Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 15
New Discovered Adipokines Associated with the Pathogenesis of Obesity and Type 2 Diabetes
Received 25 May 2022
Accepted for publication 26 July 2022
Published 7 August 2022 Volume 2022:15 Pages 2381—2389
DOI https://doi.org/10.2147/DMSO.S376163
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Jia-Xue Cheng, Ke Yu
Center for Endocrine Metabolism and Immune Diseases, Beijing Luhe Hospital, Capital Medical University, Beijing, People’s Republic of China
Correspondence: Ke Yu, Center for Endocrine Metabolism and Immune Diseases, Beijing Luhe Hospital, Capital Medical University, No. 82, Xinhua South Road, Tongzhou District, Beijing, People’s Republic of China, Tel +86 13811657618, Email [email protected]
Abstract: Obesity is defined as abnormal or excessive accumulation of adipose tissue, closely associated with the increased risk of various comorbidities, especially type 2 diabetes mellitus (T2DM). Adipose tissue is a complex structure responsible for not only fat storage but also releasing adipokines which may play roles in the pathogenesis and could be developed into biomarkers for diagnosis, treatment and prognosis of obesity-related metabolic diseases. This review aims to summarize several adipokines discovered recently that have promising functions in obesity and T2DM. Among them, the levels of FSTL1, WISP1 and Asprosin in subjects with obesity or diabetes are commonly higher than in normal controls, suggesting that they may be pathogenic. Inversely, SFRP5, Metrnl, NRG4 and FAM19A5 may serve as the protective factors.
Keywords: adipokines, obesity, diabetes
Introduction
Obesity has become a global epidemic, which is spiraling out of control. With the prevalence of obesity, obesity-related problems such as T2DM, are also rapidly increasing. To discover the pathophysiological mechanisms between obesity and T2DM becomes particularly important for preventing and alleviating obesity related diabetes. Studies have found that adipose tissue can not only store fat, but also be an endocrine organ,1 secreting a variety of bioactive factors, collectively known as adipokines. As early as 1987, adipose tissue was identified as a main site for metabolism of sex steroids2 and production of adipsin,3 an endocrine factor that is markedly down-regulated in rodent obesity. The subsequent identification and characterization of leptin4 in 1994 firmly established adipose tissue as an endocrine organ. Adipokines secreted by adipose tissue have been identified that either promote inflammatory responses or contribute to the resolution of inflammation. An imbalance of pro- and anti-inflammatory adipokines leads to obesity-linked metabolic dysfunction.5 Restoring the balance would be the method for treating obesity and its complications. Our review focuses on describing a few newly discovered adipokines in chronological order of being identified as an “adipokine”. Their functions in metabolism have been proved in past studies, but some are disputable or the specific action pathway is still unclear. A summary of researches progress so far seems necessary to provide the basis for further exploration on their potentials as biomarkers for diagnosis, treatment, and prognosis in obesity and T2DM (Table 1).
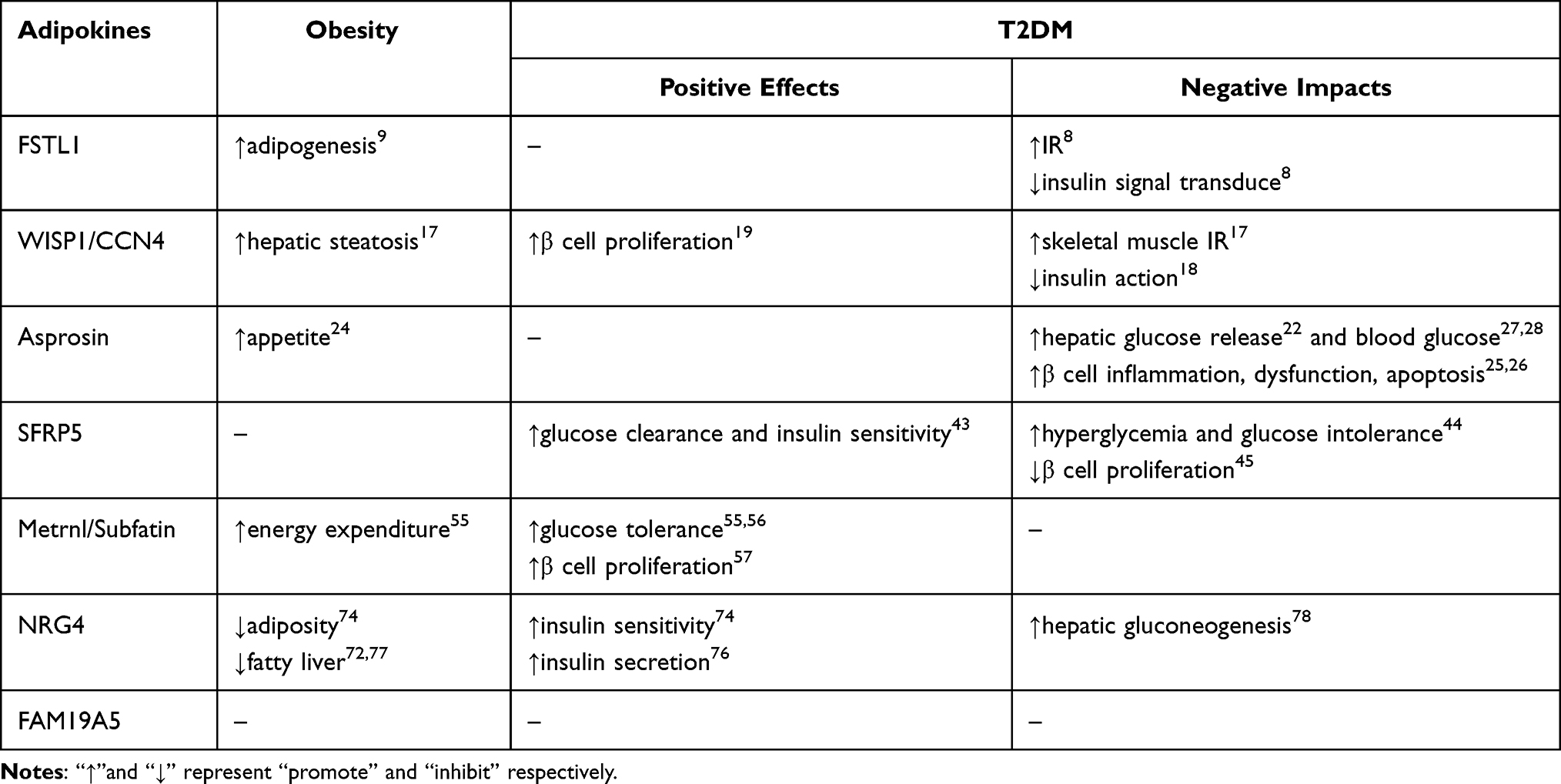 |
Table 1 Newly Discovered Adipokines’ Effect on the Pathogenesis of Obesity and T2DM |
Pro-Inflammatory Adipokines
The upregulated adipokines in the state of obesity and insulin resistance (IR) generally have pro-inflammatory effects, leading to the form of a chronic inflammatory state and contributing to metabolic dysfunction.
FSTL1 Promotes Adipogenesis, Inflammation and IR
Preclinical Studies
Follistatin like 1 (FSTL1) was originally cloned from an osteoblast cell line as transforming growth factor-β (TGF-β) stimulated clone 36 (TSC-36) in 1993.6 Later, it was reported as a pro-inflammatory molecule in 2006, which led to severe paw swelling and arthritis in mouse paws.7 FSTL1 expression was increased in adipose tissues of obese mice.8 During the switch from chow diet to high fat diet (HFD), FSTL1 deletion mice gained less body weight, fat mass, and glucose level than the control group. FSTL1 promoted adipogenesis by inhibiting the conversion of PPARγ to p-PPARγ through the integrin/FAK/ERK signaling pathway,9 and could activate NFκB and JNK signaling pathways, critical in obesity-induced inflammation and IR, in adipocytes and macrophages.8 Insulin-stimulated phosphorylation of both Akt and IRS-1 was markedly reduced by FSTL1 treatment, which impaired insulin signal transduction in 3T3-L1 adipocytes.8 Further, FSTL1 expression in adipose tissue10 and circulation11 rose dramatically in response to acute physical activity in rodents.
Clinical Studies
Serum levels of FSTL1 were significantly higher in patients with overweight/obese8,10 or newly diagnosed T2DM10 than in control subjects. Furthermore, a positive correlation between FSTL1 levels and body mass index (BMI), waist-to-hip ratio (WHR), fasting blood glucose (FBG), 2-hour post–glucose load blood glucose (2h-BG), glycated hemoglobin (HbA1c), triglyceride (TG), total cholesterol (TC), and HOMA-IR was observed. On the other hand, morbid and super obesity were potentially associated with a decline in plasma FSTL1 levels.12 In the intervention study, acute physical activity was found to significantly increase the circulating FSTL1 concentration in young, healthy participants.10,13
Together, FSTL1 is a potential mediator of adipogenesis, inflammation and IR. Prospective cohort studies are warranted to gain more evidence of the causality between FSTL1 and metabolic disorders. In addition, adenovirus mediated overexpression of FSTL1 or blocking its actions through neutralizing antibodies in animals will directly elucidate the role of FSTL1 in the pathogenesis.
WISP1 /CCN4 Boosts IR and Islet β Cell Proliferation
Preclinical Studies
Wingless-type inducible signaling pathway protein 1 (WISP1, also known as CCN4), a target gene of the canonical Wnt signaling pathway and a member of the CCN family of extracellular matrix proteins,14,15 has been confirmed as a pro‐inflammatory adipokine in 2015.16 HFD-fed mice demonstrated upregulated WISP1 expression in epididymal adipose tissue.16 Knockdown of WISP1 in HFD-fed mice significantly attenuated hepatic steatosis and skeletal muscle IR via reversing inflammation-associated JNK phosphorylation. Treatment with WISP1 significantly increased lipogenesis-associated gene expression and TG accumulation in hepatocytes and suppressed insulin signaling in C2C12 skeletal muscle cells, which was abrogated after NFκB-, JNK-, and TLR4-knockdown.17 In vitro studies on primary human skeletal muscle cells (hSkMCs) and murine AML12 hepatocytes also showed that recombinant WISP1 directly impaired insulin action by inhibiting the Akt signaling pathway.18 Meanwhile, WISP1 promoted endogenous and transplanted adult mice pancreatic β cell proliferation depending on Akt signaling, exhibiting potential therapeutic use to prevent or delay the appearance of diabetes.19
Clinical Studies
WISP1 mRNA expression was elevated in visceral adipose tissue (VAT) rather than in subcutaneous adipose tissue (SAT) in human subjects, correlated positively with fasting insulin and negatively with insulin sensitivity. Reduction of WISP1 mRNA expression in SAT was observed after the weight loss with a low-calorie diet.16 Both mRNA expression in VAT and serum levels of WISP1 were increased in obese men.18 Patients with T2DM had higher levels of circulating WISP1, associated with central abdominal fat mass.20 A conflicting study reported no difference in WISP1 concentrations between individuals with normal glucose tolerance (NGT) and with T2DM but revealing the positive correlations between circulating WISP1 with BMI, body fat percentage, TG, hip circumference and fatty liver index.21
More prospective clinical studies would be valuable to establish the causal relationship of WISP1 on obesity and T2DM. Further cell or molecule studies are needed to precisely determine the role of this promising adipokine in the pathogenesis of diseases.
Asprosin Elevates Blood Glucose and Impairs β Cells
Preclinical Studies
Asprosin, the C-terminal cleavage product of profibrillin, was found as a novel adipokine in patients with Neonatal Progeroid Syndrome (NPS) in 2016.22 It performs two critical fasting-related functions (hepatic glucose production and appetite stimulation) using the same cAMP second messenger system, although using different spatiotemporal mechanisms at two distinct organs. Upon secretion by white adipose, Asprosin travels to the liver, stimulating the release of glucose by binding to the OLFR734 receptor.22,23 It also crosses the blood-brain barrier to hypothalamus, where it stimulates appetite by activating orexigenic AgRP neurons and inhibiting anorexigenic POMC neurons.24 Animal experiments demonstrated that Asprosin could induce islet β cell inflammation, dysfunction and apoptosis through TLR4/JNK-mediated signaling25 and promote β cell apoptosis by inhibiting the autophagy of β‐cell via AMPK‐mTOR pathway.26 The administration of Asprosin increased blood glucose level in healthy mice while there was no change in diabetic ones.27 On the other hand, intraperitoneal injection of Asprosin-specific monoclonal antibody could drop plasma Asprosin levels, lower appetite and body weight and reduce blood glucose in mouse models.22,28 Asprosin-neutralizing antibody is a kind of dual-effect pharmacologic therapy that targets at both overnutrition and hyperglycemia.
Clinical Studies
Decreased levels of plasma Asprosin have been observed in NPS patients associated with reduced appetite and extreme leanness22,24 and in anorexia patients,29 whereas pathological increase in circulating Asprosin is related to obesity30,31 and T2DM.32–34 Asprosin is also expressed in human placenta and elevated in the plasma of pregnant women with gestational diabetes (GDM) and their offspring.35 The postprandial Asprosin level is apparently lower than the fasting in individuals with NGT, but not in T2DM patients. In another word, the alteration of meal-related circadian oscillation of Asprosin may be affected by T2DM.36,37 Clinical trials have explored the influences of diet, exercise, drugs and surgery on circulating Asprosin. A trial showed that rapid coffee consumption led to lower energy, fat intake and circulatory Asprosin. Rapid caffeine metabolizers were more likely to benefit from the consumption of more than two cups of coffee per day (≥15cpw) by reducing their BMI.38 An 8-week Nordic Walking training at maximal fat oxidation intensity decreased the concentration of Asprosin in the blood as well as visceral obesity in young women with metabolic disorders.39 Blood samples of 10 men and 10 women who performed a single 20-s bicycle sprint were collected before exercise, in the 3s, 15s, 30s, and 60s of recovery, and 24h after competition. Whereas the single anaerobic effort induced an increase in Asprosin secretion only in women.40 Metformin or SGLT2 inhibitors treatment could lower circulating Asprosin levels in patients with newly diagnosed T2DM.41,42 Blood Asprosin levels decreased significantly 6 months after bariatric surgery, and Asprosin concentrations before bariatric surgery were associated with the weight reduction magnitude.31
Despite advances in the understanding of Asprosin’s function, the reproducibility of some data produced in this field is waiting for proof. The secretion and action mechanism and the regulating factors are unclear.
Anti-Inflammatory Adipokines
In addition to the pro-inflammatory adipokines described above, adipose tissues also secrete a number of anti-inflammatory factors, which have shown beneficial effects on adiposity and insulin action.
SFRP5ʹs Effect on Moderating IR and Blood Glucose is Controversial
Preclinical Studies
Secreted frizzled-related protein 5 (SFRP5) is an anti-inflammatory adipokine discovered in 2010.43 Its expression was reduced in white adipose tissue of obese HFD mice. SFRP5-deficient mice fed with HFD exhibited elevated F4/80 and CD68, macrophage-mediated inflammation markers, in epididymal adipose tissue and impaired glucose clearance and insulin sensitivity compared with the wildtype mice, which was significantly improved after two weeks’ intravenous injection of SFRP5. In vitro, upregulation expression of SFRP5 in 3T3-L1 adipocytes prevented the inflammatory and insulin-resistant state by binding with Wnt5a and neutralizing JNK activation in macrophages and adipocytes via paracrine and autocrine mechanisms.43 However, a report provided contradictory findings that elevated hyperglycemia and glucose intolerance was observed by overexpressing SFRP5 in obese, prediabetic mice. Conversely, anti-SFRP5 monoclonal antibody (mAb) therapy improved these phenotypes in vivo.44 In addition, SFRP5 was downregulated in pancreatic islets from obese rodents and humans, correlated with activated canonical Wnt signaling, promoting proliferation in primary islet cells and in the β cell line INS1E. Its expression in β cells could be positively modulated by IGF binding protein 3 (IGFBP3) secreted from visceral adipose tissue.45
Clinical Studies
In human studies, individuals with obesity,46–48 T2DM,47–50 metabolic syndrome (Mets),51 or Polycystic ovary syndrome (PCOS)47 generally exhibited lower SFRP5 levels in blood than normal controls. Circulating SFRP5 levels were positively associated with insulin sensitivity, high density lipoprotein cholesterol (HDL-C) and adiponectin, but negatively with BMI, WHR, HbA1c, FBG, 2h-BG and HOMA-IR. Another study showed that increasing concentrations of SFRP5 were independently and significantly associated with T2DM.52 After treated with metformin53 for 3 months, serum SFRP5 of PCOS patients significantly increased than that before administration. 16 weeks’ treatment with liraglutide47 rose plasma SFRP5 levels and reduced HOMA-IR and BMI moderately, suggesting increases in insulin secretion and sensitivity and decreases in weight.
SFRP5 is an adipokine which acts as an inhibitor of Wnt signaling pathway. It has been suggested to exert anti-inflammatory and insulin-sensitizing effects, however, contradictory data has also been reported. Prospective studies will improve our understanding of its functions in metabolism. Further exploration of the biological mechanisms may pave the way for SFRP5 to serve as a potential novel treatment option for obesity and T2DM.
Metrnl/Subfatin Improves Glucose Tolerance and β Cell Proliferation
Preclinical Studies
Meteorin-Like (Metrnl/Subfatin) was identified as a novel adipokine in 2014, dramatically expressed in subcutaneous fat of both rodents and humans.54 It can be induced in muscle after exercise and adipose tissue upon cold exposure, and is present in the blood. Increasing circulating levels of Metrnl stimulated energy expenditure and the gene expression associated with anti-inflammatory cytokines and improved glucose tolerance in obese/diabetic mice.55 An intraperitoneal injection of recombinant Metrnl improved glucose tolerance in mice with HFD-induced obesity or T2DM via a Ca2+-CAMKK2-AMPK-HDAC5-GLUT4-p38-TBC1D1 signaling pathway.56 Metrnl could also ameliorate β cell function by inhibiting apoptosis and promoting proliferation of it through activating the Wnt/β-catenin pathway in T2DM mice.57 Global Metrnl knockout increased blood TG by 14% and decreased TC by 16% and HDL-C by 24%, reflecting Metrnl ‘s beneficial aspect on the regulation of lipid metabolism.58
Clinical Studies
The clinical evidences regarding its circulating levels in obesity and T2DM are conflicting. Some studies showed less circulatory Metrnl levels in obese or T2DM patients compared with the control group,59–62 correlated with higher FBG, 2h-BG, fasting insulin, HOMA-IR, HbA1c, high-sensitive C-reactive protein (hs-CRP), interleukin-6 (IL-6), and tumor necrosis factor-a (TNF-a). Others demonstrated either an increase63–65 or no significant change.66,67 Clinical trials have revealed that the weight loss via low calorie diet (LCD), combined training (CT) or bariatric surgery (BS) could rise Metrnl levels, in correlation to the improvement in glucose and lipid homeostasis. In LCD and BS patients, serum Metrnl concentrations significantly increased after 3 months, but returned to baseline after 12 months.68 Besides, another study reported remarkably enhanced circulatory Metrnl levels 12 months after BS.69 CT for 16 weeks increased brown adipose tissue (BAT) thermogenic activity as well as serum Metrnl levels.70 Metformin treatment did not increase the serum Metrnl levels after 12 weeks.60
Although Metrnl has shown emerging effects in obesity, T2DM and dyslipidemia, there are conflicts in the clinical results. Besides, a myriad of work still needs to be done to explore its structure-function relationship and regulatory mechanism with various signaling pathways in related diseases.
NRG4 Ameliorates IR, but Augments Hepatic Gluconeogenesis
Preclinical Studies
Neuregulin-4 (NRG4), a member of the ErbB ligand family, was firstly described in 1999.71 Then it was identified as a previously unknown BAT-enriched secreted factor attenuating hepatic lipogenic signaling and preserving glucose and lipid homeostasis in obesity in 2014.72 NRG4-deficient mice upon HFD gained more body weight, higher plasma TG concentrations, pronounced hepatic steatosis and exacerbated glucose intolerance and insulin resistance compared with controls.72 Similarly, mice with ErbB4 deletion developed into Mets when fed with a medium-fat diet (MFD).73 On the contrary, transgenic expression of NRG4 resulted in the prevention of HFD-induced adiposity and fatty liver, and the improvement of insulin sensitivity.72,74,75 In vitro experiments, NRG4 gave a pronounced effect on insulin secretion in the rat insulinoma cell line.76 The promotion of adipocyte browning by n-3 polyunsaturated fatty acids was accompanied by an elevation of NRG4 expression via the PPARG pathway. NRG4 directly prevented lipid accumulation in HepG2 cells.77 Collectively, these findings provided the evidence in support of the potential health benefits of NRG4 in managing obesity and obesity-associated diseases. On the other hand, NRG4 knockdown in liver attenuated hepatic gluconeogenesis via suppressing PEPCK, G6Pase and PGC-1α expression in diabetic mice, reminding that NRG4 specific-silencing in liver will provide a potential therapeutic strategy for T2DM.78
Clinical Studies
Several observational studies have showed that circulating NRG4 concentrations were inversely associated with the risk of obesity,79 T2DM72,80 or Mets.81,82 Subjects with lower NRG4 levels had higher FBG, fasting insulin, HOMA-IR, HbA1c, TC, TG, and hs-CRP. Controversially, other researchers found that serum NRG4 level was elevated in T2DM.83–85 Serum NRG4 increased significantly in response to a 3-week ω-3 polyunsaturated fatty acids dietary protocol.86 Plasma levels of NRG4 were improved in three training protocols: high-intensity interval training (HIIT), circuit resistance training (CRT), and moderate intensity continuous training (MICT) compared with the control group. What is more, the increase was greater in HIIT and CRT compared with the MICT.87 After 24 weeks, metformin therapy resulted in a significant increase of NRG4 levels compared with the baseline and the placebo group.88
The phenotype of mice with either a gain or loss of NRG4 function suggest that reduced NRG4 may be causally linked to obesity-related impaired glucose metabolism. Prospective cohort studies are warranted to gain more evidence in humans. In addition, the exact mechanisms of how NRG4 exerts these beneficial effects are not entirely clear.
FAM19A5ʹs Impact on T2DM is Unclear
Preclinical Studies
In 2004, a small secretory protein family was found in humans and mice, which is composed of five highly homologous genes, called TAFA1-5. TAFA mRNAs are highly expressed in specific brain regions, but rarely in other tissues.89 In 2018, researchers found that family with sequence similarity 19 member A5 (FAM19A5/TAFA5) was a new type of protective factor, highly expressed in human and mouse adipose tissue. It significantly inhibited the proliferation and migration of vascular smooth muscle cells and the proliferation of carotid intima after balloon injury via the binding receptor S1PR2 in mice. HFD could induce the downregulation of FAM19A5 expression in adipose tissue.90
Clinical Studies
Plasma FAM19A5 in patients with nonalcoholic fatty liver was significantly lower than that in the control group, and there was a significant negative correlation between FAM19A5 and BMI, visceral fat, alanine aminotransferase, aspartate aminotransferase, liver hardness and carotid intima-media thickness.91 The latest research showed that serum FAM19A5 levels were apparently decreased in the obese children compared with healthy controls. Negative correlations were detected between FAM19A5 and BMI as well as FBG and fasting insulin.92 Inversely, it was found that serum FAM19A5 concentrations in T2DM patients were higher than that in non-diabetic subjects, and positively correlated with WC, WHR, FBG and HbA1c.93
What we can see is that FAM19A5 is so novel that there are few researches about it. Whether and how FAM19A5 participates in obesity, IR and T2DM are waiting for our attention in the future.
Conclusion
Taken together, adipokines act as the promising candidates which have been shown to possess properties of mediating glycolipid metabolism. Generally, overexpression of pro-inflammatory adipokines or lack of anti-inflammatory adipokines in rodent experiments are causally linked to the occurrence and development of obesity and T2DM. The pro-inflammatory adipokines increase whereas the anti-inflammatory adipokines decrease in obese rodents and humans, associated with corresponding metabolic indicators of adiposity and T2DM. It is worth discussing that, as previously reviewed,94 adipokines, such as, IFN-γ and IL-10, can be raised in T2D patients, but not generally favor pro-inflammation. The impaired Th1/Th2 ratios were implicated with a delicate balance existing within diverse metabolic conditions. Consistent with this view, our review provides controversial human studies about the four anti-inflammatory adipokines, proving that their circulating levels are abnormally elevated in patients with T2DM. Based on this point, maybe we should pay more attention to the relationship between the balance of multiple adipokines and metabolic diseases in the future, rather than just focusing on one factor. There is still a lot waiting to be explored. For example, some contradictory data need to be corroborated in large sample sizes. Clinical cohort studies to demonstrate the causal relationship between adipokines and metabolic diseases are required to carry out. Few preclinical studies about the pathophysiological molecular mechanisms by which adipokines act have been conducted. There will be a long way to go before adipokines can be put into clinical trials and applied in humans.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab. 2004;89(6):2548–2556. doi:10.1210/jc.2004-0395
2. Siiteri PK. Adipose tissue as a source of hormones. Am J Clin Nutr. 1987;45(1 Suppl):277–282.
3. Flier JS, Cook KS, Usher P, Spiegelman BM. Severely impaired adipsin expression in genetic and acquired obesity. Science. 1987;237(4813):405–408.
4. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372(6505):425–432.
5. Ouchi N, Parker JL, Lugus JJ, Walsh K. Adipokines in inflammation and metabolic disease. Nat Rev Immunol. 2011;11(2):85–97.
6. Shibanuma M, Mashimo J, Mita A, Kuroki T, Nose K. Cloning from a mouse osteoblastic cell line of a set of transforming-growth-factor-beta 1-regulated genes, one of which seems to encode a follistatin-related polypeptide. Eur J Biochemistry. 1993;217(1):13–19.
7. Miyamae T, Marinov AD, Sowders D, et al. Follistatin-like protein-1 is a novel proinflammatory molecule. J Immunol. 2006;177(7):4758–4762.
8. Fan N, Sun H, Wang Y, et al. Follistatin-like 1: a potential mediator of inflammation in obesity. Mediators Inflamm. 2013;2013:752519.
9. Fang D, Shi X, Jia X, et al. Ups and downs: the PPARγ/p-PPARγ seesaw of follistatin-like 1 and integrin receptor signaling in adipogenesis. Mol Metab. 2022;55:101400.
10. Xu X, Zhang T, Mokou M, et al. Follistatin-like 1 as a Novel Adipomyokine Related to Insulin Resistance and Physical Activity. J Clin Endocrinol Metab. 2020;105(12):87.
11. Kon M, Tanimura Y, Yoshizato H. Effects of acute endurance exercise on follistatin-like 1 and apelin in the circulation and metabolic organs in rats. Arch Physiol Biochem. 2020;1:1–5.
12. Horak M, Kuruczova D, Zlamal F, Tomandl J, Bienertova-Vasku J. Follistatin-Like 1 Is Downregulated in Morbidly and Super Obese Central-European Population. Dis Markers. 2018;2018:4140815.
13. Kon M, Ebi Y, Nakagaki K. Effects of acute sprint interval exercise on follistatin-like 1 and apelin secretions. Arch Physiol Biochem. 2021;127(3):223–227.
14. Katoh M, Katoh M. WNT signaling pathway and stem cell signaling network. Clin Cancer Res. 2007;13(14):4042–4045.
15. Yeger H, Perbal B. The CCN family of genes: a perspective on CCN biology and therapeutic potential. J Cell Commun Signal. 2007;1(3–4):159–164.
16. Murahovschi V, Pivovarova O, Ilkavets I, et al. WISP1 is a novel adipokine linked to inflammation in obesity. Diabetes. 2015;64(3):856–866.
17. Jung TW, Kang C, Goh J, et al. WISP1 promotes non-alcoholic fatty liver disease and skeletal muscle insulin resistance via TLR4/JNK signaling. J Cell Physiol. 2018;233(8):6077–6087.
18. Hörbelt T, Tacke C, Markova M, et al. The novel adipokine WISP1 associates with insulin resistance and impairs insulin action in human myotubes and mouse hepatocytes. Diabetologia. 2018;61(9):2054–2065.
19. Fernandez-Ruiz R, García-Alamán A, Esteban Y, et al. Wisp1 is a circulating factor that stimulates proliferation of adult mouse and human beta cells. Nat Commun. 2020;11(1):5982.
20. Klimontov VV, Bulumbaeva DM, Fazullina ON, et al. Circulating Wnt1-inducible signaling pathway protein-1 (WISP-1/CCN4) is a novel biomarker of adiposity in subjects with type 2 diabetes. J Cell Commun Signal. 2020;14(1):101–109.
21. Tacke C, Aleksandrova K, Rehfeldt M, et al. Assessment of circulating Wnt1 inducible signalling pathway protein 1 (WISP-1)/CCN4 as a novel biomarker of obesity. J Cell Commun Signal. 2018;12(3):539–548.
22. Romere C, Duerrschmid C, Bournat J, et al. Asprosin, a Fasting-Induced Glucogenic Protein Hormone. Cell. 2016;165(3):566–579.
23. Li E, Shan H, Chen L, et al. OLFR734 Mediates Glucose Metabolism as a Receptor of Asprosin. Cell Metab. 2019;30(2):319–328.e318.
24. Duerrschmid C, He Y, Wang C, et al. Asprosin is a centrally acting orexigenic hormone. Nat Med. 2017;23(12):1444–1453.
25. Lee T, Yun S, Jeong JH, Jung TW. Asprosin impairs insulin secretion in response to glucose and viability through TLR4/JNK-mediated inflammation. Mol Cell Endocrinol. 2019;486:96–104.
26. Wang R, Hu W. Asprosin promotes β-cell apoptosis by inhibiting the autophagy of β-cell via AMPK-mTOR pathway. J Cell Physiol. 2021;236(1):215–221.
27. Hekim MG, Kelestemur MM, Bulmus FG, et al. Asprosin, a novel glucogenic adipokine: a potential therapeutic implication in diabetes mellitus. Arch Physiol Biochem. 2021;1–7.
28. Mishra I, Duerrschmid C, Ku Z, et al. Asprosin-neutralizing antibodies as a treatment for metabolic syndrome. eLife. 2021;10.
29. Du C, Wang C, Guan X, et al. Asprosin is associated with anorexia and body fat mass in cancer patients. Supportive Care Cancer. 2021;29(3):1369–1375.
30. Ugur K, Aydin S. Saliva and Blood Asprosin Hormone Concentration Associated with Obesity. Int J Endocrinol. 2019;2019:2521096.
31. Wang C-Y, Lin T-A, Liu K-H, et al. Serum asprosin levels and bariatric surgery outcomes in obese adults. Int j Obesity. 2019;43(5):1019–1025.
32. Naiemian S, Naeemipour M, Zarei M, et al. Serum concentration of asprosin in new-onset type 2 diabetes. Diabetol Metab Syndr. 2020;12:65.
33. Zhang L, Chen C, Zhou N, Fu Y, Cheng X. Circulating asprosin concentrations are increased in type 2 diabetes mellitus and independently associated with fasting glucose and triglyceride. Clin Chim Acta. 2019;489:183–188.
34. Wang Y, Qu H, Xiong X, et al. Plasma Asprosin Concentrations Are Increased in Individuals with Glucose Dysregulation and Correlated with Insulin Resistance and First-Phase Insulin Secretion. Mediators Inflamm. 2018;2018:9471583.
35. Zhong L, Long Y, Wang S, et al. Continuous elevation of plasma asprosin in pregnant women complicated with gestational diabetes mellitus: a nested case-control study. Placenta. 2020;93:17–22.
36. Zhang X, Jiang H, Ma X, Wu H. Increased serum level and impaired response to glucose fluctuation of asprosin is associated with type 2 diabetes mellitus. J Diabetes Investig. 2020;11(2):349–355.
37. Corica D, Pepe G, Aversa T, et al. Meal-Related Asprosin Serum Levels Are Affected by Insulin Resistance and Impaired Fasting Glucose in Children With Obesity. Front Endocrinol (Lausanne). 2021;12:805700.
38. Gkouskou KG, Georgiopoulos G, Vlastos I, et al. CYP1A2 polymorphisms modify the association of habitual coffee consumption with appetite, macronutrient intake, and body mass index: results from an observational cohort and a cross-over randomized study. Int j Obesity. 2022;46(1):162–168.
39. Kantorowicz M, Szymura J, Szygula Z, Kusmierczyk J, Maciejczyk M, Wiecek M. Nordic Walking at Maximal Fat Oxidation Intensity Decreases Circulating Asprosin and Visceral Obesity in Women With Metabolic Disorders. Front Physiol. 2021;12:726783.
40. Wiecek M, Szymura J, Maciejczyk M, Kantorowicz M, Szygula Z. Acute Anaerobic Exercise Affects the Secretion of Asprosin, Irisin, and Other Cytokines - A Comparison Between Sexes. Front Physiol. 2018;9:1782.
41. Gozel N, Kilinc F. Investigation of plasma asprosin and saliva levels in newly diagnosed type 2 diabetes mellitus patients treated with metformin. Endokrynol Pol. 2021;72(1):37–43.
42. Jiang A, Feng Z, Yuan L, Zhang Y, Li Q, She Y. Effect of sodium-glucose co-transporter-2 inhibitors on the levels of serum asprosin in patients with newly diagnosed type 2 diabetes mellitus. Diabetol Metab Syndr. 2021;13(1):34.
43. Ouchi N, Higuchi A, Ohashi K, et al. Sfrp5 is an anti-inflammatory adipokine that modulates metabolic dysfunction in obesity. Science. 2010;329(5990):454–457.
44. Rulifson IC, Majeti JZ, Xiong Y, et al. Inhibition of secreted frizzled-related protein 5 improves glucose metabolism. Am J Physiol Endocrinol Metab. 2014;307(12):E1144–1152.
45. Rebuffat SA, Oliveira JM, Altirriba J, et al. Downregulation of Sfrp5 promotes beta cell proliferation during obesity in the rat. Diabetologia. 2013;56(11):2446–2455.
46. Rydzewska M, Nikołajuk A, Matulewicz N, Stefanowicz M, Karczewska-Kupczewska M. Serum secreted frizzled-related protein 5 in relation to insulin sensitivity and its regulation by insulin and free fatty acids. Endocrine. 2021;74(2):300–307.
47. Hu W, Li L, Yang M, et al. Circulating Sfrp5 is a signature of obesity-related metabolic disorders and is regulated by glucose and liraglutide in humans. J Clin Endocrinol Metab. 2013;98(1):290–298.
48. Hu Z, Deng H, Qu H. Plasma SFRP5 levels are decreased in Chinese subjects with obesity and type 2 diabetes and negatively correlated with parameters of insulin resistance. Diabetes Res Clin Pract. 2013;99(3):391–395.
49. Carstensen-Kirberg M, Kannenberg JM, Huth C, et al. Inverse associations between serum levels of secreted frizzled-related protein-5 (SFRP5) and multiple cardiometabolic risk factors: KORA F4 study. Cardiovasc Diabetol. 2017;16(1):109.
50. He X, Ma H. Correlation Between Circulating Levels of Secreted Frizzled-Related Protein 5 and Type 2 Diabetic Patients and Subjects with Impaired-Glucose Regulation. Diabetes Metabolic Syndrome Obesity. 2020;13:1243–1250.
51. Xu Q, Wang H, Li Y, et al. Plasma Sfrp5 levels correlate with determinants of the metabolic syndrome in Chinese adults. Diabetes Metab Res Rev. 2017;33(6):875.
52. Lu Y-C, Wang C-P, Hsu -C-C, et al. Circulating secreted frizzled-related protein 5 (Sfrp5) and wingless-type MMTV integration site family member 5a (Wnt5a) levels in patients with type 2 diabetes mellitus. Diabetes Metab Res Rev. 2013;29(7):551–556.
53. Zhang Y, Ran Y, Kong L, et al. Decreased SFRP5 correlated with excessive metabolic inflammation in polycystic ovary syndrome could be reversed by metformin: implication of its role in dysregulated metabolism. J Ovarian Res. 2021;14(1):97.
54. Li Z-Y, Zheng S-L, Wang P, et al. Subfatin is a novel adipokine and unlike Meteorin in adipose and brain expression. CNS Neurosci Ther. 2014;20(4):344–354.
55. Rao RR, Long JZ, White JP, et al. Meteorin-like is a hormone that regulates immune-adipose interactions to increase beige fat thermogenesis. Cell. 2014;157(6):1279–1291.
56. Lee JO, Byun WS, Kang MJ, et al. The myokine meteorin-like (metrnl) improves glucose tolerance in both skeletal muscle cells and mice by targeting AMPKα2. FEBS J. 2020;287(10):2087–2104.
57. Hu W, Wang R, Sun B. Meteorin-Like Ameliorates β Cell Function by Inhibiting β Cell Apoptosis of and Promoting β Cell Proliferation via Activating the WNT/β-Catenin Pathway. Front Pharmacol. 2021;12:627147.
58. Qi Q, Hu W-J, Zheng S-L, et al. Metrnl deficiency decreases blood HDL cholesterol and increases blood triglyceride. Acta Pharmacol Sin. 2020;41(12):1568–1575.
59. Dadmanesh M, Aghajani H, Fadaei R, Ghorban K. Lower serum levels of Meteorin-like/Subfatin in patients with coronary artery disease and type 2 diabetes mellitus are negatively associated with insulin resistance and inflammatory cytokines. PLoS One. 2018;13(9):e0204180.
60. Lee JH, Kang YE, Kim JM, et al. Serum Meteorin-like protein levels decreased in patients newly diagnosed with type 2 diabetes. Diabetes Res Clin Pract. 2018;135:7–10.
61. El-Ashmawy HM, Selim FO, Hosny TAM, Almassry HN. Association of low serum Meteorin like (Metrnl) concentrations with worsening of glucose tolerance, impaired endothelial function and atherosclerosis. Diabetes Res Clin Pract. 2019;150:57–63.
62. Fadaei R, Dadmanesh M, Moradi N, et al. Serum levels of subfatin in patients with type 2 diabetes mellitus and its association with vascular adhesion molecules. Arch Physiol Biochem. 2020;126(4):335–340.
63. Chung HS, Hwang SY, Choi JH, et al. Implications of circulating Meteorin-like (Metrnl) level in human subjects with type 2 diabetes. Diabetes Res Clin Pract. 2018;136:100–107.
64. AlKhairi I, Cherian P, Abu-Farha M, et al. Increased Expression of Meteorin-Like Hormone in Type 2 Diabetes and Obesity and Its Association with Irisin. Cells. 2019;8(10):325.
65. Cherian P, Al-Khairi I, Jamal M, et al. Association Between Factors Involved in Bone Remodeling (Osteoactivin and OPG) With Plasma Levels of Irisin and Meteorin-Like Protein in People With T2D and Obesity. Front Endocrinol (Lausanne). 2021;12:752892.
66. Wu Q, Dan Y-L, He Y-S, et al. Circulating Meteorin-like Levels in Patients with Type 2 Diabetes Mellitus: a Meta-Analysis. Curr Pharm Des. 2020;26(44):5732–5738.
67. Ferns GA, Fekri K, Abadi MSS, Dehkordi MB, Arjmand M-H. A meta-analysis of the relationship between serums metrnl-like protein/subfatin and risk of type 2 diabetes mellitus and coronary artery disease. Arch Physiol Biochem. 2021;1–7.
68. Schmid A, Karrasch T, Schäffler A. Meteorin-Like Protein (Metrnl) in Obesity, during Weight Loss and in Adipocyte Differentiation. J Clin Med. 2021;10(19):87.
69. Pellitero S, Piquer-Garcia I, Ferrer-Curriu G, et al. Opposite changes in meteorin-like and oncostatin m levels are associated with metabolic improvements after bariatric surgery. Int j Obesity. 2018;42(4):919–922.
70. Bonfante ILP, Monfort-Pires M, Duft RG, et al. Combined training increases thermogenic fat activity in patients with overweight and type 2 diabetes. Int j Obesity. 2022;1:56.
71. Harari D, Tzahar E, Romano J, et al. Neuregulin-4: a novel growth factor that acts through the ErbB-4 receptor tyrosine kinase. Oncogene. 1999;18(17):2681–2689.
72. Wang G-X, Zhao X-Y, Meng Z-X, et al. The brown fat-enriched secreted factor Nrg4 preserves metabolic homeostasis through attenuation of hepatic lipogenesis. Nat Med. 2014;20(12):1436–1443.
73. Zeng F, Wang Y, Kloepfer LA, Wang S, Harris RC. ErbB4 deletion predisposes to development of metabolic syndrome in mice. Am J Physiol Endocrinol Metab. 2018;315(4):E583–E593.
74. Ma Y, Gao M, Liu D. Preventing High Fat Diet-induced Obesity and Improving Insulin Sensitivity through Neuregulin 4 Gene Transfer. Sci Rep. 2016;6:26242.
75. Chen Z, Wang G-X, Ma SL, et al. Nrg4 promotes fuel oxidation and a healthy adipokine profile to ameliorate diet-induced metabolic disorders. Mol Metab. 2017;6(8):863–872.
76. South JCM, Blackburn E, Brown IR, Gullick WJ. The neuregulin system of ligands and their receptors in rat islets of Langerhans. Endocrinology. 2013;154(7):2385–2392.
77. Yang F, Zhou N, Zhu X, Min C, Zhou W, Li X. n-3 PUFAs protect against adiposity and fatty liver by promoting browning in postnatally overfed male rats: a role for NRG4. J Nutr Biochem. 2021;93:108628.
78. Zhang L, Bai M, Tang H, et al. Role of hepatic neuregulin 4 in the regulation of gluconeogenesis in mice. Life Sci. 2019;217:185–192.
79. Guo D, Liu J, Zhang P, et al. Adiposity Measurements and Metabolic Syndrome Are Linked Through Circulating Neuregulin 4 and Adipsin Levels in Obese Adults. Front Physiol. 2021;12:667330.
80. Zhang L, Fu Y, Zhou N, Cheng X, Chen C. Circulating neuregulin 4 concentrations in patients with newly diagnosed type 2 diabetes: a cross-sectional study. Endocrine. 2017;57(3):535–538.
81. Cai C, Lin M, Xu Y, Li X, Yang S, Zhang H. Association of circulating neuregulin 4 with metabolic syndrome in obese adults: a cross-sectional study. BMC Med. 2016;14(1):165.
82. Yan P, Xu Y, Wan Q, et al. Plasma Neuregulin 4 Levels Are Associated with Metabolic Syndrome in Patients Newly Diagnosed with Type 2 Diabetes Mellitus. Dis Markers. 2018;2018:6974191.
83. Kang YE, Kim JM, Choung S, et al. Comparison of serum Neuregulin 4 (Nrg4) levels in adults with newly diagnosed type 2 diabetes mellitus and controls without diabetes. Diabetes Res Clin Pract. 2016;117:1–3.
84. Chen -L-L, Peng -M-M, Zhang J-Y, et al. Elevated circulating Neuregulin4 level in patients with diabetes. Diabetes Metab Res Rev. 2017;33(4):87.
85. Kocak MZ, Aktas G, Erkus E, et al. Neuregulin-4 is associated with plasma glucose and increased risk of type 2 diabetes mellitus. Swiss Med Wkly. 2019;149:w20139.
86. Żebrowska A, Hall B, Stolecka-Warzecha A, Stanula A, Sadowska-Krępa E. The Effect of Omega-3 Fatty Acid Supplementation on Serum Adipocytokines, Lipid Profile and Biochemical Markers of Inflammation in Recreational Runners. Nutrients. 2021;13(2):87.
87. Saeidi A, Shishvan SR, Soltani M, et al. Differential Effects of Exercise Programs on Neuregulin 4, Body Composition and Cardiometabolic Risk Factors in Men With Obesity. Front Physiol. 2021;12:797574.
88. Elbarbary NS, Ismail EAR, Ghallab MA. Effect of metformin as an add-on therapy on neuregulin-4 levels and vascular-related complications in adolescents with type 1 diabetes: a randomized controlled trial. Diabetes Res Clin Pract. 2022;186:109857.
89. Tang YT, Emtage P, Funk WD, et al. TAFA: a novel secreted family with conserved cysteine residues and restricted expression in the brain. Genomics. 2004;83(4):727–734.
90. Wang Y, Chen D, Zhang Y. Novel Adipokine, FAM19A5, Inhibits Neointima Formation After Injury through Sphingosine-1-Phosphate Receptor 2. Circulation. 2018;138(1):48–63.
91. Yari FA, Shabani P, Karami S, Sarmadi N, Poustchi H, Bandegi AR. Circulating levels of FAM19A5 are inversely associated with subclinical atherosclerosis in non-alcoholic fatty liver disease. BMC Endocr Disord. 2021;21(1):153.
92. Xie K, Liu L, Yin C, et al. Follistatin-like 1 and family with sequence similarity to 19 member A5 levels are decreased in obese children and associated with glucose metabolism. Ann Nutr Metab. 2022.
93. Lee YB, Hwang HJ, Kim JA, et al. Association of serum FAM19A5 with metabolic and vascular risk factors in human subjects with or without type 2 diabetes. Diab Vasc Dis Res. 2019;16(6):530–538.
94. Mahlangu T, Dludla PV, Nyambuya TM, et al. A systematic review on the functional role of Th1/Th2 cytokines in type 2 diabetes and related metabolic complications. Cytokine. 2020;126:154892.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Understanding the Role of Metabolic Syndrome as a Risk Factor for Hepatocellular Carcinoma
Chavez-Tapia NC, Murúa-Beltrán Gall S, Ordoñez-Vázquez AL, Nuño-Lambarri N, Vidal-Cevallos P, Uribe M
Journal of Hepatocellular Carcinoma 2022, 9:583-593
Published Date: 5 July 2022
Variations of Ghrelin and Obestatin Hormones During the Menstrual Cycle of Women of Different BMIs
Salem AM, Latif R, Rafique N, Aldawlan MI, Almulla LB, Alghirash DY, Fallatah OA, Alotaibi FM, Aljabbari FH, Yar T
International Journal of Women's Health 2022, 14:1297-1305
Published Date: 7 September 2022
Associations of Obesity Indices with Bone Mineral Densities and Risk of Osteoporosis Stratified Across Diabetic Vascular Disease in T2DM Patients
Zheng S, Zhou J, Wang K, Wang X, Li Z, Chen N
Diabetes, Metabolic Syndrome and Obesity 2022, 15:3459-3468
Published Date: 3 November 2022
The Effectiveness of Indigenous Knowledge-Based Lifestyle Interventions in Preventing Obesity and Type 2 Diabetes Mellitus in Indigenous Children in Canada: A Systematic Review
Kshatriya M, Wang KW, Hildebrand J, Crawford R, Nadarajah A, Youssef M, Rivas A, Kaushal A, Banfield LE, Thabane L, Samaan MC
Adolescent Health, Medicine and Therapeutics 2023, 14:175-193
Published Date: 28 September 2023
Knowledge of Cardiovascular Disease Risk Factors and Its Primary Prevention Practices Among the Saudi Public – A Questionnaire-Based Cross-Sectional Study
Bashatah A, Syed W, Al-Rawi MBA
International Journal of General Medicine 2023, 16:4745-4756
Published Date: 20 October 2023