Back to Journals » Journal of Pain Research » Volume 13
Parietal Block Using Bupivacaine in the Rat: An Anatomical, Behavioral and Histological Study
Authors Kfoury TT ![]() , Ben Rehouma M, Zetlaoui PJ, Benhamou D, Mazoit JX
, Ben Rehouma M, Zetlaoui PJ, Benhamou D, Mazoit JX
Received 29 April 2019
Accepted for publication 2 August 2019
Published 8 January 2020 Volume 2020:13 Pages 17—24
DOI https://doi.org/10.2147/JPR.S213820
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Überall
Toni T Kfoury,1,2 Mouna Ben Rehouma,2 Paul J Zetlaoui,1 Dan Benhamou,1 Jean Xavier Mazoit1,2
1Département d’Anesthésie-Réanimation, Hôpitaux Universitaires Paris-Sud AP-HP, Paris, France; 2Laboratoire d’Anesthésie, Faculté de Médecine, INSERM UMR 1195, Université Paris-Sud, Paris, France
Correspondence: Jean Xavier Mazoit
Laboratoire d’Anesthésie, Faculté de Médecine, INSERM UMR 1195, Université Paris-Sud, 63 Rue Gabriel Péri, Bicêtre Cedex 94276, France
Email [email protected]
Purpose: Animal models of regional anaesthesia are useful for studying the effects of blocks and improve their efficacy. The aim of our experiments was to validate a multi-site paravertebral block in the rat.
Material and methods: Dissection and indigo carmine dye injection were performed in five rats (3 rats were dissected and 2 were dye injected). In other groups (n=7rats/group), after inflammation inductive carrageenan injection in the abdominal wall, bupivacaine or saline was injected laterally to the spinal column at the T5, T10, L1, L4 and S1 level. The efficacy of the block on mechanical nociception was measured using von Frey hairs. In addition, we measured c-Fos immunoreactive nuclei in the cord.
Results: The multi-site injection showed a perinervous distribution of the injected solution without intra-thoracic, intra-abdominal or epidural diffusion. Bilateral block with a relatively small volume of bupivacaine (0.5 mL) significantly increased the threshold to mechanical pain as compared to control (p=0.007) and significantly decreased the number of c-Fos immunoreactive nuclei in the posterior horn of the spinal cord (p<0.0001).
Conclusion: This study shows that a parietal abdominal wall block is easy to perform in the rat. This block allows investigators to explore the mechanisms of action of abdominal parietal wall blocks.
Keywords: parietal block, rat, animal model
Introduction
Parietal abdominal nerve blocks improve analgesia in patients undergoing abdominal, gynecological and urological surgery.1–5 During abdominal surgery, pain has several components; the first is due to abdominal wall injury, and the second and third are related to visceral and parietal peritoneal inflammation, respectively.6–8
Animal models of regional anesthesia are useful for studying the effects of the blocks on nociception, behavior and inflammation.1,9 With that respect, models have been developed in rodents.9,10 However, because of anatomical, behavioral and biochemical differences between humans and rodents, it is necessary to perform experiments in the animal before any extrapolation to human should be made.9–12 In addition, discrimination between the effects of sensory block and anti-inflammatory properties of local anesthetics needs adapted models aiming at discriminate between different mechanisms. Rats (usually of the Sprague-Dawley strain) are the most commonly used animals to study pain because of their size adapted to behavioral experiments.9 Thalhammer et al performed a neurologic evaluation of the rat during sciatic nerve block.10 They showed that this strain was an excellent model for the study of behavior after the realization of a nerve block.
Because anatomical differences between rats and humans at the spine and abdominal wall muscles level may induce discrepancies between human and rat models,11–13 we wanted to validate a new model of abdominal wall regional block in the rat. We performed a multi-injection block from T5 to S1. In the first part, we studied the anatomy of parietal wall innervation and in a second part, we studied the effect of the block on an inflammatory injury of the abdominal wall using bupivacaine.
Methods
This placebo-controlled experiment was performed on male Sprague-Dawley rats weighing 175 to 225 g (Elevage Janvier, Le Genest-St-Isle, France). Rats were housed with food and water available ad libitum and maintained on a 12-hrs light-dark cycle. Rats were handled repeatedly over at least 3 days before experiments to habituate them to investigators and to the testing paradigm. Ethical approval for this study was provided by the Comité d’Ethique en Expérimentation animale N°26 (CEEA 26, Paris-Sud, N° 2012_089). Experiments followed the ARRIVE (Animal Research Reporting of In Vivo Experiments) guidelines. After testing, all animals were euthanized using an overdose of pentobarbital sodium.
Drugs and Chemicals
Bupivacaine 0.25% with epinephrine 1:200,000 was from Mylan, Saint Priest, France; carrageenan (2.5% wt/vol solution of lambda carrageenan in saline) was from Sigma-Aldrich, Saint Quentin Fallavier France; indigo carmine was from SERB laboratories, Paris, France; pentobarbital sodium was from Centravet, Taden, France. Buffers and other chemicals were from Sigma-Aldrich, Saint Quentin Fallavier France.
Anatomical Study
Rats were anesthetized with intraperitoneal (i.p.) pentobarbital, 50mg/kg. The abdominal wall and the back were shaved and the block was performed with the rat in a ventral position (on their belly). Injections were performed on both sides of the vertebral column 3 mm lateral to the spinal process of T5, T10, L1, L4 and S1 (Figure 1). The last rib, and then, the spinal process of T13 vertebra were first identified by palpation. The spinal processes of T5, T10, L1, L4 and S1 vertebra were identified and the skin in front of these vertebras was labeled with a skin pencil. Indigo carmine (1:6000 in normal saline) was injected on both sides of each labeled vertebra in a perpendicular direction to the skin using a 23 gauge needle. The solution was injected once the needle crossed the first muscular layer. Thoracic and abdominal cavities of the rats were systematically opened to check any solution diffusion.
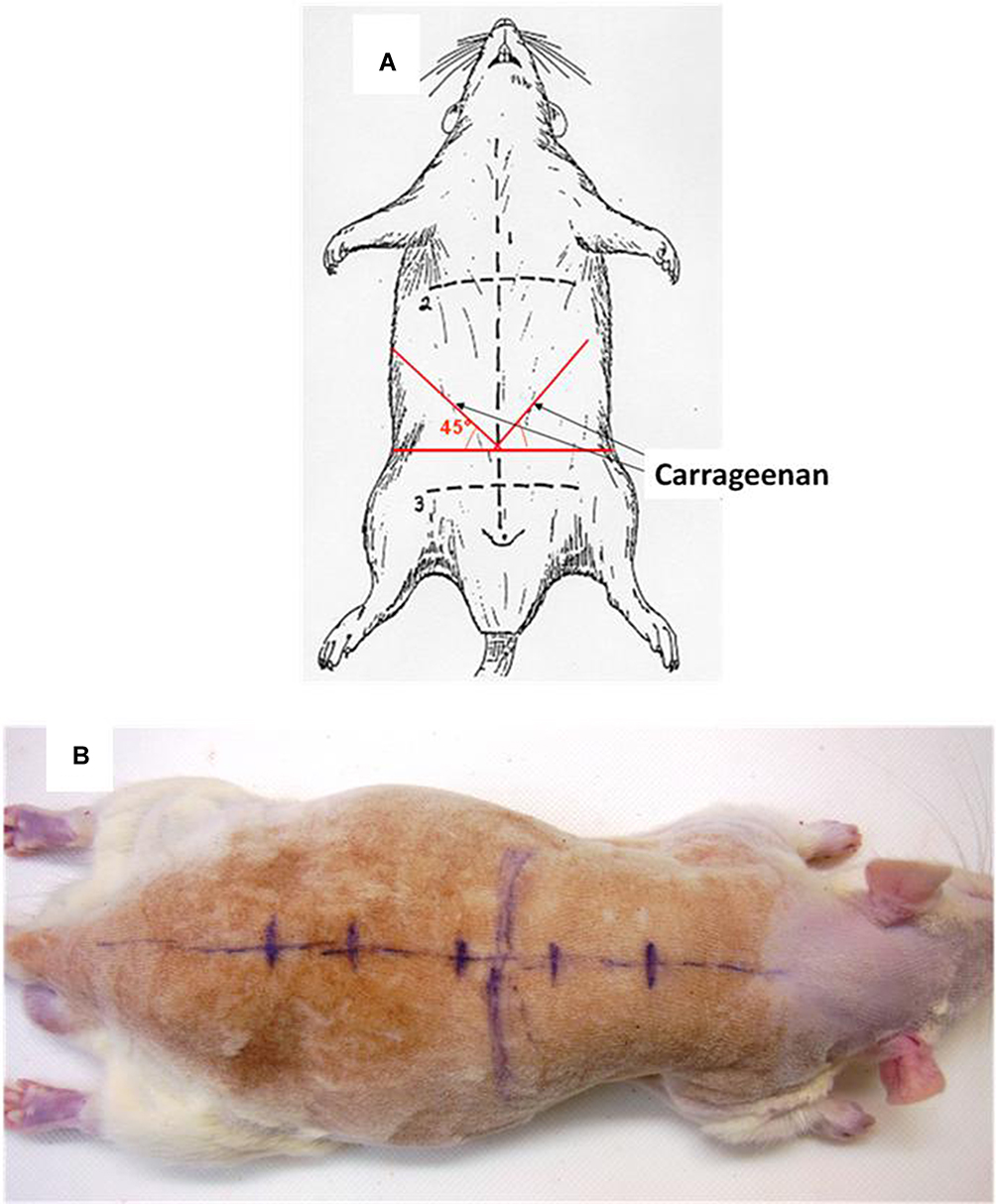 |
Figure 1 (A) Injection sites of carrageenan in the abdominal wall. Carrageenan or saline was injected at a point 1/3 from midline on the bisector of the angle formed by the intersection between the line joining the two anterior superior iliac crests (horizontal line, mark 3) and the linea Alba (mark 1). (B) Landmarks for the parietal abdominal nerve block; Spine, ribs and vertebrae (T5, T10, L1, L4 and S1) are marked with a pen. |
Abdominal Wall Block and Carrageenan Injection
Rats were anesthetized using pentobarbital sodium 50 mg/kg i.p. Rats were not shaved. The block was performed as previously described and bupivacaine was equally distributed between injection sites. Once bupivacaine was injected, the rat was settled on the back. Carrageenan was injected using a 23-gauge needle inserted at the intersection between the line joining the two anterior superior iliac crests and the linea Alba and directed on a cranial and lateral way. This point is situated on the bisector of the angle formed by both lines already described on each side of the abdominal wall (Figure 1). 0.5 mL freshly prepared carrageenan was injected on each side of the abdominal wall between abdominal wall layers.
Experimental Groups
Five adult Sprague-Dawley rats were included in the anatomical study. For the behavioral experiment, 14 animals (n = 7/group) were randomly assigned to receive saline (control group) or bupivacaine 15 mg/kg for the block.14 The same pattern of the experiment was repeated for c-Fos study using 16 animals (n = 8/group). The number of animals needed was calculated considering a power of 80% and a type 1 error of 5% (bilateral). Expecting a Cohen D of 1.5 for mechanical pain experiment and of 0.6 for cFos experiment, seven and eight animals/group were considered, respectively.
Parietal Pain Assessment
Mechanical pain was assessed by the application of calibrated von Frey filaments 6 hrs after inflammatory insult once the rats had well recovered from anesthesia. von Frey filaments were applied vertically to the abdominal wall on two different points on both sides of the abdominal wall where carrageenan was injected. Testing began with a small (10 g/mm2) filament. The filaments were applied in increasing order until the abdominal wall contraction was elicited, which was considered as a positive response. The maximum force applied was 137.3 g.mm-2. The contraction threshold was determined three times, at 10-mins intervals, and the mean contraction threshold was used for data analysis.
c-Fos Experiment
Two hours following carrageenan administration, rats were deeply anaesthetized with sodium pentobarbital (50 mg kg−1 i.p.) and perfused intracardially with 300 mL of 0.6 M phosphate-buffered saline (PBS) followed by 300 mL of ice-cold fixative (4% paraformaldehyde in 0.6 M phosphate buffer – PAF 4%) at a rate of 1 mL s−1. The spinal cord was removed and postfixed in the same fixative overnight. The spinal cord was then cut into slices of two metamers before paraffin inclusion. Serial transverse sections of 8 µm were cut using a vibratom (Thermo Microtome, Microm HM340E, Thermo Fischer, Les Ulis France). After wax removal and rehydration, the sections were washed twice with 0.15 M PBS solution and immersed in 2% H2O2 for 30 min. The sections were pre-treated with 1% normal goat serum (Vector Laboratories, Eurobio Courtaboeuf France) and 0.3% Triton X-100 in 0.15 M PBS (NGST) for 2 hrs, then incubated for 20 hrs at room temperature with rabbit polyclonal antibody directed against c-Fos protein at a concentration of 50 µg/mL (Merck Millipore, Fontenay sous bois, France). The incubated sections were washed three times in PBS and incubated in biotinylated goat anti-rabbit 1:500 (Vector Laboratories, Eurobio Courtaboeuf France) in NGST for 1 hr at room temperature, then washed twice in PBS and incubated for 1 hr in avidin–biotin-peroxidase complex (Vectastain Elite ABC PK-6100, Vector Laboratories, Eurobio Courtaboeuf France). Finally, the sections were developed in diaminobenzidine (0.035%) solution (SK-4600; Vector Laboratories, Eurobio Courtaboeuf France). The sections were mounted on adhesion microscope slides SuperFrost plus (Roth laboratories, Lauterbourg, France), and coverslipped. Spinal cord sections from the different experimental conditions were processed concurrently to control for variability in staining intensity. Four sections corresponding to each two-metamer segment and exhibiting the greatest number of labeled cells were selected from each rat. The selection of sections and quantification of the number of c-Fos positive cells in the superficial (I-II) and deep (V-VI) laminae of the dorsal horn were performed by an investigator blind to the experimental condition. c-Fos positive cells were localized under light-field microscopy at 10x et 40x. Labeled nuclei were counted using a microscope (Nikon, Eclipse E2000) regardless of staining intensity.
Statistical Analysis
The mechanical withdrawal threshold after von Frey hairs application was compared between groups using the Mann–Whitney test. The number of c-Fos reactive nuclei in the spinal cord was compared between groups using the Kruskal–Wallis test followed by the Dunn test with Holm’s correction. Data are reported as medians and centiles.
Results
Anatomical Study (Figures 1 and 2)
The dissection of three adult rats confirmed that innervation of the abdominal wall (including the anterior pelvic wall) arises from T5 to S1. Nerves pass under the paravertebral muscles before entering the abdominal wall between the transversus abdominis and the internal oblique abdominal muscles (Figure 2A).
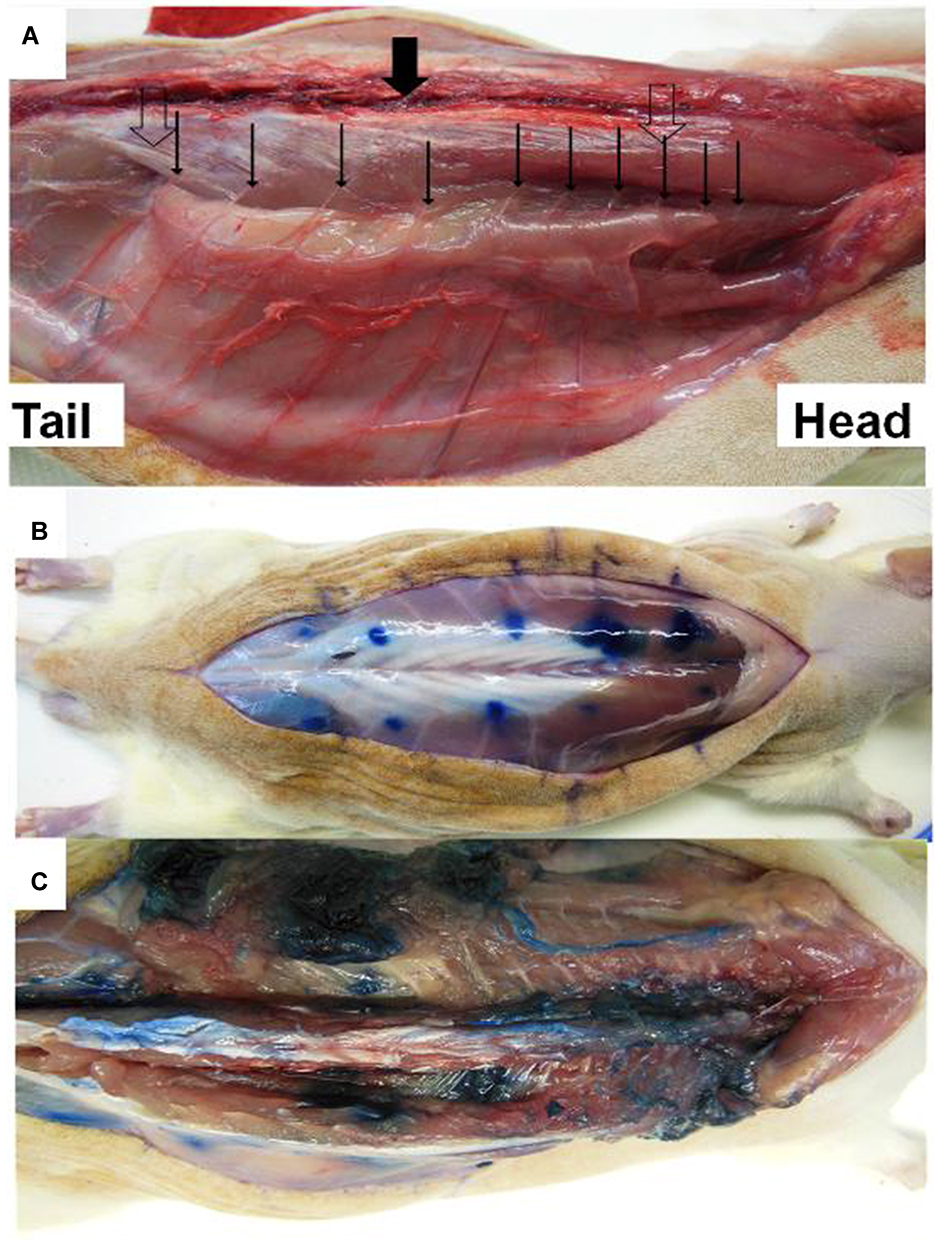 |
Figure 2 (A) Nerves (large clear arrows) emerging from the spine (large solid arrow). Nerves pass under the paravertebral muscles (thin solid arrows) before entering the abdominal wall between parietal muscles. (B) Blue dots show injection location for parietal abdominal nerve block injection. (C) Diffusion of indigo carmine dye solution after muscular dissection. |
After the injection of indigo carmine dye, the dissection of two other rats showed that injection below paravertebral muscles insures a perinervous distribution of the injected solution (Figure 2C). A multi-site injection with a relatively small volume allows a perinervous distribution of the injected solution without intra-thoracic, intra-abdominal or epidural diffusion.
Parietal Pain Assessment
Mechanical withdrawal thresholds: All animals in both groups reached the threshold of 137g/mm2 before carrageenan injection. Six hours after inflammatory insult induction, animals in the bupivacaine group showed a significantly higher withdrawal threshold than those in the control group (Figure 3A).
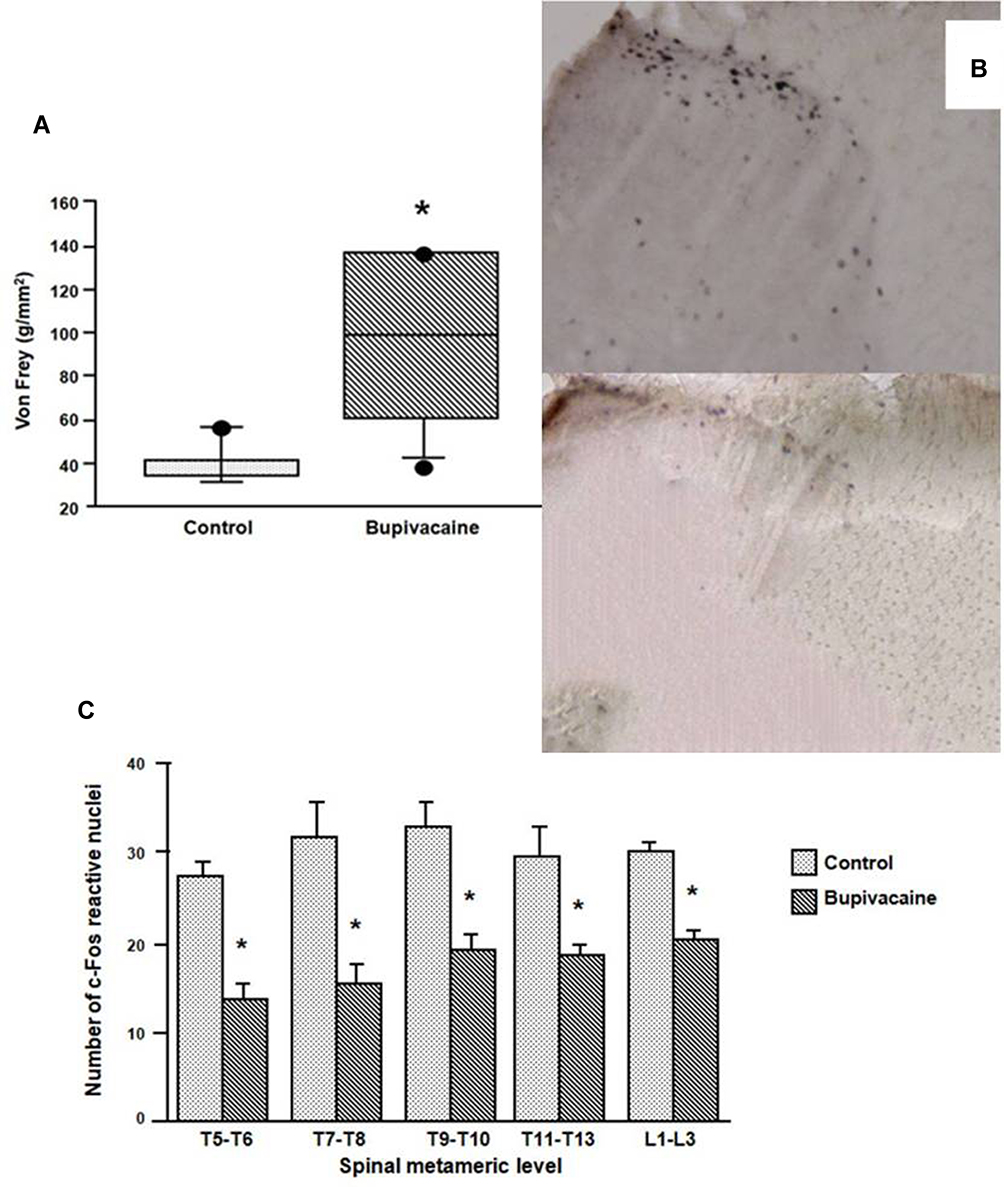 |
Figure 3 (A) Mechanical pain threshold in the two groups after parietal nerve block. The bupivacaine group shows a significant increase in threshold as compared to the placebo group. Box plots are median, 10th, 25th, 75th and 90th percentile, dots are lower and upper range. *p=0.007 vs control. (B) c-Fos reactive nuclei 2 hrs after injury in the dorsal horn at the T9-T10 level in two representative animals (control group in the upper part and bupivacaine group in the lower part). (C) Number of c-Fos immunoreactive nuclei in the spinal cord measured at different metameric levels. c-Fos reactive nuclei were significantly less numerous in the bupivacaine group than in the control group from T5 to L3. *p<0.0001 bupivacaine group vs control group. |
Immunohistological Staining
The expression of c-Fos at different levels of the spinal cord was significantly inhibited in rats receiving bupivacaine in comparison with rats receiving normal saline in their parietal block (Figure 3B and C). No statistical difference between laminae I-II and V-VI was observed.
Discussion
The present observation of the abdominal wall innervation corresponds to the descriptions of Greene and of Rizk.11,12 These authors described marked anatomical differences between humans and rats. This finding allows us to imagine and propose a multisite injection nerve block of the abdominal wall in the rat. The small volume of local aesthetic solution injected at each site guarantees a limited distribution to the perineural space (across nerves). It is likely that the use of larger local aesthetic volume may induce the risk of intra-abdominal, intra-thoracic or even epidural diffusion.
Carrageenan has been widely used to induce an inflammatory insult in both peripheral and peritoneal pain model.15–18 In a peripheral pain model, hyperalgesia at the site of injection and increased c-Fos immunoreactive nuclei in the spinal cord were recorded 2 hrs after carrageenan injection.19 Sciatic nerve block using bupivacaine has proved efficacy to treat and prevent pain and hyperalgesia induced by carrageenan injection in rat hind paw.15 Our results show that our block with a five-site injection has an antinociceptive effect similar in intensity to that observed at the lower limb after a sciatic nerve block. c-Fos staining in the cord also confirms that the abdominal wall innervation in the rat extends from T5 to S1.
c-Fos, the protein of the proto-oncogene c-fos has been extensively used as a marker of activation of nociceptive neurons in the spinal cord for more than 20 years since Hunt et al first reported that parietal noxious stimulation to a hind paw of rats leads to a marked induction of c-Fos in superficial and deep dorsal horn neurons.20 More generally, c-Fos is a marker of neural activation following noxious stimulation and tissue injury.21 Our proposed parietal abdominal nerve block using bupivacaine decreased the number of immunoreactive c-Fos nuclei in the cord. Despite a marked difference in the spinal cord anatomy between rats and humans, the validation of this multi-site block in the rat could open the door to experimental studies, with the possibility of linking the results between the two species.
In conclusion, this study confirms that a parietal abdominal wall block is easy to perform in the rat. This block allows investigators to explore the mechanisms of action of abdominal parietal wall blocks, with possible implications in humans.
Author Contributions
All authors contributed towards data analysis, drafting and critically revising the paper, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Coutaux A, Adam F, Willer JC, Le Bars D. Hyperalgesia and allodynia: peripheral mechanisms. Joint Bone Spine. 2005;72(5):359–371. doi:10.1016/j.jbspin.2004.01.010
2. McDonnell JG, O’Donnell B, Curley G, Heffernan A, Power C, Laffey JG. The analgesic efficacy of transversus abdominis plane block after abdominal surgery: a prospective randomized controlled trial. Anesth Analg. 2007;104(1):193–197. doi:10.1213/01.ane.0000250223.49963.0f
3. Mishriky BM, George RB, Habib AS. Transversus abdominis plane block for analgesia after Cesarean delivery: a systematic review and meta-analysis. Can J Anaesth. 2012;59(8):766–778. doi:10.1007/s12630-012-9729-1
4. Baik JS, Oh AY, Cho CW, Shin HJ, Han SH, Ryu JH. Thoracic paravertebral block for nephrectomy: a randomized, controlled, observer-blinded study. Pain Med. 2014;15(5):850–856. doi:10.1111/pme.12320
5. Bhattacharjee S, Ray M, Ghose T, Maitra S, Layek A. Analgesic efficacy of transversus abdominis plane block in providing effective perioperative analgesia in patients undergoing total abdominal hysterectomy: a randomized controlled trial. J Anaesthesiol Clin Pharmacol. 2014;30(3):391–396. doi:10.4103/0970-9185.137274
6. Williams PL. Gray’s Anatomy.
7. Boeckxstaens GE, de Jonge WJ. Neuroimmune mechanisms in postoperative ileus. Gut. 2009;58(9):1300–1311. doi:10.1136/gut.2008.169250
8. Tanaka K, Matsugami T, Chiba T. The origin of sensory innervation of the peritoneum in the rat. Anat Embryol (Berl). 2002;205(4):307–313. doi:10.1007/s00429-002-0254-9
9. Le Bars D, Gozariu M, Cadden SW. Animal models of nociception. Pharmacol Rev. 2001;53(4):597–652.
10. Thalhammer JG, Vladimirova M, Bershadsky B, Strichartz GR. Neurologic evaluation of the rat during sciatic nerve block with lidocaine. Anesthesiology. 1995;82(4):1013–1025. doi:10.1097/00000542-199504000-00026
11. Greene EC. Anatomy of the Rat. Vol. XXVII. Philadelphia, PA: Transactions of the American Philosophical Society, New Series; 1935.
12. Rizk NN. A new description of the anterior abdominal wall in man and mammals. J Anat. 1980;131(Pt3):373–385.
13. Brown SH, Banuelos K, Ward SR, Lieber RL. Architectural and morphological assessment of rat abdominal wall muscles: comparison for use as a human model. J Anat. 2010;217(3):196–202. doi:10.1111/j.1469-7580.2010.01271.x
14. Kohane DS, Sankar WN, Shubina M, Hu D, Rifai N, Berde CB. Sciatic nerve blockade in infant, adolescent, and adult rats: a comparison of ropivacaine with bupivacaine. Anesthesiology. 1998;89(5):1199–1208. doi:10.1097/00000542-199811000-00021
15. Gentili ME, Mazoit JX, Samii KK, Fletcher D. The effect of a sciatic nerve block on the development of inflammation in carrageenan injected rats. Anesth Analg. 1999;89(4):979–984. doi:10.1097/00000539-199910000-00029
16. Pagano RL, Dias MA, Dale CS, Giorgi R. Neutrophils and the calcium-binding protein MRP-14 mediate carrageenan-induced antinociception in mice. Mediators Inflamm. 2002;11(4):203–210. doi:10.1080/0962935029000050
17. Beloeil H, Asehnoune K, Moine P, Benhamou D, Mazoit JX. Bupivacaine’s action on the carrageenan-induced inflammatory response in mice: cytokine production by leukocytes after ex-vivo stimulation. Anesth Analg. 2005;100(4):1081–1106. doi:10.1213/01.ANE.0000146964.05212.65
18. Beloeil H, Ji RR, Berde CB. Effects of bupivacaine and tetrodotoxin on carrageenan-induced hind paw inflammation in rats (Part 2): cytokines and p38 mitogen-activated protein kinases in dorsal root ganglia and spinal cord. Anesthesiology. 2006;105(1):139–145. doi:10.1097/00000542-200607000-00023
19. Pham-Marcou TA, Beloeil H, Sun X, et al. Antinociceptive effect of resveratrol in carrageenan-evoked hyperalgesia in rats: prolonged effect related to COX-2 expression impairment. Pain. 2008;140(2):274–283. doi:10.1016/j.pain.2008.08.010
20. Hunt SP, Pini A, Evan G. Induction of c-fos-like protein in spinal cord neurons following sensory stimulation. Nature. 1987;328(6131):632–634. doi:10.1038/328632a0
21. Gao YJ, Ji RR. c-Fos and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury?. Open Pain J. 2009;2:11–17. doi:10.2174/1876386300902010011
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.