Back to Journals » Journal of Pain Research » Volume 11
Neuropathic pain modulation after spinal cord injury by breathing-controlled electrical stimulation (BreEStim) is associated with restoration of autonomic dysfunction
Authors Karri J ![]() , Li S, Zhang L, Chen YT, Stampas A
, Li S, Zhang L, Chen YT, Stampas A ![]() , Li S
, Li S
Received 17 May 2018
Accepted for publication 13 July 2018
Published 12 October 2018 Volume 2018:11 Pages 2331—2341
DOI https://doi.org/10.2147/JPR.S174475
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Überall
Jay Karri,1 Shengai Li,1 Larry Zhang,1 Yen-Ting Chen,1 Argyrios Stampas,1 Sheng Li1,2
1Department of Physical Medicine and Rehabilitation, McGovern Medical School, The University of Texas Health Science Center at Houston, Houston, TX, USA; 2TIRR Memorial Hermann Research Center, TIRR Memorial Hermann Hospital, Houston, TX, USA
Background: Recent findings have implicated supraspinal origins from the pain neuromatrix–central autonomic network (PNM–CAN) in the generation of neuropathic pain (NP) after spinal cord injury (SCI). The aim of this study was to further investigate the theorized PNM–CAN mechanisms in persons with SCI by using a centrally directed pain intervention, provided by breathing-controlled electrical stimulation (BreEStim), to measure resultant autonomic changes measured by time and frequency domain heart rate variability (HRV) analysis.
Methods: Null and active BreEStim interventions were administered to SCI+NP subjects (n=10) in a random order. HRV data and VAS pain scores were collected at resting pre-test and 30 minutes post-test time points. Resting HRV data were also collected from SCI–NP subjects (n=11).
Results: SCI+NP subjects demonstrated a lower baseline HRV and parasympathetic tone, via SD of the normal-to-normal intervals (SDNN) and low frequency (LF) parameters, compared with SCI–NP subjects. However, following active BreEStim, SCI+NP subjects exhibited an increase in HRV and parasympathetic tone, most notably via pairs of successive R–R beat lengths varying by greater than 50 ms (NN50) and proportion of NN50 for total number of beats (pNN50) parameters along with lower VAS scores. Additionally, the post-test SCI+NP group was found to have a statistically comparable autonomic profile to the SCI–NP group across all HRV variables, including SDNN and LF parameters.
Conclusion: The analgesic effects of active BreEStim in SCI+NP subjects were associated with restoration of autonomic dysfunction in this population.
Keywords: autonomic dysfunction, spinal cord injury, neuropathic pain, BreEStim, electrical stimulation, heart rate variability
Introduction
Neuropathic pain (NP) is a chronic pain condition that is often caused by damage to the somatosensory system, such as that observed with spinal cord injury (SCI).1 NP is prevalent in up to 80% of the persons with SCI, which contributes to suffering and a low quality of life.2 Managing NP in SCI subjects is clinically challenging, partly because the underlying mechanisms of this condition are multifactorial in nature and have yet to be clearly delineated.3,4 Rather, the precise pathophysiology of chronic NP after SCI is controversial with many differing theories in existence. Conventional thought based on historical findings suggests that NP originates at the level of the SCI.3,4 Moreover, there exists increasing evidence suggesting that supraspinal centers may have a vital role in NP pathogenesis.5–8 Furthermore, numerous recent studies have also demonstrated associations between these supraspinal centers and the central autonomic network (CAN) in the context of NP generation after SCI.8–11 However, these proposed autonomic correlates to NP after SCI have been less investigated.
Following a landmark series of studies, Melzack et al12–14 identified a pain neuromatrix (PNM) composed of numerous cortical and subcortical structures responsible for mediating the multidimensional nature of pain. Functional MRI (fMRI) investigations have supported these findings by demonstrating increased activity diffusely across the PNM structures of healthy controls after induction of painful stimuli.5,6 These structures are varied and extensive; the most implicated areas include the insular cortex, anterior cingulate cortex, posterior parietal cortex, prefrontal cortex, and thalamus.5,6 Interestingly, similar fMRI approaches found that pain induction in persons with chronic SCI increased the activity in certain PNM regions to a greater extent as in healthy controls.8 These SCI-specific regions of increased activity included regions of the insular cortex, anterior cingulate cortex, and prefrontal cortex.8 In particular, these pain-specific PNM regions in persons with chronic SCI were highly associated and shared much overlap with the CAN.8 Their relations are schematically represented in Figure 1.5–11
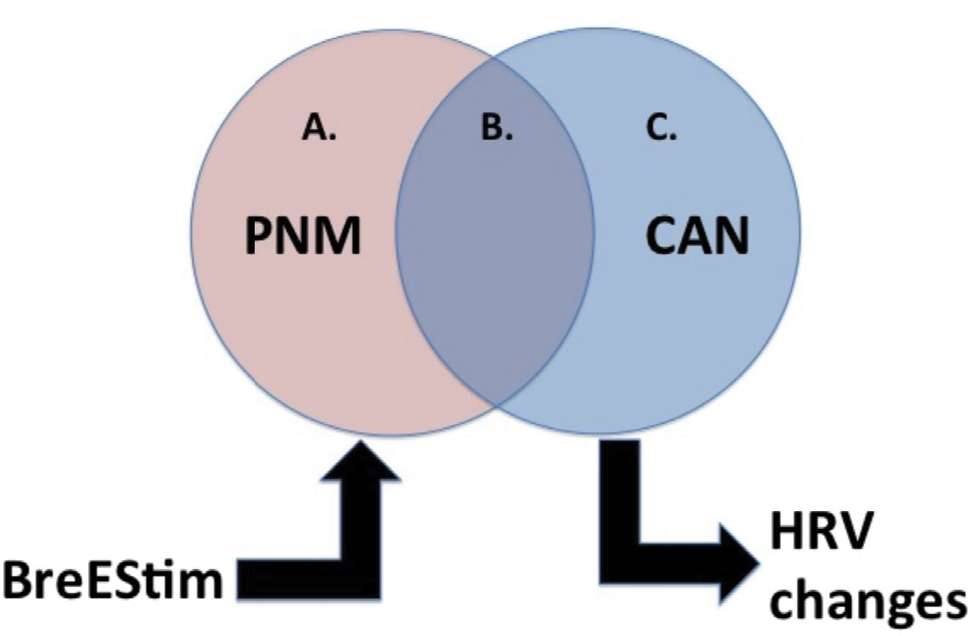 | Figure 1 Schematic depicting the proposed relationship among the PNM and the CAN. Notes: (A) Upon pain induction, the PNM is diffusely activated. It is susceptible to BreEStim influence. (B) The PNM–CAN of shared structures is specifically activated in chronic SCI. (C) The CAN helps regulate the autonomic system. BreEStim is hypothesized to modulate the PNM, including the shared PNM–CAN areas, to produce analgesic effects, and thus changes in HRV parameters as well. Abbreviations: BreEStim, breathing-controlled electrical stimulation; CAN, central autonomic network; HRV, heart rate variability; PNM, pain neuromatrix; SCI, spinal cord injury. |
These findings support the fact that NP after chronic SCI may be the result of maladaptive neuroplasticity and central sensitization occurring at the supraspinal level and that these alterations may be the resultant of injury chronicity.3,4,7–9,15 Moreover, it can also be suggested that autonomic activity may represent a surrogate measure to detect NP activity, as produced via the shared PNM–CAN, and prove to be useful in NP diagnostics in SCI populations.8–11
Heart rate variability (HRV), the physiological variance in the interbeat interval, is an increasingly documented modality for quantifying autonomic activity and thus reflecting CAN function.16,17 Numerous studies have even characterized HRV differences in subjects with various acute and chronic pain syndromes that include but are not limited to irritable bowel syndrome, lower back pain, fibromyalgia, and so on.18–22 Koenig et al23 have also established that HRV parameters can capture diminished parasympathetic influence with induction of experimental pain in healthy adults. We recently published an innovative study that evidenced the capacity of baseline HRV parameters, namely time domain variables, to differentiate chronic SCI subjects with and without NP.24 Additionally, we demonstrated that those HRV differences were unlikely to be influenced by the neurologic level or severity of the SCI itself.24
In this study, we intended to further explore modulation of chronic NP following SCI as detected by HRV changes. Essentially, we aimed to explore the utility of HRV differences to capture acute autonomic responses to treatment of chronic NP following SCI. To investigate this, we utilized centrally directed NP analgesia to measure changes in CAN activity. Breathing-controlled electrical stimulation (BreEStim) is a novel non-invasive intervention using electrical stimulation (EStim) that is triggered by voluntary breathing and delivered to a peripheral nerve transcutaneously. BreEStim has been shown to be effective for the treatment of chronic NP following SCI.25,26 Moreover, we have also presented evidence suggesting that BreEStim operates via selective modulation of various cortical and subcortical structures responsible for voluntary respiration and noxious stimuli processing.27–29 These neural structures are explicably the same components utilized in the PNM machinery and thus also the shared PNM–CAN. We hypothesized that centrally directed NP analgesia via BreEStim would affect PNM–CAN activity, and thus HRV measures, in persons with chronic NP following SCI. As such, BreEStim-induced analgesia was hypothesized to be accompanied by restoration of automatic dysfunctions in these persons with NP following SCI as seen in our study.24 In particular, we hypothesized that those HRV parameters produced by time domain analysis would be sensitive to parasympathetic tone changes associated with NP and with BreEStim treatment for NP.
Methods
Subject population
This study was conducted with the necessary approvals and recommendations of the University of Texas Health Sciences Center at Houston institutional review board. All subjects were recruited at our outpatient specialty SCI rehabilitation clinic, located in an academic setting. Inclusion and exclusion criteria for the study were as follows: 1) persons with capacity of providing consent, 2) who were between 18 and 75 years of age, 3) with an SCI for >6 months, 4) without clinically significant or unstable medical or neuropsychiatric disorders, 5) without a history of intracranial pathology, such as traumatic brain injury, stroke, or brain tumors, 6) without a history of intracranial intervention or surgery, 7) without a history of cardiac pathology or implanted pacemakers, 8) without the current usage of inotropic or chronotropic medications, such as beta-blockers, that may artificially alter the heart rate and/or rhythm, and 9) on a stable pain medication regimen, if needed, for at least 2 weeks prior to the experiment. All subjects provided written informed consent prior to the experiments.
A physician with subspecialty training and board certification in SCI medicine then assigned those subjects who met the primary inclusion/exclusion criteria to either SCI+NP or SCI–NP groups. Subjects in the SCI+NP group were determined to have chronic NP for >3 months, whereas subjects in the SCI–NP group were determined to be either without pain or with a nociceptive pain condition, ie, musculoskeletal in origin. Testing was excluded from those who were suspected to have autonomic dysreflexia in the preceding 24 hours. The primary experimental portion of the study was conducted in the SCI+NP group (n=10). To further interrogate the autonomic profiles of SCI subjects with and without pain, the SCI+NP group was then compared with the SCI–NP group (n=11). Of note, both SCI groups comprise a subset of subjects from our prior study exploring baseline HRV differences between SCI+NP and SCI–NP groups.24
VAS scoring and HRV collection
For SCI–NP subjects, HRV data were only collected at resting baseline (BS SCI–NP). For SCI+NP subjects, VAS scores and HRV data were collected at resting baseline (BS SCI+NP or pre-test SCI+NP) and at 30 minutes following the intervention (post-test SCI+NP). At both time points, the subjects were asked to rate their current pain level using the VAS pain scoring system, which designates scores from 0 through 10, with “0” described as no pain and a score of “10” described as agonizing, distressing, and/or the worst pain ever felt. Following pain reporting, the subjects were prepared for electrocardiogram (ECG) collection for HRV analysis and given a 5-minute resting period. After the skin was cleaned with an alcohol wipe to ensure appropriate electrode contact, disposable adhesive electrodes attached to ECG leads were placed on the subject’s chest. The white “right arm” and black “left arm” electrodes were placed in the first intercostal spaces along the midclavicular lines on the right and left chest wall, respectively. The red “left leg” electrode was placed in the lower intercostal spaces along the left mid-axillary line. After instructing subjects to remain calm, seated, and relaxed with limited movement, a 5-minute ECG recording was collected using a heart rhythm scanner (Biocom 5000 Wireless ECG Recorder; Biocom Technologies, Poulsbo, WA, USA). ECG heart rhythms were saved for off-line HRV analysis. The same ECG recording protocol was used as in our recent study.24
Breathing only (null) and BreEStim (active) interventions
All SCI+NP subjects participated in two experiments, which were administered in a random order, on different days, and at least 3 days apart. In Experiment 1 (null), a total of 120 voluntary, paced breaths were administered to subjects. In Experiment 2 (active), a total of 120 electrical stimuli during BreEStim were delivered to all subjects. Note that both the null and active interventions were each only performed once for each subject.
For Experiment 1 (null), subjects were seated comfortably and fitted with a leak-proof face mask, which was connected to a Pneumotach system (Hans Rudolph Inc., Shawnee, KS, USA) to record the airflow rate. Subjects were instructed to breathe using fast, strong, and deep inhalations. Data from the Pneumotach apparatus were displayed on the computer screen to ensure sufficient inhalation effort. After a brief training tutorial, a total of 120 breaths meeting experimental criteria were conducted.
For Experiment 2 (active), subjects followed the same protocol for deep, voluntary breathing as described earlier. Additionally, a pair of trimmed surface electrodes ~2 cm×2 cm separated by ~1 cm was placed over the ventral aspect of the distal forearm along the path of the median nerve. A single pulse (0.1 ms square wave) of EStim was delivered to the median nerve transcutaneously when the airflow rate reached the preset threshold level of 40% peak rate. This occurred when subjects took a fast, strong, and deep inhalation as instructed. Subjects were encouraged to gradually increase the intensity of EStim, before the halfway point of 60 breaths, to a painful yet tolerable pain level equivalent to a VAS score of 7–8. Subjects were explicitly instructed that EStim averseness was important for the BreEStim intervention. After a brief training tutorial, a total of 120 breathing-mediated EStim impulses were delivered.
Prior to the intervention in both experiments, subjects had a familiarization session to ensure that they understood the equipment, procedure, and requirements. For each treatment session that lasted ~30 minutes, the subjects were encouraged and allowed to take breaks to ensure sufficient rest. These settings and protocols were similar to our recent series of BreEStim experiments, which include studies evidencing BreEStim as an effective analgesic intervention of NP associated with SCI.25 Further technical details of BreEStim are available online in a methodology video article: http://www.jove.com/video/50077/.1
Data analysis
All demographic data were provided by the subjects. Clinical variables, particularly those relevant to the SCI, were gathered from the most recent International Standards for Neurological Classification of SCI examination documented by a physician with specialty training and board certification in SCI medicine. Relevant SCI-associated clinical variables gathered included the date of the injury, neurologic injury level, and severity of injury. Active prescription medications were surveyed in the electronic health record to determine the type, frequency, and dosage of NP medications being used. NP medications were assigned to categories of GABA analogs, which include gabapentin and pregabalin, atypical antidepressants, which include tricyclic antidepressants (TCA) and serotonin–norepinephrine reuptake inhibitors (SNRI), or opiates. As is conventional for quantifying opiates in scientific publications, daily morphine equivalents are reported.30
Kubios HRV analysis software (University of Eastern Finland, Joensuu, Finland) was used to evaluate the ECG recording.31 The heart rhythms were analyzed via time and frequency domain approaches to obtain various HRV parameters.21–24 The time domain parameters include SD of the normal-to-normal intervals (SDNN), root mean squared of successive differences (RMSSD), pairs of successive R–R beat lengths varying by greater than 50 ms (NN50), and proportion of NN50 for total number of beats (pNN50). SDNN reflects overall HRV, whereas RMSSD, NN50, and pNN50 reflect parasympathetic tone. The frequency domain parameters include low frequency (LF) band, high frequency (HF) band, total power, and low to high frequency ratio (LF/HF). Standard frequency stratification designating LF as 0.04–0.15 Hz and HF as 0.15–0.40 Hz, as determined by the fast Fourier transformation algorithm, was utilized. LF is an LF spectral component representing sympathetic-driven (baroreflex-mediated) vagal outflow, HF reflects parasympathetic tone, and total power reflects overall HRV, whereas LF/HF reflects degree of autonomic balance.
Statistical analyses
Statistical analyses were performed similar to those described in our recent publication.24 The major dependent variables were as follows: 1) VAS scores, 2) average HR, and 3) mean R–R length; the major HRV-dependent variables from the time domain analysis were as follows: 1) SDNN, 2) RMSSD, 3) NN50, and 4) pNN50; the major HRV-dependent variables from frequency domain analysis were as follows: 1) LF, 2) HF, 3) total power, and 4) LF/HF. We chose to analyze 1) demographics and SCI parameters for SCI+NP vs SCI–NP groups, 2) BS SCI+NP vs post-test SCI+NP groups for both null and active experiments, 3) the change between BS SCI+NP and post-test SCI+NP values across null or active experiments, and 4) BS SCI+NP active group vs post-test SCI+NP active group vs BS SCI–NP group.
For use in statistical analyses, all HRV parameters, expect SDNN, are non-normally distributed. For two group comparisons of similar groups, paired t-tests and Wilcoxon signed-rank tests were used for parametric and non-parametric variables, respectively. For two group comparisons of non-similar groups, independent samples t-tests and Mann–Whitney U tests were used for parametric and non-parametric variables, respectively. Two-way ANOVA tests and the Kruskal–Wallis tests with Scheirer–Ray–Hare extension were conducted to measure the interaction of time (BS and post-test) and treatment (null and active) factors in producing changes in parametric and non-parametric variables, respectively.
Statistical analysis was performed using the STATA Version 12.1 (StataCorp LP, College Station, TX, USA). An alpha level of 0.05 was used as threshold for significance for all statistical tests. Data were reported as mean±SD within the text and as mean±standard error of the mean (SEM) in the figures. Only the significant main effects were presented, unless otherwise noted.
Results
Study subjects
Our study cohort included 21 predominantly male, middle-aged subjects with SCI, who were dichotomized into SCI+NP (n=10) or SCI–NP (n=11) groups (Tables 1 and 2). Both groups were similar in age, gender, and SCI-relevant clinical parameters, which include duration of injury, tetraplegia (C8 and above), T4 and above injuries, and complete injury prevalence (Table 3). The clinically relevant cutoffs of C8 and above, and T4 and above, were chosen to reveal the number of persons in our study cohort with tetraplegia and disrupted sympathetic innervation to the heart, respectively.
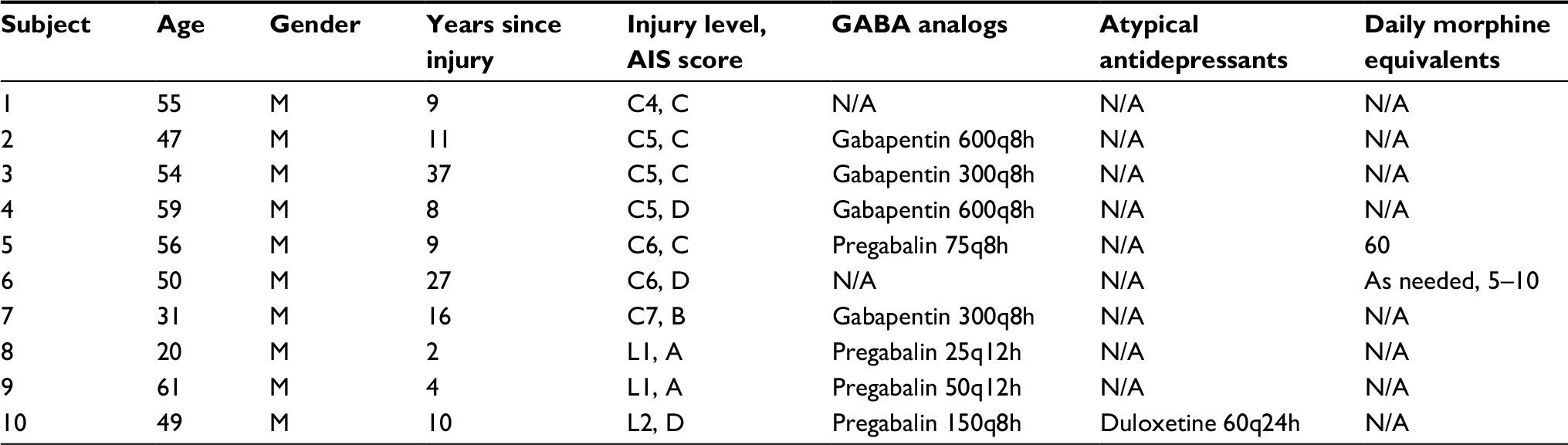 | Table 1 Pertinent demographics, SCI-relevant parameters, and pain medications, along with their milligram dosage and frequency, of each patient in the SCI+NP study group Abbreviations: AIS, American spinal injury association impairment scale; F, female; M, male; NP, neuropathic pain; SCI, spinal cord injury. |
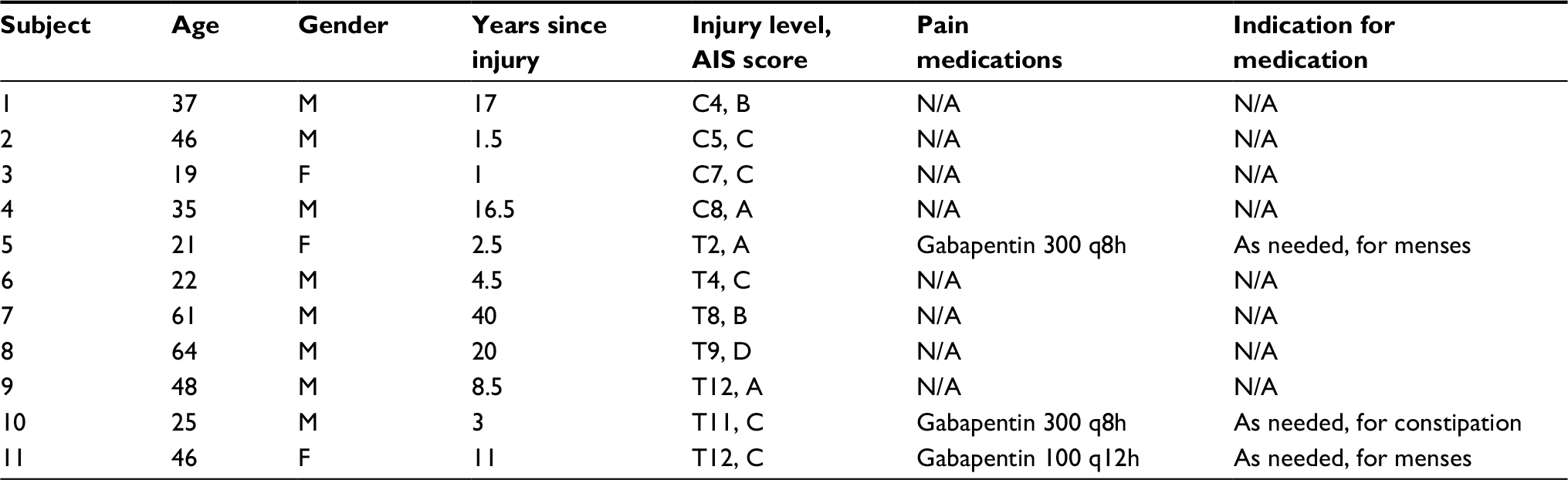 | Table 2 Pertinent demographics, SCI-relevant parameters, and pain medications, along with their milligram dosage, frequency, and indication, of each patient in the SCI–NP study group Abbreviations: AIS, American spinal injury association impairment scale; F, female; M, male; NP, neuropathic pain; SCI, spinal cord injury. |
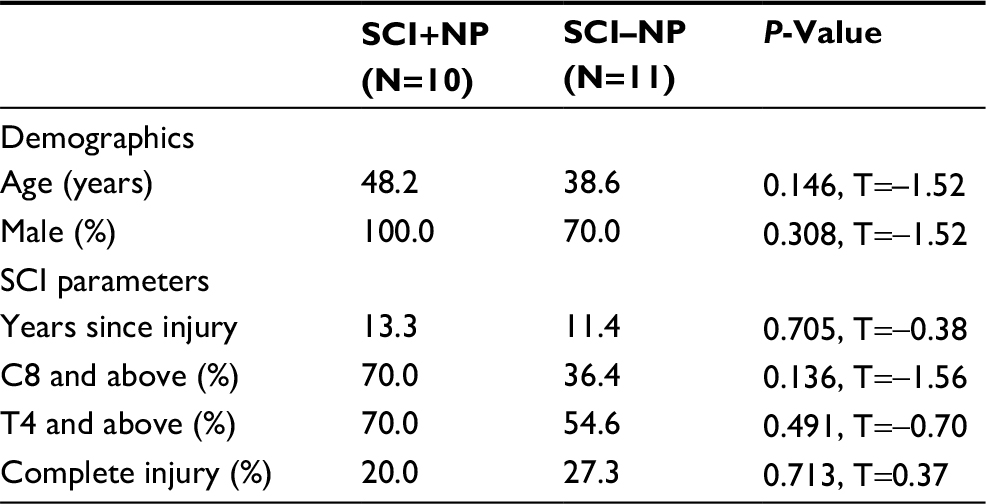 | Table 3 Pertinent demographics and SCI-relevant parameters of the SCI+NP and SCI–NP study groups Abbreviations: NP, neuropathic pain; SCI, spinal cord injury. |
In regard to NP treatment, most SCI+NP subjects were utilizing pain medications. The most commonly utilized medications were the GABA analogs, gabapentin (n=4) and pregabalin (n=4). Three subjects used other medications – the SNRI duloxetine (n=1) and an opioid (n=2). One patient (subject 1) was not using pain medications. In the SCI–NP group, only three subjects were using pain medications; they were using only gabapentin on an as-needed basis for indications other than NP. Regarding the interventions, all subjects tolerated treatment sessions well; no adverse events including episodes of hyperventilation or hypoxia were reported.
VAS, HRV changes for the null vs active experiments
The VAS scores reported during the baseline phase of both null and active interventions were comparable for SCI+NP subjects (Table 3 and Figure 2A). However, there was a significant interaction between the effects of time and treatment on VAS scores (F[1, 36]=4.42, P=0.04). Simple main effects analysis showed that VAS scores significantly decreased across time with the active treatment (P<0.01), but there were no differences across time with the null treatment.
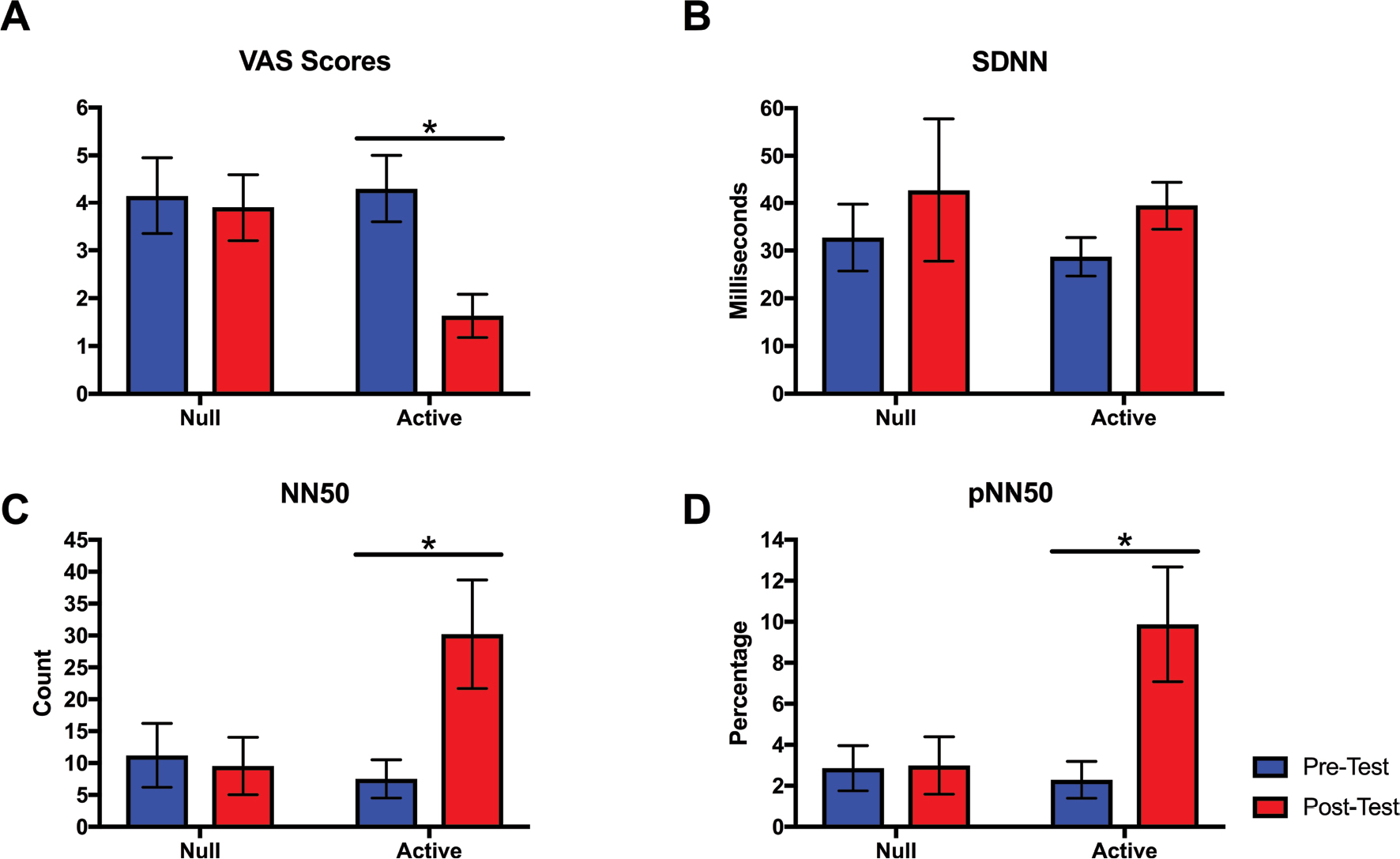 | Figure 2 The changes in various parameters across pre-test and post-test time points for null and active experiments in the SCI+NP study group (n=10). Notes: Parameters displayed include (A) VAS, (B) SDNN, (C) NN50: pairs of successive R–R beat lengths varying by greater than 50 ms, and (D) pNN50: proportion of NN50 for total number of beats. Standard error bars are shown; asterisk denotes statistically significant differences. Abbreviations: NP, neuropathic pain; SCI, spinal cord injury; SDNN, SD of the normal-to-normal intervals. |
The recorded mean HR and R–R length were comparable across both time and interventions. In regard to HRV time domain parameters, however, several key differences were appreciated (Table 3 and Figure 2B–D). Parasympathetic tone, as measured by NN50 and pNN50, was found to profoundly increase across time only for the active intervention. There was a significant interaction between the effects of time and treatment for both parameters (NN50: F[1, 36]=4.92, P=0.03; pNN50: F[1, 36]=4.57, P=0.04). Simple main effects analysis showed that the values of both parameters significantly increased across time with active treatment (NN50: P=0.03; pNN50: P=0.02), but there were no differences across time with null treatment. HRV frequency domain parameters did not change across time for either null or active treatments (Table 4).
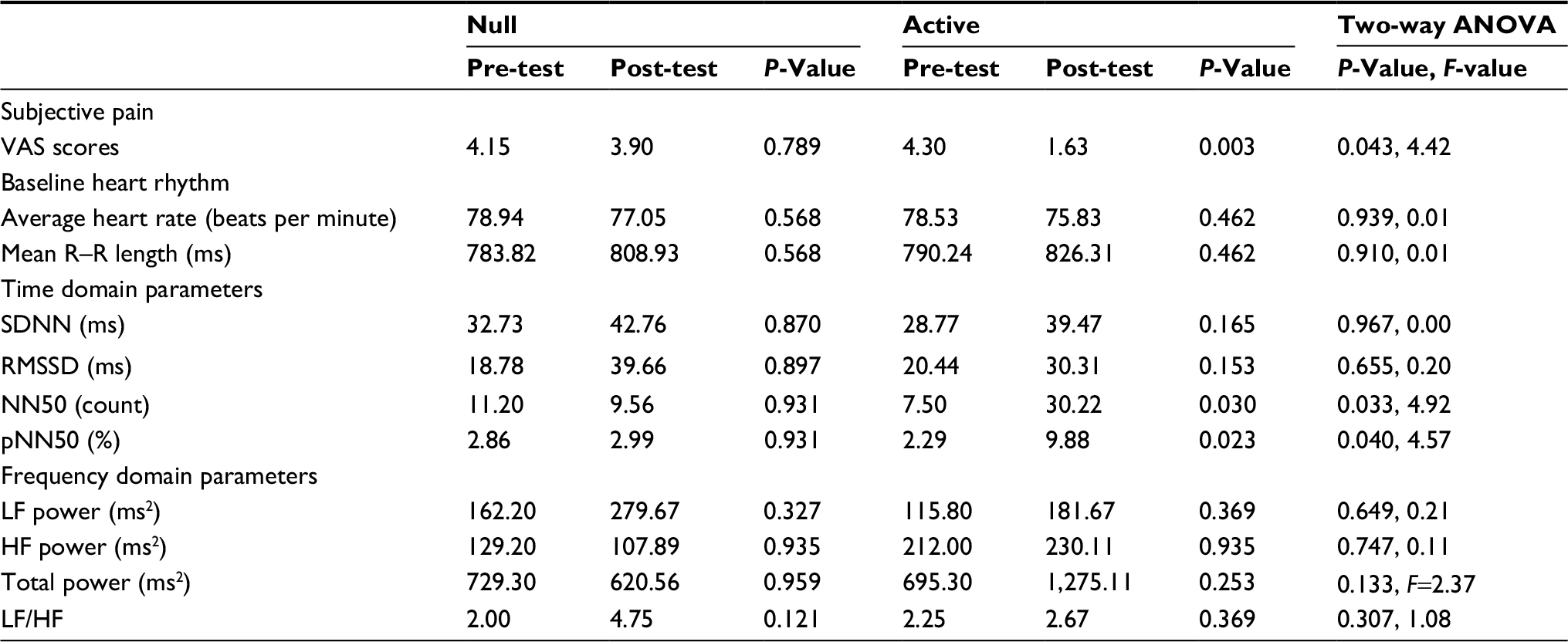 | Table 4 VAS scores, heart rhythm, and time domain and frequency domain parameters of the SCI+NP study groupa Notes: aValues obtained during baseline and post-test time points during the null and active experiments. NN50: pairs of successive R–R beat lengths varying by greater than 50 ms and pNN50: proportion of NN50 for total number of beats. Abbreviations: HF, high frequency; LF, low frequency; LF/HF, low to high frequency ratio; NP, neuropathic pain; RMSSD, root mean squared of successive differences; SCI, spinal cord injury; SDNN, SD of the normal-to-normal intervals. |
SCI+NP active treatment response vs SCI–NP baseline
To examine whether HRV parameters were restored after BreEStim along with the analgesic effects, we compared heart rhythm and HRV parameters for the active BS (or pre-test) SCI+NP group and the active post-test SCI+NP group each vs the BS SCI–NP group. There were no differences in the baseline heart rhythm for either group comparison (Table 5). In comparing HRV time domain parameters, however, the active pre-test SCI+NP group was found to have a lower HRV than the baseline SCI–NP group (P=0.01) (Table 5 and Figure 3), as determined by SDNN. For all other time domain parameters, both the SCI+NP groups were found to be comparable to the SCI–NP group. Additionally, HRV frequency domain analysis revealed the active pre-test SCI+NP group to have a lower overall HRV than the baseline SCI–NP group (P<0.05), as determined by LF (Table 5 and Figure 4). For all other frequency domain parameters, both the SCI+NP groups were found to be comparable to the SCI–NP group. Taken together, these comparisons suggested acute restoration of some autonomic function after the active treatment with BreEStim.
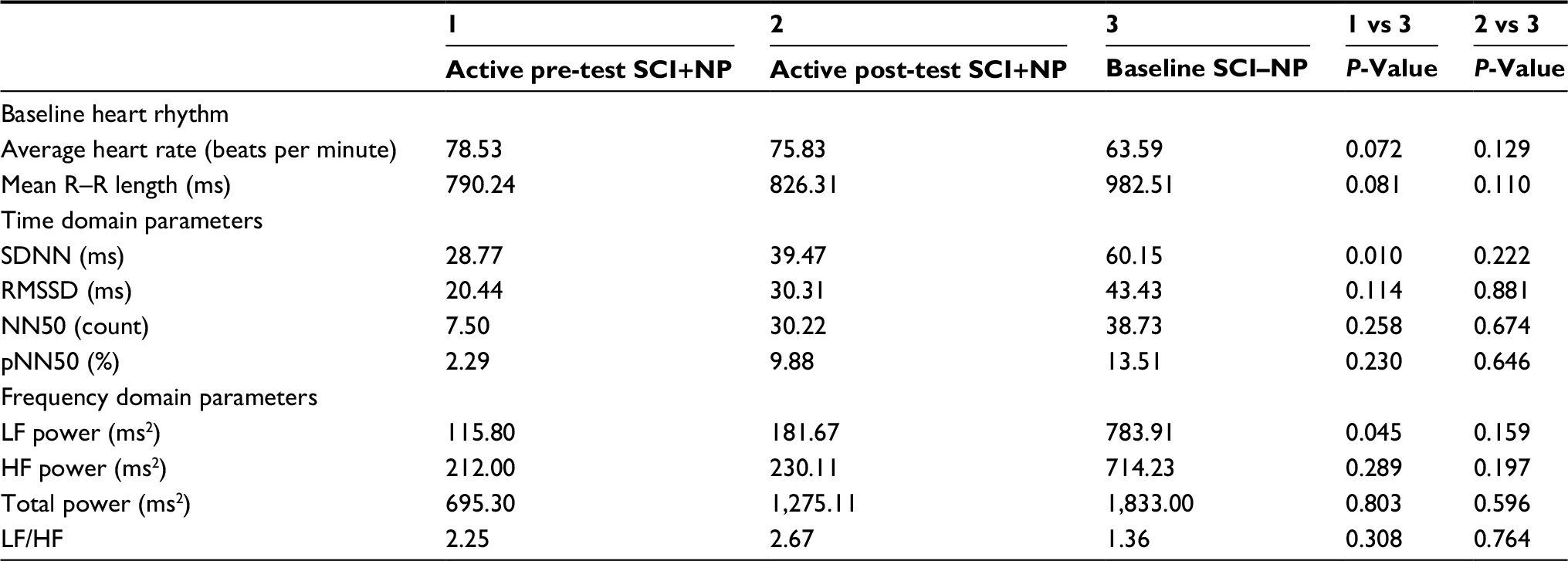 | Table 5 Heart rhythm, time domain parameters, and frequency domain parameters of the SCI+NP group during baseline and post-test time points and the SCI–NP group at baseline Note: NN50: pairs of successive R–R beat lengths varying by greater than 50 ms and pNN50: proportion of NN50 for total number of beats. Abbreviations: HF, high frequency; LF, low frequency; LF/HF, low to high frequency ratio; NP, neuropathic pain; RMSSD, root mean squared of successive differences; SCI, spinal cord injury; SDNN, SD of the normal-to-normal intervals. |
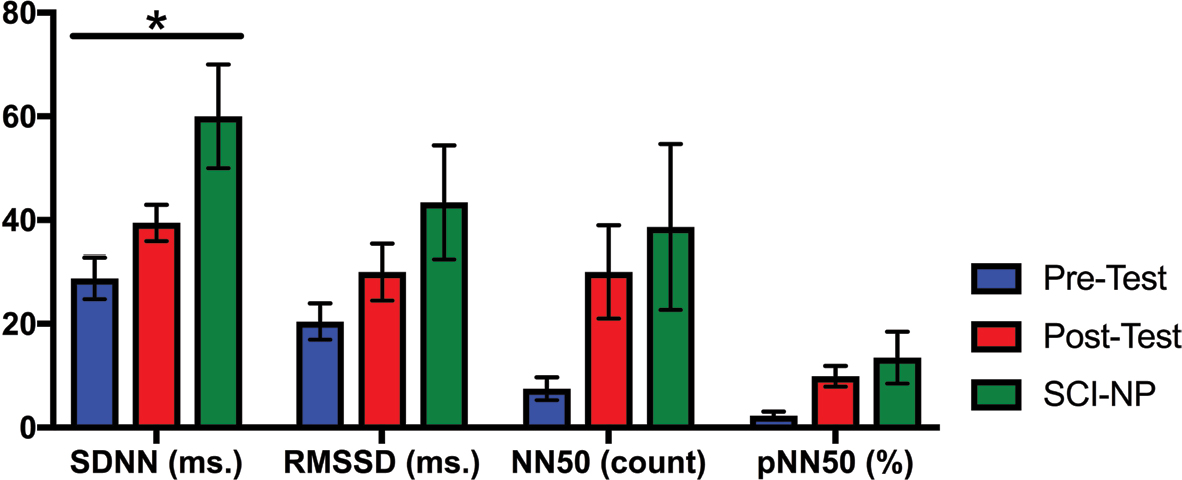 | Figure 3 The values of the HRV time domain parameters for the active pre-test, active post-test, and SCI–NP baseline tests are shown. Notes: Time domain parameters displayed include SDNN, RMSSD, NN50: pairs of successive R–R beat lengths varying by greater than 50 ms, and pNN50: proportion of NN50 for total number of beats. SDNN reflects overall HRV, whereas RMSSD, NN50, and pNN50 reflect parasympathetic tone. Standard error bars are shown; asterisk denotes statistically significant differences. Abbreviations: HRV, heart rate variability; NP, neuropathic pain; RMSSD, root mean squared of successive differences; SCI, spinal cord injury; SDNN, SD of the normal-to-normal intervals. |
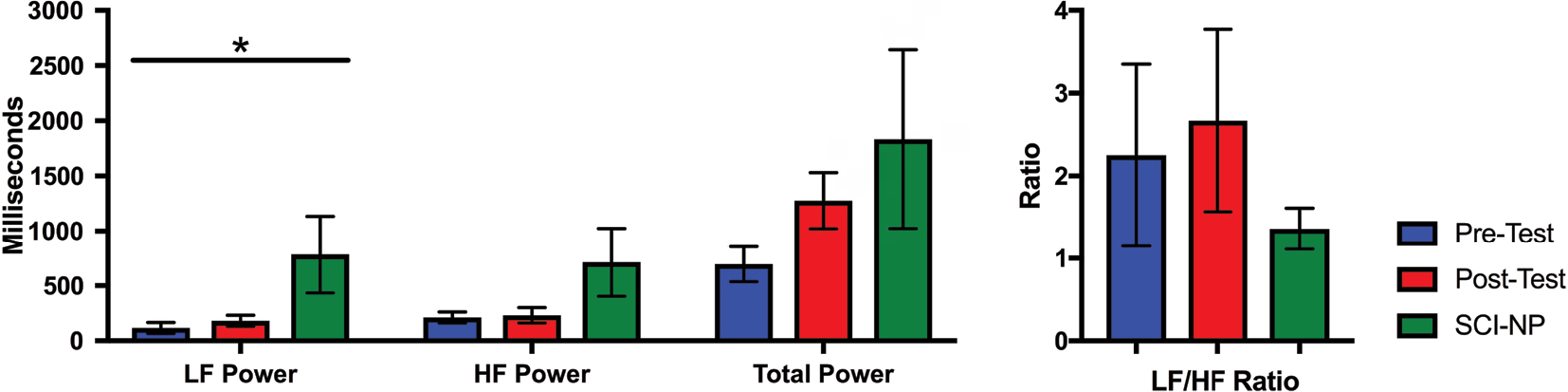 | Figure 4 The values of the frequency domain parameters for the active pre-test, active post-test, and SCI–NP baseline tests are shown. Note: Frequency domain parameters displayed include LF band, HF band, total power, and LF/HF. Standard error bars are shown; asterisk denotes statistically significant differences. Abbreviations: HF, high frequency; LF, low frequency; LF/HF, low to high frequency ratio; NP, neuropathic pain; SCI, spinal cord injury. |
Discussion
We have recently reported novel findings demonstrating the capability of certain baseline time domain HRV parameters to reflect autonomic aberrations specifically associated with chronic NP in an SCI cohort.24 Additionally, the findings suggested that those distinct HRV disparities were unlikely to be influenced by the neurological level of injury or SCI status itself. This succeeding study characterizes the capacity of HRV parameters to reflect autonomic restoration in response to a centrally targeted analgesic intervention in a cohort of subjects with SCI and chronic NP, concomitantly with analgesic effects. As such, the current findings further highlight central autonomic influences in the context of the supraspinal PMN–CAN-associated pathogenesis of chronic NP after SCI.
The crossover study design provided both null and active analgesic interventions, in a random order, to all subjects in the SCI+NP group. The experimental group of SCI+NP subjects was similar to our SCI–NP cohort in demographics and SCI clinical characteristics (Table 3). The novel and unique findings in this study were that certain HRV parameters, namely NN50 and pNN50, were able to capture the increase in parasympathetic tone following active analgesic intervention (Table 4, Figures 3 and 4). The increases in parasympathetic modulation across time were specific for active intervention and were associated with patient-reported analgesic effect. Additionally, other HRV parameters demonstrated baseline differences in autonomic profiles between SCI+NP and SCI–NP subjects (Table 5, Figures 3 and 4). Namely, SCI+NP subjects were found to exhibit a lower overall HRV and parasympathetic tone compared with SCI–NP subjects, as determined by lower values of SDNN and LF, respectively. Following active intervention, however, we observed that SCI+NP subjects displayed an increased overall HRV and parasympathetic tone (Table 4, Figures 3 and 4). This autonomic restoration, specific to active intervention, is associated with NP analgesic treatment response. Furthermore, the SCI+NP subjects following active intervention were found to be comparable to SCI–NP subjects regarding heart rhythm variability time and frequency domain parameters.
Sensitivity of time domain HRV parameters
These collective findings led us to suspect that there may be a degree of sensitivity for various HRV parameters. In other words, just as SDNN and LF were apt at measuring baseline differences between SCI+NP and SCI–NP subjects, they may not be sensitive to capturing acute changes in autonomic activity. Similarly, the NN50 and pNN50 variables, which were unable to differentiate baseline differences between SCI–NP and SCI+NP groups, were capable of measuring active treatment response. Thus, NN50 and pNN50 may be much more sensitive to acute changes in autonomic activity relative to chronic changes. The phenomenon of HRV changes during acute vs chronic stress has been reported in both human and animal studies.32–34 Moreover, these studies also find overall interbeat variance, SDNN, to be more susceptible to chronic stressors. Our recent studies further support sensitivity of time domain HRV parameters from 5-minute ECG recordings in capturing autonomic dysfunction after SCI.35,36 For example, as compared to frequency domain HRV parameters, SDNN/NN50 were able to better track autonomic responses to transcutaneous tibial nerve stimulation of neurogenic bladder based on urodynamic outcomes.35,36
BreEStim modulation of NP after SCI is associated with restoration of autonomic dysfunction
Our finding of active treatment response associated with an increase in parasympathetic tone can be supported by the normal mechanisms of the CAN machinery. In the absence of pain pathology, the CAN acts to regulate the sympathetic and parasympathetic output in response to the physiological environment.37,38 Numerous studies exploring HRV changes in the healthy adults found that a robust parasympathetic tone was a marker of healthy status.23,38–40 These findings are congruent with our study with decreased parasympathetic tone in persons with concomitant SCI and NP relative to persons with SCI but without NP or healthy persons.24 Therefore, it follows that the decreased parasympathetic tone in persons with SCI and NP may be secondary to altered mechanisms in the pathological CAN, which may be absent in persons with SCI and without NP and healthy persons.
The central thrust of this study – that NP modulations after SCI have supraspinal origins that are strongly associated with central autonomic pathogenesis – can also be substantiated by recent advances in the understanding of NP following amputation. Just as implicated in SCI, central neuroplastic changes have also been suggested to drive the pathogenesis of NP following amputations.41,42 Namely, the similar fMRI studies used to interrelate NP and SCI have also shown strong cortical and subcortical restructuring patterns in persons with NP after amputation, including structures involved in autonomic modulation. Even further, treating phantom pain has been found to correlate with the degree of somatosensory smudging.43 These similarities of supraspinal modulation of NP after SCI (injury at the central nervous system) and amputation (injury of the peripheral nerves) are further evidenced by reports of BreEStim providing efficacious analgesic effect despite treating both conditions via peripherally driven stimulation at the median nerve, rather than the injury foci.25,44 Moreover, because BreEStim to the median nerve on one side has also been shown to alter pain threshold capacity on both sides in healthy persons, it is thought to modulate activity at the central level, likely including the PNM. In the context of our current study, the extensively established BreEStim intervention for supraspinal analgesia substantiates our theorized paradigm schematically represented in Figure 1. Furthermore, the analgesia-associated autonomic changes as quantified by HRV in this study serve to demonstrate the interconnected PNM–CAN machinery.10,11
Additionally, the mechanistic actions of GABA analog medications also serve to further substantiate our findings. GABA analog medications, including gabapentin and pregabalin, are first-line treatments for NP.45 Gabapentin has been demonstrated to act by increasing the spinal-derived parasympathetic outflow by way of inhibiting glutamatergic transmission.46,47 Additionally, pregabalin functions by decreasing the release of norepinephrine, among other neurotransmitters, to ameliorate sympathetically modulated pain processes.48,49 Consequently, our findings suggest that increases in parasympathetic tone, as determined by HRV, may be correlated with analgesic perception. Collectively, our findings provide experimental evidence supporting the notion that chronic NP in persons with SCI is likely to be modulated at the supraspinal level.
Limitations
The primary limitation of this study was a small sample size; this study was powered primarily based on sample sizes of convenience. As such, further analyses exploring the impact of the neurological level of injury on HRV modulations after intervention were prevented. Findings from this work will help future studies in estimating the appropriate power and sample sizes required. Future higher powered studies exploring HRV changes associated with different NP treatments would provide useful information regarding the autonomic pathophysiology of NP-associated analgesia. Additionally, the use of fMRI in conjunction to analgesic experiments in subjects with SCI and NP may provide instrumental knowledge regarding neural activity in PNM–CAN.
Of note, many experimental pain studies including this study utilize a 5-minute ECG approach for HRV analysis for logistical ease. However, it has been suggested that frequency domain parameters may be more appropriately sensitive with longer recordings, including 24-hour ECG monitoring.16,17 Regardless, studies have shown that the capacity of frequency domain parameters extrapolated 5-minute ECG to reflect chronic pain, which substantiated the use of a 5-minute ECG approach in this study.19–22 Finally, restoration of autonomic dysfunction was observed concomitantly with analgesic effects after only one session of BreEStim treatment. It would be important to know for future clinical use whether this effect will be accumulated and sustained after long-term BreEStim treatment.
Conclusion
Increasing evidence suggests that supraspinal modulation of chronic NP after SCI is highly associated with central autonomic circuitry. However, the role of autonomic correlates associated in this context has been less investigated. In an experimental model, we found that centrally delivered analgesia, as provided by BreEStim, in a cohort of persons with chronic NP and SCI is associated with autonomic restoration, as measured by HRV changes. Our results provide investigational evidence supportive of the high degree of interconnectedness between the PNM and the CAN in modulating chronic NP following SCI and NP response to neuromodulatory interventions, such as BreEStim.
Acknowledgments
This study was funded in part by the Rehabilitation Research Experience for Medical Students (RREMS) Program of the Association of Academic Physiatrists (AAP); Mission Connect, a program of TIRR Foundation (015–116); and an NIH/NICHD/NCMRR grant (R21HD087128). The authors also acknowledge the subjects who took part in the study.
Author contributions
JK, SAL, LZ, YC, AS, and SL designed, discussed, and interpreted data. JK, SAL, LZ, YC, and AS performed the experiments and conducted necessary data analysis. JK, AS, and SL wrote the manuscript. All coauthors approved the final version of the manuscript. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
Sheng Li (the corresponding author) holds US Patent No 8,229,566 ‘‘Method and Apparatus of Breathing-Controlled Electrical Stimulation for Skeletal Muscles’’, issued on July 24, 2012, and US Patent No 8,588,919 ‘‘Method and Apparatus of Breathing-Controlled Electrical Stimulation for Skeletal Muscles’’ Divisional of Application No 12/146,176 (issued as US Patent 8,229,566). Sheng Li was blinded to all experiments and did not have direct patient contact in this study. All other authors declare that this study was conducted in the absence of any commercial or financial relationships that could be construed as a conflict of interest. The authors report no other conflicts of interest in this work.
References
Siddall PJ, Mcclelland JM, Rutkowski SB, Cousins MJ. A longitudinal study of the prevalence and characteristics of pain in the first 5 years following spinal cord injury. Pain. 2003;103(3):249–257. | ||
Jensen MP, Chodroff MJ, Dworkin RH. The impact of neuropathic pain on health-related quality of life: review and implications. Neurology. 2007;68(15):1178–1182. | ||
Yezierski RP, Robert P. Spinal cord injury pain: spinal and supraspinal mechanisms. J Rehabil Res Dev. 2009;46(1):95–108. | ||
Siddall PJ. Management of neuropathic pain following spinal cord injury: now and in the future. Spinal Cord. 2009;47(5):352–359. | ||
Seifert F, Schuberth N, de Col R, et al. Brain activity during sympathetic response in anticipation and experience of pain. Hum Brain Mapp. 2013;34(8):1768–1782. | ||
Duerden EG, Albanese MC. Localization of pain-related brain activation: a meta-analysis of neuroimaging data. Hum Brain Mapp. 2013;34(1):109–149. | ||
Komaki Y, Hikishima K, Shibata S, et al. Functional brain mapping using specific sensory-circuit stimulation and a theoretical graph network analysis in mice with neuropathic allodynia. Sci Rep. 2016;6(1). | ||
Gustin SM, Wrigley PJ, Henderson LA, Siddall PJ. Brain circuitry underlying pain in response to imagined movement in people with spinal cord injury. Pain. 2010;148(3):438–445. | ||
Zeilig G, Enosh S, Rubin-Asher D, Lehr B, Defrin R. The nature and course of sensory changes following spinal cord injury: predictive properties and implications on the mechanism of central pain. Brain. 2012;135(Pt 2):418–430. | ||
Appelhans BM, Luecken LJ. Heart rate variability and pain: associations of two interrelated homeostatic processes. Biol Psychol. 2008;77(2):174–182. | ||
Napadow V, Dhond R, Conti G, et al. Brain correlates of autonomic modulation: combining heart rate variability with fMRI. Neuroimage. 2008;42(1):169–177. | ||
Melzack R, Wall PD. Pain mechanisms: a new theory. Survey Anesthesiol. 1967;11:89–90. | ||
Melzack R, Loeser JD. Phantom body pain in paraplegics: Evidence for a central “pattern generating mechanism” for pain. Pain. 1977;4(Supp C):195–210. | ||
Melzack R. Pain and the neuromatrix in the brain. J Dent Educ. 2001;65(12):1378–1382. | ||
Hulsebosch CE. Pathophysiology and animal models of spinal cord injury pain. In: Yezierski RP, Burchiel K. Spinal Cord Injury Pain: Assessment, Mechanisms, Management. Seattle, WA: IASP Press; 2002:117–136. | ||
Sztajzel J. Heart rate variability: a noninvasive electrocardiographic method to measure the autonomic nervous system. Swiss Med Wkly. 2004;134(35–36):514–522. | ||
Task Force of the European Society of Cardiology. Heart rate variability: standards of measurement, physiological interpretation, and clinical use. Circulation. 1996;93:1043–1065. | ||
Storella RJ, Shi Y, O’Connor DM, Pharo GH, Abrams JT, Levitt J. Relief of chronic pain may be accompanied by an increase in a measure of heart rate variability. Anesth Analg. 1999;89(2):448–450. | ||
Adeyemi EO, Desai KD, Towsey M, Ghista D. Characterization of autonomic dysfunction in patients with irritable bowel syndrome by means of heart rate variability studies. Am J Gastroenterol. 1999;94(3):816–823. | ||
Kalezic N, Åsell M, Kerschbaumer H, Lyskov E. Physiological reactivity to functional tests in patients with chronic low back pain. J Musculoskelet Pain. 2007;15(1):29–40. | ||
Cohen H, Neumann L, Shore M, Amir M, Cassuto Y, Buskila D. Autonomic dysfunction in patients with fibromyalgia: application of power spectral analysis of heart rate variability. Semin Arthritis Rheum. 2000;29(4):217–227. | ||
Evans S, Seidman LC, Tsao JC, Lung KC, Zeltzer LK, Naliboff BD. Heart rate variability as a biomarker for autonomic nervous system response differences between children with chronic pain and healthy control children. J Pain Res. 2013;6:449. | ||
Koenig J, Jarczok MN, Ellis RJ, Hillecke TK, Thayer JF. Heart rate variability and experimentally induced pain in healthy adults: a systematic review. Eur J Pain. 2014;18(3):301–314. | ||
Karri J, Zhang L, Li S, Chen YT, Stampas A, Li S. Heart Rate Variability: A novel modality for diagnosing neuropathic pain after spinal cord injury. Front Physiol. 2017;8:495. | ||
Li S, Davis M, Frontera JE, Li S. A novel nonpharmacological intervention - breathing-controlled electrical stimulation for neuropathic pain management after spinal cord injury - a preliminary study. J Pain Res. 2016;9:933–940. | ||
Li S. Breathing-controlled Electrical Stimulation (BreEStim) for management of neuropathic pain and spasticity. J Vis Exp. 2013:71. | ||
Li S, Berliner JC, Melton DH, Li S. Modification of electrical pain threshold by voluntary breathing-controlled electrical stimulation (BreEStim) in healthy subjects. PLoS One. 2013;8(7):e70282. | ||
Li S, Hu T, Beran MA, Li S. Habituation to experimentally induced electrical pain during voluntary-breathing controlled electrical stimulation (BreEStim). PLoS One. 2014;9(8):e104729. | ||
Hu H, Li S, Li S. Pain modulation effect of breathing-controlled electrical stimulation (BreEStim) is not likely to be mediated by deep and fast voluntary breathing. Sci Rep. 2015;5:14228. | ||
Schneider C, Yale SH, Larson M. Principles of pain management. Clin Med Res. 2003;1(4):337–340. | ||
Tarvainen MP, Niskanen JP, Lipponen JA, Ranta-Aho PO, Karjalainen PA. Kubios HRV—heart rate variability analysis software. Comput Methods Programs Biomed. 2014;113(1):210–220. | ||
Bigger JT, Fleiss JL, Steinman RC, Rolnitzky LM, Schneider WJ, Stein PK. RR variability in healthy, middle-aged persons compared with patients with chronic coronary heart disease or recent acute myocardial infarction. Circulation. 1995;91(7):1936–1943. | ||
Coppens M, Vindla S, James DK, Sahota DS. Computerized analysis of acute and chronic changes in fetal heart rate variation and fetal activity in association with maternal smoking. Am J Obstet Gynecol. 2001;185(2):421–426. | ||
Cyr NE, Dickens MJ, Romero LM. Heart rate and heart-rate variability responses to acute and chronic stress in a wild-caught passerine bird. Physiol Biochem Zool. 2009;82(4):332–344. | ||
Stampas A, Zhu L, Li S. Randomized trial of bladder neuromodulation with transcutaneous tibial nerve stimulation in acute spinal cord injury: evidence of autonomic nervous system changes. Neurourol Urodynamics. 2018. | ||
Stampas A, Zhu L, Li S. Orthostatic hypotension is a significant confounder in heart rate variability analyses of spinal cord injury. Spinal Cord Ser Cas. 2018. | ||
Benarroch EE. The central autonomic network: functional organization, dysfunction, and perspective. Mayo Clin Proc. 1993;68(10):988–1001. | ||
Macey PM, Ogren JA, Kumar R, Harper RM. Functional imaging of autonomic regulation: methods and key findings. Front Neurosci. 2015;9:513. | ||
Kristal-Boneh E, Raifel M, Froom P, Ribak J. Heart rate variability in health and disease. Scand J Work Environ Health. 1995;21(2):85–95. | ||
Schubert C, Lambertz M, Nelesen RA, Bardwell W, Choi JB, Dimsdale JE. Effects of stress on heart rate complexity—a comparison between short-term and chronic stress. Biol Psychol. 2009;80(3):325–332. | ||
Yanagisawa T, Fukuma R, Seymour B, et al. Induced sensorimotor brain plasticity controls pain in phantom limb patients. Nat Commun. 2016;7:13209. | ||
Lyu Y, Guo X, Bekrater-Bodmann R, Flor H, Tong S. Phantom limb perception interferes with motor imagery after unilateral upper-limb amputation. Sci Rep. 2016;6:21100. | ||
Flor H, Denke C, Schaefer M, Grüsser S. Effect of sensory discrimination training on cortical reorganisation and phantom limb pain. Lancet. 2001;357(9270):1763–1764. | ||
Li S, Melton DH, Berliner JC. Breathing-controlled electrical stimulation could modify the affective component of neuropathic pain after amputation: a case report. J Pain Res. 2012;5:71. | ||
Hatch MN, Cushing TR, Carlson GD, Chang EY. Neuropathic pain and SCI: Identification and treatment strategies in the 21st century. J Neurol Sci. 2018;384:75–83. | ||
Coderre TJ, Kumar N, Lefebvre CD, Yu JS, Js Y. Evidence that gabapentin reduces neuropathic pain by inhibiting the spinal release of glutamate. J Neurochem. 2005;94(4):1131–1139. | ||
Maiorov DN, Krenz NR, Krassioukov AV, Weaver LC. Role of spinal NMDA and AMPA receptors in episodic hypertension in conscious spinal rats. Am J Physiol. 1997;273(3 Pt 2):H1266–H1274. | ||
Han DW, Kweon TD, Lee JS, Lee YW. Antiallodynic effect of pregabalin in rat models of sympathetically maintained and sympathetic independent neuropathic pain. Yonsei Med J. 2007;48(1):41–47. | ||
Harris RE, Napadow V, Huggins JP, et al. Pregabalin rectifies aberrant brain chemistry, connectivity, and functional response in chronic pain patients. Anesthesiology. 2013;119(6):1453–1464. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.