Back to Journals » Infection and Drug Resistance » Volume 13
Plasmid-Encoded blaNDM-5 Gene That Confers High-Level Carbapenem Resistance in Salmonella Typhimurium of Pork Origin
Authors Gao Y, Wen J ![]() , Wang S, Xu X, Zhan Z, Chen Z, Bai J, Qu X, Zhang H, Zhang J, Liao M
, Wang S, Xu X, Zhan Z, Chen Z, Bai J, Qu X, Zhang H, Zhang J, Liao M
Received 11 February 2020
Accepted for publication 28 April 2020
Published 19 May 2020 Volume 2020:13 Pages 1485—1490
DOI https://doi.org/10.2147/IDR.S249357
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Yuan Gao,1 Junping Wen,1 Shaojun Wang,1 Xuebin Xu,2 Zeqiang Zhan,1 Zhengquan Chen,1 Jie Bai,1 Xiaoyun Qu,1 Hongxia Zhang,1 Jianmin Zhang,1 Ming Liao1
1National and Regional Joint Engineering Laboratory for Medicament of Zoonoses Prevention and Control, Guangdong Laboratory for Lingnan Modern Agriculture, Key Laboratory of Zoonoses, Ministry of Agriculture, Key Laboratory of Animal Vaccine Development, Key Laboratory of Zoonoses Prevention and Control of Guangdong Province, College of Veterinary Medicine, South China Agricultural University, Guangzhou, Guangdong, People’s Republic of China; 2Department of Microbiology, Shanghai Municipal Centre for Disease Control and Prevention, Shanghai, People’s Republic of China
Correspondence: Ming Liao; Jianmin Zhang
National and Regional Joint Engineering Laboratory for Medicament of Zoonoses Prevention and Control, Guangdong Laboratory for Lingnan Modern Agriculture, Key Laboratory of Zoonoses, Key Laboratory of Animal Vaccine Development, Ministry of Agriculture, Key Laboratory of Zoonoses Prevention and Control of Guangdong Province, College of Veterinary Medicine, South China Agricultural University, No. 483 Wushan Road, Tianhe District, Guangzhou 510642, Guangdong, People’s Republic of China
Tel +86 020 85290240; +86 020 85280240
Fax +86 020 85290240; +86 020 85285282
Email [email protected]; [email protected]
Purpose: Carbapenem resistance is rarely reported in Salmonella Typhimurium, especially from a food origin. Here, we report a plasmid-mediated mobile carbapenem-resistant blaNDM-5 gene in Salmonella Typhimurium isolated from pork in Shanghai, China in 2016.
Patients and Methods: In July 2016, the S. Typhimurium SH160 strain was recovered from minced pork meat purchased from a supermarket in Yangpu District, Shanghai, China. Antimicrobial susceptibility testing, multi-locus sequence typing, conjugation, S1-PFGE, southern hybridization, whole-genome sequencing and data analysis were performed.
Results: This isolate was found to be a ST34 strain and resistant to carbapenems, cephalosporins, and most other commonly used antibiotics. The blaNDM-5 gene was harbored by a 46161-bp IncX3 plasmid which was found to be transferable. The IncX3 plasmid contains a composite cassette, consisting of ISSwil-IS3000-ΔISAba125-IS5-blaNDM-5-bleMBL-trpF-dsbC-IS26-ctuA1-ΔumuD. In addition, this strain was found to harbor an additional 161706-bp IncHI2 plasmid which carries nine resistant genes, such as aadA1, aadA3, aph(3ʹ)-la, sul1, sul2, sul3, floR, cmlA and dfrA12.
Conclusion: We reported the S. Typhimurium with transferable IncX3 plasmid harboring blaNDM-5 gene from minced pork. We characterized the complete genetic features of the plasmid, which demonstrated the potential for spreading in different bacterial pathogens. Therefore, extensive surveillance and monitoring for carbapenem-resistant bacterium in the food chain and public health are urgently required.
Keywords: S. Typhimurium, pork, blaNDM-5, public health
Introduction
Salmonella is an important foodborne pathogenic bacterium that causes considerable morbidity and mortality. The burden of foodborne non-typhoidal serovars of Salmonella enterica (NTS) infections is extremely high, with a global estimation of 78.4 million cases, 28,693 deaths, and over 2 million disability adjusted life years (DALYs) in 2010.1 The emergence and spread of multidrug resistance (MDR) have occurred in Salmonella enterica serovar Typhimurium strains, which has had a significant impact on the effectiveness of current strategies to control and manage diseases associated with foodborne infections.1,2
The trend of MDR in Salmonella has increased each year in China.3 Of particular concern is the emerging resistance to carbapenems, since these agents are often regarded as the ‘last-line’ of effective therapy for treating infections caused by MDR Gram-negative bacteria.4 Moreover, there has been an increase in the spread of carbapenemase-encoding genes among nosocomial enteric bacteria, particularly Klebsiella pneumoniae and Escherichia coli.5 Although carbapenemase-producing bacteria have been reported in clinical infections, there are sporadic reports of Salmonella, especially those involving a food origin. Due to the emergence of such resistance in foodborne pathogens, active surveillance and monitoring for carbapenem-resistant bacteria in the food chain is urgently required, as they may be transferred to both humans and the environment.6
Therefore, this study aims to describe the occurrence of carbapenemase-carrying and MDR Salmonella Typhimurium isolated from retail pork in Shanghai and investigate the mechanism of the plasmid-mediated carbapenem-resistant Salmonella enterica serovar. Whole-genome-sequencing (WGS) was used to determine the complete plasmid profile, followed by a comparative genetic analysis, to lay the foundation for follow-up studies related to food safety and public health issues caused by Salmonella.
Materials and Methods
Specimen Collection, Isolate Identification and Multilocus Sequence Typing (MLST)
In July 2016, the S. Typhimurium SH160 strain was recovered from minced pork meat in a supermarket in Yangpu District, Shanghai, China. Salmonella was isolated according to the US FDA Bacteriological Analytical Manual.7 Isolates with typical Salmonella phenotypes were further confirmed using API identification kits (bioMérieux, France). The O and H antigens were characterized using slide agglutination with Salmonella diagnostic serum (S&A Reagents Lab, Bangkok, Thailand). The serological determination of Salmonella serotypes was performed in accordance with the Kauffmann-White scheme.8 Multi-locus sequence typing (MLST) was performed using primer sets (Supplemental Table 1). Sequences of seven housekeeping genes (aroC, dnaN, hemD, hisD, purE, sucA, and thrA) were compared with the available MLST database (http://mlst.warwick.ac.uk/mlst/dbs/Senterica) to get the allele number and sequence typing (ST) number for each isolate. Sequence information for newly assigned alleles and STs was deposited in the MLST database.
Antimicrobial Susceptibility Testing and Identification of the blaNDM-5 Strain
The susceptibility of S. Typhimurium SH160 strain to different antibiotics was determined using the standard agar dilution method and interpreted in accordance with the Clinical and Laboratory Standards Institute (CLSI) standard.9 E. coli strain ATCC25922 was used as a quality control strain. The PCR amplification of the blaNDM-5 gene was performed to identify the presence of blaNDM-5 gene (Primers are in the Supplemental Table 2).
Conjugation, S1-PFGE, and Southern Hybridization Assay
To test the transferability of the blaNDM-5-bearing plasmid, conjugation by filter mating was performed between S. Typhimurium SH160 strain and streptomycin-resistant E. coli EC600. Both the donor and recipient strains were cultured to the exponential phase in LB broth. The donor and recipient bacteria were mixed at a 1:4 ratio and incubated overnight at 37°C. The mixture was then spread onto a selective MacConkey agar plate containing imipenem (2 mg/L), and streptomycin (3000 mg/L) to select for transconjugants that had acquired the blaNDM-5 bearing plasmid. To estimate the size and location of blaNDM-5 in the parental strain and the transconjugants, DNA linearization with S1 nuclease followed by PFGE (S1-PFGE) and Southern hybridization analyses were performed. Southern hybridization was performed in accordance with the manufacturer’s instructions of the digoxigenin (DIG)-High Prime DNA Labelling and Detection Starter Kit II (Roche Diagnostics) using blaNDM-5 digoxigenin-labelled probes (primers for the probe are in the Supplemental Table 2).
Whole-Genome Sequencing and Comparison Analysis
The whole S. Typhimurium SH160 genome was sequenced using PacBio RS II Single Molecule Real-Time (SMRT) and Illumina sequencing platforms. The raw reads were then assembled into a contig using the hierarchical genome assembly process (HGAP) and Canu.10,11 The complete plasmid sequence was predicted with GLIMMER and annotated with the NR, Swiss-Prot, Pfam, GO, COG, and KEGG databases using sequence alignment tools, including BLAST, Diamond and HMMER.12–15 Easyfig was used in comparative analysis (Genbank accession number KY435936, KF220657, KF877335).16,17 ResFinder and PlasmidFinder were used to detect drug-resistant genes and the plasmid sequencing type.18,19
Results
S. Typhimurium Isolation and Antimicrobial Susceptibility Testing
Multi-locus sequence typing analysis revealed that SH160 belonged to sequence-type 34 (ST34). The S. Typhimurium strain, SH160, was found to be resistant to a wide range of antibiotics, including imipenem, ampicillin, cefotaxime, cefepime, gentamycin, nalidixic acid, ofloxacin, florfenicol, and sulfisoxazole (Table 1). However, the strain was susceptible to amikacin, ciprofloxacin tetracycline, streptomycin, and polymyxin B.
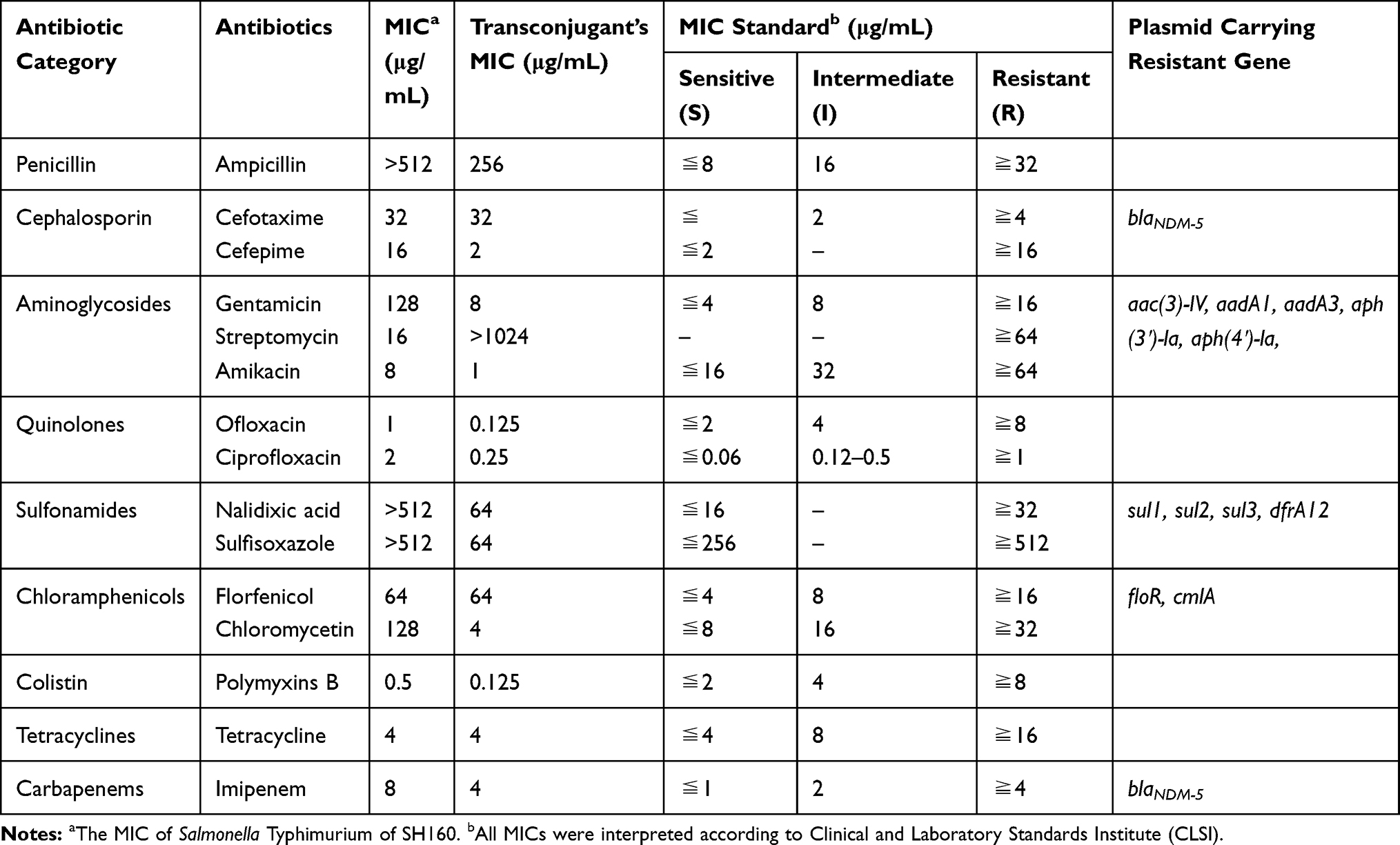 |
Table 1 The Minimum Inhibitory Concentration (MIC) of High-Resistant Salmonella Typhimurium SH160 |
Conjugation, S1-PFGE, and Southern Hybridization
The blaNDM-5 gene was successfully transferred to E. coli EC600. The transconjugant was resistant to cephalosporin, imipenem, ampicillin, gentamycin, florfenicol, and sulfisoxazole. S1-PFGE and Southern hybridization indicated that the blaNDM-5 gene was located on a conjugative plasmid, with a size of approximately 46 kb, designated pNDM5_SH160 (Figure 1).
 |
Figure 1 S1-PFGE and hybridization indicating the transfer of single plasmid harbouring blaNDM-5. Lane 1, M, H-9812 (PFGE marker strain, S. Braenderup H9812; Lane 2, SH160, S. Typhimurium SH160 (donor); Lane 3, Tr160, EC600 (transconjugant)). |
Whole-Genome Sequencing and Comparison Analysis
A 46161-bp blaNDM-5-carrying IncX3 plasmid, pNDM5_SH160, and a 161706-bp IncHI2 plasmid were identified in SH160, which exhibited G+C content of 46.74% and 46.48%, respectively. These plasmids also contained 62 and 163 predicted coding sequences (CDSs), respectively. The blaNDM-5 gene is adjacent to an incomplete ISAba125, which was interrupted by the insertion of IS5 and truncated by an insertion of IS3000 at its left end. A zinc metalloproteinase-encoding gene, mpr, lies further upstream of blaNDM-5. Several putative open reading frames (ORFs) of an unknown function and a truncated ISSwil was found to be present between mpr and IS3000. There are bleMBL (mediating bleomycin resistance), trpF (encoding a phosphoribosyl anthranilate isomerase), dsbC (encoding an oxidoreductase), a remnant of ctuA1 (encoding an iron-tolerant protein), and a truncated umuD gene (encoding a mutagenesis protein) nearby (Figure 2). In addition to the blaNDM-5 gene, multiple resistance genes were identified, including aadA1, aadA3, aph(3ʹ)-la, aph(4ʹ)-la, sul1, sul2, sul3, floR, cmlA, and dfrA12, which were all located in the IncHI2 plasmid (Table 1).
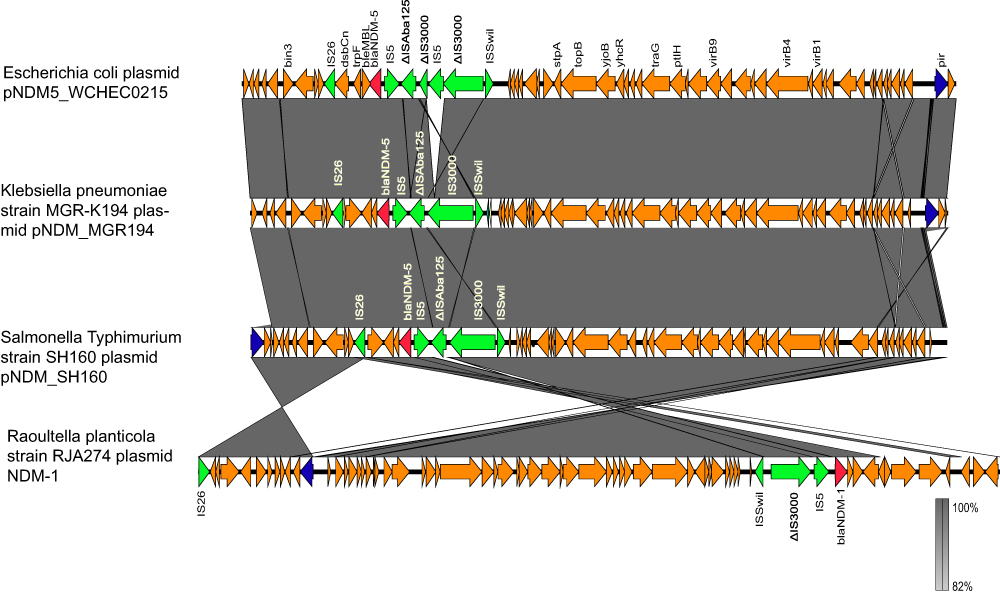 |
Figure 2 Features of the genetic structure of pNDM5_WCHEC0215 (Genbank accession number KY435936), pNDM_MGR194 (Genbank accession number KF220657), pNDM5_SH160, and RJA274 plasmid (Genbank accession number KF877335). The blaNDM-1/5 gene is shown in red. IS3000, ISAba125, and other insert sequences are shown in green. The replicons are shown in blue. The rest of the genes are shown in orange. |
The results showed that the blaNDM-5-bearing plasmid shared 100% query coverage with several other blaNDM-5-harbouring plasmids, as well as >99% sequence identity with the plasmid, pNDM_MGR194, which was isolated in India.20 The genetic background of blaNDM-5 in these two plasmids is identical (ISSwil-IS3000-ΔISAba125-IS5-blaNDM-5-bleMBL-trpF-dsbC-IS26-ctuA1-ΔumuD). Such a background of the blaNDM-5 gene in IncX3 plasmids is highly similar to that of blaNDM-1 and blaNDM-5 in several other IncX3 plasmids (Figure 2).
Discussion
Carbapenems are critical ‘last-resort’ antibiotics reserved for the treatment of serious infections caused by certain highly resistant Gram-negative bacteria. In contrast to other Enterobacteriaceae (eg, K. pneumoniae, E. coli, and Enterobacter spp.), carbapenem-resistant Salmonella has rarely been reported.21–23 We report the successful isolation and characterization of a metallo-beta-lactamase (MβL) encoding blaNDM-5 gene harboured in S. Typhimurium isolated from pork meat in Shanghai, China. This strain together with the previous isolation from Jiangsu province confirms the prevalence of blaNDM-5 in food chains.24 Therefore, there is a potential risk of cross-contamination in food, animals, and humans. Hence, identifying the mechanism of the spread of carbapenem-resistant Salmonella in the environment has become a substantial global health concern.
According to the MLST results, the SH160 strain belongs to ST34, which is one of the most prevalent types of S. Typhimurium. Moreover, ST34 clones have raised international concern regarding their rapid prevalence and high level of drug resistance. A previous study demonstrated that S. Typhimurium ST34 clones experienced a rapid expansion in China and exhibited resistance to cephalosporin antibiotics.25 Notably, S. Typhimurium ST34 SH160 strain is not only resistant to several common antibiotics due to harbouring of a variety of resistant genes in the IncHI2 plasmid, but also resistant to carbapenem and cephalosporins due to the blaNDM-5 gene in IncX3 plasmid. Therefore, the surveillance of S. Typhimurium ST34 should be strengthened to reduce the probability of drug-resistant plasmid transmission.
In this study, we identified the pNDM5_SH160 and the IncHI2 plasmids were transferable indicating the potential risk of horizontal transmission of drug resistance. In addition, the comparison of pNDM5_SH160 showed that E. coli, Klebsiella pneumoniae, and Raoutella planticola share a mostly conserved plasmid backbone (ISSwil-IS3000-ΔISAba125-IS5-blaNDM-1/5-bleMBL-trpF-dsbC-IS26-ctuA1-ΔumuD), including the blaNDM-5/blaNDM-1 gene, which shows the prevalence of the plasmid with a strong transmissibility among different species widely (Figure 2). On the other hand, IncX-type plasmids are narrow-host range plasmids of Enterobacteriaceae and include at least five subtypes: IncX1-IncX5. In addition, some IncX3 plasmids carrying ESBL and/or the carbapenemase gene from various species of Enterobacteriaceae found in several countries have been completely sequenced.26
Since the blaNDM−5 gene was first identified from E. coli in the UK,26 strains containing the blaNDM-5 have emerged in several countries.20,27 Nevertheless, the blaNDM-5 gene has not been identified in Salmonella enterica of pork origin in Shanghai.6 NDM-5 differs from NDM-1 by two amino acid substitutions (Val88Leu and Met154Leu) and confers high-level resistance to carbapenems and broad-spectrum cephalosporins.28 As a result, plasmids carrying NDM variants might be widely disseminated, posing a potential public health threat.
Conclusion
We reported the carbapenem-resistant Salmonella Typhimurium from minced pork and identified that carbapenem-resistance was mediated by a transferable plasmid. Complete genetic features of pNDM5_SH160 and comparison were performed. Such emergence of NDM carrier plasmids indicates that we should develop strategies to control the spread. Extensive surveillance and monitoring of the carbapenem-resistant bacterium in the food chain are urgently required.
Acknowledgments
This research was supported by the National Key R&D Program of China (2017YFC1600101, 2018YFD0500500); National Natural Science Foundation of China (31972762); Pearl River S&T Nova Program of Guangzhou (201806010183); Province Science and Technology of Guangdong Research Project (2017A020208055); Guangdong Key S&T Program (Grant no. 2019B020217002) from the Department of Science and Technology of Guangdong Province; Walmart Foundation (SA1703162); National Broiler Industry Technology System Project (cARS-41-G16).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kirk MD, Pires SM, Black RE, et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: a data synthesis. PLoS Med. 2015;12:1–21.
2. Scallan E, Hoekstra RM, Mahon BE, Jones TF, Griffin PM. An assessment of the human health impact of seven leading foodborne pathogens in the United States using disability adjusted life years. Epidemiol Infect. 2015;143:2795–2804. doi:10.1017/S0950268814003185
3. Wang J, Li Y, Xu X, et al. Antimicrobial resistance of Salmonella enterica serovar typhimurium in Shanghai, China. Front Microbiol. 2017;8:1–10. doi:10.3389/fmicb.2017.00001
4. Kumarasamy KK, Toleman MA, Walsh TR, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect Dis. 2010;10:597–602. doi:10.1016/S1473-3099(10)70143-2
5. Zhang Q, Lv L, Huang X, et al. Rapid increase in carbapenemase-producing enterobacteriaceae in retail meat driven by the spread of the bla NDM-5 -carrying IncX3 plasmid in China from 2016 to 2018. Antimicrob Agents Chemother. 2019;63. doi: 10.1128/AAC.00573-19
6. Fernández J, Guerra B, Rodicio MR. Resistance to carbapenems in non-typhoidal Salmonella enterica serovars from humans, animals and food. Vet Sci. 2018;5.
7. Andrews WH, Jacobson A, T H. Bacteriological and analytical method. Salmonella Bacteriol Anal Method. 2009;1–21.
8. Popoff MY, Bockemühl J, Gheesling LL. Supplement 2002 (no. 46) to the Kauffmann-White scheme. Res Microbiol. 2004;155:568–570. doi:10.1016/j.resmic.2004.04.005
9. Dolinsky AL. M100 performance standards for antimicrobial susceptibility testing. Clin Lab Stand Ins. 2017; 8:0956–4624.
10. Ellington MJ, Kistler J, Livermore DM, Woodford N. Multiplex PCR for rapid detection of genes encoding acquired metallo-β-lactamases [1]. J Antimicrob Chemother. 2007;59:321–322. doi:10.1093/jac/dkl481
11. Chin CS, Alexander DH, Marks P, et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat Methods. 2013;10:563–569. doi:10.1038/nmeth.2474
12. Koren S, Walenz BP, Berlin K, Miller JR, Bergman NH, Phillippy AM. Canu: scalable and accurate long-read assembly via adaptive k -mer weighting and repeat separation. Genome Res. 2017;27:722–736. doi:10.1101/gr.215087.116
13. Jensen LJ, Julien P, Kuhn M, et al. eggNOG: automated construction and annotation of orthologous groups of genes. Nucleic Acids Res. 2008;36:250–254. doi:10.1093/nar/gkm796
14. Finn RD, Bateman A, Clements J, et al. Pfam: the protein families database. Nucleic Acids Res. 2014;42:222–230. doi:10.1093/nar/gkt1223
15. Kanehisa M. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28:27–30. doi:10.1093/nar/28.1.27
16. Bairoch A. The SWISS-PROT protein. Nucleic Acids Res. 1994;22:3578–3580.
17. Salzberg SL, Deicher AL, Kasif S, White O. Microbial gene identification using interpolated Markov models. Nucleic Acids Res. 1998;26:544–548. doi:10.1093/nar/26.2.544
18. Sullivan MJ, Petty NK, Beatson SA. Easyfig: A genome comparison visualizer. Bioinformatics. 2011;27:1009–1010. doi:10.1093/bioinformatics/btr039
19. Carattoli A, Zankari E, Garciá-Fernández A, et al. In Silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58:3895–3903. doi:10.1128/AAC.02412-14
20. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67:2640–2644. doi:10.1093/jac/dks261
21. Krishnaraju M, Kamatchi C, Jha AK, et al. Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J Med Microbiol. 2015;33:30–38. doi:10.4103/0255-0857.148373
22. Borowiak M, Szabo I, Baumann B, et al. VIM-1-producing Salmonella Infantis isolated from swine and minced pork meat in Germany. J Antimicrob Chemother. 2017;72:2131–2133. doi:10.1093/jac/dkx101
23. Fischer J, Rodrìguez I, Schmoger S, et al. Salmonella enterica subsp. enterica producing VIM-1 carbapenemase isolated from livestock farms. J Antimicrob Chemother. 2013;68:478–480. doi:10.1093/jac/dks393
24. Wang W, Baloch Z, Peng Z, et al. Genomic characterization of a large plasmid containing a bla NDM-1 gene carried on Salmonella enterica serovar Indiana C629 isolate from China. BMC Infect Dis. 2017;17:1–8. doi:10.1186/s12879-017-2515-5
25. Wang Z, He J, Li Q, et al. First detection of NDM-5-positive salmonella enterica serovar typhimurium isolated from retail pork in China. Microb Drug Resist. 2019;00:5–8.
26. Wong MHY, Yan M, Chan EWC, Liu LZ, Kan B, Chen S. Expansion of salmonella enterica serovar typhimurium ST34 clone carrying multiple resistance determinants in China. Antimicrob Agents Chemother. 2013;57:4599–4601. doi:10.1128/AAC.01174-13
27. Hornsey M, Phee L, Wareham DW. A novel variant, NDM-5, of the New Delhi metallo-β-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob Agents Chemother. 2011;55:5952–5954. doi:10.1128/AAC.05108-11
28. Yang L, Hu X, Xu X, et al. Salmonella enterica serovar Typhimurium ST34 co-expressing bla NDM-5 and bla CTX-M-55 isolated in China. Emerg. Microbes Infect. 2017;6:e61–3.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.