Back to Journals » OncoTargets and Therapy » Volume 16
Noncoding RNA-Mediated High Expression of PFKFB3 Correlates with Poor Prognosis and Tumor Immune Infiltration of Lung Adenocarcinoma
Authors Gu X, Li X, Zhang X, Tong L, Feng R, Liu L, Sun H, Zhang Q, Bian T, Zhang J, Gao L, Zhang C, Liu J, Liu Y ![]()
Received 7 April 2023
Accepted for publication 10 September 2023
Published 22 September 2023 Volume 2023:16 Pages 767—783
DOI https://doi.org/10.2147/OTT.S416155
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Daniel Neureiter
Xue Gu,1,* Xiaoli Li,1,* Xue Zhang,2,* Li Tong,3 Ran Feng,1 Lei Liu,1 Hui Sun,1 Qing Zhang,1 Tingting Bian,1 Jianguo Zhang,1 Lihua Gao,2 Chenxi Zhang,4 Jian Liu,2 Yifei Liu1
1Department of Pathology, Affiliated Hospital of Nantong University, Medical School of Nantong University, Nantong, 226001, People’s Republic of China; 2Department of Oncology, Affiliated Hospital of Nantong University, Medical School of Nantong University, Nantong, 226001, People’s Republic of China; 3Department of Pathology, Affiliated Hospital of Nantong University, Dalian Medical University, Nantong, 226001, People’s Republic of China; 4Medical School of Nantong University, Nantong, 226001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yifei Liu; Jian Liu, Email [email protected]; [email protected]
Background: There is growing evidence showing that 6-phosphofructo-2-kinase (PFKFB3) plays crucial roles in different types of human cancers, including LUAD; however, the specific mechanism by which PFKFB3 plays a role in LUAD remains unclear.
Methods: We investigated the expression of PFKFB3 and explored the underlying mechanism as well as the correlation with immune markers using several online datasets, such as Tumor Immune Estimate Resource (TIMER), UALCAN, and the Cancer Genome Atlas (TCGA) databases, miRWalk, Targetscan, MiRDB and starBase database. Western blot and immunohistochemistry analysis were performed to verify the corresponding outcomes.
Results: It was shown that the mRNA expression of PFKFB3 was lower in LUAD than in the normal tissues, while its protein expression was not consistent with the mRNA level. The expression of PFKFB3 was correlated with clinicopathological parameters and several signaling pathways. The potential long chain (lnc)RNA/microRNA/PFKFB3 axis and the possible mechanism by which tumor progression in LUAD is promoted was predicted. We obtained the LINC01798/LINC02086/AP000845.1/HAGLR-miR-17-5p-PFKFB3 axis after comprehensive analyses of expression, correlation, and survival. Moreover, the expression of PFKFB3 was positively correlated with immune cells and immune checkpoint expression, including PD-1, PD-L1 and CTLA-4.
Conclusion: The present study demonstrated that noncoding RNAs mediated the upregulation of PFKFB3 and was associated with a poor prognosis and immune tumor infiltration in LUAD.
Keywords: PFKFB3, miR-17-5p, lncRNA, tumor immune infiltration, PD-1/PD-L1
Introduction
In accordance with 2020 global cancer statistics, the incidence of lung cancer accounted for 11.4% out of 19.3 million new cases, and its mortality rate was higher than that of all human cancers in both men and women.1 LUAD is the most common histological subtype and is considered to be closely related to smoking habits. Currently, comprehensive treatment is the main therapy for lung cancer. Targeted and immunized drugs are available; however, the therapy for lung cancer depends on its subtype and stage, as well as tumor size and the general condition of the patients.2 Even though much progress has been made in the treatment of lung cancer, the prognosis for patients is still disappointing; therefore, searching for a new prognostic molecular marker to help doctors diagnose and treat lung cancer is essential.
6-phosphofructo-2-kinase (PFKFB3), as well as fructose-2,6-bisphosphatase 3 (F26BP3) with a molecular weight of ~58 kDa belongs to the PFKFB family, which is a key enzyme in glycolysis progression.3 PFKFB3 has been reported to be upregulated in a majority of human cancers, such as breast cancer (BC),4 gastric cancer (GC),5 colorectal cancer (CRC),6 and nasopharyngeal carcinoma (NPC).7 Recently, PFKFB3 has been identified as one of the new kinases involved in cell metabolism and plays an important role in regulating cell growth and the proliferation in GC.5 The overexpression or overactivation of PFKFB3 is closely related to many tumor diseases and has become a new target for antitumor drug research and development; however, the specific mechanism is less clear. Therefore, exploring its pathogenic mechanism in LUAD is of great significance for clinical treatment.
In the present study, we analyzed the expression of PFKFB3 along with its clinicopathological relationship and function in LUAD. We also investigated the upstream molecular of PFKFB3 and determined its correlation with immune cells or immune checkpoints. Taken together, the noncoding (nc)RNA-mediated axis related to PFKFB3 may promote tumor progression in LUAD.
Materials and Methods
TIMER Database
TIMER (https://cistrome.shinyapps.io/timer/) is a web resource for systematic evaluations of the clinical impact of different immune cells in diverse cancer types. The abundance of six immune infiltrates (B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells) are estimated using the TIMER (https://cistrome.shinyapps.io/timer/) algorithm. We analyzed the mRNA expression of PFKFB3 in various human cancer types and the correlation between PFKFB3 and immune cell markers. P < 0.05 was considered statistically significant.
UALCAN Database
The UALCAN database (http://ualcan.path.uab.edu/) is a comprehensive, user-friendly, and interactive web resource that can be used to perform in-depth analysis of TCGA data (https://portal.gdc.cancer.gov/) of gene expression.8 Here, we investigated the mRNA and protein level of PFKFB3 along with the relationship between PFKFB3 and the clinicopathological parameters.
LinkedOmics Database
LinkedOmics database (www.linkedomics.org/) contains multi-omics data and clinical data for 32 cancer types and 11,158 patients from the TCGA project.9 We used it to analyze the function of PFKFB3 in LUAD using 515 patients.
The miRWalk, Targetscan, MiRDB and starBase Database
MiRWalk (http://mirwalk.umm.uni-heidelberg.de/),10–12 Targetscan (http://www.targetscan.org/), and miRDB (http://www.mirdb.org/)13–15 databases are online resources specialized in predicting microRNA (miRNA) target and function annotation. StarBase (https://starbase.sysu.edu.cn/) is an online website dedicated to analyzing the interaction between RNA and RNA, including mRNA, miRNA, and lncRNA.16 It is so portable that users can easily find the possible upstream molecules of target gene. Here, after comprehensive analyses of those possible miRNAs, only those that appeared in all three databases were selected as candidate upstream molecules of PFKFB3. We then analyzed a series of data comprising the correlation expression between miRNA-PFKFB3, lncRNA-miRNA, and the expression of each of miRNA in LUAD and the normal control.
Gene Expression Profiling Interactive Analysis Database
GEPIA (http://gepia.cancer-pku.cn/) is an online database based on TCGA and GTEx data that provides crucial interactive and customizable functions, such as differential expression analysis in multiple types of human cancers and their adjacent normal tissues, patient survival analysis, and correlation analysis between any two molecules.17 GEPIA was used to analyze the correlation expression between PFKFB3 and immune cells and immune checkpoint markers in LUAD. |R| > 0.1 and p < 0.05 were set as the selection criteria for identifying as statistically significant.
Cell Culture
A549 (cat. no. SCSP-503) and H1299 (cat. no. SCSP-589) were purchased from the cell bank of the Chinese Academy of Sciences (Shanghai, China). 16HBE (cat. no. CL-0249) was purchased from Procell Life Science & Technology Company (Wuhan, China). Cells were maintained in RPMI-1640 medium (Servicebio, Wuhan, China) supplemented with 10% fetal bovine serum (FBS; AusGeneX, Brisbane, Australia) and 1% penicillin/streptomycin solution.
Western Blotting
Protein was extracted from cells and tissues using RIPA lysis buffer supplemented with phenylmethylsulfonyl fluoride (Biotite Biotechnology, Shanghai, China). After incubating at 4°C for 30 min, the proteins were centrifuged at 12,000 rpm for 30 min. Thirty micrograms of protein were separated using sodium dodecyl sulphate–polyacrylamide gel electrophoresis gel, and then transferred onto polyvinylidene difluoride (PVDF) membranes. After blocking with 5% skim milk at room temperature for 2 h, the membranes were incubated with rabbit anti-GAPDH polyclonal antibody (cat. no. 10494-1-AP, Proteintech, Wuhan, China) and rabbit anti-PFKFB3 polyclonal antibody (cat. no. 13763-1-AP, Proteintech, Wuhan, China) at 4°C overnight, respectively. The membranes were then incubated with donkey HRP-conjugated anti-rabbit secondary antibody (cat. no. ab216773, Abcam, Cambridge, MA, USA) for 2 h at room temperature. The bands were detected by enhanced chemiluminescence (ECL) kit (BIO-RAD, USA).
Immunohistochemistry
After dewaxing, hydration, antigen retrieval, and adding endogenous peroxidase and a series of treatments, the LUAD and adjacent normal tissues were stained with rabbit anti-PFKFB3 (cat. no. 13763-1-AP, Proteintech, Wuhan, China) at room temperature for 1 h, followed by staining with anti-rabbit secondary antibody (HRP) at room temperature for ~30 min. Finally, the images were captured using a Smart digital camera (JEDA, Nanjing, China).
Statistical Analysis
All experiments were conducted at least three times. Data are presented as the mean ± standard error of mean (SEM). GraphPad Prism (version 6.01; GraphPad Software, Inc, San Diego, CA, USA) was used to create the visualizing graphs. For statistical analyses between different treatment groups, a parametric two-sided t-test was used. The statistical analyses in this study were calculated using the online database mentioned above. P < 0.05 or log rank p < 0.05 was considered statistically significant.
Results
Expression of PFKFB3 mRNA in Human Cancers
To analyze mRNA expression in different types of human cancers, we used the Tumor Immune Estimation Resource (TIMER) database (https://cistrome.shinyapps.io/timer/) and found that PFKFB3 mRNA expression was significantly downregulated in bladder urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA), kidney chromophobe (KICH), kidney renal papillary cell carcinoma (KIRP), liver hepatocellular carcinoma (LIHC), LUAD, lung squamous cell carcinoma (LUSC), and prostate adenocarcinoma (PRAD). In addition, the mRNA expression of PFKFB3 was significantly higher in cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), head and neck squamous cell carcinoma (HNSC), kidney renal clear cell carcinoma (KIRC), stomach adenocarcinoma (STAD), and thyroid carcinoma (THCA) than in their paired normal tissues (Figure 1A). To verify the accuracy of the analysis from the database, we downloaded LUAD data from the Cancer Genome Atlas (TCGA) database (https://portal.gdc.cancer.gov/) and discovered that the level of PFKFB3 mRNA was lower in LUAD than in paracancerous tissues (Figure 1B). Similar results were obtained from the UALCAN database (http://ualcan.path.uab.edu/) (Figure 1C). In brief, the result was inconsistent among previous research and multiple online databases.
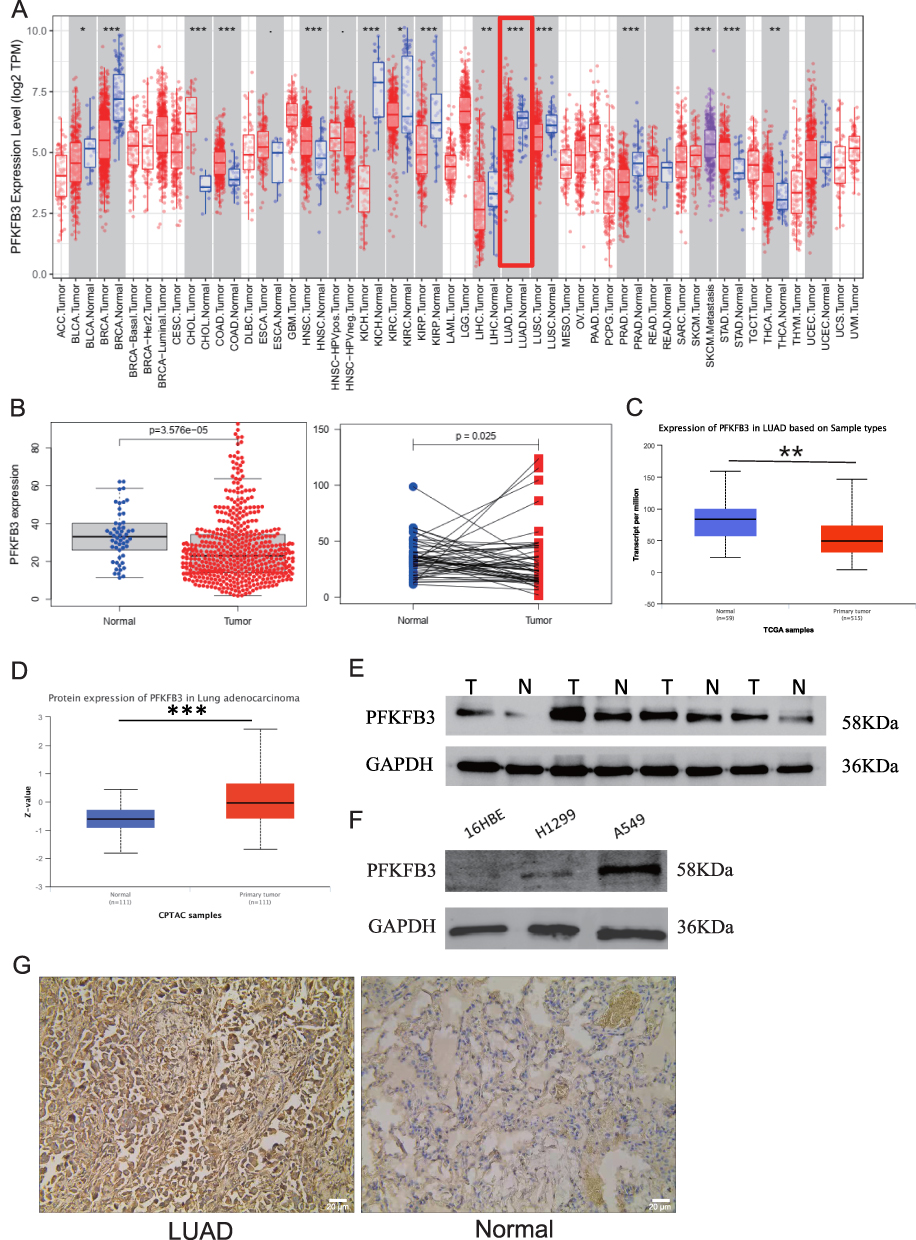 |
Figure 1 Expression analysis of 6-phosphofructo-2-kinase (PFKFB3) mRNA and protein in lung adenocarcinoma (LUAD). Notes: (A) Expression of PFKFB3 mRNA in multiple human cancers based on the Cancer Genome Atlas (TCGA) database (ttps://portal.gdc.cancer.gov/) of cancer and normal tissues. (B) PFKFB3 mRNA expression in LUAD from data downloaded from the TCGA database. (C) Expression of PFKFB3 mRNA in LUAD from the UALCAN database (http://ualcan.path.uab.edu/). (D) Expression of PFKFB3 protein in LUAD from the UALCAN database (http://ualcan.path.uab.edu/). (E) Expression of PFKFB3 protein in LUAD and adjacent normal tissues as determined by Western blotting. (F) Expression of PFKFB3 protein in various LUAD cell lines (H1299, A549) and in a normal lung cell line (16HBE) as determined by Western blotting. (G) Immunohistochemistry stain used to detected PFKFB3 protein levels in LUAD tissues and normal lung tissues. Scale bars: 20μm. *p < 0.05, **p < 0.01, ***p < 0.001. |
Protein Expression of PFKFB3 in LUAD
To investigate the level of PFKFB3 protein expression, we examined the UALCAN database (http://ualcan.path.uab.edu/) and discovered that the PFKFB3 protein level was higher in LUAD than in normal tissues (Figure 1D). In support of this, Western blotting and immunohistochemistry (IHC) were conducted (Figure 1E–G). PFKFB3 staining was localized in the cell membrane and cytoplasm.
Relationship Between PFKFB3 Expression and Clinicopathological Parameters
We also used the UALCAN database (http://ualcan.path.uab.edu/) to analyze the relationship between PFKFB3 expression and the clinicopathological parameters in LUAD patients. Obviously, the differences in PFKFB3 mRNA were related to age, sex, race, smoking habits, disease stage, and nodal metastasis status (Figure 2).
 |
Figure 2 Correlation analysis between 6-phosphofructo-2-kinase (PFKFB3) and clinicopathological parameters in lung adenocarcinoma (LUAD). Notes: (A–F) Expression of PFKFB3 in LUAD based on patient age, sex, race, smoking habits, disease stage, and nodal metastasis status. *p < 0.05, **p < 0.01, ***p < 0.001. |
PFKFB3 Enrichment
Gene ontology (GO) analysis was conducted using the LinkedOmics database (www.linkedomics.org/) to clarify the biological function of PFKFB3 expression. As shown in Figure S1A–C, PFKFB3 expression was most positively related to extracellular structure organization and negatively associated with the mitochondrial respiratory chain complex assembly as they related to biological progress. From the perspective of cellular components, ECM and the mitochondrial protein complex were the most prominent signaling pathways related positively and negatively to PFKFB3, respectively. Molecular function showed that PFKFB3 expression had a positive connection to cytokine binding and a negative connection to rRNA binding.
Prediction and Analyses of Upstream PFKFB3 miRNAs
It has been widely acknowledged that ncRNAs are responsible for the regulation of gene expression.18,19 To ascertain whether PFKFB3 was modulated by some specific ncRNAs, we first predicted upstream miRNAs that could potentially bind to PFKFB3 and chose seven miRNAs as molecular candidates. To improve visualization, a miRNA–PFKFB3 network was established using Cytoscape (https://cytoscape.org/) (Figure 3A). We then analyzed the correlation between PFKFB3 and each of candidate miRNAs using starBase website (https://starbase.sysu.edu.cn/). The results in Figure 3B–H show a positive correlation between miR-133b and PFKFB3. The remaining five miRNAs were negatively correlated with PFKFB3 after ruling out miR-3666, which showed no relationship to PFKFB3. In addition, there was no statistical relationship between PFKFB3 and miR-20b-5p. We then attempted to determine whether each miRNA had a significant difference in expression between cancer and adjacent normal tissues as well as the overall survival rate. Last, we determined the expression and prognostic value of miR-17-5p (Figure 4A–I). To sum, miR-17-5p was markedly upregulated in LUAD and this upregulation was positively linked to patient prognosis. These findings suggested that miR-17-5p might be the most potential regulatory PFKFB3 miRNA in LUAD.
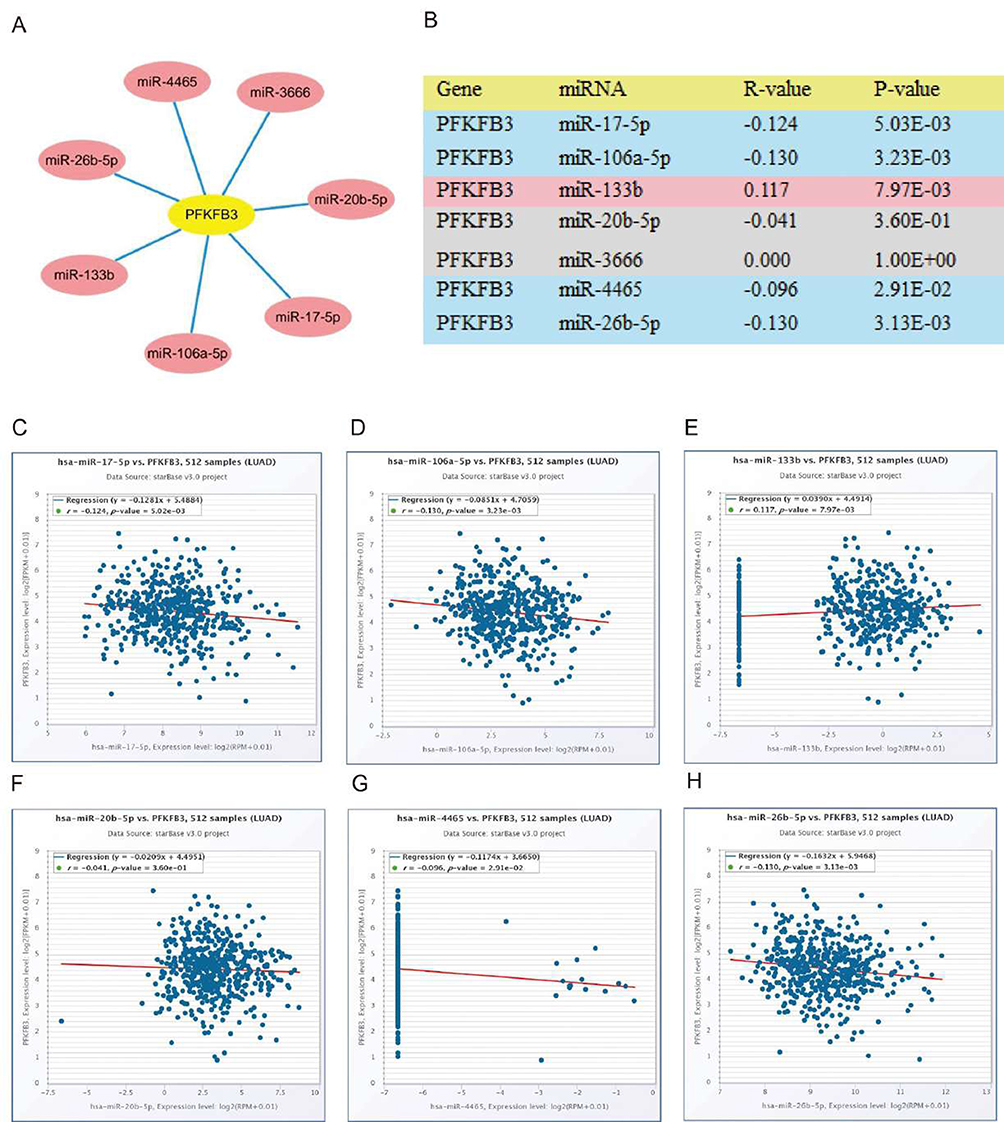 |
Figure 3 Identification of miR-17-5p as a potential upstream 6-phosphofructo-2-kinase (PFKFB3) miRNA in lung adenocarcinoma (LUAD). Notes: (A) The miRNA–PFKFB3 regulatory network established by Cytoscape (https://cytoscape.org/). (B) The expression correlation between predicted miRNAs and PFKFB3 in LUAD analyzed using starBase database. (C–H) Correlation analysis between PFKFB3 and miRNAs in LUAD using starBase database. |
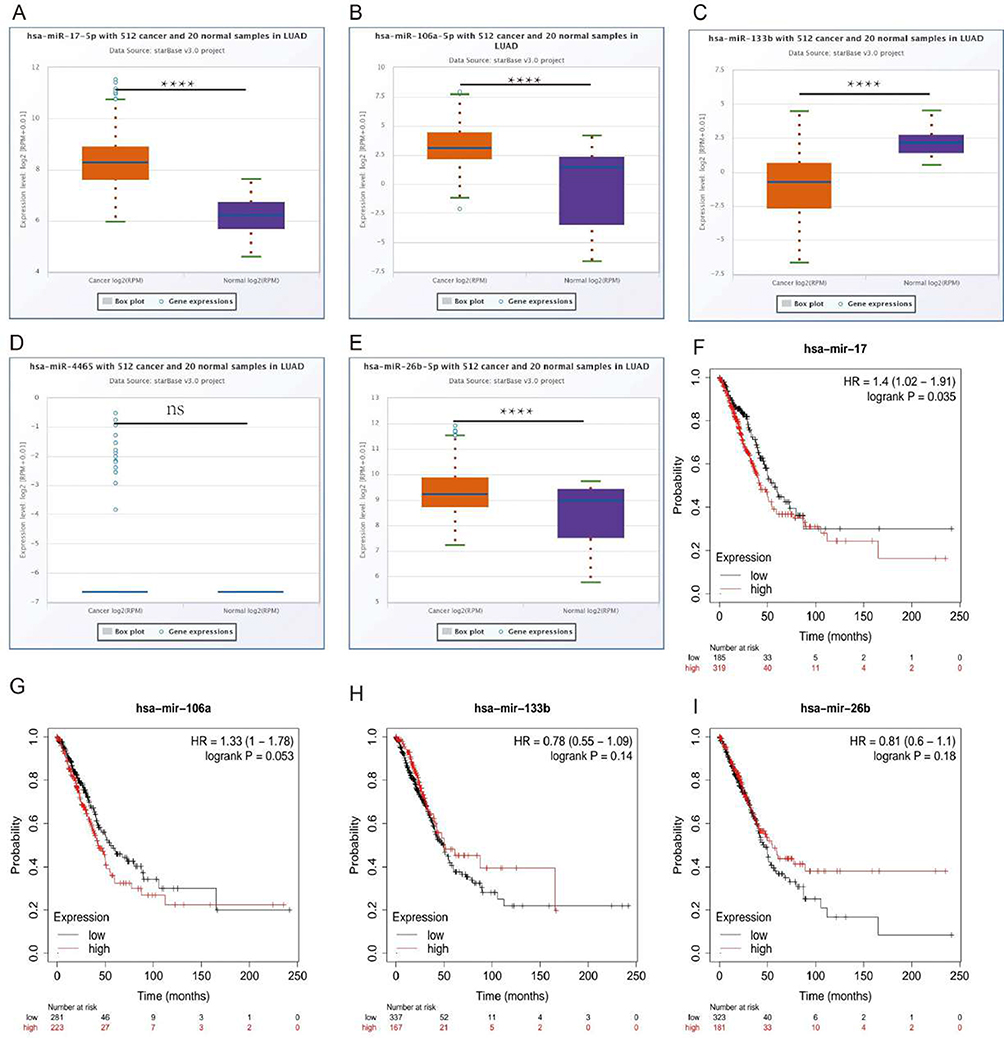 |
Figure 4 Differential expression analysis and survival analysis of microRNAs. Notes: (A–E) Expression of each miRNA in LUAD and adjacent normal tissues determined by starBase database. (F–I) Survival analysis of each miRNA in LUAD. ****p < 0.0001. |
Prediction and Analysis of Upstream Long Noncoding RNAs of miR-17-5p
We then predicted the potential long noncoding (lnc)RNAs of miR-17-5p using starBase v 3.0 and found 216 possible lncRNAs. Considering the correlation between each lncRNA and each miRNA, the differential expression of each lncRNA between LUAD and normal tissues and whether their overall survival is meaningful, AP000845.1, HAGLR, LINC01798, and LINC02086 were chosen as candidate lncRNAs (Figure 5A–L). To further investigate, AP000845.1 and LINC02086 were positively correlated with miR-17-5p, while the other two were negatively correlated. In addition, the expressions of HAGLR and LINC01798 were downregulated in LUAD, and their downregulation was associated with a poor positive prognosis; however, there were exceptions. Even though the expression of AP000845.1 was higher in cancer than in adjacent normal tissue, its upregulation was negatively correlated with a poor prognosis. We could conclude that AP000845.1, HAGLR, LINC01798, and LINC02086 were the most potential upstream lncRNAs of the miR-17-5p/PFKFB3 axis in LUAD.
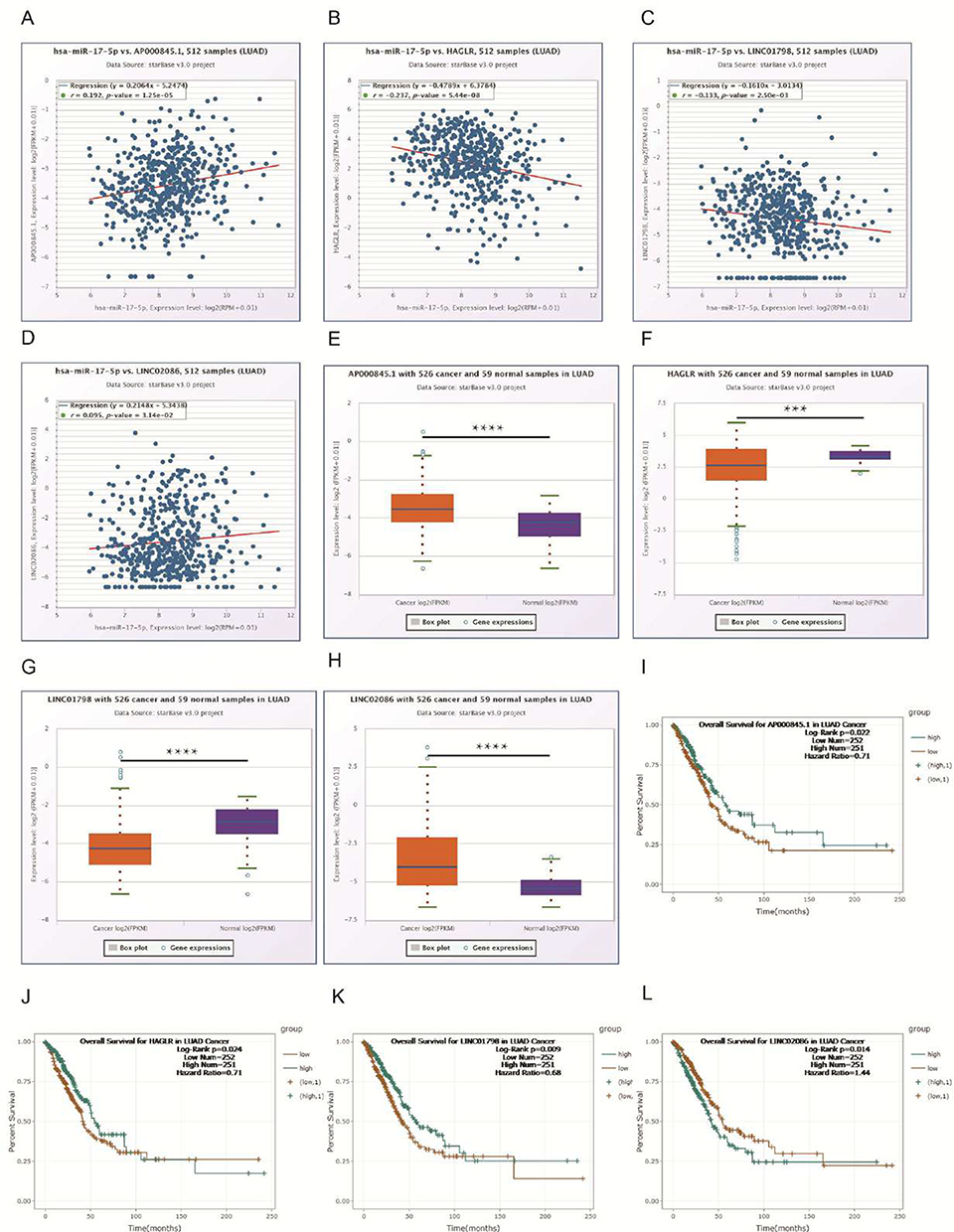 |
Figure 5 Expression and survival analyses for upstream long noncoding (lnc)RNAs of miR-17-5p in lung adenocarcinoma (LUAD). Notes: (A–D) Correlation expression analyses between miR-17-5p and predicted lncRNAs. (E–H) Expression of each candidate lncRNA in LUAD and normal controls. (I–L) Survival analyses of candidate lncRNAs in LUAD determined using starBase. ***p < 0.001, ****p < 0.0001. |
PFKFB3 Positively Correlates with Immune Cell Infiltration in LUAD
As a key regulator of glycolysis, PFKFB3 plays an important role in oncogenesis and TME. As shown in Figure 6A, compared with the normal level, the arm-level gain appeared in the B cells, CD4+ T cells, and dendritic cells. At the tumor-infiltration level in the macrophage cell, deep deletion and high amplification were obtained at the same time. There were no significant changes in immune cell infiltration level in the CD8+ T cells and neutrophils. Next, we investigated the correlation between PFKFB3 and each immune cell. It was obvious that PFKFB3 had a positive relationship with B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells. Among these, PFKFB3 had the most significant correlation with dendritic cells in LUAD (Figure 6B–G).
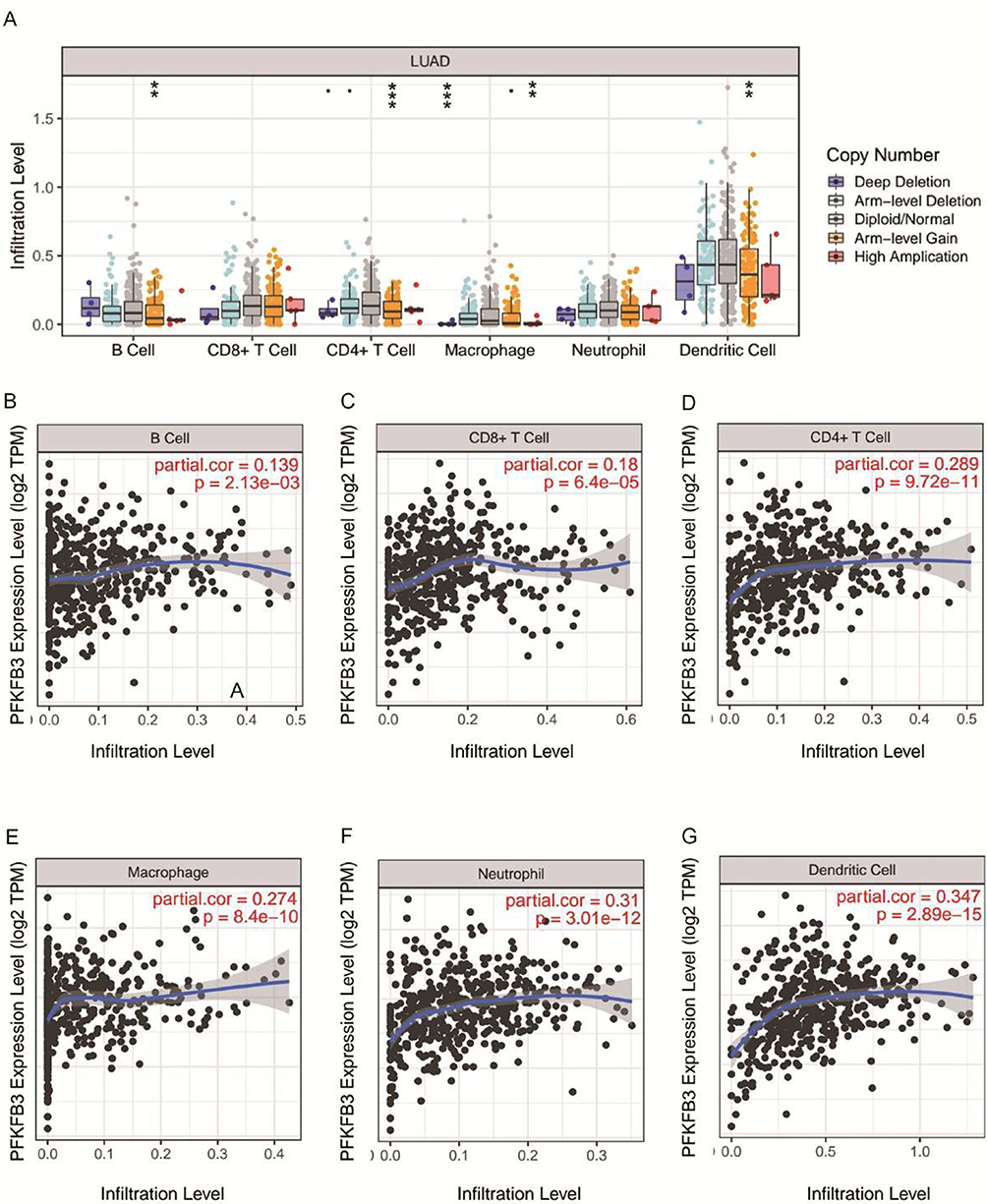 |
Figure 6 Relationship of immune cell infiltration with 6-phosphofructo-2-kinase (PFKFB3) levels in LUAD. Notes: (A) Infiltration level of various immune cells under different copy numbers of PFKFB3 in LUAD. (B–G) Correlation of PFKFB3 expression level with B cells (B), CD8+ T cells (C), CD4+ T cells (D), macrophages (E), neutrophils (F) or dendritic cells (G) infiltration levels in LUAD. **p <0.01, ***p < 0.001. |
Expression Correlation of PFKFB3 and Biomarkers of Immune Cells in LUAD
To explore the role of PFKFB3 in tumor immunity, we verified the expression correlation of PFKFB3 with biomarkers of immune cells in LUAD using the Gene Expression Profiling Interactive Analysis (GEPIA) database (http://gepia.cancer-pku.cn/). As listed in Table 1, PFKFB3 had a positive correlation with the biomarker of CD4+ T cells (CD4), M1 macrophage cells (NOS2, IRF5, PTGS2), M2 macrophage cells (CD163, MS4A4A), neutrophil cells (CEACAM8), and dendritic cells (CDC1, NRP1, ITGAX), respectively; however, the expression of PFKFB3 was negatively correlated with that of B cells (CD19, CD79A). Furthermore, there was no correlation between the levels of PFKFB3 and CD8+ T cells. To further confirm the findings above, we analyzed the correlation between PFKFB3 expression and the other cells mentioned above in the TIMER database (https://cistrome.shinyapps.io/timer/) (Table 2). To some extent, the results were roughly consistent with findings above except that the correlation between PFKFB3 and B cell was positive and PFKFB3 had a positive connection with CD8A.
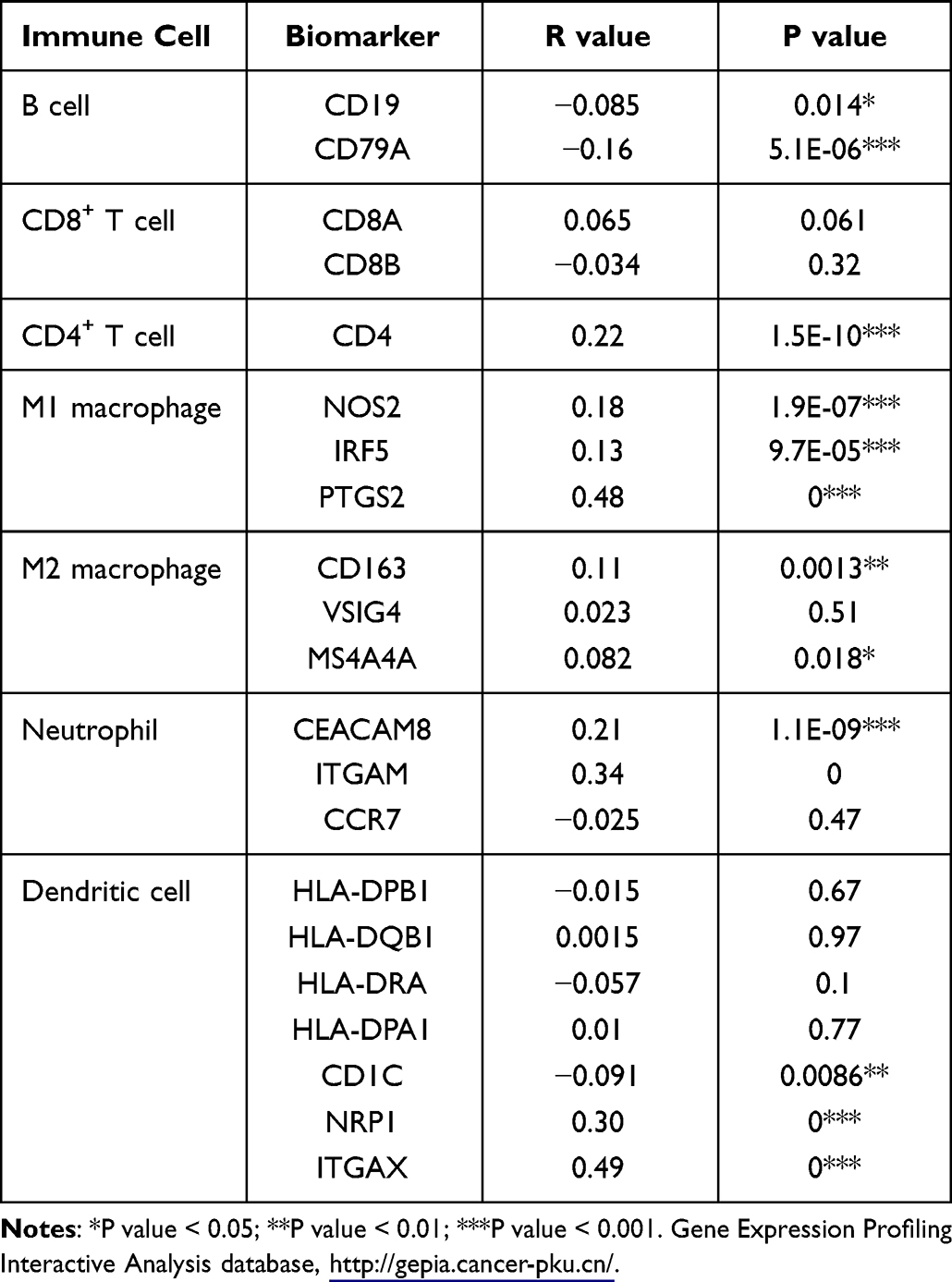 |
Table 1 Correlation Analysis Between 6-Phosphofructo-2-Kinase (PFKFB3) and Biomarkers of Immune Cells in Lung Adenocarcinoma (LUAD) Determined Using the Gene Expression Profiling Interactive Analysis Database (GEPIA) |
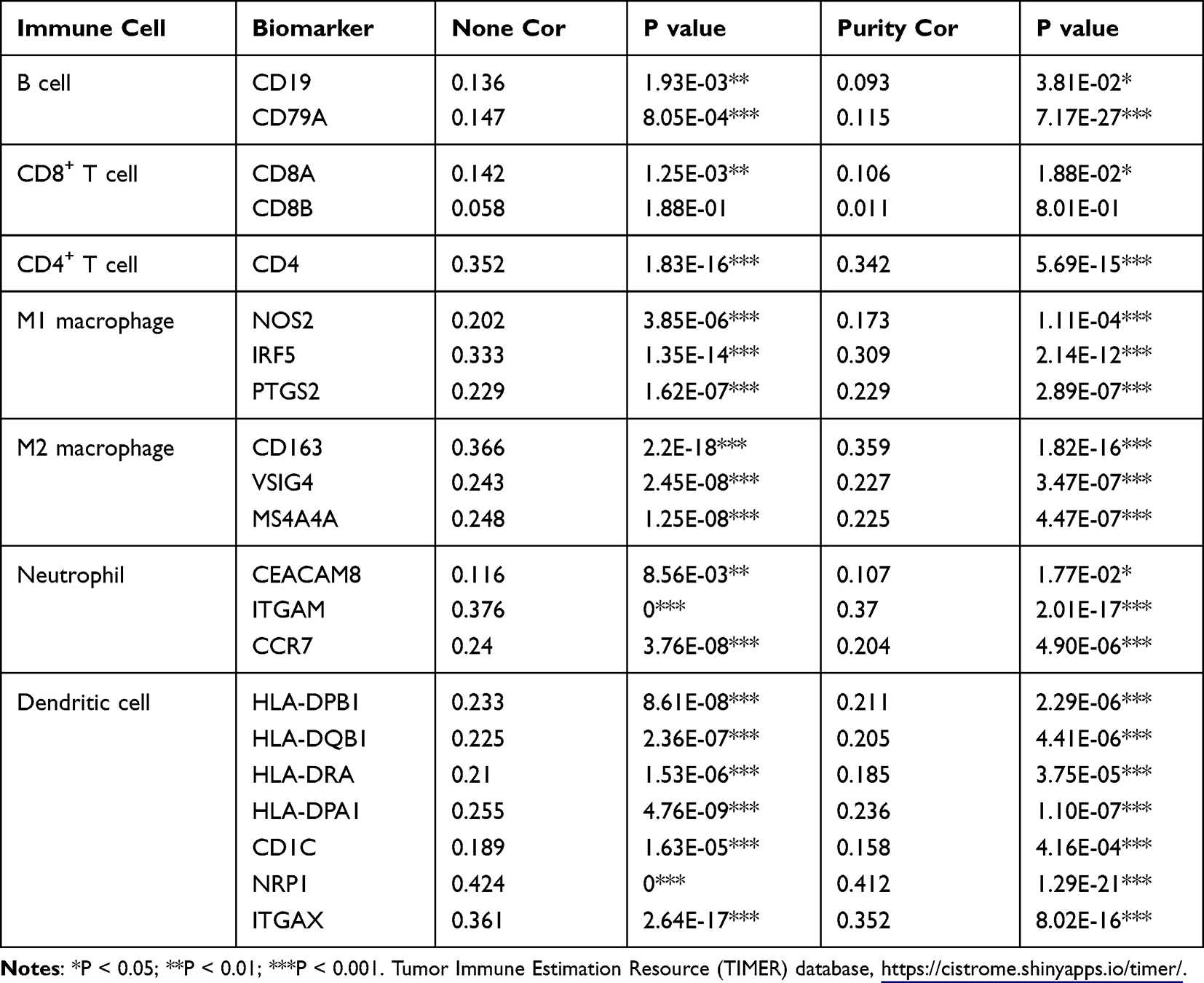 |
Table 2 Correlation Analysis Between 6-Phosphofructo-2-Kinase (PFKFB3) and Biomarkers of Immune Cells in Lung Adenocarcinoma (LUAD) Determined Using Tumor Immune Estimation Resource (TIMER) Database |
Relationship Between PFKFB3 and Immune Checkpoints in LUAD
It is well known that PD1/PD-L1 and CTLA-4 are very important immune checkpoints that could be used by tumors to avoid immunity and be able to evade immune surveillance.20 Because PFKFB3 was considered to have a potential oncogenic role in LUAD, the relationship between PFKFB3 and PD1, PD-L1, or CTLA-4 was assessed. The expression of PFKFB3 that was positively correlated with PD1, PD-L1, and CTLA-4 was observed (Figure 7A–C). Similar results were shown in GEPIA (Figure 7D–F); therefore, it could be concluded that tumor immune escape might play a significant role in PFKFB3-mediated carcinogenesis of LUAD.
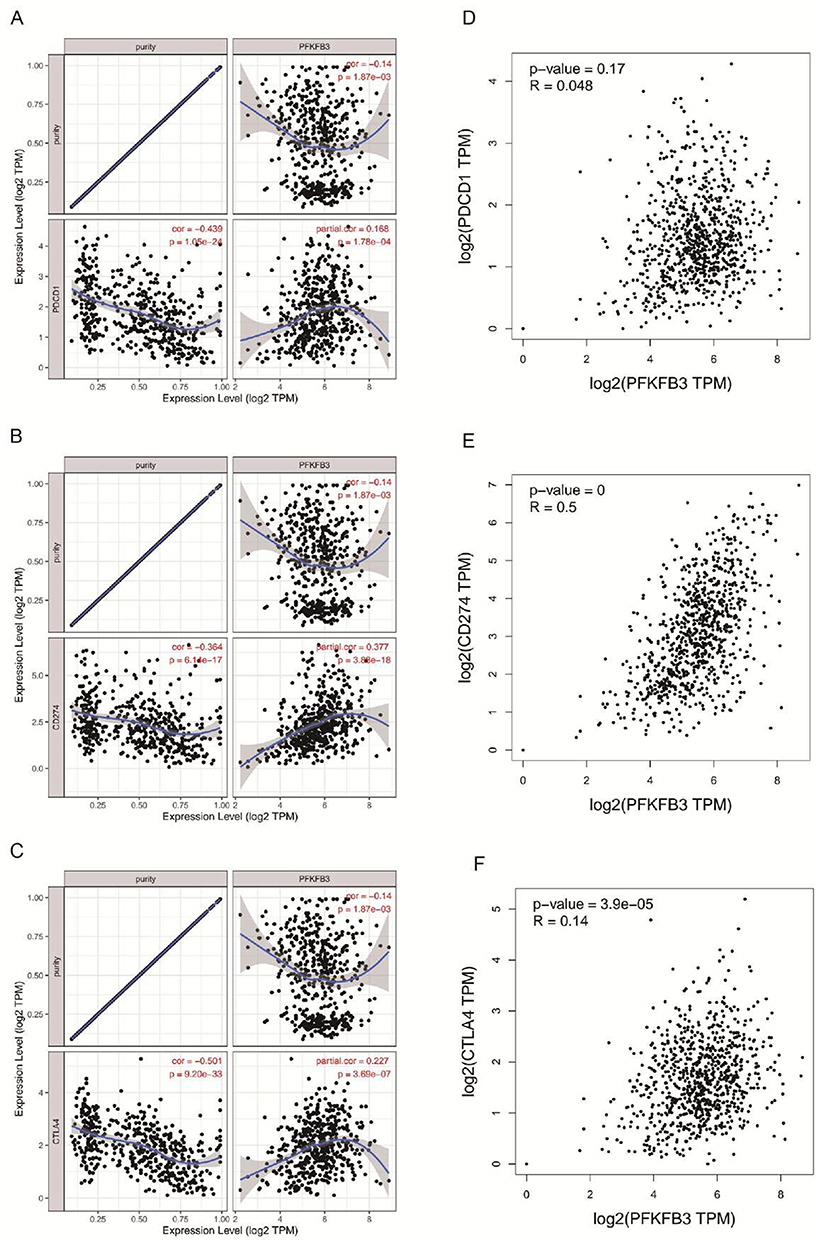 |
Figure 7 Correlation of 6-phosphofructo-2-kinase (PFKFB3) expression with PD-1, PD-L1 and CTLA-4 expression in lung adenocarcinoma (LUAD). Notes: (A) Spearman correlation of PFKFB3 with expression of PD-1 in LUAD adjusted by purity using the Tumor Immune Estimation Resource (TIMER) database (https://cistrome.shinyapps.io/timer/). (B) Spearman correlation of PFKFB3 with expression of PD-L1 in LUAD adjusted by purity using TIMER. (C) Spearman correlation of PFKFB3 with expression of CTLA-4 in LUAD adjusted by purity using TIMER. (D) The expression correlation of PFKFB3 with PD-1 in LUAD determined using the Gene Expression Profiling Interactive Analysis (GEPIA) database (http://gepia.cancer-pku.cn/). (E) The expression correlation of PFKFB3 with PD-L1 in LUAD determined by GEPIA database. (F) Expression correlation of PFKFB3 with CTLA-4 in LUAD determined by GEPIA database. |
Discussion
As we know, the prognosis of patients with LUAD is still not satisfactory because of the efficacy of treatment and whether there is brain metastasis. Understanding its mechanism of occurrence and development may provide new methods and ideas for the treatment of LUAD; therefore, seeking new molecular prognostic markers is essential. There is growing evidence demonstrating that PFKFB3 plays an important role in promoting tumor progression in different kinds of human cancers,21 such as ESCC,22 LUAD. In addition, in our previous study, we have found that PFKFB3 is an oncogene in LUAD tumor progression and concluded that targeting PFKFB3 may become a promising therapy,23 however, fewer samples were collected, and the clinical value was not analyzed.
In the present study, we conducted the mRNA expression of PFKFB3 in LUAD from the TIMER database (https://cistrome.shinyapps.io/timer/) and validated by TCGA LUAD (https://portal.gdc.cancer.gov/) data in the UALCAN database (http://ualcan.path.uab.edu/). Interestingly, the outcomes obtained from several databases were different from the studies we found before. Considering the count and purity of lung samples, we presume that this may be one of the reasons for the difference; therefore, the sample size should be increased for verification in the future. Next, we explored the protein level of PFKFB3 in LUAD from the UALCAN database (http://ualcan.path.uab.edu/) and conducted Western blotting and immunohistochemistry (IHC) analyses for further validation. We proved that the mRNA expression level of PFKFB3 was definitely downregulated in LUAD and concluded that the mRNA and protein expression of PFKFB3 were contrary. This situation may be explained from the following aspects. First, deubiquitination has been reported to be an important post-translational modification and plays a vital role in tumor progression and multiple metabolic pathways.24,25 Deubiquitination is catalyzed by deubiquitinating enzymes (DUBs) which could remove ubiquitin from ubiquitinated proteins and rescue proteins from degradation. 25 Second, RNA methylation modification may become another possibility. For example, m1A methylation modification has been found to increase translation rates.26 Moreover, m6A methylation modification could also promote translation efficiency by binding to a m6A site.27 In terms of analysis related to clinical characteristics, the expression of PFKFB3 generally increased with tumor stage and nodal metastasis of LUAD. Even though related research has demonstrated that rho-associated coiled-coil-containing protein kinase 2 could stabilize PFKFB3 expression and promote osteosarcoma growth and metastasis,28 how PFKFB3 regulates tumor progression through certain signaling pathways remains unclear.
It has been acknowledged that ncRNA contains miRNA and lncRNA. They have been reported to function by regulating the expression of angiogenesis-related factors, endothelial cell proliferation, cell migration, and tube formation.29 The competitive endogenous RNAs (ceRNA) hypothesis was first proposed in 2011 and demonstrated that ceRNAs could regulate the expression of downstream mRNA by combining shared miRNAs and may become an ignored factor resulting in diseases.30 MiRNA is a class of noncoding single-stranded RNA molecules with a length of approximately 22 nucleotides encoded by endogenous genes. They are involved in post-transcriptional gene expression regulation in animals and plants.31 To explore the possible upstream molecule of PFKFB3, we introduced three programs—miRDB, Targetscan, and miRWalk databases. At the end, seven miRNAs were obtained. Most miRNAs have been reported to promote the progression of human cancer. For example, miR-17-5p promoted gastric cancer progression by targeting runt-related transcription factor 3,32 miR-106a-5p could promote 5-FU resistance and the metastasis of CRC,33 and miR-133b could inhibit proliferation, invasion, and migration of LUAD cells by targeting CDCA8.34 After conducting expression and survival analyses, only miR-17-5p proved to be significant; therefore, we chose it as the most potential upstream molecule that could bind to PFKFB3. Currently, miR-17-5p was reported to play a role in the progression of hepatocellular carcinoma,35 gastric cancer32 and others.
PFKFB3 is a key enzyme in the process of glycolysis, and its production of fructose-2,6-bisphosphatase (F26BP) promotes the glycolysis mechanism by activating PFK-1.36 The role of PFKFB3 was different from other biomarkers such as GLUT and p16.37 Even though studies on upstream PFKFB3 have been reported in other cancers,38–40 there are few reports on how the miR-17-5p/PFKFB3 axis regulates lung tumor progression; therefore, we predicted the upstream lncRNAs of miR-17-5p and selected 216 lncRNAs for further investigation. By analyzing the correlation, expression, and survival analyses, we found that LINC01798, LINC02086, AP000845.1, and HAGLR were the most promising lncRNAs. According to the latest research, the LINc02086/miR-770-5p/SLC26A2 axis promoted migration in laryngeal squamous cell carcinoma,41 and HAGLR could promote the development of CRC by the miR-185-5p/CDK4/CDK6 axis and exacerbate the progression of hepatocellular carcinoma by negatively regulating miR-6785-5p.42,43 Taken together, LINC01798, LINC02086, AP000845.1, and the HAGLR/miR-17-5p/PFKFB3 axis may be the potential regulatory pathways in LUAD.
Increasing evidence has suggested that TME is a vital component in promoting tumor progression.44,45 In our research, the expression of PFKFB3 was significantly positively correlated with most immune cells, including B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells in LUAD. In addition, PFKFB3 had a positive correlation with several immune cell biomarkers. These findings indicated that tumor immune infiltration might play an important role in PFKFB3-mediated tumorigenesis.
Several studies have demonstrated that immunotherapy, such as PD-1/PD-L1 immune checkpoint inhibitors, have had some success in solid tumors, but in lung cancer, especially in advanced LUAD, the prognosis and survival benefit are still poor.46,47 Here, we also assessed the relationship between PFKFB3 and immune checkpoints, including PD-1, PD-L1 and CTLA-4. It was obvious that the expression of PFKFB3 was positively correlated with immune checkpoints, indicating that targeting PFKFB3 might become a direction of immunotherapy, thereby improving the effectiveness of immunotherapy. According to the latest analysis of the neoadjuvant therapy for lung cancer, compared with patients receiving single-agent immunotherapy, patients treated with neoadjuvant immunotherapy presented significant advantages in the respect of pathological response. Moreover the safety and validity could be better than traditional therapy.48
Conclusion
Overall, we demonstrated that the mRNA of PFKFB3 was upregulated in LUAD while its protein level was higher in LUAD than in adjacent normal tissues. We identified possible upstream regulatory mechanisms of PFKFB3 in LUAD, the LINC01798/LINC02086/AP000845.1/HAGLR-miR-17-5p-PFKFB3 axis (Figure 8). Furthermore, our research illustrated that PFKFB3 might exert its oncogenic roles by increasing tumor immune cell infiltration and immune checkpoint expression; however, much more basic studies conducted and clinical data must be compiled in the future.
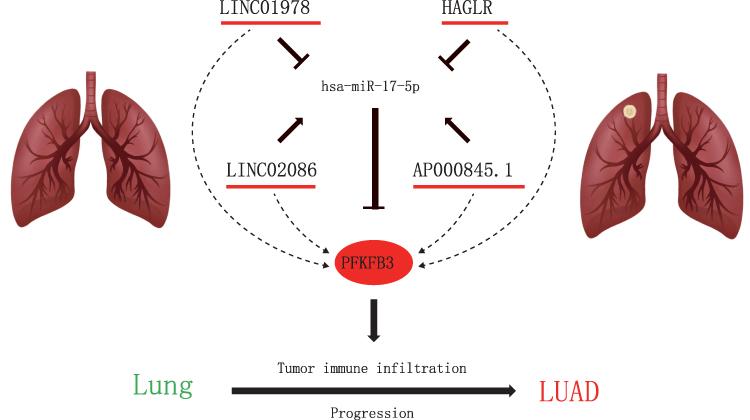 |
Figure 8 Model of LINC01978/LINC2086/AP000845.1/HAGLR-miR-17-5p-PFKFB3 axis in carcinogenesis of LUAD. |
Data Sharing Statement
The databases presented in this study were obtained from online repositories. All the data could be retrieved from following databases: TCGA (https://portal.gdc.cancer.gov/), TIMER (https://cistrome.shinyapps.io/timer/), UALCAN (http://ualcan.path.uab.edu/), GEPIA (http://gepia.cancer-pku.cn/), LinkedOmics (http://www.linkedomics.org/), miRWalk (http://mirwalk.umm.uni-heidelberg.de/), Targetscan (http://www.targetscan.org/), MiRDB (http://www.mirdb.org/) and Starbase (https://starbase.sysu.edu.cn/).
Ethics Approval and Consent to Participate
All patients consented to the use of their biological material for research in written form. The Ethics Committee of Affiliated Hospital of Nantong University approved our study.
Acknowledgments
We thank Taoming Mo for his contributions to the revision of the manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approve of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by grants from National Natural Science Foundation of China (No. 82273422) Jiangsu Graduate Research and Practice Innovation Program (SJCX21_1473, SJCX22_1639), Basic Science Research Project in Nantong City, Jiangsu, China (No.JC2021029), Nantong Municipal Science and Technology Project (No. JCZ19103), 2022 Nantong Basic Science Research Plan-Young Science and Technology Talents Innovation Special Project (No. JC12022016), 2022 Nantong City science and Technology Guidance Plan (No. JCZ2022016).
Disclosure
The authors declare that there are no competing interests.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. House CM. 中华医学会肺癌临床诊疗指南(2019版) [Chinese Medical Association guidelines for clinical diagnosis and treatment of lung cancer (2019 edi tion)]. Zhonghua Zhong Liu Za Zhi. 2020;42(4):257–287. Chinese. doi:10.3760/cma.j.cn112152-20200120-00049
3. Shi L, Pan H, Liu Z, Xie J, Han W. Roles of PFKFB3 in cancer. Signal Transduct Target Ther. 2017;2:17044. doi:10.1038/sigtrans.2017.44
4. O’Neal J, Clem A, Reynolds L, et al. Inhibition of 6-phosphofructo-2-kinase (PFKFB3) suppresses glucose metabolism and the growth of HER2+ breast cancer. Breast Cancer Res Treat. 2016;160(1):29–40. doi:10.1007/s10549-016-3968-8
5. Han J, Meng Q, Xi Q, Wang H, Wu G. PFKFB3 was overexpressed in gastric cancer patients and promoted the proliferation and migration of g astric cancer cells. Cancer Biomark. 2017;18(3):249–256. doi:10.3233/CBM-160143
6. Han J, Meng Q, Xi Q, et al. Interleukin-6 stimulates aerobic glycolysis by regulating PFKFB3 at early stage of colorectal cancer. Int J Oncol. 2016;48(1):215–224. doi:10.3892/ijo.2015.3225
7. Gu M, Li L, Zhang Z, et al. PFKFB3 promotes proliferation, migration and angiogenesis in nasopharyngeal carcinoma. J Cancer. 2017;8(18):3887–3896. doi:10.7150/jca.19112
8. Chandrashekar DS, Bashel B, Balasubramanya SAH, et al. UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017;19(8):649–658. doi:10.1016/j.neo.2017.05.002
9. Vasaikar SV, Straub P, Wang J, Zhang B. LinkedOmics: analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018;46(D1):D956–d963. doi:10.1093/nar/gkx1090
10. Dweep H, Gretz N, Sticht C. miRWalk database for miRNA-target interactions. Methods Mol Biol. 2014;1182:289–305.
11. Dweep H, Sticht C, Pandey P, Gretz N. miRWalk--database: prediction of possible miRNA binding sites by “walking” the genes of three genomes. J Biomed Inform. 2011;44(5):839–847. doi:10.1016/j.jbi.2011.05.002
12. Sticht C, De La Torre C, Parveen A, Gretz N. miRWalk: an online resource for prediction of microRNA binding sites. PLoS One. 2018;13(10):e0206239. doi:10.1371/journal.pone.0206239
13. Chen Y, Wang X. miRDB: an online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020;48(D1):D127–d131. doi:10.1093/nar/gkz757
14. Wang X. miRDB: a microRNA target prediction and functional annotation database with a wiki interface. RNA. 2008;14(6):1012–1017. doi:10.1261/rna.965408
15. Wong N, Wang X. miRDB: an online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015;43(Database issue):D146–52. doi:10.1093/nar/gku1104
16. Yang JH, Li JH, Shao P, Zhou H, Chen YQ, Qu LH. StarBase: a database for exploring microRNA-mRNA interaction maps from Argonaute CLIP-Seq and Degrado me-Seq data. Nucleic Acids Res. 2011;39(Database issue):D202–9. doi:10.1093/nar/gkq1056
17. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–w102. doi:10.1093/nar/gkx247
18. Huang Z, Zhou JK, Peng Y, He W, Huang C. The role of long noncoding RNAs in hepatocellular carcinoma. Mol Cancer. 2020;19(1):77. doi:10.1186/s12943-020-01188-4
19. Xie CR, Wang F, Zhang S, et al. Long noncoding RNA HCAL facilitates the growth and metastasis of hepatocellular carcinoma by acting a s a ceRNA of LAPTM4B. Mol Ther Nucleic Acids. 2017;9:440–451. doi:10.1016/j.omtn.2017.10.018
20. Shi L, Yang Y, Li M, et al. LncRNA IFITM4P promotes immune escape by up-regulating PD-L1 via dual mechanism in oral carcinogenesi s. Mol Ther. 2022;30:1564–1577. doi:10.1016/j.ymthe.2022.01.003
21. Lu L, Chen Y, Zhu Y. The molecular basis of targeting PFKFB3 as a therapeutic strategy against cancer. Oncotarget. 2017;8(37):62793–62802. doi:10.18632/oncotarget.19513
22. Bao J, Wu Y, Wang L, Zhu Y. The role of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-3 in esophageal squamous cell carcin oma. Medicine. 2020;99(15):e19626. doi:10.1097/MD.0000000000019626
23. Li X, Liu J, Qian L, et al. Expression of PFKFB3 and Ki67 in lung adenocarcinomas and targeting PFKFB3 as a therapeutic strategy. Mol Cell Biochem. 2018;445(1–2):123–134. doi:10.1007/s11010-017-3258-8
24. Antao AM, Tyagi A, Kim KS, Ramakrishna S. Advances in deubiquitinating enzyme inhibition and applications in cancer therapeutics. Cancers. 2020;12(6):1579. doi:10.3390/cancers12061579
25. Sun T, Liu Z, Yang Q. The role of ubiquitination and deubiquitination in cancer metabolism. Mol Cancer. 2020;19(1):146. doi:10.1186/s12943-020-01262-x
26. Oerum S, Dégut C, Barraud P, Tisné C. m1A post-transcriptional modification in tRNAs. Biomolecules. 2017;7(1):20. doi:10.3390/biom7010020
27. Zhao BS, Roundtree IA, He C. Post-transcriptional gene regulation by mRNA modifications. Nat Rev Mol Cell Biol. 2017;18(1):31–42.
28. Deng X, Yi X, Deng J, et al. ROCK2 promotes osteosarcoma growth and metastasis by modifying PFKFB3 ubiquitination and degradation. Exp Cell Res. 2019;385(2):111689. doi:10.1016/j.yexcr.2019.111689
29. Zhao Z, Sun W, Guo Z, Zhang J, Yu H, Liu B. Mechanisms of lncRNA/microRNA interactions in angiogenesis. Life Sci. 2020;254:116900. doi:10.1016/j.lfs.2019.116900
30. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
31. Liu SH, Hsu KW, Lai YL, et al. Systematic identification of clinically relevant miRNAs for potential miRNA-based therapy in lung ade nocarcinoma. Mol Ther Nucleic Acids. 2021;25:1–10. doi:10.1016/j.omtn.2021.04.020
32. Song J, Liu Y, Wang T, Li B, Zhang S. MiR-17-5p promotes cellular proliferation and invasiveness by targeting RUNX3 in gastric cancer. Biomed Pharmacother. 2020;128:110246. doi:10.1016/j.biopha.2020.110246
33. Liu J, Huang Y, Wang H, Wu D. MiR-106a-5p promotes 5-FU resistance and the metastasis of colorectal cancer by targeting TGF¦ÂR2. Int J Clin Exp Pathol. 2018;11(12):5622–5634.
34. Hu C, Wu J, Wang L, et al. miR-133b inhibits cell proliferation, migration, and invasion of lung adenocarcinoma by targeting CDC A8. Pathol Res Pract. 2021;223:153459. doi:10.1016/j.prp.2021.153459
35. Yang F, Yin Y, Wang F, et al. miR-17-5p Promotes migration of human hepatocellular carcinoma cells through the p38 mitogen-activate d protein kinase-heat shock protein 27 pathway. Hepatology. 2010;51(5):1614–1623. doi:10.1002/hep.23566
36. Imbert-Fernandez Y, Clem BF, O’Neal J, et al. Estradiol stimulates glucose metabolism via 6-phosphofructo-2-kinase (PFKFB3). J Biol Chem. 2014;289(13):9440–9448. doi:10.1074/jbc.M113.529990
37. Pezzuto A, D’Ascanio M, Ricci A, Pagliuca A, Carico E. Expression and role of p16 and GLUT1 in malignant diseases and lung cancer: a review. Thorac Cancer. 2020;11(11):3060–3070. doi:10.1111/1759-7714.13651
38. Ge X, Lyu P, Cao Z, et al. Overexpression of miR-206 suppresses glycolysis, proliferation and migration in breast cancer cells v ia PFKFB3 targeting. Biochem Biophys Res Commun. 2015;463(4):1115–1121. doi:10.1016/j.bbrc.2015.06.068
39. Wang J, Li X, Xiao Z, et al. MicroRNA-488 inhibits proliferation and glycolysis in human prostate cancer cells by regulating PFKFB 3. FEBS Open Bio. 2019;9(10):1798–1807. doi:10.1002/2211-5463.12718
40. Zheng WD, Zhou FL, Lin N. MicroRNA-26b inhibits osteosarcoma cell migration and invasion by down-regulating PFKFB3 expression. Genet Mol Res. 2015;14(4):16872–16879. doi:10.4238/2015.December.14.14
41. Shi Y, Yang D, Qin Y. Identifying prognostic lncRNAs based on a ceRNA regulatory network in laryngeal squamous cell carcino ma. BMC Cancer. 2021;21(1):705. doi:10.1186/s12885-021-08422-2
42. Li YH, Huang GM, Wang W, Zang HL. LncRNA HAGLR exacerbates hepatocellular carcinoma through negatively regulating miR-6785-5p. Eur Rev Med Pharmacol Sci. 2020;24(18):9353–9360. doi:10.26355/eurrev_202009_23018
43. Sun W, Nie W, Wang Z, Zhang H, Li Y, Fang X. Lnc HAGLR promotes colon cancer progression through sponging miR-185-5p and activating CDK4 and CDK6 in vitro and in vivo. Onco Targets Ther. 2020;13:5913–5925. doi:10.2147/OTT.S246092
44. Huang J, Li J, Zheng S, et al. Tumor microenvironment characterization identifies two lung adenocarcinoma subtypes with specific imm une and metabolic state. Cancer Sci. 2020;111(6):1876–1886. doi:10.1111/cas.14390
45. Sadeghi Rad H, Monkman J, Warkiani ME, et al. Understanding the tumor microenvironment for effective immunotherapy. Med Res Rev. 2021;41(3):1474–1498. doi:10.1002/med.21765
46. Jiang L, Huang J, Jiang S, et al. The surgical perspective in neoadjuvant immunotherapy for resectable non-small cell lung cancer. Cancer Immunol Immunother. 2021;70(8):2313–2321. doi:10.1007/s00262-021-02847-1
47. Steven A, Fisher SA, Robinson BW. Immunotherapy for lung cancer. Respirology. 2016;21(5):821–833. doi:10.1111/resp.12789
48. Jiang J, Wang Y, Gao Y, et al. Neoadjuvant immunotherapy or chemoimmunotherapy in non-small cell lung cancer: a systematic review an d meta-analysis. Transl Lung Cancer Res. 2022;11(2):277–294. doi:10.21037/tlcr-22-75
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.