Back to Journals » Journal of Inflammation Research » Volume 17
Origin and Function of Monocytes in Inflammatory Bowel Disease
Authors Liao X ![]() , Liu J, Guo X, Meng R, Zhang W, Zhou J, Xie X, Zhou H
, Liu J, Guo X, Meng R, Zhang W, Zhou J, Xie X, Zhou H ![]()
Received 20 November 2023
Accepted for publication 23 April 2024
Published 13 May 2024 Volume 2024:17 Pages 2897—2914
DOI https://doi.org/10.2147/JIR.S450801
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Xiping Liao,1,2,* Ji Liu,3,* Xiaolong Guo,1 Ruiping Meng,1 Wei Zhang,1 Jianyun Zhou,1 Xia Xie,1,2 Hongli Zhou1
1Clinical Medical Research Center, the Second Affiliated Hospital, Army Medical University, Chongqing, People’s Republic of China; 2Department of Gastroenterology, the Second Affiliated Hospital, Army Medical University, Chongqing, People’s Republic of China; 3Sun Yat-sen University Cancer Center, State Key Laboratory of Oncology in South China, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xia Xie, Department of Gastroenterology, the Second Affiliated Hospital, Army Medical University, No. 83 Xinqiao Main Street, Shapingba District, Chongqing, 400037, People’s Republic of China, Email [email protected] Hongli Zhou, Clinical Medical Research Center, the Second Affiliated Hospital, Army Medical University, No. 83 Xinqiao Main Street, Shapingba District, Chongqing, 400037, People’s Republic of China, Email [email protected]
Abstract: Inflammatory bowel disease (IBD), including Crohn’s disease (CD) and ulcerative colitis (UC), is a chronic disease resulting from the interaction of various factors such as social elements, autoimmunity, genetics, and gut microbiota. Alarmingly, recent epidemiological data points to a surging incidence of IBD, underscoring an urgent imperative: to delineate the intricate mechanisms driving its onset. Such insights are paramount, not only for enhancing our comprehension of IBD pathogenesis but also for refining diagnostic and therapeutic paradigms. Monocytes, significant immune cells derived from the bone marrow, serve as precursors to macrophages (M&phis;s) and dendritic cells (DCs) in the inflammatory response of IBD. Within the IBD milieu, their role is twofold. On the one hand, monocytes are instrumental in precipitating the disease’s progression. On the other hand, their differentiated offsprings, namely moM&phis;s and moDCs, are conspicuously mobilized at inflammatory foci, manifesting either pro-inflammatory or anti-inflammatory actions. The phenotypic spectrum of these effector cells, intriguingly, is modulated by variables such as host genetics and the subtleties of the prevailing inflammatory microenvironment. Notwithstanding their significance, a palpable dearth exists in the literature concerning the roles and mechanisms of monocytes in IBD pathogenesis. This review endeavors to bridge this knowledge gap. It offers an exhaustive exploration of monocytes’ origin, their developmental trajectory, and their differentiation dynamics during IBD. Furthermore, it delves into the functional ramifications of monocytes and their differentiated progenies throughout IBD’s course. Through this lens, we aspire to furnish novel perspectives into IBD’s etiology and potential therapeutic strategies.
Keywords: IBD, monocytes, inflammatory response
Introduction
Inflammatory Bowel Disease (IBD) is a chronic and complex intestinal inflammatory disorder. Recent clinical data have shown promising results in IBD treatment through the combination of Granulocyte-Monocytes Apheresis with certain pharmacological agents.1–4 This emerging evidence highlights the potential efficacy of monocytes-based interventions in IBD, despite the absence of well-established therapeutic guidelines. Moreover, extensive basic research has elucidated the pivotal role of monocytes in the pathogenesis and progression of IBD.
Monocytes, integral components of the immune system, are produced in the bone marrow and play a vital role in maintaining intestinal homeostasis through processes such as recruitment, activation and differentiation.5–8 In patients with IBD, there is typically an overabundance of highly active monocytes in the intestinal milieu. These monocytes contribute significantly to the inflammatory cascade by releasing pro-inflammatory cytokines and chemokines, thereby recruiting additional immune cells to the site of inflammation.9–14 For example, monocytes-derived cytokines, including IL-1β, IL-6, and IL-23, have the capacity to activate T cells within the innate layer of the intestinal mucosa. This, in turn, results in the production of additional inflammatory cytokines, thereby amplifying the inflammatory response within the intestinal tract.14–16 Furthermore, chemokines released by monocytes, such as CCL11, can attract other immune cells, including neutrophils and eosinophils, to sites of inflammation, exacerbating tissue damage within the intestine (Figure 1).17,18
 |
Figure 1 Recruitment of Immune Cells by Monocytes in IBD. Through the secretion of cytokines, including chemokines and other inflammatory mediators, monocytes facilitate the congregation of these immune cells. These cells collectively contribute to either the amplification or the regulation of the inflammatory response integral to the pathogenesis of IBD, thereby influencing the disease’s progression and outcomes. |
Throughout the course of IBD, monocytes process a plethora of signals, such as inflammatory mediators and pattern recognition receptors.1–11 These signals subsequently lead to the release of cytokines, which play a crucial role in regulating tissue damage and repair, as well as various cellular activities, including differentiation, activation, apoptosis, phagocytosis, and immune responses.12–17 Given the central role of monocytes in the pathogenesis of IBD, they have emerged as a focal point in pathological studies aimed at elucidating the onset and progression of the disease. Consequently, there is a burgeoning interest in delineating the immunobiology of monocytes in IBD, and unraveling the intricacies of their functional alterations in the context of the disease.
Originate of Monocytes
The conventional model of monocytes generation posits that hematopoietic stem cells (HSCs) in the bone marrow differentiate into multipotent progenitors (MPP), which subsequently give rise to common myeloid progenitors (CMP). These CMPs then differentiate further into granulocyte-monocytes progenitors (GMP), monocytes-dendritic cell progenitors (DMP), and common monocytes progenitors (cMoP), ultimately resulting in the development of monocytes.19,20 However, recent studies in mice have challenged this linear model, proposing two parallel pathways originating from the CMP stage: one pathway leads to the formation of neutrophil-like monocytes (NeuMo) and neutrophils via GMP and monocytes progenitors (MP); the other pathway leads to dendritic cell-like monocytes (DCMo) and dendritic cells (DC) via DMP and cMoP. Furthermore, in vitro studies have shown that human GMPs failed to produce DCs, suggesting a potential dual-pathway in human monocytes development as well.21,22 This raises the question of whether MP and cMoP, derivatives of GMP and DMP respectively, can generate additional monocytes subgroups.
While non-classical monocytes are known to evolve from classical monocytes,23–25 evidence suggests other precursor cells might be involved.26–28 Their survival in the bone marrow is dependent on the nuclear receptor subfamily 4 group A member 1 (Nr4a1), a transcription factor essential for their development in blood and tissues.29 An intermediate monocytes subset, bridging classical and non-classical monocytes, has been identified in both the bone marrow and blood.24,30,31 Upon reaching a certain developmental stage in the bone marrow, CCR2 and its ligands CCL2 and CCL7 direct these monocytes to migrate into blood and tissues, playing innate immune roles against invading antigens.32 In summary, monocytes mature in the bone marrow, enter circulation, and are recruited to various tissues based on ecological demands, thus providing essential innate immune functions. However, the developmental relationships between different monocytes subsets remain unclear and require further investigation (Figure 2).
 |
Figure 2 Development and Migration of Monocytes in IBD Pathogenesis. Monocytes originate from hematopoietic stem cells (HSC) in the bone marrow and undergo a sequential process of differentiation, culminating in the formation of monocytes. These monocytes primarily access the circulation through the CCL/CCR2 axis. In the context of IBD, a range of cytokines, chemokines, integrins, adhesion molecules, and gut microenvironmental cues mediate the migration of circulating monocytes to sites of inflammation. Once monocytes infiltrate the gut, they differentiate into macrophages and dendritic cells, ultimately leading to immune response. |
Clinical Relevance of Monocytes in IBD
Grimm et al identified an increase in CCL2 production by macrophages, smooth muscle cells, and endothelial cells in IBD patients. Elevated CCL2 levels recruited monocytes to the intestinal mucosa, where they differentiated into macrophages, releasing inflammatory mediators like MIP, RANTES, and CCL2, exacerbating mucosal inflammation. These monocytes also showed increased sensitivity to LPS, contributing to IBD’s persistent inflammation.33–35 Clinical data from CD patients show higher monocytes counts during relapses compared to remission.36 Patients with monocytes counts over 8.15% are more likely to relapse within six months, even if in deep remission.37 In UC patients with low CRP levels, higher monocytes counts correlate negatively with mucosal healing.38 Hence, monocytes counts and lymphocyte-to-monocytes ratios could predict disease activity in UC.39
Further research identified RhoB, CTSD, and ZYX as key genes in UC, associated with monocytes infiltration.40 Polymorphisms in IBD-related risk genes affect cytokine secretion and signaling in monocytes-derived macrophages, regulating inflammation in IBD.41–43 Frame-shift mutations in CSF2RB and CARD15 reduce monocytes response to GM-CSF, increasing IBD susceptibility.44,45 Autophagy gene variants ATG16L1 and NOD2 enhance monocytes phagocytic capacity in IBD patients.46 These findings highlight altered monocytes recruitment and differentiation in IBD and show how genetic markers affect monocytes functions, disrupting mucosal barriers and intestinal homeostasis.
Migration and Recruitment of Monocytes During IBD
Important Mediators of Monocytes Migration and Recruitment in the Course of IBD
The migration and recruitment of monocytes within the intestine differ markedly between homeostatic and inflammatory conditions. Under homeostatic conditions, monocytes utilize CCR2 and β7-integrin to facilitate their homing to the intestine. Conversely, during episodes of intestinal inflammation, monocytes amplify their recruitment through enhanced expression of CCR2 and no longer rely on β7-integrin for this process.6 In animal models of acute colitis, CCR2 expression has been found to be pivotal in recruiting Ly6Chi monocytes to the inflamed intestine, rendering Ly6Chi monocytes the predominant cell type within the lamina propria.47 CCL2, a well-documented ligand for CCR2, is primarily produced by monocytes and “classically activated” pro-inflammatory macrophages, although CCL7 and CCL8 are also recognized as ligands for CCR2.48–50 Researches indicate that macrophage-expressed Dectin-1 and the highly expressed Engulfment and Cell Motility protein 1 in intestinal tissues facilitate the infiltration of Ly6ChiCCR2hi monocytes subsets from the blood into inflamed colons by upregulating CCL2.51–53 Furthermore, studies have shown that pathogen infection-induced activation of NOD2 is associated with the development of CD, partially attributed to NOD2-mediated recruitment of CCL2-CCR2-dependent inflammatory monocytes.54 Additionally, CD30L has been shown to activate circulating classical monocytes and upregulate CCR2 through the NF-κB pathway. This, in turn, induces monocytes homing in a colitis mouse model via the CCL2/CCR2 axis.55,56 Importantly, researches have indicated that the chemotactic cytokine PSMP is upregulated earlier than CCL2 in experimental colitis and promotes the expression of CCL2. This suggests that PSMP may initiate the early recruitment of circulating Ly6ChiCCR2+ monocytes to the tissue, working in concert with CCL2 and CCL7 to drive colitis. Nevertheless, the chemotactic action remains dependent on CCR2.50 Therefore, CCR2 emerges as a crucial regulator in directing monocytes chemotaxis to lesion sites in IBD.
Monocytes migration in IBD is a complex process that extends beyond the CCL2/CCR2 axis. In active IBD, there is an observed upregulation of α4 integrin and CX3CR1, which facilitates the infiltration of CD14+CD16+ monocytes into the mucosa.57 Furthermore, CD14+ monocytes with high HLA-DR expression can re-localize to the intestinal mucosa via interactions with CCR9 and CCL25.58 Employing a TNBS-induced chronic colitis rat model, Ajuebor et al demonstrated that RANTES plays a role in promoting monocytes recruitment to inflammatory sites during the transition from acute to chronic colitis.59 In studies involving DSS-induced colitis and bone marrow transplantation, Fpr2/3 was shown to modulate the CCL20-CCR6 interaction, thereby driving monocytes chemotaxis to intestinal mucosa.60 Moreover, GM-CSF activation of monocytes enhances their migration, chemotaxis, adhesion, and recruitment to inflamed sites, which also correlates with the upregulation of chemokines.7 Thus, a comprehensive understanding of monocytes chemokine expression in IBD could better address their recruitment dynamics, beyond a singular pathway focus.
Beyond the influence of chemokines and their receptors on monocytes migration, cell adhesion molecules play a pivotal role in monocytes recruitment in IBD. Early research identified significant monocytes migration to the intestines in CD patients, with β2 integrins playing a crucial role.61 Recent findings indicate that α4β7 integrin regulates non-classical monocytes adhesion to MAdCAM-1 and E-cadherin, influencing intestinal homing.62 Furthermore, compounds derived from chemical and traditional herbal medicines that inhibit the expression of adhesion molecules such as ICAM-1, VCAM-1, and MAdCAM-1 have been shown to obstruct monocytes adhesion to inflamed intestines, providing relief from IBD.7,63–65 Moreover, in a DSS-induced murine colitis model, Abe et al demonstrated that TLR9-activated conventional dendritic cells regulate monocytes transport to the inflamed colon through the induction of IFN-1.66 Additionally, the glucocorticoid-induced TNF receptor family-related protein ligand, GITR-L, modulates the transport of monocytes from splenic reservoirs to inflammatory sites.24 Notably, recent research on experimental colitis revealed that the impaired migratory capability of Trim33−/− circulating monocytes was not related to defects in the CCL2/CCR2 axis, but rather to the diminished migratory ability of bone marrow-derived precursor cells with a Trim33 deficiency.60 These findings highlight the necessity for research on monocytes recruitment in IBD to extend beyond the CCL2/CCR2 axis and proteins expressing chemotactic or adhesive properties at inflammatory sites. Furthermore, there is a need to consider potential abnormalities in the differentiation of monocytes precursor cells.
Effect of Blood Compositions on Monocytes Migration and Recruitment
Blood components such as platelets and angiotensin affect monocytes migration in intestinal inflammation. PDE-3 inhibitors in murine models reduce monocytes recruitment by interrupting platelet-monocytes interactions.67 Angiotensin II limits CCL2 production, hindering Ly6C+CCR2+ monocytes recruitment.68 The peptide SRSRY reduces inflammatory monocytes recruitment to inflamed tissues.69 Cobalt protoporphyrin IX increases monocytes entry into the bloodstream but disrupts the CCL2 gradient in the colon, reducing monocytes recruitment and inflammatory cytokine expression.70 Apolipoprotein AI mimetic peptide 5A downregulates plasma CCL2 levels, reducing pro-inflammatory monocytes and suppressing monocytes adhesion and migration in human intestinal microvascular endothelial cell layers.71 These findings show that blood components impact monocytes migration, but their roles and mechanisms during IBD are not fully understood. The potential of blood analogues targeting monocytes infiltration for IBD therapy requires further research.
The Intestinal Microenvironment and the Migration and Recruitment of Monocytes During IBD
The intestinal microenvironment plays a crucial role in the migration and recruitment of monocytes in IBD. Structural disruptions to tight junction proteins such as occludin and impaired epithelial barrier function can enhance monocytes migration.8 Mucosal injuries stimulate the production of chemokines, such as RANTES, CCL20-CCR6, and CCL8, promoting monocytes recruitment to the colon.59,60,72 From a microbiota perspective, commensal Gram-positive bacteria can attract monocytes into the colon, inducing inflammation.73 Bacterial infections can stimulate NOD2-mediated CCL2-CCR2-dependent inflammatory monocytes recruitment.54
Dietary and metabolic product studies indicate that non-digestible polysaccharides stimulate macrophages to produce chemokines like CCL5 and CXCL8 in vitro, enhancing monocytes recruitment.67 Conversely, polyphenols significantly inhibit CCL2 expression and the NF-κB pathway,74,75 and butyrate suppresses TNF-α-induced VCAM-1 expression,76 reducing monocytes adhesion to intestinal epithelial cells (IECs). In the SAMP3/Yit ileitis mouse model, Ω-3 polyunsaturated fatty acids mitigate monocytes recruitment by decreasing intestinal CCL2 expression,77 highlighting the importance of dietary intake in clinical IBD management. In terms of metabolic products, spermine-treated cells exhibit reduced IFN-γ-induced CCL2 expression in THP-1 cells, subsequently decreasing monocytes recruitment.78 Furthermore, cholecystectomy-induced accumulation of secondary bile acids regulates chemokines like CCL2, CCL7, and CCL8 to suppress monocytes recruitment.79 In summary, alterations in intestinal structure, microbiota, diet, and metabolic byproducts influence monocytes recruitment to the colon. Although the intestinal microbiota appears to be a dominant factor in this process, further research is needed to validate these findings. Table 1
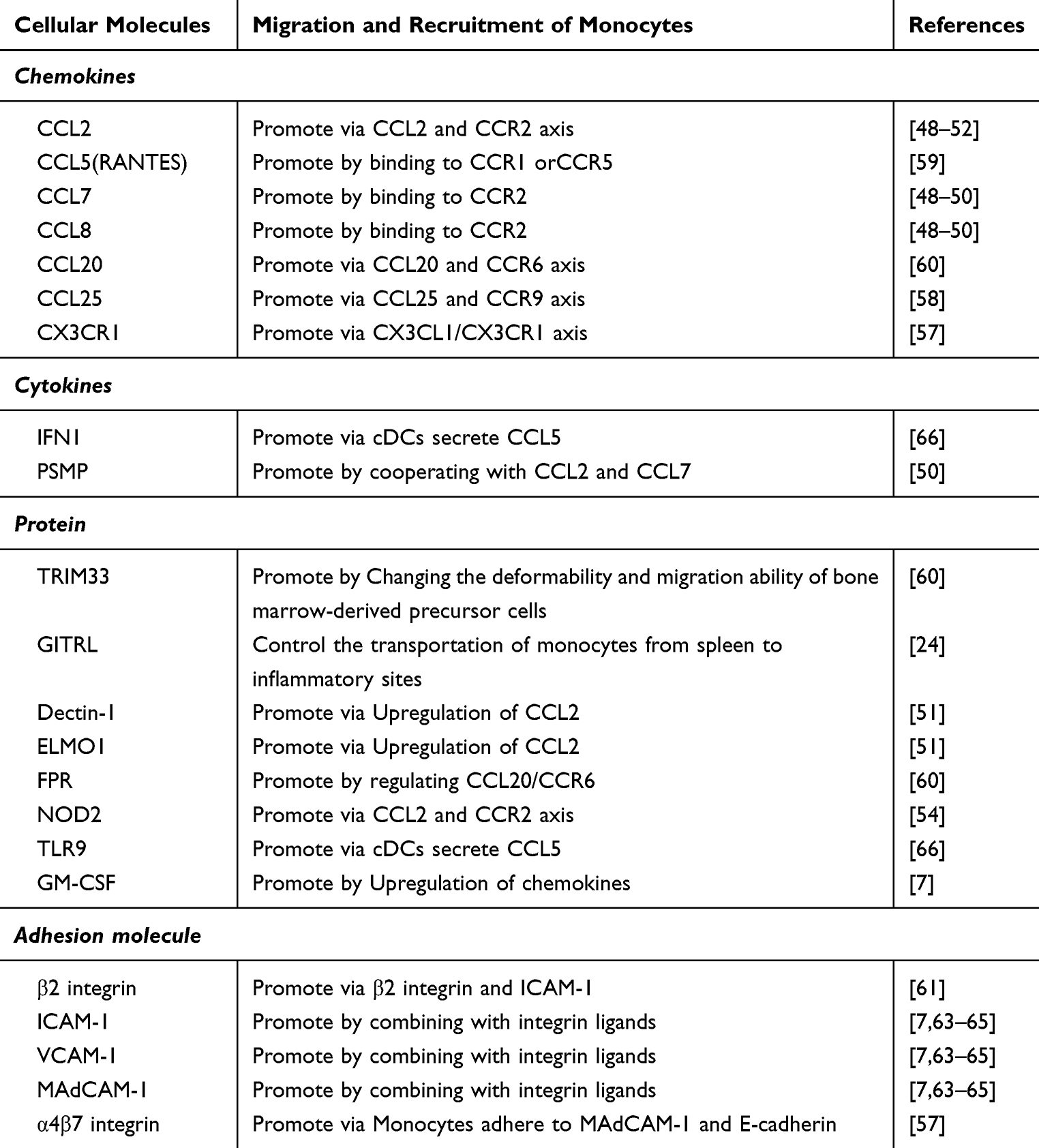 |
Table 1 Cellular Molecules Associated with the Migration and Recruitment of Monocytes in IBD |
Changes in both the blood and intestinal microenvironments, largely mediated by chemokines and adhesion molecules, are critical factors influencing monocytes migration during the progression of IBD. Banerjee et al discussed the role of biomechanical forces in regulating integrins in autoimmunity, shedding light on the importance of mechanical forces in autoimmune conditions.80 Studies have demonstrated that monocytes migration and adhesion are closely related to ICAM1/VCAM1 expression as well as the intrinsic mechanical strength of the cells themselves.81 However, this field remains relatively underexplored. The intrinsic biomechanical movement capabilities of monocytes and their impact on migration are intriguing and represent a novel avenue of investigation. Delving deeper into the field of biomechanics, including the effects on monocytes stiffness and uncovering the mechanisms of dysregulation during IBD, will provide valuable insights into monocytes migration and recruitment in IBD. Understanding these biomechanical aspects may offer new opportunities for therapeutic intervention, ultimately contributing to better management and treatment of IBD.
Differentiation of Monocytes During IBD
The intestinal inflammatory environment during IBD profoundly impacts the fate of monocytes. Once recruited from the bloodstream, these monocytes exhibit their unique subset characteristics, but they also further differentiate into intestinal macrophages and dendritic cells within specific tissue microenvironments.82 This process mirrors the adaptive response of monocytes to immunological stimuli. However, the complexity of IBD arises from the interplay of multiple factors. The heterogeneity observed in monocytes differentiation within the IBD milieu suggests that they may adopt diverse cellular fates influenced by various determinants.83 While our current understanding is limited regarding whether monocytes differentiate into cell types other than macrophages and dendritic cells, available evidence suggests that their differentiation is intricately linked with factors such as the intestinal microenvironment, inflammatory responses, bio-signaling molecules, genetic predispositions and immunoregulation. For a comprehensive understanding of the role and functionality of monocytes in IBD, in-depth studies focusing on their differentiation pathways and mechanisms in inflammatory settings are crucial. These studies will shed light on their pivotal role in the pathogenesis of IBD and could provide essential insights for developing novel therapeutic strategies.
Effect of Inflammatory Microenvironment on Monocytes Differentiation
Under physiological homeostasis, monocytes predominantly replenish the pool of intestinal macrophages, typically differentiating into tolerogenic macrophages.6,83 However, the delicate balance between monocytes and macrophages is perturbed in IBD, resulting in a substantial influx of monocytes to the intestines and mesenteric lymph nodes. Once recruited, these monocytes differentiate into inflammatory macrophages, characterized by robust phagocytic capabilities and the capacity to produce inducible nitric oxide synthase (iNOS).84,85 This shift is hypothesized to be a consequence of an interruption in the monocytes maturation process, culminating in the accumulation of intermediate form macrophages. This aberration subsequently leads to an augmented population of inflammatory macrophages following Toll-like receptor (TLR) stimulation.83 Intriguingly, this disruption in equilibrium may be intricately linked to the absence of IL-10 receptor (IL-10R) expression.86 Furthermore, it has been demonstrated that IFN-γ plays a pivotal role in maintaining the inflammatory macrophages within the inflamed intestinal milieu.87
The process of monocytes differentiation extends beyond the simple production of inflammatory macrophages, as demonstrated by experimental mouse colitis models. In these models, in the presence of CD4+ T cells, IFN-γ-stimulated monocytes-derived cells represent a distinct population of newly activated macrophages.88 Recent research has elucidated that during the resolution phase of gut muscular inflammation, activated enteric glial cells facilitate monocytes infiltration via the release of CCL2 and CSF1, ultimately leading to their differentiation into anti-inflammatory macrophages.89 Moreover, it has been observed that non-classical monocytes tend to differentiate into macrophages that play a pivotal role in promoting wound healing.90,91 Over time, monocytes also possess the capacity to differentiate into migratory antigen-presenting cells with dendritic cell characteristics, referred to as moDCs.47
Consequently, within the inflammatory milieu of IBD, monocytes differentiation emerges as a complex and multifaceted process. While contemporary researches suggest that differentiated inflammatory macrophages are predominant in such inflammatory conditions, a comprehensive examination of additional subsets is indispensable. A meticulous investigation of these various subsets could shed light on the intricacies governing the diverse differentiation pathways of monocytes, thereby elucidating the intricate mechanisms underpinning this process. Such insights could potentially pave the way for novel therapeutic interventions aimed at modulating monocytes differentiation to ameliorate IBD pathology.
Relationship Between Intestinal Microbes and Monocytes Differentiation
During acute inflammation in IBD patients, infiltrating monocytes within the lamina propria alter tight junctions, inducing epithelial cell apoptosis and disrupting mucosal integrity through inflammatory factors. This leads to the exposure of the intestinal mucosa to gut microbiota.92 In the case of animal models with acute colitis, Ly6ChiCX3CR1int monocytes accumulate and activate TLR and NOD2 recognition pathways. They respond to bacterial products and differentiate into pro-inflammatory effector cells secreting IL-6 and IL-23, as well as cells with antigen-presenting characteristics,47 potentially referred to as moMφs and moDCs respectively. Research indicates that fecal bacteria from CD patients induce monocytes differentiation into macrophages that express high levels of pro-inflammatory cytokines.93 Conversely, the probiotic strain Saccharomyces boulardii can induce the proliferation of classical monocytes, which, upon infiltrating the lamina propria, differentiate into CX3CR1+Mφ.94 Moreover, gut microbiota like M. Capsulatus Bath affect the activation and maturation of moDCs.95–97 Due to the rich diversity of gut microbiota, combined with the heterogeneity of monocytes differentiation, examining a single microbial group or species might not sufficiently elucidate their interrelationships.
The Relationship Between Cytokines and Monocytes Differentiation
Recent research highlights the role of cytokines and growth factors in directing monocytes differentiation. Classical monocytes, when activated by CD30L, tend towards inflammatory monocytes through the CCL2/CCR2 and NF-κB signaling pathways.56 Schridde et al demonstrated that TGF-β is pivotal for intestinal monocytes differentiation into macrophages.98 Furthermore, during IBD inflammation, moMφs’ TGF-β activation ability via integrin αvβ8 diminishes.10 Recent findings emphasize GM-CSF-induced STAT5 tetramerization as critical for monocytes differentiation. STAT5 tetramers are essential for monocytes to become moDCs. Aberrations in these tetramers shift differentiation towards functionally distinct moMφs, with heightened arginase I expression and reduced LPS-induced NO synthesis.99 A noteworthy observation is that moMφs, when treated with GH within a GM-CSF milieu, demonstrate a pronounced bias towards anti-inflammatory gene signatures. This shift is attributed to a decline in activin A mediated via the PI3K pathway and an upsurge in MAFB expression, collectively impeding the polarization towards pro-inflammatory macrophages.100
In-depth examinations of experimental colitis paradigms suggest that IFN-γ orchestrates the differentiation trajectory of monocytes towards macrophages. This orchestration occurs via an epigenetic mechanism, specifically through the acetylation of the promoter region of the NOS2 gene, mediated by the STAT-1 signaling pathway.87 Within the monocytes-macrophage axis, the expression of IRF5 predominantly channels the differentiation of Ly6Chi monocytes into the CD11c+moMφs phenotype.101 Interestingly, when exposed to GM-CSF, monocytes exhibiting IRF4 anomalies also favor macrophage differentiation. Contextualizing this within the broader cytokine-mediated modulation landscape, it is postulated that IRF4 may function downstream of the STAT5 tetramer assembly, though this intricate mechanism merits comprehensive exploration.99,102 Moreover, the genetic backdrop characterized by miR-223 deficiency manifests in a pronounced decrement in the CR3CX1+MΦ population, contrasted by an augmentation in moDCs differentiation trajectory.103 Recent discoveries underscore that in monocytes, NOD2 activation exerts an inhibitory effect on macrophage differentiation, operating through the mTOR signaling cascade and modulated by the TNF-α axis.104 This insight potentially elucidates the pathophysiological accumulation of deleterious moMφs observed in CD patients harboring risk-associated NOD2 alleles or specific missense mutations. Lu et al using murine models, shown that the Monocytes Chemotactic Protein-Induced Protein 1 is upregulated through the ATF3-AP1S2 pathway, promoting pro-inflammatory polarization of moMφs in the intestinal mucosa.105 Conversely, Zhou et al reported that the herbal compound, Xianglian Wan, restores colonic immune balance by disrupting the STAT1 and PPARγ interaction, reducing moMφs production, and favoring anti-inflammatory macrophage polarization, validated in IBD patients.106 Table 2
 |
Table 2 Differentiation of Mouse Monocytes During Intestinal Inflammation |
In summary, monocytes differentiation within the ambit of IBD embodies a multifaceted biological trajectory. Contemporary research avenues have yet to distinctly demarcate whether moMφs and moDCs possess overlapping developmental continuums. A long-standing focal point in the discourse remains the identification of pivotal signaling events that underpin the heterogeneity in monocytes differentiation. The pertinent query centers on whether these signals arrest moMφs and moDCs at specific developmental junctures or if they directly modulate the differentiation trajectory of monocytes.
Function of Monocytes in IBD
In 2010, the International Federation of Immunological Societies’ Nomenclature Committee formally classified human monocytes into three subgroups: classical (CD14++CD16−), intermediate (CD14++CD16+), and non-classical (CD14+CD16++).107 The primary functions of classical monocytes include potent phagocytosis, tissue repair, production of inflammatory mediators such as TNF-α, IL-1β, ROS in response to microbial pathogens, and the capability to migrate to inflamed lesions. Intermediate monocytes display production of inflammatory mediators such as TNF-α, IL-1β, IL-6, proliferation during infections, and antigen-presentation roles. Meanwhile, non-classical monocytes mainly perform “patrolling” of endothelial cells, Fcγ-mediated phagocytosis, and secretion of TNF-α, IL-1β, CCL3 under immunogenic stimuli.108
Though all three subgroups can produce pro-inflammatory cytokines, classical monocytes are the best producers, followed by intermediate monocytes, and the least effective are the non-classical monocytes.109 This further elucidates the functional heterogeneity of monocytes. For instance, the deficiency of COMMD10 enhances the inflammatory mediator activity of classical monocytes,110 the downregulation of Tim-3 leads to higher expression of pro-inflammatory cytokines in classical monocytes.111 Furthermore, in inflamed colonic venous endothelium, non-classical monocytes accelerate subsequent leukocyte activation and infiltration by locally producing inflammatory cytokines and chemokines.112 They also significantly express α4β7 integrins, promoting wound healing at the lesion sites.62,113 Notably, in UC patients with anxiety and depression symptoms, the percentage of intermediate and non-classical monocytes in peripheral blood is higher, their phagocytic function is reduced, and these cells’ functions are impaired.114 This indicates that the phenotype of monocytes subgroups can vary with or without coexisting diseases. Due to the significant phenotypic and functional differences between monocytes subgroups, the immune response mechanisms induced by monocytes remain intricate.
Monocytes Directly Regulate Intestinal Inflammation
Monocytes occupy a pivotal position in orchestrating the inflammatory response characteristic of IBD, primarily through the induction of iNOS/NO and the secretion of pro-inflammatory cytokines including IFN-γ, TNF-α, IL-1β, IL-6, IL-8, IL-12 and IL-23.9–14,115–117
On the one hand, IBD is marked by a surge of monocytes that release oxygen radicals and enzymes, culminating in tissue damage.118 The resultant injury to the mucosal tissue, when exposed to the diverse microbial milieu of the gut, leads to an augmented activation of monocytes STAT3. This, in turn, compromises the patient’s innate defenses, potentiates low-level bacterial infections, and serves as a catalyst for the disease.119 Illustratively, chronic infection with Toxoplasma gondii intensifies monocytes activation, thereby escalating the release of the inflammatory mediator NO and the concomitant secondary environmental damage.120 The activation of the iNOS/NO pathway by monocytes amplifies the inflammatory response in IBD, perpetuating tissue damage. Notably, the attenuation of iNOS/NO activity generated by monocytes has been shown to mitigate experimental colitis.121,122 Conversely, VEGFR1+ cells, derived from the monocytes lineage, foster the accumulation of Treg cells in the ulcerated regions of DSS-induced colitis via tyrosine kinase signaling.123 In the inflamed milieu, monocytes release microparticles that express pro-coagulant tissue factors, thereby facilitating the healing of the damaged mucosa.124
On the other hand, alterations in the surface expression of cytokine receptors on monocytes (eg, IL-1R, CXCR3, TNF-related receptors),9,125,126 the activation of inflammatory cytokine pathways (eg, NF-κB, MAPK, JAK/STAT signaling pathways),115,125,127–132 and the subsequent regulation of pro-inflammatory genes12,14 collectively contribute to an amplification of the inflammatory response mediated by monocytes.110,133 However, the intricacies of the molecular mechanisms governing the dysregulation of cytokine secretion in monocytes during IBD remain to be fully elucidated. While the administration of anti-inflammatory drugs and certain synthetic compounds that inhibit inflammatory mediators has demonstrated efficacy in ameliorating experimental colitis, the translatability of these findings to clinical IBD treatment necessitates further exploration.63,134–136 This exploration may potentially unravel the interplay between inflammatory cytokines and extrinsic factors that modulate the inflammatory regulatory role of monocytes. Exogenous IL-10 significantly inhibits IL-12 release from LPMC stimulated with IFN-γ-treated LPS. Yet, PGE2’s inhibition of IL-12 could only be partially reversed by an anti-IL-10 monoclonal antibody,137 highlighting the intricate interplay among inflammatory cytokines. Studies also indicated that platelet-derived hyaluronidase 2 cleaves HA into fragments that directly stimulate monocytes in the inflammatory milieu to produce pro-inflammatory cytokines.138 In murine colitis models, GM-CSF modulates monocytes to upregulate cytokines like IL-4, IL-10 and IL-13, promoting Th2 T-cell differentiation.7 Vagus nerve stimulation ameliorates colonic damage, attributed to enhanced epithelial barriers and suppressed monocytes JAK1/STAT6 signaling.139–141 In light of these complex networks and interactions that characterize the interplay between inflammatory cytokines, the ideal therapeutic strategy for IBD would encompass compounds that restore or recalibrate the delicate balance between pro-inflammatory and anti-inflammatory cytokines. This balanced modulation has the potential to substantially mitigate the inflammatory cascade characteristic of IBD, thereby ameliorating the clinical manifestations of the disease (Table 3).
 |
Table 3 The Main Inflammatory Cytokines Produced by Monocytes During IBD |
Regulation of Leukocyte Function
In the context of IBD, monocytes regulate other immune cells (eg, T lymphocytes, neutrophils, eosinophils) primarily through antigen presentation and cytokine secretion. Experiments using peripheral blood mononuclear cell isolated from UC patients have shown that CD14+ monocytes expressing CD1a stimulate the activation and differentiation of CD4+ T cells into Th cells.15 Furthermore, when treated with the Hymenolepis diminuta antigen HdAg, monocytes present antigens that enhance the production of IL-10 and IL-4 by activated T cells, alleviating DSS-induced colitis.142
In IBD, there is a significant inverse correlation between the expression of miR-374a-5p in monocytes and pro-inflammatory genes (eg, TNFα, IL1A, IL6, and OSM), as well as the ability to activate T cells.14 Early studies postulated that in the pathogenesis of CD, lamina propria monocytes produce IL-12, which, through an unknown mechanism, drives a pathogenic Th1-associated immune response.143 IL-12, IL-1β, and CD163-negative monocytes-like cells induce the expression of IL-8 in colonic CD4+ T cells.16 Similarly, monocytes-derived IL-12 and IL-23 contribute to the promotion of Th1/Th17 immune responses in the colonic mucosa.144 Moreover, IECs induce monocytes TGF-β expression via thrombospondin-1, fostering the generation of induced regulatory T cells and mitigating experimental colitis in mice.145
In addition to modulating T lymphocytes, monocytes also participated in regulating other innate immune cells. For example, low concentrations of Clostridium difficile toxin effectively stimulate monocytes to release IL-8, promoting neutrophil extravasation and tissue infiltration.17 In studies of DSS-induced colitis, inflammatory monocytes express CCL11, leading to an increase in eosinophilic granulocytes.18 These findings underscore the complexity of monocytes-mediated immune regulation in IBD, encompassing cytokines, chemokines, T cells, and a myriad of interacting mechanisms (Figure 3).
 |
Figure 3 Functional heterogeneity and duality of monocytes in IBD. (A) Monocytes exhibit functional heterogeneity, with different subtypes displaying distinct functional characteristics. (B) The stimulation of various cytokine receptors, triggered by the underlying causes of IBD, activates downstream inflammatory signaling pathways, which increasing pro-inflammatory gene expression. This results in heightened inflammation and, through interactions with other immune cells, exacerbates intestinal damage. (C) Monocytes also produce anti-inflammatory cytokines during IBD, fostering immune regulatory interactions that aid in intestinal repair. |
Conclusion
Monocytes, pivotal constituents of the immune system, play an indispensable role in the pathogenesis of IBD. Originating in the bone marrow, these cells navigate through the bloodstream, targeting inflamed sites where they orchestrate inflammatory responses. Within the IBD milieu, monocytes undergo further differentiation, engaging in inflammatory reactions, tissue repair, and immune modulation through their recruitment, tissue infiltration, and cytokine release.
A particularly noteworthy aspect of monocytes in IBD is their role in bridging intestinal inflammation with the neuroendocrine system. Recent evidence from colitis mouse models reveals that α4β7 integrin-expressing monocytes are instrumental in recruiting neutrophils to the brain vasculature. This interaction precipitates heightened cytokine concentrations, with a marked surge in IL-1β levels—a cytokine implicated in mediating anxiety-like behaviors.146 Furthermore, specific commensal bacteria, namely B. vulgatus and F. varium, incite human moDCs, culminating in augmented corticotropin-releasing factor (CRF)/Urocortin 1 (UCN1) 2 levels. As modulators of brain stress networks, CRF/UCN1 exacerbates anxiety-like behaviors.147,148 Corroborating this link, IBD patients exhibit infiltration of monocytes, along with other immune cells, in the enteric ganglia.149 External stressors, via the hypothalamic-pituitary-adrenal axis, escalate glucocorticoid concentrations. Prolonged exposure to these elevated glucocorticoid levels catalyzes the genesis of pro-inflammatory subsets of intestinal glial cells, which in turn, amplify TNF-α-mediated inflammation in monocytes via CSF1.150 This revelation poses implications for the clinical administration of corticosteroids in IBD management, suggesting that while they may transiently dampen inflammation, their chronic use could potentially exacerbate the disease.
The intricate crosstalk between the neuroendocrine system and gut immunity, orchestrated by monocytes, underscores their paramount significance in IBD. As we chart the future course of research, it becomes imperative to probe deeper into the regulatory dynamics governing monocytes differentiation and function, and their interplay in IBD. Such insights could unveil novel therapeutic strategies and pathways for the prevention and management of IBD. Understanding the nuanced roles and functionalities of monocytes and their progenies can illuminate the pathogenesis of IBD, heralding innovative therapeutic avenues. Elucidating the regulatory and operational mechanisms of monocytes can potentially unveil therapeutic targets, thereby catalyzing the design of inhibitors or activators that modulate their activity in intestinal inflammation. The ongoing clinical development of novel therapeutic agents—including selective Janus kinase inhibitors, sphingosine-1-phosphate receptor modulators, SMAD7 antisense oligonucleotides, phosphodiesterase 4 inhibitors, IL-12/IL-23 inhibitors, and integrin inhibitor biologics—coupled with advances in drug combination strategies and nanoparticle targeting technologies, promises to significantly enhance the treatment landscape for patients with IBD. Additionally, insights into the reparative role of monocytes in tissue injury could pave the way for therapeutic interventions focused on tissue repair in IBD. An expanded understanding of how monocytes modulate systemic immunity may also engender pioneering immunotherapeutic approaches, enhancing the overall treatment efficacy for IBD.
Data Sharing Statement
No new data were generated or analyzed in support of this research.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of China (grant number 82302060), Chongqing Natural Science Foundation (grant number CSTB2023NSCQ-MSX0171), the key project of QingBo plan of XinQiao Hospital(grant number 2023YQB020), Key project of Chongqing Natural Science Foundation 375(grant number CSTB2023NSCQ-ZDJ0014), Key project of Chongqing Natural Science Foundation (grant number CSTB2023NSCQ-ZDX0007) and Research Project of Chongqing Science and Technology Bureau (grant number cstc2020jscx-msxmX0129).
Disclosure
The authors declare no competing interests in this work.
References
1. Noguchi Y, Shimazu K, Totani T, Komura K, Tanaka A. Comparison of adsorption efficiency of leukocytes in single needle GMA with or without PSL treatment in patients with active ulcerative colitis. Transfus Apher Sci. 2023;62(2):103581. doi:10.1016/j.transci.2022.103581
2. Tanida S, Ozeki K, Katano T, et al. Induction Therapy With a Combination of Weekly Adalimumab Plus Intensive Granulocyte and Monocyte Adsorptive Apheresis in Patients With Ulcerative Colitis and Failure of Conventional Agents, Biologics and Janus Kinase Inhibitor. J Clin Med Res. 2023;15(3):181–186. doi:10.14740/jocmr4887
3. Tsuzuki Y, Shiomi R, Matsumoto H, et al. Combined effects of granulocyte and monocyte adsorption apheresis and corticosteroids on ulcerative colitis. J Clin Biochem Nutr. 2023;72(1):68–73. doi:10.3164/jcbn.22-52
4. Ueno N, Sugiyama Y, Kobayashi Y, et al. Concomitant pharmacologic medications influence the clinical outcomes of granulocyte and monocyte adsorptive apheresis in patients with ulcerative colitis: a multicenter retrospective cohort study. J Clin Apher. 2023. doi:10.1002/jca.22040
5. Williams H, Mack C, Baraz R, et al. Monocyte Differentiation and Heterogeneity: inter-Subset and Interindividual Differences. Int J Mol Sci. 2023;24(10):8757. doi:10.3390/ijms24108757
6. Desalegn G, Pabst O. Inflammation triggers immediate rather than progressive changes in monocyte differentiation in the small intestine. Nat Commun. 2019;10(1):3229. doi:10.1038/s41467-019-11148-2
7. Däbritz J, Weinhage T, Varga G, et al. Reprogramming of monocytes by GM-CSF contributes to regulatory immune functions during intestinal inflammation. J Immunol. 2015;194(5):2424–2438. doi:10.4049/jimmunol.1401482
8. Bao J, Yura RE, Matters GL, et al. Meprin A impairs epithelial barrier function, enhances monocyte migration, and cleaves the tight junction protein occludin. Am J Physiol Renal Physiol. 2013;305(5):F714–26. doi:10.1152/ajprenal.00179.2012
9. Kaneda K, Saitoh F, Shibusawa H, et al. Correlation of neutrophil and monocyte derived interleukin-1 receptor antagonist and interleukin-8 with colitis severity in the rabbit. Cytokine. 2011;56(2):508–514. doi:10.1016/j.cyto.2011.07.008
10. Jones G-R, Bain CC, Fenton TM, et al. Dynamics of Colon Monocyte and Macrophage Activation During Colitis. Front Immunol. 2018;9:2764. doi:10.3389/fimmu.2018.02764
11. Chen J, Lv YP, Dai QM, Hu ZH, Liu ZM, Li JH. Host defense peptide LEAP-2 contributes to monocyte/macrophage polarization in barbel steed (Hemibarbus labeo). Fish Shellfish Immunol. 2019;87:184–192. doi:10.1016/j.fsi.2019.01.015
12. Chulkina MM, Pichugin AV, Ataullakhanov RI. Pharmaceutical grade synthetic peptide Thr-Glu-Lys-Lys-Arg-Arg-Glu-Thr-Val-Glu-Arg-Glu-Lys-Glu ameliorates DSS-induced murine colitis by reducing the number and pro-inflammatory activity of colon tissue-infiltrating Ly6G(+) granulocytes and Ly6C(+) monocytes. Peptides. 2020;132:170364. doi:10.1016/j.peptides.2020.170364
13. Geyer CE, Newling M, Sritharan L, et al. C-Reactive Protein Controls IL-23 Production by Human Monocytes. Int J Mol Sci. 2021;22(21).
14. Perez-Sanchez C, Barbera Betancourt A, Lyons PA, et al. miR-374a-5p regulates inflammatory genes and monocyte function in patients with inflammatory bowel disease. J Exp Med. 2022;219(5):1136. doi:10.1084/jem.20211366
15. Al-Amodi O, Jodeleit H, Beigel F, Wolf E, Siebeck M, Gropp R. CD1a-Expressing Monocytes as Mediators of Inflammation in Ulcerative Colitis. Inflamm Bowel Dis. 2018;24(6):1225–1236. doi:10.1093/ibd/izy073
16. Chapuy L, Bsat M, Rubio M, et al. IL-12 and Mucosal CD14+ Monocyte-Like Cells Induce IL-8 in Colonic Memory CD4+ T Cells of Patients With Ulcerative Colitis but not Crohn’s Disease. J Crohns Colitis. 2020;14(1):79–95. doi:10.1093/ecco-jcc/jjz115
17. Linevsky JK, Pothoulakis C, Keates S, et al. IL-8 release and neutrophil activation by Clostridium difficile toxin-exposed human monocytes. Am J Physiol. 1997;273(6):G1333–40. doi:10.1152/ajpgi.1997.273.6.G1333
18. Waddell A, Ahrens R, Steinbrecher K, et al. Colonic eosinophilic inflammation in experimental colitis is mediated by Ly6C(high) CCR2(+) inflammatory monocyte/macrophage-derived CCL11. J Immunol. 2011;186(10):5993–6003. doi:10.4049/jimmunol.1003844
19. Ożańska A, Szymczak D, Rybka J. Pattern of human monocyte subpopulations in health and disease. Scand J Immunol. 2020;92(1):e12883. doi:10.1111/sji.12883
20. Trzebanski S, Jung S. Plasticity of monocyte development and monocyte fates. Immunol Lett. 2020;227:66–78. doi:10.1016/j.imlet.2020.07.007
21. Yáñez A, Coetzee SG, Olsson A, et al. Granulocyte-Monocyte Progenitors and Monocyte-Dendritic Cell Progenitors Independently Produce Functionally Distinct Monocytes. Immunity. 2017;47(5):890–902.e4. doi:10.1016/j.immuni.2017.10.021
22. Kawamura S, Onai N, Miya F, et al. Identification of a Human Clonogenic Progenitor with Strict Monocyte Differentiation Potential: a Counterpart of Mouse cMoPs. Immunity. 2017;46(5):835–848.e4. doi:10.1016/j.immuni.2017.04.019
23. Yona S, Kim KW, Wolf Y, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity. 2013;38(1):79–91. doi:10.1016/j.immuni.2012.12.001
24. Hilgendorf I, Gerhardt LM, Tan TC, et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ Res. 2014;114(10):1611–1622. doi:10.1161/circresaha.114.303204
25. Varol C, Landsman L, Fogg DK, et al. Monocytes give rise to mucosal, but not splenic, conventional dendritic cells. J Exp Med. 2007;204(1):171–180. doi:10.1084/jem.20061011
26. Kurotaki D, Osato N, Nishiyama A, et al. Essential role of the IRF8-KLF4 transcription factor cascade in murine monocyte differentiation. Blood. 2013;121(10):1839–1849. doi:10.1182/blood-2012-06-437863
27. Alder JK, Georgantas RW, Hildreth RL, et al. Kruppel-like factor 4 is essential for inflammatory monocyte differentiation in vivo. J Immunol. 2008;180(8):5645–5652. doi:10.4049/jimmunol.180.8.5645
28. Carlin LM, Stamatiades EG, Auffray C, et al. Nr4a1-dependent Ly6C(low) monocytes monitor endothelial cells and orchestrate their disposal. Cell. 2013;153(2):362–375. doi:10.1016/j.cell.2013.03.010
29. Ginhoux F, Greter M, Leboeuf M, et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science. 2010;330(6005):841–845. doi:10.1126/science.1194637
30. Ruiz-Rosado Jde D, Olguín JE, Juárez-Avelar I, et al. MIF Promotes Classical Activation and Conversion of Inflammatory Ly6C(high) Monocytes into TipDCs during Murine Toxoplasmosis. Mediators Inflamm. 2016;2016:9101762. doi:10.1155/2016/9101762
31. Shi C, Velázquez P, Hohl TM, Leiner I, Dustin ML, Pamer EG. Monocyte trafficking to hepatic sites of bacterial infection is chemokine independent and directed by focal intercellular adhesion molecule-1 expression. J Immunol. 2010;184(11):6266–6274. doi:10.4049/jimmunol.0904160
32. Tsou CL, Peters W, Si Y, et al. Critical roles for CCR2 and MCP-3 in monocyte mobilization from bone marrow and recruitment to inflammatory sites. J Clin Invest. 2007;117(4):902–909. doi:10.1172/jci29919
33. Grimm MC, Doe WF. Chemokines in Inflammatory Bowel Disease Mucosa: expression of RANTES, Macrophage Inflammatory Protein (MIP)-1α, MIP-1β, and γ-Interferon-Inducible Protein-10 by Macrophages, Lymphocytes, Endothelial Cells, and Granulomas. Inflamm Bowel Dis. 1996;2(2):88–96.
34. Grimm MC, Pavli P, Van de Pol E, Doe WF. Evidence for a CD14+ population of monocytes in inflammatory bowel disease mucosa--implications for pathogenesis. Clin Exp Immunol. 1995;100(2):291–297. doi:10.1111/j.1365-2249.1995.tb03667.x
35. Grimm MC, Elsbury SK, Pavli P, Doe WF. Enhanced expression and production of monocyte chemoattractant protein-1 in inflammatory bowel disease mucosa. J Leukoc Biol. 1996;59(6):804–812. doi:10.1002/jlb.59.6.804
36. Ferreiro-Iglesias R, Barreiro-de Acosta M, López-Díaz J, Bastón Rey I, Domínguez-Muñoz JE. Usefulness of peripheral blood monocyte count to predict relapse in patients with inflammatory bowel disease: a prospective longitudinal cohort study. Rev Esp Enferm Dig. 2022;114(1):10–15. doi:10.17235/reed.2021.7683/2020
37. Kwon Y, Kim YZ, Choe YH, Kim MJ. Increased monocyte abundance as a marker for relapse after discontinuation of biologics in inflammatory bowel disease with deep remission. Front Immunol. 2022;13:996875. doi:10.3389/fimmu.2022.996875
38. Furukawa S, Ikeda Y, Yagi S, et al. Association Between Peripheral Blood Monocyte Count and Mucosal Healing in Japanese Patients With Ulcerative Colitis. Clin Transl Gastroenterol. 2021;12(11):e00429. doi:10.14309/ctg.0000000000000429
39. Cherfane CE, Gessel L, Cirillo D, Zimmerman MB, Polyak S. Monocytosis and a Low Lymphocyte to Monocyte Ratio Are Effective Biomarkers of Ulcerative Colitis Disease Activity. Inflamm Bowel Dis. 2015;21(8):1769–1775. doi:10.1097/mib.0000000000000427
40. Dai YC, Qiao D, Fang CY, et al. Single-cell RNA-sequencing combined with bulk RNA-sequencing analysis of peripheral blood reveals the characteristics and key immune cell genes of ulcerative colitis. World J Clin Cases. 2022;10(33):12116–12135. doi:10.12998/wjcc.v10.i33.12116
41. Hedl M, Sun R, Abraham C. Disease Risk-Associated Genetic Variants in STAT1 and STAT4 Function in a Complementary Manner to Increase Pattern-Recognition Receptor-Induced Outcomes in Human Macrophages. J Immunol. 2020;205(5):1406–1418. doi:10.4049/jimmunol.1901112
42. Hedl M, Lahiri A, Ning K, Cho JH, Abraham C. Pattern recognition receptor signaling in human dendritic cells is enhanced by ICOS ligand and modulated by the Crohn’s disease ICOSLG risk allele. Immunity. 2014;40(5):734–746. doi:10.1016/j.immuni.2014.04.011
43. Hedl M, Abraham C. A TPL2 (MAP3K8) disease-risk polymorphism increases TPL2 expression thereby leading to increased pattern recognition receptor-initiated caspase-1 and caspase-8 activation, signalling and cytokine secretion. Gut. 2016;65(11):1799–1811. doi:10.1136/gutjnl-2014-308922
44. Halme L, Turunen U, Paavola-Sakki P, et al. CARD15 frameshift mutation in patients with CROHN disease is associated with immune dysregulation. Scand J Gastroenterol. 2004;39(12):1243–1249. doi:10.1080/00365520410003489
45. Chuang LS, Villaverde N, Hui KY, et al. A Frameshift in CSF2RB Predominant Among Ashkenazi Jews Increases Risk for Crohn’s Disease and Reduces Monocyte Signaling via GM-CSF. Gastroenterology. 2016;151(4):710–723.e2. doi:10.1053/j.gastro.2016.06.045
46. Wolfkamp SC, Verseyden C, Vogels EW, et al. ATG16L1 and NOD2 polymorphisms enhance phagocytosis in monocytes of Crohn’s disease patients. World J Gastroenterol. 2014;20(10):2664–2672. doi:10.3748/wjg.v20.i10.2664
47. Zigmond E, Varol C, Farache J, et al. Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells. Immunity. 2012;37(6):1076–1090. doi:10.1016/j.immuni.2012.08.026
48. Takada Y, Hisamatsu T, Kamada N, et al. Monocyte chemoattractant protein-1 contributes to gut homeostasis and intestinal inflammation by composition of IL-10-producing regulatory macrophage subset. J Immunol. 2010;184(5):2671–2676. doi:10.4049/jimmunol.0804012
49. Yadav A, Saini V, Arora S. MCP-1: chemoattractant with a role beyond immunity: a review. Clin Chim Acta. 2010;411(21–22):1570–1579. doi:10.1016/j.cca.2010.07.006
50. Pei X, Zheng D, She S, et al. The PSMP-CCR2 interactions trigger monocyte/macrophage-dependent colitis. Sci Rep. 2017;7(1):5107. doi:10.1038/s41598-017-05255-7
51. Rahabi M, Jacquemin G, Prat M, et al. Divergent Roles for Macrophage C-type Lectin Receptors, Dectin-1 and Mannose Receptors, in the Intestinal Inflammatory Response. Cell Rep. 2020;30(13):4386–4398.e5. doi:10.1016/j.celrep.2020.03.018
52. Kimura Y, Taniguchi M, Okuda T. Acertannin Prevented Dextran Sulfate Sodium-induced Colitis by Inhibiting the Colonic Expression of IL-23 and TNF-α in C57BL/6J Mice. Planta Med. 2023;89(7):746–753. doi:10.1055/a-2037-2995
53. Haberkamp C, Allabauer I, Blaha N, et al. TNF promotes DECTIN2 family C-type lectin receptor expression in human macrophages. J Leukoc Biol. 2023. doi:10.1093/jleuko/qiad029
54. Kim YG, Kamada N, Shaw MH, et al. The Nod2 sensor promotes intestinal pathogen eradication via the chemokine CCL2-dependent recruitment of inflammatory monocytes. Immunity. 2011;34(5):769–780. doi:10.1016/j.immuni.2011.04.013
55. Mei C, Wang X, Meng F, et al. CD30L(+) classical monocytes play a pro-inflammatory role in the development of ulcerative colitis in patients. Mol Immunol. 2021;138:10–19. doi:10.1016/j.molimm.2021.06.016
56. Mei C, Meng F, Wang X, et al. CD30L is involved in the regulation of the inflammatory response through inducing homing and differentiation of monocytes via CCL2/CCR2 axis and NF-κB pathway in mice with colitis. Int Immunopharmacol. 2022;110:108934. doi:10.1016/j.intimp.2022.108934
57. Takeda S, Sato T, Katsuno T, et al. Adsorptive depletion of alpha4 integrin(hi)- and CX3CR1hi-expressing proinflammatory monocytes in patients with ulcerative colitis. Dig Dis Sci. 2010;55(7):1886–1895. doi:10.1007/s10620-009-0974-2
58. Linton L, Karlsson M, Grundström J, et al. HLA-DR(hi) and CCR9 Define a Pro-Inflammatory Monocyte Subset in IBD. Clin Transl Gastroenterol. 2012;3(12):e29. doi:10.1038/ctg.2012.23
59. Ajuebor MN, Hogaboam CM, Kunkel SL, Proudfoot AE, Wallace JL. The chemokine RANTES is a crucial mediator of the progression from acute to chronic colitis in the rat. J Immunol. 2001;166(1):552–558. doi:10.4049/jimmunol.166.1.552
60. Birkl D, O’Leary MN, Quiros M, et al. Formyl peptide receptor 2 regulates monocyte recruitment to promote intestinal mucosal wound repair. FASEB j. 2019;33(12):13632–13643. doi:10.1096/fj.201901163R
61. Liu ZX, Noguchi M, Hiwatashi N, Toyota T. Monocyte aggregation and multinucleated giant-cell formation in vitro in Crohn’s disease. The effect of cell adhesion molecules. Scand J Gastroenterol. 1996;31(7):706–710. doi:10.3109/00365529609009154
62. Schleier L, Wiendl M, Heidbreder K, et al. Non-classical monocyte homing to the gut via α4β7 integrin mediates macrophage-dependent intestinal wound healing. Gut. 2020;69(2):252–263. doi:10.1136/gutjnl-2018-316772
63. Park SW, Banskota S, Gurung P, et al. Synthesis and evaluation of 6-heteroarylamino-2,4,5-trimethylpyridin-3-ols as inhibitors of TNF-α-induced cell adhesion and inflammatory bowel disease. Medchemcomm. 2018;9(8):1305–1310. doi:10.1039/c8md00156a
64. Kadayat TM, Banskota S, Gurung P, et al. Discovery and structure-activity relationship studies of 2-benzylidene-2,3-dihydro-1H-inden-1-one and benzofuran-3(2H)-one derivatives as a novel class of potential therapeutics for inflammatory bowel disease. Eur J Med Chem. 2017;137:575–597. doi:10.1016/j.ejmech.2017.06.018
65. Kadayat TM, Banskota S, Bist G, et al. Synthesis and biological evaluation of pyridine-linked indanone derivatives: potential agents for inflammatory bowel disease. Bioorg Med Chem Lett. 2018;28(14):2436–2441. doi:10.1016/j.bmcl.2018.06.012
66. Abe K, Nguyen KP, Fine SD, et al. Conventional dendritic cells regulate the outcome of colonic inflammation independently of T cells. Proc Natl Acad Sci U S A. 2007;104(43):17022–17027. doi:10.1073/pnas.0708469104
67. Matsunaga H, Hokari R, Higashiyama M, et al. Cilostazol, a specific PDE-3 inhibitor, ameliorates chronic ileitis via suppression of interaction of platelets with monocytes. Am J Physiol Gastrointest Liver Physiol. 2009;297(6):G1077–84. doi:10.1152/ajpgi.00240.2009
68. Hachiya K, Masuya M, Kuroda N, et al. Irbesartan, an angiotensin II type 1 receptor blocker, inhibits colitis-associated tumourigenesis by blocking the MCP-1/CCR2 pathway. Sci Rep. 2021;11(1):19943. doi:10.1038/s41598-021-99412-8
69. Genua M, Ingangi V, Fonteyne P, et al. Treatment with a Urokinase Receptor-derived Cyclized Peptide Improves Experimental Colitis by Preventing Monocyte Recruitment and Macrophage Polarization. Inflamm Bowel Dis. 2016;22(10):2390–2401. doi:10.1097/mib.0000000000000896
70. Schaefer REM, Callahan RC, Atif SM, et al. Disruption of monocyte-macrophage differentiation and trafficking by a heme analog during active inflammation. Mucosal Immunol. 2022;15(2):244–256. doi:10.1038/s41385-021-00474-8
71. Nowacki TM, Remaley AT, Bettenworth D, et al. The 5A apolipoprotein A-I (apoA-I) mimetic peptide ameliorates experimental colitis by regulating monocyte infiltration. Br J Pharmacol. 2016;173(18):2780–2792. doi:10.1111/bph.13556
72. Asano K, Takahashi N, Ushiki M, et al. Intestinal CD169(+) macrophages initiate mucosal inflammation by secreting CCL8 that recruits inflammatory monocytes. Nat Commun. 2015;6:7802. doi:10.1038/ncomms8802
73. Nakanishi Y, Sato T, Ohteki T. Commensal Gram-positive bacteria initiates colitis by inducing monocyte/macrophage mobilization. Mucosal Immunol. 2015;8(1):152–160. doi:10.1038/mi.2014.53
74. Babu D, Lee JS, Park SY, et al. Involvement of NF-kappaB in the inhibitory actions of Platycarya strobilacea on the TNF-alpha-induced monocyte adhesion to colon epithelial cells and chemokine expression. Arch Pharm Res. 2008;31(6):727–735. doi:10.1007/s12272-001-1219-0
75. Babu D, Thapa D, Lee JS, et al. Inhibitory effects of an aqueous extract of Cornus kousa Burg. leaves on TNF-alpha-induced chemokine expression and monocyte adhesion to human colonic epithelial cells. Arch Pharm Res. 2009;32(1):91–98. doi:10.1007/s12272-009-1122-4
76. Menzel T, Lührs H, Zirlik S, et al. Butyrate inhibits leukocyte adhesion to endothelial cells via modulation of VCAM-1. Inflamm Bowel Dis. 2004;10(2):122–128. doi:10.1097/00054725-200403000-00010
77. Govers C, Tang Y, Stolte EH, Wichers HJ, Mes JJ. Wheat-derived arabinoxylans reduced M2-macrophage functional activity, but enhanced monocyte-recruitment capacity. Food Funct. 2020;11(8):7073–7083. doi:10.1039/d0fo00316f
78. Morón B, Spalinger M, Kasper S, et al. Activation of protein tyrosine phosphatase non-receptor type 2 by spermidine exerts anti-inflammatory effects in human THP-1 monocytes and in a mouse model of acute colitis. PLoS One. 2013;8(9):e73703. doi:10.1371/journal.pone.0073703
79. Liu Y, Xu J, Ren X, et al. Cholecystectomy-induced secondary bile acids accumulation ameliorates colitis through inhibiting monocyte/macrophage recruitment. Gut Microbes. 2022;14(1):2107387. doi:10.1080/19490976.2022.2107387
80. Banerjee S, Nara R, Chakraborty S, Chowdhury D, Haldar S. Integrin Regulated Autoimmune Disorders: understanding the Role of Mechanical Force in Autoimmunity. Front Cell Dev Biol. 2022;10:852878. doi:10.3389/fcell.2022.852878
81. Chen C, Bai X, Ding Y, Lee IS. Electrical stimulation as a novel tool for regulating cell behavior in tissue engineering. Biomater Res. 2019;23:25. doi:10.1186/s40824-019-0176-8
82. Little MC, Hurst RJ, Else KJ. Dynamic changes in macrophage activation and proliferation during the development and resolution of intestinal inflammation. J Immunol. 2014;193(9):4684–4695. doi:10.4049/jimmunol.1400502
83. Bain CC, Scott CL, Uronen-Hansson H, et al. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 2013;6(3):498–510. doi:10.1038/mi.2012.89
84. Tamoutounour S, Henri S, Lelouard H, et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur J Immunol. 2012;42(12):3150–3166. doi:10.1002/eji.201242847
85. Pohl JM, Gutweiler S, Thiebes S, et al. Irf4-dependent CD103(+)CD11b(+) dendritic cells and the intestinal microbiome regulate monocyte and macrophage activation and intestinal peristalsis in postoperative ileus. Gut. 2017;66(12):2110–2120. doi:10.1136/gutjnl-2017-313856
86. Zigmond E, Bernshtein B, Friedlander G, et al. Macrophage-restricted interleukin-10 receptor deficiency, but not IL-10 deficiency, causes severe spontaneous colitis. Immunity. 2014;40(5):720–733. doi:10.1016/j.immuni.2014.03.012
87. Nakanishi Y, Sato T, Takahashi K, Ohteki T. IFN-γ-dependent epigenetic regulation instructs colitogenic monocyte/macrophage lineage differentiation in vivo. Mucosal Immunol. 2018;11(3):871–880. doi:10.1038/mi.2017.104
88. Brem-Exner BG, Sattler C, Hutchinson JA, et al. Macrophages driven to a novel state of activation have anti-inflammatory properties in mice. J Immunol. 2008;180(1):335–349. doi:10.4049/jimmunol.180.1.335
89. Stakenborg M, Abdurahiman S, De Simone V, et al. Enteric glial cells favor accumulation of anti-inflammatory macrophages during the resolution of muscularis inflammation. Mucosal Immunol. 2022;15(6):1296–1308. doi:10.1038/s41385-022-00563-2
90. Heidbreder K, Sommer K, Wiendl M, et al. Nr4a1-dependent non-classical monocytes are important for macrophage-mediated wound healing in the large intestine. Front Immunol. 2022;13:1040775. doi:10.3389/fimmu.2022.1040775
91. Sommer K, Heidbreder K, Kreiss L, et al. Anti-β7 integrin treatment impedes the recruitment on non-classical monocytes to the gut and delays macrophage-mediated intestinal wound healing. Clin Transl Med. 2023;13(4):e1233. doi:10.1002/ctm2.1233
92. Lissner D, Schumann M, Batra A, et al. Monocyte and M1 Macrophage-induced Barrier Defect Contributes to Chronic Intestinal Inflammation in IBD. Inflamm Bowel Dis. 2015;21(6):1297–1305. doi:10.1097/mib.0000000000000384
93. Gu J, Liu G, Xing J, Song H, Wang Z. Fecal bacteria from Crohn’s disease patients more potently activated NOD-like receptors and Toll-like receptors in macrophages, in an IL-4-repressible fashion. Microb Pathog. 2018;121:40–44. doi:10.1016/j.micpath.2018.05.009
94. Ibáñez L, Pontier-Bres R, Larbret F, et al. Saccharomyces boulardii Strain CNCM I-745 Modifies the Mononuclear Phagocytes Response in the Small Intestine of Mice Following Salmonella Typhimurium Infection. Front Immunol. 2019;10:643. doi:10.3389/fimmu.2019.00643
95. Indrelid S, Kleiveland C, Holst R, Jacobsen M, Lea T. The Soil Bacterium Methylococcus capsulatus Bath Interacts with Human Dendritic Cells to Modulate Immune Function. Front Microbiol. 2017;8:320. doi:10.3389/fmicb.2017.00320
96. Jensen SR, Mirsepasi-Lauridsen HC, Thysen AH, et al. Distinct inflammatory and cytopathic characteristics of Escherichia coli isolates from inflammatory bowel disease patients. Int J Med Microbiol. 2015;305(8):925–936. doi:10.1016/j.ijmm.2015.10.002
97. Radwan P, Radwan-Kwiatek K, Tabarkiewicz J, Radej S, Rolinski J. Enhanced phenotypic and functional maturation of monocyte-derived dendritic cells from patients with active Crohn’s disease and ulcerative colitis. J Physiol Pharmacol. 2010;61(6):695–703.
98. Schridde A, Bain CC, Mayer JU, et al. Tissue-specific differentiation of colonic macrophages requires TGFβ receptor-mediated signaling. Mucosal Immunol. 2017;10(6):1387–1399. doi:10.1038/mi.2016.142
99. Monaghan KL, Zheng W, Akhter H, et al. Tetramerization of STAT5 regulates monocyte differentiation and the dextran sulfate sodium-induced colitis in mice. Front Immunol. 2023;14:1117828. doi:10.3389/fimmu.2023.1117828
100. Soler Palacios B, Nieto C, Fajardo P, et al. Growth Hormone Reprograms Macrophages toward an Anti-Inflammatory and Reparative Profile in an MAFB-Dependent Manner. J Immunol. 2020;205(3):776–788. doi:10.4049/jimmunol.1901330
101. Corbin AL, Gomez-Vazquez M, Berthold DL, et al. IRF5 guides monocytes toward an inflammatory CD11c(+) macrophage phenotype and promotes intestinal inflammation. Sci Immunol. 2020;5(47).
102. Briseño CG, Haldar M, Kretzer NM, et al. Distinct Transcriptional Programs Control Cross-Priming in Classical and Monocyte-Derived Dendritic Cells. Cell Rep. 2016;15(11):2462–2474. doi:10.1016/j.celrep.2016.05.025
103. Zhou H, Xiao J, Wu N, et al. MicroRNA-223 Regulates the Differentiation and Function of Intestinal Dendritic Cells and Macrophages by Targeting C/EBPβ. Cell Rep. 2015;13(6):1149–1160. doi:10.1016/j.celrep.2015.09.073
104. Chauvin C, Alvarez-Simon D, Radulovic K, et al. NOD2 in monocytes negatively regulates macrophage development through TNFalpha. Front Immunol. 2023;14:1181823. doi:10.3389/fimmu.2023.1181823
105. Lu H, Zhang C, Wu W, et al. MCPIP1 restrains mucosal inflammation by orchestrating the intestinal monocyte to macrophage maturation via an ATF3-AP1S2 axis. Gut. 2023;72(5):882–895. doi:10.1136/gutjnl-2022-327183
106. Zhou HF, Yang C, Li JY, et al. Quercetin serves as the major component of Xiang-lian Pill to ameliorate ulcerative colitis via tipping the balance of STAT1/PPARγ and dictating the alternative activation of macrophage. J Ethnopharmacol. 2023:116557. doi:10.1016/j.jep.2023.116557
107. Ziegler-Heitbrock L, Ancuta P, Crowe S, et al. Nomenclature of monocytes and dendritic cells in blood. Blood. 2010;116(16):e74–80. doi:10.1182/blood-2010-02-258558
108. Gren ST, Grip O. Role of Monocytes and Intestinal Macrophages in Crohn’s Disease and Ulcerative Colitis. Inflamm Bowel Dis. 2016;22(8):1992–1998. doi:10.1097/mib.0000000000000824
109. Boyette LB, Macedo C, Hadi K, et al. Phenotype, function, and differentiation potential of human monocyte subsets. PLoS One. 2017;12(4):e0176460. doi:10.1371/journal.pone.0176460
110. Mouhadeb O, Ben Shlomo S, Cohen K, et al. Impaired COMMD10-Mediated Regulation of Ly6C(hi) Monocyte-Driven Inflammation Disrupts Gut Barrier Function. Front Immunol. 2018;9:2623. doi:10.3389/fimmu.2018.02623
111. Wang S, Liu Y, Yuan X, Yao L, Yang S, Ma R. Fecal bacteria from ulcerative colitis patients downregulate Tim-3-mediated inhibitory effects on monocytes in vitro. Microb Pathog. 2019;128:147–152. doi:10.1016/j.micpath.2018.12.043
112. Kuboi Y, Nishimura M, Ikeda W, et al. Blockade of the fractalkine-CX3CR1 axis ameliorates experimental colitis by dislodging venous crawling monocytes. Int Immunol. 2019;31(5):287–302. doi:10.1093/intimm/dxz006
113. Du K, Foote MS, Mousavi S, et al. Combination of organic acids benzoate, butyrate, caprylate, and sorbate provides a novel antibiotics-independent treatment option in the combat of acute campylobacteriosis. Front Microbiol. 2023;14:1128500. doi:10.3389/fmicb.2023.1128500
114. Gao X, Duan S, Cao Y, Zhang Y. Change of monocytes/macrophages in ulcerative colitis patients with symptoms of anxiety and depression. BMC Gastroenterol. 2023;23(1):67. doi:10.1186/s12876-023-02693-8
115. Andoh A, Ogawa A, Kitamura K, et al. Suppression of interleukin-1beta- and tumor necrosis factor-alpha-induced inflammatory responses by leukocytapheresis therapy in patients with ulcerative colitis. J Gastroenterol. 2004;39(12):1150–1157. doi:10.1007/s00535-004-1464-0
116. Aparicio-Pagés MN, Verspaget HW, Peña AS, et al. Natural, lectin- and phorbol ester-induced cellular cytotoxicity in Crohn’s disease and ulcerative colitis. J Clin Lab Immunol. 1988;27(3):109–113.
117. Warny M, Keates AC, Keates S, et al. p38 MAP kinase activation by Clostridium difficile toxin A mediates monocyte necrosis, IL-8 production, and enteritis. J Clin Invest. 2000;105(8):1147–1156. doi:10.1172/jci7545
118. Brown SJ, Mayer L. The immune response in inflammatory bowel disease. Am J Gastroenterol. 2007;102(9):2058–2069. doi:10.1111/j.1572-0241.2007.01343.x
119. Li Y, de Haar C, Peppelenbosch MP, van der Woude CJ. New insights into the role of STAT3 in IBD. Inflamm Bowel Dis. 2012;18(6):1177–1183. doi:10.1002/ibd.21884
120. Saraav I, Cervantes-Barragan L, Olias P, et al. Chronic Toxoplasma gondii infection enhances susceptibility to colitis. Proc Natl Acad Sci U S A. 2021;118(36).
121. Rachmilewitz D, Stamler JS, Karmeli F, et al. Peroxynitrite-induced rat colitis--a new model of colonic inflammation. Gastroenterology. 1993;105(6):1681–1688. doi:10.1016/0016-5085(93)91063-n
122. Giraud AS, Pereira PM, Thim L, Parker LM, Judd LM. TFF-2 inhibits iNOS/NO in monocytes, and nitrated protein in healing colon after colitis. Peptides. 2004;25(5):803–809. doi:10.1016/j.peptides.2004.01.019
123. Betto T, Amano H, Ito Y, et al. Vascular endothelial growth factor receptor 1 tyrosine kinase signaling facilitates healing of DSS-induced colitis by accumulation of Tregs in ulcer area. Biomed Pharmacother. 2019;111:131–141. doi:10.1016/j.biopha.2018.12.021
124. Williams JC, Klein TW, Goldberger BA, Sleasman JW, Mackman N, Goodenow MM. Δ(9)-Tetrahydrocannabinol (THC) enhances lipopolysaccharide-stimulated tissue factor in human monocytes and monocyte-derived microvesicles. J Inflamm. 2015;12:39. doi:10.1186/s12950-015-0084-1
125. Zhao Q, Kim T, Pang J, et al. A novel function of CXCL10 in mediating monocyte production of proinflammatory cytokines. J Leukoc Biol. 2017;102(5):1271–1280. doi:10.1189/jlb.5A0717-302
126. O’Dwyer AM, Lajczak NK, Keyes JA, Ward JB, Greene CM, Keely SJ. Ursodeoxycholic acid inhibits TNFα-induced IL-8 release from monocytes. Am J Physiol Gastrointest Liver Physiol. 2016;311(2):G334–41. doi:10.1152/ajpgi.00406.2015
127. Benard C, Cultrone A, Michel C, et al. Degraded carrageenan causing colitis in rats induces TNF secretion and ICAM-1 upregulation in monocytes through NF-kappaB activation. PLoS One. 2010;5(1):e8666. doi:10.1371/journal.pone.0008666
128. Frasch SC, McNamee EN, Kominsky D, et al. G2A Signaling Dampens Colitic Inflammation via Production of IFN-γ. J Immunol. 2016;197(4):1425–1434. doi:10.4049/jimmunol.1600264
129. Gren ST, Janciauskiene S, Sandeep S, et al. The protease inhibitor cystatin C down-regulates the release of IL-β and TNF-α in lipopolysaccharide activated monocytes. J Leukoc Biol. 2016;100(4):811–822. doi:10.1189/jlb.5A0415-174R
130. Gerner RR, Klepsch V, Macheiner S, et al. NAD metabolism fuels human and mouse intestinal inflammation. Gut. 2018;67(10):1813–1823. doi:10.1136/gutjnl-2017-314241
131. Cordes F, Lenker E, Weinhage T, et al. Impaired IFN-γ-dependent STAT3 Activation Is Associated With Dysregulation of Regulatory and Inflammatory Signaling in Monocytes of Ulcerative Colitis Patients. Inflamm Bowel Dis. 2021;27(6):887–901. doi:10.1093/ibd/izaa280
132. Seidelin JB, Jensen S, Hansen M, et al. IAPs and RIPK1 mediate LPS-induced cytokine production in healthy subjects and Crohn’s disease. Clin Exp Immunol. 1093.
133. Juchem KW, Gounder AP, Gao JP, et al. NFAM1 Promotes Pro-Inflammatory Cytokine Production in Mouse and Human Monocytes. Front Immunol. 2021;12:773445. doi:10.3389/fimmu.2021.773445
134. Bruin KF, Hommes DW, Jansen J, Tytgat GN, Wouter ten Cate J, van Deventer SJ. Modulation of cytokine release from human monocytes by drugs used in the therapy of inflammatory bowel diseases. Eur J Gastroenterol Hepatol. 1995;7(8):791–795.
135. Elfiky AMI, Ghiboub M, Li Yim AYF, et al. Carboxylesterase-1 Assisted Targeting of HDAC Inhibitors to Mononuclear Myeloid Cells in Inflammatory Bowel Disease. J Crohns Colitis. 2022;16(4):668–681. doi:10.1093/ecco-jcc/jjab176
136. Elfiky AMI, Hageman IL, Becker MAJ, et al. A BET Protein Inhibitor Targeting Mononuclear Myeloid Cells Affects Specific Inflammatory Mediators and Pathways in Crohn’s Disease. Cells. 2022;11(18).
137. Monteleone G, Parrello T, Monteleone I, Tammaro S, Luzza F, Pallone F. Interferon-gamma (IFN-gamma) and prostaglandin E2 (PGE2) regulate differently IL-12 production in human intestinal lamina propria mononuclear cells (LPMC). Clin Exp Immunol. 1999;117(3):469–475. doi:10.1046/j.1365-2249.1999.00991.x
138. de la Motte C, Nigro J, Vasanji A, et al. Platelet-derived hyaluronidase 2 cleaves hyaluronan into fragments that trigger monocyte-mediated production of proinflammatory cytokines. Am J Pathol. 2009;174(6):2254–2264. doi:10.2353/ajpath.2009.080831
139. Meregnani J, Clarençon D, Vivier M, et al. Anti-inflammatory effect of vagus nerve stimulation in a rat model of inflammatory bowel disease. Auton Neurosci. 2011;160(1–2):82–89. doi:10.1016/j.autneu.2010.10.007
140. Meroni E, Stakenborg N, Gomez-Pinilla PJ, et al. Vagus Nerve Stimulation Promotes Epithelial Proliferation and Controls Colon Monocyte Infiltration During DSS-Induced Colitis. Front Med Lausanne. 2021;8:694268. doi:10.3389/fmed.2021.694268
141. de Jonge WJ, van der Zanden EP, The FO, et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nat Immunol. 2005;6(8):844–851. doi:10.1038/ni1229
142. Reyes JL, Lopes F, Leung G, et al. Treatment with Cestode Parasite Antigens Results in Recruitment of CCR2+ Myeloid Cells, the Adoptive Transfer of Which Ameliorates Colitis. Infect Immun. 2016;84(12):3471–3483. doi:10.1128/iai.00681-16
143. Monteleone G, Biancone L, Marasco R, et al. Interleukin 12 is expressed and actively released by Crohn’s disease intestinal lamina propria mononuclear cells. Gastroenterology. 1997;112(4):1169–1178. doi:10.1016/s0016-5085(97)70128-8
144. Nakagawa T, Katsuno T, Noguchi Y, et al. Irsogladine Maleate Prevents Colitis in Interleukin-10 Gene-Deficient Mice by Reducing Interleukin-12 and −23 Production. Biol Pharm Bull. 2015;38(11):1681–1688. doi:10.1248/bpb.b15-00189
145. Fang LL, Yu HQ, Wu RJ, et al. Thrombospondin 1 Modulates Monocyte Properties to Suppress Intestinal Mucosal Inflammation. J Innate Immun. 2015;7(6):601–611. doi:10.1159/000398799
146. Koido S, Ohkusa T, Kan S, et al. Production of corticotropin-releasing factor and urocortin from human monocyte-derived dendritic cells is stimulated by commensal bacteria in intestine. World J Gastroenterol. 2014;20(39):14420–14429. doi:10.3748/wjg.v20.i39.14420
147. Mönnikes H, Schmidt BG, Taché Y. Psychological stress-induced accelerated colonic transit in rats involves hypothalamic corticotropin-releasing factor. Gastroenterology. 1993;104(3):716–723. doi:10.1016/0016-5085(93)91006-4
148. Greenwood-Van Meerveld B, Johnson AC, Cochrane S, Schulkin J, Myers DA. Corticotropin-releasing factor 1 receptor-mediated mechanisms inhibit colonic hypersensitivity in rats. Neurogastroenterol Motil. 2005;17(3):415–422. doi:10.1111/j.1365-2982.2005.00648.x
149. Wiese JJ, Manna S, Kühl AA, et al. Myenteric plexus immune cell infiltrations and neurotransmitter expression in Crohn’s disease and ulcerative colitis. J Crohns Colitis. 2023. doi:10.1093/ecco-jcc/jjad122
150. Schneider KM, Blank N, Alvarez Y, et al. The enteric nervous system relays psychological stress to intestinal inflammation. Cell. 2023;186(13):2823–2838.e20. doi:10.1016/j.cell.2023.05.001
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.