Back to Journals » Cancer Management and Research » Volume 18

Piper nigrum L. Extract Modulates Cell Cycle and Promotes Apoptosis-Like Cell Death in BT-474 Breast Cancer Cells with Similar Effects in MCF-7 Cells
Authors Wiraswati HL ![]() , Ramadhanti J
, Ramadhanti J ![]() , Rohmawaty E
, Rohmawaty E ![]() , Berbudi A
, Berbudi A ![]() , Mayasari W, Laelalugina A
, Mayasari W, Laelalugina A ![]() , Afif AA, Setiawan AMD
, Afif AA, Setiawan AMD ![]() , Sofwah NFM, Edzana RA, Vučević K, Hidayati NA, Mochida K, Ma'ruf IF
, Sofwah NFM, Edzana RA, Vučević K, Hidayati NA, Mochida K, Ma'ruf IF
Received 19 February 2026
Accepted for publication 8 July 2026
Published 18 July 2026 Volume 2026:18 604276
DOI https://doi.org/10.2147/CMAR.S604276
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Seema Singh
Hesti Lina Wiraswati,1,2 Julia Ramadhanti,1,2 Enny Rohmawaty,1,2 Afiat Berbudi,1,2 Wulan Mayasari,3 Amila Laelalugina,2 Ali Abdullah Afif,4 Audrey Madina Dewi Setiawan,4 Naila Fathiyah Mufarrihah Sofwah,4 Raina Athaayaa Edzana,4 Katarina Vučević,5 Nur Akmalia Hidayati,6 Keiichi Mochida,7 Ilma Fauziah Ma’ruf8
1Department of Biomedical Sciences, Faculty of Medicine, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 2Oncology and Stem Cell Working Group, Faculty of Medicine, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 3Department of Forensic Medicine and Medicolegal Study, Faculty of Medicine, Universitas Padjadjaran, Bandung, West Java, Indonesia; 4Undergraduate Program, Faculty of Medicine, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 5Faculty of Medicine, University of Belgrade, Belgrade, Serbia; 6Research Center for Applied Microbiology, National Research and Innovation Agency, Bogor, Indonesia; 7RIKEN Center for Sustainable Resource Science, Yokohama City University, Yokohama, Japan; 8Research Center for Pharmaceutical Ingredients and Traditional Medicine, National Research and Innovation Agency, Bogor, West Java, Indonesia
Correspondence: Hesti Lina Wiraswati, Department of Biomedical Sciences, Faculty of Medicine, Universitas Padjadjaran, Sumedang, West Java, 45363, Indonesia, Tel/Fax +62 22 84288888, Email [email protected]
Purpose: Cancer therapy remains challenging because of recurrence, drug resistance, and severe side effects, driving the search for novel complementary strategies. In this study, we explore, for the first time, the potential of Piper nigrum L. (P. nigrum L. black pepper), a widely used spice known for its bioactive properties, in treating BT-474 luminal B and MCF-7 luminal A breast cancer cells.
Materials and Methods: BT-474 cells were treated with P. nigrum L. ethanolic extract (PNEE). Cytotoxicity, cell morphology, gene and protein expression, cell death and cell cycle progression were evaluated using the MTT, microscopy, Reverse Transcriptase Quantitative Polymerase Chain Reaction (RT-qPCR), Enzyme-Linked Immunosorbent Assay (ELISA), and flow cytometry, respectively. MCF-7 cells were included as a comparative breast cancer model, whereas 3T3-L1 cells were used as a non-cancerous model.
Results: Our findings demonstrate PNEE’s multimodal activity in BT-474 cells, decreasing cell viability with an IC50 of 58 ppm, and significantly suppressed colony formation. PNEE induced S-phase cell cycle arrest, and modulated several cancer-associated markers, including EGFR, PI3K/AKT pathway components, cyclin A, MMP-9, HIF-1α, and BCL2. Furthermore, PNEE elicited a regulated cell death phenotype with apoptotic features, distinct Annexin V/PI profiles in BT-474 luminal B and MCF-7 luminal A breast cancer cells. In addition, PNEE showed lower cytotoxicity toward non-cancerous 3T3-L1 cells under experimental condition tested.
Conclusion: PNEE exerts multiple anticancer effects in BT-474 luminal B breast cancer cells and induces a regulated cell death phenotype that does not fully conform to a canonical caspase-dependent apoptotic pattern. Distinct cell death responses observed between BT-474 and MCF-7 cells further suggest that breast cancer subtype-specific factors may influence the cellular response to PNEE. These findings support further investigation of PNEE, including mechanistic studies and evaluation of efficacy and safety in appropriate in vivo models.
Keywords: Piper nigrum L., black pepper, luminal B breast cancer, regulated cell death, cell cycle arrest, PI3K/AKT signaling
Introduction
Breast cancer is currently the most commonly diagnosed malignancy in women, with approximately 2.30 million new cases and 666,103 deaths worldwide.1 Among its molecular subtypes, Luminal B tumor has a poorer prognosis, with a higher tendency to metastasize to distant organs, leading to significantly lower post-relapse survival rates than Luminal A.2 Although significant progress has been achieved in breast cancer management, current treatments still face considerable challenges, including severe side effects such as fatigue, loss of appetite or diarrhea that often diminish patients’ quality of life.3 Therefore, there is continued interest in exploring new drug candidates to improve the therapeutic effect of existing cancer drugs, improving patients’ quality of life, and preventing cancer.
Natural substances derived from medicinal plants have attracted increasing scientific attention due to their diverse bioactive compounds and potential to modulate cancer-related pathways.4–6 Several studies have explored their roles as complementary approaches in cancer treatment, including in aggressive breast cancer subtypes such as luminal B.7 Herbal medicine is widely used as a forms of Complementary and Alternative Medicine (CAM) among cancer patients, including those with breast cancer, with reported roles in symptom management and supportive therapy.8–11 Numerous studies have demonstrated that herbal-derived compounds exhibit anticancer properties through multiple mechanisms, including the inhibition of the cell proliferation and modulation of cell death pathways.5,12–14 Emerging evidence also highlights potential synergistic interactions between herbal compounds and conventional chemotherapy agents.15–17
This transition from palliative to therapeutic exploration has drawn attention to botanicals with potential biological bioactivity. Among these, Piper nigrum L. (P. nigrum L., black pepper), which is one of the world’s most popular natural spice products, has received considerable attention due to its reported pharmacological effects, including potential anticancer activity.18,19 The pharmacological activities of P. nigrum L. are largely associated with its bioactive compounds, particularly alkaloids. Among these compounds, piperine and piperidine derivatives have been extensively studied and are well-documented to regulate cell proliferation, induce apoptosis, and inhibit metastasis processes.20,21 In addition to these majors constituents, several other alkaloids and alkaloid-like compounds, including piperamide, piperamine, sarmentine, trichostachine, and piperettine, have been reported to exhibit anticancer activity.22–26 However, compared with piperine, these compounds remain relatively underexplored, and their molecular mechanisms responsible for their biological activities are still not fully understood.
Although numerous ethnomedicinal plants remain unexplored, the selection of P. nigrum L. has attracted interest because of its reported reproducible anticancer activity, a long history of human consumption, and a relatively well-established safety profile. However, most previous studies have focused on isolated compounds such as piperine, whereas the biological activity and underlying mechanisms of P. nigrum L. ethanolic extract (PNEE) remains poorly understood, particularly in molecularly distinct breast cancer subtypes such as luminal B tumors.
Previous studies have demonstrated the anticancer potential of PNEE through multiple mechanisms, including induction of cell death, reduction of cell proliferation, and generation of oxidative stress in various cancer cell models.27–31 In breast cancer, PNEE exhibits significant anticancer activity through reduce cell viability, induce cell death, and modulate cell cycle progression in MCF-7 cells.27,28 In addition, chemopreventive effects have been reported in NMU-induced mammary tumor rat models, where P. nigrum L. treatment was associated with reduced expression of MMP-2, MMP-9, and VEGF, suggesting potential anti-metastatic and anti-angiogenic activities.27,29 However, its effects in luminal B breast cancer remain poorly characterized. Given the heterogeneity of breast cancer subtypes, it is critical to assess whether the observed anticancer effects of P. nigrum L. extracts are conserved across different molecular profiles. Furthermore, EGFR and PI3K/AKT signaling pathways have emerged as important molecular targets for anticancer drug development. While previous studies have demonstrated that P. nigrum L. extracts and their major alkaloid constituents exert antiproliferative and pro-apoptotic effects in various cancer models, the involvement of EGFR and PI3K/AKT signaling in the anticancer activity of crude PNEE remains largely unexplored.26 Therefore, these molecular markers were included in the present study to investigate potential mechanisms underlying PNEE activity in luminal B breast cancer cells.
Despite these promising findings, the anticancer effects of PNEE on luminal B breast cancer cells, particularly BT-474 cells, remain unexplored. Deng et al27 reported potent activity of piperine-free P. nigrum fruit extract (PFPE) against ZR-75-1 cells, another luminal B subtype, however, the effect of PNEE on BT-474 cells have not been characterized. Given the established roles of EGFR/PI3K/AKT signaling, Cyclin A-mediated cell cycle progression, MMP-9-associated metastasis, BCL2-regulated cell survival, and HIF-1α-mediated stress responses in breast cancer progression, these molecular markers were selected to further investigate the mechanisms underlying PNEE activity. Therefore, this study aimed to address this gap by evaluating the potential of PNEE as a therapeutic agent against BT-474 breast cancer cells. MCF-7 cells, a luminal A subtype, were included for comparative cell death profiling, whereas non-cancerous 3T3-L1 fibroblasts were used to assess the selectivity of PNEE cytotoxicity.
Materials and Methods
Materials
The materials used in this research include the human breast cancer cell lines BT-474 (#HTB-20, ATCC, USA) and MCF-7 (#HTB-22, ATCC, USA), as well as the non-cancerous mouse fibroblast cell line 3T3-L1 (CL-173TM, ATCC, USA) cell lines, Roswell Park Memorial Institute (RPMI, Cat. No. SA R8758, Sigma Aldrich, Germany), fetal bovine serum (FBS, Cat. No. SA F7524, Sigma Aldrich, Germany), penicillin-streptomycin (Cat. No. SA P4333, Sigma Aldrich, Germany), dimethyl sulfoxide (DMSO) (Sigma Aldrich, Germany), trypsin (GIBCO, USA), Dulbecco’s phosphate buffered saline (DPBS, Cat. No. D8662, Sigma Aldrich, Germany), ethanol 70% (LKPI, Indonesia), 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (Cat. No. M2003, MTT, Sigma-Aldrich, St. Louis, MO, USA), RNase A II (Geneaid, Taiwan), propidium iodide (Cat. No. P4170, Sigma Aldrich, Germany), radioimmunoprecipitation assay (RIPA) (Sigma Aldrich, Germany), Dulbecco’s Phosphate-Buffered Saline (DPBS) (Sigma Aldrich, Germany). Protein expression analyses were performed using Human EGFR ELISA Kit (Cat. No. SEA757Hu, CCC, USA), Human AIF (Apoptosis-Inducing Factor) ELISA Kit (Cat. No. EH2606, CCC, USA), and Human Caspase-3 ELISA Kit (Cat. No. E-EL-H0017, Elabscience, USA). The Total RNA Mini Kit (RB100, Geneaid, Taiwan) and primers for gene amplification (β-actin, cyclin A, PI3K, AKT, MMP-9, EGFR, and HIF1α) were obtained from Integrated DNA Technologies (USA). Ethanol (96%), methanol (99.8%), sodium carbonate (Na2CO3), sodium nitrite (NaNO2), sodium hydroxide (NaOH), ammonia solution (NH3), chloroform (CHCl3), glacial acetic acid (CH3COOH), magnesium powder (Mg), and sulfuric acid (H2SO3) were obtained from Merck (Darmstadt, Germany). Phytochemical-specific reagents, including iron (III) chloride (FeCl3), 2.2-diphenyl-1-picrylhydrazyl (DPPH), Folin-Ciocalteu reagent, gallic acid (C7H6O5), aluminum chloride (AlCl3), quercetin (C15H10O7), and polyvinylpolypyrrolidone (PVPP), were obtained from Sigma-Aldrich (St. Louis, USA). Alkaloid detection reagents (Wagner’s and Dragendorff’s) were freshly prepared in-house using analytical-grade iodine (I2), potassium iodide (KI), bismuth subnitrate (BiONO3·H2O), and acetic acid (Merck, Germany). Primers used in this study are listed in Table 1.
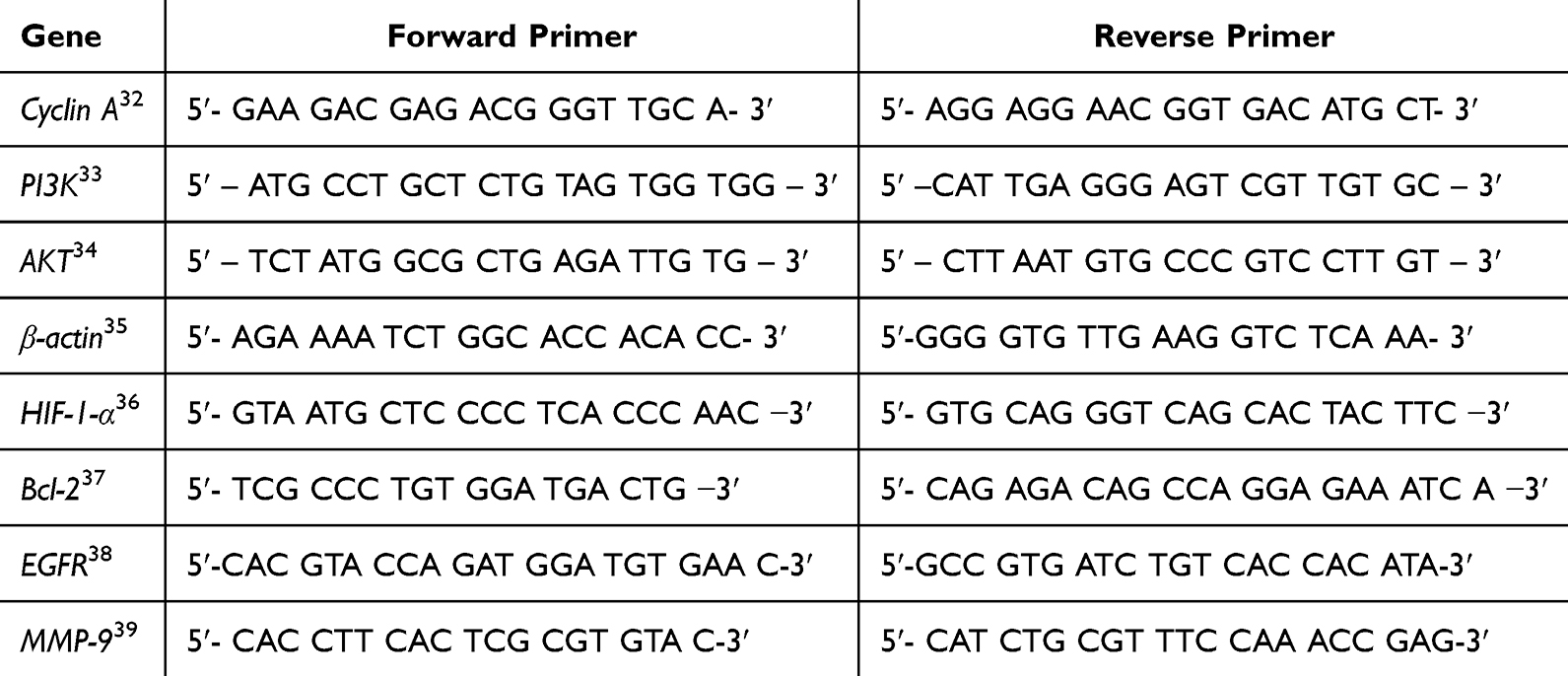 |
Table 1 The Primers Used in This Study |
Ethical Approval
P. nigrum L. plants were collected from a plantation in West Bandung, West Java Province, Indonesia with permission from the plantation owner. This permit is sufficient under Indonesian regulations on biodiversity use, as P. nigrum L. is a cultivated species that is not listed as protected by Government Regulation No. 7 of 1999. All experimental procedures involving plant materials complied with institutional approval from the Faculty of Medicine, Universitas Padjadjaran, as well as national legislation on biodiversity and conservation of natural resources, including Law No. 32 of 2009 on Environmental Protection and Management and Law No. 11 of 2013 (ratification of the Nagoya Protocol on Access and Benefit Sharing) and the international frameworks of the Convention on Biological Diversity (CBD) and the Nagoya Protocol. A voucher specimen of P. nigrum L. (Voucher No. 581, Plant Identification Form No. 464/LBM/IT/XI/2024) was deposited in the Herbarium Jatinangoriense (Biosystematics and Molecular Laboratory, Biology Department, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Indonesia). Taxonomic identification of the plant material was conducted by taxonomist Joko Kusmoro, following the methodologies described in Anderson’s Flora Taxonomica40 and augmented with data from the Global Biodiversity Information Facility.41 A representative specimen of Piper nigrum L. used in this study is shown in Figure 1.
 |
Figure 1 Piper nigrum L. plant used as the source of peppercorns for extract preparation in this study. |
P. Nigrum L. Ethanol Extract (PNEE) Preparation
To prepare the extract, 200 g of dried peppercorn from P. nigrum L., was finely ground to enhance the solvent penetration. Black pepper plants were macerated in 96% ethanol at room temperature for 48 hours. After maceration, the solution was filtered sequentially through filter paper and 0.45 µm syringe to remove particulates, and the filtrate was then concentrated using a rotary evaporator (40–50°C under reduced pressure) in a fume hood until a viscous crude extract (PNEE) were obtained. The final extract was transferred in vials at 4°C or −20°C for long-term stability. To formulate the 100,000 ppm PNEE stock solution, 100 mg of the extract was homogenized in 1 mL dimethyl sulfoxide (DMSO) using a vortex mixer. The stock solution was aliquoted into light-protected vials and refrigerated (4°C) until further use. The stock solution was diluted in the RPMI-1640 complete medium to achieve the desired working concentrations, prior to cellular treatment. The extract was evaluated as a crude preparation without quantitative standardization of specific marker compounds, such as piperine. Consequently, the observed biological effects reflect the combined activity of multiple constituents present in the extract.
Phytochemical Screening
Phytochemical screening was conducted to identify the secondary metabolites in PNEE using qualitative colorimetric assays. For each test, 2 g of the extract was dissolved in distilled water or ethanol, followed by reagent-specific reactions: phenolics were detected via 5% FeCl3 (bluish-black coloration), tannins with 1% FeCl3 (green, red, purple, blue, or black hues), and flavonoids through sequential addition of magnesium powder, concentrated HCl, 2N H2SO4, and 10% NaOH (orange, pink, or dark red coloration persisting ≥3 minutes). Alkaloids were detected using Wagner’s reagent (chocolate-brown precipitate), whereas saponins were confirmed by the formation of stable foam after shaking with 2N HCl. Color changes, precipitates, or foam stability were recorded immediately after reagent application, enabling preliminary characterization of bioactive compounds in the extract. This approach provided a preliminary qualitative characterization of the major phytochemical classes present in PNEE.
Cancer Cell Culture and Treatment
The BT-474 luminal B breast cancer, MCF-7 luminal A breast cancer, and normal fibroblast 3T3-L1 cell lines were cultured in RPMI medium supplemented with 10% FBS 100 IU/mL and 5% PS (100 µg/mL). Cells were maintained in a CO2 (5%) incubator at 37°C. For cytotoxicity studies, BT-474 and 3T3-L1 cells were treated with PNEE at the indicated concentrations. DMSO at the corresponding final concentration served as the negative (vehicle) control. BT-474 cells were used as the primary model for evaluating the cytotoxic and mechanistic effects of PNEE. Non-cancerous 3T3-L1 fibroblasts were included to assess the selectivity of PNEE cytotoxicity. MCF-7 cells were used solely as a comparative breast cancer model for cell death profiling by Annexin V/PI flow cytometry and were not included in cytotoxicity assays.
MTT Assay
BT-474 or 3T3-L1 cells were seeded in 96 well plates at a density of 10,000 cells/well and treated with various doses of PNEE. Extracts were obtained according to the methods described above. PNEE (50 mg) was dissolved in 500 µL dimethyl sulfoxide (DMSO), and cells treated with the corresponding concentration of DMSO alone served as the vehicle control (0 ppm PNEE). The experimental design consisted of one control group and 8 PNEE treatment groups (1, 5, 10, 25, 50, 75, 100, and 200 ppm). Tests were performed at the indicated doses for 24 h. Toxicity tests were conducted using the MTT assay kit. The absorbance was measured using an ELISA reader (Multiscan EX, Thermo Scientific®, Singapore) at a wavelength of 550 nm.
Microscopical Observation of Changes in Cell Morphology
The treated BT-474 cells were then observed under an inverted microscope to observe the morphological changes that marked apoptosis, including the formation of apoptotic bodies, rounding of the cells, or membrane blebbing. In addition, colony growth was monitored by serial microscopic imaging. Due to the tendency of BT-474 cells to grow as compact three-dimensional colony-like aggregates, colony expansion was assessed by measuring colony area rather than by conventional colony counting. The colony size was observed under an inverted microscope and the relative colony area was quantified using ImageJ software (NIH, USA) and expressed as a percentage of the control group.42
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
After treatment, RNA was extracted using a Total RNA Mini Kit (Geneaid, Taiwan). mRNA expression of Cyclin A, PI3K, AKT, EGFR, MMP9, BCL2, and HIF1α was determined by RT-qPCR using the SensiFAST™ SYBR® No-ROX One-Step Kit (Meridian Biosciences) and specific primers in the AriaMx Real-Time PCR System (Agilent Technologies, USA). The PCR conditions were as follows: 45°C for 10 min for reverse transcription, 95°C for 2 min, and 40 cycles of 95°C for 5 s, and 60°C for 20s. Data were analyzed by the 2−ΔΔCT method, using the housekeeping gene β-actin as the internal control. However, the use of a single housekeeping gene may represent a limitation for normalization.
Enzyme-Linked Immunosorbent Assay (ELISA)
BT-474 cells were seeded into 24-well plates at a density of 40,000 cells/well. After treatment, cellular proteins were extracted by lysis in RIPA buffer to generate protein lysates. The expression levels of EGFR, Apoptosis-Inducing Factor (AIF), and Caspase-3 proteins were quantified using commercially available ELISA kits specific to each target following the manufacturer’s standardized protocol. Protein concentrations were normalized prior to ELISA analysis to ensure comparability between samples.
Flow Cytometry
BT-474 and MCF-7 cells were seeded in 24-well plates at a density of 30,000 cells per well and allowed to adhere overnight. The cells were treated with PNEE at the specified concentrations for 24 h. Following treatment, the spent medium was aspirated and pooled into a 15 mL falcon tube. Cells were washed with 1 mL PBS, and the wash solution was transferred to the collected medium. Adherent cells were detached using 200 µL of 0.25% trypsin-EDTA solution (5-minute incubation at 37°C), neutralized with complete medium, and transferred to the same falcon tube. The cell suspension was centrifuged (1,500 rpm, 5 min), and the pellet was resuspended in 1 mL DPBS for subsequent analysis.
Cell death was evaluated via cytofluorometric assessment using fluorescein isothiocyanate (FITC)-labeled Annexin V to detect phosphatidylserine externalization and Propidium Iodide (PI) to identify membrane-permeabilized cells. Initial gating excluded debris based on the forward scatter (FSC) and side scatter (SSC) parameters. Apoptotic populations were identified using dual-parameter plots: Annexin V-FITC (FL1 channel) versus PI (FL3 channel). Cells were classified as PI-/Annexin V− (viable), PI-/Annexin V+ (early apoptotic), PI+/Annexin V+ (late apoptotic or secondary necrotic), and PI+/Annexin V− (necrotic). Cell size alterations were additionally evaluated using PI versus forward scatter (FSC) plots, where reduced FSC was interpreted as a decrease in cell size, a morphological feature commonly associated with apoptosis. Selected Annexin V/PI and PI/FSC subpopulations were quantified for comparative analysis.
For cell cycle analysis, ethanol-fixed cells were stained with PI to quantify the DNA content. Cell cycle distribution was determined from DNA content histograms and quantified as the percentage of cells in G0/G1, S, and G2/M phases. The samples were analyzed using a BD FACSLyric™ Flow Cytometry System (BD Biosciences) equipped with FlowJo™ software (version 10.8; BD Biosciences).
Statistical Analysis
All data were obtained from three independent biological replicates, each of which contained triplicate technical measurements. Results are presented as mean ± standard error of the mean (SEM). Statistical analysis was performed based on predefined comparisons between each group and its corresponding control using two-tailed Student’s t-test in Microsoft Excel (Microsoft Office Home 2024, Version 2506). The threshold for statistical significance was set at p-value of < 0.05.
Results
Phytochemical Constituents of PNEE
Qualitative phytochemical screening of PNEE indicated the presence of flavonoids and alkaloids, whereas phenolics, tannins, and saponins were not detected (Table 2). Flavonoids were confirmed by the formation of a persistent orange-red coloration following sequential treatment with magnesium powder, concentrated HCl, 2N H2SO3, and 10% NaOH. Alkaloids were identified by the appearance of a distinct chocolate-brown precipitate following addition of Wagner’s reagent. Tests for phenolic compounds, tannins, and saponins were negative: no bluish-black color was observed with 5% FeCl3, no color changes indicating tannins with 1% FeCl3, and no stable foam was produced in the 2N HCl shake test. The absence of these compounds indicates that flavonoids and alkaloids represent the major classes of secondary metabolites detected in PNEE. These findings provide preliminary phytochemical basis for further investigation of the biological activities of PNEE.
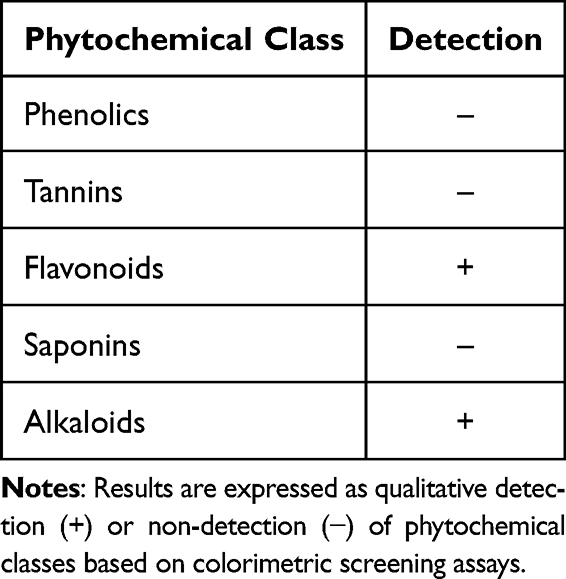 |
Table 2 Qualitative Phytochemical Screening of PNEE |
PNEE Inhibits Cell Proliferation and Induces S-Phase Cell Cycle Arrest
The cytotoxic effect of PNEE on BT-474 breast cancer cells was assessed using a MTT assay (Figure 2). Cell viability decreased with increasing PNEE concentration, yielding an IC50 value of 58 ppm, indicating a concentration-dependent response. At concentrations ≤10 ppm, cell viability remained near 100%, indicating minimal toxicity. The decrease in cell viability began at 25 ppm (80.9%), dropping significantly to 58.4% at 50 ppm and 36.2% at 75 ppm. No further substantial reduction in viability was observed at concentrations above 200 ppm, suggesting that the cytotoxic effect had reached a plateau (Figure 2A). Based on these findings, subsequent experiments were performed to determine whether this reduction in cell viability was associated with inhibition of cell proliferation, cell cycle perturbation, and/or induction of cell death, with significant bioactivity thresholds identified between 25 and 100 ppm where the biological effects became evident.
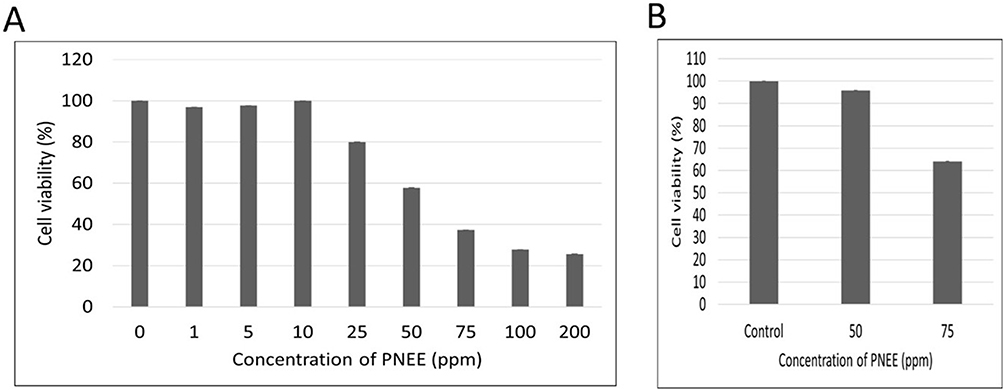 |
Figure 2 Cytotoxic effects of PNEE on (A) HER2-positive BT-474 breast cancer cells and (B) 3T3-L1 fibroblasts. Cells were treated with the indicated concentrations of PNEE for 24 h, followed by MTT viability assay (absorbance measured at λ=550 nm). The 0 ppm group represents the DMSO vehicle control. Values represent mean ±SD of three independent experiments. |
To evaluate potential selective toxicity, we examined the effect of PNEE in non-cancerous 3T3-L1 fibroblast cells at two relevant concentrations. Results showed that 50 ppm PNEE did not induce cytotoxicity (98.2% cell viability), whereas 75 ppm PNEE induced 35% toxicity (Figure 2B). Considering that 50 ppm markedly reduced the viability of BT-474 cells while exerting minimal effects on normal fibroblasts, we selected 50 ppm PNEE for all subsequent experiments.
To assess long-term survival, a colony formation analysis was performed, as illustrated in Figure 3. At 0 h, the initial colony sizes were comparable between the control and treated groups. However, after 24 h, the control colonies showed substantial expansion, whereas the PNEE-treated colonies remained compact and showed minimal growth. This divergence became more pronounced at 72 h; control colonies occupied larger confluent areas, whereas PNEE-treated colonies remained sparse, small, and fragmented (Figure 3A). ImageJ quantification demonstrated that the relative colony area of PPNE-treated cells was reduced approximately 74% to 25% at 24h and from 100% to 30% at 72h compared with the corresponding control groups (Figure 3B). These findings indicate that PNEE exerts a potent time-dependent inhibitory effect on BT-474 cell proliferation and impairs clonogenic survival, thus strengthening its cytotoxic and antiproliferative potential against BT-474 breast cancer cells.
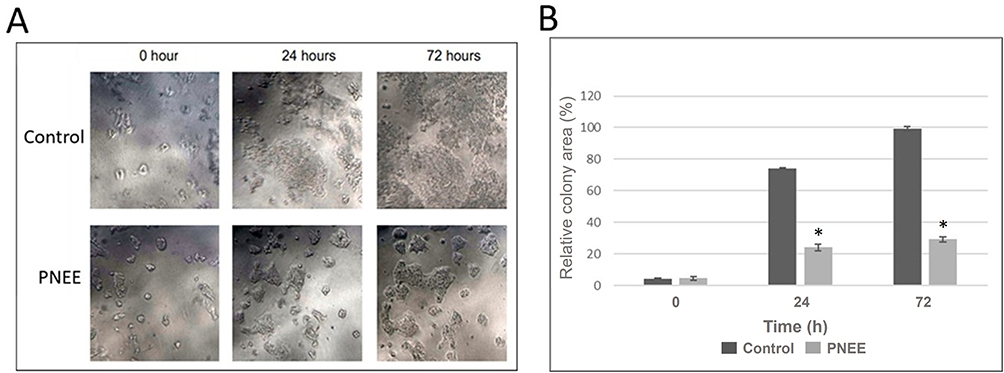 |
Figure 3 PNEE reduced the colony area of BT-474 cells. (A) Representative bright-field microscopy images captured using an inverted phase-contrast microscope (100× magnification) showing the colony morphology of untreated (control) and PNEE-treated (50 ppm) BT-474 cells at 0, 24, and 72 h post-treatment. (B) Quantitative analysis of relative colony area of PNEE treatment compared to control, using ImageJ software. Data are presented as mean ± SEM from three independent experiments. Statistical comparisons between control and PNEE-treated groups at each time point were performed using a two-tailed Student’s t-test (*p < 0.05). |
Flow cytometric analysis revealed that treatment with 50 ppm PNEE for 24 h increased the proportion of BT-474 cells in the S-phase from 16.4% in control cells to 27.2% in PNEE-treated cells, indicating S-phase cell cycle arrest. This increase was accompanied by a reduction in the G0/G1 population (58.5% to 51.2%) and the G2/M population (18.5% to 16.4%) (Figure 4A and B). Consistent with these findings, RT-qPCR analysis showed significant downregulation of the cell cycle regulator cyclin A (0.02-fold), PI3K (0.18-fold), AKT (0.14-fold), and EGFR (0.04-fold) (Figure 4C). ELISA analysis further demonstrated a reduction in EGFR protein level from approximately 19.5 ng/mL in control cells to 4.5 ng/mL in PNEE-treated cells, following PNEE treatment (Figure 4D), consistent with the decrease of EGFR mRNA expression. RT-qPCR analysis further demonstrated an increase in HIF-1α expression following PNEE treatment (Figure 4C), suggesting activation of a cellular stress-response despite the simultaneous suppression of EGFR/PI3K/AKT pathway components and Cyclin A. In addition, expression of the metastasis-related gene MMP9 was reduced to 0.82-fold relative to the control group (Figure 4C), indicating a potential inhibitory effect of PNEE on metastasis-related signaling.
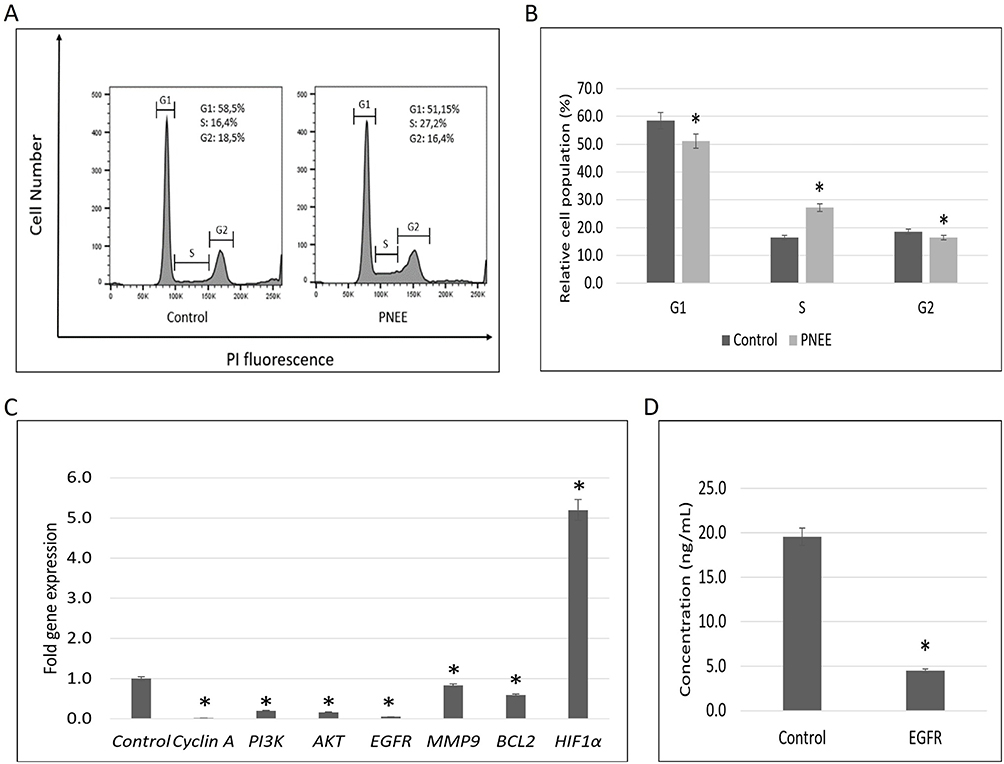 |
Figure 4 PNEE induced cell cycle arrest and modulated growth-related molecular markers in BT-474 cells. BT-474 cells were treated with 50 ppm PNEE for 24 h. (A and B) Cells were fixed with 70% ethanol, stained with propidium iodide (PI), and analyzed by flow cytometry to determine cell cycle distribution. The x-axis indicates the cell cycle phase, and the y-axis represents the relative cell population. (C) Gene expression levels of cyclin A, PI3K/AKT, EGFR, MMP9, BCL-2, and HIF1α were measured by RT-qPCR, using β-actin as the housekeeping gene. Expression data were based on CT values from the control and treated groups. (D) EGFR protein levels were quantified by ELISA and absorbance was measured at 450 nm. All results are presented as the mean ± SD from three independent experiments. Statistical significance (triplicate experiments) analyzed by two-tailed t-test is denoted as *p < 0.05 compared to the control. |
PNEE Promotes Apoptosis-Like Cell Death in BT-474 Cells and Early Apoptotic Features in MCF-7 Cells
Flow cytometric analysis demonstrated that treatment with 50 ppm PNEE for 24 h induced distinct Annexin V/PI staining profiles in both BT-474 luminal B and MCF-7 luminal A breast cancer cells (Figure 5A, B and Table S1). Quantitative analysis of Annexin V/PI staining revealed significant alterations in cell population distribution following PNEE treatment. In both cell lines, apoptosis represented the predominant mode of cell death, whereas the necrotic population remained comparatively low (Table S1).
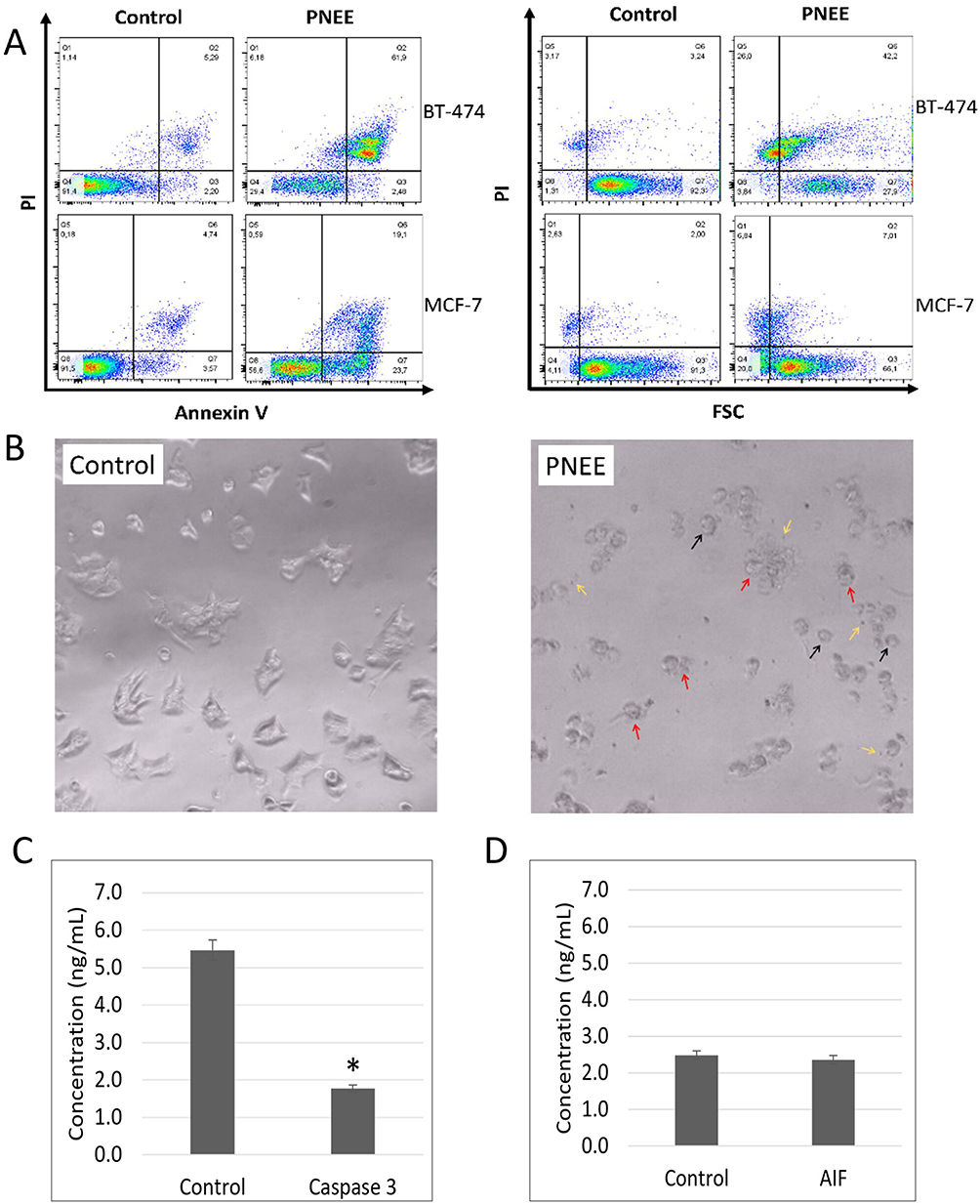 |
Figure 5 PNEE induced cell death in BT-474 luminal B and MCF-7 luminal A breast cancer cells. (A) Representative Annexin V/PI and PI/FSC flow cytometry plots. (B) Bright-field microscopy showing cell rounding (black arrows), membrane blebbing (red arrows), and apoptotic body formation (yellow arrows), in BT-474 cells treated with 50 ppm PNEE for 24 h compared to solvent control. (C) Caspase 3 and (D) AIF protein levels were quantified using ELISA in BT-474 cells following protein extraction, with absorbance measured at 450 nm. Statistical significance (triplicate experiments) analyzed by two-tailed t-test is denoted as *p < 0.05 compared to the control. |
In BT-474 cells, PNEE markedly increased the PI+/Annexin V+ population from 3.00±1.43% in the control group to 63.77±1.47% following treatment, whereas the PI−/Annexin V+ population remained low (2.21±0.26%) (Figure 5A and Table S1). The predominance of PI⁺/Annexin V⁺ cells, together with the increased PI⁺/FSC− population, is consistent with a late apoptotic phenotype characterized by lost membrane integrity, phosphatidylserine externalization, and reduced cell size (Figure 5A and Table S1, S2). Quantitative analysis further demonstrated a marked increase in the late apoptosis-like population from 3.00 ± 0.43% in untreated cells to 63.77 ± 1.47% following PNEE treatment, whereas the necrotic population remained comparatively low (1.29 ± 5.77%) (Table S1). Bright-field microscopy confirmed these findings, with PNEE-treated BT-474 cells displaying classical apoptotic morphology, including cell round-up, membrane blebbing, apoptotic body formation, which were absent in control groups (Figure 5B). However, ELISA demonstrated no upregulation of caspase-3 (5.5 ng/mL in control cells to 1.8 ng/mL following PNEE treatment) or AIF protein levels (2.5 ng/mL in control cells versus 2.4 ng/mL following PNEE treatment) (Figures 5C and D), suggesting that the observed apoptosis-like phenotype is not fully explained by canonical caspase-dependent or AIF-mediated cell death pathways.
In contrast, MCF-7 cells exhibited a distinct apoptotic response to PNEE treatment (Figure 5A and Table S1, S2). Although a PI+/Annexin V+ population was observed (18.15±0.71%), this proportion was substantially lower than that detected in BT-474 cells (63.77±1.47%) (Figure 5A and Table S1). In addition, the proportion of PI+/FSC− cells was lower in MCF-7 cells (7.07±0.46%) than in BT-474 cells (25.90±0.08%), suggesting less progression toward advanced apoptotic morphology (Figure 5A and Table S2). Conversely, MCF-7 cells exhibited a higher proportion of PI−/Annexin V+ cells (56.50%) than BT-474 cells (2.21%), indicating a greater representation of early apoptotic cells (Figure 5A and Table S1). This observation was further supported by a larger PI−/FSC− population in MCF-7 cells (19.37±0.90%) compared with BT-474 cells (3.35±0.71%), consistent with increased cell shrinkage in the absence of membrane permeabilization (Table S2). Together, these results suggest that PNEE induced a distinct apoptotic response in MCF-7 cells, characterized by a greater representation of early apoptotic features and reduced progression to late stages than in BT-474 cells.
Discussion
This study demonstrates the anticancer potential of P. nigrum L. ethanolic extract (PNEE) against BT-474 luminal B breast cancer cells for the first time. Our findings highlight that PNEE not only inhibits cell proliferation and colony formation but also induces S-phase cell cycle arrest and features consistent with apoptosis-like cell death, with molecular changes suggestive of stress-related pathways, HIF-1α upregulation. These findings expand the therapeutic potential of P. nigrum L. beyond that of previously studied cancer models, especially in tackling luminal B breast cancer, which is known for its aggressive behavior and treatment resistance.2,30 P. nigrum L. has long been utilized in traditional medicinal systems, including Ayurveda and traditional Asian medicine, for the management of various ailments. It has also been traditionally used in several regions as a complementary herbal remedy for tumor-related conditions, and its ethnomedicinal use has motivated further investigations into its anticancer potential. The present findings further support these reports by demonstrating the antiproliferative effects, S-phase cell cycle arrest, and apoptosis-like cell death induced by PNEE in BT-474 luminal B breast cancer cells.
Phytochemical screening of PNEE identified the presence of flavonoids and alkaloids, whereas phenolic compounds, tannins, and saponins were notably absent. Previous studies have reported variability in the phytochemical composition of P. nigrum L. extracts depending on the plant material, extraction solvent, and analytical methods employed. Nevertheless, alkaloids and flavonoids are among the most consistently reported phytochemical classes in P. nigrum L., and have been widely associated with its anticancer activity through several well-established pathways.6 Alkaloids have been reported to modulate key oncogenic signaling cascades, particularly the PI3K/AKT31 and EGFR43 pathways, can suppress STAT3 activation,44 and promote mitochondria-mediated apoptosis by disrupting the balance between pro-apoptotic Bax and antiapoptotic Bcl-2 proteins triggering the apoptotic cascade.45 Alkaloids reported to interfere cell cycle progression by modulating cyclin-dependent kinases, resulting in cell cycle arrest at the S-phase,46 activated programmed cell death through both intrinsic (mitochondrial) and extrinsic (death receptor) pathways by regulating Bcl-2 family proteins and caspase activation,47,48 and demonstrated antimetastatic properties by inhibiting epithelial-mesenchymal transition and matrix metalloproteinase activity.49 Caspase-independent apoptosis has also been reported by flavonoid or alkaloid induction in various cancer cells.40,50,51 Therefore, the phytochemical profile observed in the present study provides a plausible biochemical basis for the antiproliferative activity, S-phase cell cycle arrest, and apoptosis-like phenotype induced by PNEE in BT-474 cells.
The differential sensitivity observed between BT-474 and 3T3-L1 cells is consistent with previous reports showing that cancer cells are generally more susceptible to phytochemicals because of their altered metabolism, dysregulated proliferative signaling, and increased oxidative stress.41,42 However, because 3T3-L1 fibroblasts are not normal mammary epithelial cells, these findings should be interpreted as preliminary evidence of differential sensitivity rather than definitive tumor selectivity.
The antiproliferative activity of PNEE observed in the MTT and colony formation assays is consistent with previous studies identifying alkaloids as a major bioactive constituents of P. nigrum L. with cytotoxic and growth-inhibitory properties.21,22,26 In the present study, PNEE significantly suppressed BT-474 cell proliferation in dose- and time-dependent manners. These findings are consistent with previous reports describing the antiproliferative effects of PNEE in various cancer models,26 confirming its broad-spectrum efficacy and establishing its potency against the more clinically challenging luminal B subtype. Our results also agree with a recent work on Piperine-Free P. nigrum Fruit Extract (PFPE), an ethanol-based fraction that exhibited antiproliferative effects in ZR-75-1 luminal B cells.27 Together, these findings suggest that the anticancer activity of P. nigrum L., is not solely attributable to single bioactive compound but rather to the combined effects of multiple constituents in the extract.
At the cellular level, PNEE-induced cell cycle arrest was specific to the S phase in BT-474 luminal B breast cancer cells, accompanied by downregulation of cyclin A and key regulators of the PI3K/AKT pathway. As a receptor tyrosine kinase (RTK), EGFR activation typically triggers the downstream PI3K/AKT and Ras/MAPK pathways, promoting cyclin-CDK complex formation and cell cycle progression.52 Therefore, suppression of these molecules provides a plausible molecular basis for the antiproliferative effects observed in the present study. These findings are consistent with previous reports demonstrating that PNEE induces cell cycle arrest in several cancer models, although the affected phase appears to be cell type-dependent. Whereas G0/G1 arrest has been reported in MCF-7 (breast), PANC-1 (pancreatic), HeLa (cervix), and KKU-100 (CCA) cancer cell lines,53–55 PNEE induced S-phase arrest in BT-474 cells, A similar S-to-G2/M cell cycle blockade has also been reported in KKU-M452 cholangiocarcinoma (CCA) cells following PNEE treatment,53 supporting the notion that the effects of PNEE on cell cycle progression are context-dependent. The observed suppression of EGFR is consistent with our in silico predictions and other studies, wherein non-piperine P. nigrum derivatives, including piperamide and piperamine derivates may interact with EGFR.22,56,57
To our knowledge, this is the first study demonstrating coordinated suppression of EGFR and PI3K/AKT signaling pathway by PNEE in BT-474 luminal B breast cancer cells. Although inhibition of this pathway has previously been reported for isolated P. nigrum L. constituents, particularly piperine, in SNU-16 gastric cancer cells and in a xenograft tumor model,58 evidence for the crude ethanolic extract has remained limited. Therefore, our findings expand the current understanding of the molecular mechanisms underlying the antiproliferative activity of PNEE. Furthermore, the downregulation of MMP-9 expression is consistent with previous studies in MCF-7 luminal A breast cancer cells53 and NMU-treated mammary tumor rats treated with piperine-free P. nigrum fruit extract (PFPE),27 suggesting that PNEE may also influence metastasis-related pathways. Nevertheless, whether these molecular changes translate into functional inhibition of migration, invasion, or metastasis remains to be determined.
Importantly, our study revealed that PNEE induced distinct apoptosis in BT 474 luminal B and MCF-7 luminal A breast cancer cells. Although both cell types exhibited shrinkage and phosphatidylserine exposure, hallmarks of apoptosis, their apoptotic profiles differed. BT-474 cells displayed rapid progression to late apoptosis, whereas MCF-7 cells showed early apoptotic dominance, but minimal late apoptosis, suggesting a delayed or incomplete apoptotic program. This differential response may reflect intrinsic variations in the apoptotic machinery between luminal A and B subtypes, which may influence therapeutic outcomes. Notably, no upregulation of caspase-3 or AIF protein was observed in BT-474 cells. As caspase-3 and AIF are key mediators of canonical apoptosis and caspase-independent cell death, respectively,59 these findings suggest that the observed cell death may not be fully explained by increased expression of these pathways alone. Our findings align with previous reports that revealed that certain phytochemicals can trigger alternative cell death pathways, particularly in apoptosis-resistant cancer cells.60
The upregulation of HIF-1α and suppression of Bcl-2 further support activation of cellular stress responses and mitochondrial destabilization, following PNEE treatment61,62 Elevated HIF-1α expression implies cellular adaptation to metabolic stress and regulation of cell death under adverse conditions.62,63 Concurrently, suppression of Bcl-2 supports mitochondrial destabilization, tipping the balance toward cell death, a process that has been reported to occur even in the absence of caspase activation.51,62,64,65 In the present study, these molecular changes, together with reduced caspase-3 expression and unchanged AIF levels, suggest that PNEE induces a regulated cell death that does not fully conform to the classical caspase-dependent pathway. The predominance of early apoptosis in MCF-7 cells may also be influenced by the well-documented deficiency of functional caspase-3 in this cell line,66 whereas BT-474 cells progressed more rapidly to late apoptosis. Nevertheless, the precise molecular mechanisms underlying these subtype-specific responses require further investigation.
Collectively, our findings provide new evidence for the anticancer activity of PNEE by demonstrating its ability to inhibit cell proliferation, disrupt oncogenic signaling, modulate metastasis-associated markers, and induce regulated apoptosis-like cell death in BT-474 breast cancer cells. Comparation with MCF-7 luminal A cells further demonstrated subtype-specific differences in apoptotic responses, emphasizing the influence of molecular context on the cellular effects of PNEE. These actions, together with its favorable selectivity toward cancer cells over non-cancerous 3T3-L1 cells, support the potential of PNEE as a promising candidate for further preclinical development. Nevertheless, the present study evaluated acute cellular responses following 24-hour treatment and did not address long-term therapeutic effects. Although a standard chemotherapeutic agent was not included, future studies should compare PNEE with established anticancer drugs, such as cisplatin, paclitaxel, or doxorubicin. Further investigations should also focus on identifying and quantitatively the bioactive compounds responsible for these activities, elucidating the molecular mechanisms underlying the observed cell death responses, combining PNEE with standard anticancer agents, and evaluating the efficacy and safety of PNEE in appropriate in vivo models.
Conclusion
This study provides the new insights into the anticancer potential of PNEE in BT-474 luminal B breast cancer cells. PNEE exhibited selective cytotoxic and antiproliferative effects against BT-474 luminal B breast cancer cells while showing minimal toxicity toward normal fibroblast cells under the experimental conditions tested. PNEE arrested cell cycle progression at the S-phase and modulated several cancer-associated molecular markers, including EGFR, PI3K/AKT pathway components, cyclin A, MMP-9, HIF-1α, and BCl-2. In addition, PNEE triggered a regulated apoptosis-like cell death phenotype that did not conform to the canonical caspase-3-dependent mechanism. Comparative analysis with MCF-7 luminal A cells further demonstrated subtype-specific differences in apoptotic responses, suggesting that the cellular effects of PNEE may depend on the molecular characteristics of breast cancer subtypes. These findings support the further investigation potential of PNEE as a source of bioactive compounds for breast cancer therapy. Future studies should focus on elucidating the underlying cell death mechanism and evaluating the efficacy and safety of PNEE in appropriate in vivo models, and determining its optimal dosage and potential interactions with existing therapies.
Abbreviation
AlCl3, aluminum chloride; AIF, Apoptosis-Inducing Factor; ATCC, American Type Culture Collection;BiONO3·H2O, bismuth subnitrate; CAM, Complementary and Alternative Medicine; CBD, Convention on Biological Diversity; CE, Catechin Equivalent; C7H6O5, gallic acid; C15H10O7, quercetin; CHCl3, chloroform; CH3COOH, glacial acetic acid; DMSO, dimethyl sulfoxide; DPPH, diphenyl-1-picrylhydrazyl;ELISA, Enzyme-Linked Immunosorbant Assay; EndoG, endonuclease G;FBS, fetal bovine serum; FeCl3, iron (III) chloride; FSC, forward scatter; GAE, Gallic Acid Equivalent; GLOBOCAN, Global Cancer Observatory; H2SO 3, sulfuric acid; KI, potassium iodide; Mg, magnesium powder; MTT, 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide; Na2CO3, sodium carbonate; NaNO2, sodium nitrite; NaOH, sodium hydroxide; NH3, ammonia solution; PBS, phosphate buffered saline; PI, Propidium Iodide; PI3K/AKT, Phosphoinositide 3-kinase/Protein kinase B; PFPE, piperine-free P. nigrum fruit extract; PNEE, P. nigrum ethanolic extract; PVPP, polyvinylpolypyrrolidone; QE, Quercetin Equivalent; RIPA, radioimmunoprecipitation assay; ROS, Reactive Oxygen Species; RPMI, Roswell Park Memorial Institute; RT-qPCR, Reverse Transcriptase Quantitative Polymerase Chain Reaction; RTK, receptor tyrosine kinase; SEM, standard error of the mean; SSC, side scatter; TAE, Tannic Acid Equivalent.
Acknowledgments
The authors would like to thank the Biomedical laboratory, Faculty of Medicine and the Chemistry Services Laboratory, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, for their technical assistance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Faculty of Medicine, Universitas Padjadjaran (Grant No. 5317/UN6.C/PT.02/2025). The publication of this article was funded by the Indonesian Endowment Fund for Education (LPDP), on behalf of the Indonesian Ministry of Higher Education, Science and Technology and managed under the EQUITY Program (Contract No. 4303/B3/DT.03.08/2025 and 3927/UN6.RKT/HK.07.00/2025).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gu H, Wang R, Beeraka NM, et al. Global burden and trends of breast cancer: GLOBOCAN 2022 estimates of incidence and mortality in 185 countries. Chin Med J. 2026;139(3):404–16. doi:10.1097/CM9.0000000000003921
2. Orrantia-Borunda E, Anchondo-Nuñez P, Acuña-Aguilar LE, Gómez-Valles FO, Ramírez-Valdespino CA. Subtypes of Breast Cancer. In: Mayrovitz HN, editor. Breast Cancer. Brisbane (AU): Exon Publications; 2022.
3. Katta B, Vijayakumar C, Dutta S, et al. The incidence and severity of patient-reported side effects of chemotherapy in routine clinical care: a Prospective Observational Study. Cureus. 2023;15(4):e38301. doi:10.7759/cureus.38301
4. Naeem A, Hu P, Yang M, et al. Natural products as anticancer agents: current status and future perspectives. Molecules. 2022;27(23):8367. doi:10.3390/molecules27238367
5. Rizki Budiman M, Wiraswati HL, Rezano A. Purple sweet potato phytochemicals: potential chemo-preventive and anticancer activities. Open Access Maced J Med Sci. 2021;9(F):288–298. doi:10.3889/oamjms.2021.6784
6. Azzam MH, Wiraswati FN, L H. The anticancer effect of phytochemicals and potential of breynia cernua: an overview. Biomed Pharmacol J. 2022;15(4):2259–2278.
7. da Silva FC, Brandao DC, Ferreira EA, et al. Tailoring potential natural compounds for the treatment of luminal breast cancer. Pharmaceuticals; 2023: 16.
8. Erdem E, Sezer Efe Y, Bayat M, et al. Complementary and alternative medicine methods used among Turkish pediatric oncology patients. J Pediatr Nurs. 2020;52:e103–e107. doi:10.1016/j.pedn.2019.11.013
9. Gottschling S, Meyer S, Längler A, et al. Differences in use of complementary and alternative medicine between children and adolescents with cancer in Germany: a population based survey. Pediatr Blood Cancer. 2014;61(3):488–492. doi:10.1002/pbc.24769
10. Wanchai A, Armer JM, Stewart BR. Complementary and alternative medicine use among women with breast cancer: a systematic review. Clin J Oncol Nurs. 2010;14(4):E45–55. doi:10.1188/10.CJON.E45-E55
11. Hasanah SN, Widowati L. Jamu pada pasien tumor/kanker sebagai terapi komplementer. Jurnal Kefarmasian Indonesia. 2016;6(1):49–59. doi:10.22435/jki.v6i1.5469.49-59
12. Wiraswati HL, Ekawardhani S, Rohmawaty E, et al. Antioxidant, antiinflammation, and antifibrotic activity of ciplukan (Physalis angulata L). Extract J Inflammat Res. 2024;17(null):6297–6306. doi:10.2147/JIR.S470318
13. Tariq A, Sadia S, Pan K, et al. A systematic review on ethnomedicines of anti-cancer plants. Phytother Res. 2017;31(2):202–264. doi:10.1002/ptr.5751
14. Yadav N, Deshmukh R, Mazumder R. A comprehensive review on the use of traditional Chinese medicine for cancer treatment. Pharmacol Res. 2024;11:100423.
15. Jung T, Cheon C. Synergistic and additive effects of herbal medicines in combination with chemotherapeutics: a scoping review. Integr Cancer Ther. 2024;23:15347354241259416. doi:10.1177/15347354241259416
16. Cheon C. Synergistic effects of herbal medicines and anticancer drugs: a protocol for systematic review and meta-analysis. Medicine. 2021;100(46):e27918. doi:10.1097/MD.0000000000027918
17. Fasinu PS, Rapp GK. Herbal interaction with chemotherapeutic drugs-a focus on clinically significant findings. Front Oncol. 2019;9:1356. doi:10.3389/fonc.2019.01356
18. Chaveerach A, Mokkamul P, Sudmoon R, et al. Ethnobotany of the genus piper (Piperaceae) in Thailand. Ethnobotany Res Applicat. 2006;4:223–231. doi:10.17348/era.4.0.223-231
19. Bezerra DP, Pessoa C, de Moraes MO, et al. Overview of the therapeutic potential of piplartine (piperlongumine). Eur J Pharm Sci. 2013;48(3):453–463. doi:10.1016/j.ejps.2012.12.003
20. Mitra S, Anand U, Jha NK, et al. Anticancer applications and pharmacological properties of piperidine and piperine: a comprehensive review on molecular mechanisms and therapeutic perspectives. Front Pharmacol. 2021;12:772418. doi:10.3389/fphar.2021.772418
21. Wiraswati HL, Ma’ruf IF, Sharifi-Rad J, et al. Piperine: an emerging biofactor with anticancer efficacy and therapeutic potential. BioFactors. 2025;51(1):e2134. doi:10.1002/biof.2134
22. Wiraswati H, Rohmawaty E, Ramadhanti J, et al. Anticancer potential of secondary metabolites of Piper nigrum against MOA B, AIF, CYP and EGFR proteins: in silico study. J Pharm Pharmacogn Res. 2025;13:816–835. doi:10.56499/jppres24.2138_13.3.816
23. Ahmad N, Fazal H, Abbasi BH, et al. Biological role of Piper nigrum L. (Black pepper): a review. Asian Pac J Trop Biomed. 2012;2(3):S1945–S1953. doi:10.1016/S2221-1691(12)60524-3
24. Barata LM, Andrade EH, Ramos AR, et al. Secondary metabolic profile as a tool for distinction and characterization of cultivars of black pepper (Piper nigrum L.) cultivated in Pará State. Brazil Int J Mol Sci. 2021;22(2).
25. Luo Y, Liu H-M, Su M-B, et al. Synthesis and biological evaluation of piperamide analogues as HDAC inhibitors. Bioorg Med Chem Lett. 2011;21(16):4844–4846. doi:10.1016/j.bmcl.2011.06.046
26. Wiraswati HL, Ma’ruf IF, Hidayati NA, et al. Harnessing the anticancer potential of Piper nigrum: a synergistic approach to chemotherapy enhancement and reduced side effects. Discover Oncol. 2025;16(1):10. doi:10.1007/s12672-024-01716-4
27. Deng Y, Sriwiriyajan S, Tedasen A, et al. Anti-cancer effects of Piper nigrum via inducing multiple molecular signaling in vivo and in vitro. J Ethnopharmacol. 2016;188:87–95. doi:10.1016/j.jep.2016.04.047
28. de Souza Grinevicius VM, Kviecinski MR, Santos Mota NSR, et al. Piper nigrum ethanolic extract rich in piperamides causes ROS overproduction, oxidative damage in DNA leading to cell cycle arrest and apoptosis in cancer cells. J Ethnopharmacol. 2016;189:139–147. doi:10.1016/j.jep.2016.05.020
29. Saetang J, Tedasen A, Sangkhathat S, et al. Low piperine fractional Piper nigrum extract enhanced the antitumor immunity via regulating the Th1/Th2/Treg cell subsets on NMU-Induced tumorigenesis rats. Planta Med. 2022;88(7):527–537. doi:10.1055/a-1458-5646
30. Szostakowska M, Trębińska-Stryjewska A, Grzybowska EA, et al. Resistance to endocrine therapy in breast cancer: molecular mechanisms and future goals. Breast Cancer Res Treat. 2019;173(3):489–497. doi:10.1007/s10549-018-5023-4
31. Li HW, Zou T-B, Jia Q, et al. Anticancer effects of morin-7-sulphate sodium, a flavonoid derivative, in mouse melanoma cells. Biomed Pharmacother. 2016;84:909–916. doi:10.1016/j.biopha.2016.10.001
32. Mittendorf EA, Liu Y, Tucker SL, et al. A novel interaction between HER2/neu and cyclin E in breast cancer. Oncogene. 2010;29(27):3896–3907. doi:10.1038/onc.2010.151
33. Hsu YC, Chen MJ, Huang TY. Inducement of mitosis delay by cucurbitacin E, a novel tetracyclic triterpene from climbing stem of Cucumis melo L. through GADD45γ in human brain malignant glioma (GBM) 8401 cells. Cell Death Dis. 2014;5(2):e1087. doi:10.1038/cddis.2014.22
34. Zhang J, Zhang -L-L, Shen L, et al. Regulation of AKT gene expression by cisplatin. Oncol Lett. 2013;5(3):756–760. doi:10.3892/ol.2013.1132
35. Ku M, Kang M, Suh J-S, et al. Effects for sequential treatment of siakt and paclitaxel on gastric cancer cell lines. Int J Med Sci. 2016;13(9):708–716. doi:10.7150/ijms.15501
36. Aslan C, Maralbashi S, Kahroba H, et al. Docosahexaenoic acid (DHA) inhibits pro-angiogenic effects of breast cancer cells via down-regulating cellular and exosomal expression of angiogenic genes and microRNAs. Life Sci. 2020;258:118094. doi:10.1016/j.lfs.2020.118094
37. Yan W, Ma X, Zhao X, et al. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des Devel Ther. 2018;12:3961–3972. doi:10.2147/DDDT.S181939
38. Mason JK, Fu M, Chen J, et al. Flaxseed oil enhances the effectiveness of trastuzumab in reducing the growth of HER2-overexpressing human breast tumors (BT-474). J Nutr Biochem. 2015;26(1):16–23. doi:10.1016/j.jnutbio.2014.08.001
39. Alrehaili AA, Gharib AF, Karam RA, et al. Clinical significance of plasma MMP-2 and MMP-9 levels as biomarkers for tumor expression in breast cancer patients in Egypt. Mol Biol Rep. 2020;47(2):1153–1160. doi:10.1007/s11033-019-05216-5
40. Lou J, Zhou Y, Feng Z, et al. Caspase-Independent regulated necrosis pathways as potential targets in cancer management. Front Oncol. 2021;10:616952.
41. Tang JY, Ou-Yang F, Hou M-F, et al. Oxidative stress-modulating drugs have preferential anticancer effects - involving the regulation of apoptosis, DNA damage, endoplasmic reticulum stress, autophagy, metabolism, and migration. Semin Cancer Biol. 2019;58:109–117. doi:10.1016/j.semcancer.2018.08.010
42. Snezhkina AV, Kudryavtseva AV, Kardymon OL, et al. ROS generation and antioxidant defense systems in normal and malignant cells. Oxid Med Cell Longev. 2019;2019:6175804. doi:10.1155/2019/6175804
43. Rajendran P, Maheshwari U, Muthukrishnan A, et al. Myricetin: versatile plant based flavonoid for cancer treatment by inducing cell cycle arrest and ROS-reliant mitochondria-facilitated apoptosis in A549 lung cancer cells and in silico prediction. Mol Cell Biochem. 2021;476(1):57–68. doi:10.1007/s11010-020-03885-6
44. Fan JJ, Hsu W-H, Lee K-H, et al. Dietary flavonoids luteolin and quercetin inhibit migration and invasion of squamous carcinoma through reduction of Src/Stat3/S100A7 signaling. Antioxidants. 2019;8(11):557. doi:10.3390/antiox8110557
45. Vrhovac Madunić I, Madunić J, Antunović M, et al. Apigenin, a dietary flavonoid, induces apoptosis, DNA damage, and oxidative stress in human breast cancer MCF-7 and MDA MB-231 cells. Naunyn-Schmiedeberg’s Arch Pharmacol. 2018;391(5):537–550. doi:10.1007/s00210-018-1486-4
46. Pal HC, Katiyar SK. Cryptolepine, a plant alkaloid, inhibits the growth of non-melanoma skin cancer cells through inhibition of topoisomerase and induction of DNA damage. Molecules; 2016:21.
47. Grebinyk A, Prylutska S, Grebinyk S, et al. Antitumor efficiency of the natural alkaloid berberine complexed with C60 fullerene in Lewis lung carcinoma in vitro and in vivo. Cancer Nanotechnol. 2021;12(1):24. doi:10.1186/s12645-021-00096-6
48. Al-Rashed S, Baker A, Ahmad SS, et al. Vincamine, a safe natural alkaloid, represents a novel anticancer agent. Bioorg Chem. 2021;107:104626. doi:10.1016/j.bioorg.2021.104626
49. Delaney LM, Farias N, Ghassemi Rad J, et al. The natural alkaloid piperlongumine inhibits metastatic activity and epithelial-to-mesenchymal transition of triple-negative mammary carcinoma cells. Nutr Cancer. 2021;73(11–12):2397–2410. doi:10.1080/01635581.2020.1825755
50. Wang L, Liu L, Shi Y, et al. Berberine induces caspase-independent cell death in colon tumor cells through activation of apoptosis-inducing factor. PLoS One. 2012;7(5):e36418. doi:10.1371/journal.pone.0036418
51. Gupta G, Samuel VP, M RM, Rani B, et al. Caspase-independent cell death in lung cancer: from mechanisms to clinical applications. Naunyn-Schmiedeberg’s Arch Pharmacol. 2025;398.
52. Wee P, Wang Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers. 2017;9(5). doi:10.3390/cancers9050052
53. Buranrat BS, Laddawan P, Auemduan K, Kukongviriyapan, Veerapol V. Anticancer effects of Piper nigrum extract against cholangiocarcinoma cells. Pharmacogn Mag. 2022;18(77):160. doi:10.4103/pm.pm_521_21
54. Buranrat B. Proliferative inhibition, apoptotic induction, migratory suppression of Piper nigrum extract on hela cervical cell line. Indian J Pharm Sci. 2022;84(5):1210–1217. doi:10.36468/pharmaceutical-sciences.1016
55. Jeong JH, Ryu J-H, Lee HJ. In vitro inhibition of Piper nigrum and piperine on growth, migration, and invasion of PANC-1 human pancreatic cancer cells. Nat Prod Commun. 2021;16(11):1934578X211057694. doi:10.1177/1934578X211057694
56. Kiranmayee M, Rajesh N, Vidya Vani M, et al. Green synthesis of Piper nigrum copper-based nanoparticles: in silico study and ADMET analysis to assess their antioxidant, antibacterial, and cytotoxic effects. Front Chem. 2023;1:1218588.
57. Elekofehinti OO, Adetoyi IR, Popoola HO, et al. Discovery of potential epidermal growth factor receptor inhibitors from black pepper for the treatment of lung cancer: an in-silico approach. Silico Pharmacol. 2024;12(1):28. doi:10.1007/s40203-024-00197-1
58. Chen H, Sheng H, Zhao Y, et al. Piperine inhibits cell proliferation and induces apoptosis of human gastric cancer cells by downregulating phosphatidylinositol 3-kinase (PI3K)/Akt pathway. Med Sci Monit. 2020;26:e928403. doi:10.12659/MSM.928403
59. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
60. Dirsch VM, Kirschke SO, Estermeier M, et al. Apoptosis signaling triggered by the marine alkaloid ascididemin is routed via caspase-2 and JNK to mitochondria. Oncogene. 2004;23(8):1586–1593. doi:10.1038/sj.onc.1207281
61. Yang Z, Zhao T-Z, Zou Y-J, et al. Hypoxia induces autophagic cell death through hypoxia-inducible Factor 1α in Microglia. PLoS One. 2014;9(5):e96509. doi:10.1371/journal.pone.0096509
62. Wiraswati HL, Hangen E, Sanz AB, et al. Apoptosis inducing factor (AIF) mediates lethal redox stress induced by menadione. Oncotarget. 2016;7(47):76496–76507. doi:10.18632/oncotarget.12562
63. Zhao X, Liu L, Li R, et al. Hypoxia-Inducible factor 1-α (HIF-1α) induces apoptosis of human uterosacral ligament fibroblasts through the death receptor and mitochondrial pathways. Med Sci Monit. 2018;24:8722–8733. doi:10.12659/MSM.913384
64. Chipuk JE, Green DR. How do BCL-2 proteins induce mitochondrial outer membrane permeabilization? Trends Cell Biol. 2008;18(4):157–164. doi:10.1016/j.tcb.2008.01.007
65. Qian S, Wei Z, Yang W, et al. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front Oncol. 2022;12:985363. doi:10.3389/fonc.2022.985363
66. Tian T. MCF-7 cells lack the expression of Caspase-3. Int J Biol Macromol. 2023;231:123310. doi:10.1016/j.ijbiomac.2023.123310
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.