Back to Journals » Journal of Inflammation Research » Volume 18
Plasma Exosomal miR-17-5p Regulates Macrophage Polarization by Targeting Bcl11b in Sepsis-Induced Lung Injury
Authors Xian Y ![]() , Sun Y, Wang L, Lin L, Wu Z, Zhang K, Chen R
, Sun Y, Wang L, Lin L, Wu Z, Zhang K, Chen R
Received 24 February 2025
Accepted for publication 19 July 2025
Published 8 August 2025 Volume 2025:18 Pages 10651—10668
DOI https://doi.org/10.2147/JIR.S524742
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Tara Strutt
Ying Xian,1,2,* Yinfang Sun,2,* Lifu Wang,3 Lin Lin,1 Zhongdao Wu,4 Kouxing Zhang,2 Rui Chen1
1Department of Respiratory Diseases, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou, Guangdong, People’s Republic of China; 2Department of General Intensive Care Unit, The Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China; 3KingMed School of Laboratory Medicine, Guangzhou Medical University, Guangzhou, Guangdong, People’s Republic of China; 4Department of Parasitology, Zhongshan School of Medicine, Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Rui Chen, Department of Respiratory Diseases, Sun Yat-sen Memorial Hospital, Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China, Email [email protected] Kouxing Zhang, Department of General Intensive Care Unit, The Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China, Email [email protected]
Background: Sepsis is a life-threatening syndrome characterized by overwhelming inflammation and immune dysregulation, commonly complicated by acute lung injury. Patients with underlying conditions such as diabetes, malignancy, and chronic liver disease are particularly vulnerable. Dysregulated macrophage polarization plays a pivotal role in sepsis progression. Although exosomal microRNAs (miRNAs) have emerged as key immune modulators, the precise role of plasma-derived exosomal miR-17-5p in this process remains poorly defined. The transcription factor Bcl11b, previously linked to immune cell regulation, has not yet been studied in the context of sepsis-associated macrophage reprogramming.
Methods: Extracellular vehicles (EVs) were isolated from the plasma of sepsis patients and healthy controls. A series of in vitro and in vivo experiments were conducted to investigate the effect of exosomal miR-17-5p on macrophage polarization, using qRT-PCR, flow cytometry, ELISA, and Western blot analyses. Transcriptome sequencing and dual-luciferase reporter assays were used to explore the regulatory relationship between miR-17-5p and Bcl11b.
Results: Plasma exosomes derived from sepsis patients exhibited reduced levels of miR-17-5p and promoted M1 macrophage polarization, characterized by increased iNOS and pro-inflammatory cytokines. Overexpression of miR-17-5p inhibited M1 polarization and alleviated inflammatory injury both in LPS-treated macrophages and in a CLP-induced mouse model. Mechanistically, miR-17-5p directly targeted the 3′UTR of Bcl11b, suppressing its expression. Restoration of Bcl11b reversed the anti-inflammatory effects of miR-17-5p, reinforcing M1 polarization and exacerbating lung injury.
Conclusion: Plasma exosomal miR-17-5p promotes macrophage M1 polarization by targeting Bcl11b and contributes to sepsis-induced lung injury. These findings highlight a previously unrecognized miR-17-5p–Bcl11b regulatory axis and suggest a potential biomarker and therapeutic target for sepsis.
Keywords: miR-17-5p, Bcl11b, extracellular vesicles, macrophages polarization, sepsis, acute lung injury
Introduction
Sepsis is a life-threatening condition caused by a dysregulated and harmful host response to infection with pathogenic microorganisms, leading to systemic inflammation and multiple organ dysfunction.1–3 The lungs are especially susceptible to injury due to sepsis, increasing the risk of sepsis patients developing acute lung injury (ALI) and, in extreme cases, acute respiratory distress syndrome (ARDS).4 Due to lack of effective clinical medical treatments, both the incidence and mortality rates are high.5 The etiology of sepsis-triggered ALI encompasses the rupture of alveolar walls, the impairment of the alveolar-capillary barrier, and the outpouring of protein-rich fluid. This sequence of events leads to heightened permeability in alveolar blood vessels and a reduction in surfactant within the alveoli, adversely affecting pulmonary gas exchange and inflicting substantial damage on the lungs, as well as diminishing pulmonary function.6 Therefore, it is necessary to delve into the lung damage caused by sepsis to uncover novel diagnostic and therapeutic approaches. In addition to structural lung injury, the host’s underlying health status plays a crucial role in determining susceptibility and response to sepsis-related complications.
It is also worth noting that individuals with underlying conditions—such as diabetes, malignancies, or chronic liver disease—are more vulnerable to the onset and complications of sepsis, and their immune responses may vary accordingly, further complicating the inflammatory progression of sepsis-induced ALI. To understand this complexity, it is essential to examine the cellular and molecular mechanisms driving inflammation during sepsis.
Sepsis is characterized by an imbalanced immune reaction to infection in the host.7 Multiple pro-inflammatory substances that accumulate during an inflammatory response, in addition to impaired immune function, are involved in the pathophysiology of sepsis.8 Macrophages are essential components of the innate immune system, where M1 and M2 phenotypes play significant roles in inflammation. M1 macrophages are primarily associated with promoting inflammation, while M2 macrophages are typically involved in downregulating inflammatory responses.9 Key inflammatory markers involved in M1 polarization include TNF-α, IL-1β, iNOS, and GBP5, which are crucial mediators in amplifying local and systemic inflammatory signaling. Conversely, M2 markers such as CD206, TGF-β, and Arg-1 contribute to immune resolution and tissue repair. Remarkably, survivors of sepsis have been observed to experience prolonged immunosuppression, leading to macrophage dysfunction and inadequate wound healing.10 An imbalance between M1 and M2 macrophages can promote sepsis occurrence and exacerbate its progression.11 Alveolar macrophages act as the initial defense against inhaled particles and microbes, being vital in preserving the balance of the pulmonary immune system.12 Research has shown that in mice with sepsis-induced ALI, pyroptosis of alveolar macrophages is associated with their polarization towards towards M1 phenotype.13 Therefore, it might be essential to create treatment plans for sepsis that focus on employing macrophages to modify the immune response.
Extracellular vehicles (EVs) represent a variety of cell-derived membranous structures, which include exosomes and macrovesicles, emerging either from the endosomal compartment or directly from the cell’s plasma membrane.14 These vesicles facilitate the conveyance of a range of cellular constituents, including proteins, mRNA, miRNAs, DNA, and lipids, across extended distances, thereby impacting a multitude of physiological and pathological processes.15 EVs play a broad role in sepsis, promoting inflammation, oxidation, and apoptosis, mediating microvascular dysfunction and multi-organ damage.14 A recent study revealed that approximately 46 miRNAs were markedly elevated in lung tissue of mice following lipopolysaccharide (LPS) stimulation, highlighting their potential involvement in the pathogenesis of lung injury.16 EVs from sepsis plasma are known to mediate neuronal inflammation in the brain, which is facilitated by miRNAs and innate immune signaling.17 Research has shown that exosomal miR-21 from mesenchymal stem cells could drive M2 polarization of macrophages and mitigate the effects of sepsis.18 Additionally, neutrophil-derived exosomal miR-30d-5p has been shown to induce a shift towards M1 phenotype in macrophages and to trigger pyroptosis in the context of sepsis-related ALI.19 Among the many miRNAs transported by EVs, miR-17-5p has drawn increasing attention due to its regulatory roles in inflammation and tissue injury.
Among the many miRNAs implicated in sepsis, miR-17-5p has been identified as a key regulator of gene expression, particularly affecting vascular and pulmonary processes. However, its immune-modulatory role remains context-dependent, with unclear upstream triggers and cellular targets in septic inflammation. MiR-17-5p was identified to be a vital regulator of gene expressions in sepsis,20 and its deregulated biological processes included blood vessel development and lung development in sepsis-induced ALI.21 We previously indicated miR-17-5p in bronchoalveolar lavage fluid-EVs could potentially serve as a diagnostic marker and may play a role in lung inflammation.22 A decrease in exosomal miR-17-5p levels has been associated with fibrosis development in individuals with sustained heightened sensitivity to inflammatory triggers after experiencing COVID-19-related ARDS.23 In studies examining the effects of miR-17-5p, it has been demonstrated that increased levels of this miRNA can lead to a decrease in inflammatory cytokines levels, specifically TNF-α and IL-1β, within macrophages activated by LPS.24 Despite these findings, its role in exosome-mediated macrophage polarization during sepsis-induced ALI has not been well elucidated. This gap in understanding forms the basis of our current investigation.
Given that miR-17-5p is known to regulate gene expression post-transcriptionally, we hypothesized it might target key transcription factors involved in macrophage activation. Bcl11b, a zinc finger transcription factor primarily known for its function in T cell development and neuroimmune modulation, has recently been implicated in macrophage phenotype regulation.25 However, its role in sepsis-related innate immunity remains underexplored. To date, no study has directly examined the potential regulatory relationship between exosomal miR-17-5p and Bcl11b in alveolar macrophages under septic conditions.
Based on the above background, the objective of this research was to clarify the mechanisms by which miR-17-5p influences macrophage activation and to enhance our comprehension of the development of sepsis-induced ALI. In our research, we explored the role of plasma-derived EVs from sepsis patients by inducing macrophage M1 polarization. Our results indicate a novel pathway that may enhance macrophage activation and death, thereby amplifying inflammation following sepsis and causing damage to the lungs.
Materials and Methods
Collection of Clinical Samples
The research included 46 samples (19 health control and 27 patients with sepsis) at the Third Affiliated Hospital of Sun Yat-sen University from October 2021 to October 2023. Participants were categorized into two groups: sepsis group and control group. Blood samples were obtained from sepsis patients and controls. The patients (aged 18–70 years) were diagnosed with sepsis in the ICU of the Third Affiliated Hospital. Sepsis was identified based on the updated consensus definition established by the Third International Task Force for Sepsis and Septic Shock (Sepsis-3 definition).26 Inclusion criteria were as follows: (1) ICU admission due to infection with a confirmed source; (2) hospitalization exceeding 24 h; (3) diagnosis of sepsis based on the Sepsis-3.0 definition; for septic shock, mean arterial pressure ≤65 mmHg (1 mmHg = 1.33 kPa) and serum lactate ≥2 mmol/L after adequate fluid resuscitation were required; (4) availability of complete clinical data. Exclusion criteria included: (1) ICU stay less than 24 h; (2) age under 18 years; (3) pregnant or lactating women; (4) patients with autoimmune diseases or malignancies receiving immunosuppressive therapy. Patients with common chronic comorbidities such as hypertension, diabetes mellitus, chronic obstructive pulmonary disease (COPD), or renal disease were not excluded unless they met the above exclusion criteria. No significant differences were found between the healthy control group and patients with sepsis in terms of age, gender, or common comorbidities, including hypertension, diabetes, COPD, and renal disease (P > 0.05 for all; Table S1). After diagnosis within 24 h, plasma from 10 mL of whole blood was obtained and stored in a −80 °C refrigerator for further EVs extraction. All participants provided their written consent after being fully informed about the study. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. Ethical approval was obtained from the Ethics Committee of the Third Affiliated Hospital of Sun Yat-sen University (No. [2022]02–216-01).
Purification and Identification of EVs
EVs were isolated via ultracentrifugation. Plasma samples were melted at 37°C. The sample was moved to a fresh centrifuge tube and subjected to centrifugation at 2, 000 × g at a temperature of 4°C for a duration of 30 min. The resulting supernatant was meticulously decanted into another centrifuge tube and centrifuged at 10, 000 × g at 4°C for 45 min to pellet larger vesicles. Supernatant was then passed through a 0.45 μm filter to remove any remaining debris, and filtrate was collected. This filtrate was subsequently transferred to a new centrifuge tube and centrifuged at 100, 000 × g for 70 min at 4°C. After aspirating supernatant, pellet was re-suspended in 10 mL of pre-cooled PBS and centrifuged again at 100, 000 × g for a duration of 70 min at 4°C. After supernatant was discarded, EVs were resuspended in 200 μL of pre-chilled PBS, and EVs were stored at −80°C for preservation. Transmission electron microscopy (TEM) was employed for EVs analysis. Additionally, EVs were analyzed using nanoparticle tracking analysis (NTA). Furthermore, EV markers (CD63 and CD81) were confirmed using flow cytometry.27
Cell Culture and Treatment
The murine alveolar macrophages (MH-S), acquired from ATCC (CRL-2019, USA), were cultured in RPMI 1640 enriched with 10% FBS and 1% penicillin/streptomycin. MH-S were maintained in a humidified atmosphere at 37°C with 5% CO2. MH-S were treated with LPS (1 μg/μL) to simulate sepsis, and peripheral blood exosomes from healthy and sepsis patients were treated respectively to investigate the function of EVs from different sources, which were divided into LPS+N-EXO group and LPS+S-EXO group. Protein samples extracted from the peripheral blood EVs were measured by a BCA kit (Beyotime, China) for protein concentration evaluation. Based on exosome protein concentration, 10 μg of exosomes were added to a 6-well plate per well and incubated with MH-S cells for 24 h. In addition, MH-S were treated with 1 μg/μL LPS to simulate sepsis, and mimic-negative control (NC) and mimic-miR-17-5p were transfected into Control, LPS+mimic-NC, and LPS+mimic-miR-17-5p groups. Furthermore, we performed overexpression of Bcl11b, and the experimental groups were divided as follows: NC group (MH-S transfected with PCDH-NC and mimics NC, with the solvent of LPS added), LPS+NC group (MH-S transfected with PCDH-NC and mimics NC, with 1 μg/mL LPS added), LPS+NC+miR-17-5p group (MH-S transfected with PCDH-NC and miR-17-5p mimics, with 1 μg/mL LPS added), and LPS+Bcl11b-OE+miR-17-5p group (MH-S transfected with PCDH-Bcl11b and miR-17-5p mimics, with 1 μg/mL LPS added).
Cell Transfection
MH-S were plated in 6-well plates with a concentration of 1×105 cells/mL, and the transfection process was initiated once the cells reached a confluence level of approximately 60–70%. 100 pmol of miR-17-5p mimics or NC, PCDH-Bcl11b or NC was combined with 250 μL of Opti-MEM, while 5 μL of Lipofectamine RNAiMax was separately mixed with 250 μL of Opti-MEM for a duration of 5 min. Subsequently, a mixture of RNA and liposomes was incubated for 20 min prior to transfection into cells. Cells were harvested for further analysis 48 h post-transfection.
Establishment of Septic ALI Model
Male C57BL/6 mice (8–10 weeks and 21–23 g) were procured from Laboratory Animal Center of Sun Yat-sen University in China. Upon arrival, the mice were allowed to acclimate for a week in a climate-controlled vivarium, maintained at 22–25°C and 50% relative humidity, with a 12-h alternating light-dark cycle. They had free access to food and water throughout this period. Septic ALI model was induced by cecal ligation and puncture (CLP).28 The experiment was randomized into Sham, CLP, and CLP +agomir-17-5p groups, with 15 mice in each group. The animals were anesthetized using an intraperitoneal injection of sodium pentobarbital (50 mg/kg), followed by transcardiac perfusion with 100 mL of 0.9% saline. In the septic C57BL/6 mice model, a single intravenous injection of miR-17-5p agomir or agomir-NC was administered via tail vein. In brief, miR-17-5p agomir or agomir-NC was injected into mice 3 consecutive days before CLP surgery through tail vein at a total injection volume of 0.25mg/day/mouse. The survival rate of CLP mice with or without miR-17-5p agomir was analyzed by Log rank test. Lung tissues were then immediately dissected at 48 h post-injury and snap-frozen in liquid nitrogen. All animal procedures for the study were conducted in compliance with protocol approved by the Institutional Animal Care and Use Committee at the Medical College of Sun Yat-sen University (Approval Number SYSU-IACUC-2022-000786). Animal experiments were performed in accordance with ARRIVE guidelines and the IACUC Handbook (third edition).
Quantitative Reverse Transcription PCR (qRT-PCR)
qRT-PCR was utilized to measure miR-17-5p, miR-193a-5p, iNOS, Arg-1, IL-1β, IL-6, TGF-β, YM1, CCL24, Serpina1c, Zfyve9, Gm20708, Cebpb, Cxcr3, Aak1, Kcanb1, Bcl11b, and Bal6 levels in plasma-EVs, MH-S, or lung tissues. Total RNA was extracted by TriQuick Reagent (R1100, Solarbio). Further, RNA samples (2 µg) were reverse transcribed into cDNA. 5×RT SuperMix for qPCR (K1074, APEXbio) and 2×SYBR Green qPCR Master Mix (K1070, APEXbio) were applied in iQ5 Multicolor Real-Time PCR Detection System (582BR 005500, BIO-RAD) to assess genes expression. The relative gene levels were analyzed using 2−ΔΔCt method. GAPDH or U6 served as the internal control. Primers are shown in Tables 1 and 2.
 |
Table 1 The mRNA-Specific Primer Sequences |
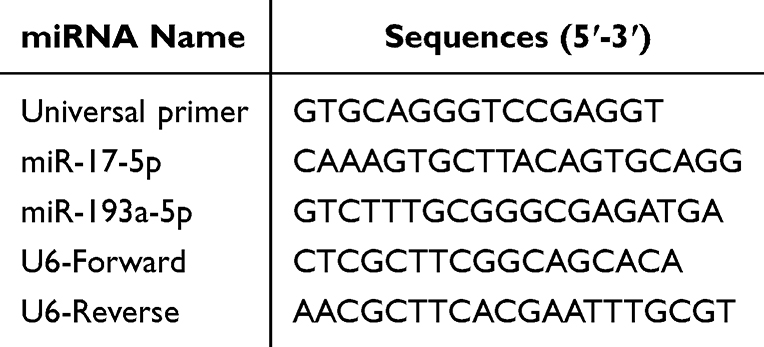 |
Table 2 The miRNA-Specific Primer Sequences |
Enzyme-Linked Immunosorbent Assay (ELISA)
Mouse ELISA Kits from RUIXIN BIOTECH (RX203063M and RX202412M) were utilized to measure IL-1β and TNF-α levels. The procedure involved measuring the optical density (OD) of plasma samples or MH-S supernatants at a wavelength of 450 nm. IL-1β and TNF-α levels were then determined through comparing these OD values to the respective standard curves.
Received Operating Characteristic (ROC) Curve
The R package pROC was utilized to plot ROC curve for miR-17-5p in plasma-EVs. The diagnostic performance of miR-17-5p as a biomarker was assessed by calculating area under the curve (AUC).
Flow Cytometry Analysis for Macrophage Polarization
Flow cytometry was performed to evaluate macrophage polarization in MH-S cells and lung tissue samples. M1-type macrophages were identified as F4/80⁺CD86⁺, and M2-type macrophages as F4/80⁺CD206⁺. Single-cell suspensions were incubated with the following fluorochrome-conjugated monoclonal antibodies: anti-mouse F4/80-APC (clone BM8, BioLegend, Cat# 123116), anti-mouse CD86-PE (clone GL1, BioLegend, Cat# 105008), anti-mouse CD80-PE (clone 16–10A1, Thermo Fisher, Cat# 12–0801-82), and anti-mouse CD206-FITC (clone C068C2, BioLegend, Cat# 141704). After staining for 30 minutes at 4 °C in the dark, cells were washed and resuspended in PBS containing 2% FBS. Data were acquired using a BD Celesta™ flow cytometer (BD Biosciences, USA), and analysis was conducted using FlowJo software (version 10.0, Tree Star Inc., USA).
Hepatic and Kidney Function Analysis
Mouse blood samples were collected and centrifuged, plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST), blood urea nitrogen (BUN) and serum creatinine (Scr) were monitored using biochemical kits per manufacturer’s instructions.
Histological Evaluation
For histopathological examination, lung specimens were removed and immersed in 4% paraformaldehyde solution for a period of 24 h. Subsequently, tissues were fixed in paraffin, sectioned to a thickness of 4–6 mm, and subjected to hematoxylin and eosin (H&E) staining. Lung injury in this study was evaluated using a scoring system that ranged from 0, signifying no damage, to 4, indicating the most severe damage. The scoring took into account several factors including neutrophil infiltration, hemorrhage presence, alveolar congestion, and alveolar wall damage. The overall lung injury score was derived by aggregating the scores for each of these factors.29,30
RNA Sequencing and Identification of Related Differentially Expressed Genes (DEGs)
RNA was isolated from MH-S using TRIzol in conjunction with DNase I. The integrity and purity of RNA were evaluated via electrophoresis on a 1% agarose gel. RNA integrity was further assessed with a 2100 Bioanalyzer (Agilent Technologies, USA), and quantified with a NanoDrop ND-2000 spectrophotometer (Nanodrop Technologies, USA). RNA samples were chosen for sequencing based on their high quality, which included an OD260/280 ratio ranging from 1.8 to 2.2, an OD260/230 ratio no less than 2.0, an RNA Integrity Number of at least 8.0, and a 28S:18S ratio of at least 1.0. These RNA samples underwent purification, reverse transcription, and the construction of cDNA libraries, which were then sequenced. RNA sequencing data from this study are accessible in online databases. R language package “limma (v 3.48.3)” was utilized to identify DEGs, using an adjusted P ≤ 0.05 and |log2FC| ≥ 2 as criteria for selection. A heatmap was generated to visualize DEGs. Additionally, R package ClusterProfiler was utilized for the functional annotation of DEGs through Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, and to map out the significantly enriched pathways.
Western Blot
Western blot was performed to assess Bcl11b, IL-1β, IL-6, TGF-β, YM1, and CCL24 levels. Protein extracts from both cells and lung tissue were quantified using BCA kit (P0012, Beyotime) to determine protein concentration. Each sample, containing 20 μg of protein, was combined with loading buffer, heated at 100°C for a duration of 5 min to denature, and subsequently resolved through SDS-PAGE. The membranes were incubated with primary antibodies as follows: Bcl11b (55414-1-AP, 1:1000, Proteintech), IL-1β (A1112, 1:1000, ABCLONAL), IL-6 (21,865-1-AP, 1:1000, Proteintech), TGF-β (19,999-1-AP, 1:1000, Proteintech), YM1 (ab192029, 1:1000, Abcam), CCL24 (22,306-1-AP, 1:1000, Proteintech), and GAPDH (60004-1-Ig, 1: 50000, Proteintech). Following an overnight incubation at 4°C, membranes were incubated with secondary antibodies goat anti-mouse IgG (BL001A, 1:50000, Biosharp) or goat anti-rabbit IgG (BL003A, 1:50000, Biosharp) at ambient temperature for 2 h. Chemiluminescent signals were captured using an automated imaging system (OI-X6, Guangzhou, China) with enhanced chemiluminescence reagents from Millipore, USA. The OD of each protein band was normalized against the corresponding GAPDH band for comparison.
Dual-Luciferase Reporter Assay
Firstly, bioinformatics was conducted to predict binding sites of miR-17-5p with 3’untranslated region (UTR) of Bcl11b. Then, a dual-luciferase assay was employed to validate interaction between miR-17-5p and 3’UTR of Bcl11b. To pinpoint exact binding site, 3’UTR of Bcl11b, with either the wild-type (WT) or mutant (MUT) sequence of predicted interaction site, was synthesized and inserted into pmirGLO vector, a dual-luciferase reporter system from Promega, USA. 293T cell line was employed for these assays. Using Luc-Pair™ Duo-Luciferase Assay Kit 2.0 (LF001, GeneCopoeia, USA), 293T cells were seeded in 24-well plates at a concentration of 5×104 cells per well and, when they reached 60–70% confluence, were transfected with miR-17-5p mimics or NC at a final concentration of 20 pmol for a period of 24 h. Subsequently, the cells were co-transfected with either the WT or MUT Bcl11b 3’UTR construct and a Renilla luciferase-expressing control vector using Lipofectamine Transfection Reagent (11668019, Life Technologies) for an additional 48 h. Cells were collected utilizing 300 μL of Passive Lysis Buffer, and luciferase activities were quantified using microplate reader.31 The firefly luciferase activity was standardized against Renilla luciferase using dual-luciferase assay system (Thermo, USA).
Statistical Analysis
Data analysis was conducted using SPSS 19 and GraphPad Prism 10.4.1. The mean ± standard deviation (SD) of the measurement data was calculated. The Shapiro–Wilk test was used to assess the normality of data distribution. For data following a normal distribution, Student’s t-test or one-way/two-way ANOVA was used. For non-normally distributed data, the Mann–Whitney U-test (for two groups) and Kruskal–Wallis test (for more than two groups) were applied. A P value < 0.05 was considered statistically significant.
Results
Exosomes from Different Sources Were Involved in LPS-Induced Macrophage Polarization
Blood is the most commonly used body fluid to obtain and study EVs.32 First, we collected peripheral blood from healthy individuals and sepsis patients and extracted exosomes. TEM results showed that Con-plasma-exosomes and Sepsis-plasma-exosomes had a cup-shaped structure (Figure 1A). NTA detection revealed that the particle sizes ranged between 100–200 nm (Figure 1B), and flow cytometry detection showed positivity for exosomal markers CD63 and CD81 (Figure 1C), confirming successful exosome isolation. Next, alveolar macrophages (MH-S) were treated with LPS to simulate sepsis, followed by incubation with 10 μg/well of exosomes derived from the peripheral blood of healthy individuals or sepsis patients for 24 h (Figure 1D). In contrast to LPS+N-EXO group, LPS+S-EXO group exhibited an upregulation in M1 marker iNOS and a downregulation in M2 marker Arg-1. There was a notable rise in pro-inflammatory cytokines TNF-α and IL-1β levels (Figure 1E and F). These results suggested that N-EXO treatment may suppress polarization of M1 macrophages and production of pro-inflammatory factors, while promoting polarization of M2 macrophages, thereby reducing overall inflammatory response. Our results indicated that exosomes from different sources were involved in LPS-induced macrophage polarization.
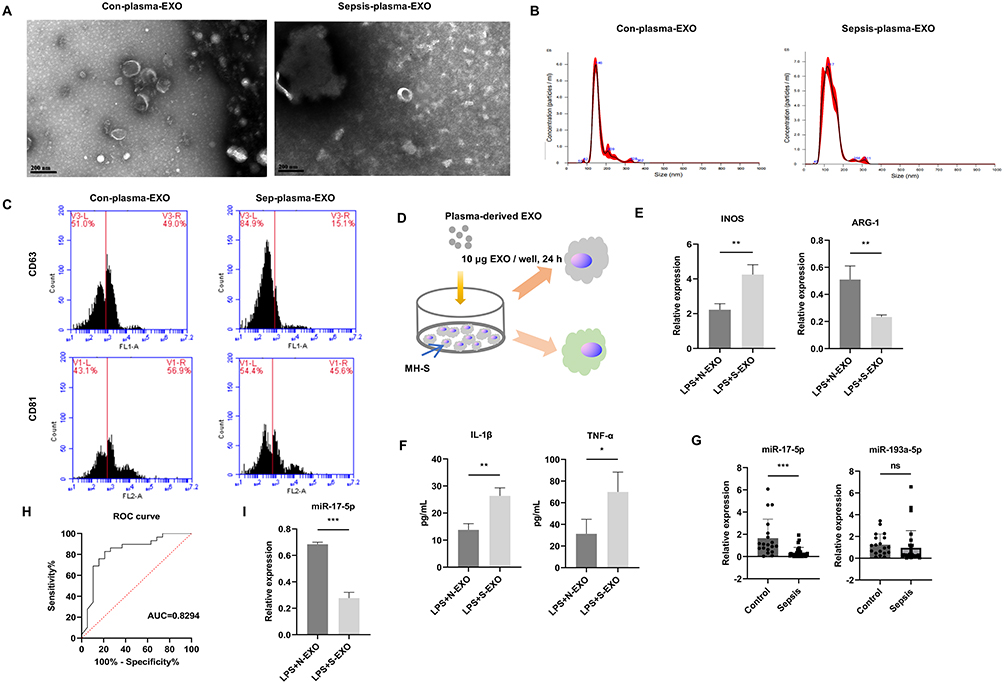 |
Figure 1 Exosomes from different sources were involved in LPS-induced macrophage polarization. (A) TEM images of plasma-EVs. Scale bars, 200 nm. (B) EVs particles were assessed using NTA. (C) Markers for EVs (CD63 and CD81) were quantified through flow cytometry. (D) MH-S cells were treated with 10 μg/well of plasma-derived exosomes from sepsis patients or healthy controls in combination with LPS (1 μg/μL) for 24 h. (E) Detection of Arg-1 and iNOS levels by qRT-PCR. (F) Detection of inflammatory cytokines (TNF-α and IL-1β) levels in supernatant of MH-S through ELISA. (G) Comparison of miR-17-5p and miR-193a-5p in plasma-EVs between control group (n = 19) and sepsis group (n = 27). (H) Diagnostic value of miR-17-5p of plasma-EVs using received operating characteristic (ROC) curve techniques. (I) Detection of miR-17-5p expression in macrophages via qRT-PCR. *P < 0.05, **P < 0.01, ***P < 0.001; ns, not significant. |
We previously found that miR-193a-5p and miR-17-5p were differentially expressed in exosomes from bronchoalveolar lavage fluid, and a series of experiments confirmed that miR-193a-5p and miR-17-5p could serve as diagnostic markers for pneumonia.22 Pneumonia is a major cause of sepsis.33 Therefore, after discovering that exosomes from different sources could induce polarization and inflammatory response changes in sepsis macrophage models, we detected miR-193a-5p and miR-17-5p levels in exosomes from peripheral blood of multiple healthy individuals and patients with sepsis. Compared to Con-plasma-exosomes group, miR-17-5p expression in Sepsis-plasma-exosomes group was significantly reduced, while miR-193a-5p showed no significant difference (Figure 1G). Given its lack of statistical significance, miR-193a-5p was not pursued for further functional analysis. The construction of ROC curve revealed that the AUC value for miR-17-5p was 0.8294, indicating that it may serve as a potential biomarker for early diagnosis and therapeutic monitoring of sepsis (Figure 1H). Therefore, we chose miR-17-5p to study further. Additionally, we detected miR-17-5p levels in macrophages treated with exosomes from peripheral blood of healthy individuals and patients with sepsis. Compared to the LPS+N-EXO group, miR-17-5p levels in LPS+S-EXO group was significantly reduced (Figure 1I). To further explore how miR-17-5p expression varied among different clinical backgrounds, we performed subgroup analyses based on age, sex, and hypertension, and diabetes. As shown in Supplementary Table S2, exosomal miR-17-5p levels were lower in sepsis patients across both age groups (<65 years: P = 0.005; ≥65 years: P = 0.009) and in both males (P = 0.043) and females (P = 0.003). Among patients with hypertension, miR-17-5p was also markedly reduced compared to controls (P = 0.004). In the diabetes subgroup, due to limited cases, no significant difference was observed (P = 0.117). Interaction analysis showed no significant subgroup effects (P > 0.05), suggesting consistent downregulation of miR-17-5p in sepsis.
MiR-17-5p Was Involved in LPS-Induced Macrophage Polarization
Studies have shown that exosomes can carry miRNAs for long-distance targeted transport, affecting the function of recipient cells.34,35 To investigate whether miR-17-5p modulates macrophage polarization, we overexpressed miR-17-5p in MH-S cells and examined its impact on inflammatory activation. Transfection efficiency was first validated by qRT-PCR, showing significantly elevated miR-17-5p levels in the mimic group compared to the negative control (Figure 2A). LPS stimulation alone markedly decreased endogenous miR-17-5p expression, which was successfully restored upon mimic transfection (Figure 2B). Functionally, LPS induced a pronounced increase in the secretion of IL-1β and TNF-α, consistent with a pro-inflammatory response; these elevations were significantly attenuated by miR-17-5p overexpression (Figure 2C). Expression of the M1-associated marker iNOS was upregulated following LPS exposure, while the M2 marker ARG-1 was suppressed—both of which were reversed upon miR-17-5p transfection, suggesting a phenotypic shift away from M1 polarization (Figure 2D). Flow cytometry further demonstrated that LPS promoted an increase in the proportion of F4/80⁺CD86⁺ (M1-type) macrophages and a decrease in F4/80⁺CD206⁺ (M2-type) cells. These trends were significantly corrected by miR-17-5p, which reduced CD86⁺ and elevated CD206⁺ populations (Figure 2E). Together, these findings indicate that miR-17-5p inhibits LPS-induced M1 polarization and inflammatory cytokine production while enhancing M2-type differentiation in macrophages.
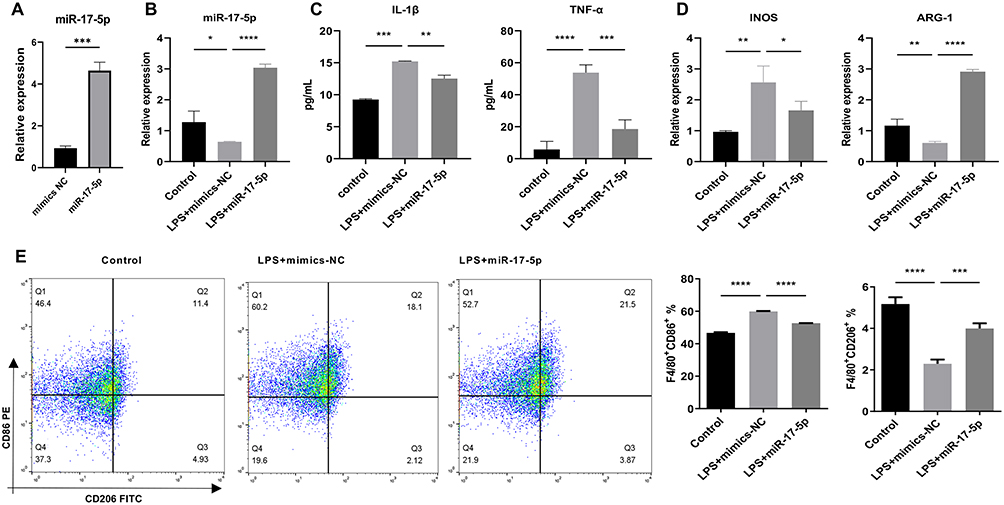 |
Figure 2 MiR-17-5p was involved in LPS-induced macrophage polarization. (A) qRT-PCR was used to confirm the efficiency of miR-17-5p overexpression after mimic transfection. (B) qRT-PCR was utilized to assess miR-17-5p levels. (C) ELISA was employed to measure TNF-α and IL-1β levels in cell supernatant. (D) qRT-PCR was utilized to assess iNOS, and Arg-1 levels. (E) Flow cytometry was utilized to assess the polarization of M1-type macrophages (F4/80+CD86+) and M2-type macrophages (F4/80+CD206+). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
MiR-17-5p Regulated Macrophage Polarization Involved in Sepsis-Induced Organ Damage
Next, we intervened miR-17-5p expression at animal level by tail vein injecting miR-17-5p agomir into CLP mice to observe the phenotypic changes in sepsis. The experimental endpoint showed a survival rate of 47% in the CLP group, which increased to 60% following administration of miR-17-5p agomir (Figure 3A). Liver and kidney function tests revealed that, compared to Sham group, ALT, AST, BUN, and Scr levels all increased in CLP group. After treatment with miR-17-5p agomir, ALT, AST, BUN, and Scr levels significantly decreased (Figure 3B). Additionally, compared to Sham group, TNF-α and IL-1β levels were elevated in CLP group. Treatment with miR-17-5p agomir significantly reduced TNF-α and IL-1β levels induced by CLP (Figure 3C). H&E staining revealed that the lungs of CLP group showed obvious alveolar disruption and damage, extensive inflammatory cell infiltration, and thickened alveolar septa. Following administration of miR-17-5p agomir, the degree of inflammatory cell infiltration was reduced, and alveolar septa became thinner (Figure 3D). Furthermore, compared to Sham group, proportion of M1-type macrophages M1 (F4/80+CD86+) was increased in CLP group, M1 polarization marker iNOS levels were increased, proportion of M2-type macrophages (F4/80+CD206+) was decreased, M2 polarization marker Arg-1 levels were decreased, and miR-17-5p expression was reduced. After treatment with miR-17-5p agomir, proportion of F4/80+CD86+ was decreased, M1 polarization marker iNOS levels were decreased, proportion of F4/80+CD206+ was elevated, M2 polarization marker Arg-1 levels were elevated, and miR-17-5p expression was elevated (Figure 3E and F). Our results indicated miR-17-5p regulated macrophage polarization and was involved in sepsis-induced organ damage.
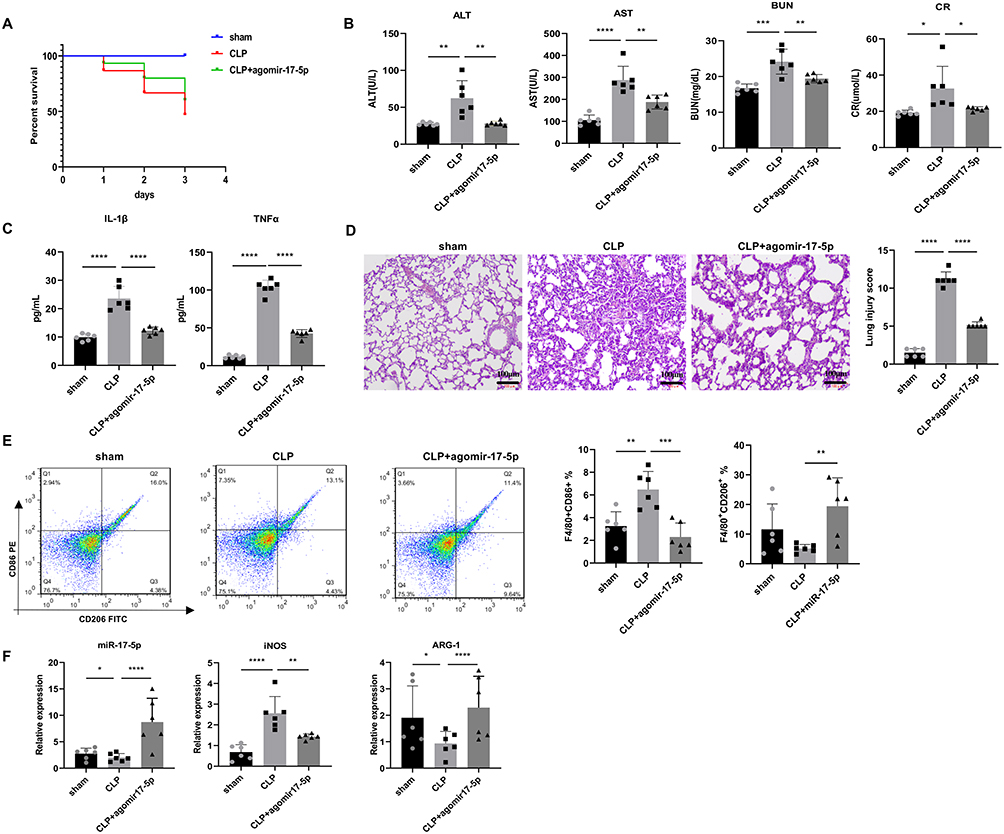 |
Figure 3 MiR-17-5p regulated macrophage polarization involved in sepsis-induced organ damage. (A) The survival rate of CLP mice with or without miR-17-5p agomir (n = 15). (B) ALT, AST, BUN, and Scr levels in the serum of septic mice (n = 6). (C) Serum cytokines (TNF-α and IL-1β) levels evaluation by ELISA. (D) H&E staining of CLP-induced lung damages for septic mice (n = 6). Scale bars, 100 μm. (E) Representative flow cytometry plots and quantification of CD86⁺ (M1) and CD206⁺ (M2) macrophages from lung single-cell suspensions (n = 6 mice per group). (F) qRT-PCR was employed to assess miR-17-5p, iNOS, and Arg-1 levels in lung tissue. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
Transcriptome Sequencing Analysis of the Mechanism of miR-17-5p Regulation of Macrophage Polarization
Next, we explored the mechanism by which miR-17-5p regulated macrophage polarization through transcriptome sequencing, in order to screen for targets regulated through miR-17-5p and to clarify pathways modulated via miR-17-5p. Figure 4A showed a heatmap of DEGs. KEGG enrichment analysis revealed DEGs were primarily enriched in Cytokine-cytokine receptor interaction, Systemic lupus erythematosus, Rheumatoid arthritis, and Osteoclast differentiation (Figure 4B). GO analysis indicated DEGs were mainly enriched in biological processes (BP) of cellular processes, biological regulation, and response to stimulus, cellular components (CC) of cell, cell part, and membrane and molecular functions (MF) of binding, catalytic activity, and molecular transducer activity (Figure 4C). By taking the intersection of genes from Control VS LPS+mimic-NC and LPS+mimic-NC VS LPS+mimic-miR-17-5p, we selected genes with high fold changes and those that have binding sites with miR-17-5p, including Serpina1c, Zfyve9, Gm20708, Cebpb, Cxcr3, Aak1, Kcanb1, Bcl11b, and Bal6. qRT-PCR validation revealed compared to Control group, Serpina1c, Zfyve9, Gm20708, Cebpb, Cxcr3, Aak1, Kcanb1, Bcl11b, and Bal6 levels were up-regulated in LPS+mimic-NC group. After overexpressing miR-17-5p, Serpina1c, Zfyve9, Gm20708, Cxcr3, Aak1, Kcanb1, Bcl11b, and Bal6 levels were down-regulated, while Cebpb levels were up-regulated (Figure 4D). Their expression trends were consistent with sequencing-based read counts (Figure 4E). Western blot further confirmed that compared to Control group, Bcl11b levels were up-regulated in LPS+mimic-NC group. After overexpressing miR-17-5p, Bcl11b levels were down-regulated (Figure 4F). Ultimately, Bcl11b was identified as a potential target that may bind to miR-17-5p and participate in macrophage polarization for further experimental investigation.
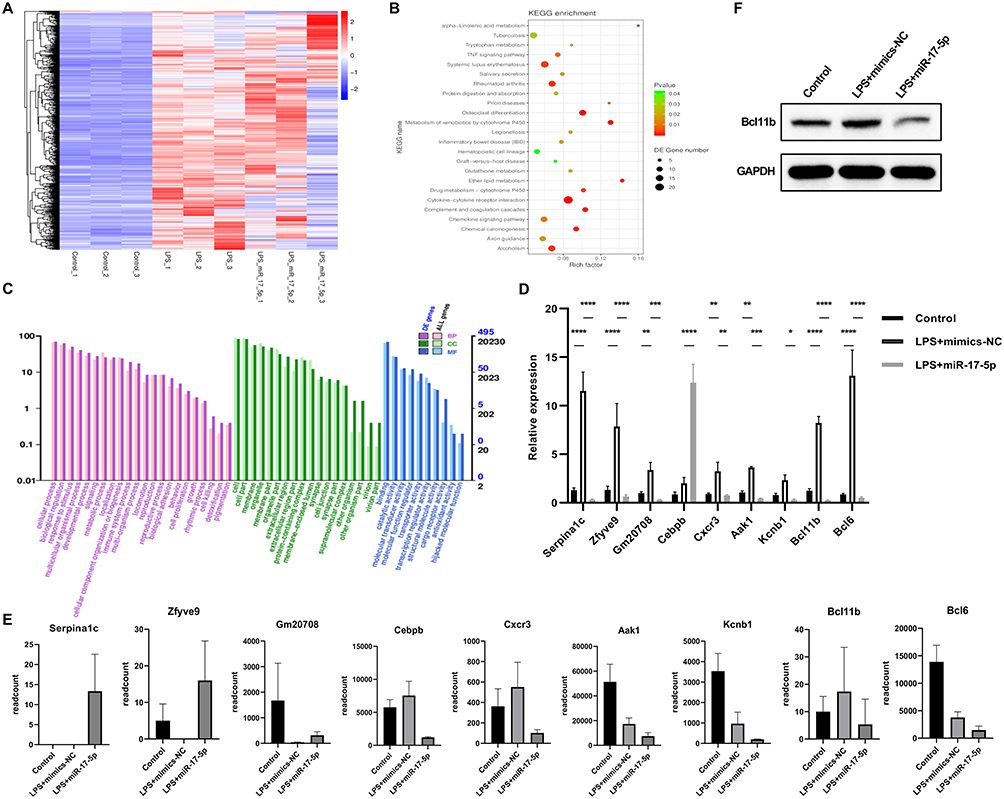 |
Figure 4 Transcriptome sequencing analysis of the mechanism of miR-17-5p regulation of macrophage polarization. (A) Heatmap of DEGs. (B) KEGG enrichment analysis of DEGs. (C) GO enrichment analysis of DEGs. (D) The expression of DEGs (Serpina1c, Zfyve9, Gm20708, Cebpb, Cxcr3, Aak1, Kcanb1, Bcl11b, and Bal6) was validated by qRT-PCR. (E) RNA-seq–based read counts for corresponding DEGs, indicating consistent expression trends with qRT-PCR results. (F) Western blot was employed to measure changes in Bcl11b under miR-17-5p involvement in LPS-induced macrophage polarization. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
MiR-17-5p Targeted Bcl11b to Regulate Macrophage Polarization and Participate in Sepsis Injury
Finally, we investigated the impact of miR-17-5p and Bcl11b in mediating macrophage polarization and their mechanisms in sepsis-induced injury at the cellular level. We performed overexpression of Bcl11b. Compared to NC group, Bcl11b levels in Bcl11b-OE group significantly increased (Figure 5A), indicating successful transfection of Bcl11b-OE. Furthermore, compared to NC group, pro-inflammatory factors IL-1β and TNF-α levels were elevated in the LPS+NC group (Figure 5B), M1 polarization marker iNOS levels were up-regulated, while M2 polarization marker Arg-1 levels decreased (Figure 5C), and M1-type marker genes (IL-6 and IL-1β) levels were elevated, while M2-type marker genes (TGF-β, YM1, and CCL24) levels were down-regulated (Figure S1B and S1C), proportion of M1-type macrophages M1 (F4/80+CD86+) was increased, and proportion of M2-type macrophages (F4/80+CD206+) was repressed (Figure 5D). Under the influence of miR-17-5p mimics, pro-inflammatory factors IL-1β and TNF-α levels decreased (Figure 5B), M1 polarization marker iNOS levels decreased, while M2 polarization marker Arg-1 levels increased (Figure 5C), and M1-type marker genes (IL-1β and IL-6) levels decreased, while M2-type marker genes (TGF-β, YM1, and CCL24) levels increased (Figure S1B and S1C), and proportion of F4/80+CD86+ decreased, while proportion of F4/80+CD206+ increased (Figure 5D). Overexpression of Bcl11b could reverse the effects of miR-17-5p mimics (Figures 5B-D, S1B and S1C), suggesting that Bcl11b may play a crucial regulatory role in macrophage polarization, affecting regulatory effects of miR-17-5p on macrophage polarization, and there may be a regulatory relationship between the two. Furthermore, through bioinformatics analysis, we found that miR-17-5p had binding sites with 3’UTR of Bcl11b (Figure 5E). Co-transfection of the WT and MUT constructs of Bcl11b-3’UTR with miR-17-5p mimics and subsequent dual-luciferase assay showed miR-17-5p had no significant regulatory effect on Bcl11b-mut-1, but had a significant inhibitory effect on Bcl11b-WT-1 (Figure 5F), indicating that Bcl11b-WT-1 was a target sequence of miR-17-5p. Although multiple binding sites were predicted, we selected the most conserved and strongly predicted site for validation, which was functionally confirmed in our assay. Our results demonstrated miR-17-5p targeted Bcl11b to regulate macrophage polarization and participate in sepsis-induced injury. In summary, our results suggested that sepsis plasma-derived exosomal miR-17-5p inhibited M1 macrophage polarization in sepsis-associated ALI by suppressing its target gene Bcl11b (Figure 6).
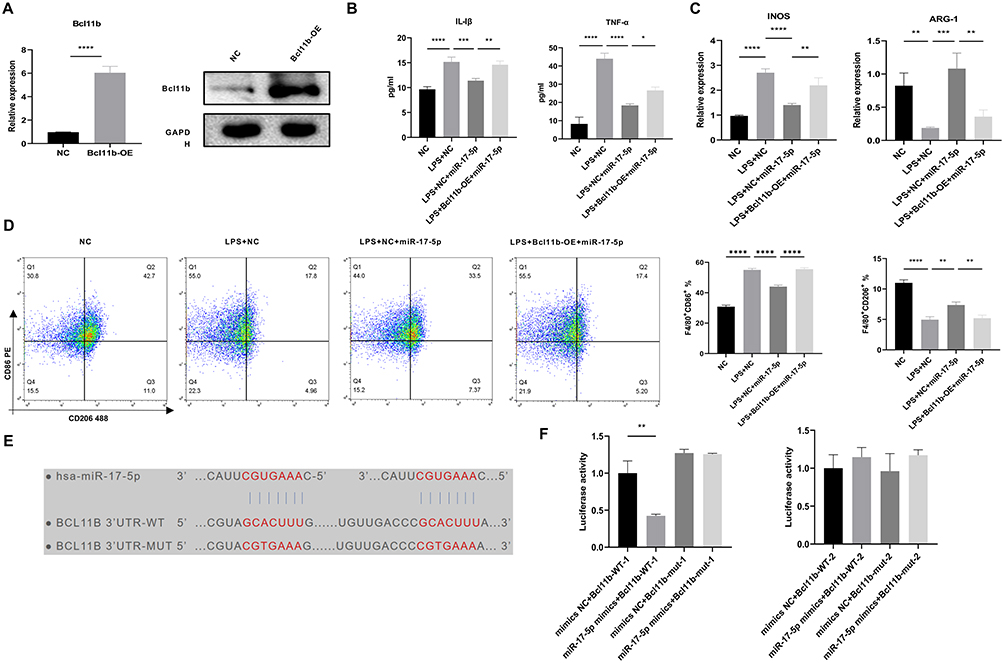 |
Figure 5 MiR-17-5p targeted Bcl11b to regulate macrophage polarization and participate in sepsis injury. (A) qRT-PCR and Western blot were utilized to assess overexpression effect of Bcl11b. (B) ELISA was conducted to measure TNF-α and IL-1β levels in cell supernatant. (C) qRT-PCR was employed to measure M1 polarization marker iNOS and the M2 polarization marker Arg-1 levels. (D) Flow cytometry was utilized to assess polarization of M1-type macrophages (F4/80+CD86+) and M2-type macrophages (F4/80+CD206+). (E) Prediction of the binding sites of miR-17-5p with the 3’UTR of Bcl11b. (F) Dual-luciferase assay was utilized to validate interaction between miR-17-5p and 3’UTR of Bcl11b. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
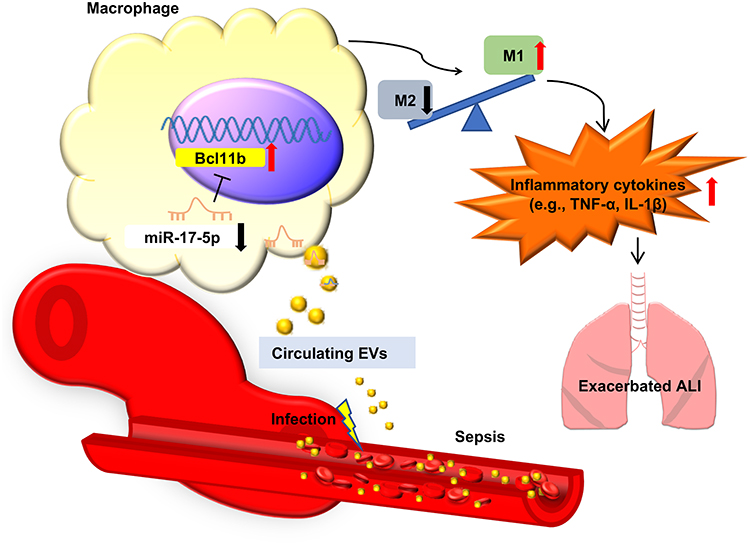 |
Figure 6 Schematic diagram illustrating the proposed mechanism by which plasma-derived exosomal miR-17-5p regulates macrophage polarization during sepsis-induced ALI. Infection triggers the release of EVs into circulation, where miR-17-5p levels are reduced. The downregulation of miR-17-5p leads to the upregulation of its target gene BCL11B in macrophages, which promotes M1 polarization and enhances the production of pro-inflammatory cytokines such as TNF-α and IL-1β. This process contributes to aggravated pulmonary inflammation and worsened ALI. |
Discussion
Sepsis is characterized by a systemic inflammatory response triggered by an infection.36 A range of bioactive substances released into blood during endotoxin stimulation contribute to the initiation and escalation of sepsis.37 Given that pneumonia is one of the most common causes of sepsis and shares overlapping inflammatory pathways, it is biologically plausible to explore whether pneumonia-associated miRNAs also play a role in the progression of sepsis. Our research identified that the miR-17-5p/Bcl11b axis modulated MH-S polarization, suggesting its potential as a therapeutic target for treating airway inflammation and remodeling associated with sepsis-induced ALI. This is also the first time to report the study of miR-17-5p/Bcl11b and macrophage polarization in sepsis-induced lung injury.
EVs are a heterogeneous population of spherical membrane structures released by virtually all types of cells, encompassing a variety of cellular components. EVs activities modulating macrophage- and tissue-mediated injury and repair responses.38 MiRNAs from EVs of different origins participate in the modulation of biological functions of pulmonary epithelial cells, endothelial cells, and phagocytes during sepsis-associated ALI through EV-mediated miRNA transfer, which holds significant potential for diagnosis and therapy.39 A previous investigation has indicated that certain exosomal miRNAs in sepsis patients are predictive of the condition, with miR-26b-5p and miR-125b-5p being specifically modulated in exosomes and serum.40 Additionally, exosomal miRNAs that are down-regulated, such as hsa-let-7f-5p, miR-331-3p, miR-301a-3p, and miR-335-5p, have been identified as independent prognostic markers for in-hospital and 90-day mortality rates among sepsis patients.41 Nevertheless, there is a scarcity of research on the involvement of peripheral circulating serum EVs in sepsis-related ALI.42 In the present study, a reduction in the levels of plasma-derived exosomal miR-17-5p has been observed in sepsis patients as opposed to those without infection, indicating a potential functional role for miR-17-5p in sepsis-associated ALI. The observed downregulation of miR-17-5p in sepsis may be attributed to the complex immune dysregulation that characterizes the condition. Sepsis is often accompanied by overwhelming inflammation, oxidative stress, and elevated levels of cytokines such as TNF-α and IL-6, all of which can disrupt the cellular machinery responsible for miRNA synthesis, maturation, or sorting into exosomes.43–45 These pathological factors may act in concert to reduce the packaging efficiency or stability of miR-17-5p in circulating extracellular vesicles. Additionally, we discovered that EVs derived from the plasma of sepsis patients could effectively drive M1 polarization of macrophages, as indicated by decreased expression of Arg1 and the increased expression of iNOS. The overexpression or dysregulation of iNOS is associated with numerous diseases, including sepsis.46 The inflammatory response characterized by IL-1β, TNF-α, and IL-6 is intimately associated with M1 phenotype of macrophages.47 M1-polarized macrophages are known to initiate sustained inflammatory reactions, releasing a variety of cytokines, including IL-1β, TNF-α, and IL-6, as well as chemokines, which contribute to chronic inflammation.48 This aligns with our observations of increased IL-1β and TNF-α in MH-S treated with peripheral blood EVs from sepsis patients. These findings suggested miR-17-5p could be a potential diagnostic marker and plays a crucial regulatory role in sepsis-associated ALI.
MiRNAs are known to regulate a variety of cellular processes, including the polarization of macrophages.49 Research has demonstrated that exosome-mediated miR-93-TXNIP-NLRP3 apoptosis contributes to the functional differences between M1 and M2 macrophages in sepsis-induced acute kidney injury.50 Roquin-1 within macrophages can enhance the differentiation of Tregs and reduce M1 macrophage polarization through miRNAs in macrophage-derived EVs.51 Moreover, in sepsis, upregulation of miR-17-5p can suppress inflammatory cytokines (IL-1β and TNF-α) levels induced by LPS in macrophages.24 In our research, an in vitro experiment with MH-S transfected with a miR-17-5p mimic demonstrated elevated miR-17-5p levels notably inhibited the M1 polarization of MH-S. In vivo studies, the administration of miR-17-5p agomir markedly mitigated production of inflammatory cytokines and chemokines, and altered macrophage polarization. Consistently, the enhanced expression of miR-17-5p was related to reduced sepsis severity in a mouse model, marked by lower TNF-α levels, decreased hepatic and renal damage, and reduced mortality. In sepsis-induced organ dysfunction, the lungs are among the earliest organs to be impacted.52 Our study demonstrated a decrease in lung injury scores and a reduction in CD86⁺ M1 macrophages, which served as biomarkers for the protective effects of miR-17-5p in mice subjected to CLP-induced sepsis. The inflammatory cytokines secretion and upregulation of iNOS are associated with M1 macrophages, while the increase in Arg1 is linked to M2 macrophages,38 which supports our observations.
MiR-17-5p plays a role in cancer development through targeting genes involved in tumor suppression and apoptosis throughout the progression of the disease. The concurrent decrease in miR-17-5p and increase in Bcl11b suggests a potential regulatory function of miR-17-5p on Bcl11b in T-cell acute lymphoblastic leukemia.53 Bcl11b, also referred to as CTIP2, plays a role in various biological processes including cell division,54 apoptosis,55 and immune reactions,56 in several pathological states, including inflammation,57 heart-related conditions,58 and the dormant phase of HIV infection.59 In HIV-1-infected human macrophages and microglial cells, reports showed that Bcl11b abrogates cell cycle arrest.60 Furthermore, Bcl11b plays an essential part in the generation and secretion of cytokines such as IL-6, IFN-γ, and TNF-α.56,61,62 In the context of ALI, macrophages and the cytokines they secrete are pivotal in modulating the condition. In our research, we discovered an inverse relationship between Bcl11b expression and miR-17-5p levels in macrophages activated by LPS. RNA sequencing analysis revealed possible target sites for miR-17-5p within the Bcl11b transcript. Our results indicated miR-17-5p may function as a modulator of Bcl11b, with upregulated expression resulting in reduced Bcl11b levels. Consequently, this downregulation of Bcl11b impacts cytokine and chemokine release, as well as the polarization state of macrophages. Additionally, the expression of TGF-β, YM1, and CCL24 was found to be inversely correlated with Bcl11b levels, suggesting that the suppression of Bcl11b regulates polarization signaling in sepsis. Our study has uncovered a distinct role for Bcl11b, which deviates from its recognized function as a tumor suppressor55 and its involvement in immune response induction.63 We have discovered new mechanisms involving miR-17-5p and a novel regulatory function for Bcl11b, where increased expression of miR-17-5p leads to a reduction in Bcl11b levels, influencing cytokine and chemokine release as well as macrophage polarization.
Based on the current body of research, our study is among the first to establish a mechanistic link between the elevation of miR-17-5p and the progression of sepsis-related ALI, mediated through its regulation of Bcl11b and the resulting impact on macrophage polarization. We found that increased expression of miR-17-5p exerts a protective effect by downregulating Bcl11b, thereby dampening the inflammatory response and promoting M2 macrophage polarization. These results suggest that miR-17-5p may serve as a potential biomarker for sepsis-associated ALI and offer a theoretical basis for the development of targeted therapeutic strategies.
Several limitations of this study should be acknowledged. First, the patient samples were derived from a single medical center, and the relatively limited cohort size may affect the generalizability of the findings. Although plasma exosomes were isolated using a standardized protocol, donor-to-donor variability cannot be completely ruled out. To mitigate this, we selected individuals based on consistent clinical criteria and confirmed vesicle quality across samples; however, pooled sampling and validation in larger cohorts will be necessary in future investigations. Second, although we employed both qRT-PCR and flow cytometry to evaluate macrophage polarization, additional validation using protein-level techniques such as Western blotting or immunofluorescence, alongside expanded marker panels including GBP5, CD80, and TGF-β, would further strengthen the robustness of our conclusions. While flow cytometry provided insights into macrophage phenotypes, spatial localization within lung tissue remains unexplored. Immunohistochemical staining for markers such as CD68 or F4/80 could provide anatomical context and will be pursued in subsequent studies. Third, the cross-species application of human plasma-derived exosomes to murine macrophage models may introduce biological variability. Although many miRNA sequences and their regulatory targets are conserved across species, potential differences in exosome uptake and intracellular processing between humans and mice could influence the observed outcomes. This methodological limitation should be considered when interpreting the translational relevance of our findings. Fourth, while our data suggest that exosomal miR-17-5p regulates macrophage polarization, we lack direct evidence confirming cell-specific delivery of these exosomes in vivo. Furthermore, although multiple putative binding sites for miR-17-5p on the Bcl11b transcript were computationally predicted, only the most conserved site was experimentally validated in this study. The contribution of other potential sites remains to be elucidated. Additionally, the functional role of Bcl11b in vivo was not directly examined in the current study, leaving open questions regarding its precise mechanistic involvement in sepsis-induced lung injury. Finally, the exclusive use of young adult male mice may not fully capture the biological complexity of sepsis. Age-related differences in immune function and disease progression are well established, and whether the miR-17-5p–Bcl11b axis functions similarly in aged or female animals remains unknown. Future studies incorporating aged models, female mice, and multi-center clinical cohorts will be essential to improve the rigor and translational potential of our findings.
Conclusion
This study identifies a novel regulatory axis involving plasma exosomal miR-17-5p and Bcl11b that modulates macrophage polarization during sepsis-induced lung injury. Our findings demonstrate that miR-17-5p is reduced in exosomes derived from septic patients and promotes M1 macrophage polarization by relieving the suppression of Bcl11b. These results suggest that miR-17-5p holds potential as a diagnostic marker for sepsis and as a mechanistic target in regulating inflammation. However, considering the modest sample size and the single-center design, these conclusions should be interpreted with appropriate caution. Further studies involving larger and more diverse patient populations, along with in-depth mechanistic exploration, will be important to establish the broader applicability of this regulatory pathway.
Data Sharing Statement
The data generated in the present study are included in the figures and tables of this article.
Ethics Approval Statements and Informed Consent
All participants provided their written consent after being fully informed about the study. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. Ethical approval was obtained from the Ethics Committee of The Third Affiliated Hospital of Sun Yat-sen University (No. [2022]02-216-01).
All animal procedures for the study were conducted in compliance with protocol approved by the Institutional Animal Care and Use Committee at the Medical College of Sun Yat-sen University (Approval Number SYSU-IACUC-2022-000786). Animal experiments were performed in accordance with ARRIVE guidelines and the IACUC Handbook (third edition).
Author Contributions
Ying Xian and Yinfang Sun contributed equally to this work. Rui Chen and Kouxing Zhang are co-corresponding authors.
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
No funding was received.
Disclosure
The authors have no conflicts of interest to declare.
References
1. Saxena J, Das S, Kumar A, et al. Biomarkers in sepsis. Clin Chim Acta. 2024;562:119891. doi:10.1016/j.cca.2024.119891
2. Lin Y, Liu L, Lin Y, et al. MiR-145 alleviates sepsis-induced inflammatory responses and organ injury by targeting ADAM17. Front Biosci. 2024;29(1):44. doi:10.31083/j.fbl2901044
3. Pei Z, Yao W, Wang S, Wu Y. Regulation of sirtuins in sepsis-induced myocardial damage: the underlying mechanisms for cardioprotection. Front Biosci. 2024;29(2):54. doi:10.31083/j.fbl2902054
4. Qiao X, Yin J, Zheng Z, Li L, Feng X. Endothelial cell dynamics in sepsis-induced acute lung injury and acute respiratory distress syndrome: pathogenesis and therapeutic implications. Cell Commun Signal. 2024;22(1):241. doi:10.1186/s12964-024-01620-y
5. Xu Y, Xin J, Sun Y, et al. Mechanisms of sepsis-induced acute lung injury and advancements of natural small molecules in its treatment. Pharmaceuticals. 2024;17(4):472. doi:10.3390/ph17040472
6. Mohsin M, Tabassum G, Ahmad S, Ali S, Ali Syed M. The role of mitophagy in pulmonary sepsis. Mitochondrion. 2021;59:63–75. doi:10.1016/j.mito.2021.04.009
7. Cajander S, Kox M, Scicluna BP, et al. Profiling the dysregulated immune response in sepsis: overcoming challenges to achieve the goal of precision medicine. Lancet Respir Med. 2024;12(4):305–322. doi:10.1016/s2213-2600(23)00330-2
8. Prescott HC, Angus DC. Enhancing recovery from sepsis: a review. JAMA. 2018;319(1):62–75. doi:10.1001/jama.2017.17687
9. Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization. Eur J Pharmacol. 2020;877:173090. doi:10.1016/j.ejphar.2020.173090
10. Davis FM, Schaller MA, Dendekker A, et al. Sepsis induces prolonged epigenetic modifications in bone marrow and peripheral macrophages impairing inflammation and wound healing. Arteriosclerosis Thrombosis Vasc Biol. 2019;39(11):2353–2366. doi:10.1161/atvbaha.119.312754
11. Wang Z, Wang Z. The role of macrophages polarization in sepsis-induced acute lung injury. Front Immunol. 2023;14:1209438. doi:10.3389/fimmu.2023.1209438
12. Wang L, Wang D, Zhang T, Ma Y, Tong X, Fan H. The role of immunometabolism in macrophage polarization and its impact on acute lung injury/acute respiratory distress syndrome. Front Immunol. 2023;14:1117548. doi:10.3389/fimmu.2023.1117548
13. Zhou S, Yang X, Mo K, Ning Z. Pyroptosis and polarization of macrophages in septic acute lung injury induced by lipopolysaccharide in mice. Immun Inflamm Dis. 2024;12(3):e1197. doi:10.1002/iid3.1197
14. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
15. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9(1):27. doi:10.1038/s41392-024-01735-1
16. Shi Q, Fijten RR, Spina D, et al. Altered gene expression profiles in the lungs of benzo[a]pyrene-exposed mice in the presence of lipopolysaccharide-induced pulmonary inflammation. Toxicol Appl Pharmacol. 2017;336:8–19. doi:10.1016/j.taap.2017.09.023
17. Park C, Lei Z, Li Y, et al. Extracellular vesicles in sepsis plasma mediate neuronal inflammation in the brain through miRNAs and innate immune signaling. J Neuroinflammation. 2024;21(1):252. doi:10.1186/s12974-024-03250-0
18. Yao M, Cui B, Zhang W, Ma W, Zhao G, Xing L. Exosomal miR-21 secreted by IL-1β-primed-mesenchymal stem cells induces macrophage M2 polarization and ameliorates sepsis. Life Sci. 2021;264:118658. doi:10.1016/j.lfs.2020.118658
19. Jiao Y, Zhang T, Zhang C, et al. Exosomal miR-30d-5p of neutrophils induces M1 macrophage polarization and primes macrophage pyroptosis in sepsis-related acute lung injury. Crit Care. 2021;25(1):356. doi:10.1186/s13054-021-03775-3
20. Zhang Z, Chen L, Xu P, Xing L, Hong Y, Chen P. Gene correlation network analysis to identify regulatory factors in sepsis. J Transl Med. 2020;18(1):381. doi:10.1186/s12967-020-02561-z
21. Acosta-Herrera M, Lorenzo-Diaz F, Pino-Yanes M, et al. Lung transcriptomics during protective ventilatory support in sepsis-induced acute lung injury. PLoS One. 2015;10(7):e0132296. doi:10.1371/journal.pone.0132296
22. Sun Y, Xian Y, Duan Z, et al. Diagnostic potential of microRNAs in extracellular vesicles derived from bronchoalveolar lavage fluid for pneumonia-a preliminary report. Cells. 2022;11(19):2961. doi:10.3390/cells11192961
23. Curcio R, Poli G, Fabi C, et al. Exosomal miR-17-5p, miR-146a-3p, and miR-223-3p correlate with radiologic sequelae in survivors of COVID-19-related acute respiratory distress syndrome. Int J Mol Sci. 2023;24(17):13037. doi:10.3390/ijms241713037
24. Ji ZR, Xue WL, Zhang L. Schisandrin B attenuates inflammation in LPS-induced sepsis through miR-17-5p downregulating TLR4. Inflammation. 2019;42(2):731–739. doi:10.1007/s10753-018-0931-3
25. Dong Y, Hu K, Zhang J, et al. ScRNA-seq of gastric cancer tissues reveals differences in the immune microenvironment of primary tumors and metastases. Oncogene. 2024;43(20):1549–1564. doi:10.1038/s41388-024-03012-5
26. Shankar-Hari M, Phillips GS, Levy ML, et al. Developing a new definition and assessing new clinical criteria for septic shock: for the third international consensus definitions for sepsis and septic shock (Sepsis-3). JAMA. 2016;315(8):775–787. doi:10.1001/jama.2016.0289
27. Crescitelli R, Lässer C, Szabó TG, et al. Distinct RNA profiles in subpopulations of extracellular vesicles: apoptotic bodies, microvesicles and exosomes. J Extracell Vesicles. 2013;2. doi:10.3402/jev.v2i0.20677
28. Siempos II, Lam HC, Ding Y, Choi ME, Choi AM, Ryter SW. Cecal ligation and puncture-induced sepsis as a model to study autophagy in mice. J Visualized Exp. 2014;(84):e51066. doi:10.3791/51066
29. Hirano Y, Aziz M, Yang WL, et al. Neutralization of osteopontin attenuates neutrophil migration in sepsis-induced acute lung injury. Crit Care. 2015;19(1):53. doi:10.1186/s13054-015-0782-3
30. Aziz M, Matsuda A, Yang WL, Jacob A, Wang P. Milk fat globule-epidermal growth factor-factor 8 attenuates neutrophil infiltration in acute lung injury via modulation of CXCR2. J Immunol. 2012;189(1):393–402. doi:10.4049/jimmunol.1200262
31. Zeng T, Wang X, Wang W, et al. Endothelial cell-derived small extracellular vesicles suppress cutaneous wound healing through regulating fibroblasts autophagy. Clin Sci. 2019;133(9). doi:10.1042/cs20190008
32. Nieuwland R, Siljander PR. A beginner’s guide to study extracellular vesicles in human blood plasma and serum. J Extracell Vesicles. 2024;13(1):e12400. doi:10.1002/jev2.12400
33. Guzzardella A, Motos A, Vallverdú J, Torres A. [Corticosteroids in sepsis and community-acquired pneumonia]. Kortikosteroide bei Sepsis und ambulant erworbener Pneumonie. Med Klin Intensivmed Notfmed. 2023;118(Suppl 2):86–92. doi:10.1007/s00063-023-01093-w
34. Fang Z, Zhang X, Huang H, Wu J. Exosome based miRNA delivery strategy for disease treatment. Chin. Chem. Lett. 2022;33(4):1693–1704. doi:10.1016/j.cclet.2021.11.050
35. Nail HM, Chiu -C-C, Leung C-H, Ahmed MMM, Wang H-MD. Exosomal miRNA-mediated intercellular communications and immunomodulatory effects in tumor microenvironments. J Biomed Sci. 2023;30(1):69. doi:10.1186/s12929-023-00964-w
36. Zheng Y, Gao Y, Zhu W, Bai XG, Qi J. Advances in molecular agents targeting toll-like receptor 4 signaling pathways for potential treatment of sepsis. Eur J Med Chem. 2024;268:116300. doi:10.1016/j.ejmech.2024.116300
37. Mera S, Tatulescu D, Cismaru C, et al. Multiplex cytokine profiling in patients with sepsis. APMIS. 2011;119(2):155–163. doi:10.1111/j.1600-0463.2010.02705.x
38. Hu Q, Lyon CJ, Fletcher JK, Tang W, Wan M, Hu TY. Extracellular vesicle activities regulating macrophage- and tissue-mediated injury and repair responses. Acta Pharm Sin B. 2021;11(6):1493–1512. doi:10.1016/j.apsb.2020.12.014
39. Xiong C, Huang X, Chen S, Li Y. Role of extracellular microRNAs in sepsis-induced acute lung injury. J Immunol Res. 2023;2023:5509652. doi:10.1155/2023/5509652
40. Reithmair M, Buschmann D, Märte M, et al. Cellular and extracellular miRNAs are blood-compartment-specific diagnostic targets in sepsis. J Cell Mol Med. 2017;21(10):2403–2411. doi:10.1111/jcmm.13162
41. Shin B, Lee JY, Im Y, et al. Prognostic implication of downregulated exosomal miRNAs in patients with sepsis: a cross-sectional study with bioinformatics analysis. J Inten Care. 2023;11(1):35. doi:10.1186/s40560-023-00683-2
42. Jiang K, Yang J, Guo S, Zhao G, Wu H, Deng G. Peripheral circulating exosome-mediated delivery of miR-155 as a novel mechanism for acute lung inflammation. Mol Ther. 2019;27(10):1758–1771. doi:10.1016/j.ymthe.2019.07.003
43. O’Connell RM, Rao DS, Baltimore D. Baltimore D. microRNA regulation of inflammatory responses. Annu Rev Immunol. 2012;30:295–312. doi:10.1146/annurev-immunol-020711-075013
44. Squadrito ML, Baer C, Burdet F, et al. Endogenous RNAs modulate microRNA sorting to exosomes and transfer to acceptor cells. Cell Rep. 2014;8(5):1432–1446. doi:10.1016/j.celrep.2014.07.035
45. Liu D, Huang SY, Sun JH, et al. Sepsis-induced immunosuppression: mechanisms, diagnosis and current treatment options. Mil Med Res. 2022;9(1):56. doi:10.1186/s40779-022-00422-y
46. Cinelli MA, Do HT, Miley GP, Silverman RB. Inducible nitric oxide synthase: regulation, structure, and inhibition. Med Res Rev. 2020;40(1):158–189. doi:10.1002/med.21599
47. Wang L, He C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front Immunol. 2022;13:967193. doi:10.3389/fimmu.2022.967193
48. Zhang Q, Sun W, Li T, Liu F. Polarization behavior of bone macrophage as well as associated osteoimmunity in glucocorticoid-induced osteonecrosis of the femoral head. J Inflamm Res. 2023;16:879–894. doi:10.2147/jir.S401968
49. Wang Q, Hong L, Chen M, et al. Targeting M2 macrophages alleviates airway inflammation and remodeling in asthmatic mice via miR-378a-3p/GRB2 pathway. Front Mol Biosci. 2021;8:717969. doi:10.3389/fmolb.2021.717969
50. Juan CX, Mao Y, Cao Q, et al. Exosome-mediated pyroptosis of miR-93-TXNIP-NLRP3 leads to functional difference between M1 and M2 macrophages in sepsis-induced acute kidney injury. J Cell Mol Med. 2021;25(10):4786–4799. doi:10.1111/jcmm.16449
51. Zheng L, Ling W, Zhu D, et al. Roquin-1 resolves sepsis-associated acute liver injury by regulating inflammatory profiles via miRNA cargo in extracellular vesicles. iScience. 2023;26(8):107295. doi:10.1016/j.isci.2023.107295
52. Cheng H, Wang X, Yao J, Yang C, Liu J. Mitophagy and ferroptosis in sepsis-induced ALI/ARDS: molecular mechanisms, interactions and therapeutic prospects of medicinal plants. J Inflamm Res. 2024;17:7819–7835. doi:10.2147/jir.S488655
53. He Z, Liao Z, Chen S, et al. Downregulated miR-17, miR-29c, miR-92a and miR-214 may be related to BCL11B overexpression in T cell acute lymphoblastic leukemia. Asia-Pac J Clin Oncol. 2018;14(5):e259–e265. doi:10.1111/ajco.12979
54. Shen Q, Huang X, Chen S, et al. BCL11B suppression does not influence CD34(+) cell differentiation and proliferation. Hematology. 2012;17(6):329–333. doi:10.1179/1024533212z.000000000145
55. Abe H, Kamimura K, Okuda S, et al. BCL11B expression in hepatocellular carcinoma relates to chemosensitivity and clinical prognosis. Cancer Med. 2023;12(14):15650–15663. doi:10.1002/cam4.6167
56. Hirose S, Touma M, Go R, et al. Bcl11b prevents the intrathymic development of innate CD8 T cells in a cell intrinsic manner. Int Immunol. 2015;27(4):205–215. doi:10.1093/intimm/dxu104
57. Drashansky TT, Helm E, Huo Z, et al. Bcl11b prevents fatal autoimmunity by promoting T(reg) cell program and constraining innate lineages in T(reg) cells. Sci Adv. 2019;5(8):eaaw0480. doi:10.1126/sciadv.aaw0480
58. Rimpelä JM, Pörsti IH, Jula A, et al. Genome-wide association study of nocturnal blood pressure dipping in hypertensive patients. BMC Med. Genet. 2018;19(1):110. doi:10.1186/s12881-018-0624-7
59. Cismasiu VB, Paskaleva E, Suman Daya S, Canki M, Duus K, Avram D. BCL11B is a general transcriptional repressor of the HIV-1 long terminal repeat in T lymphocytes through recruitment of the NuRD complex. Virology. 2008;380(2):173–181. doi:10.1016/j.virol.2008.07.035
60. Cherrier T, Suzanne S, Redel L, et al. p21(WAF1) gene promoter is epigenetically silenced by CTIP2 and SUV39H1. Oncogene. 2009;28(38):3380–3389. doi:10.1038/onc.2009.193
61. Desplats P, Dumaop W, Smith D, et al. Molecular and pathologic insights from latent HIV-1 infection in the human brain. Neurology. 2013;80(15):1415–1423. doi:10.1212/WNL.0b013e31828c2e9e
62. Vanvalkenburgh J, Albu DI, Bapanpally C, et al. Critical role of Bcl11b in suppressor function of T regulatory cells and prevention of inflammatory bowel disease. J Exp Med. 2011;208(10):2069–2081. doi:10.1084/jem.20102683
63. Walker JA, Oliphant CJ, Englezakis A, et al. Bcl11b is essential for group 2 innate lymphoid cell development. J Exp Med. 2015;212(6):875–882. doi:10.1084/jem.20142224
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.