Back to Journals » Infection and Drug Resistance » Volume 13
Phenotypic and Molecular Characteristics of Community-Associated Staphylococcus aureus Infection in Neonates
Authors Dong Q, Liu Y, Li W, Chen M, Li W, Wang X, Fu J, Ye X ![]()
Received 2 October 2020
Accepted for publication 27 November 2020
Published 22 December 2020 Volume 2020:13 Pages 4589—4600
DOI https://doi.org/10.2147/IDR.S284781
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Qian Dong,1 Yanling Liu,1 Wenhui Li,1 Minqi Chen,1 Wenyu Li,1 Xulin Wang,1 Jinjian Fu,2 Xiaohua Ye1
1School of Public Health, Guangdong Pharmaceutical University, Guangzhou, People’s Republic of China; 2Department of Prevention and Health Care, Liuzhou Maternity and Child Health Care Hospital, Liuzhou, People’s Republic of China
Correspondence: Jinjian Fu; Xiaohua Ye Email [email protected]; [email protected]
Background: Staphylococcus aureus is an important human pathogen causing a wide variety of diseases ranging from mild skin infections to life-threatening bacteremia. This study aimed to provide insight into the phenotype–genotype characteristics of S. aureus isolated from neonates.
Methods: A hospital-based retrospective study was conducted to recruit neonatal patients with community-associated S. aureus infection. All 92 S. aureus clinical isolates were tested for antimicrobial susceptibility, resistance genes, virulence genes, and multilocus sequence typing.
Results: A total of 92 S. aureus isolates were included in this study, including 29 (31.5%) MRSA isolates. Most S. aureus isolates were susceptible to gentamicin, and most methicillin-resistant S. aureus (MRSA) isolates were susceptible to vancomycin and linezolid. The predominant S. aureus genotypes were CC188 (ST188), CC59 (ST338), CC5 (ST5), CC88 (ST8), and CC6 (ST6), with CC59 (ST338) as the most prevalent MRSA. We observed a significantly corresponding relationship between clonal complexes and resistance patterns (such as CC45/CC59/CC121/CC630 associated with multidrug resistance) or the number of virulence genes (such as CC1/CC5/CC45/CC59/CC88 associated with harboring 5– 7 virulence genes, and CC22/CC25/CC121 associated with carrying 8– 13 genes). Moreover, both univariate and multivariate analyses indicated that invasive S. aureus isolates were significantly associated with resistance to several classes of antibiotics and carrying specific virulence factors.
Conclusion: Our findings provide insight into phenotype–genotype characteristics for S. aureus isolates so as to provide new ideas for differentiating potential genetic backgrounds and monitoring new epidemiologic trends.
Keywords: Staphylococcus aureus, antimicrobial susceptibility, molecular characterization, virulence genes
Introduction
Staphylococcus aureus (S. aureus) is an important human pathogen causing a wide variety of diseases ranging from mild skin infections to life-threatening bacteremia and contributes to both community-associated and healthcare-associated infections with a high clinical burden for infants worldwide.1 Nearly 120,000 S. aureus bloodstream infections and 20,000 associated deaths occurred in the United States in 2017, indicating that S. aureus infections account for significant morbidity and mortality.2 A multicenter study in China showed that the incidence of community-associated S. aureus from skin and soft tissue infections was significantly higher in pediatric patients than in adult patients, suggesting that S. aureus is an important causative agent in children.3 Moreover, several studies have reported that outbreaks and transmission events due to S. aureus and methicillin-resistant S. aureus (MRSA) occurred in neonatal intensive care units (NICUs).4–6 Therefore, it is urgent to study phenotypic and genotypic characteristics of S. aureus in neonates, which may promote an increased understanding of its genetic backgrounds and pathogenesis.
The incidence of S. aureus (especially for MRSA) infection and colonization have been explored for neonates across the globe.7–10 For example, MRSA colonization was detected in 6.7% of infants, and MRSA infection occurred in 2.0% of infants in NICUs of the US.7 Similarly, rates of MRSA colonization and infection were found to be 5.2% and 2.6% in Chinese NICUs, respectively.8 These findings suggest that active surveillance may detect MRSA affected neonates earlier and limit nosocomial spread. To note, previous studies have mainly focused on MRSA isolates in neonates or infants,6,9,11,12 whereas little is known about methicillin-sensitive S. aureus (MSSA). In terms of virulence and pathogenicity, S. aureus infection is characterized by virulence factors and antibiotic resistance.13 However, limited data on phenotypic and molecular characteristics of neonatal S. aureus are available for China, and the potential relationship between clonal complexes (CCs) and phenotype–genotype characteristics of S. aureus is still unclear. Therefore, we undertook a hospital-based retrospective study of neonatal S. aureus infection in Liuzhou, China. The primary purpose of this study was to describe antimicrobial susceptibility, virulence genes, and molecular typing of both MRSA and MSSA isolates. The second purpose was to reveal the potential corresponding relationships between CCs and phenotype–genotype characteristics of S. aureus isolates.
Materials and Methods
Patient Enrollment and Clinical Data Collection
This hospital-based retrospective study was conducted from April 2014 to May 2017 at the sole tertiary children’s hospital in Liuzhou, China. Our children’s hospital has around 10,000–20,000 admissions of children annually. The target population was neonates ≤28 days of age and with community-associated S. aureus infections. The eligibility criteria for community-associated S. aureus infections included: (1) a culture sample was obtained within 48h of hospitalization or during an outpatient visit; (2) the patient had no history of known risk factors for S. aureus infections in the past year; and (3) the patient had no history of medical facility visits in the past year.14 Infections in which any positive culture was obtained from cerebrospinal fluid (CSF), blood, sterile fluid, or an abscess were considered to be invasive. Infections in which all positive cultures were obtained from the trachea, urine, conjunctiva, or a wound were considered to be non-invasive. We used a case report form for the collection of demographic information (sex and age), clinical symptoms, types of infection, antibiotic use, and clinical outcomes.
Bacterial Isolation and Identification
Specimens were collected from infection sites of each neonatal patient enrolled and cultured on blood agar. To avoid overrepresentation, we included only the first isolate from each patient. Preliminary S. aureus identification was performed based on a combination of bacterial morphology, Gram staining, hemolysis, catalase test, and tube coagulase test. Then, Polymerase Chain Reaction (PCR) assays were used to test the staphylococci 16SrRNA, nuc, and mecA genes (Supplementary Table S1).15 All S. aureus isolates carried the 16S rRNA and nuc genes. The mecA-positive or cefoxitin-resistant S. aureus was identified as MRSA. So MRSA included genotypic (mecA-positive) and phenotypic (cefoxitin-resistant) MRSA isolates.
Antimicrobial Susceptibility Tests
The minimum inhibitory concentrations of antimicrobial agents were determined according to the Clinical and Laboratory Standards Institute.16 The following drugs were tested: penicillin, cefoxitin, erythromycin, clindamycin, tetracycline, trimethoprim-sulfamethoxazole, rifampin, chloramphenicol, ciprofloxacin, gentamicin, vancomycin, and linezolid. S. aureus ATCC 29,213 was used as a control strain. Multidrug-resistant (MDR) S. aureus (MDRSA) was defined as being non-susceptible to ≥1 agent in ≥3 antimicrobial categories.17 The resistance pattern was categorized into three groups: susceptible to all antibiotics, non-MDR (non-susceptible to 1–2 antimicrobial categories), and MDR (non-susceptible to ≥3 antimicrobial categories).
MLST Typing and SCCmec Analysis
For all S. aureus isolates, multilocus sequence typing (MLST) was analyzed by sequencing seven housekeeping genes.18 Sequence types (STs) were confirmed by comparing with the MLST database (http://saureus.mlst.net). Clonal complexes (CCs) were inferred using the eBURST algorithm, based on our data and a list of distinct ST types with six of the seven loci.19 MRSA isolates carrying the mecA gene were subjected to SCCmec typing, which is based on a set of multiplex PCR reactions (Supplementary Table S1).20
Antibiotic-Resistant Genes and Virulence Genes
For S. aureus isolates, specific PCR tests were used to detect the presence of tetracycline-resistant genes including tet (M) and tet (K), erythromycin-resistant genes including erm (A) and erm (C), and virulence genes including the Panton–Valentine leucocidin (pvl) toxin gene, the toxic shock syndrome toxin-1 gene (tsst1), the staphylococcal enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sej, sek, sel, sem, sen, seo, sep, seq, ser, and seu), the immune evasion cluster (IEC) genes (scn, chp, and sak), and the hemolysin genes (hla, hlb, and hld).21–26 The detailed information for PCR tests was presented in the supplementary Table S1.
Statistical Analyses
Briefly, a two-stage analysis process was employed to explore specific characteristics for invasive isolates. First, Pearson’s chi-squared test or Fisher’s exact test was used to test the differences in phenotype–genotype characteristics between invasive and non-invasive isolates. Second, the random forest model (machine learning approach) was used to evaluate the importance of each characteristic variable, which may be associated with invasive S. aureus isolates. Variable importance scores (mean decrease Gini, MDG) for characteristic variables were estimated and ranked in a descending order. A greater MDG value indicated that the degree of impurity arising from a category could be reduced the most by a single variable, suggesting an important variable. The random forest was implemented in the R package (Breiman and Cutler’s Random Forests for Classification and Regression, version 4.6–14).
In addition, we used the correspondence analysis to test two-dimensional relationships and the cluster analysis to explore high-dimensional relationships. Correspondence analysis provides a graphic and statistical method for exploring the internal relationship between categorical variables, so we used the correspondence analysis to explore potential corresponding relationships between CCs and phenotypic or genotypic characteristics of S. aureus isolates. Correspondence analyses were performed using STATA version 14.0 (StataCorp LP, College Station, TX, USA), and a two-sided P-value of <0.05 was defined as being of statistical significance. Moreover, the unweighted pair-group method with arithmetic means (UPGMA) cluster analysis was used to construct the UPGMA tree based on the mismatch matrix of alleles between ST types, simultaneously labeling with multiple molecular characteristics.27 The UPGMA tree was also marked and visualized by iTOL (https://itol.embl.de/) to compare the phenotype–genotype characteristics of S. aureus isolates.28
Ethics Statement
This study was approved by the Ethics Committee of Guangdong Pharmaceutical University and was conducted in accordance with the approved guidelines (No. 2015–22). Before participating, all parents or legal guardians of the neonates signed an informed consent form regarding the goals of the study and the willingness to participate. So this study complied with the Declaration of Helsinki.
Results
S. aureus and MRSA Detection
A total of 92 S. aureus isolates (13 invasive isolates and 79 non-invasive isolates) were identified from the study neonates during the study period, including 29 MRSA isolates. A total of 52 (56.5%) neonates were male, the average age was 15.8±7.4 days old, and the average length of hospital stay was 10.3±5.4 days. A few neonates (8 of 92 neonates, 8.7%) were transferred to the Neonatal Intensive Care Unit (NICU).
Antimicrobial Susceptibility Testing
All S. aureus isolates were susceptible to vancomycin, and most of S. aureus isolates were susceptible to chloramphenicol and linezolid (Table 1). But most isolates were resistant to penicillin (96.7%), and nearly half of the isolates were resistant to erythromycin (40.2%) and clindamycin (40.2%). Notably, 33 (35.9%) isolates were classified as MDRSA, with the most common resistance pattern being non-susceptible to clindamycin, erythromycin, and cefoxitin (9/33, 27.3%). In addition, one linezolid-resistant isolate was classified as MDRSA, which was non-susceptible to cefoxitin, erythromycin, clindamycin, tetracycline, and linezolid. For 26 phenotypic tetracycline-resistant isolates, 17 (65.4%) carried tet (K). But the tet (M) gene was absent from all phenotypic tetracycline-resistant isolates. For 37 phenotypic erythromycin-resistant isolates, 19 (51.4%) carried erm (C) and 1 (2.7%) carried erm (A).
 |
Table 1 Phenotypic and Molecular Characteristics of Invasive and Non-Invasive S. aureus Isolates |
Molecular Typing and Virulence Genes
Among all S. aureus isolates, we observed 28 unique STs belonging to 18 CCs (Figure 1), except for three non-typeable isolates. The most common S. aureus CCs were CC188, CC59, CC5, and CC88, with the predominant MRSA being CC59-MRSA. The most prevalent S. aureus STs were ST188, ST338, ST5, ST88, and ST6, with the predominant MRSA being ST338-MRSA. There were 29 MRSA isolates, including 9 genotypic (mecA-positive) and 20 phenotypic (cefoxitin-resistant) MRSA isolates. Among all 9 genotypic MRSA isolates, the predominant SCCmec type was SCCmec IVd (n=3), followed by SCCmec IVa (n=2), SCCmec I (n=2), and untypeable (n=2).
 |
Figure 1 Clonal dendrogram and detailed molecular characteristics of 89 ST-typeable S. aureus isolates. Abbreviations: ST, sequence type; CC, clonal complex; IEC, immune evasion cluster |
As to the IEC genes (Table 1), the predominant IEC gene was sak (95.7%), followed by chp (21.7%) and scn (14.1%). In terms of the enterotoxin genes, the most abundant gene was seg (22.8%), followed by sei (20.7%), sek (17.4%), seb (14.1%), sem (14.1%) and seo (12.0%). Note that several enterotoxin genes (including see, sel and ser) were absent from all S. aureus isolates. With regard to the hemolysin genes, hla was detected in 97.8%, hlb in 53.3%, and hld in 94.6% of S. aureus isolates. In addition, tsst1 was present in 24 (26.1%) isolates, and pvl was found in 16 (17.4%) isolates.
Relationships Between CCs and Phenotype–Genotype Characteristics
The first correspondence analysis indicated a significantly corresponding relationship between CCs and resistance patterns (χ2=56.00, P=0.003; Figures 2A and 3), suggesting that CC45/CC59/CC121/CC630 isolates were associated with being MDR, CC7/CC8/CC12/CC25/CC509 isolates were associated with being non-MDR, and CC1/CC5/CC6/CC22/CC88/CC188/CC398 isolates were associated with being susceptible to all antibiotics. The second correspondence analysis revealed a significantly corresponding relationship between CCs and the number of virulence genes (such as CC6/CC7/CC8/CC188/CC398/CC509 and harboring 1–4 virulence genes, CC1/CC5/CC12/CC45/CC59/CC88/CC630 and harboring 5–7 virulence genes, and CC22/CC25/CC121 and carrying 8–13 genes; χ2= 67.97, P<0.001; Figures 2B and 3). According to Figure 1, CC6/CC188 isolates were associated with harboring sak-hla-hld, CC59 (including ST338, ST59 and ST537) isolates were associated with harboring sak-seb-sek-hla-hlb-hld-pvl, CC5 isolates were associated with harboring sak-seg-sei-sej-sem-seo-hla-hlb–hld, and CC88 isolates were associated with harboring sak-sep-hla-hlb-hld-pvl. In addition, CC59/CC88/CC25 isolates were associated with carrying pvl, and CC630 and CC45 isolates were associated with carrying tsst1.
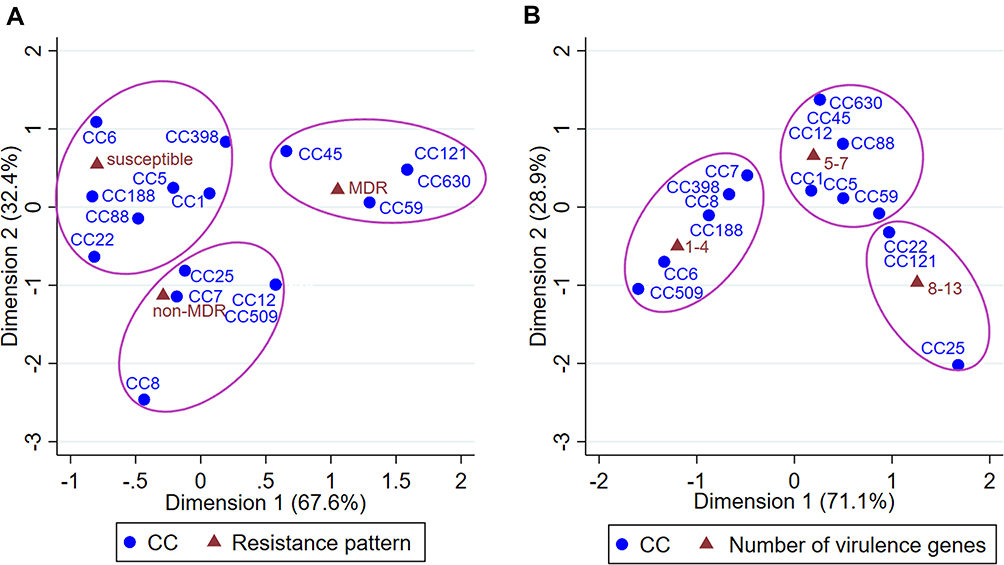 |
Figure 2 Correspondence analysis for testing the relationship between CCs and resistance patterns (A) or the number of virulence genes (B) of S. aureus isolates. Abbreviations: CCs, clonal complexes; MDR, multidrug-resistant. |
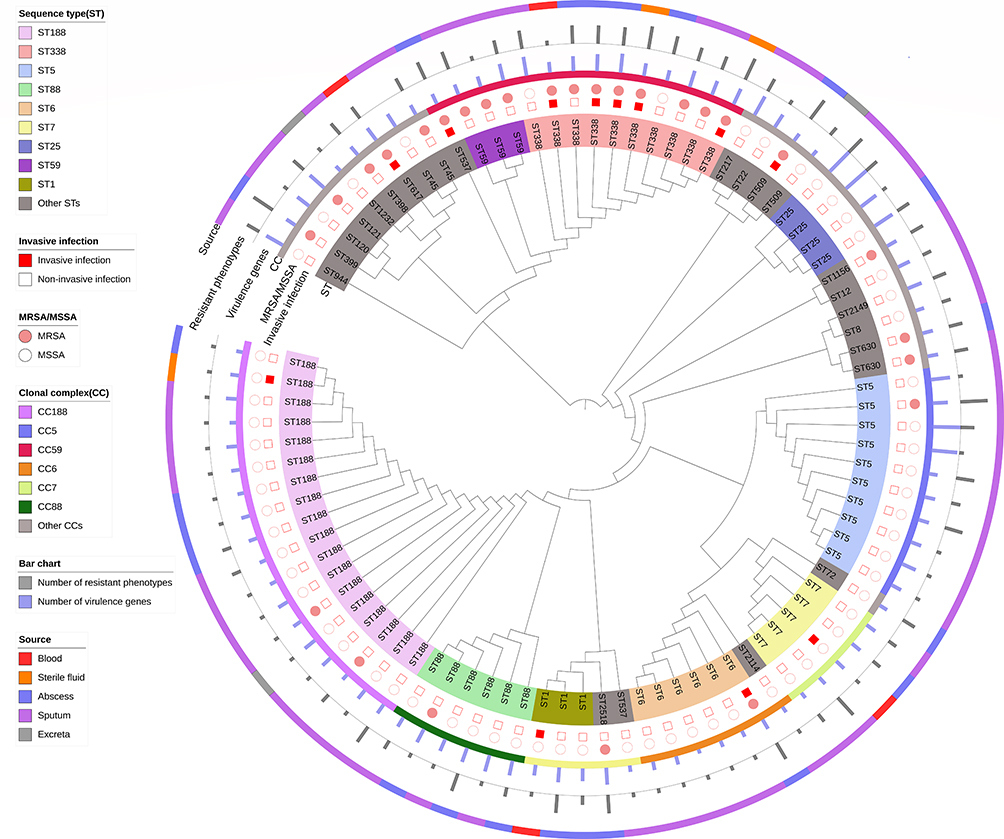 |
Figure 3 Clonal dendrogram for describing the relationship between genotypes and detailed phenotype–genotype characteristics of 89 ST-typeable S. aureus isolates. |
Differences in Phenotype–Genotype Characteristics Between Invasive and Non-Invasive S. aureus Isolates
When comparing the resistance characteristics between invasive and non-invasive isolates (Table 1), we observed that invasive S. aureus isolates were significantly associated with higher rates of resistance to cefoxitin (69.2% vs 22.8%, P=0.001) and tetracycline (53.8% vs 24.1%, P=0.027). When comparing the virulence genes between invasive and non-invasive isolates, invasive S. aureus isolates were significantly associated with carrying sak (76.9% vs 98.7%, P=0.008), seg (0.0% vs 26.6%, P=0.034), sei (0.0% vs 24.1%, P=0.047), sek (46.2% vs 12.7%, P=0.003), and pvl (38.5% vs 13.9%, P=0.031). Notably, invasive S. aureus isolates were significantly associated with CC59 (46.2% vs 10.1%, P=0.001). Among 13 invasive S. aureus isolates, the predominant linage was MRSA CC59 (ST388) carrying several virulence genes (Table 2).
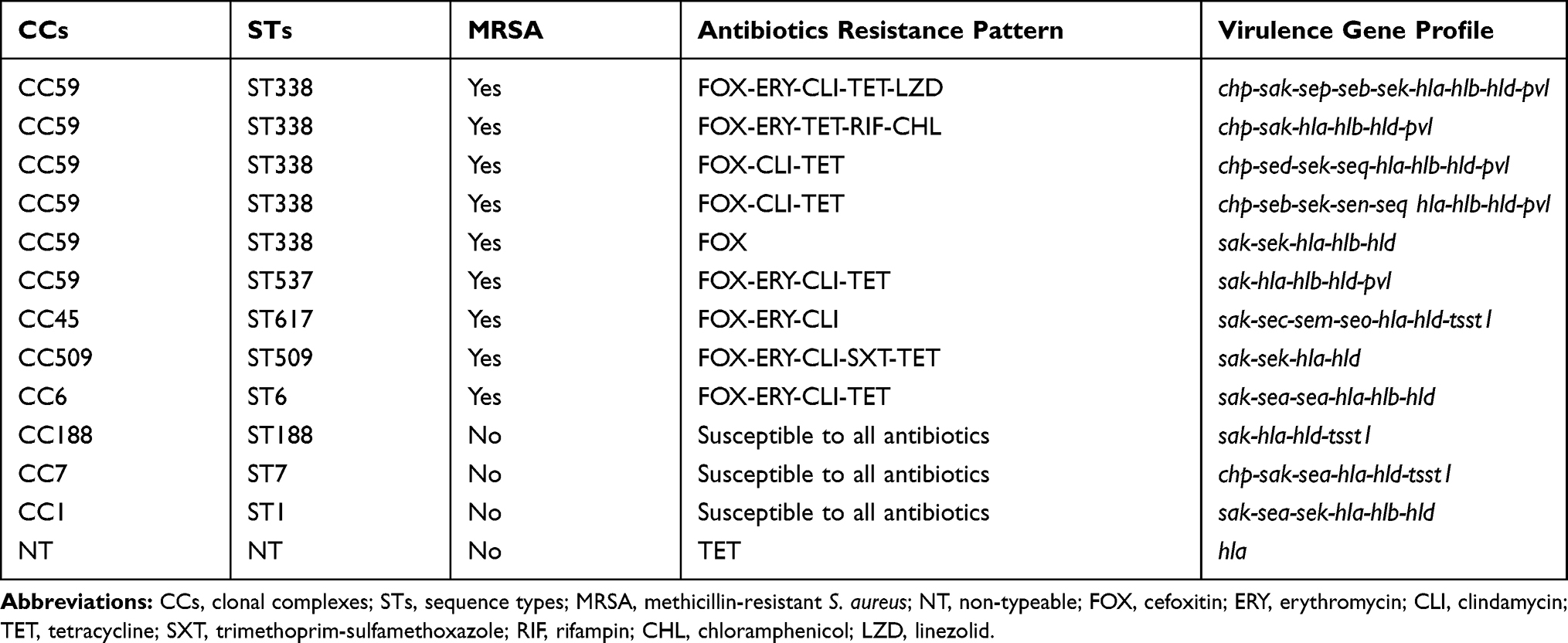 |
Table 2 Genotypic and Phenotypic Characteristics of 13 Invasive S. aureus Isolates |
In addition, random forest model was used to evaluate the importance of phenotypic and molecular characteristics influencing invasive S. aureus isolates (Figure 4). The results of random forest showed that the top-10 important variables were sak, cefoxitin, sek, CC59, tsst1, sea, pvl, chp, rifampin, and tetracycline.
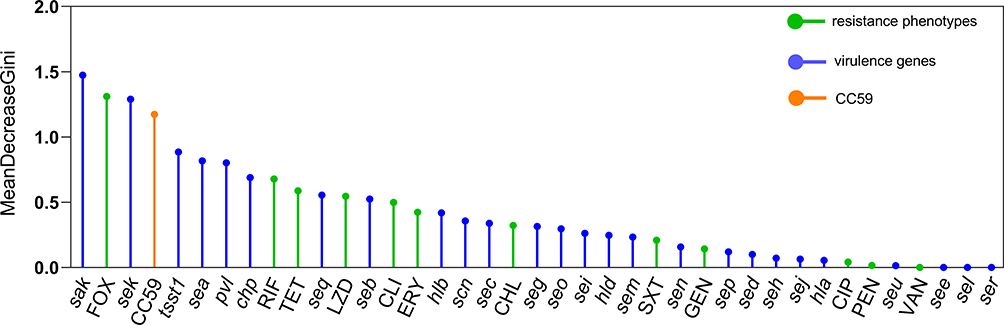 |
Figure 4 Importance rank of phenotypic and molecular characteristics influencing invasive S. aureus isolates. Abbreviations: FOX, cefoxitin; RIF, rifampin; TET, tetracycline; LZD, linezolid; CLI, clindamycin; ERY, erythromycin; CHL, chloramphenicol; GEN, gentamicin; CIP, ciprofloxacin; SXT, trimethoprim-sulfamethoxazole; PEN, penicillin; VAN, vancomycin. |
Discussion
The antimicrobial resistance of neonatal S. aureus and MRSA has become an important public health issue. The World Health Organization recommends penicillin in combination with gentamicin as first-line antibiotics for newborns with serious bacterial infection or sepsis.29 But, worryingly, we observed a high rate of resistance to penicillin (96.7%) among S. aureus isolates, which is in line with recent reports on neonatal S. aureus septicemia in China (96.0%) and Gambia (92.5%).30,31 Fortunately, the present study demonstrated low-level resistance to gentamicin (7.6%), which is similar to the results from China (0.8%) and Gambia (5.5%).31,32 These findings suggest that gentamicin is still the first choice for the treatment of S. aureus infections, especially for serious bacterial infection or sepsis. MRSA has become a significant cause of morbidity and mortality in infants worldwide. It is well known that vancomycin and linezolid are the few clinically effective agents against MRSA infections. In this study, most MRSA isolates were susceptible to vancomycin and linezolid, which is consistent with previous studies in China, Japan, and Colombia.12,30,33 These findings suggest that vancomycin and linezolid are still used as the first-line antibiotic treatment for MRSA infections. Notably, MDRSA infections are associated with worse health outcomes and higher expenditures. In this study, 35.9% S. aureus isolates were classified as MDRSA, with the most common resistance pattern being non-susceptible to clindamycin, erythromycin, and cefoxitin. Similar rates of MDRSA among S. aureus isolates were found in Guangzhou (30.5%), but significantly higher rates were observed in Shanghai (51.3%) and Iran (53.1%).32,34,35 Therefore, the prevalence and resistance patterns of MDRSA isolates must be monitored in prospective surveillance programs so as to guide rational use of antibiotics.
The results of molecular characteristics may broaden our insights into neonatal S. aureus and potential transmission mechanisms. In this study, the most common S. aureus genotypes were CC188 (ST188), CC59 (ST338), CC5 (ST5), CC88 (ST8), and CC6 (ST6), with CC59 (ST338) as the predominant MRSA, which is similar to the latest reports from China.32,34 However, the most common S. aureus CCs in Germany were CC15, CC8, CC25, and CC7, and the dominant CCs in the US were CC30, CC5, and CC8.36,37 These findings exhibit marked clonal diversity and regional differences in genotype distribution. Notably, the present study found that ST188 was the most prevalent in neonatal isolates, which was also reported as the major host-adapted lineage causing human and livestock infections in China.38 These findings suggest that the burden of S. aureus ST188 infections in humans has been increasing, especially in neonates. Besides, ST398/ST7/ST59 were the most prevalent S. aureus isolated from breast milk samples,39 and CC5/CC59/CC45 were the most predominant clones observed in both mother and neonate isolates,40 which were also found in our study. These findings may provide evidence for S. aureus transmission through breastfeeding and the birth canal.
The potential relationships between CCs and phenotype–genotype characteristics of S. aureus isolates may provide more implications for virulent properties. However, previous studies were limited to use descriptive tables or figures only, so this study builds on previous literature to use correspondence analysis to test the potential relationships between two-dimensional characteristic variables. Our correspondence analyses revealed significantly corresponding relationships between CCs and resistance patterns or the number of virulence genes, suggesting that CC45/CC59/CC121/CC630 isolates were associated with being MDR, CC6/CC188 were associated with carrying 1–4 virulence genes (sak-hla-hld), and CC1/CC5/CC45/CC59/CC88 were associated with harboring 5–7 virulence genes (sak-seb-sek-hla-hlb-hld-pvl for CC59, sak-seg-sei-sej-sem-seo-hla-hlb–hld for CC5, and sak-sep-hla-hlb-hld-pvl for CC88). Strikingly, the CC5/USA100, CC8/USA300, CC1/USA400, and CC45/USA600 lineages are frequently associated with life-threatening S. aureus infections and widespread in both the community and hospital settings,41 which are also found in neonatal infections in our study. Importantly, CC5/USA100 is a leading cause of invasive diseases and represents the majority of vancomycin-resistant/-intermediate isolates,41 and CC5 isolates in the present study also carried several virulence genes but were susceptible to all antibiotics. Moreover, multidrug-resistant CC5 isolates reported as livestock-associated MRSA have been identified in livestock and related workers in China and Germany.42,43 Conflicting outcomes obscure the identification of virulence and resistance of S. aureus isolates, which may lead to increase morbidity and mortality. Notably, the toxic shock syndrome toxin gene tsst1 was mainly observed in CC45 isolates, the pvl gene was mainly found in CC59 and CC88 isolates, and IEC genes were mainly found in CC88, which are consistent with the latest reports.32,41,44,45 These findings reveal the necessity of differentiating S. aureus clones based on multiple phenotype–genotype characteristics, which may provide new ideas for monitoring new epidemiologic trends.
S. aureus (especially for MRSA) is a major cause of community-associated and hospital-associated invasive infections globally. When exploring invasive S. aureus and MRSA isolates in depth, we observed a significant relationship between CC59 carrying multiple virulence factors and invasive MRSA infections, and the random forest also revealed that CC59 isolates play an important role in neonatal invasive infections. Similarly, other studies found an interaction between specific CCs and clinical outcomes.46,47 For example, there was a strong association between CC1/CC121/CC30 and invasive S. aureus infections.46 These findings suggest that distinct combinations of virulence genes may be present in specific invasive CCs. However, few studies have systematically compared phenotypic and molecular characteristics between invasive and non-invasive S. aureus isolates. This study adds to existing knowledge by providing insight into the genotypic and phenotypic characteristics for invasive isolates. The results from univariate analyses revealed that invasive S. aureus isolates were significantly associated with resistance to specific classes of antibiotics (including cefoxitin and tetracycline) and carrying or non-carrying specific virulence factors (including sak, seg, sei, sek, and pvl). In addition, the random forest indicated that invasive S. aureus isolates were associated with resistance to several classes of antibiotics and carrying specific virulence factors, which provides more evidence for revealing the specific phenotype–genotype characteristics for invasive S. aureus isolates. These findings provide implications for differentiating the genetic background and pathogenicity of invasive isolates based on multiple phenotype–genotype characteristics.
This study is a new attempt to reveal potential relationships between CCs and phenotype–genotype characteristics of S. aureus isolates by using both correspondence analysis and cluster analysis, which provides implications for monitoring new epidemiologic trends. Moreover, this study provides insight into the phenotypic and molecular characteristics for invasive S. aureus by using the multivariate random forest model, which provides implications for the genetic background and pathogenicity of invasive isolates. However, there are some limitations to this study. First, S. aureus carriage of healthcare workers could not be taken into consideration because of limited funding, so it could not determine whether exists the risk of neonatal S. aureus transmission by healthcare workers. Second, no samples were obtained from mothers, so it could not determine whether exists the risk of S. aureus transmission from mothers to neonates. Future multi-object cohort studies are required to better understand the transmission routes of neonatal infections.
Conclusion
This study demonstrated that most MRSA isolates were susceptible to vancomycin and linezolid, suggesting that vancomycin and linezolid are still used as the first-line antibiotic treatment for MRSA infections. Notably, this study contributes to the literature by revealing a significantly corresponding relationship between CCs and phenotype–genotype characteristics, which provides new ideas for monitoring new epidemiologic trends. Interestingly, we found that invasive S. aureus isolates were significantly associated with resistance to several classes of antibiotics and carrying specific virulence factors, which provides implications for differentiating the genetic background and pathogenicity of invasive S. aureus isolates based on multiple phenotype–genotype characteristics.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Nos. 81973069 and 81602901), the Guangdong Basic and Applied Basic Research Foundation (No. 2019A1515010915), and Liuzhou Science and Technology Bureau Project (No. 2018BJ10504). The funders had no role in study design, data collection and analysis, and interpretation of the data.
Disclosure
The authors have no conflicts of interest to disclose.
References
1. Tong SYC, Davis JS, Eichenberger E, Holland TL, Fowler VG. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28:603–661. doi:10.1128/CMR.00134-14
2. Kourtis AP, Hatfield K, Baggs J, Mu Y, See I, Epson E. Vital signs: epidemiology and recent trends in methicillin-resistant and in methicillin-susceptible Staphylococcus aureus bloodstream infections — united States. MMWR Morb Mortal Wkly Rep. 2019;68:214–219.
3. Liu Y, Xu Z, Yang Z, Sun J, Ma L. Characterization of community-associated Staphylococcus aureus from skin and soft-tissue infections: a multicenter study in China. Emerg Microbes Infect. 2016;5:e127. doi:10.1038/emi.2016.128
4. Harris SR, Cartwright EJP, Török ME, et al. Whole-genome sequencing for analysis of an outbreak of meticillin-resistant Staphylococcus aureus: a descriptive study. Lancet Infect Dis. 2013;13:130–136. doi:10.1016/S1473-3099(12)70268-2
5. Brown NM, Reacher M, Rice W, et al. An outbreak of meticillin-resistant Staphylococcus aureus colonization in a neonatal intensive care unit: use of a case-control study to investigate and control it and lessons learnt. J Hosp Infect. 2019;103(1):35–43. doi:10.1016/j.jhin.2019.05.009
6. Cheng VCC, Wong SC, Cao H, et al. Whole-genome sequencing data-based modeling for the investigation of an outbreak of community-associated methicillin-resistant Staphylococcus aureus in a neonatal intensive care unit in Hong Kong. Eur J Clin Microbiol Infect Dis. 2019;38:563–573.
7. Maraqa NF, Aigbivbalu L, Masnita-Iusan C, et al. Prevalence of and risk factors for methicillin-resistant Staphylococcus aureus colonization and infection among infants at a level III neonatal intensive care unit. Am J Infect Control. 2011;39:35–41. doi:10.1016/j.ajic.2010.07.013
8. de Lencastre H, Geng W, Qi Y, et al. Epidemiology of Staphylococcus aureus in neonates on admission to a Chinese neonatal intensive care unit. PLoS One. 2020;15:e0211845.
9. Washam M, Woltmann J, Haberman B, Haslam D, Staat MA. Risk factors for methicillin-resistant Staphylococcus aureus colonization in the neonatal intensive care unit: A systematic review and meta-analysis. Am J Infect Control. 2017;45:1388–1393. doi:10.1016/j.ajic.2017.06.021
10. Giuffrè M, Amodio E, Bonura C, et al. Methicillin-resistant Staphylococcus aureus nasal colonization in a level III neonatal intensive care unit: incidence and risk factors. Am J Infect Control. 2015;43:476–481. doi:10.1016/j.ajic.2014.12.027
11. Lavie-Nevo K, Srigley JA, Al-Rawahi GN, et al. Prevalence and clinical impact of methicillin-resistant Staphylococcus aureus colonization among infants at a level III neonatal intensive care unit. Am J Infect Control. 2019;47:1336–1339. doi:10.1016/j.ajic.2019.04.173
12. Murai T, Okazaki K, Kinoshita K, et al. Comparison of USA300 with non-USA300 methicillin-resistant Staphylococcus aureus in a neonatal intensive care unit. Int J Infect Dis. 2019;79:134–138. doi:10.1016/j.ijid.2018.11.020
13. Laabei M, Recker M, Rudkin JK, et al. Predicting the virulence of MRSA from its genome sequence. Genome Res. 2014;24:839–849. doi:10.1101/gr.165415.113
14. Centers for Disease Control and Prevention. Community-associated MRSA information for clinicians. infection control topics. centers for disease control and prevention. Atlanta, GA. Available from: http://www.cdc.gov/ncidod/dhqp/ar_mrsa_ca_clinicians.html#4.
15. Zhang K, Sparling J, Chow BL, et al. New quadriplex PCR assay for detection of methicillin and mupirocin resistance and simultaneous discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J Clin Microbiol. 2004;42:4947–4955. doi:10.1128/JCM.42.11.4947-4955.2004
16. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; 27th Informational Supplement. M100-S28. Wayne, PA: Clinical and Laboratory Standards Institute; 2018.
17. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18:268–281.
18. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones ofStaphylococcus aureus. J Clin Microbiol. 2000;38:1008–1015. doi:10.1128/JCM.38.3.1008-1015.2000
19. Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol. 2004;186:1518–1530. doi:10.1128/JB.186.5.1518-1530.2004
20. Zhang K, McClure J-A, Elsayed S, Louie T, Conly JM. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2005;43:5026–5033. doi:10.1128/JCM.43.10.5026-5033.2005
21. Lina G, Quaglia A, Reverdy ME, Leclercq R, Vandenesch F, Etienne J. Distribution of genes encoding resistance to macrolides, lincosamides, and streptogramins among staphylococci. Antimicrob Agents Chemother. 1999;43:1062–1066. doi:10.1128/AAC.43.5.1062
22. Aarestrup FM, Agerso Y, Gerner-Smidt P, Madsen M, Jensen LB. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn Microbiol Infect Dis. 2000;37:127–137. doi:10.1016/S0732-8893(00)00130-9
23. Wamel W, Rooijakkers SHM, Ruyken M, Kessel K, Strijp J. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J Bacteriol. 2006;188:1310–1315. doi:10.1128/JB.188.4.1310-1315.2006
24. Omoe K, Hu DL, Takahashi-Omoe H, Nakane A, Shinagawa K. Comprehensive analysis of classical and newly described staphylococcal superantigenic toxin genes in Staphylococcus aureus isolates. FEMS Microbiol Lett. 2005;246:191–198. doi:10.1016/j.femsle.2005.04.007
25. Jarraud S, Mougel C, Thioulouse J, et al. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun. 2002;70:631–641. doi:10.1128/IAI.70.2.631-641.2002
26. McClure J-A, Conly JM, Lau V, et al. Novel multiplex PCR assay for detection of the staphylococcal virulence marker Panton-Valentine leukocidin genes and simultaneous discrimination of methicillin-susceptible from -resistant staphylococci. J Clin Microbiol. 2006;44:1141–1144. doi:10.1128/JCM.44.3.1141-1144.2006
27. Jolley KA, Feil EJ, Chan MS, Maiden MC. Sequence type analysis and recombinational tests (START). Bioinformatics. 2001;17:1230–1231. doi:10.1093/bioinformatics/17.12.1230
28. Letunic I, Bork P. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47:W256–W9. doi:10.1093/nar/gkz239
29. Pocket book of hospital care for children: guidelines for the management of common childhood illnesses. 2nd edition. Geneva: World Health Organization; 2013. Available from: https://www.ncbi.nlm.nih.gov/books/NBK154447/.
30. Li JY, Chen SQ, Yan YY, et al. Identification and antimicrobial resistance of pathogens in neonatal septicemia in China—A meta-analysis. Int J Infect Dis. 2018;71:89–93. doi:10.1016/j.ijid.2018.04.794
31. Darboe S, Dobreniecki S, Jarju S, et al. Prevalence of Panton-Valentine Leukocidin (PVL) and antimicrobial resistance in community-acquired clinical Staphylococcus aureus in an urban gambian hospital: a 11-year period retrospective pilot study. Front Cell Infect Microbiol. 2019;9:170.
32. Liang B, Mai J, Liu Y, et al. Prevalence and characterization of Staphylococcus aureus isolated from women and children in Guangzhou, China. Front Microbiol. 2018;9:2790. doi:10.3389/fmicb.2018.02790
33. Salazar-Ospina L, Jiménez JN. High frequency of methicillin-susceptible and methicillin-resistant Staphylococcus aureus in children under 1 year old with skin and soft tissue infections. J Pediatr. 2018;94:380–389. doi:10.1016/j.jped.2017.06.020
34. Wang X, Liu Q, Zhang H, et al. Molecular characteristics of community-associated Staphylococcus aureus isolates from pediatric patients with bloodstream infections between 2012 and 2017 in Shanghai, China. Front Microbiol. 2018;9:1211. doi:10.3389/fmicb.2018.01211
35. Imani Fooladi AA, Ashrafi E, Tazandareh SG, et al. The distribution of pathogenic and toxigenic genes among MRSA and MSSA clinical isolates. Microb Pathog. 2015;81:60–66. doi:10.1016/j.micpath.2015.03.013
36. Deinhardt-Emmer S, Sachse S, Geraci J, et al. Virulence patterns of Staphylococcus aureus strains from nasopharyngeal colonization. J Hosp Infect. 2018;100:309–315. doi:10.1016/j.jhin.2017.12.011
37. Hulten KG, Mason EO, Lamberth LB, Forbes AR, Revell PA, Kaplan SL. Analysis of invasive community-acquired methicillin-susceptible staphylococcus aureus infections during a period of declining community acquired methicillin-resistant staphylococcus aureus infections at a large Children’s Hospital. Pediatr Infect Dis J. 2018;37:235–241. doi:10.1097/INF.0000000000001753
38. Wang Y, Liu Q, Liu Q, et al. Phylogenetic analysis and virulence determinant of the host-adapted Staphylococcus aureus lineage ST188 in China. Emerg Microbes Infect. 2018;7:45. doi:10.1038/s41426-018-0048-7
39. Li X, Zhou Y, Zhan X, Huang W, Wang X. Breast milk is a potential reservoir for livestock-associated staphylococcus aureus and community-associated Staphylococcus aureus in Shanghai, China. Front Microbiol. 2017;8:2639.
40. Lin J, Wu C, Yan C, et al. A prospective cohort study of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus carriage in neonates: the role of maternal carriage and phenotypic and molecular characteristics. Infect Drug Resist. 2018;11:555–565. doi:10.2147/IDR.S157522
41. King JM, Kulhankova K, Stach CS, Vu BG, Salgado-Pabón W. Phenotypes and Virulence among Staphylococcus aureus USA100, USA200, USA300, USA400, and USA600 Clonal Lineages. mSphere. 2016;1:e00071–16. doi:10.1128/mSphere.00071-16
42. Guo D, Liu Y, Han C, Chen Z, Ye X. Phenotypic and molecular characteristics of methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolated from pigs: implication for livestock-association markers and vaccine strategies. Infect Drug Resist. 2018;11:1299–1307. doi:10.2147/IDR.S173624
43. Köck R, Schaumburg F, Mellmann A, et al. Livestock-associated methicillin-resistant Staphylococcus aureus (MRSA) as causes of human infection and colonization in Germany. PLoS One. 2013;8:e55040.
44. Eibach D, Nagel M, Hogan B, et al. Nasal Carriage of Staphylococcus aureus among children in the ashanti region of ghana. PLoS One. 2017;12:e0170320. doi:10.1371/journal.pone.0170320
45. Mrochen DM, Grumann D, Schulz D, et al. Global spread of mouse-adapted Staphylococcus aureus lineages CC1, CC15, and CC88 among mouse breeding facilities. Int J Med Microbiol. 2018;308:598–606. doi:10.1016/j.ijmm.2017.11.006
46. Williamson DA, Ritchie SR, Roberts SA, et al. Clinical and molecular epidemiology of community-onset invasive Staphylococcus aureus infection in New Zealand children. Epidemiol Infect. 2014;142:1713–1721. doi:10.1017/S0950268814000053
47. Kurt K, Rasigade J-P, Laurent F, et al. Subpopulations of Staphylococcus aureus clonal complex 121 are associated with distinct clinical entities. PLoS One. 2013;8:e58155.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.