")
Back to Journals » Cancer Management and Research » Volume 12
NRP1 is a Prognostic Factor and Promotes the Growth and Migration of Cells in Intrahepatic Cholangiocarcinoma
Authors Wu YN, He LH, Bai ZT, Li X
Received 26 April 2020
Accepted for publication 8 July 2020
Published 7 August 2020 Volume 2020:12 Pages 7021—7032
DOI https://doi.org/10.2147/CMAR.S260091
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Bilikere Dwarakanath
Yong-Na Wu,1– 4,* Li-Hong He,1,2,5,* Zhong-Tian Bai,1,2,5,* Xun Li1,2,5
1Department of General Surgery, The First Hospital of Lanzhou University, Lanzhou 730000, Gansu Province, People’s Republic of China; 2Key Laboratory of Biological Therapy and Regenerative Medicine Transformation Gansu Province, Lanzhou 730000, Gansu Province, People’s Republic of China; 3Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, Gansu Province, People’s Republic of China; 4University of Chinese Academy of Sciences, Beijing 100049, People’s Republic of China; 5The First Clinical Medical College, Lanzhou University, Lanzhou 730000, Gansu Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xun Li
Department of General Surgery, The First Hospital of Lanzhou University, Lanzhou University, No. 1 Donggang West Road, Lanzhou 730000, Gansu Province, People’s Republic of China
Email [email protected]
Background: Neuropilin-1 (NRP-1) participates in cancer cell proliferation and metastasis as a multifunctional co-receptor by interacting with multiple signaling pathways. However, few studies have addressed the precise function and prognosis analysis of NRP1 in intrahepatic cholangiocarcinoma (ICC). We aimed to study the correlations between NRP1 and clinicopathological characteristics and NRP1 effect on ICC cell line functions.
Methods: NRP1 mRNA and its protein levels in human ICC tissues and cell lines were detected by IHC, qRT-PCR, and WB method. Transwell, wound healing, and CCK-8 assays were performed to verify the effects of NRP1 knockdown and overexpression on cell migration and proliferation capability.
Results: NRP1 proteins and mRNA levels increased in ICC tissues compared to those in paired adjacent non-tumor tissues. High NRP1 expression of ICC tissues was related to poor prognosis. NRP1 expression level was expected to be an independent prognosticator for overall survival and cumulative tumor recurrence, and was closely related to tumor number (P=0.047). Knockdown of NRP1 inhibited cell proliferation and migration capability of RBE cells in vitro, and NRP1 overexpression in 9810 cells accelerated proliferation and migration. Additionally, NRP1 may promote cell proliferation and migration in ICC via the FAK/PI3-K/AKT pathway.
Conclusion: As an oncogene, NRP1 may function as a candidate target and prognostic biomarker of value for ICC therapy.
Keywords: neuropilin-1, prognostic factor, cell growth and migration, intrahepatic cholangiocarcinoma
Introduction
Intrahepatic cholangiocarcinoma (ICC), known as the second most common liver cancer, accounts for approximately 10% of primary liver cancer. Most ICCs originate from intrahepatic bile duct epithelial cells, with strong metastatic and invasion ability and poor prognosis.1,3 Currently, the best way to treat ICC is through surgical resection; however, only 15% of ICC patients present with resectable ICC.4,5 In addition, radiotherapy and chemotherapy offer limited benefits for advanced ICC. The mortality and incidence of ICC is on the rise worldwide, and the median survival is within 3 years, seriously endangering human health.6 Therefore, identification of novel therapeutic molecular targets is essential for improving the prognosis of ICC.7,8
NRP1 was first discovered as an adhesion molecule in the frog nervous system in 1987.9 It is a non-tyrosine kinase transmembrane glycoprotein located on the cell surface and plays a role of a co-receptor for secreted Semaphorin-3A (Sema3A). Previous studies have shown that NRP1 gene expression is widespread over a variety of cells and tissues such as endothelial cells, and the heart, liver, lung, kidney, pancreas, and skeletal muscle.10 NRP1 plays a vital role in the nervous, digestive, and immune systems, etc. through interaction with various ligands, promoting angiogenesis, neural development, cytoskeleton remodeling, inflammation, initial immune response, as well as occurrence and development of tumors.11,14 Noteworthily, the role of NRP1 in the occurrence and development of human malignant tumors has arisen as an interesting research topic recently.15,16 Increasing evidence suggests that higher expression or mutations of NRP1 are closely related to initiation, progression, and prognosis of varieties of types of malignant tumors in humans, including hepatocellular carcinoma,17 gastric cancer,18 breast cancer,19,20 prostatic cancer, and pancreatic cancer.21,22
However, so far the expression of NRP1 and its underlying role in the occurrence, development, and prognosis of ICC have been rarely discussed to date. The important role of NRP1 in many malignant tumors has prompted us to investigate the biological effects and clinical relevance of NRP1 in human ICC. In comparison with paired adjacent non-tumor tissues, we found significantly increased expression of NRP1 protein in the tumor tissues of ICC patients. We also found that the upregulation of NRP1 was an independent prognosticator for overall survival and cumulative recurrence. In addition, in vitro experiment indicated that NRP1 depletion significantly inhibited the proliferation and migration capability of human ICC cell lines; however, overexpression of NRP1 resulted in opposite results. Based on these results, we conducted further experiments and found that NRP1 possibly enrolled in the net of the FAK/PI3-K/AKT pathway. To sum up, NRP1 is a valuable promoting factor in ICC and a potential target for further drug development.
Materials and Methods
Patients and Clinical Tissue Samples
Tumor tissues and paired adjacent non-tumor tissues were collected from 291 ICC patients treated at Zhongshan Hospital of Fudan University from 2010 to 2014. All patients’ tissue specimens were classified as ICC through pathological examination. Fresh tissue specimens were obtained, formalin-fixed, and paraffin embedded according to a previously reported method.23 The study was approved by the Ethics Committee, and all the participants provided informed consent under Institutional Review Board protocols before tissue specimen collection. Patients’ clinicopathological data including gender, age, tumor number, tumor size, embolus, tumor encapsulation, TNM, liver cirrhosis, NRP1, lymphonodus, capsule, serum alpha fetoprotein (AFP), CA199, and HBsAg concentration were collected before surgery for analysis.
Immunohistochemistry (IHC)
IHC was conducted on the basis of a formerly reported method with slight modifications.24,25 In brief, tissue microarrays were constructed using BioChip (Shanghai, China). The rabbit monoclonal antibody against human NRP1 (1:200 dilution) was purchased from Abcam (Cambridge, UK). The treated sections were placed under an optical microscope (Olympus Corp., Tokyo, Japan; magnification, ×200) to observe the experimental results. For comprehensive analysis of the results, in terms of staining intensity and stained cell quantity, two specialist pathologists who lacked information about the patients separately evaluated the tissue sections. The intensity was scored as follows: 0, negative; 1, weak; 2, moderate; and 3, strong. The frequency of positive cells was defined as follows: 0, less than 5%; 1, 5–25%; 2, 26–50%; 3, 51–75%; and 4, greater than 75%.
ICC Cell Lines and Culture
Human RBE and 9810 ICC cell lines were obtained from Shanghai Cell Bank (Shanghai, China). They were cultured under standard condition using Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Shanghai, China) complemented with 10% fetal bovine serum (FBS; Gibco) and 1% Streptomycin-Penicillin (100 IU/mL; Gibco) at 37°C with 5% CO2. This was followed by 1-day invasion and migration assays, 2-day qRT-PCR and WB analysis, 3-day lentivirus transfection and CCK-8 assay experiments. All operations were conducted in a biosafety cabinet after ultraviolet sterilization.
Cell Transfection
The lentiviral packaging system was composed of the vectors pGCL-puro, pHelper 1.0, and pHelper 2.0, provided by Hanyin Biotech Genechem (Shanghai, China). Three plasmids (pHelper 1.0, pHelper 2.0, and pGCL-puro or pGCL-puro-NRP1) were co-transfected into 293T cells using Lipofectamine 2000 (Invitrogen, USA). After 12 hours, the cell culture medium was replaced with fresh medium containing 10% fetal calf serum. The lentivirus supernatant containing NRP1 cDNA was collected at 48 hours after co-transfection. Lentivirus supernatant was transfected into 9810 cells with 8 μg/mL polybrene (Sigma-Aldrich, ST. Louis, MO, USA) to obtain a stable cell line, named 9810-OE, the 9810-NC was used as a control. A set of shRNA (short hairpin RNA) lentiviral vectors differed in NRP1-targeting sequences (Supplementary Table S1) and pLKO.1-puro empty vector with green fluorescent protein (GFP) were purchased from Genechem (Shanghai, China). The lentiviral packaging system was the same as mentioned above. Lentivirus supernatant was transfected into RBE cells with 8 μg/mL polybrene (Sigma-Aldrich) to obtain a stable cell line, named RBE-shNRP1. The RBE-NC was used as a control.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
In our study, qRT-PCR was performed according to a previously reported method.22 NRP1 and GAPDH (inner control) primers were synthesized and purchased from Sangon Biotech (Shanghai, China): NRP1 sense, 5′-ATCACGTGCAGCTCAAGTGG-3′, and antisense, 5′-TCATGCAGTGGGCAGAGTTC-3′; GAPDH sense, 5′-GGTATGACAACGAATTTGGC-3′, and antisense, 5′-GAGCACAGGGTACTTTATTG-3′. The experiments were repeated in triplicate.
WB Analysis
In this study, WB was performed according to a previously reported method with slight modification.24,25 Briefly, RIPA lysis buffer containing protease and phosphatase inhibitors for extracting total protein from tissues and cells was purchased from Sigma-Aldrich. Anti-human NRP1 antibody (1:2000 dilution) was purchased from Abcam. Finally, relative proteins were quantified using Image-pro plus 6.0 (Media Cybernetics, USA).
Cell Proliferation, Invasion, and Migration Assays
To explore cell proliferation activity, we performed CCK-8, assay experiments in six replicates. To evaluate cell migration capability, we performed transwell and wound healing assays separately, and the experiments were performed in triplicate. CCK-8, transwell, and wound healing assays were carried out according to a previously reported method.24 We blocked proliferation in the wound healing assays with 2% serum concentration. The variation in migration capability of cells can be determined by observing and photographing using a light microscope (Olympus Corp.; magnification, ×100).
Statistical Analysis
Data were analyzed using SPSS v.21.0 software (IBM Corp., Chicago, IL, USA). The correlation between NRP1 expression and clinicopathological characteristics were analyzed using Fisher’s exact test or Pearson chi-squared test. We used multi-factor or one-way ANOVA to analyze the significance of differences among groups. Overall survival and cumulative recurrence were described using Kaplan-Meier plots (Log rank tests). The data for cell functional assays, described as means±standard deviation, were evaluated by Mann–Whitney U-test. P<0.05 was regarded as representing a statistically significant difference.
Results
Expression of NRP1 in ICC Tissues and Its Correlation with Prognosis and Clinicopathological Characteristics
Figure 1 shows the immunohistochemical expression of NRP1 in ICC tissues and adjacent non-tumor tissues. Light microscopy showed that the expression levels of NRP1 protein between ICC and adjacent tissues were different. In total, 188 samples (64.60%) showed high NRP1 expression, while 103 samples (35.40%) had low NRP1 expression. The overall survival and cumulative tumor recurrence in ICC patients were evaluated using Kaplan-Meier analysis, thus revealing that high NRP1 expression was significantly linked with decreased overall survival and high risk of cumulative tumor recurrence (Figure 2). Further, the high NRP1 expression always indicates a poor prognosis. The results of correlation between NRP1 expression and clinicopathological characteristics (Table 1) showed that NRP1 upregulation was significantly associated with tumor number (P=0.047). No significant difference showed in age, gender, tumor size, AFP expression, CA199 expression, and TNM of NRP1 expression in patients with ICC (P>0.05). Furthermore, univariate and multivariate Cox regression analyses (Table 2) demonstrated that NRP1 is an independent prognosticator of overall survival (P=0.000) and cumulative tumor recurrence (P=0.021).
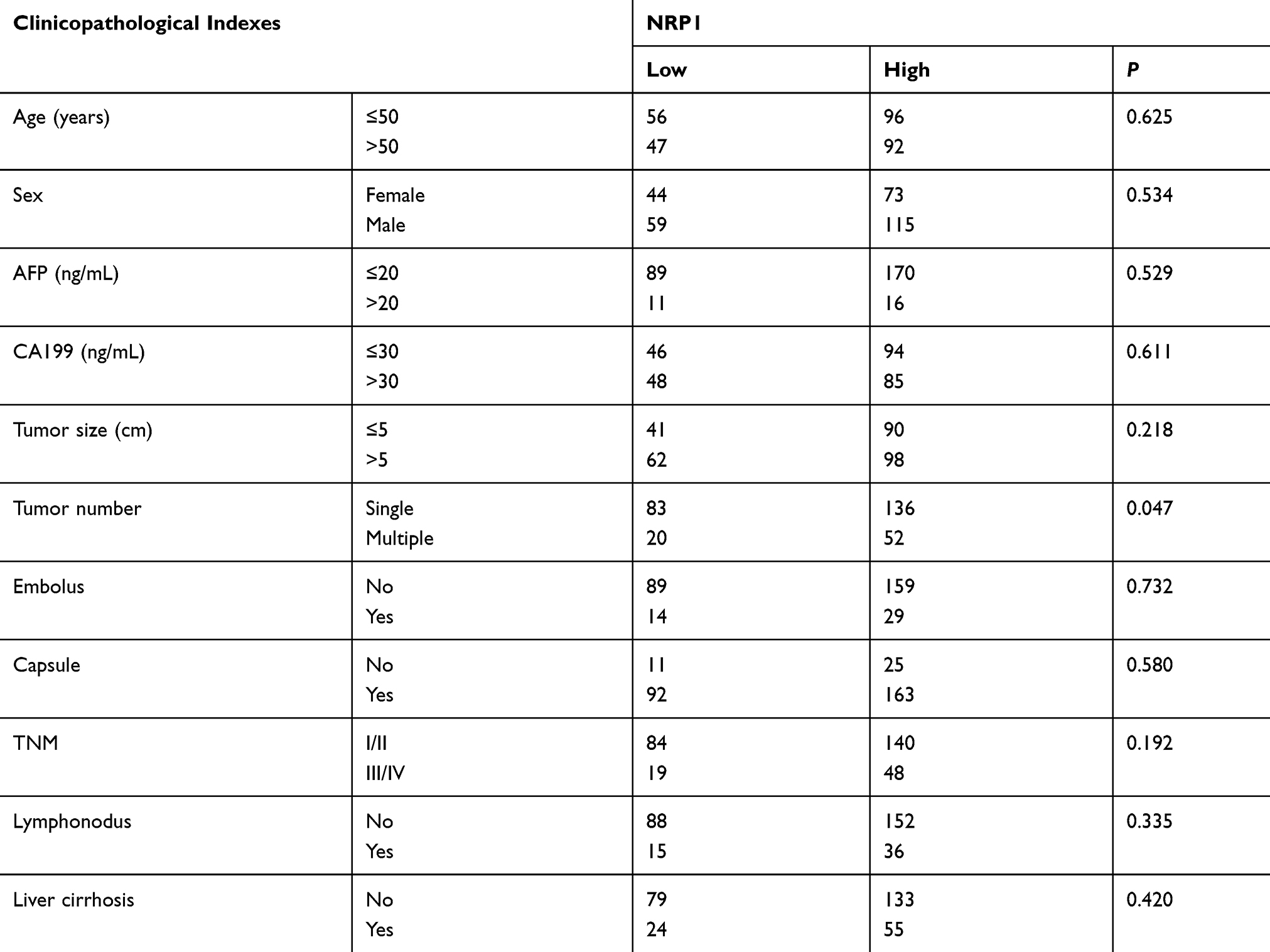 |
Table 1 Correlations Between the Factors and Clinicopathological Characteristics in ICC (n=291) |
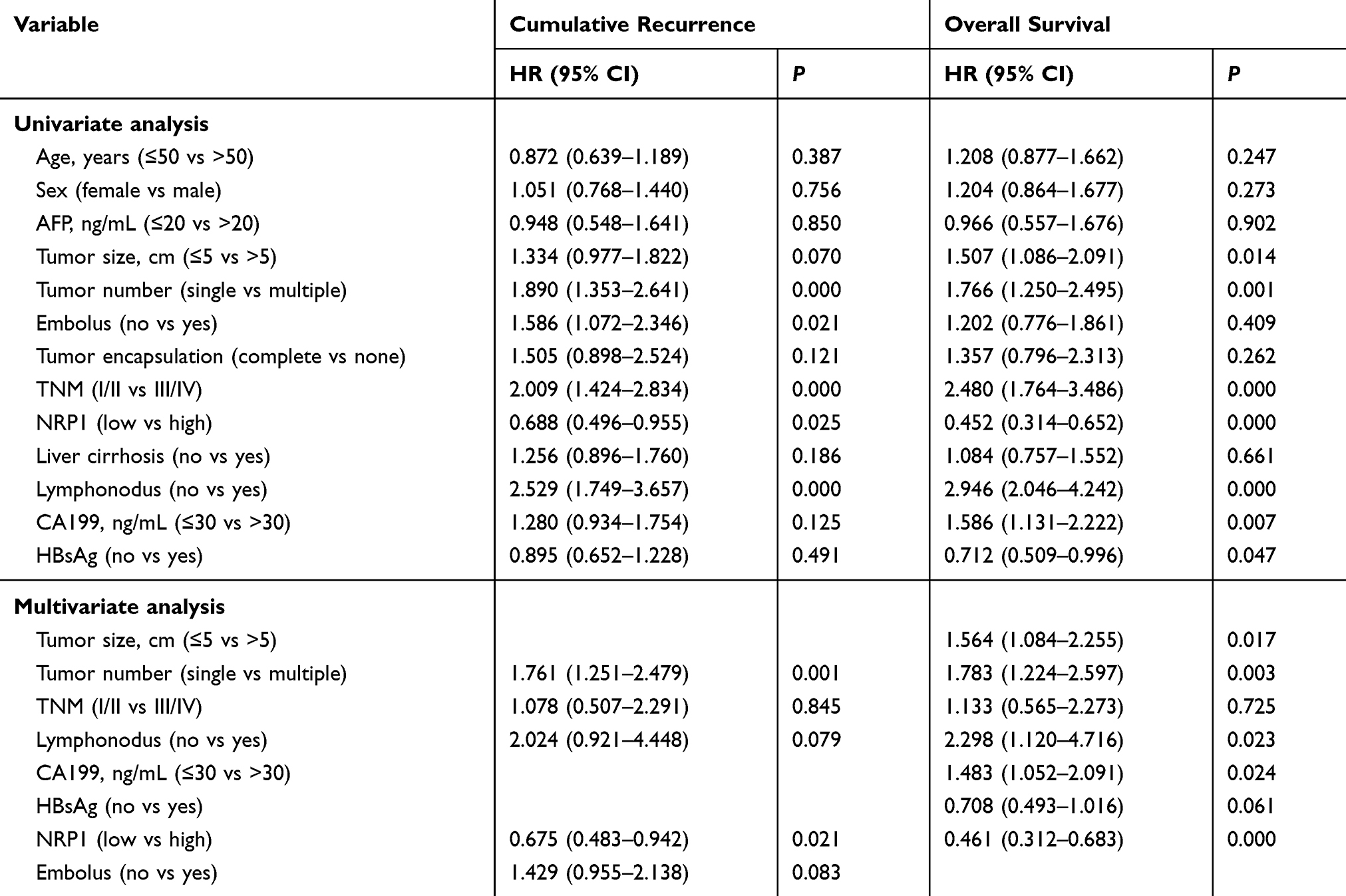 |
Table 2 Univariate and Multivariate Analyses of Prognostic Factors in ICC (n=291) |
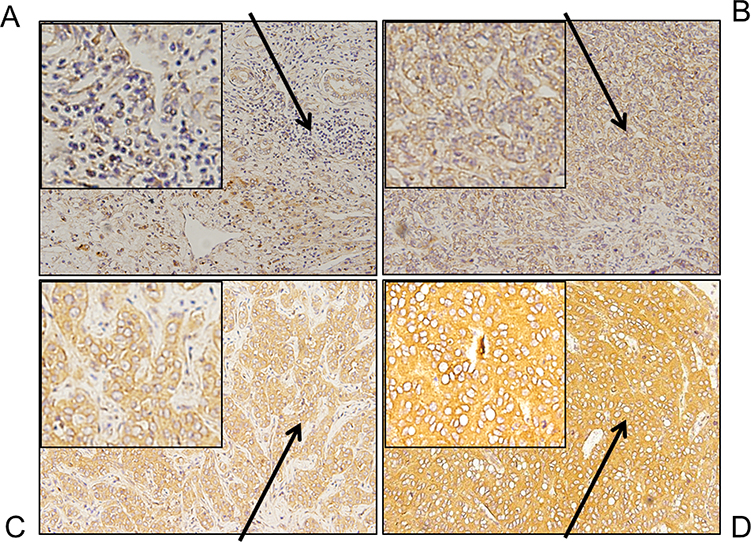 |
Figure 1 NRP1 expression detection in both ICC tissues and adjacent non-tumor tissues using IHC analysis; Magnification, ×200. (A) The staining in adjacent non-tumor tissue is weak, indicating that the expression level of NRP1 is low. (B) Weak staining in tumor tissues, indicating that the expression level of NRP1 is low. (C) Moderate staining in tumor tissues, indicating the general expression level of NRP1. (D) Strong staining in tumor tissues, indicating that the expression level of NRP1 is high. |
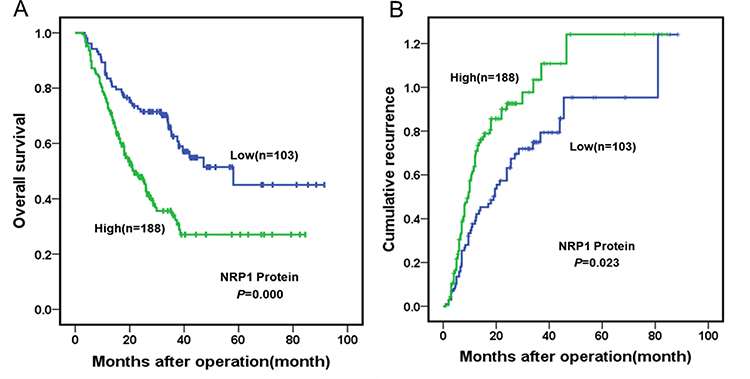 |
Figure 2 IHC staining level showing overall survival and cumulative tumor recurrence in ICC patients, evaluated using Kaplan–Meier analysis. (A) The overall survival of 103 ICC patients (35.40%) with low NRP1 expression was higher than that of 188 ICC patients (64.60%) with high NRP1 expression (P<0.001). (B) The cumulative recurrence of 103 ICC patients (35.40%) with low NRP1 expression was lower than that of 188 ICC patients (64.60%) with high NRP1 expression (P=0.023). |
Expression of NRP1 is Upregulated in Human ICCs
The expression of NRP1 was markedly increased in ICC tissues than that in adjacent non-tumor tissues at protein and mRNA levels. Specifically, NRP1 mRNA expression in 55 ICC tissues and their paired adjacent tissues were evaluated by qRT-PCR. Among the 55 groups, cancer tissues had higher levels of NRP1 mRNA than those in adjacent tissues in 33 pairs (60.00%) (Figure 3A). At the protein level, cancer tissues had higher expression levels of NRP1 proteins than those in adjacent tissues in eight out of 15 pairs (53.33%) (Figure 3D and E). Also, we assessed NRP1 protein expression levels in ICC cell lines by Western blot (WB), which revealed that the highest NRP1 expression level was found in the RBE cell line, whereas the lowest was observed in the 9810 cell line (Figure 3B and C).
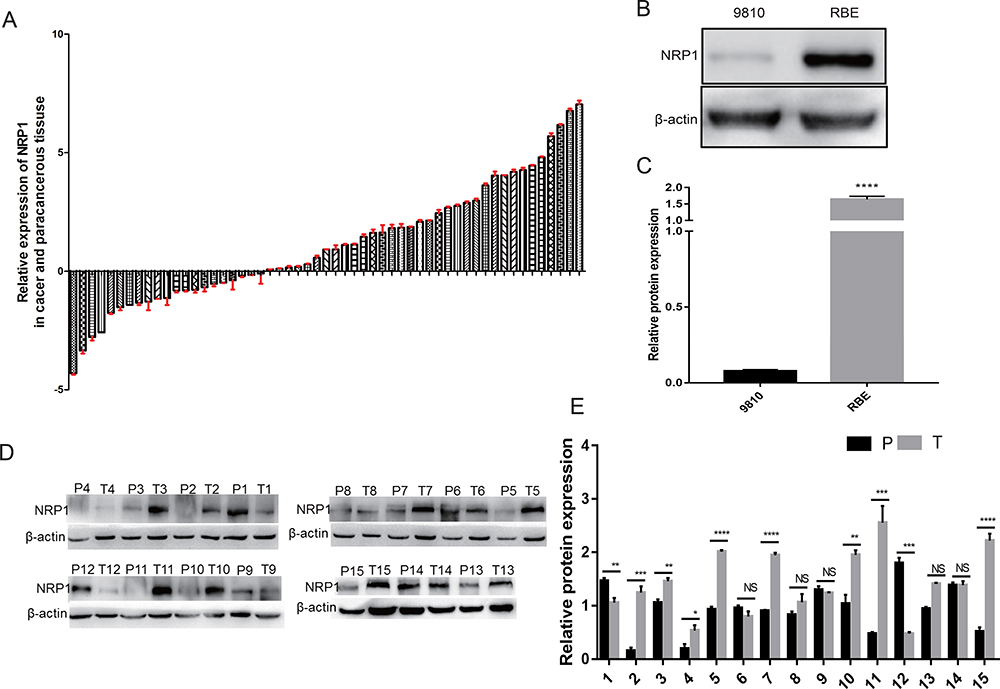 |
Figure 3 Expression difference of NRP1 mRNAs and its proteins between ICC tissues and adjacent non-tumor tissues. (A) The expression level of NRP1 mRNAs in 55 paired groups was detected using qRT-PCR (T, tumor tissue; P, adjacent non-tumor tissue). (B and C) The expression level of NRP1 proteins in ICC cell lines. (D and E) Relative NRP1 expression in 15 paired groups was analyzed using WB. * P<0.05, ** P<0.01, *** P<0.001, and **** P<0.0001. |
NRP1 Depletion Impairs Proliferation, Invasion, and Migration of RBE Cell Line in vitro
To explore the role of NRP1 in ICC cell lines based on the above experimental results, we knocked out NRP1 from the RBE cell line having high expression of NRP1. As shown in Figure 4, the normal control group was defined as the RBE-NC group and the NRP1 depletion group was defined as the RBE-shNRP1 group. After silencing, the expression of NRP1 mRNA level in the RBE-shNRP1 group decreased significantly (Figure 4A). Figure 4C presents the validation of transfection efficiency at the protein level. Figure 4B shows CCK-8, Figure4D and E show transwell, and Figure4F and G show wound healing assay results, respectively. CCK-8 assay was used to analyze cell proliferation. Transwell and wound healing assays were used to evaluate cell invasion and migration capability. The results revealed that eliminating NRP1 impairs proliferation, invasion, and migration in RBE cells.
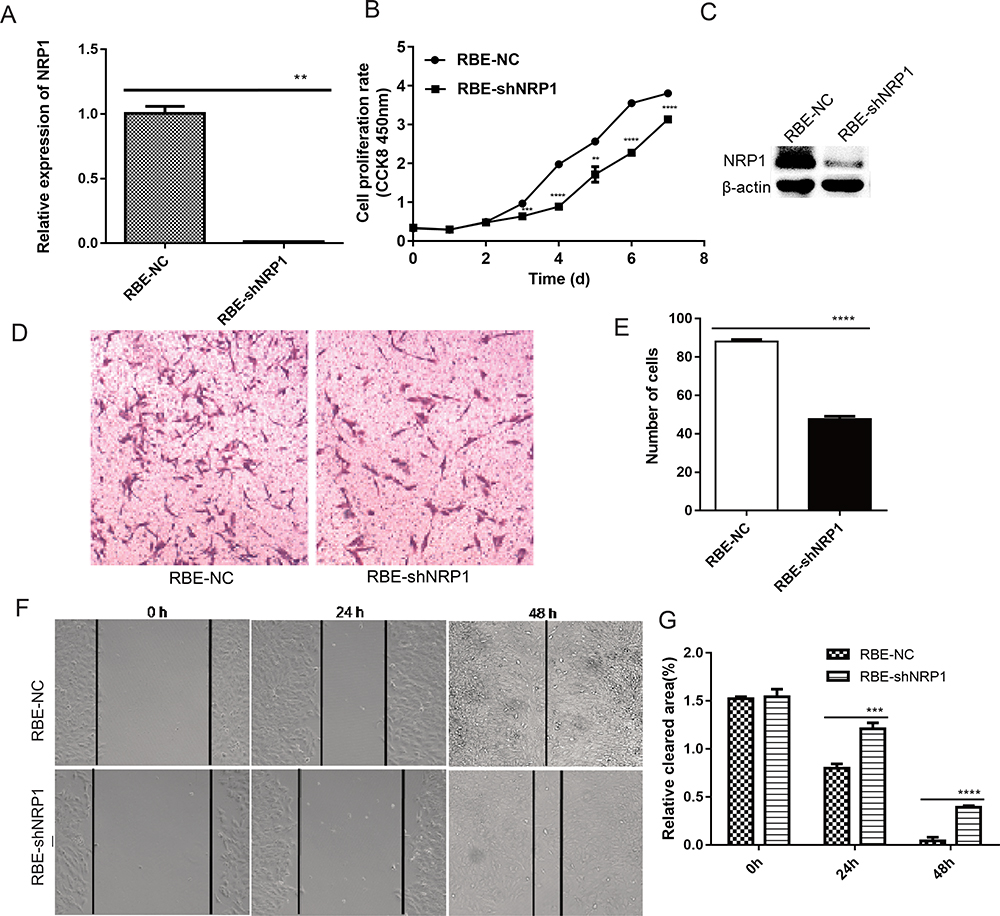 |
Figure 4 Cellular proliferation and migration comparison after NRP1 knockdown in the RBE cell line. (A) NRP1 expression levels in stably transfected RBE cells was confirmed by qRT-PCR. (B) CCK-8 assay confirmed that the proliferation ability of RBE-NC cells was better than that of RBE-shNRP1 cells. (C) Western blotting confirmed NRP1 expression levels in stably transfected ICC cells. (D and E) Transwell and (F and G) Wound healing assays of RBE-shNRP1 cells showed a considerable inhibitory effect on cell migration. Magnification, ×100. ** P<0.01, *** P<0.001, and **** P<0.0001. |
NRP1 Overexpression Promotes Proliferation, Invasion, and Migration of 9810 Cell Line in vitro
To further prove the role of NRP1 in ICC cell lines, we overexpressed NRP1 in the 9810 cell line, which has the lowest expression of NRP1. In Figure 5, the normal control group was defined as the 9810-NC group and the NRP1 overexpression group was defined as the 9810-OE-NRP1 group. After overexpression, the expression of NRP1 mRNA in the 9810-OE-NRP1 group increased significantly (Figure 5A). Figure 5C shows the validation of transfection efficiency at the protein level. Figure 5B shows the results of CCK-8, Figure 5D and E showthe results of transwell, and Figure 5F and G showwound healing assays. The results indicate that overexpression of NRP1 promoted proliferation, migration, and invasion of 9810 cells.
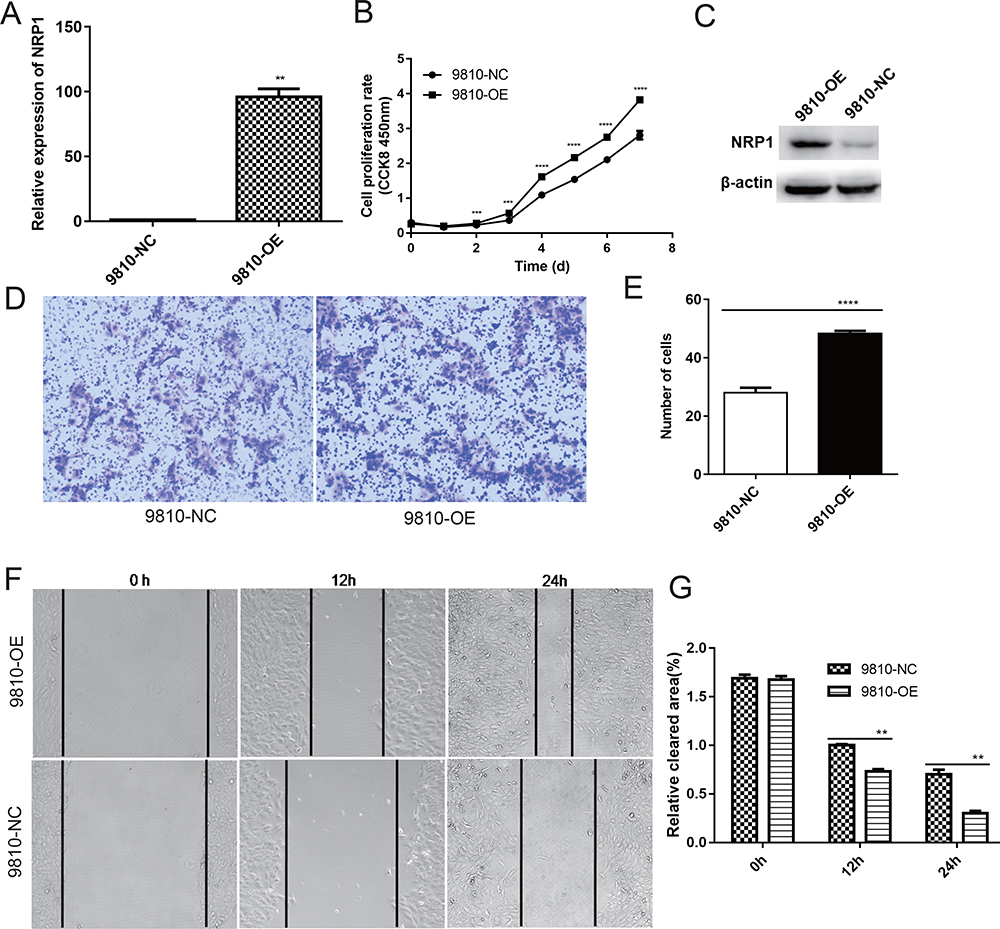 |
Figure 5 Cellular proliferation and migration comparison after NRP1 overexpression in the 9180 cell line. (A) qRT-PCR confirmed NRP1 expression levels in stably transfected ICC cells. (B) CCK-8 experiment confirmed that the proliferation ability of 9810-OE-NRP1 cells was better than that of 9810-NC cells. (C) Western blotting confirmed NRP1 expression levels in stably transfected ICC cells. (D and E) Transwell and (F and G) wound healing assays of 9810-OE-NRP1 cells showed a considerable inhibitory effect on cell migration capability. Magnification, ×100. ** P<0.01, *** P<0.001, and **** P<0.0001. |
p-FAK/p-PI3-K/p-AKT Pathway Could Be One of the Possible Mechanisms Behind the Effects of NRP1 on Cell Function
To ascertain the signaling pathways that contribute to NRP1-related ICC transformation, we used WB to analyze the expression of key proteins including NK-κB, p-FAK, p-PI3-K, Paxillin, and p-AKT in common pathways, and the relative proteins were normalized to β-actin levels. As shown in Figure 6, downregulation of NRP1 in RBE cells by shRNA significantly reduced p-FAK, p-PI3-K, and p-AKT pathway activities compared to those in RBE-NC cells. Conversely, FAK, PI3-K, and AKT were clearly activated in 9810-OE cells relative to 9810-NC cells (Figure 6). The above results indicate that NRP1 depletion inhibits the activation of the p-FAK/p-PI3-K/p-AKT pathway, while high expression of NRP1 promotes the activation of the p-FAK/p-PI3-K/p-AKT pathway.
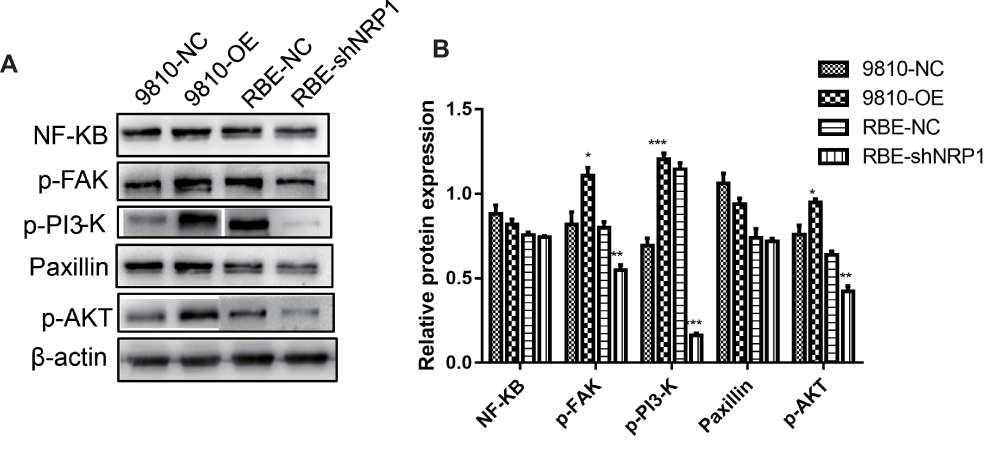 |
Figure 6 The expression level of NK-κB, p-FAK, p-PI3-K, Paxillin, and p-AKT. (A) The proteins in four types of ICC cells were detected using WB. (B) The relative proteins and the density of each band were normalized to β-actin levels. * P<0.05, ** P<0.01, and *** P<0.001. |
Discussion
ICC is a highly invasive malignant tumor associated with an extremely poor prognosis, and radical resection is the only effective treatment presently. However, a vast majority of patients have different sites of invasion and metastasis at the time of identification. Therefore, early diagnosis and prevention of ICC invasion and metastasis are key steps to enhance the efficacy of treatment.26,28 Several reports have shown that mutations or elevated expression of NRP1 are correlated with many human malignancies.17,18,20,22,29 However, the expression of NRP1 and its possible role in the occurrence, development, and prognosis of ICC have been rarely discussed. The relationship between NRP1 expression and clinicopathology in ICC remains unclear.
In our research, based on a large number of organization samples from 291 ICC patients, we preliminarily studied the expression of NRP1 in ICC tissues and its correlation with prognosis and clinicopathological characteristics, and we found that NRP1 is associated with poor prognosis and may be a cancer promoting factor. Although a previous study30 reported that NRP1 is increased in human cholangiocarcinoma tissue, they chose NRP1 based on the previous gastric cancer study, and only pay attention to the effect of NRP1 silencing on cell proliferation. More perfectly, we focused on the effect of NRP1 silencing and overexpressing on proliferation, invasion, and migration of ICC cell lines. Moreover, this paper only measured the expression of NRP1 in five patients with cholangiocarcinoma, while our research measured the expression of NRP1 in 55 patients with carcinoma and paired normal tissues.
The important role of NRP1 in many malignant tumors has prompted us to investigate the clinical relevance, prognostic evaluation, and biological effects of NRP1 in human ICC. In our study, we demonstrated the relationship between high NRP1 expression and decreased overall survival and high risk of cumulative recurrence in ICC patients. Interestingly, although 35.40% of ICC patients had lower NRP1 levels in tumors than those in the adjacent non-tumor tissues, ICC generally showed an upregulation of NRP1, and high NRP1 expression indicated poor prognosis in all ICC patients. This suggested that NRP1 upregulation might occur in parallel contributing to the oncogenic process. It is worth mentioning that the NRP1 expression was significantly correlated with tumor number, and was an independent predictor of overall survival and cumulative recurrence. Patients with low NRP1 expression were 0.461 and 0.675 times less likely to suffer from tumor recurrence and death, respectively, than those with high NRP1 expression.
However, the specific effects of NRP1 on cell function are still unclear. Accordingly, further verification was essential. Based on the above results showing that the highest NRP1 expression was found in the RBE cell line and the lowest in 9810 cells, we overexpressed the NRP1 gene in the 9810 cell line and silenced it in the RBE cell line. The cell function experiments showed that overexpression of NRP1 increased the proliferation, invasion, and migration abilities of the 9180 cell line. Conversely, knockdown of NRP1 in the RBE cell line impaired the proliferation and migration abilities of cells. These findings are in agreement with the findings of Zhu et al,30 who showed that NRP1 depletion inhibits cell migration, proliferation, and angiogenesis.
Previous research has shown that focal adhesion kinase (FAK) is a cytoplasmic non-receptor tyrosine kinase and plays a pivotal role in cell migration, adhesion, and infiltration.31 FAK is involved in many cancer cells signaling pathways, which can transmit extracellular signals to cells via integrins and growth factor receptors. It is also associated with the PI3-K/AKT signaling pathway and can promote cytoskeletal reconstruction, formation, and renewal of adhesion plaques, and the expression of matrix metalloproteinase on the cell surface, thus promoting the migration and infiltration of cancer cells.32 Numerous research has shown that the FAK/PI3-K/AKT signaling pathway is intimately connected to the occurrence and development of ICC as well as the effectiveness of chemotherapy on ICC.33,34 In this study, we further elucidated the signal transduction pathway and molecular mechanism of NRP1 in ICC. Based on the above findings, we speculated and verified that NRP1 may promote the proliferation, and migration of ICC cells via the p-FAK/p-PI3-K/p-AKT pathway, which provides a new candidate target for inhibiting ICC proliferation and migration. Similarly, tolerance of ICC to conventional chemotherapy drugs is a major challenge during ICC treatment, and the mechanism of the tolerance may be related to the activation of the PI3-K/AKT signaling pathway.35 Reducing the expression of NRP1 and inhibiting PI3-K/AKT signaling may solve the problem of ICC tolerance to conventional chemotherapeutic drugs.
It is worth mentioning that previous studies have shown that NRP-1 can result in increased phosphorylation of FAK through the VEGF/VEGFR2 pathway, which is essential cancer metastasis.36,37 NRP-1 can also activate the HGF/c-Met pathway, leading to AKT activation and downregulation of p27.37,39 In addition to the PI3-K/AKT pathway, NRP-1 can promote cholangiocarcinoma cells proliferation and metastasis through the MEK/ERK pathway.30 Although the above signaling pathways were not investigated in this study, the crosstalk among the signaling pathways may promote the proliferation, invasion, and metastasis of ICC cells.
In conclusion, our study demonstrated that NRP1 is a valuable tumor-promoting and prognostic factor. The enigma behind NRP1 upregulation needs further studies to deeply investigate, which could be useful in the development of more effective targeted strategies to prevent ICC progression. To understand the function of NRP1 in ICC more comprehensively, more research will be needed, such as in vivo animal experiments and upstream and downstream signaling pathway experiments.
Data Sharing Statement
All data generated or analyzed during the present study are included in this published article.
Ethics Approval and Consent
All experiments conducted in the present study were approved by the Ethics Committee of the First Hospital of Lanzhou University (Gansu, China), and written informed consent was obtained.
Acknowledgments
This study was supported by grants from Gansu Province Science and Technology Major Project (1602FKDA001), Lanzhou Talent Innovation and Entrepreneurship Project (2016-RC-57), Gansu Health Industry Scientific Research Plan Management Project (GWGL2014-57, GSWSKY-2015-49), and Chengguan District Science and Technology Bureau Project (2017SHFZ0014). We also thank Dr. Wen-Bo Meng and Dr. Ping Yue for their professional contributions to this study.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests.
References
1. Khan AS, Dageforde LA. Cholangiocarcinoma. Surg Clin North Am. 2019;99(2):315–335. doi:10.1016/j.suc.2018.12.004
2. Lunsford KE, Javle M, Gaber AO, Vauthey JN, Ghobrial RM. Liver transplantation for locally advanced intrahepatic cholangiocarcinoma Reply. Lancet Gastroenterol. 2018;3(8):529–530. doi:10.1016/S2468-1253(18)30167-5
3. Fujita T, Razumilava N, Gores GJ. Liver transplantation for intrahepatic cholangiocarcinoma Reply. Lancet. 2014;384(9949):1182–1183. doi:10.1016/S0140-6736(14)61716-5
4. Buettner S, van Vugt JLA, Ijzermans JN, Groot Koerkamp B. Intrahepatic cholangiocarcinoma: current perspectives. Onco Targets Ther. 2017;10:1131–1142. doi:10.2147/OTT.S93629
5. Wang K, Zhang H, Xia Y, Liu J, Shen F. Surgical options for intrahepatic cholangiocarcinoma. Hepatobiliary Surg Nutr. 2017;6(2):79–90. doi:10.21037/hbsn.2017.01.06
6. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
7. Soejima Y, Takeuchi M, Akashi T, Sawabe M, Fukusato T. beta4 and beta6 integrin expression is associated with the subclassification and clinicopathological features of intrahepatic cholangiocarcinoma. Int J Mol Sci. 2018;19(4):1004. doi:10.3390/ijms19041004
8. Yang SZ, Wang AQ, Du J, et al. Low expression of ARID1A correlates with poor prognosis in intrahepatic cholangiocarcinoma. World J Gastroenterol. 2016;22(25):5814–5821. doi:10.3748/wjg.v22.i25.5814
9. Tsai Y-CI, Fotinou C, Rana R, et al. Structural studies of neuropilin-2 reveal a zinc ion binding site remote from the vascular endothelial growth factor binding pocket. FEBS J. 2016;283(10):1921–1934. doi:10.1111/febs.13711
10. Wild JR, Staton CA, Chapple K, Corfe BM. Neuropilins: expression and roles in the epithelium. Int J Exp Pathol. 2012;93(2):81–103. doi:10.1111/j.1365-2613.2012.00810.x
11. Roy S, Bag AK, Singh RK, Talmadge JE, Batra SK, Datta K. Multifaceted role of neuropilins in the immune system: potential targets for immunotherapy. Front Immunol. 2017;8:1228. doi:10.3389/fimmu.2017.01228
12. Napolitano V, Tamagnone L. Neuropilins controlling cancer therapy responsiveness. Int J Mol Sci. 2019;20(8):2049. doi:10.3390/ijms20082049
13. Hu C, Jiang X. Role of NRP-1 in VEGF-VEGFR2-independent tumorigenesis. Target Oncol. 2016;11(4):501–505. doi:10.1007/s11523-016-0422-0
14. Prud’homme GJ, Glinka Y. Neuropilins are multifunctional coreceptors involved in tumor initiation, growth, metastasis and immunity. Oncotarget. 2012;3(9):921–939. doi:10.18632/oncotarget.626
15. Torres-Salido MT, Sanchis M, Sole C, et al. Urinary neuropilin-1: a predictive biomarker for renal outcome in lupus nephritis. Int J Mol Sci. 2019;20(18):4601. doi:10.3390/ijms20184601
16. Chaudhary B, Khaled YS, Ammori BJ, Elkord E. Neuropilin 1: function and therapeutic potential in cancer. Cancer Immunol Immunother. 2014;63(2):1–99. doi:10.1007/s00262-013-1500-0
17. Bergé M, Allanic D, Bonnin P, et al. Neuropilin-1 is upregulated in hepatocellular carcinoma and contributes to tumour growth and vascular remodelling. J Hepatol. 2011;55(4):866–875. doi:10.1016/j.jhep.2011.01.033
18. Peng Y, Liu Y-M, Li L-C, Wang -L-L, Wu X-L. MicroRNA-338 inhibits growth, invasion and metastasis of gastric cancer by targeting NRP1 expression. PLoS One. 2014;9(4):e94422–e94422. doi:10.1371/journal.pone.0094422
19. Glinka Y, Mohammed N, Subramaniam V, Jothy S, Prud’homme GJ. Neuropilin-1 is expressed by breast cancer stem-like cells and is linked to NF-kappaB activation and tumor sphere formation. Biochem Biophys Res Commun. 2012;425(4):775–780. doi:10.1016/j.bbrc.2012.07.151
20. Naik A, Al-Zeheimi N, Bakheit CS, et al. Neuropilin-1 associated molecules in the blood distinguish poor prognosis breast cancer: a cross-sectional study. Sci Rep. 2017;7(1):3301. doi:10.1038/s41598-017-03280-0
21. Tse BWC, Volpert M, Ratther E, et al. Neuropilin-1 is upregulated in the adaptive response of prostate tumors to androgen-targeted therapies and is prognostic of metastatic progression and patient mortality. Oncogene. 2017;36(24):417–3427. doi:10.1038/onc.2016.482
22. Shi S, Lu Y, Qin Y, et al. miR-1247 is Correlated with Prognosis of Pancreatic Cancer and Inhibits Cell Proliferation by Targeting Neuropilins. Curr Mol Med. 2014;14(3):316–327. doi:10.2174/1566524014666140228120014
23. Bi C, Liu M, Rong W, et al. High Beclin-1 and ARID1A expression corelates with poor survival and high recurrence in intrahepatic cholangiocarcinoma: a histopathological retrospective study. BMC Cancer. 2019;19(1):213. doi:10.1186/s12885-019-5429-3
24. Gao ZF, Wu YN, Bai ZT, Zhang L, Zhou Q, Li X. Tumor-suppressive role of HACE1 in hepatocellular carcinoma and its clinical significance. Oncol Rep. 2016;36(6):3427–3435. doi:10.3892/or.2016.5205
25. Ohara K, Horibe T, Kohno M, Kawakami K. Characterization of antilytic peptide antibody: application for the detection of lytic-based hybrid peptide in serum samples. J Pept Sci. 2011;17(7):493–498. doi:10.1002/psc.1349
26. Rahnemai-Azar AA, Weisbrod AB, Dillhoff M, Schmidt C, Pawlik TM. Intrahepatic cholangiocarcinoma: current management and emerging therapies. Expert Rev Gastroenterol Hepatol. 2017;11(5):439–449. doi:10.1080/17474124.2017.1309290
27. Zhu AX, Hezel AF. Development of molecularly targeted therapies in biliary tract cancers: reassessing the challenges and opportunities. Hepatology. 2011;53(2):695–704. doi:10.1002/hep.24145
28. Guion-Dusserre JF, Lorgis V, Vincent J, Bengrine L, Ghiringhelli F. FOLFIRI plus bevacizumab as a second-line therapy for metastatic intrahepatic cholangiocarcinoma. World J Gastroenterol. 2015;21(7):2096–2101. doi:10.3748/wjg.v21.i7.2096
29. Ueyama H, Horibe T, Nakajima O, Ohara K, Kohno M, Kawakami K. Semaphorin 3A lytic hybrid peptide binding to neuropilin-1 as a novel anti-cancer agent in pancreatic cancer. Biochem Biophys Res Commun. 2011;414(1):60–66. doi:10.1016/j.bbrc.2011.09.021
30. Zhu H, Jiang X, Zhou X, et al. Neuropilin-1 regulated by miR-320 contributes to the growth and metastasis of cholangiocarcinoma cells. Liver Int. 2018;38(1):125–135. doi:10.1111/liv.13495
31. Zhou J, Yi Q, Tang L. The roles of nuclear focal adhesion kinase (FAK) on cancer: a focused review. J Exp Clin Cancer Res. 2019;38(1):250. doi:10.1186/s13046-019-1265-1
32. Lee BY, Timpson P, Horvath LG, Daly RJ. FAK signaling in human cancer as a target for therapeutics. Pharmacol Ther. 2015;146:132–149. doi:10.1016/j.pharmthera.2014.10.001
33. Chen M-H, Chiang K-C, Cheng C-T, et al. Antitumor activity of the combination of an HSP90 inhibitor and a PI3K/mTOR dual inhibitor against cholangiocarcinoma. Oncotarget. 2014;5(9):2372–2389. doi:10.18632/oncotarget.1706
34. Yothaisong S, Dokduang H, Techasen A, et al. Increased activation of PI3K/AKT signaling pathway is associated with cholangiocarcinoma metastasis and PI3K/mTOR inhibition presents a possible therapeutic strategy. Tumor Biol. 2013;34(6):3637–3648. doi:10.1007/s13277-013-0945-2
35. McRee AJ, Sanoff HK, Carlson C, Ivanova A, O’Neil BH. A Phase I trial of mFOLFOX6 combined with the oral PI3K inhibitor BKM120 in patients with advanced refractory solid tumors. Invest New Drugs. 2015;33(6):1225–1231. doi:10.1007/s10637-015-0298-3
36. Hamerlik P, Lathia JD, Rasmussen R, et al. Autocrine VEGF - VEGFR2 – neuropilin-1 signaling promotes glioma stem-like cell viability and tumor growth. J Exp Med. 2012;209(3):507–520. doi:10.1084/jem.20111424
37. Barr MP, Gray SG, Gately K, et al. Vascular endothelial growth factor is an autocrine growth factor, signaling through neuropilin-1 in non-small cell lung cancer. Mol Cancer. 2015;14:45. doi:10.1186/s12943-015-0310-8
38. Shang N, Arteaga M, Zaidi A, et al. FAK is required for c-Met/beta-catenin-driven hepatocarcinogenesis. Hepatology. 2015;61(1):214–226. doi:10.1002/hep.27402
39. Barat S, Bozko P, Chen X, et al. Targeting c-MET by LY2801653 for treatment of cholangiocarcinoma. Mol Carcinog. 2016;55(12):2037–2050. doi:10.1002/mc.22449
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.