")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
Orexin/Hypocretin Type 2 Receptor (HCRTR2) Gene as a Candidate Gene in Sertraline-Associated Insomnia in Depressed Patients
Authors Firouzabadi N , Navabzadeh N, Moghimi-Sarani E, Haghnegahdar M
Received 19 February 2020
Accepted for publication 22 April 2020
Published 4 May 2020 Volume 2020:16 Pages 1121—1128
DOI https://doi.org/10.2147/NDT.S250141
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Negar Firouzabadi,1,2 Niloofar Navabzadeh,1 Ebrahim Moghimi-Sarani,3 Maral Haghnegahdar1
1Department of Pharmacology and Toxicology, School of Pharmacy, Shiraz University of Medical Sciences, Shiraz, Iran; 2Pharmaceutical Sciences Research Center, Shiraz University of Medical Sciences, Shiraz, Iran; 3Department of Psychiatry, School of Medicine, Shiraz University of Medical Sciences, School of Medicine, Shiraz, Iran
Correspondence: Negar Firouzabadi
Department of Pharmacology and Toxicology, School of Pharmacy, Shiraz University of Medical Sciences, Shiraz, Iran
Tel +98 917-314-5303
Fax +98 713-2424128
Email [email protected]
Background: Selective serotonin reuptake inhibitors (SSRIs) are considered as first-line drugs for treating depressive disorders. Among the adverse effects reported with sertraline is sleep disturbances; however, the etiology lying beneath is obscure. Orexin, the most recently discovered neurotransmitter, is involved in the sleep cycle. It exerts its physiological actions through orexin or hypocretin type 1 and 2 receptors (HCRTR1 and HCRTR2). Dysfunction of the orexin system contributes to various psychiatric, neurologic and neuropsychiatric disorders. Thus, our study aimed to assess the possible association of genetic variation of HCRTR2 G1246A with hypersomnia reported with sertraline in a group of major depressive disorder (MDD) patients.
Patients and Methods: Ninety-six newly diagnosed MDD patients were enrolled in our cohort study. MDD was assessed using DSM-V criteria. Insomnia Severity Index (ISI) was used to assess insomnia at baseline (week 0) and week 4. Blood samples were collected for further genotyping of HCRTR2 G1246A (rs2653349) using polymerase chain reaction-restriction fragment length polymorphism.
Results: A significant association between G1264A polymorphism of HCRTR2 and insomnia was observed. Insomnia with sertraline happens by 2.5-fold (P=0.022; odds ratio (OR)=2.5; 95% confidence interval (CI): 1.1– 5.7) in patients having GG genotype. Patients with G allele experience insomnia by 2.1-fold more than A allele carriers (P=0.022; OR=2.1; 95% CI= 1.1– 4.0). Subgroup analysis showed a significant association between GG genotype as well as the G allele and insomnia only in female MDD patients (P=0.011; OR=4.0; 95% CI=1.3– 12.0 and P=0.033; OR=2.4; 95% CI=1.02– 5.7, respectively).
Conclusion: In conclusion, the G1246A variant might be a predictor for insomnia in MDD patients treated with sertraline. Our findings support the idea that some variants of the HCRTR might contribute to inter-individual variability in the sleep pattern of patients receiving antidepressants.
Keywords: orexin receptor 2, sleep, insomnia, selective serotonin reuptake inhibitors, sertraline
Introduction
Selective serotonin reuptake inhibitors (SSRIs) have emerged as first-line drugs for treating depressive disorders especially due to their minimal adverse effects and good tolerability.1 However, despite their superior side effects profile over older antidepressants, they are still associated with various adverse drug reactions. Among these side effects, sleep disturbances have been frequently reported with SSRIs. Comparing two of the most widely used SSRIs, fluoxetine and sertraline, sertraline-treated patients exhibit a greater frequency of somnolence.2 Sleep disturbances among patients treated with sertraline were as follows: 35.6% insomnia, 26.5% hypersomnia, 39.3% mixed insomnia-hypersomnia and 18.8% no sleep disturbance.3
Many neurotransmitters in the brain such as ɣ-aminobutyric acid (GABA), serotonin and norepinephrine are responsible for sleep control. Orexin/hypocretin has been found in the hypothalamus areas rich in sleep-active neurons.4 This discovery adds to the list of neurotransmitters involved in the sleep cycle. It has been proposed that orexin and its receptors have a substantial role in conducting the sleep/wakefulness cycle.5–11 An important issue is the projection of orexin neurons to norepinephrine and serotonin neurons responsible for arousal.12,13 Serotonergic neurons located in the nucleus dorsalis raphe play a pivotal role in many physiological processes including sleep/arousal and mood control.14 Orexin stimulates serotonergic neurons in the dorsal raphe nucleus.13 Two types of orexin receptors, orexin/hypocretin type 1 receptor (HCRTR1) and orexin/hypocretin type 2 receptor (HCRTR2) were also found to be lavishly expressed in these monoaminergic nuclei.15,16 Dysfunction of the orexin system contributes to various psychiatric, neurologic and neuropsychiatric disorders.9 Changes in serum the levels of orexin are related to neuronal activity and sleep cycle.17,18 Orexin levels display a diurnal pattern in which, orexinergic neurons are activated during wakefulness and being practically silent during normal sleep.18–20 Although not yet fully understood, aberrant activation of the orexin neurons during the night might contribute to insomnia.21 Orexin receptor agonists would be of potential value for treating narcolepsy and conditions of excessive daytime sleepiness in humans. Likewise, dual orexin receptor antagonists, have potential as new medications for the treatment of insomnia.22
Orexin receptors (HCRTR1 and HCRTR2) are the G-protein coupled receptors with a seven-membrane domain. Generally, HCRTR1 is coupled to Gq, and HCRTR2 signals through Gq or Gi/Go. However, the coupling mechanisms appear to be different in various cell types and it is not thoroughly studied in neurons.23,24 Although the genetic deletion of HCRTR1 in mice shows no impact on sleep/wakefulness pattern, disruption of HCRTR2 causes modest sleepiness.25
Among the many factors affecting drug response and adverse drug reactions, genetics is presumed to be an accountable parameter.26 Reports have studied HCRTR2 polymorphisms.27–37 G1246A (rs2653349) on HCRTR2 causes an amino acid replacement of valine with isoleucine at position 308, which could alter receptor function. Thus we performed an association study in a cohort of MDD patients treated with sertraline to evaluate whether a certain genotype or allele of HCRTR2 (G1246A) would influence the occurrence of insomnia in this group of MDD patients.
Patients and Methods
Patients
The study was approved by the Ethics committee of Shiraz University of Medical Sciences and was following the Declaration of Helsinki. Written patient informed consent was obtained from each individual. A group of 96 unrelated, newly diagnosed MDD patients, 57 females and 39 males with a mean age of 38.3±12.6 years (mean age ± SD), recruited from Hafez Hospital, Shiraz University of Medical Sciences, were selected for the study and followed during 4 weeks by an experienced psychiatrist. The diagnosis of MDD was based on DSM-5 criteria.38 Patients had no previous history of sleep disturbances during the last 6 months. Newly diagnosed was defined as no previous history of MDD and no concomitant antidepressant use. Patients with either of the following conditions were excluded from our study. Family history and/or personal history of schizophrenia, family history and/or personal history of bipolar disorder, anxiety disorders, manic or hypomanic episode, mood incongruent psychotic symptoms, and active substance dependence, no cigarette smoking, no history of alcohol abuse or dependence and current treatment with antipsychotics or mood stabilizers. No history of conditions that may predispose to depression or sleepiness/insomnia/hypersomnia such as hypothyroidism, etc. Patients were asked not to use any alcohol and caffeine-containing beverages/food during the study. None of the patients experienced any sleep disturbances at baseline (week 0).
Insomnia Severity Index (ISI)
At the time of MDD diagnosis (week 0) and after 4 weeks of sertraline use, the ISI was calculated for each patient. ISI is a reliable and valid instrument for evaluation and quantification of insomnia severity and is a clinically suitable tool for measuring treatment-related insomnia outcomes.39
Drug Administration
Sertraline (SERTRALINE-ABIDI®, Abidi factory Tehran, Iran) was administered at a fixed-dose regimen of 50 mg for one week initially as previously described.40 The dose could be increased by 50 mg/day per week to a maximum of 200 mg/day according to clinical response over the 4 weeks of treatment duration.
Evaluation of insomnia was done at baseline, weeks 0 and 4 of treatment. Before interviewing and after obtaining the informed written consent, 5mL of venous blood samples were collected for further genotyping.
Genotyping
A blood sample was taken at baseline (week 0) for further genotyping assays. DNA extraction was carried out using a standard protocol.41 Genotyping of HCRTR2 (G1246A) was performed using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method (Table 1) and in a final volume of 25 µL and as described by Pinessi et al.28 PCR products were digested with Mbo I restriction enzyme (Thermo Fisher Scientific, Waltham, Massachusetts). Electrophoresis on 2.5% agarose (Invitrogen® Ultra Pure, Waltham, Massachusetts) gel was used to separate digested fragments. They were then stained by ethidium bromide and visualized in a UV transilluminator. All samples were genotyped at least twice and reconfirmed.
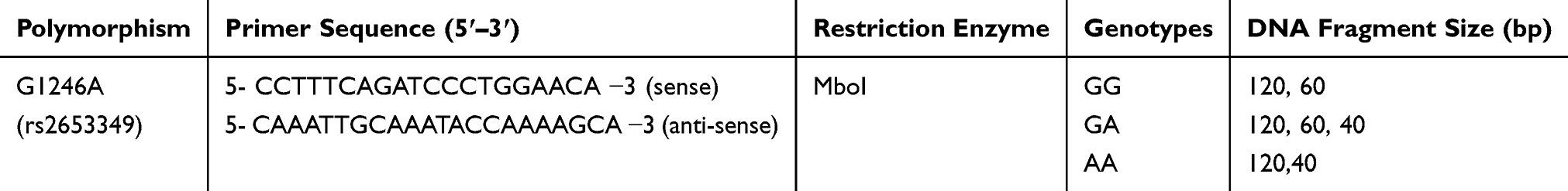 |
Table 1 Primer Sequences and G1246A Polymorphism Properties |
Statistical Analysis
Data analysis was performed using SPSS® 21.0 for windows® (SPSS Inc., Chicago, Illinois). Continuous variables are demonstrated as mean ± S.D. Genotype frequencies are presented in percentage (%). Hardy–Weinberg equilibrium (HWE) for the distribution of genotypes was calculated. Kolmogorov–Smirnov test was used to test the normal distribution of continuous variables. Associations between categorical variables were calculated by Pearson’s chi-square or Fisher’s exact test and for interval data by Student’s t-test. χ2 test was performed for univariate analysis of genotypes. Odds ratio (OR) and 95% confidence intervals (CI) were calculated. P-value <0.05 was considered as statistically significant.
Results
Demographic data are presented in Table 2. As it is demonstrated no significant difference was observed regarding sex and age between groups with insomnia and No insomnia group (P=0.658 and 0.321 respectively). As shown in Table 3, estimating ISI scores reveals a significant change between week 0 and week 4 in the insomnia group (P=0.001). Table 4 demonstrates genetic variations in two groups of patients receiving sertraline, one group experiencing insomnia after a 4-week course of sertraline treatment and the other one not experiencing insomnia. As shown in Table 4, our study indicates a significant association between G1264A polymorphism (rs2653349) of HCRTR2 and insomnia with sertraline. Insomnia happens by 2.5-fold (P=0.022; OR=2.5; 95% CI: 1.1–5.7) in patients with GG genotype. Patients with the G allele experience insomnia 2.1 fold more than carriers of the A allele (P=0.022; OR=2.1, 95% CI= 1.06–4.0). As shown in Table 5, subgroup analysis showed a significant association between GG genotype as well as the G allele and insomnia only in female MDD patients (P=0.011; OR=4.0, 95% CI=1.3–12.0 and P=0.033; OR=2.4, 95% CI=1.02–5.7 respectively).
 |
Table 2 Clinical and Demographic Characteristics of the Study Samples |
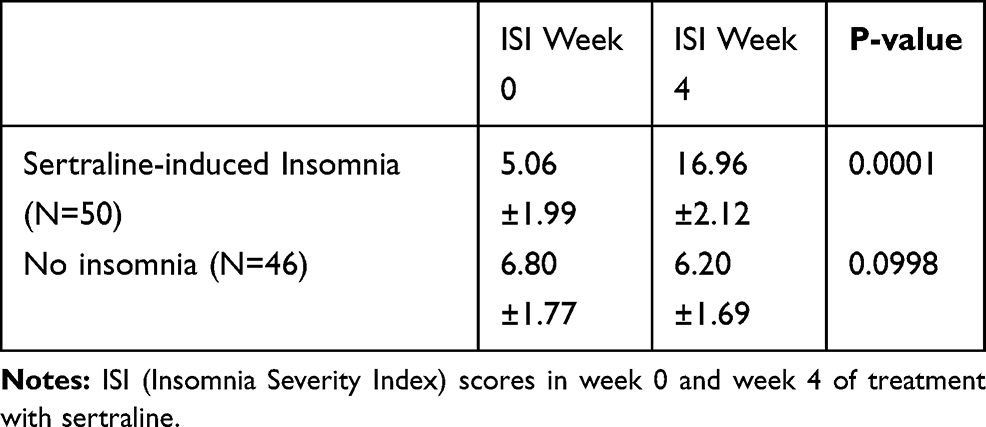 |
Table 3 ISI Scores of Enrolled Patients at Baseline (Week 0) and Week 4 of Treatment with Sertraline |
 |
Table 4 Genotype and Allele Distribution of G1246A Variant in Study Groups |
 |
Table 5 Genotype and Allele Distribution of G1246A Variant in Female and Male Patients |
Discussion
The results of our study indicate a significant association between G1264A polymorphism of HCRTR2 (rs2653349) and insomnia. Insomnia with sertraline happens by 2.5-fold in patients having the GG genotype. Patients with the G allele experience insomnia 2.1-fold more than carriers of the A allele. Subgroup analysis showed a significant association between GG genotype as well as the G allele and insomnia only in female MDD patients (P=0.011; OR=4.0, 95% CI=1.3–12.0 and P=0.033; OR=2.4, 95% CI=1.02–5.7 respectively).
Our data suggest that the orexin system may be involved in the process of sleep and arousal in MDD patients treated with sertraline. Orexins have been shown to influence a wide range of physiologic and behavioral processes. Aberrant activation of the orexin neurons at night might contribute to signs of insomnia such as increased natural metabolic rate.42 The orexin peptides apply their most prominent behavioral effects on sleep and awakening through HCRTR2 particularly.21 Suvorexant and almorexant, two orexin receptor antagonists, have been shown to stimulate sleep in both animals and men.22,43,44 In general, orexins depolarize neurons and induce neural excitability and firing. Several ionic channels are involved in their signaling mechanisms. Rise in intracellular calcium through voltage-gated calcium channels and activation of sodium/calcium exchanger contribute to the excitatory mechanism of orexins and their role in arousal.45,46 Although not much is known about the exact role of HCRTRs in sleep and arousal, several genetic studies have revealed their part in this matter. For example, functional mutations in the HCRTR2 gene were found in dogs and mice with familial narcolepsy.5,25 Orexin B failed to depolarize orexin neurons in the HCRTR2 knock-out mice. Immunoelectron microscopic observations have shown tight contacts among orexin neurons which unveiled structural resemblances to the glutamatergic synapses.47 All these reports suggest that HCRTR2 is a primary receptor in sleep and arousal. Though this causal link between narcolepsy/hypersomnia and loss of orexin signaling sometimes through alteration in HCRTR2 brought about the possibility that some mutations on HCRTR2 might contribute to this matter.
Polymorphism of G1246A (rs2653349) on HCRTR2 has been previously reported to be associated with different neurologic and psychiatric disorders.27,29,32,48 The 1246G→A polymorphism (rs2653349) of the HCRTR2 gene makes an amino acid substitution of valine to isoleucine at position 308, which could cause alteration in receptor function. Different studies reveal a decline of orexin neurons and HCRTR2 in narcolepsy and depression respectively.9 The result of our study shows that carriers of the G allele confront insomnia 2.1 times more than carriers of the A allele. Since binding of orexins to HCRTR2 stimulates calcium influx into the neurons and causes post-synaptic excitatory effects49,50 it may be postulated that mutation in the HCRTR2 gene might cause changes in HCRTR2 binding sites and interrupt calcium influx. As a consequence, the HCRTR2 will be less stimulated and arousal is perturbed. So based on our results it is presumed that patients with the wild allele and genotype do not encounter perturbation in arousal but experience insomnia. The latest studies suggest that G-protein coupled receptors such as HCRTR2 may form dimers as part of their regular cellular trafficking and function.51,52 Substitution of valine –isoleucine might be at the dimer interface and influence the dimerization process of the receptor,30 hence affecting the signaling process of HCRTR2 and as a consequence influence the sleep-wake process.
Dividing our study population to men and women revealed a significant association between GG genotype as well as the G allele with insomnia in women treated with sertraline and not men. Since G1246A polymorphism (rs2653349) is located on HCRTR2 within the chromosome 6p12 and not the sex chromosomes, it could be postulated that this finding might be due to higher frequency of enrolled female patients (57female/39male). It is worth mentioning that women are approximately twice as likely as are men to develop depression during their lifetime.53 So it was logical that enrollment of MDD patients leads to a higher frequency in women rather than men. Additionally, it is worth mentioning that the regulation of the orexin/hypocretin system is sexually dimorphic and that sexual hormones modulate this system and affect HCRTRs expression. On the other hand, orexins participate in the regulation of the hypothalamic-pituitary-gonadal axis.54,55 A previous study indicates that the administration of an HCRTR2 antagonist, JNJ-10397049, resulted in decreased serum levels of gonadotropins and ova numbers in female rats.56 Another study in postmortem human brain reported that hypothalamic orexin/hypocretin is increased in depressed women and not depressed men and HCRTR257 mRNA expression was elevated in depressed men who committed suicide and not in depressed women.58 However, how the variants of rs2653349 affect the levels of gonadal hormones as well as how sex hormones affect expression of orexin receptors in carriers of different genotypes of G1246A is still unclear.
As a limitation of our study, it must be addressed that measuring serum levels of sertraline at the baseline and after the course of treatment would have provided many vivid results. Moreover, sertraline is mainly metabolized by CYP2C19. Individuals carrying one or two alleles encoding defective CYP2C19 metabolism show significantly higher serum levels of sertraline.59 This parameter, as well as other genetic parameters, may influence serum levels of sertraline and thus affect both clinical response and extent of side effects such as insomnia. It can be speculated that patients with the GG genotype of G1246A might also have the allele of CYP2C19 which causes higher serum levels of sertraline and as a result, higher chance of insomnia. However, this hypothesis should be further studied. Therefore, it is suggested to measure serum levels of sertraline in patients with different genotypes of G1246A on HCRTR2 to assess the relationship between these variants, sertraline serum level and prevalence of insomnia.
In summary, we provide preliminary data that genetic variation in the HCRTR2 gene is associated with insomnia in MDD patients taking sertraline. Analysis of this gene in larger sample sets will provide a much subtle picture of the contribution of this variant in insomnia associated with sertraline.
Acknowledgments
This work was financially supported by a grant from the Pharmaceutical Sciences Research Center, Shiraz University of Medical Sciences, Shiraz, Iran (Grant No. 1396-01-05-14116). The project was a part of a Pharm.D, thesis by Niloofar Navabzadeh, Department of Pharmacology & Toxicology, School of Pharmacy, Shiraz University of Medical Sciences, Shiraz, Iran.
Author Contributions
Negar Firouzabadi was responsible for study design, providing the funding for this study, statistical analysis, and manuscript preparation. Niloofar Navabzadeh was responsible for recruiting the subjects, methodology, performing the clinical rating, collecting the samples and editing the manuscript. Ebrahim Moghimi Sarani was responsible for collecting the samples, drafting or revising the article and gave the final approval of the version to be published. Maral Haghnegahdar was responsible for methodology, drafting or revising the article and gave the final approval of the version to be published. Negar Firouzabadi wrote the paper. All authors have contributed to and have approved the final manuscript. All authors declare that they have no conflicts of interest. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known competing interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Rush AJ, Trivedi MH, Wisniewski SR, et al. Bupropion-SR, sertraline, or venlafaxine-XR after failure of SSRIs for depression. N Eng J Med. 2006;354(12):1231–1242. doi:10.1056/NEJMoa052963
2. Aguglia E, Casacchia M, Cassano GB, et al. Double-blind study of the efficacy and safety of sertraline versus fluoxetine in major depression. Int Clin Psychopharmacol. 1993;8(3):197–202. doi:10.1097/00004850-199300830-00010
3. Liu X, Buysse DJ, Gentzler AL, et al. Insomnia and hypersomnia associated with depressive phenomenology and comorbidity in childhood depression. Sleep. 2007;30(1):83–90. doi:10.1093/sleep/30.1.83
4. Siegel JM. The neurotransmitters of sleep. J Clin Psychiatry. 2004;65(Suppl 16):4–7.
5. Lin L, Faraco J, Li R, et al. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell. 1999;98(3):365–376. doi:10.1016/S0092-8674(00)81965-0
6. Chemelli RM, Willie JT, Sinton CM, et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell. 1999;98(4):437–451. doi:10.1016/S0092-8674(00)81973-X
7. de Lecea L, Sutcliffe JG. The hypocretins and sleep. FEBS J. 2005;272(22):5675–5688. doi:10.1111/j.1742-4658.2005.04981.x
8. De Lecea L, Huerta R. Hypocretin (orexin) regulation of sleep-to-wake transitions. Front Pharmacol. 2014;12(5):16.
9. Chen Q, de Lecea L, Hu Z, Gao D. The hypocretin/orexin system: an increasingly important role in neuropsychiatry. Med Res Rev. 2015;35(1):152–197.
10. de Lecea L. Optogenetic control of hypocretin (orexin) neurons and arousal circuits. InSleep Neuronal Plas Brain Funct. 2014;25:367–378.
11. Jennings KJ, de Lecea L. Hypocretins (Orexins): Twenty Years of Dissecting Arousal Circuits. InThe Orexin/Hypocretin System. Academic Press; 2019 Jan 1:1–29.
12. Bayer L, Eggermann E, Saint-Mleux B, et al. Selective action of orexin (hypocretin) on nonspecific thalamocortical projection neurons. J Neurosci. 2002;22(18):7835–7839. doi:10.1523/JNEUROSCI.22-18-07835.2002
13. Brown RE, Sergeeva O, Eriksson KS, Haas HL. Orexin A excites serotonergic neurons in the dorsal raphe nucleus of the rat. Neuropharmacology. 2001;40(3):457–459. doi:10.1016/S0028-3908(00)00178-7
14. Jacobs BL, Fornal CA. Activity of brain serotonergic neurons in the behaving animal. Pharmacol Rev. 1991;43(4):563–578.
15. Trivedi P, Yu H, MacNeil DJ, Van der Ploeg LH, Guan XM. Distribution of orexin receptor mRNA in the rat brain. FEBS Lett. 1998;438(1–2):71–75. doi:10.1016/S0014-5793(98)01266-6
16. Marcus JN, Aschkenasi CJ, Lee CE, et al. Differential expression of orexin receptors 1 and 2 in the rat brain. J Comp Neurol. 2001;435(1):6–25. doi:10.1002/cne.1190
17. Yoshida Y, Fujiki N, Nakajima T, et al. Fluctuation of extracellular hypocretin‐1 (orexin A) levels in the rat in relation to the light–dark cycle and sleep–wake activities. Eur J Neurosci. 2001;14(7):1075–1081. doi:10.1046/j.0953-816x.2001.01725.x
18. Lee MG, Hassani OK, Jones BE. Discharge of identified orexin/hypocretin neurons across the sleep-waking cycle. J Neurosci. 2005;25(28):6716–6720. doi:10.1523/JNEUROSCI.1887-05.2005
19. Fujiki N, Yoshida Y, Ripley B, Honda K, Mignot E, Nishino S. Changes in CSF hypocretin-1 (orexin A) levels in rats across 24 hours and in response to food deprivation. Neuroreport. 2001;12(5):993–997. doi:10.1097/00001756-200104170-00026
20. Grady SP, Nishino S, Czeisler CA, Hepner D, Scammell TE. Diurnal variation in CSF orexin-A in healthy male subjects. Sleep. 2006;29(3):295–297. doi:10.1093/sleep/29.3.295
21. Scammell TE, Winrow CJ. Orexin receptors: pharmacology and therapeutic opportunities. Annu Rev Pharmacol Toxicol. 2011;10(51):243–266. doi:10.1146/annurev-pharmtox-010510-100528
22. Brisbare-Roch C, Dingemanse J, Koberstein R, et al. Promotion of sleep by targeting the orexin system in rats, dogs and humans. Nat Med. 2007;13(2):150–155. doi:10.1038/nm1544
23. Randeva HS, Karteris E, Grammatopoulos D, Hillhouse EW. Expression of orexin-A and functional orexin type 2 receptors in the human adult adrenals: implications for adrenal function and energy homeostasis. J Clin Endocrinol Metab. 2001;86(10):4808–4813. doi:10.1210/jcem.86.10.7921
24. Karteris E, Randeva HS, Grammatopoulos DK, Jaffe RB, Hillhouse EW. Expression and coupling characteristics of the CRH and orexin type 2 receptors in human fetal adrenals. J Clin Endocrinol Metab. 2001;86(9):4512–4519. doi:10.1210/jcem.86.9.7849
25. Willie JT, Chemelli RM, Sinton CM, et al. Distinct narcolepsy syndromes in Orexin receptor-2 and Orexin null mice: molecular genetic dissection of Non-REM and REM sleep regulatory processes. Neuron. 2003;38(5):715–730. doi:10.1016/S0896-6273(03)00330-1
26. Hawcutt DB, Thompson B, Smyth RL, Pirmohamed M. Paediatric pharmacogenomics: an overview. Arch Dis Child. 2013;98(3):232–237. doi:10.1136/archdischild-2012-302852
27. Rainero I, Gallone S, Valfre W, et al. A polymorphism of the hypocretin receptor 2 gene is associated with cluster headache. Neurology. 2004;63(7):1286–1288. doi:10.1212/01.WNL.0000142424.65251.DB
28. Pinessi L, Binello E, Martino PD, et al. The 1246 G/A polymorphism of the HCRTR2 gene is not associated with migraine. Cephalalgia. 2007;27(8):945–949. doi:10.1111/j.1468-2982.2007.01347.x
29. Schürks M, Kurth T, Geissler I, Tessmann G, Diener HC, Rosskopf D. Cluster headache is associated with the G1246A polymorphism in the hypocretin receptor 2 gene. Neurology. 2006;66(12):1917–1919. doi:10.1212/01.wnl.0000215852.35329.34
30. Rainero I, Gallone S, Rubino E, et al. Haplotype analysis confirms the association between the HCRTR2 gene and cluster headache. Headache. 2008;48(7):1108–1114. doi:10.1111/j.1526-4610.2008.01080.x
31. Rainero I, Rubino E, Valfrè W, et al. Association between the G1246A polymorphism of the hypocretin receptor 2 gene and cluster headache: a meta-analysis. J Headache Pain. 2007;8(3):152. doi:10.1007/s10194-007-0383-x
32. Annerbrink K, Westberg L, Olsson M, et al. Panic disorder is associated with the Val308Iso polymorphism in the hypocretin receptor gene. Psychiatr Genet. 2011;21(2):85–89. doi:10.1097/YPG.0b013e328341a3db
33. Baumber L, Sjöstrand C, Leone M, et al. A genome-wide scan and HCRTR2 candidate gene analysis in a European cluster headache cohort. Neurology. 2006;66(12):1888–1893. doi:10.1212/01.wnl.0000219765.95038.d7
34. Schürks M, Limmroth V, Geissler I, et al. Association between migraine and the G1246A polymorphism in the hypocretin receptor 2 gene. Headache. 2007;47(8):1195–1199. doi:10.1111/j.1526-4610.2007.00863.x
35. Schürks M, Kurth T, Geissler I, Tessmann G, Diener HC, Rosskopf D. The G1246A polymorphism in the hypocretin receptor 2 gene is not associated with treatment response in cluster headache. Cephalalgia. 2007;27(4):363–367. doi:10.1111/j.1468-2982.2007.01287.x
36. Fourier C, Ran C, Steinberg A, Sjöstrand C, Waldenlind E, Belin AC. Analysis of HCRTR2 gene variants and cluster headache in Sweden. Headache. 2019;59(3):410–417. doi:10.1111/head.13462
37. Cengiz M, Karaj V, Kocabasoğlu N, Gozubatik-Celik G, Dirican A, Bayoglu B. Orexin/hypocretin receptor, Orx1, gene variants are associated with major depressive disorder. Int J Psychiatry Clin Pract. 2019;23(2):114–121. doi:10.1080/13651501.2018.1551549
38. DSM-5 American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Arlington: American Psychiatric Publishing; 2013.
39. Bastien CH, Vallières A, Morin CM. Validation of the Insomnia Severity Index as an outcome measure for insomnia research. Sleep Med. 2001;2(4):297–307. doi:10.1016/S1389-9457(00)00065-4
40. Firouzabadi N, Raeesi R, Zomorrodian K, Bahramali E, Yavarian I. Beta adrenoceptor polymorphism and clinical response to sertraline in major depressive patients. J Pharm Pharm Sci. 2017;16(20):1–7. doi:10.18433/J3W31F
41. MWer S, Dykes D, Polesky H. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988;16(3):1215. doi:10.1093/nar/16.3.1215
42. Bonnet MH, Arand DL. Insomnia, metabolic rate and sleep restoration. J Intern Med. 2003;254(1):23–31. doi:10.1046/j.1365-2796.2003.01176.x
43. Herring W, Budd K, Hutzelmann J, et al. Efficacy and tolerability of the dual orexin receptor antagonist MK-4305 in patients with primary insomnia: a randomized, controlled, adaptive crossover polysomnography study: O182. J Sleep Res. 2010;19:44–45.
44. Winrow CJ, Gotter AL, Cox CD, et al. Promotion of sleep by suvorexant—a novel dual orexin receptor antagonist. J Neurogenet. 2011;25(1–2):52–61. doi:10.3109/01677063.2011.566953
45. Van Den Pol AN, Gao XB, Obrietan K, Kilduff TS, Belousov AB. Presynaptic and postsynaptic actions and modulation of neuroendocrine neurons by a new hypothalamic peptide, hypocretin/orexin. J Neurosci. 1998;18(19):7962–7971. doi:10.1523/JNEUROSCI.18-19-07962.1998
46. Kohlmeier KA, Inoue T, Leonard CS. Hypocretin/orexin peptide signaling in the ascending arousal system: elevation of intracellular calcium in the mouse dorsal raphe and laterodorsal tegmentum. J Neurophysiol. 2004;92(1):221–235. doi:10.1152/jn.00076.2004
47. Yamanaka A, Tabuchi S, Tsunematsu T, Fukazawa Y, Tominaga M. Orexin directly excites orexin neurons through orexin 2 receptor. J Neurosci. 2010;30(38):12642–12652. doi:10.1523/JNEUROSCI.2120-10.2010
48. Nishizawa D, Kasai S, Hasegawa J, et al. Associations between the orexin (hypocretin) receptor 2 gene polymorphism Val308Ile and nicotine dependence in genome-wide and subsequent association studies. Mol Brain. 2015;8(1):50. doi:10.1186/s13041-015-0142-x
49. De Lecea L, Kilduff TS, Peyron C, et al. The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc National Acad Sci. 1998;95(1):322–327. doi:10.1073/pnas.95.1.322
50. Näsman J, Bart G, Larsson K, Louhivuori L, Peltonen H, Åkerman KE. The orexin OX1 receptor regulates Ca2+ entry via diacylglycerol-activated channels in differentiated neuroblastoma cells. J Neurosci. 2006;26(42):10658–10666. doi:10.1523/JNEUROSCI.2609-06.2006
51. Filizola M, Weinstein H. The study of G‐protein coupled receptor oligomerization with computational modeling and bioinformatics. FEBS J. 2005;272(12):2926–2938. doi:10.1111/j.1742-4658.2005.04730.x
52. Guo W, Shi L, Filizola M, Weinstein H, Javitch JA. Crosstalk in G protein-coupled receptors: changes at the transmembrane homodimer interface determine activation. Proc National Acad Sci. 2005;102(48):17495–17500. doi:10.1073/pnas.0508950102
53. Kuehner C. Why is depression more common among women than among men? Lancet Psychiatry. 2017;4(2):146–158. doi:10.1016/S2215-0366(16)30263-2
54. Pu S, Jain MR, Kalra PS, Kalra SP. Orexins, a novel family of hypothalamic neuropeptides, modulate pituitary luteinizing hormone secretion in an ovarian steroid-dependent manner. Regul Pept. 1998;78(1–3):133–136. doi:10.1016/s0167-0115(98)00128-1
55. Silveyra P, Lux-Lantos V, Libertun C. Both orexin receptors are expressed in rat ovaries and fluctuate with the estrous cycle: effects of orexin receptor antagonists on gonadotropins and ovulation. Am J Physiol Endocrinol Metab. 2007;293(4):E977–E985. doi:10.1152/ajpendo.00179.2007
56. Kok SW, Roelfsema F, Overeem S, et al. Pulsatile LH release is diminished, whereas FSH secretion is normal, in hypocretin-deficient narcoleptic men. Am J Physiol Endocrinol Metab. 2004;287(4):E630–E636. doi:10.1152/ajpendo.00060.2004
57. Weller CM, Wilbrink LA, Houwing-Duistermaat JJ, et al. Cluster headache and the hypocretin receptor 2 reconsidered: a genetic association study and meta-analysis. Cephalalgia. 2015;35(9):741–747. doi:10.1177/0333102414557839
58. Lu J, Zhao J, Balesar R, et al. Sexually dimorphic changes of hypocretin (orexin) in depression. EBioMedicine. 2017;1(18):311–319. doi:10.1016/j.ebiom.2017.03.043
59. Rudberg I, Hermann M, Refsum H, Molden E. Serum concentrations of sertraline and N-desmethyl sertraline in relation to CYP2C19 genotype in psychiatric patients. Eur J Clin Pharmacol. 2008;64(12):1181. doi:10.1007/s00228-008-0533-3
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.