.gene.6364283
24. Jo EK. Mycobacterial interaction with innate receptors: tLRs, C-type lectins, and NLRs. Curr Opin Infect Dis. 2008;21(3):279–286. doi:10.1097/QCO.0b013e3282f88b5d
25. Sasindran SJ, Torrelles JB. Mycobacterium Tuberculosis Infection and Inflammation: what is Beneficial for the Host and for the Bacterium? Front Microbiol. 2011;2:2. doi:10.3389/fmicb.2011.00002
26. Carvalho NB, Oliveira FS, Duraes FV, et al. Toll-like receptor 9 is required for full host resistance to mycobacterium avium infection but plays no role in induction of Th1 Responses. Infect Immun. 2011;79(4):1638–1646. doi:10.1128/IAI.01030-10
27. Bowker N, Salie M, Schurz H, et al. Polymorphisms in the pattern recognition receptor mincle gene (CLEC4E) and association with tuberculosis. Lung. 2016;194(5):763–767. doi:10.1007/s00408-016-9915-y
28. Lai YF, Lin T-M, Wang C-H, et al. Functional polymorphisms of the TLR7 and TLR8 genes contribute to Mycobacterium tuberculosis infection. Tuberculosis. 2016;98:125–131. doi:10.1016/j.tube.2016.03.008
29. Mortaz E, Adcock IM, Tabarsi P, et al. Interaction of Pattern Recognition Receptors with Mycobacterium Tuberculosis. J Clin Immunol. 2015;35(1):1–10. doi:10.1007/s10875-014-0103-7
30. Olvany JM, Sausville LN, White MJ, et al. CLEC4E (Mincle) genetic variation associates with pulmonary tuberculosis in Guinea-Bissau (West Africa). Infect Genet Evol. 2020;85:104560. doi:10.1016/j.meegid.2020.104560
31. Zhang Y, Jiang T, Yang X, et al. Toll-like receptor −1, −2, and −6 polymorphisms and pulmonary tuberculosis susceptibility: a systematic review and meta-analysis. PLoS One. 2013;8(5):e63357. doi:10.1371/journal.pone.0063357
32. Puig-Kröger A, Serrano-Gómez D, Caparrós E, et al. Regulated expression of the pathogen receptor dendritic cell-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing nonintegrin in THP-1 human leukemic cells, monocytes, and macrophages. J Biol Chem. 2004;279(24):25680–25688. doi:10.1074/jbc.M311516200
33. Azad AK, Torrelles JB, Schlesinger LS. Mutation in the DC-SIGN cytoplasmic triacidic cluster motif markedly attenuates receptor activity for phagocytosis and endocytosis of mannose-containing ligands by human myeloid cells. J Leukoc Biol. 2008;84(6):1594–1603. doi:10.1189/jlb.0308192
34. Barreiro LB, Ben-Ali M, Quach H, et al. Evolutionary dynamics of human Toll-like receptors and their different contributions to host defense. PLoS Genet. 2009;5(7):e1000562. doi:10.1371/journal.pgen.1000562
35. Tailleux L, Pham-Thi N, Bergeron-Lafaurie A, et al. DC-SIGN induction in alveolar macrophages defines privileged target host cells for mycobacteria in patients with tuberculosis. PLoS Med. 2005;2(12):e381. doi:10.1371/journal.pmed.0020381
36. Boily-Larouche G, Zijenah LS, Mbizvo M, et al. DC-SIGN and DC-SIGNR genetic diversity among different ethnic populations: potential implications for pathogen recognition and disease susceptibility. Hum Immunol. 2007;68(6):523–530. doi:10.1016/j.humimm.2007.02.002
37. Vidal SM, Malo D, Vogan K, et al. Natural resistance to infection with intracellular parasites: isolation of a candidate for Bcg. Cell. 1993;73(3):469–485. doi:10.1016/0092-8674(93)90135-D
38. Gruenheid S, Pinner E, Desjardins M, et al. Natural resistance to infection with intracellular pathogens: the nramp1 protein is recruited to the membrane of the phagosome. J Exp Med. 1997;185(4):717–730. doi:10.1084/jem.185.4.717
39. Serrano-Gómez D, Dominguez-Soto A, Ancochea J, et al. Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin mediates binding and internalization of aspergillus fumigatus conidia by dendritic cells and macrophages. J Immunol. 2004;173(9):5635–5643. doi:10.4049/jimmunol.173.9.5635
40. Quesniaux VJ, Nicolle DM, Torres D, et al. Toll-like receptor 2 (TLR2)-dependent-positive and TLR2-independent-negative regulation of proinflammatory cytokines by mycobacterial lipomannans. J Immunol. 2004;172(7):4425–4434. doi:10.4049/jimmunol.172.7.4425
41. Lu B, Rutledge BJ, Gu L, et al. Abnormalities in monocyte recruitment and cytokine expression in monocyte chemoattractant protein 1-deficient mice. J Exp Med. 1998;187(4):601–608. doi:10.1084/jem.187.4.601
42. Li HT, Zhang TT, Zhou YQ, et al. SLC11A1 (formerly NRAMP1) gene polymorphisms and tuberculosis susceptibility: a meta-analysis. Int J Tuberc Lung Dis. 2006;10(1):3–12.
43. Blackwell JM, Jamieson SE, Burgner D. HLA and infectious diseases. Clin Microbiol Rev. 2009;22(2):370–385. doi:10.1128/CMR.00048-08
44. Ates O, Dalyan L, Müsellim B, et al. NRAMP1 (SLC11A1) gene polymorphisms that correlate with autoimmune versus infectious disease susceptibility in tuberculosis and rheumatoid arthritis. Int J Immunogenet. 2009;36(1):15–19. doi:10.1111/j.1744-313X.2008.00814.x
45. Song W, Li D, Tao L, et al. Solute carrier transporters: the metabolic gatekeepers of immune cells. Acta Pharm Sin B. 2020;10(1):61–78. doi:10.1016/j.apsb.2019.12.006
46. Searle S, Blackwell JM. Evidence for a functional repeat polymorphism in the promoter of the human NRAMP1 gene that correlates with autoimmune versus infectious disease susceptibility. J Med Genet. 1999;36(4):295–299. doi:10.1136/jmg.36.4.295
47. Berrington WR, Hawn TR. Mycobacterium tuberculosis, macrophages, and the innate immune response: does common variation matter? Immunol Rev. 2007;219(1):167–186. doi:10.1111/j.1600-065X.2007.00545.x
48. Selvaraj P, Alagarasu K, Harishankar M, et al. Cytokine gene polymorphisms and cytokine levels in pulmonary tuberculosis. Cytokine. 2008;43(1):26–33. doi:10.1016/j.cyto.2008.04.011
49. Ben-Ali M, Barbouche M-R, Bousnina S, et al. Toll-like receptor 2 Arg677Trp polymorphism is associated with susceptibility to tuberculosis in Tunisian patients. Clin Diagn Lab Immunol. 2004;11(3):625–626. doi:10.1128/CDLI.11.3.625-626.2004
50. Velez DR, Wejse C, Stryjewski ME, et al. Variants in toll-like receptors 2 and 9 influence susceptibility to pulmonary tuberculosis in Caucasians, African-Americans, and West Africans. Hum Genet. 2010;127(1):65–73. doi:10.1007/s00439-009-0741-7
51. Newport MJ, Allen A, Awomoyi AA, et al. The toll-like receptor 4 Asp299Gly variant: no influence on LPS responsiveness or susceptibility to pulmonary tuberculosis in The Gambia. Tuberculosis. 2004;84(6):347–352. doi:10.1016/j.tube.2004.02.001
52. Olesen R, Wejse C, Velez DR, et al. DC-SIGN (CD209), pentraxin 3 and vitamin D receptor gene variants associate with pulmonary tuberculosis risk in West Africans. Genes Immun. 2007;8(6):456–467. doi:10.1038/sj.gene.6364410
53. Khor CC, Chapman SJ, Vannberg FO, et al. A Mal functional variant is associated with protection against invasive pneumococcal disease, bacteremia, malaria and tuberculosis. Nat Genet. 2007;39(4):523–528. doi:10.1038/ng1976
54. Jones BW. Differential roles of Toll-like receptors in the elicitation of proinflammatory responses by macrophages. Ann Rheum Dis. 2001;60(Suppl 3):6–12. doi:10.1136/ard.60.1.6
55. Fitness J, Hill A, Fine P, et al. Large-scale candidate gene study of leprosy susceptibility in the Karonga district of northern Malawi. Am J Trop Med Hyg. 2004;71(3):330–340. doi:10.4269/ajtmh.2004.71.330
56. Chensue SW, Warmington KS, Allenspach EJ, et al. Differential expression and cross-regulatory function of RANTES during mycobacterial (type 1) and schistosomal (type 2) antigen-elicited granulomatous inflammation. J Immunol. 1999;163(1):165–173. doi:10.4049/jimmunol.163.1.165
57. van Crevel R, Ottenhoff TH, Van Der Meer JW. Innate immunity to Mycobacterium tuberculosis. Clin Microbiol Rev. 2002;15(2):294–309. doi:10.1128/CMR.15.2.294-309.2002
58. Saukkonen JJ, Bazydlo B, Thomas M, et al. β-Chemokines Are Induced by Mycobacterium tuberculosis and Inhibit Its Growth. Infect Immun. 2002;70(4):1684–1693. doi:10.1128/IAI.70.4.1684-1693.2002
59. Ben-Selma W, Harizi H, Bougmiza I, et al. Polymorphisms in the RANTES Gene Increase Susceptibility to Active Tuberculosis in Tunisia. DNA Cell Biol. 2011;30(10):789–800. doi:10.1089/dna.2010.1200
60. Lin Y, Gong J, Zhang M, et al. Production of monocyte chemoattractant protein 1 in tuberculosis patients. Infect Immun. 1998;66(5):2319–2322. doi:10.1128/IAI.66.5.2319-2322.1998
61. Flores-Villanueva PO, Ruiz-Morales JA, Song C-H, et al. A functional promoter polymorphism in monocyte chemoattractant protein–1 is associated with increased susceptibility to pulmonary tuberculosis. J Exp Med. 2005;202(12):1649–1658. doi:10.1084/jem.20050126
62. Thye T, Nejentsev S, Intemann CD, et al. MCP-1 promoter variant −362C associated with protection from pulmonary tuberculosis in Ghana, West Africa. Hum Mol Genet. 2009;18(2):381–388. doi:10.1093/hmg/ddn352
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.