Back to Journals » Stem Cells and Cloning: Advances and Applications » Volume 14
Phenotypic and Functional Responses of Human Decidua Basalis Mesenchymal Stem/Stromal Cells to Lipopolysaccharide of Gram-Negative Bacteria
Authors Alshareef GH, Mohammed AE ![]() , Abumaree M, Basmaeil YS
, Abumaree M, Basmaeil YS ![]()
Received 14 August 2021
Accepted for publication 11 October 2021
Published 2 November 2021 Volume 2021:14 Pages 51—69
DOI https://doi.org/10.2147/SCCAA.S332952
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Bernard Binetruy
Ghofran Hasan Alshareef,1 Afrah E Mohammed,1 Mohammed Abumaree2,3 ,† Yasser S Basmaeil2
1Biology Department, College of Science, Princess Nourah Bint Abdulrahman University, Riyadh, 84428, Saudi Arabia; 2Stem Cell & Regenerative Medicine Department, King Abdullah International Medical Research Center, King Saud bin Abdulaziz University for Health Sciences, Ministry of National Guard Health Affairs, Riyadh, Saudi Arabia; 3College of Science and Health Professions, King Saud bin Abdulaziz University for Health Sciences, King Abdulaziz Medical City, Ministry of National Guard Health Affairs, Riyadh, 11481, Saudi Arabia
†Prof. Mohammed Abumaree passed away on 26.07.2019.
Correspondence: Afrah E Mohammed; Yasser S Basmaeil
Stem Cell & Regenerative Medicine Department, King Abdullah International Medical Research Center, Ministry of National Guard Health Affairs, Riyadh, Saudi Arabia
Tel +966 11 8011111 Ext: 94573
Email [email protected]; [email protected]
Introduction: Human decidua basalis mesenchymal stem cells (DBMSCs) are potential therapeutics for the medication to cure inflammatory diseases, like atherosclerosis. The current study investigates the capacity of DBMSCs to stay alive and function in a harmful inflammatory environment induced by high levels of lipopolysaccharide (LPS).
Methods: DBMSCs were exposed to different levels of LPS, and their viability and functional responses (proliferation, adhesion, and migration) were examined. Furthermore, DBMSCs’ expression of 84 genes associated with their functional activities in the presence of LPS was investigated.
Results: Results indicated that LPS had no significant effect on DBMSCs’ adhesion, migration, and proliferation (24 h and 72 h) (p > 0.05). However, DBMSCs’ proliferation was significantly reduced at 10 μg/mL of LPS at 48 h (p < 0.05). In addition, inflammatory cytokines and receptors related to adhesion, proliferation, migration, and differentiation were significantly overexpressed when DBMSCs were treated with 10 μg/mL of LPS (p < 0.05).
Conclusion: These results indicated that DBMSCs maintained their functional activities (proliferation, adhesion, and migration) in the presence of LPS as there was no variation between the treated DBMSCs and the control group. This study will lay the foundation for future preclinical and clinical studies to confirm the appropriateness of DBMSCs as a potential medication to cure inflammatory diseases, like atherosclerosis.
Keywords: placenta, endothelial cells, proliferation, adhesion, migration, real-time PCR
Introduction
Stem cell-based therapy and regenerative medicine have potentials to treat multiple diseases, including cardiovascular diseases (CVD), respiratory, neurological, skeletal, and autoimmune diseases. Moreover, what makes stem cells unique is their ability to release chemokines and receptors. These secretomes are needed for tissue regeneration, differentiation into various cell types, and self-renewal.1 There are two classes of stem cells. They can be categorized to adult stem cells and embryonic stem cells.2 Both have the capacity to differentiate to varied types of cells; therefore, they are suitable to use in regenerative medicine and cellular therapy.3 Varied types of stem cells have been tested and evaluated, examples of which are embryonic stem cells (ESCs), bone marrow stem cells (BMSCs), induced pluripotent stem cells (iPSCs), hematopoietic stem cells (HSCs), neural stem cells (NSCs), very small embryonic-like stem cells (VSELs), mesenchymal stem cells (MSCs), and others.4–6 Due to ethical controversies, applications of embryonic stem cells in clinical settings are restricted.6,7
Nevertheless, mesenchymal stem cells (MSCs) gained attentions in medicine and were applied to stem cellular therapies because they have several advantages over other stem cells.1 MSCs can directly differentiate when implanted in the damaged tissue.8–10 MSCs regulate paracrine effects as they secrete cytokines and growth factors, thus increase inflammatory reaction or tissue fibrosis at the site of injury, which then stimulate endogenous progenitor cells and new blood vessels formation (angiogenesis).11,12
Moreover, at the site of injury, damaged tissues release micro-environmental factors that are related to ischemia gradients and inflammation; MSCs respond well to those factors released at the site of inflammation and migrate to the affected area.13–15 Furthermore, MSCs can be found in various sources in the body including, bone marrow, muscles, umbilical cord, and other areas and have potential to differentiate and heal damaged tissues.16–18 MSCs can be isolated from decidua basalis, which is a thin layer on the maternal tissue of the human placenta. These MSCs are called decidua basalis mesenchymal stem cells (DBMSCs).19,22 They have several important functions in response to any injury and one of which is their capability to differentiate into numerous cell lineages, such as chondrocytes, adipocytes, and osteocytes.16,20 DBMSCs are subjected to oxidative stress mediators and inflammation due to their significant location. They are located near the maternal circulation in their placental vascular microenvironment niche.21–23 Recently, it has been shown that DBMSCs maintain their functions in oxidative stress environments induced by exposure to high levels of hydrogen peroxide (H2O2).24 In addition, DBMSCs can preserve endothelial cell functions from damage resulting from exposure to elevated levels of oxidative stress inducers, including H2O2 and monocytes.23,25
Another form of oxidative stress or induced inflammatory microenvironment is the presence of endotoxins found on the outer membrane of gram-negative bacteria. Gram-negative organisms can be found in different locations on or in the human body. They are normally found in the gastrointestinal tract, genitourinary tract, and respiratory tract; and gram-negative aerobic and anaerobic bacteria colonize the human guts, which makes the human guts the largest reservoir of these microorganisms.26–29 Endotoxin or lipopolysaccharide (LPS) has potential roles in the structural integrity of the gram-negative bacterial walls.30 LPS is a complex glycolipid molecule.31,32 The LPS aids these organisms by shielding them from environmental impacts as well as antimicrobial or antibacterial products.30 Microorganisms produce LPS not only in infections but can leak LPS through the healthy and intact mucosal lines of the guts.33–36 Furthermore, the presence of harmful bacteria in the gut and LPS in the body is associated with several health conditions, such as inflammation, obesity, and metabolic disorder.31,32,37 It is well known that persistent inflammation can potentially develop many diseases in the body, such as autoimmune diseases, arthritis, allergic reactions, cancer, and vascular diseases.35,38,39 Continuous vascular inflammation can develop atherosclerosis, which is a chronic coronary heart disease, and potentially lead to health problems like cardiovascular diseases.39–41
Low concentrations of LPS can induce vascular inflammation and endothelial injury, which is characterized by dysfunction and phenotypic changes, including overexpression of inflammatory and adhesion molecules. Typical dysfunction includes increased endothelial cell proliferation and increased permeability, which is characterized by increased barrier functional activity of the endothelial cell layer allowing plasma macromolecules, such as lipoproteins to enter the intimal layer more easily.42–44 LPS can be detected in the blood of healthy people at low concentrations ranging from 3 to 200 pg/mL.33–36 This level of LPS in healthy individuals has the potential to activate endothelial cells and cause inflammatory responses, thus can lead to atherosclerosis.35,45–47 Even if the individuals have healthy guts, low levels of LPS leak through the intact mucosa and end up in the splanchnic circulation.29,48,49
In this study, DBMSCs’ functional responses in pro-inflammatory environments induced by LPS were evaluated. Several studies evaluated DBMSCs’ responses to oxidative stress and found that under oxidative stress environment induced by H2O2 as well as monocytes, DBMSCs’' functional activities including adhesion, proliferation, and migration were maintained.23,25 These significant results make DBMSCs as a suitable therapeutic target to repair endothelial cell injury and treat diseases caused by inflammation, such as atherosclerosis. Therefore, it was expected that DBMSCs would survive and function in harsh inflammatory environments induced by elevated levels of LPS.
In the current study, the main objective was to examine the ability of DBMSCs to survive in harsh inflammatory environments induced by high levels of LPS. Furthermore, the objectives were to study the consequences of LPS effects on DBMSCs’ functional activities including adhesion, proliferation, and migration and to evaluate DBMSCs' expression of genes associated with inflammation and their functional activities.
Materials and Methods
Ethics of Experimentation
This study was approved by the institutional research board (KAIMRC project # RC15/110/R & PNU Project # IRBC/246/13) at King Abdullah International Medical Research Centre (KAIMRC)/King Abdulaziz Medical City, Riyadh, Saudi Arabia. All placentae were obtained with informed consent from healthy patients. The study was conducted in accordance with the Declaration of Helsinki.
Isolation and Culture of DBMSCs
MSCs have been isolated from the decidua basalis (DBMSCs) of human term placenta right after delivery, using a previously published method.19 DBMSCs were cultured in a complete DBMSCs culture medium in flask T25 [DMEM-F12 medium containing 10% MSC FBS (mesenchymal stem cell certified fetal bovine serum, catalogue number 12-662-011, Life Technologies, Grand Island, USA), and antibiotics (100 µg/mL streptomycin and 100 U/mL penicillin)]. Incubation of the cells at 37 °C was performed in a humidified atmosphere consisting of 5% CO2 and 95% air, in a cell culture incubator.
Culture of DBMSCs with Different Concentrations of LPS
DBMSCs were cultured in varied concentrations (0, 0.50, 1, 10, 50, 100, 200 µg/mL) of LPS (Catalogue number TLRL-B5LPS, InvivoGen, USA). Experiments were completed in triplicate as well as repetition was performed independently with nine DBMSCs (passage 3) preparations.
DBMSCs’ Adhesion and Proliferation Assay Using the xCELLigence System
The xCELLigence system (RTCA-DP version; Roche Diagnostics, Mannheim, Germany) was used to evaluate the adhesion and proliferation of DBMSCs as this device automatically records and monitors real-time cell viability. The method used in this study was followed as previously published.23,25,50–52 The experiments were initiated by addition of 100 μL of complete DBMSC growth medium in 16-well culture plates (Catalogue number 05469813001, E-Plate 16, Roche Diagnostics) in presence and absence of different concentrations (0, 0.50, 1, 10, 50, 100, 200 µg/mL) of LPS. Plates were incubated at RT for 30 min then plates were placed in xCELLigence and instrument was set to blank. Plates were then removed from the machine. Next, the plates were seeded with 1×104 DBMSCs in each well and were incubated for 30 min at RT in the biosafety cabinet. The culture plates were then placed in the xCELLigence system at 37°C in a cell culture incubator, and the DBMSCs’ index was automatically monitored every 15 minutes for 72 h and analyzed through RTCA software [version 1.2.1.1002]; the data for cell adhesion was recorded at (2 h) and proliferation was recorded at (24, 48 and 72 h).23,25,50–52 LPS treatments were changed every 12 hours. Each experiment has been done and repeated in triplicate.
DBMSCs’' Migration Assay Using xCELLigence System
DBMSCs’ migration has been tested by applying the xCELLigence system, and CIM-16-well plates (Catalogue number 05665825001, CIM-16, Roche Diagnostics GmbH) as previously noted.23,25,50–52 The migration experiments have been started by addition of growth medium with 10% 'fetal bovine serum (FBS) in the presence and absence of different concentrations (0, 0.50, 1, 10, 50, 100, 200 µg/mL) of LPS in the lower chamber and free serum medium has been added in the upper chamber. The CIM-plate 16 was placed in the incubator at 37°C for 1 hour then the plate was placed in xCELLigence and instrument was set to blank. Next, the CIM-plate 16 was seeded with 1×104 DBMSCs in each well in the upper chamber and has been incubated for 30 min. Then, plate was placed in the xCELLigence system and DBMSCs migration has been monitored automatically every 15 min for 24 h using the xCELLigence system and RTCA software [version 1.2.1.1002]; then data have been presented as a cell index value.23,25,50–52 Migration of DBMSCs with or without 10% FBS has been acted as a positive and negative control, respectively. Experiment has been done and repeated in triplicate.
Gene Expression by Real-Time Polymerase Chain Reaction (RT-PCR)
DBMSCs’ expression of RT2 profiler PCR Arrays 84 genes related to Human Inflammatory Cytokines and Receptors (Catalog number PAHS-011Z, Qiagen, Hilden, Germany) has been detected as previously published.19,23,24,52 There were two DBMSC samples: the control samples (untreated) and samples that were treated with LPS. LPS treatments were changed every 12 hours during the 48-hours incubation period. After 48 h, the total RNA from DBMSCs [untreated DBMSCs, and DBMSCs treated with LPS] was extracted by using RNeasy Mini Kit (Qiagen, Hilden, Germany). Next, cDNA has been synthesized and utilized in a QuantiTect Reverse Transcription Kit (Qiagen, Hilden, Germany). Then, 20 µL of the mixture was added to each well in the RT2 profiler PCR Arrays. The 96-well plate was then placed in the Thermal Cycle. The real-time polymerase chain reaction (RT- PCR) has been done in triplicate on the CFX96 real-time PCR detection system (BIO-RAD, California, USA). Then, the data were analyzed.19,23,24,52 The ΔΔ−2 values have been then estimated to represent the findings as fold changes. The relative expression of internal controls (housekeeping genes) was used as provided in the RT2 profiler PCR Arrays kit. GAPDH was used as a housekeeping gene for humans as provided in the kit. Trials were performed in triplicate using DBMSCs obtained from three placentae independently.
Statistical Analysis
All the data shown in the bar graphs are presented as means ± standard deviation (SD) of three independent experiments. To avoid bias, the experiments were performed three times independently. Using GraphPad Prism 9, we analyzed the data sets of two experimental groups using t- test, two tailed. Data deemed statistically significant if p < 0.05.
Results
Isolation and Culture of DBMSCs
MSCs from the decidua basalis (DBMSCs) were successfully isolated from human term placentas and cultured in complete DBMSCs DMEM/F12 supplemented with penicillin at 100µg/mL and streptomycin at 100U/mL culture medium in a T75 cell culture flask, filter cap at 37°C in a cell culture incubator was previously described.25 At passage 2, DBMSCs were positive for (>95%) MSC markers, and negative for haematopoietic markers (Figure 1). DBMSCs were able to differentiate into adipocytes, chondrocytes and osteocytes. These properties of DBMSCs were consistent with what we have reported previously.19 In all experiments, DBMSCs were used at passage 2.
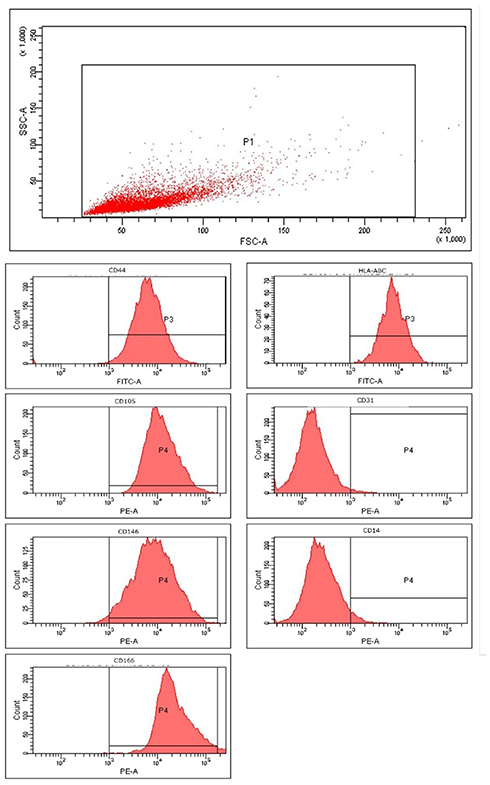 |
Figure 1 Characterization of decidua basalis mesenchymal stem/stromal cells (DBMSC's) for mesenchymal stem cell (MSC) markers. Flow cytometric analysis of DBMSCs for MSC markers (CD44, CD105, CD146, CD166 and HLA-ABC); and MSC-negative markers (CD31 (endothelial cell) and CD14 (macrophage/monocyte marker)). |
LPS Dose Response and Time Course for Experimental Standardization
To assess the impact of LPS on DBMSCs’ proliferation, DBMSCs have been cultured and treated at different concentrations of LPS and monitored for 72 h by using xCelligence system. After 24 h, the proliferation of treated DBMSCs with concentrations of 0.50, 1, 10, 50, 100, 200 µg/mL LPS showed no significant changes in comparison with the control (p > 0.05) (Figure 2B). After 48 h, the proliferation of treated DBMSCs was significantly decreased at 10 µg/mL LPS (p < 0.05) while had no significant changes at 0.5, 1, 50, 100 and 200 µg/mL LPS (p > 0.05) compared with the untreated DBMSCs (Figure 2C). Lastly, after 72 h, the proliferation of treated DBMCSs with continuous exposure of LPS had no significant change in comparison with the untreated group (p >0.05) (Figure 2D). Based on the results above, the exposure time of 48 h and 10 µg/mL LPS was selected to evaluate the effects of LPS on the functions of DBMSCs (morphology, migration and gene analysis).
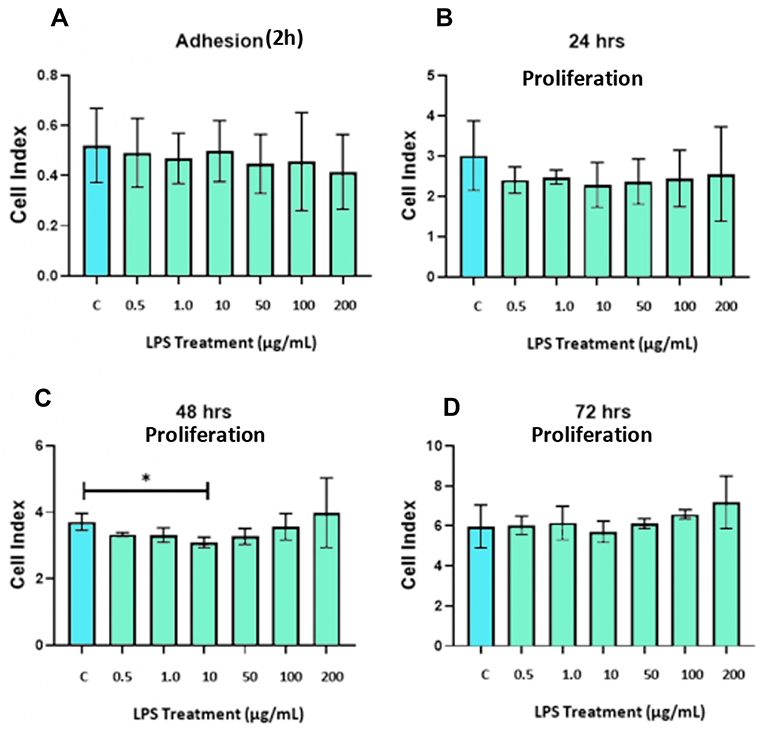 |
Figure 2 (A) LPS effects on DBMSCs’ adhesion in comparison with the control group at 2 h. (B–D) LPS effects on DBMSCs’ proliferation in comparison with the control group at 24 h, 48 h, and 72 h. Experiments were performed three times independently, and the data are represented as mean ± standard deviation. *Significant (p < 0.05). |
LPS Effects on DBMSCs’ Morphology
To see the effect of LPS on DBMSCs’ morphology, DBMSCs were cultured in 10 µg/mL LPS in complete DMEM/F12 for 48 h. Microscopic analysis showed no change in the morphology of 10 µg/mL LPS treated DBMSCs in compared with the untreated DBMSCs at 24 and 48 h (Figure 3). The viability of DBMSCs treated with 10 µg/mL LPS was >95%.
 |
Figure 3 (A1 and A2) Untreated DBMSCs vs DBMSCs treated with continuous exposure of 10 µg/mL LPS on the first hour when treatment was initiated at 60% confluency (100× magnification). (B1 and B2) Untreated DBMSCs vs DBMSCs treated with continuous exposure of 10 µg/mL LPS after 24 h of incubation period at 80% confluency (100× magnification). (C1 and C2) Untreated DBMSCs vs DBMSCs treated with continuous exposure of 10 µg/mL LPS after 48 h of incubation period at 90% confluency (100× magnification). |
LPS Effects on DBMSCs’ Migration
Under continuous exposure to LPS, the migration of DBMSCs in the upper chamber of the migration plates was examined and monitored by the xCELLigence system for 24 h. After 24 h, no significant impact of LPS on DBMSCs’ migration (p>0.05) was stated compared to the control (Figure 4).
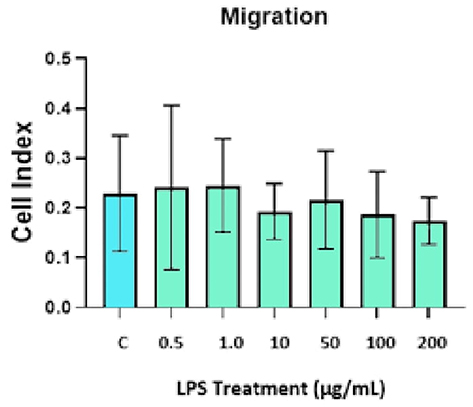 |
Figure 4 LPS effects on DBMSCs’ migration in comparison with the control group at 24 h. Experiments were performed three times independently, and the data are represented as mean ± standard deviation. |
LPS Modulates Various Gene Expression Related to Inflammatory Cytokines and Receptors in DBMSC
Total RNA was extracted from untreated and 10 µg/mL LPS treated DBMSCs for 48 h. the gene expression of inflammatory cytokines and chemokines was analyzed and assessed using RT-PCR. Then the ΔΔ−2 values were used to yield the fold change values. In Tables 1–3, as well as Figure 5, results indicated that LPS influenced and modulated DBMSCs’ expression of various genes that are related to human inflammatory cytokines and receptors. In Table 4 and Figure 6 results showed cytokines and receptors that mediate DBMSCs’ functional responses, such as proliferation, migration, and differentiation were overexpressed (p < 0.05). In Table 5 and Figure 7 genes that mediate DBMSCs’ migration properties such as CXCR2 and CXCL12 were down regulated in the presence of LPS (p < 0.05). There are other genes that were down regulated in the presence of LPS. However, they are not necessarily related to DBMSCs’ functions (Table 5). The number of overexpressed genes that are related to DBMSCs’ functional activities (proliferation, adhesion, and migration) were 31 genes (p < 0.05) (Table 4). On the other hand, the number of downregulated genes were 33 genes, 31 of which are not necessarily related to DBMSCs’ functions while two of the genes were related to DBMSCs’ migration properties (CXCR2 and CXCL12) (p < 0.05) (Table 5). Biological activities of the genes in the following Tables 1–5 were obtained from GeneBank via https via www.ncbi.nlm.nih.gov/nuccore/.
 |
Table 1 LPS Effects on DBMSCs’ Gene Expression of Cytokines and Cytokine Receptors. |
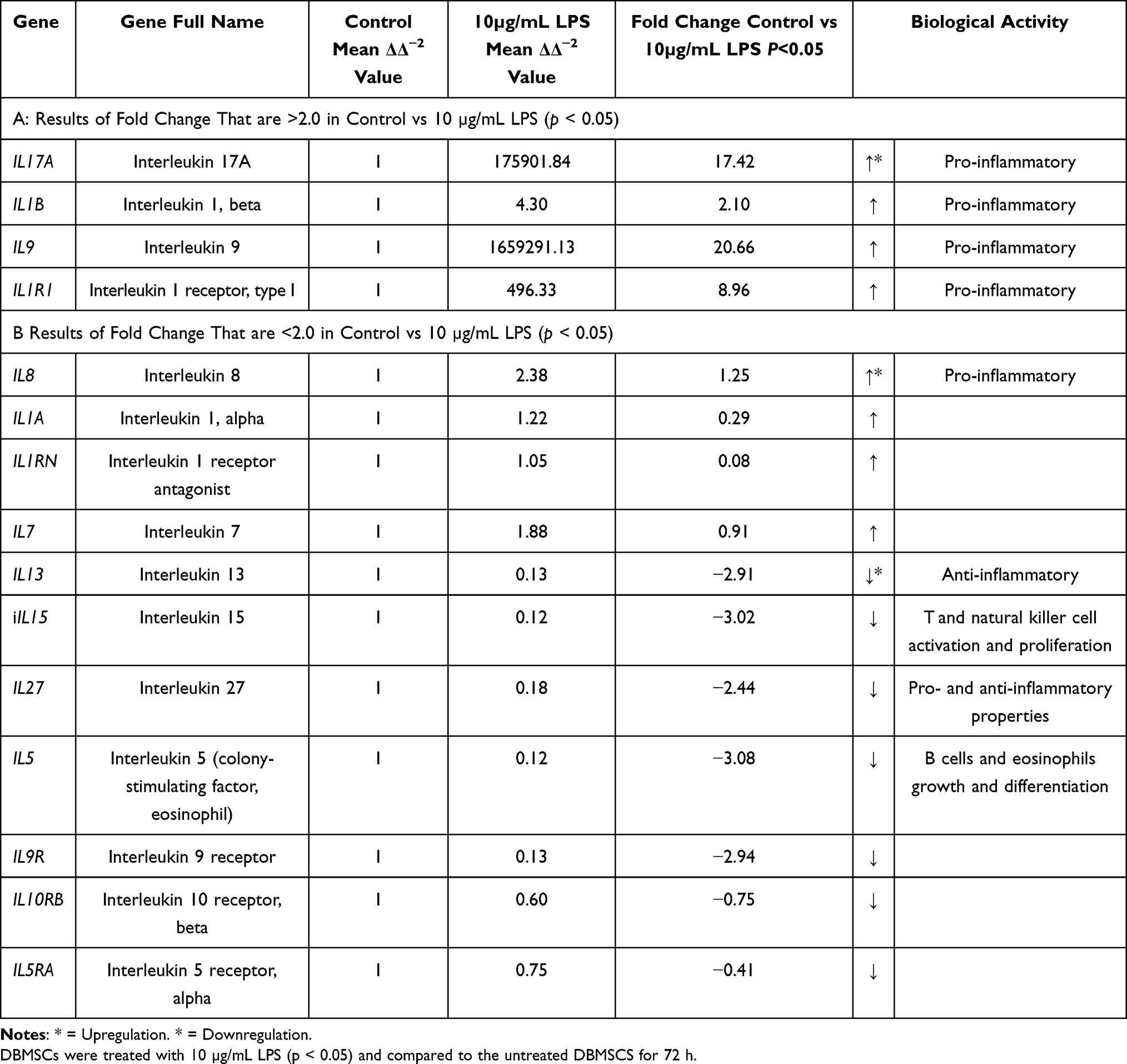 |
Table 2 LPS Effects on DBMSCs’ Gene Expression of Interleukins and Interleukin Receptors. |
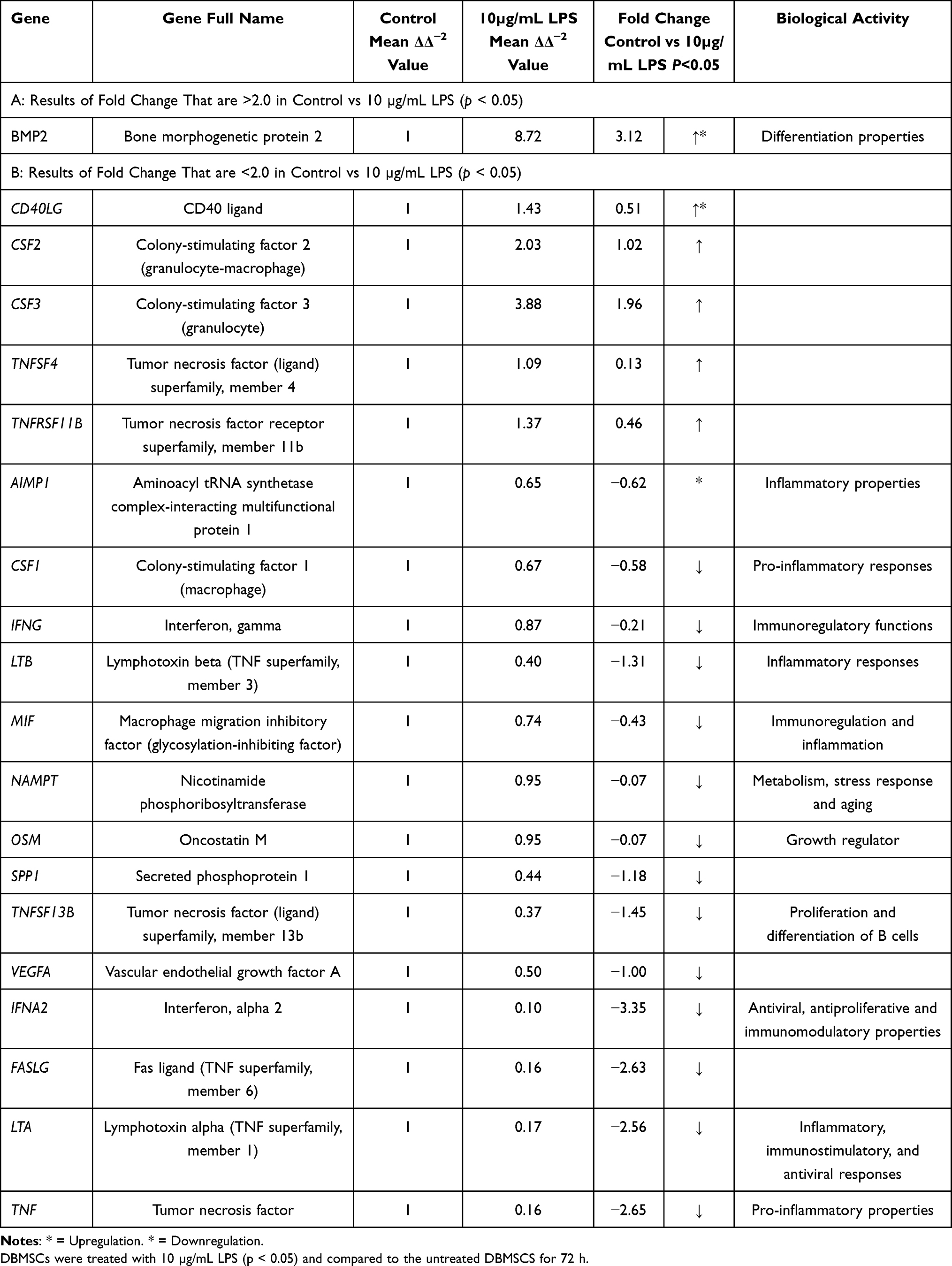 |
Table 3 LPS Effects on DBMSCs’ Gene Expression of Other Cytokines and Other Cytokine Receptors. |
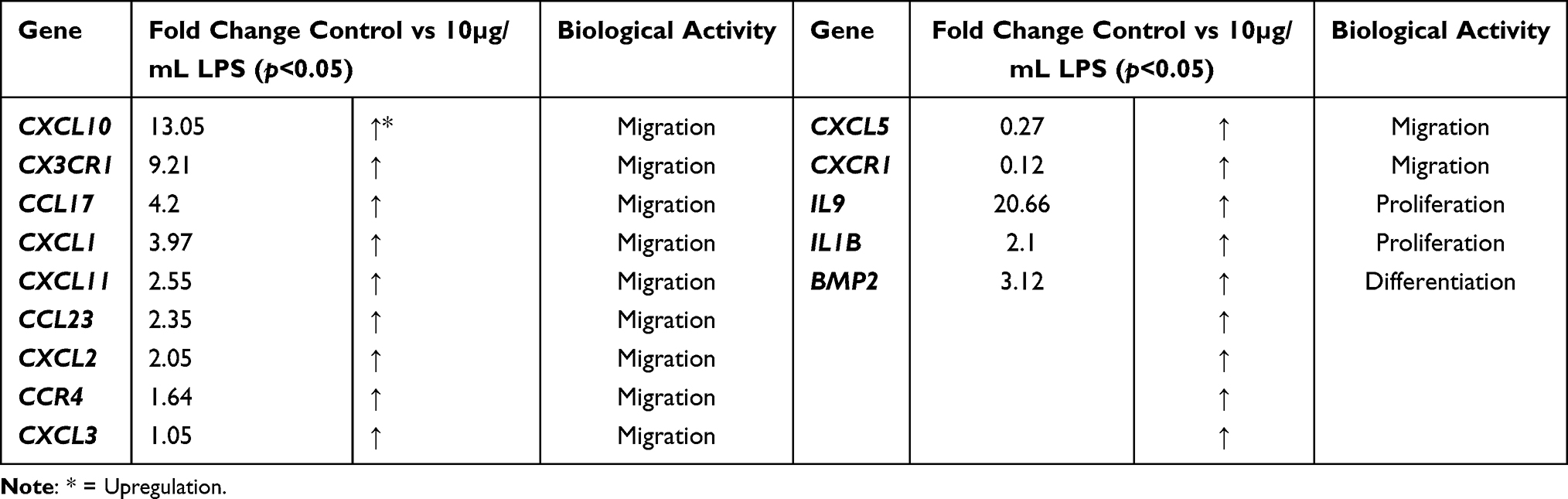 |
Table 4 Overexpressed Cytokines and Receptors That are Responsible for DBMSC Functions (Migration, Proliferation, Differentiation) in the Presence of LPS |
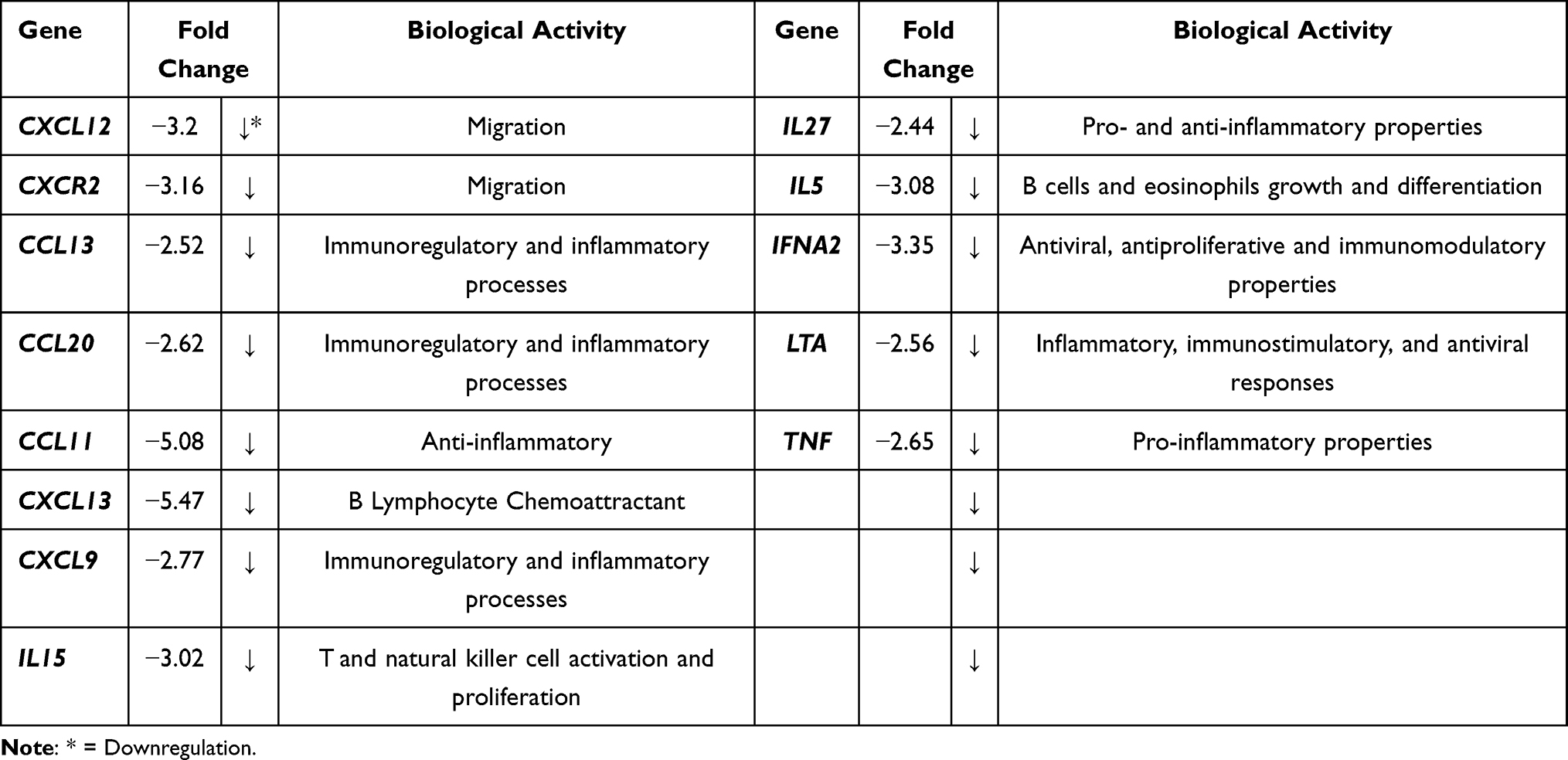 |
Table 5 Downregulated Cytokines and Receptors That are Responsible for DBMSCs’ Migration and Other Genes That are Not Necessarily Related to DBMSCs Functions in the Presence of LPS |
 |
Figure 5 RT-PCR amplification curve of the untreated DBMSCs vs DBMSCs treated with 10 µg/mL LPS (generated by CFX-96 Real-Time System). Experiments were performed three times independently, and the data are represented as mean. |
 |
Figure 6 Overexpressed cytokines and receptors that are responsible for DBMSC functions (migration, proliferation, differentiation) in the presence of LPS. Experiments were performed three times independently, and the data are represented as mean. |
 |
Figure 7 Downregulated cytokines and receptors that are responsible of DBMSCs migration and other genes that are not necessarily related to DBMSC functions in the presence of LPS. Experiments were performed three times independently, and the data are represented as mean. |
Discussion
Mesenchymal stem cells have gained a lot of attention for their therapeutic agent properties over the past decade. Moreover, MSCs have properties to regenerate and differentiate into mesodermal lineages (adipocytes, osteoblasts, and chondroblasts) as well as neuroectodermic lineages (neurons or alveolar endothelial cells).53–55 In addition, MSCs could be differentiated into non-mesodermal lineages (eg, cardiomyocytes, hepatocytes, or epithelial cells).56–58 MSCs could be originated from the bone marrow; however, they have been isolated from varied origins in humans, such as adipose tissue, hair follicles, umbilical cords, placentas, muscle, peripheral blood, amniotic fluid, and other organs, such as brain, liver, or lung.16,54,59,60 Moreover, MSCs have different sources and their multilineage differentiations are not the only potentials that make MSCs special. MSCs have potential to migrate to sites of inflammation in which they stimulate tissue regeneration by releasing paracrine factors as well as differentiating and replacing damaged tissues or cells.61 Not only MSCs have the capabilities to heal but they also are involved in paracrine signaling and cell–cell communication.62 Furthermore, MSCs have the ability to interact with the host niche at the site of injury or inflammation and promote the following functions: MSCs can inhibit the immune system, promote cell survival, and/or form new blood vessels via angiogenesis by releasing factors, such as IL-6.61,63
There are several impacts that influence the successive implications of MSCs in clinical settings when cell therapy is introduced to the patients. As a result of these challenges, 99% of MSCs will die a few days after transplantations, thus resulting in unsuccessful applications of this approach.64–73 These challenges can result from MSCs isolation techniques that will result in uncontrolled cell quality, or invasive isolation processes as there are several ways and approaches to isolate MSCs from tissues, loss of MSCs potency, limited lifespan of MSCs, or loss of MSCs properties when cells are expanding or proliferating in vitro. Other challenges could be initiated when MSCs are transplanted in the recipients’ tissues or organs, such as stressful condition (oxidative stress, inflammation, radiation, and chemotherapy).64–73 Interestingly, MSCs that are derived and isolated from maternal decidua basalis tissue of human placenta have unique phenotypic characteristics and can withstand environments that are highly stressful because pregnancy is a highly stressful condition in which the decidua basalis tissue is exposed to high amounts of oxidative stress or inflammatory environment.22–24,74–76 Previously, H2O2 and monocytes were introduced to DBMSCs, and results demonstrated that DBMSCs maintained their phenotypic properties and functions.23 LPS is another form of inflammatory environment inducer and in this study, LPS was introduced to DBMSCs, and their phenotypic functions were monitored and analyzed.
According to the International Society for Cellular Therapy (ISCT), for MSCs to be used as a therapeutic agent, they must meet the minimal criteria for defining their multipotent function; one of which is the MSCs ability to adhere to plastic surfaces when grown in vitro.77 In the current study, DBMSCs were treated with LPS without preconditioning. Results showed that after 2 h, LPS had no significant effect on DBMSCs’ adhesion (p > 0.05). Khatlani et al24 studied the multipotent properties of DBMSCs when they were preconditioned with H2O2. The authors found that DBMSCs’ adhesion in presence of H2O2 was significantly increased when they were preconditioned with H2O2 for 72 h (p < 0.05) while DBMSCs’ adhesion that were not H2O2-preconditioned remained unchanged (p > 0.05). Same trend of observation regarding DBMSCs’ adhesion when subjected to a stressful condition without preconditioning was noted. Previous studies found that preconditioned MSCs, such as DBMSCs and BMMSCs with oxidative stress mediators (eg, H2O2, glucose, hypoxia) enhanced the MSCs’ performance.24,76,78 Several studies claimed that the possible reason why MSCs’ performances were enhanced when preconditioned with hypoxia or oxidative stress could be due to activation of paracrine or pro-angiogenic pathways, so they could survive and adapt to oxidative stress environments.78–82
Next, DBMSCs’ ability to proliferate in the presence of an pro-inflammatory mediator, such as LPS is important to evaluate DBMSCs’ potential use in cellular therapies. Current results showed that LPS had no significant effect on DBMSCs’ proliferation during the 24 h and 72 h (p > 0.05). However, at 48 h, LPS significantly reduced DBMSCs’ proliferation at 10 µg/mL LPS (p < 0.05) while had no significant changes on DBMSCs’ proliferation at 0.5, 1, 50, 100, and 20,010 µg/mL LPS (p > 0.05). Khatlani et al24 reported a similar finding in which the authors evaluated the effect of H2O2 on DBMSCs’ proliferation. After 24 h and 48 h, 1, 5, 25, 50, 100, and 200 μM of H2O2 had no significant effect on DBMSCs’ proliferation (p > 0.05); however, DBMSCs’ proliferation was significantly reduced when they were treated with higher concentration of H2O2 (400 and 600 μM) (p < 0.05). A study conducted by Kurte et al83 found that MSC phenotypes can change depending on how long they were exposed to LPS. It was found that MSCs exerted different phenotypes when they were exposed to LPS at 1 h than when they were exposed to LPS at 24 h and 48 h. The authors claimed that the molecular explanation behind this phenomenon is still unclear.
In addition, results showed that IL-9 was overexpressed when DBMSCs were treated with 10 µg/mL of LPS. However, Durand et al84 found that LPS had no significant changes on IL-9 secretion by BMSCs. The variations in IL-9 expression in the presence of LPS could be related to the different types of MSCs investigated and treatment conditions. IL-9 has many important functions; it modulates and regulates several hematopoietic cells, it prevents programmed cell death (apoptosis), and most importantly it stimulates cell proliferation.85 In a study conducted by Sun et al,86 the authors found that IL-9 played an important role in trophoblast cells angiogenesis; and reduction of IL-9 led to reduction of angiogenesis in trophoblast cells. In addition, IL1-B is a pro-inflammatory cytokine that has a great effect on MSCs proliferative properties.87–89 In current study, IL1-B was overexpressed when DBMSCs were treated with 10 µg/mL LPS. This finding correlated with a study conducted by Cui et al90 in which the authors found that several pro-inflammatory cytokines, such as IL1-B that were secreted by MSCs in the lungs were upregulated under exposure of chronic airway LPS. However, additional future mechanistic study is necessary to confirm this.
To yield a successful outcome for MSCs’ clinical applications, MSCs need to be able to migrate to the sites of inflammation (target tissues).77 Results showed that LPS had no significant effect on DBMSCs’ migration (p>0.05) after 24-h incubation in the upper chamber of the migration plates. Similarly, Khatlani et al24 found that DBMSCs’ migration in the presence of H2O2 had no significant effect (p > 0.05) while preconditioning DBMSCs with H2O2 yielded a significant increase in DBMSCs’ migration (p < 0.05). Chemoattractant signals can influence the rate of MSCs’ migration to the damaged tissue; and MSCs’ migration is mediated by many chemokine ligands and receptors, such as CXCL12, CCR4, and others. These chemokines and receptors have an important effect in MSCs’ inflammatory responses and migration.91–96 Hereby, when DBMSCs were treated with 10 µg/mL of LPS, various chemokines and receptors, including CCL23, CXCL10, CXCL11, CX3CR1, CXCL2, CCR4, CXCL3, and CXCL5 were significantly overexpressed (p < 0.05). Previous studies noted that DBMSCs alone expressed CC, CXC and various chemokines and receptors family members, such as CCL1, CCL2, CCL21 and others. Nevertheless, DBMSCs alone did not express certain chemokines and receptors, such as CXCR1 and CX3CR1.19 Yet, in the presence of LPS, DBMSCs significantly overexpressed CXCR1 and CX3CR1 (p < 0.05). However, CXCL12 was downregulated in the presence of LPS, and this result is correlated to a study conducted by Wobus et al.97 In their study, the authors tested CXCL12 secretion and expression by MSCs in breast cancer cells in comparison to non-tumorigenic breast epithelial cells. They found that CXCL12 was downregulated in the breast cancer cells while the control group had no effect of CXCL12 secretion. The authors explained that cancer cells paracrine factors or small molecules derived from cancer or tumor cells could have the possible effect on this observation.97 In the current study, LPS could be the reason why CXCL12 was downregulated. It was found that overexpression of certain chemokine ligands or receptors, such as the chemokine receptor CXCR2, or chemokine ligands CXCL1, CXCL3, and CXCL5 can enhance MSCs’ migration to the target tissue and promote cell survival.98 In Shen et al study,98 MSCs were exposed to radiation/chemical-induced oral mucositis and the authors found that CXCR2 level was overexpressed after radiation and was peaked at day 7 and eventually started to decline. In addition, CXCL1, CXCL3, and CXCL5 were upregulated after exposure to radiation. However, in the presence of LPS, CXCR2 was significantly downregulated. Different trends in oxidative stress inducers could be the reason why these results did not correlate. Yet, chemokine ligands, such as, CXCL3, and CXCL5 were significantly overexpressed in the current study. These chemokines and receptors are important because they trigger factors at the site of inflammation; and with the aid of cell adhesion molecules, they contribute to the mobilization and migration of MSCs to the target tissues.91–96 However, to investigate the mechanism of migration of DBMSCs in the presence of LPS will be investigated in a later study.
Upon MSCs’ recruitment to the sites of the injury or the target tissues, MSCs secrete several mediators to initiate tissue repair and regeneration, such as anti-inflammatory, anti-apoptotic, immunomodulatory, and other agents. Both anti-inflammatory and immunomodulatory have a significant and main role in the therapeutic aspects of MSCs’ application in clinical settings.99–101 MSCs also modulate and regulate soluble factors that are related to immune responses, such as PGE2, TGF-β, IL-6, hepatocyte growth factor (HGF), or others.14,102–104 Results showed that DBMSCs treated with 10 µg/mL of LPS overexpressed certain interleukins and receptors. Du-Rocher et al found in their study through using chip array analysis that IL-17 signaling pathway is the second important pathways in MSCs.105 Moreover, IL-17 is linked to other pro-inflammatory cytokines (IFNγ and TNFα) that promote the secretion of other chemokines, such as IL-6, IL-8, ICAM-1, and PGE2.106–109 In their study, IL-17 yielded large amounts of IL-8 and IL-6.105 In the current study, IL-17A was overexpressed when DBMSCs were treated with 10 µg/mL of LPS. IL-8 was also overexpressed in the presence of LPS. These findings correlate to a study conducted by Durand et al.84 The authors evaluated the effect of certain factors, such as LPS on BMSC secretomes and they found that BMSCs secretory capacity was increased in the presence of LPS, such as IL-17A and IL-8.84 Other soluble factors were overexpressed when DBMSCs were under continual exposure of 10 µg/mL LPS, such as BMP-2. Our result correlated to a study in which the authors tested the effect of exosomes derived from human LPS-activated monocytes on MSCs osteogenic gene expression and found that BMP-2 was significantly increased.110 The BMP family which consists of 15 members can promote MSCs’ differentiation into several lineages and that depends on the microenvironment concentrations. Each member of BMP family will promote MSCs’ differentiation to certain lineages. For instance, BMP-2, BMP-4, and BMP-7 can promote MSCs’ differentiation to adipogenic lineage. BMP-2, BMP-6, and BMP-9 can modulate MSCs’ differentiation to osteogenic lineage. Lastly, BMP-2 and BMP-7 can promote MSCs to differentiate into chondrogenic lineage.111,112 The functional aspect of immune modulation of DBMSCs preconditioned with LPS will be determined in a future study.
These results and findings were in accordance with several studies on MSCs’ functional responses and gene expression of cytokines and receptors when treated with several oxidative stress inducers. These results are evident for the potential use of DBMSCS as a therapeutic agent for the treatment of inflammatory diseases, such as atherosclerosis since DBMSCs survived in harsh environments that were induced by high levels of LPS.
Conclusion
In conclusion, DBMSCs have potential functional properties that enhance their application in cell-based therapy because of their abilities to work in highly oxidative stress environments as they maintained their functional activities when treated with H2O2 and monocytes.23 LPS is another form of inflammatory environment condition. Results of this study indicated that DBMSCs’ functions (adherence, proliferation, and migration) were not significantly affected (p> 0.05) when DBMSCs were treated with continuous exposures of different concentrations (0.50, 1, 10, 50, 100, 200 µg/mL) of LPS (Figure 8). No variation between treated DBMSCs and the control group indicated that DBMSCs can survive in pro-inflammatory condition, in the presence of LPS, and can maintain their functional activities (adherence, proliferation, and migration). Moreover, many chemokines and receptors that are related to DBMSCs’ phenotypic properties, such as adherence, proliferation, differentiation, and migration were overexpressed when DBMSCs have been subjected to 10 µg/mL of LPS. Nevertheless, future in vitro and in vivo studies on DBMSCs are necessary to assess the potential use of DBMSCs in treating inflammatory diseases. For instance, DBMSCs’ ability to invade endothelial tissues or layers in LPS condition and time-related phenotype changes of DBMSCs in the presence of continuous exposure to LPS. Evaluating the functional responses of DBMSCs when preconditioned with LPS will be investigated in future studies.
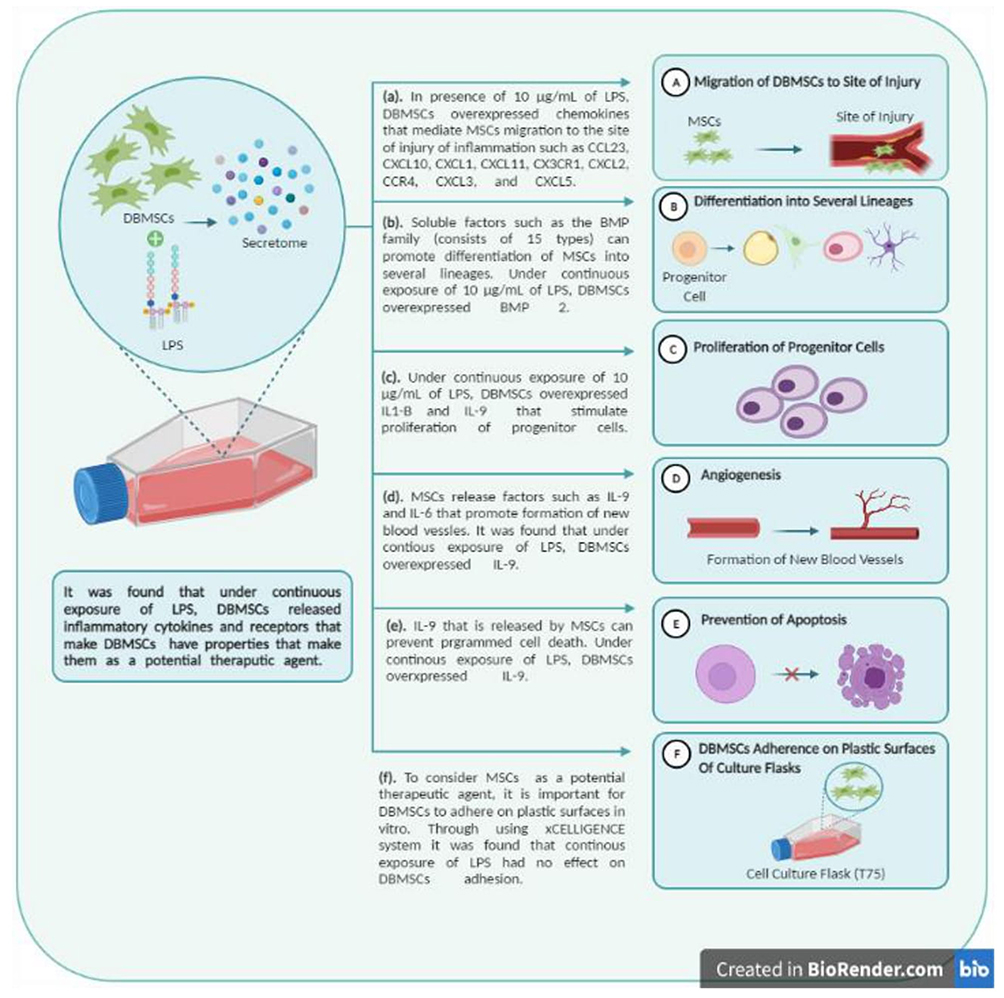 |
Figure 8 The effect of LPS on DBMSCs’ functional responses (proliferation, adhesion, and migration) (Created with BioRender.com). |
Acknowledgments
This research was funded by the Deanship of Scientific Research at Princess Nourah Bint Abdulrahman University, through the Pioneer Researcher Funding Program (Grant No# PR-1441-4) and King Abdullah International Medical Research Center for providing the supervion and facilities to support the study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Shin JH, Ryu CM, Yu HY, Shin DM, Choo MS. Current and future directions of stem cell therapy for bladder dysfunction. Stem Cell Rev Rep. 2020;16(1):82–93.
2. Bae SH. Recent achievements in stem cell therapy for pediatric gastrointestinal tract disease. Pediatr Gastroenterol Hepatol Nutr. 2013;16(1):10–16. doi:10.5223/pghn.2013.16.1.10
3. Quante M, Wang TC. Stem cells in gastroenterology and hepatology. Nat Rev Gastroenterol Hepatol. 2009;6(12):724. doi:10.1038/nrgastro.2009.195
4. Van Der Flier LG, Clevers H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu Rev Physiol. 2009;71:241–260. doi:10.1146/annurev.physiol.010908.163145
5. Ratajczak M, Suszynskan M, Pedziwiatr D, Mierzejewska K, Greco N. Umbilical cord blood-derived very small embryonic like stem cells (VSELs) as a source of pluripotent stem cells for regenerative medicine. Pediatr Endocrinol Rev. 2012;9(3):639–643.
6. Chu DT, Nguyen TT, Tien NLB, et al. Recent progress of stem cell therapy in cancer treatment: molecular mechanisms and potential applications. Cells. 2020;9(3):563. doi:10.3390/cells9030563
7. Ghafarzadeh M, Namdari P, Tarhani M, Tarhani F. A review of application of stem cell therapy in the management of Congenital Heart Disease. J Matern Fetal Neonatal Med. 2020;33(9):1607–1615.
8. Heo JS, Choi Y, Kim HS, Kim HO. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int J Mol Med. 2015;37:115–125. doi:10.3892/ijmm.2015.2413
9. Elahi KC, Klein G, Avci-Adali M, Sievert KD, Macneil S, Aicher WK. Human mesenchymal stromal cells from different sources diverge in their expression of cell surface proteins and display distinct differentiation patterns. Stem Cells Int. 2016;2016:1–9. doi:10.1155/2016/5646384
10. Nancarrow-Lei R, Mafi P, Mafi R, Khan W. A systemic review of adult mesenchymal stem cell sources and their multilineage differentiation potential relevant to musculoskeletal tissue repair and regeneration. Curr Stem Cell Res. 2017;12:601–610.
11. Olson TS, Ley K. Chemokines and chemokine receptors in leukocyte trafficking. Am J Physiol Regul Integr Comp Physiol. 2002;283(1):R7–R28. doi:10.1152/ajpregu.00738.2001
12. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stem cell signaling and therapy. Circ Res. 2008;103(11):1204–1219. doi:10.1161/CIRCRESAHA.108.176826
13. Meirelles LDS, Chagastelles PC, Nardi NB. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J Cell Sci. 2006;119:2204–2213. doi:10.1242/jcs.02932
14. Chen K, Wang D, Du WT, et al. Human umbilical cord mesenchymal stem cells hUC-MSCs exert immunosuppressive activities through a PGE2-dependent mechanism. Clin Immunol. 2010;135:448–458. doi:10.1016/j.clim.2010.01.015
15. Nitzsche F, Müller C, Lukomska B, Jolkkonen J, Deten A, Boltze J. Concise review: MSC adhesion cascade—insights into homing and transendothelial migration. Stem Cells. 2017;35:1446–1460. doi:10.1002/stem.2614
16. Friedenstein AJ, Chailakhjan RK, Lalykina KS. The development of fibroblast colonies in monolayer cultures of Guinea pig bone marrow and spleen cells. Cell Prolif. 1970;3:393–403. doi:10.1111/j.1365-2184.1970.tb00347.x
17. Wagers AJ, Weissman IL. Plasticity of adult stem cells. Cell. 2004;116(5):639–648. doi:10.1016/S0092-8674(04)00208-9
18. Bianco P, Cao X, Frenette PS, et al. The meaning, the sense and the significance: translating the science of mesenchymal stem cells into medicine. Nat Med. 2013;19(1):35–42. doi:10.1038/nm.3028
19. Abomaray FM, Al Jumah MA, Alsaad KO, et al. Phenotypic and functional characterization of mesenchymal stem/multipotent stromal cells from decidua basalis of human term placenta. Stem Cells Int. 2016;2016:1–18. doi:10.1155/2016/5184601
20. Ding DC, Shyu WC, Lin SZ. Mesenchymal stem cells. Cell Transplant. 2011;20(1):5–14. doi:10.3727/096368910X
21. Huang YC, Yang ZM, Chen XH, et al. Isolation of mesenchymal stem cells from human placental decidua basalis and resistance to hypoxia and serum deprivation. Stem Cell Rev. 2009;5:247–255. doi:10.1007/s12015-009-9069-x
22. Kusuma GD, Abumaree MH, Pertile MD, Perkins AV, Brennecke SP, Kalionis B. Mesenchymal stem/stromal cells derived from a reproductive tissue niche under oxidative stress have high aldehyde dehydrogenase activity. Stem Cell Rev Rep. 2016;12(3):285–297. doi:10.1007/s12015-016-9649-5
23. Alshabibi MA, Khatlani T, Abomaray FM, et al. Human decidua basalis mesenchymal stem/stromal cells protect endothelial cell functions from oxidative stress induced by hydrogen peroxide and monocytes. Stem Cell Res Ther. 2018;9(1). doi:10.1186/s13287-018-1021-z
24. Khatlani T, Algudiri D, Alenzi R, et al. Preconditioning by hydrogen peroxide enhances multiple properties of human decidua basalis mesenchymal stem/multipotent stromal cells. Stem Cells Int. 2018;2018:1–13. doi:10.1155/2018/6480793
25. Alshabibi MA, Al Huqail AJ, Khatlani T, et al. Mesenchymal stem/multipotent stromal cells from human decidua basalis reduce endothelial cell activation. Stem Cells Dev. 2017;26:1355–1373. doi:10.1089/scd.2017.0096
26. De Nardin E. The role of inflammatory and immunological mediators in periodontitis and cardiovascular disease. Ann Periodontol. 2001;6:30–40. doi:10.1902/annals.2001.6.1.30
27. Kuramitsu HK, Kang IC, Qi M. Interactions of Porphyromonas gingivalis with host cells: implications for cardiovascular diseases. J Periodontol. 2003;74:85–89. doi:10.1902/jop.2003.74.1.85
28. Li L, Messas E, Batista EL
29. Horseman MA, Surani S, Bowman JD. Endotoxin, toll-like receptor-4, and atherosclerotic heart disease. Curr Cardiol Rev. 2017;13(2):81–93.
30. Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol Mol Biol Rev. 2003;67:593–656.
31. Rietschel ET, Kirikae T, Schade FU, et al. Bacterial endotoxin: molecular relationships of structure to activity and function. FASEB J. 1994;8:217–225. doi:10.1096/fasebj.8.2.8119492
32. Raetz CR, Whitfield C. Lipopolysaccharide endotoxins. Annu Rev Biochem. 2002;71:635–700. doi:10.1146/annurev.biochem.71.110601.135414
33. Goto T, Eden S, Nordenstam G, Sundh V, Svanborg-Eden C, Mattsby-Baltzer I. Endotoxin levels in sera of elderly individuals. Clin Diagn Lab Immunol. 1994;1:684–688. doi:10.1128/cdli.1.6.684-688.1994
34. Niebauer J, Volk HD, Kemp M, et al. Endotoxin and immune activation in chronic heart failure: a prospective cohort study. Lancet. 1999;353:1838–1842. doi:10.1016/S0140-6736(98)09286-1
35. Wiedermann CJ, Kiechl S, Dunzendorfer S, et al. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: prospective results from the Bruneck Study. J Am Coll Cardiol. 1999;34:1975–1981. doi:10.1016/S0735-1097(99)00448-9
36. Kiechl S, Egger G, Mayr M, et al. Chronic infections and the risk of carotid atherosclerosis: prospective results from a large population study. Circulation. 2001;103(8):1064–1070. doi:10.1161/01.CIR.103.8.1064
37. Cani PD, Amar J, Iglesias MA. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56(7):1761–1772. doi:10.2337/db06-1491
38. Applebaum KM, Ray RM, Astrakianakis G. Evidence of a paradoxical relationship between endotoxin and lung cancer after accounting for left truncation in a study of Chinese female textile workers. Occup Environ Med. 2013;70(10):709–715. doi:10.1136/oemed-2012-101240
39. Jeon D, Kim SJ, Kim HS. Anti-inflammatory evaluation of the methanolic extract of Taraxacum officinale in LPS-stimulated human umbilical vein endothelial cells. BMC Complement Altern Med. 2017;17(1). doi:10.1186/s12906-017-2022-7
40. Ross R. Atherosclerosis–an inflammatory disease. N Engl J Med. 1999;340(2):115–126. doi:10.1056/NEJM199901143400207
41. Libby P. Inflammation in atherosclerosis. Arterioscler Thromb Vasc Biol. 2012;32(9):2045–2051. doi:10.1161/ATVBAHA.108.179705
42. Pfeffer R, Ganatos P, Nir A, Weinbaum S. Diffusion of macromolecules across the arterial wall in the presence of multiple endothelial injuries. J Biomech Eng. 1981;103:197–203. doi:10.1115/1.3138278
43. Stoll LL, Denning GM, Weintraub NL. Potential role of endotoxin as a proinflammatory mediator of atherosclerosis. Arterioscler Thromb Vasc Biol. 2004;24:2227–2236. doi:10.1161/01.ATV.0000147534.69062.dc
44. Weinbaum S, Tzeghai G, Ganatos P, Pfeffer R, Chien S. Effect of cell turnover and leaky junctions on arterial macromolecular transport. Am J Physiol. 1985;248:H945–960.
45. Lehr HA, Sagban TA, Ihling C, et al. Immunopathogenesis of atherosclerosis: endotoxin accelerates atherosclerosis in rabbits on hypercholesterolemic diet. Circulation. 2001;104:914–920. doi:10.1161/hc3401.093153
46. Ostos MA, Recalde D, Zakin MM, Scott-Algara D. Implication of natural killer T cells in atherosclerosis development during a LPS-induced chronic inflammation. FEBS Lett. 2002;519:23–29. doi:10.1016/S0014-5793(02)02692-3
47. Rice JB, Stoll LL, Li WG, et al. Low-level endotoxin induces potent inflammatory activation of human blood vessels: inhibition by statins. Arterioscler Thromb Vasc Biol. 2003;23:1576–1582. doi:10.1161/01.ATV.0000081741.38087.F9
48. Ghanim H, Sia CL, Upadhyay M, et al. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am J Clin Nutr. 2010;91(4):940–949. doi:10.3945/ajcn.2009.28584
49. Laugerette FC, Vors A, Geloen A. Emulsified lipids increase endotoxemia: possible role in early postprandial low-grade inflammation. J Nutr Biochem. 2011;22(1):53–59. doi:10.1016/j.jnutbio.2009.11.011
50. Abumaree MH, Abomaray FM, Alshabibi MA, AlAskar AS, Kalionis B. Immunomodulatory properties of human placental mesenchymal stem/ stromal cells. Placenta. 2017;59:87–95. doi:10.1016/j.placenta.2017.04.003
51. Abumaree MH, Hakami M, Abomaray FM, et al. Human chorionic villous mesenchymal stem/stromal cells modify the effects of oxidative stress on endothelial cell functions. Placenta. 2017;59:74–86. doi:10.1016/j.placenta.2017.05.001
52. Basmaeil YS, Al Subayyil AM, Khatlani T, et al. Human chorionic villous mesenchymal stem/stromal cells protect endothelial cells from injury induced by high level of glucose. Stem Cell Res Ther. 2018;9:238. doi:10.1186/s13287-018-0984-0
53. Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and disease. Nat Rev Immunol. 2008;8:726–736. doi:10.1038/nri2395
54. Wei X, Yang X, Han ZP, Qu FF, Shao L, Shi YF. Mesenchymal stem cells: a new trend for cell therapy. Acta Pharmacol Sin. 2013;34:747–754. doi:10.1038/aps.2013.50
55. Squillaro T, Peluso G, Galderisi U. Clinical trials with mesenchymal stem cells: an update. Cell Transplant. 2016;25:829–848. doi:10.3727/096368915X689622
56. Lee KD, Kuo TK, Whang-Peng J, et al. In vitro hepatic differentiation of human mesenchymal stem cells. Hepatology. 2004;40(6):1275–1284. doi:10.1002/hep.20469
57. Paunescu V, Deak E, Herman D, Siska IR, Tanasie G, Bunu C. In vitro differentiation of human mesenchymal stem cells to epithelial lineage. J Cell Mol Med. 2007;11:502–508. doi:10.1111/j.1582-4934.2007.00041.x
58. Quevedo HC, Hatzistergos KE, Oskouei BN, Feigenbaum GS, Rodriguez JE, Valdes D. Allogeneic mesenchymal stem cells restore cardiac function in chronic ischemic cardiomyopathy via trilineage differentiating capacity. Proc Natl Acad Sci. 2009;106:14022–14027. doi:10.1073/pnas.0903201106
59. Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV. Osteogenesis in transplants of bone marrow cells. J Embryol Exp Morphol. 1966;16:381–390.
60. Ullah I, Subbarao RB, Rho GJ. Human mesenchymal stem cells—current trends and future prospective. Biosci Rep. 2015;28:35.
61. Salgado AJ, Reis RL, Sousa NJ, Gimble JM. Adipose tissue derived stem cells secretome: soluble factors and their roles in regenerative medicine. Curr Stem Cell Res Ther. 2010;5:103–110. doi:10.2174/157488810791268564
62. Jimenez-Puerta GJ, Marchal JA, López-Ruiz E, Gálvez-Martín P. Role of mesenchymal stromal cells as therapeutic agents: potential mechanisms of action and implications in their clinical use. J Clin Med. 2020;9(2):445th ser. doi:10.3390/jcm9020445
63. Kot M, Musiał-Wysocka A, Lasota M, Ulman A, Majka M. Secretion, migration and adhesion as key processes in the therapeutic activity of mesenchymal stem cells. Acta Biochim Pol. 2019;66(4):499–507.
64. Sotiropoulou PA, Perez SA, Salagianni M, Baxevanis CN, Papamichail M. Characterization of the optimal culture conditions for clinical scale production of human mesenchymal stem cells. Stem Cells. 2006;24:462–471. doi:10.1634/stemcells.2004-0331
65. Lee KA, Shim W, Paik MJ, et al. Analysis of changes in the viability and gene expression profiles of human mesenchymal stromal cells over time. Cytotherapy. 2009;11(6):688–697. doi:10.3109/14653240902974032
66. Schoeberlein A, Mueller M, Reinhart U, Sager R, Messerli M, Surbek DV. Homing of placenta-derived mesenchymal stem cells after perinatal intracerebral transplantation in a rat model. Am J Obstet Gynecol. 2011;205(277):e1–e6. doi:10.1016/j.ajog.2011.06.044
67. Fossett E, Khan W. Optimising human mesenchymal stem cell numbers for clinical application: a literature review. Stem Cells Int. 2012;2012:1–5. doi:10.1155/2012/465259
68. Francois S, Mouiseddine M, Allenet-Lepage B. Human mesenchymal stem cells provide protection against radiation-induced liver injury by antioxidative process, vasculature protection, hepatocyte differentiation, and trophic effects. Biomed Res Int. 2013;2013:1–14. doi:10.1155/2013/151679
69. Halabian R, Tehrani HA, Jahanian-Najafabadi A, Roudkenar MH. Lipocalin-2-mediated upregulation of various antioxidants and growth factors protects bone marrow derived mesenchymal stem cells against unfavorable microenvironments. Cell Stress Chaperones. 2013;18:785–800. doi:10.1007/s12192-013-0430-2
70. Han SM, Han SH, Coh YR. Enhanced proliferation and differentiation of Oct4-and Sox2-overexpressing hu man adipose tissue mesenchymal stem cells. Exp Mol Med. 2014;46:e101. doi:10.1038/emm.2014.28
71. Amiri F, Jahanian-Najafabadi A, Roudkenar MH. In vitro augmentation of mesenchymal stem cells viability in stressful microenvironments. Cell Stress Chaperones. 2015;20:237–251. doi:10.1007/s12192-014-0560-1
72. Hagberg H, Mallard C, Ferriero DM. The role of inflammation in perinatal brain injury. Nat Rev Neurol. 2015;11:192. doi:10.1038/nrneurol.2015.13
73. Mueller M, Wolfs TG, Schoeberlein A, Gavilanes AW, Surbek D, Kramer BW. Mesenchymal stem/ stromal cells—a key mediator for regeneration after perinatal morbidity. Mol Cell Pediatr. 2016;3:6. doi:10.1186/s40348-016-0034-x
74. Raijmakers MT, Roes EM, Poston L, Steegers EA, Peters WH. The transient increase of oxidative stress during normal pregnancy is higher and persists after delivery in women with pre-eclampsia. Eur J Obstet Gynecol Reprod Biol. 2008;138(1):39–44. doi:10.1016/j.ejogrb.2007.08.005
75. Ashok A, Nabil A, Botros R. Studies on Women’s Health: Oxidative Stress in Applied Basic Research and Clinical Practice. Agarwal A, Aziz N, Rizk B, editors. New York: Humana Press; 2013.
76. Basmaeil Y, Rashid MA, Khatlani T, et al. Preconditioning of human decidua basalis mesenchymal stem/stromal cells with glucose increased their engraftment and anti-diabetic properties. Tissue Eng Regen Med. 2020;17(2):209.
77. Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8(4):315–317.
78. Peterson KM, Aly A, Lerman A, Lerman LO, Rodriguez-Porcel M. Improved survival of mesenchymal stromal cell after hypoxia preconditioning: role of oxidative stress. Life Sci. 2011;88(1–2):65–73. doi:10.1016/j.lfs.2010.10.023
79. Lai P, Li T, Yang J, et al. Upregulation of stromal cell-derived factor 1 (SDF-1) expression in microvasculature endothelial cells in retinal ischemia-reperfusion injury. Graefes Arch Clin Exp Ophthalmol. 2008;246(12):1707–1713. doi:10.1007/s00417-008-0907-3
80. Richmond A. Chemokine modulation of the tumor microenvironment. Pigment Cell Melanoma Res. 2010;23(3):312–313. doi:10.1111/j.1755-148X.2010.00714.x
81. Silva LHA, Antunes MA, Dos Santos CC, Weiss DJ, Cruz FF, Rocco PRM. Strategies to improve the therapeutic effects of mesenchymal stromal cells in respiratory diseases. Stem Cell Res Ther. 2018;9(1):45. doi:10.1186/s13287-018-0802-8
82. Zhao L, Hu C, Han F, Cai F, Wang J, Chen J. Preconditioning is an effective strategy for improving the efficiency of mesenchymal stem cells in kidney transplantation. Stem Cell Res Ther. 2020;11(1):197. doi:10.1186/s13287-020-01721-8
83. Kurte M, Vega-Letter AM, Luz-Crawford P, et al. Time-dependent LPS exposure commands MSC immunoplasticity through TLR4 activation leading to opposite therapeutic outcome in EAE. Stem Cell Res Ther. 2020;11(1). doi:10.1186/s13287-020-01840-2
84. Durand N, Russell A, Zubair AC. Effect of comedications and endotoxins on mesenchymal stem cell secretomes, migratory and immunomodulatory capacity. J Clin Med. 2019;8(4):497. doi:10.3390/jcm8040497
85. Goswami R, Kaplan MH. A brief history of IL-9. J Immunol. 2011;186:3283–3288. doi:10.4049/jimmunol.1003049
86. Sun Y, Liu S, Hu R, Zhou Q, Li X. Decreased placental IL9 and IL9R in preeclampsia impair trophoblast cell proliferation, invasion, and angiogenesis. Hypertens Pregnancy. 2020;39(3):228–235.
87. Bigildeev AE, Zezina EA, Shipounova IN, Drize NJ. Interleukin-1 beta enhances human multipotent mesenchymal stromal cell proliferative potential and their ability to maintain hematopoietic precursor cells. Cytokine. 2015;71(2):246–254. doi:10.1016/j.cyto.2014.10.018
88. Matsumura E, Tsuji K, Komori K, Koga H, Sekiya I, Muneta T. Pretreatment with IL-1β enhances proliferation and chondrogenic potential of synovium-derived mesenchymal stem cells. Cytotherapy. 2017;19(2):181–193. doi:10.1016/j.jcyt.2016.11.004
89. Amann EM, Groß A, Rojewski MT, et al. Inflammatory response of mesenchymal stromal cells after in vivo exposure with selected trauma-related factors and polytrauma serum. PLoS One. 2019;14(5):e0216862. doi:10.1371/journal.pone.0216862
90. Cui TX, Brady AE, Fulton CT, et al. CCR2 mediates chronic LPS-induced pulmonary inflammation and hypoalveolarization in a murine model of bronchopulmonary dysplasia. Front Immunol. 2020;11. doi:10.3389/fimmu.2020.579628
91. Wynn RF, Hart CA, Corradi-Perini C, O’Neill L, Evans CA, Wraith JE. A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow. Blood. 2004;104:2643–2645. doi:10.1182/blood-2004-02-0526
92. Sordi V, Malosio ML, Marchesi F, Mercalli A, Melzi R, Giordano T. Bone marrow mesenchymal stem cells express a restricted set of functionally active chemokine receptors capable of promoting migration to pancreatic islets. Blood. 2005;106:419–427. doi:10.1182/blood-2004-09-3507
93. Honczarenko M, Le Y, Swierkowski M, Ghiran I, Glodek AM, Silberstein LE. Human bone marrow stromal cells express a distinct set of biologically functional chemokine receptors. Stem Cells. 2005;24:1030–1041. doi:10.1634/stemcells.2005-0319
94. Von Luttichau I, Notohamiprodjo M, Wechselberger A, Peters C, Henger A, Seliger C. Human adult CD34- progenitor cells functionally express the chemokine receptors CCR1, CCR4, CCR7, CXCR5, and CCR10 but not CXCR4. Stem Cells Dev. 2005;14:329–336. doi:10.1089/scd.2005.14.329
95. Ringe J, Strassburg S, Neumann K, Endres M, Notter M, Burmester GR. Towards in situ tissue repair: human mesenchymal stem cells express chemokine receptors CXCR1. CXCR2 and CCR2, and migrate upon stimulation with CXCL8 but not CCL2. J Cell Biochem. 2007;101:135–146. doi:10.1002/jcb.21172
96. Ma J, Liu N, Yi B, Zhang X, Gao BB, Zhang Y. Transplanted hUCB-MSCs migrated to the damaged area by SDF-1/CXCR4 signaling to promote functional recovery after traumatic brain injury in rats. Neurol Res. 2015;37:50–56. doi:10.1179/1743132814Y.0000000399
97. Wobus M, List C, Dittrich T, Dhawan A, Duryagina R, Arabanian LS. Breast carcinoma cells modulate the chemoattractive activity of human bone marrow-derived mesenchymal stromal cells by interfering with CXCL12. Int J Cancer. 2014;136:44–54. doi:10.1002/ijc.28960
98. Shen Z, Wang J, Huang Q, et al. Genetic modification to induce CXCR2 overexpression in mesenchymal stem cells enhances treatment benefits in radiation-induced oral mucositis. Cell Death Dis. 2018;9(2). doi:10.1038/s41419-018-0310-x
99. Meirelles LDS, Fontes AM, Covas DT, Caplan AI. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009;20:419–427. doi:10.1016/j.cytogfr.2009.10.002
100. Escacena N, Quesada-Hernandez E, Capilla-Gonzalez V, Soria B, Hmadcha A. Bottlenecks in the efficient use of advanced therapy medicinal products based on mesenchymal stromal cells. Stem Cells Int. 2015;2015:895714.
101. Hmadcha A, Martin-Montalvo A, Gauthier BR, Soria B, Capilla-Gonzalez V. Therapeutic potential of mesenchymal stem cells for cancer therapy. Front Bioeng Biotechnol. 2020;8. doi:10.3389/fbioe.2020.00043
102. Di Nicola M, Carlo-Stella C, Magni M, et al. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood. 2002;99:3838–3843. doi:10.1182/blood.V99.10.3838
103. Nauta AJ, Kruisselbrink AB, Lurvink E, Willemze R, Fibbe WE. Mesenchymal stem cells inhibit generation and function of both CD34+-derived and monocyte-derived dendritic cells. J Immunol. 2006;177:2080–2087. doi:10.4049/jimmunol.177.4.2080
104. Nemeth K, Keane-Myers A, Brown JM, et al. Bone marrow stromal cells use TGF-beta to suppress allergic responses in a mouse model of ragweed-induced asthma. Proc Natl Acad Sci. 2010;107:5652–5657. doi:10.1073/pnas.0910720107
105. Du-Rocher B, Binato R, de-freitas-junior JCM, et al. IL-17 triggers invasive and migratory properties in human MSCs, while IFNγ favors their immunosuppressive capabilities: implications for the “licensing” process. Stem Cell Rev Rep. 2020;16(6):1266–1279. doi:10.1007/s12015-020-10051-4
106. Albanesi C, Cavani A, Girolomoni G. IL-17 is produced by nickel-specific T lymphocytes and regulates ICAM-1 expression and chemokine production in human keratinocytes: synergistic or antagonist effects with IFN-gamma and TNF-alpha. J Immunol. 1999;162(1):494–502.
107. Eid RE, Rao DA, Zhou J, et al. Interleukin-17 and interferon-γ are produced concomitantly by human coronary artery-infiltrating T cells and act synergistically on vascular smooth muscle cells. Circulation. 2009;119(10):1424–1432. doi:10.1161/CIRCULATIONAHA.108.827618
108. Bulek K, Liu C, Swaidani S, et al. The inducible kinase IKKi is required for IL-17-dependent signaling associated with neutrophilia and pulmonary inflammation. Nat Immunol. 2011;12(9):844–852. doi:10.1038/ni.2080
109. Gabr MA, Jing L, Helbling AR, et al. Interleukin-17 synergizes with IFNγ or TNFα to promote inflammatory mediator release and intercellular adhesion molecule-1 (ICAM-1) expression in human intervertebral disc cells. J Orthop Res. 2011;29(1):1–7. doi:10.1002/jor.21206
110. Ekström K, Omar O, Granéli C, Wang X, Vazirisani F, Thomsen P. Monocyte exosomes stimulate the osteogenic gene expression of mesenchymal stem cells. PLoS One. 2013;8(9):e75227. doi:10.1371/journal.pone.0075227
111. Kang Q, Song WX, Luo Q, et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009;18(4):545–559. doi:10.1089/scd.2008.0130
112. Chen G, Deng C, Li YP. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int J Biol Sci. 2012;8:272–288. doi:10.7150/ijbs.2929
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.