Back to Journals » The Application of Clinical Genetics » Volume 14
Osteopontin Gene Polymorphisms rs1126616 C>T and rs1126772 A>G are Associated with Atopic Dermatitis in Polish Population
Authors Kaleta B ![]() , Lachota M, Łukaszkiewicz J
, Lachota M, Łukaszkiewicz J ![]() , Woźniacka A, Bogaczewicz J
, Woźniacka A, Bogaczewicz J
Received 7 June 2021
Accepted for publication 27 August 2021
Published 5 October 2021 Volume 2021:14 Pages 417—425
DOI https://doi.org/10.2147/TACG.S323735
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Martin Maurer
Beata Kaleta,1 Mieszko Lachota,1,2 Jacek Łukaszkiewicz,3 Anna Woźniacka,4 Jarosław Bogaczewicz5
1Department of Clinical Immunology, Medical University of Warsaw, Warsaw, Poland; 2Doctoral School, Medical University of Warsaw, Warsaw, Poland; 3Department of Biochemistry and Clinical Chemistry, Medical University of Warsaw, Warsaw, Poland; 4Department of Dermatology and Venereology, Medical University of Lodz, Lodz, Poland; 5University of Economics and Human Sciences in Warsaw, Warsaw, Poland
Correspondence: Beata Kaleta
Department of Clinical Immunology, Medical University of Warsaw, Nowogrodzka 59 Street, Warsaw, 02-006, Poland
Tel +48 600 301 690
Fax +48 502 21 59
Email [email protected]
Purpose: Atopic dermatitis (AD) is a chronic, relapsing inflammatory disease, caused by environmental and genetic factors, which lead to immunological abnormalities. Osteopontin (OPN), also named secreted phosphoprotein 1 (SPP1), is a protein involved in the pathogenesis of numerous autoimmune and inflammatory conditions. However, its role in AD has not been fully elucidated. Therefore, we aim to gain an insight into the role of OPN in AD pathogenesis through investigating its gene single nucleotide polymorphisms (SNPs) and their possible associations with disease clinical features.
Patients and Methods: A total of 182 Caucasian participants (45 AD patients and 137 gender- and age-matched controls) were studied. Genomic DNA was isolated from peripheral blood samples. Genotyping for the rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G SNPs was performed by real time polymerase chain reaction (RT-PCR).
Results: The frequency of the minor TT genotype and the T allele of rs1126616 C>T was higher in AD patients compared to controls (P = 0.019, OD = 4.86, 95% CI = 1.46– 16.20, and P = 0.047, OR = 1.77, 95% CI = 1.04– 3.00, respectively) and was associated with the higher prevalence of asthma (P = 0.017, OR = 3.73, 95% CI = 0.71– 19.67, and P = 0.004, OR = 3.96, 95% CI = 1.53– 10.25, respectively). Likewise, the minor GG genotype and the G allele of rs1126772 A>G were more frequent in AD patients (P = 0.026, OR = 3.27, 95% CI = 1.29– 8.33, and P = 0.013, OR = 1.94, 95% CI = 1.18– 3.21, respectively) and were associated with the increased incidence of asthma (P = 0.016, OR = 5.06, 95% CI = 1.14– 22.49, and P = 0.002, OR = 4.40, 95% CI = 1.71– 11.35, respectively). Furthermore, haplotype frequency estimation determined the four-loci haplotype TGCT, as a significant risk factor for AD compared to controls (P = 0.031, OR = 9.48, 95% CI = 1.23– 71.91).
Conclusion: Our results suggest that the variation in the OPN gene might be associated with AD and increased incidence of asthma in Caucasians. Further studies should be conducted to look into the possible role of OPN as a biomarker for AD.
Keywords: asthma, atopic dermatitis, gene, osteopontin, polymorphism, AD, OPN
Introduction
Atopic dermatitis (AD), also called atopic eczema, is the most common skin disease worldwide.1 It is a chronic relapsing inflammatory disease, characterized by itching, swelling, cracking, and weeping skin lesions. Environmental and genetic factors associated with immunological abnormalities have been hypothesized to play a prime role in AD pathogenesis.2 Numerous studies have suggested that impaired activation of T helper (Th)1, Th2, Th17, as well as Th22 pathways is associated with disease pathogenesis and clinical manifestations.3
Multiple studies have shown the relationship between single nucleotide polymorphisms (SNPs) of immune-related genes and risk of AD.4
Osteopontin (OPN), also named secreted phosphoprotein 1 (SPP1), a member of the small integrin-binding ligand N-linked glycoprotein (SIBLING) family proteins is expressed in numerous cells and tissues, including bone cells, immune cells, neurons, adipocytes, and many others.5 Moreover, recent investigations have demonstrated that OPN is present in the basal keratinocyte layer, in hair follicles, sebaceous glands, and sweat glands.6,7 It has been proved that OPN induces angiogenesis, is involved in wound healing and biomineralization. Moreover, OPN plays an essential role in regulation of the immune responses by stimulating dendritic cells, macrophages, and B cells, and enhancing Th17 and Th1-mediated inflammatory response.5,8 Recent reports have indicated that increased OPN expression was observed in patients with infections, cancer, diabetes, asthma, some autoimmune diseases, and many other pathological conditions.8,9 In addition, functional studies have shown that allelic variabilities of the OPN gene affect its function by modulating transcription, expression, and protein secretion.8
Although OPN’s role in the regulation of immune system, and skin biology was suggested, the association of OPN gene polymorphism and AD is not fully elucidated. Moreover, to best of our knowledge, no genetic analyses of OPN polymorphism have been done in Caucasian population with AD. Based on previous studies linking OPN expression and gene polymorphisms to numerous autoimmune diseases, we hypothesized that polymorphisms in OPN gene are associated with AD. To test this hypothesis, we determined the frequency of four SNPs of the OPN gene (rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G) in patients with AD and in healthy controls, and analyzed their possible associations with disease susceptibility and clinical manifestations. The SNP rs11226616 C>T is located in exon 7 and is a synonymous variation that modifies exonic splicing enhancer.8 Rs1126772 A>G and rs9138 A>C are SNPs in the 3′ untranslated region (3ʹ UTR). It has been documented that this region controls stability, translational activity, and nuclear export of messenger RNA (mRNA).10 The SNP rs3841116 T>G is located in 5ʹ flanking region of OPN gene and together with above three SNPs was associated with some immune-related disorders in several studies.8,9,14
Materials and Methods
Subjects and Sample Collection
Venous blood samples were collected from 45 Caucasian AD patients (28 women and 17 men) and 137 gender- and age-matched Caucasian controls (85 women and 52 men). AD was diagnosed according to the Hanifin and Rajka criteria.11 The study included patients with moderate-to-severe AD, defined as a SCORing Atopic Dermatitis (SCORAD) rating 25–50 (moderate) and > 50 (severe). Exclusion criteria for the patients’ group were the presence of skin diseases other than AD, tobacco smoking and obesity (body mass index > 30).
The control group consisted of healthy volunteers, unrelated with AD patients, with no family history of atopy (including allergic rhinitis, asthma, and/or eczema) or autoimmune diseases. AD patients were age-, and gender-matched with controls at 1:3 or 1:4 ratio. Demographic and clinical features of patients with AD and controls are summarized in Table 1.
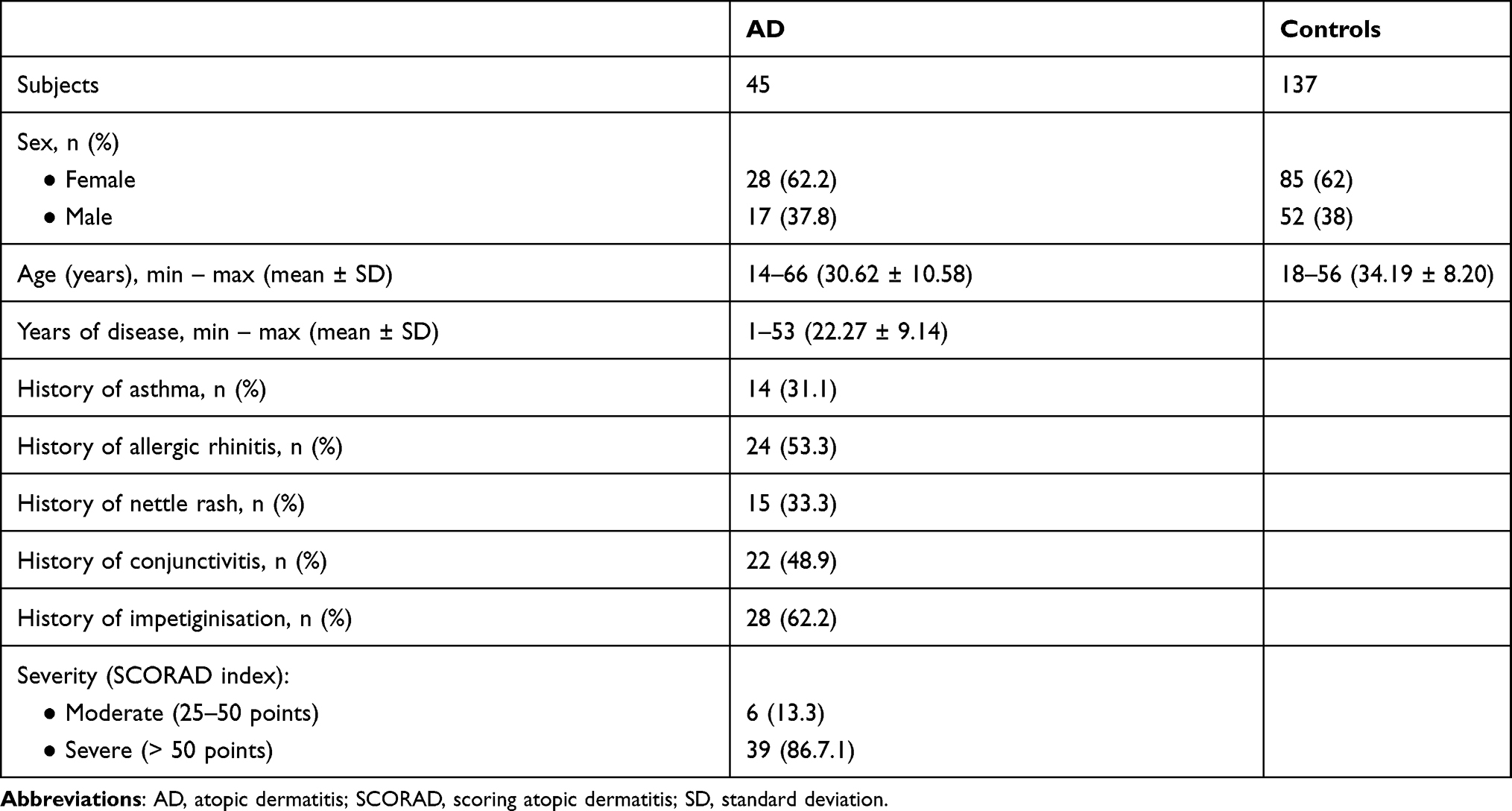 |
Table 1 Characteristics of AD Patients and Controls. Data are Expressed as Minimum – Maximum, Means ± SD or n (%) |
The study was approved by the Ethics Committee of Medical University of Lodz (No. RNN/67/08/KE) and all subjects provided written informed consent. For minor study participants statements of written informed consent from legally authorized representatives/parents were also obtained. The procedures followed were in accordance with the Helsinki Declaration of 1975, as revised in 2000.
Genetic Analysis
In the present study, we selected four SNPs (rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G) of the OPN gene from the previously published associations with autoimmune diseases,8,9,12,13 thus increasing the chance of selecting polymorphisms with phenotypic effects. Moreover, three SNPs (rs1126616 C>T, rs1126772 A>G, and rs9138 A>C) were selected because they have a minor allele frequency of >5% in Caucasian population. Another analyzed SNP, rs3841116 T>G, together with above three SNPs was associated with some immune-related disorders in several studies.8,9,14
Genomic DNA was extracted from the whole frozen blood using “Blood Mini” kit (A&A Biotechnology, Poland) according to the manufacturer’s instructions. DNA concentration and purity were determined with UV spectrophotometry. All genomic DNA samples were diluted to final concentrations of 20 ng/µL.
Rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G genotypes of the OPN were analyzed using a real time polymerase chain reaction (RT-PCR). Genotyping was carried out with the LightSNiP typing assay (TIB-MolBiol, Germany) by analyzing the melting curves with the LightCycler® 480 system available from Roche Diagnostics. All sequences of primers used for genotyping are presented in Supplemental Table S1. For all four SNPs the amplification mixture included 25ng of genomic DNA, 10µL 1x Light Cycler 480 Genotyping Master (Roche Diagnostics), 2 µL of SimpleProbe (TIB-MolBiol, Germany), containing hybridization probes and appropriate primers, and 6 µL of RNAse free water (Roche Diagnostics) in a total volume of 20 µL. RT-PCRs were performed under following conditions for all SNPs: initial denaturation at 95°C for 10 min, 45 cycles of denaturation (95°C for 10 sec), annealing at 60°C for 10 sec, extension (72°C for 15 sec). The melting curve analysis comprised an initial denaturation at 95°C for 30 sec min, lowering the temperature to 40°C for 2 min, and heating (1 acquisition/°C) increasing the temperature up to 95°C and measuring the fluorescence intensity. RT-PCR reactions were performed in 96-well plates. To confirm genotyping results, randomly selected 30 DNA samples were blindly genotyped for all 4 SNPs for the second time, and genotyping was 100% concordant.
Statistical Analysis
All genotyping results in AD patients and in controls were tested for Hardy–Weinberg Equilibrium (HWE) applying the HWSIM computer program (available at http://krunch.med.yale.edu/hwsim). Haplotype frequencies estimation and association of OPN gene haplotypes with AD were estimated using SNPStats.15 Power and sample size analysis was performed using Bioinformatics Institute’s Online Sample Size Estimator (OSSE; http://osse.bii.a-star.edu.sg/index.php). All other tests were performed by the Statistica software package, version 13.0. Allelic and genotypic frequencies were compared between AD patients and controls using the chi-square test with Yate’s correction with corresponding odds ratio (OR) and the 95% confidence interval (CI) in recessive, and allele models of inheritance. The Fisher’s exact test was used to compare the variable frequencies when the expected count was less than 5. To assess the effect of rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G genotypes on the AD severity, the ANOVA test was used. To avoid the risk of overlooking the actually existing relationships (a type II error), the Bonferroni correction was not applied.16,17 Differences were considered statistically significant at a P value < 0.05.
Results
OPN Polymorphisms Frequency
Four SNPs in the OPN gene (rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G) were successfully genotyped in 45 AD patients and 137 healthy controls. The frequencies of all four SNPs were in HWE for AD patients and controls (Supplemental Table S2), indicating that our sample population was representative. Genotypes distributions and alleles frequencies, as well as calculated ORs and 95% CI are summarized in Table 2. The distribution of CC, CT, and TT genotypes of rs1126616 C>T in the AD group was 23 (51.1%), 15 (33.3%), and 7 (15.6%), respectively, and the genotypes distribution in the control group was 85 (62%), 47 (34.3%), and 5 (3.7%), respectively. In addition, the frequencies of C and T alleles were 61 (67.8%) and 29 (32.2%) in AD patients, and 216 (78.8%) and 58 (21.2%) in controls, respectively. Statistical analysis demonstrated that the frequency of homozygous minor TT genotype of rs1126616 C>T was significantly increased in AD patients compared to healthy controls (P = 0.019, OD = 4.86, 95% CI = 1.46–16.20). Similarly, the minor T allele was more frequent in atopic patients (P = 0.047, OD = 1.77, 95% CI = 1.04–3.00).
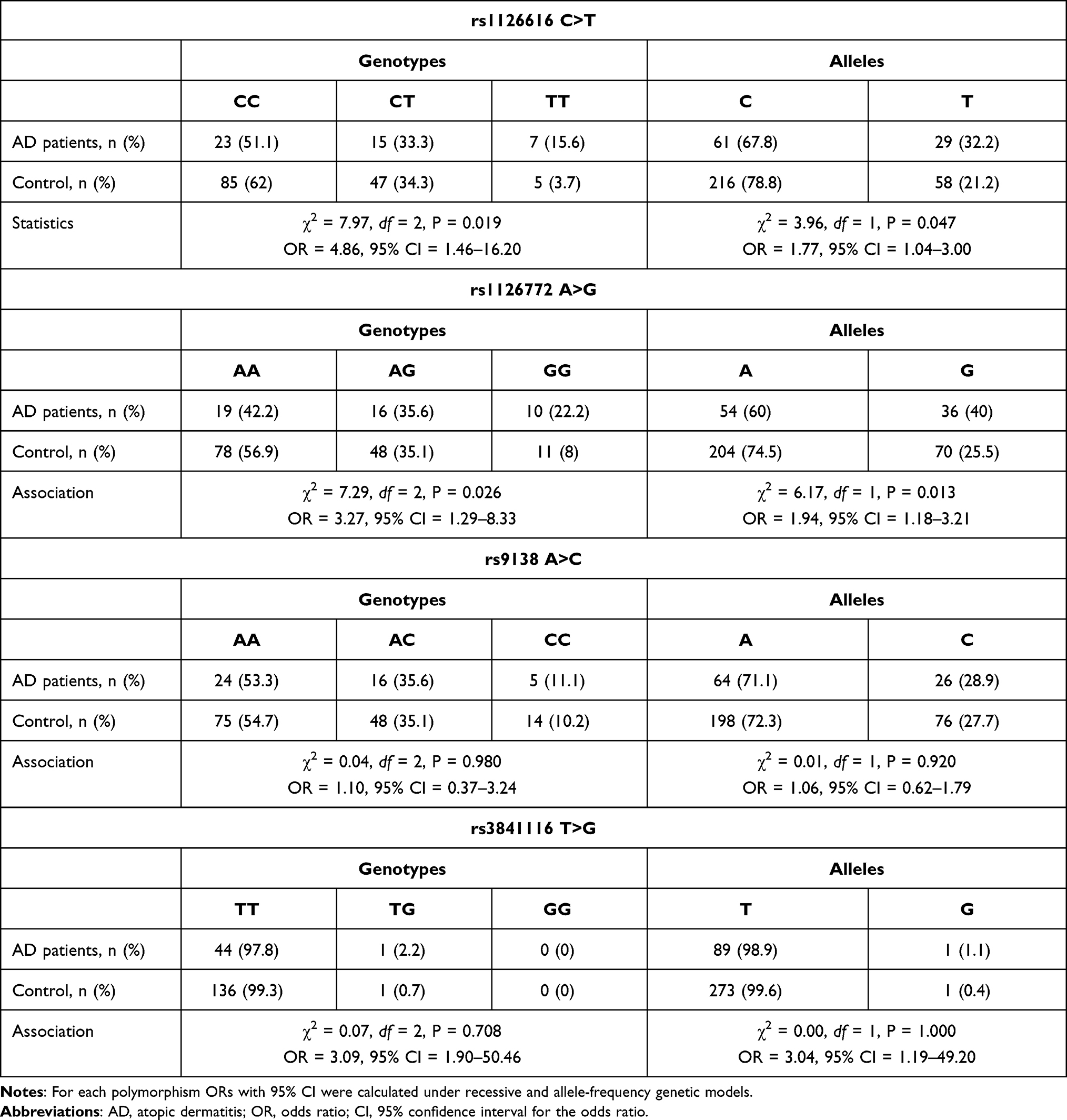 |
Table 2 Genotypes and Alleles Frequency of OPN rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G Polymorphisms in AD Patients and Controls. Data are Expressed as n (%) |
The distribution of AA, AG, and GG genotypes of rs1126772 A>G in the AD group was 19 (42.2%), 16 (35.6%), and 10 (22.2%), respectively, and the genotypes distribution in the control group was 78 (56.9%), 48 (35.1%), and 11 (8%), respectively. The frequencies of A and G alleles were 54 (60%) and 36 (40%) in AD patients, and 204 (74.5%) and 70 (25.5%) in controls, respectively. Statistical analysis demonstrated that the minor GG genotype and the G allele were more prevalent among patients than controls (P = 0.026, OD = 3.27, 95% CI = 1.29–8.33, and P = 0.013, OR = 1.94, 95% CI = 1.18–3.21, respectively). However, it is worth emphasizing that for the rs1126616 AD-association analysis, around 168 patients and 503 control probands should be genotyped to achieve a power of 80% at the desired significance level of 5% and case to control ratio of three. For rs1126772 these values are 117 and 352, respectively. Unfortunately, the numbers of study participants in AD cohort as well as the control cohort are below sample sizes sufficient to achieve the desirable statistical power.
The distribution of AA, AC, and CC genotypes of rs9138 A>C in the AD group was 24 (53.3%), 16 (35.6%), and 5 (11.1%), respectively, and the genotypes distribution in the control group was 75 (54.7%), 48 (35.1%), and 14 (10.2%), respectively. The frequencies of A and C alleles were 64 (71.1%) and 26 (28.9%) in AD patients, and 198 (72.3%) and 76 (27.7%) in controls, respectively. No significant association between this polymorphism and AD was found (for genotypes P = 0.980, OD = 1.10, 95% CI = 0.37–3.24, and for alleles P = 0.920, OD = 1.06, 95% CI = 0.62–1.79).
Similarly, genotyping results for rs3841116 T>G showed no significant differences in the comparison of genotypes and alleles frequencies between AD patients and controls (for genotypes P = 0.708, OR = 3.09, 95% CI = 1.90–50.46, and for alleles P = 1.000, OR = 3.04, 95% CI–1.19–49.20). The distribution of TT, TG, and GG genotypes of rs384116 T>G in the AD group was 44 (97.8%), 1 (2.2%), and 0 (0%), respectively, and the genotypes distribution in the control group was 136 (99.3%), 1 (0.7%), and 0 (0%), respectively. In addition, the frequencies of T and G alleles were 89 (98.9%) and 1 (1.1%) in AD patients, and 273 (99.6%) and 1 (0.4%) in controls, respectively.
Furthermore, the association between haplotypes of OPN rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G and the risk of AD was analyzed. Haplotype frequency estimation in overall population of 182 subjects showed the presence of a four-loci haplotype TGCT (rs1126616-T, rs1126772-G, rs9138-C, rs3841116-T) which frequency was significantly higher in AD patients compared to controls (P = 0.031, OR = 9.48, 95% CI = 1.23–71.91) (Supplemental Tables S3 and S4).
Influence of rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G Variants on AD Manifestations
The relationship between rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G genotypes/alleles frequencies and clinical manifestations of AD is presented in Table 3.
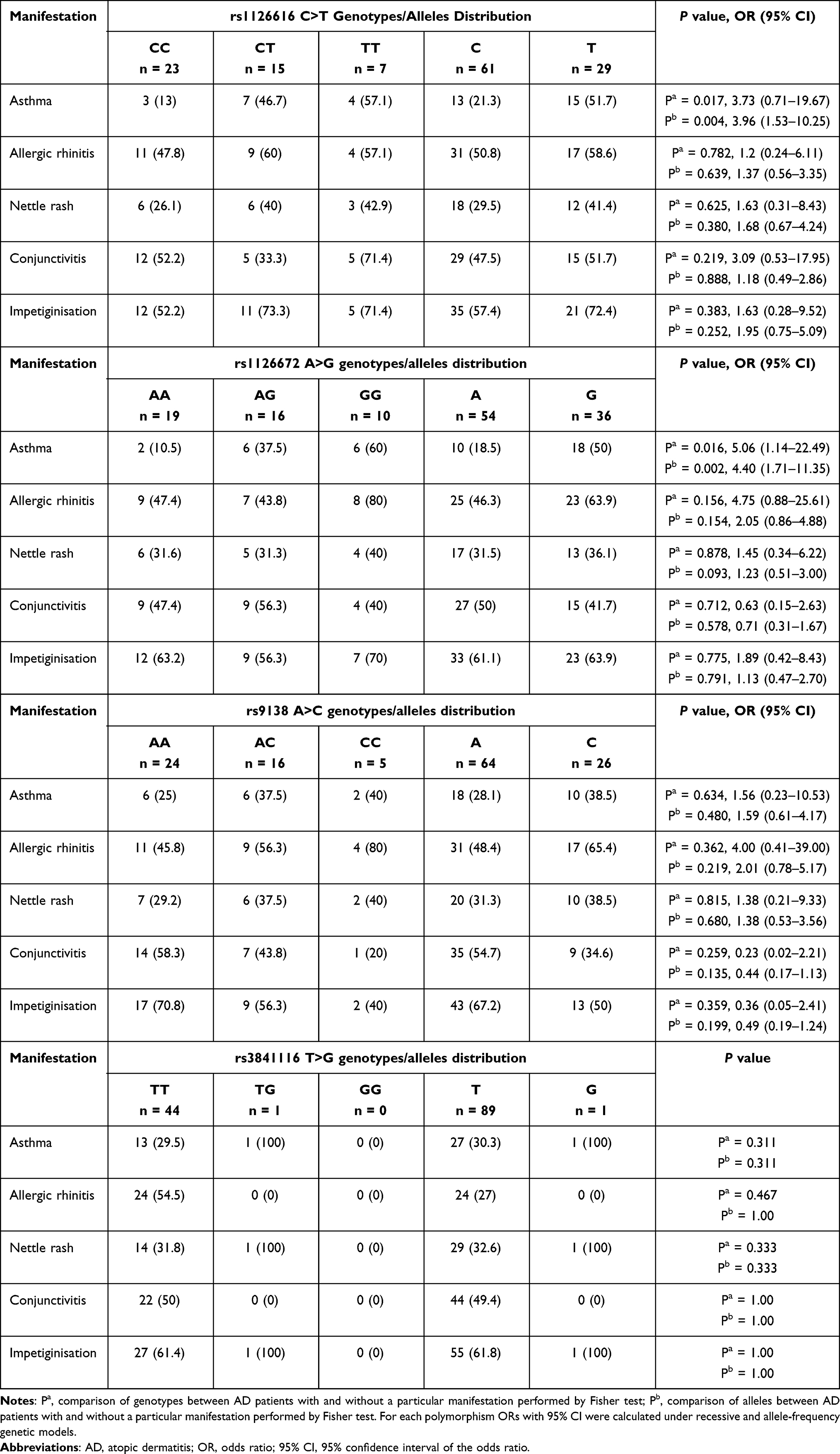 |
Table 3 Association Between Clinical Manifestations of AD and OPN rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G Genotypes/Alleles. Data are Expressed as n or n (%) |
The analysis of the distribution of rs1126616 C>T variants revealed that this polymorphism is associated with the higher frequency of asthma in AD patients. The minor TT and CT genotypes were more prevalent in AD patients with asthma compared to AD patients without asthma (P = 0.017, OR = 3.73, 95% CI – 0.71–19.67). Similarly, the T allele as more prevalent in asthmatic AD patients (P = 0.004, OR = 3.96, 95% CI = 1.53–10.25). However, no significant association of this SNP with other AD manifestations was observed.
Moreover, the minor GG/AG genotypes and the G allele of rs1126772 A>G were observed more frequently in atopic patients with asthma (P = 0.016, OR = 5.06, 95% CI = 1.14–22.49, and P = 0.002, OR = 4.40, 95% CI = 1.71–11.35, respectively). The genotyping results for rs9138 A>C, and rs3841116 T>G showed no association between these two polymorphisms and AD clinical manifestations. Furthermore, the analysis association between rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G SNPs and AD severity (assessed as SCORAD) demonstrated that these variants do not affect disease severity in the study population (P = 0.135, OR = 4.56, 95% CI = 1.99–12.65, data not showed).
Discussion
The present study was conducted to investigate the frequencies of rs1126616 C>T, rs1126772 A>G, rs9138 A>C, and rs3841116 T>G polymorphic variants of OPN gene in patients with AD and in healthy controls and their possible associations with the disease manifestations.
Genetic variations of the OPN gene affect its transcription and expression14 and have been associated with pathogenesis of numerous autoimmune and inflammatory conditions.8,11,12 The minor genotypes/alleles of three SNPs analyzed in the study (rs1126616 C>T, rs1126772 A>G, and rs9138 A>C) increase OPN mRNA stability, translational activity, and nuclear export8,10 and have been previously associated with elevated risk of autoimmune diseases.
Our results suggest that rs1126616 C>T and rs1126772 A>G polymorphisms of the OPN gene are associated with AD in Caucasians and are linked with higher incidence of asthma in AD patients. Moreover, the presence of TGCT haplotype (rs1126616-T, rs1126772-G, rs9138-C, rs3841116-T) was associated with strong AD susceptibility. Previously, the possible association of OPN SNPs and AD was examined only in two studies. Tanino et al18 analyzed five OPN SNPs in asthmatic/nonasthmatic Japanese patients with or without atopy. However, the group found no significant associations between −1687A/G, −381T/C, −94 del/G, 5891C/T and 7052T/C variants and atopy or asthma, although previous report has demonstrated that the chromosomal region of 4q24, where the OPN is mapped, is a candidate region to these pathologies.19 Our study showed that rs1126616 C>T and rs1126772 A>G polymorphisms are associated with increased prevalence of asthma in AD patients. In a similar study, Arjomandi et al20 genotyped six SNPs in OPN gene in Latino Americans, and demonstrated that rs1126616 C>T, rs1126772 A>G, and rs9138 A>C variants are associated with diagnosis and severity of asthma in Puerto Ricans, but not in Mexicans.
Our findings are also consistent with the suggested immunoregulatory role of OPN. Multiple studies revealed that OPN polymorphisms are associated with the development of inflammatory diseases, including systemic lupus erythematosus, inflammatory bowel disease, multiple sclerosis, allergy, and many other disorders.8,9 Moreover, the role of OPN in skin pathologies of immune origin was also investigated. It was found that OPN expression is elevated in Th1-mediated skin diseases, such as sarcoidosis.20–22 Lavi et al23 likewise showed that plasma OPN levels were higher in patients with sarcoidosis than in healthy subjects. In addition, the group compared the frequency of OPN rs1126616 C>T, rs1126772 A>G, and rs9138 A>C, and rs4754 C>T variants but found no differences between patients and controls. In another study Maver et al24 genotyped rs11730582 T>C, rs11728697 C>T, and rs4754 C>T OPN SNPs revealing a significant difference in genotype frequencies at rs4754 C>T in sarcoidosis patients and healthy participants. OPN and its gene polymorphisms were also implicated to participate in psoriasis. It was demonstrated that OPN expression in the skin of patients is elevated but its expression level is also associated with the severity of disease.25,26 In addition, the role of OPN variants on psoriasis susceptibility was evaluated. Abdel-Hay et al27 analyzed rs4754 C>T and rs9138 A>C, but showed no significant differences in the genotypes frequency between patients and controls. Similarly, Chen et al28 demonstrated no significantly different allelic distributions of the OPN gene between psoriasis and controls. Some authors suggested that OPN is implicated in T-cell-mediated allergic contact dermatitis, melanoma and non-melanoma skin tumors, alopecia areata, as well as skin lesions of systemic lupus erythematosus.29–32
Our study suggests an association between rs1126616 C>T and rs1126772 A>G OPN SNPs and AD susceptibility. These two polymorphisms can lead to phenotypic changes through their effect on OPN mRNA stability and processing.14 Our data indicates that the minor T allele of rs1126616 C>T, as well as the minor G allele of rs1126772 A>G are associated with increased OPN gene transcriptional activity and elevated OPN production. OPN interactions with cell surface integrins and CD44 can rapidly activate nuclear factor kappa B (NFκB) leading to enhanced cytokine production and cell survival.33 Moreover, OPN is a chemoattractant for macrophages, neutrophils, dendritic cells and natural killer cells.9 In addition, OPN it has been demonstrated that this protein is implicated in tumor necrosis factor (TNF)-α pathway and can promote the expression of adhesion molecule-1 (ICAM-1).34 ICAM-1 favors infiltration of T cells and facilitates their interactions with keratinocytes at the sites of inflammation.35 It has been also proved that OPN by regulating matrix metalloproteases (MMP) 2 and 9 function, can play a role in pathogenesis of several skin disorders, including AD.36
Conclusion
The present genetic association study suggests that OPN gene can be a risk factor for AD and may be associated with increased prevalence of asthma in atopic patients. Moreover, it has been found that TGCT haplotype in the OPN gene is associated with AD. However, the results need to be replicated on a larger sample because the numbers of study participants in AD cohort as well as the control cohort are below sample sizes sufficient to achieve a statistical power of 80%. An additional limitation of the report is that plasma OPN concentration was not measured, which could add a stronger support to the final conclusions and could be correlated with the OPN genotype-dependent differences.
The role of OPN variants in autoimmune skin diseases needs to be better explored as a diagnostic target to monitor these conditions. Advances in understanding specific SNPs may be helpful to create genetic profiles for predisposition to autoimmune diseases, including AD.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Weidinger S, Novak N. Atopic dermatitis. Lancet. 2016;387(10023):1109–1122. doi:10.1016/S0140-6736(15)00149-X
2. Hoffjan S, Stemmler S. Unravelling the complex genetic background of atopic dermatitis: from genetic association results towards novel therapeutic strategies. Arch Dermatol Res. 2015;307(8):659–670. doi:10.1007/s00403-015-1550-6
3. Werfel T, Allam JP, Biedermann T, et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J Allergy Clin Immunol. 2016;138(2):336–349. doi:10.1016/j.jaci.2016.06.010
4. Bin L, Leung DY. Genetic and epigenetic studies of atopic dermatitis. Allergy Asthma Clin Immunol. 2016;12(1):52. doi:10.1186/s13223-016-0158-5
5. Fisher LW, Torchia DA, Fohr B, Young MF, Fedarko NS. Flexible structures of SIBLING proteins, bone sialoprotein, and osteopontin. Biochem Biophys Res Commun. 2001;280(2):460–465. doi:10.1006/bbrc.2000.4146
6. Yu DW, Yang T, Sonoda T, et al. Osteopontin gene is expressed in the dermal papilla of pelage follicles in a hair-cycle-dependent manner. J Invest Dermatol. 2001;117(6):1554–1558. doi:10.1046/j.0022-202x.2001.01568.x
7. Chang PL, Harkins L, Hsieh YH, et al. Osteopontin expression in normal skin and non-melanoma skin tumors. J Histochem Cytochem. 2008;56(1):57–66. doi:10.1369/jhc.7A7325.2007
8. Subraman V, Thiyagarajan M, Malathi N, Rajan ST. OPN - revisited. J Clin Diagn Res. 2015;9:ZE10–ZE13.
9. Uede T. Osteopontin, intrinsic tissue regulator of intractable inflammatory diseases. Pathol Int. 2011;61:265–280.
10. Mazumder B, Seshadri V, Fox PL. Translational control by the 3ʹ-UTR: the ends specify the means. Trends Biochem Sci. 2003;28(2):91–98. doi:10.1016/S0968-0004(03)00002-1
11. Hanifin JM, Rajka G. Diagnostic features of atopic dermatitis. Acta Derm Venereol (Stockh). 1980;92(suppl):44–47.
12. Comi C, Cappellano G, Chiocchetti A, et al. The impact of osteopontin gene variations on multiple sclerosis development and progression. Clin Dev Immunol. 2012;2012:212893. doi:10.1155/2012/212893
13. Glas J, Seiderer J, Bayrle C, et al. The role of osteopontin (OPN/SPP1) haplotypes in the susceptibility to Crohn’s disease. PLoS One. 2011;6(12):e29309. doi:10.1371/journal.pone.0029309
14. Giacopelli F, Marciano R, Pistorio A, et al. Polymorphisms in the osteopontin promoter affect its transcriptional activity. Physiol Genomics. 2004;20(1):87–96. doi:10.1152/physiolgenomics.00138.2004
15. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. 2006;22(15):1928–1929. doi:10.1093/bioinformatics/btl268
16. Moran MD. Arguments for rejecting the sequential Bonferroni in ecological studies. Oikos. 2003;100(2):403–405. doi:10.1034/j.1600-0706.2003.12010.x
17. Nakagawa S. A farewell to Bonferroni: the problems of low statistical power and publication bias. Behav Ecol. 2004;15(6):1044–1045. doi:10.1093/beheco/arh107
18. Tanino Y, Hizawa N, Konno S, et al. Sequence variants of the secreted phosphoprotein 1 gene are associated with total serum immunoglobulin E levels in a Japanese population. Clin Exp Allergy. 2006;36(2):219–225. doi:10.1111/j.1365-2222.2006.02414.x
19. Haagerup A, Bjerke T, Schoitz PO, Binderup HG, Dahl R, Kruse TA. Allergic rhinitis - a total genome-scan for susceptibility genes suggests a locus on chromosome 4q24-q27. Eur J Hum Genet. 2001;9(12):945–952. doi:10.1038/sj.ejhg.5200753
20. Arjomandi M, Galanter JM, Choudhry S, et al. Polymorphism in osteopontin gene (SPP1) is associated with asthma and related phenotypes in a Puerto Rican population. Pediatr Allergy Immunol Pulmonol. 2011;24(4):207–214. doi:10.1089/ped.2011.0095
21. Maeda K, Takahashi K, Takahashi F, et al. Distinct roles of osteopontin fragments in the development of the pulmonary involvement in sarcoidosis. Lung. 2001;179(5):279–291. doi:10.1007/s004080000068
22. Su R, Nguyen ML, Agarwal MR, et al. Interferon-inducible chemokines reflect severity and progression in sarcoidosis. Respir Res. 2013;14(1):121. doi:10.1186/1465-9921-14-121
23. Lavi H, Assayag M, Schwartz A, Arish N, Fridlender ZG, Berkman N. The association between osteopontin gene polymorphisms, osteopontin expression and sarcoidosis. PLoS One. 2017;12(3):e0171945. doi:10.1371/journal.pone.0171945
24. Maver A, Medica I, Salobir B, Tercelj M, Peterlin B. Genetic variation in osteopontin gene is associated with susceptibility to sarcoidosis in Slovenian population. Dis Markers. 2009;27(6):295–302. doi:10.1155/2009/864313
25. Abdou AG, El Farargy S, Seleit I, Antar AG, Elhefny M, Elnaidany NF. Osteopontin expression in chronic plaque psoriasis: an association with the severity of disease. Anal Quant Cytol Histol. 2012;34:79–85.
26. Abdel-Mawla MY, El-Kasheshy KA, Ghonemy S, Al Balat W, Elsayed AA. Role of osteopontin in psoriasis: an Immunohistochemical Study. Indian J Dermatol. 2016;61(3):301–307. doi:10.4103/0019-5154.182434
27. Abdel Hay R, Nour-Edin F, Hegazy R, Khadiga S, Rashed L. Expression of osteopontin genotypes (T-4754-C and A-9138-C) in psoriasis and their relation to metabolic syndrome. J Dermatol Sci. 2014;75(2):150–153. doi:10.1016/j.jdermsci.2014.04.013
28. Chen YJ, Shen JL, Wu CY, Chen CM, Lee FY, Lee F-Y. Elevated plasma osteopontin level is associated with occurrence of psoriasis and is an unfavorable cardiovascular risk factor in patients with psoriasis. J Am Acad Dermatol. 2009;60(2):225–230. doi:10.1016/j.jaad.2008.09.046
29. Hayashi C, Rittling S, Hayata T, et al. Serum osteopontin, an enhancer of tumor metastasis to bone, promotes B16 melanoma cell migration. J Cell Biochem. 2007;101(4):979–986. doi:10.1002/jcb.21298
30. Reduta T, Śniecińska M, Pawłoś A, Sulkiewicz A, Sokołowska M. Serum osteopontin levels in disseminated allergic contact dermatitis. Adv Med Sci. 2015;60(2):273–276. doi:10.1016/j.advms.2015.05.001
31. Ganzetti G, Simonetti O, Campanati A, et al. Osteopontin: a new facilitating factor in alopecia areata pathogenesis? Acta Dermatovenerol Croat. 2015;23:19–22.
32. Kaleta B. Role of osteopontin in systemic lupus erythematosus. Arch Immunol Ther Exp (Warsz). 2014;62(6):475–482. doi:10.1007/s00005-014-0294-x
33. Green PM, Ludbrook SB, Miller DD, Horgan CM, Barry ST. Structural elements of the osteopontin SVVYGLR motif important for the interaction with α 4 integrins. FEBS Lett. 2001;503(1):75–79. doi:10.1016/S0014-5793(01)02690-4
34. Liu W, Zeng Q, Chen Y, Luo RZ. Role of leptin/osteopontin axis in the function of eosinophils in allergic rhinitis with obesity. Mediators Inflamm. 2018;2018:9138904. doi:10.1155/2018/9138904
35. Fuchs J, Zollner TM, Kaufmann R, Podda M. Redox-modulated pathways in inflammatory skin diseases. Free Radic Biol Med. 2001;30(4):337–353. doi:10.1016/S0891-5849(00)00482-2
36. Groneberg DA, Bester C, Grützkau A, et al. Mast cells and vasculature in atopic dermatitis–potential stimulus of neoangiogenesis. Allergy. 2005;60(1):90–97. doi:10.1111/j.1398-9995.2004.00628.x
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.