Back to Journals » International Journal of Nanomedicine » Volume 19
Neural Stem Cell-Derived Small Extracellular Vesicles: key Players in Ischemic Stroke Therapy – A Comprehensive Literature Review
Authors Zhu Z ![]() , Zhang Q
, Zhang Q ![]() , Feng J, Zebaze Dongmo S
, Feng J, Zebaze Dongmo S ![]() , Zhang Q, Huang S
, Zhang Q, Huang S ![]() , Liu X, Zhang G, Chen L
, Liu X, Zhang G, Chen L ![]()
Received 24 November 2023
Accepted for publication 1 May 2024
Published 14 May 2024 Volume 2024:19 Pages 4279—4295
DOI https://doi.org/10.2147/IJN.S451642
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Farooq A. Shiekh
Zhihan Zhu,1,* Qiankun Zhang,1,* Jia Feng,1 Sonia Zebaze Dongmo,1 Qianqian Zhang,1 Songze Huang,1 Xiaowen Liu,1 Guilong Zhang,2 Lukui Chen1
1Department of Neurosurgery, Southern Medical University Hospital of Integrated Traditional Chinese and Western Medicine, Southern Medical University, Guangzhou, Guangdong Province, People’s Republic of China; 2Department of Neurosurgery, The Second Affiliated Hospital of Guangzhou Medical University, Guangzhou, Guangdong Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lukui Chen, Department of Neurosurgery, Southern Medical University Hospital of Integrated Traditional Chinese and Western Medicine, Southern Medical University, Guangzhou, People’s Republic of China, Email [email protected] Guilong Zhang, Department of Neurosurgery, The Second Affiliated Hospital of Guangzhou Medical University, Guangzhou, People’s Republic of China, Email [email protected]
Abstract: Ischemic stroke, being a prominent contributor to global disability and mortality, lacks an efficacious therapeutic approach in current clinical settings. Neural stem cells (NSCs) are a type of stem cell that are only found inside the nervous system. These cells can differentiate into various kinds of cells, potentially regenerating or restoring neural networks within areas of the brain that have been destroyed. This review begins by providing an introduction to the existing therapeutic approaches for ischemic stroke, followed by an examination of the promise and limits associated with the utilization of NSCs for the treatment of ischemic stroke. Subsequently, a comprehensive overview was conducted to synthesize the existing literature on the underlying processes of neural stem cell-derived small extracellular vesicles (NSC-sEVs) transplantation therapy in the context of ischemic stroke. These mechanisms encompass neuroprotection, inflammatory response suppression, and endogenous nerve and vascular regeneration facilitation. Nevertheless, the clinical translation of NSC-sEVs is hindered by challenges such as inadequate targeting efficacy and insufficient content loading. In light of these limitations, we have compiled an overview of the advancements in utilizing modified NSC-sEVs for treating ischemic stroke based on current methods of extracellular vesicle modification. In conclusion, examining NSC-sEVs-based therapeutic approaches is anticipated to be prominent in both fundamental and applied investigations about ischemic stroke.
Keywords: neural stem cells, small extracellular vesicles, ischemic stroke, neuroprotection, neural regeneration
Introduction
Ischemic stroke refers to a clinical condition characterized by the sudden onset of neurological impairment resulting from a significant decrease or complete cessation of blood flow to the brain.1 The main pathological manifestations observed in cases of ischemic stroke encompass substantial neuronal cell death, disruption of the axonal network, generation of reactive oxygen species and free radicals, production of cerebral oedema, impairment of the blood-brain barrier (BBB), and subsequent neuroinflammatory responses, ultimately resulting in the loss of local neural tissue and associated functionality.2 The primary objective of clinical intervention for stroke is to promptly reinstate regional cerebral blood flow and minimize the occurrence and magnitude of impairment or dysfunction following stroke.3 Nevertheless, irreversible damage to neural tissue often occurs due to the limited time window for therapy and secondary brain injury.4 Furthermore, after reperfusion, the available neural cells remain inadequate for restoring and substituting necrotic functional neural tissue.
NSCs have the multidirectional potential to differentiate into astrocytes, neurons, and oligodendrocytes. Over the past decades, NSCs have served as a valuable therapeutic modality for treating neurological diseases, providing new therapeutic tools and approaches for many refractory neurological diseases,5,6 including ischemic stroke.7 However, the quick clinical translation of transplanted NSCs has been hindered by concerns over their potential oncogenicity and immunological rejection.8,9 Recent studies have revealed that paracrine effects (extracellular vesicles) are the predominant mode of action after stem cell transplantation, and extracting functional extracellular vesicles instead of NSC transplantation is promising to become a significant therapeutic approach.10
Small extracellular vesicles (sEVs) are a kind of vesicles released through exocytosis and have a variety of biologically active components, including nucleic acids, proteins and lipids.11,12 Through membrane fusion with recipient cells, sEVs facilitate intercellular communication by transferring their cargo to the recipient cells.13 As a result, sEVs show tremendous promise as a rapidly developing medical tool for treating many disorders. Extensive research has been conducted thus far on the therapeutic potential of NSC-sEVs in the context of ischemic stroke.14 The specific processes encompass neuroprotection, inflammatory response suppression, endogenous nerve regeneration facilitation, and vascular regeneration stimulation. Furthermore, the engineering of NSC-sEVs presents a potential solution to address the concerns about the inadequate targeting capability and insufficient content loading of NSC-sEVs in isolation. NSC-sEVs can precisely target cells or tissues through this engineering process, making achieving the desired therapeutic results easier.
This study aims to provide a comprehensive overview of the biological functions and therapeutic benefits of NSC-sEVs in managing ischemic stroke. Additionally, it addresses the challenges encountered in applying NSC-sEVs and suggests potential avenues for future advancements in this field.
The Ischemic Stroke and Its Treatment Modalities
Stroke is a prevalent ailment characterized by significant rates of impairment, recurrence, and mortality.15 It stands as one of the three primary diseases that have a substantial impact on human health, alongside coronary heart disease and cancer. The findings of a comprehensive examination conducted on the Global Burden of Disease Study 2019 revealed notable trends in the occurrence and impact of stroke from 1990 to 2019.16 Specifically, there was a substantial 70.0% rise in the absolute quantity of stroke incidents, an 85.0% increase in the prevalence of such events, a 43.0% surge in the number of fatalities, and a 32.0% elevation in disability resulting from a stroke on a global scale. In the year 2019, the majority of stroke episodes, precisely 62.4%, were classified as ischemic strokes. Additionally, 27.9% of the stroke cases were identified as cerebral hemorrhages. Cerebral hemorrhages accounted for 27.9% of the cases, whereas subarachnoid hemorrhages constituted 9.7% of the total.16 Ischemic stroke remains the most prevalent type of stroke, comprising the majority of newly occurring strokes. Ischemic stroke is associated with significant disability and mortality rates, resulting in substantial physical and psychological suffering for patients.
Ischemic stroke is a condition in which the blood supply to the brain tissue is drastically reduced or interrupted within a short period due to the blockage of the cerebral blood vessels, thus causing ischemic and hypoxic damage to the brain tissue and, ultimately, the patient suffers from severe neurological deficits.17 Brain tissue damage following a stroke can be directly linked to the lack of blood supply. After an episode of ischemia, neuronal cells undergo an energy reduction, resulting in the stoppage of action in the ion pump, which is highly dependent on energy.18 The process of membrane depolarization induces the release of neurotransmitters and presynaptic excitatory transmitters by neuronal cells, hence establishing a positive feedback loop characterized by persistent depolarization and neurotransmitter release.19 Extramembrane calcium ions are transported into the cell through N-methyl-D-aspartate receptor-gated channels. Due to the accumulation of intracellular lactate and the lack of adenosine triphosphate, multiple downstream signaling pathways are over-activated, resulting in calcium in-flow and intracellular calcium overload, which leads to the catabolism of membrane phospholipids and the disruption of the cytoskeleton, and the generation of a large number of free radicals.20 Simultaneously, concurrent inflammatory reactions occurring inside the brain parenchyma, such as the activation of glial cells and endothelial cells, along with the secretion of pro-inflammatory mediators,21 contribute to subsequent neuronal cell damage (Figure 1).22 The progressive evolution of these interrelated reactions results in an enhanced incidence of ischemia, leading to the damage and death of neuronal cells in the surrounding regions, especially in the area known as the ischemic penumbra, even with blood supply restored to the ischemic zone, irreversible brain tissue damage persists, eventually leading to permanent necrosis and loss of function. The prevailing therapeutic approach for ischemic stroke is promptly restoring local cerebrovascular blood flow using pharmacological or mechanical thrombolysis to save the affected tissues to the greatest extent possible, minimizing post-stroke neurological impairments.23 Tissue-type plasminogen activator (tPA) was the first pharmacological drug approved by the US Food and Drug Administration for treating ischemic stroke,24 and it serves as a standard medication for clinical revascularization. tPA catalysis converts plasminogen into the active serine protease known as plasmin via an enzymatic process. This reaction facilitates crosslinked fibrin degradation, promoting the dissolution of blood clots, a process referred to as thrombolysis. However, only a tiny proportion of patients, between 3% and 5%, benefit from this medication because of the 4.5-hour usage window and possible side effects related to tPA administration. In contrast to intravenous thrombolysis, endovascular therapies, such as mechanical and arterial thrombolysis, offer several benefits, including reduced drug dosage, increased local drug concentration, minimized systemic adverse effects, and increased recovery rates. In the context of individuals diagnosed with cerebral ischemia accompanied by significant arterial blockage, endovascular intervention has emerged as a potential therapeutic option instead of tPA. However, stroke centers usually perform endovascular intervention with the necessary neuro-interventional capabilities. This method has several drawbacks, such as an increased risk of cerebral bleeding, a narrow application range, the complexity of the process, and a lengthy recovery period. Furthermore, both techniques are limited by the therapeutic window and only apply to a subset of the patient group. There is a compelling need to develop a new therapeutic technique for cerebral infarction that can expand the temporal window, reduce side effects, and improve overall prognosis.
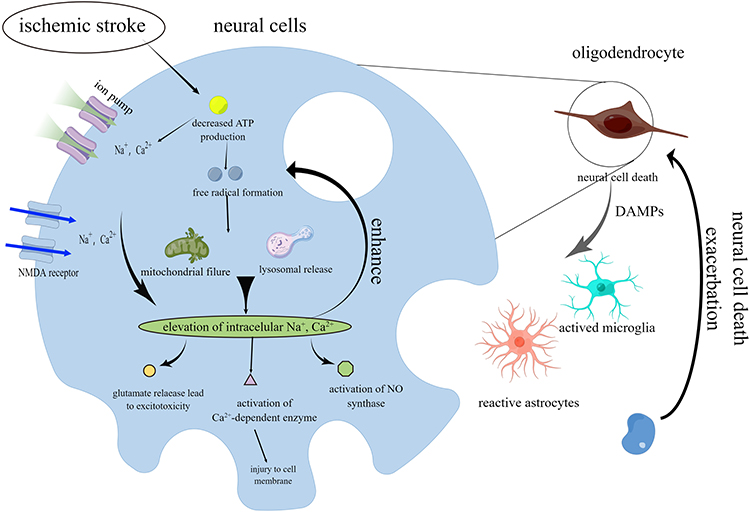 |
Figure 1 Schematic view of ischemic stroke pathophysiology. Blocking cerebral blood flow reduces neurons. Neurotransmitters and presynaptic excitatory substances are released when the cell membrane depolarizes. Calcium ions from outside enter the cell via N-methyl-D-aspartate receptor-gated channels. Calcium ions enter and accumulate in the cell, breaking down cell membrane phospholipids and disturbing the cytoskeleton. Pro-inflammatory mediators from glial and endothelial cell activation cause secondary neuronal cell damage. By Figdraw. |
The study of neuroprotective drugs and their effectiveness in treating ischemic stroke has been a significant topic of academic research. The foundation of prospective neuroprotective therapy is inhibiting or blocking the sequence of events that lead to neuronal cell injury.25 In the context of acute ischemic stroke, neuroprotection refers to the idea of minimizing damage to neurons by promptly administering medication either when symptoms appear or during the time the patient is waiting for blood reperfusion.26 In the past few years, a variety of pharmaceutical agents have been employed for neuroprotection following ischemic stroke. These agents include anti-inflammatory factors, antioxidants, glutamate antagonists, neurotrophic factors, γ-aminobutyric acid receptor agonists, calcium antagonists, and free radical scavengers.27,28 These neuroprotective medicines have shown promise in improving the prognosis of ischemic stroke in several preclinical and clinical trials; nevertheless, these results must be further validated and standardized. This necessity arises due to various factors, such as the heterogeneity of stroke in human subjects, the evaluation metrics employed in animal studies, the optimal time window for drug administration, and the influence of brain aging on treatment outcomes.29
Following cerebral ischemia, the neuronal regeneration and remodeling process is essential in treating ischemic stroke. Studies have shown that using stem cells, such as transplanting cells or promoting the natural regeneration of neural tissue, has the potential to be an effective method for repairing damaged tissues. These therapies have the potential to provide an extended therapeutic time frame for treating ischemic stroke.30
How Effective is Neural Stem Cell Therapy for Ischemic Stroke
Stem cells are a unique set of cells that can differentiate into multiple types of cells and regenerate themselves effectively. Under specific circumstances, it can undergo differentiation into a multitude of distinct tissue cells. Based on this property, transplanting stem cells and their related derivatives into patients can cure diseases by replacing damaged cells or promoting regeneration in the body.31 The advancement of stem cell technology has led to the extensive utilization of stem cells in various vital tissues and organs within the human body, as well as in numerous medical conditions encountered by individuals. This utilization offers a promising avenue for the efficacious treatment of allergic rhinitis,32 spinal cord injury,33 diabetes,34 cardiovascular diseases, bone defects,35 tumors,36,37 and other related ailments.
According to a meta-analysis, the transplantation of stem cells is efficacious in improving neurological impairments following cerebral ischemia. Additionally, it has been observed to enhance motor function and voluntary movements in the limbs of patients.38 Mesenchymal stem cells (MSCs) and NSCs are the focal points of research in the domain of ischemic stroke. Following transplantation into the body via stereotactic orientation or blood circulation, these stem cells possess specific homing characteristics that facilitate migration towards the injury site. Once positioned, they exhibit survival, growth, and differentiation into glial and neuronal cells.39
MSCs can generate a range of cytokines, including brain-derived neurotrophic factor (BDNF), basic fibroblast growth factor, and vascular endothelial growth factor (VEGF).40,41 One of the factors that plays a significant role in various physiological processes is BDNF, which promotes the survival of nerve cells and enhances synaptic plasticity and neurogenesis.42 Additionally, VEGF is known to trigger the development of blood vessels in the vicinity of cerebral ischemia. Several studies have demonstrated that transferring mitochondria from MSCs to cells damaged by stroke can have a neuroprotective impact.43 Furthermore, neural stem cell transplantation has been found to decrease the size of cerebral infarction and improve neurological function.44 Consequently, stem cell transplantation has emerged as a potent therapeutic intervention, saving patients from life-threatening conditions and enhancing their overall prognosis.
NSCs are a specific group of stem cells found in the nervous system.45 They can differentiate into multiple cell types and renew themselves. Initially, it was commonly believed that NSCs were only found during the early stages of development and in neonates. It was also thought that the adult brain and spinal cord could not repair damaged nerves. In 1992, Reynolds et al successfully extracted a population of cells from the striatum of adult mice that exhibited the ability to self-renew and differentiate into many lineages.46 Consequently, the concept of NSCs was categorically introduced, challenging the conventional belief that nerve cells are incapable of regeneration. In 1997, McKay introduced the idea of NSCs in the Journal of Science. NSCs are characterized as a population of cells that can differentiate into astrocytes, neurons, and oligodendrocytes while exhibiting self-renew ability.47 This unique property of NSCs enables them to generate a substantial quantity of nerve cells within the brain.
During the stage of adulthood, NSCs can be observed in two specific regions of the brain: the sub-granular zone of the dentate gyrus in the hippocampus region and the subventricular zone (SVZ) surrounding the lateral ventricles (Figure 2).48 NSCs undergo development and differentiation in response to the local microenvironment following migration along specific pathways towards their functional locations. Stem cells in the subgranular zone (SGZ) and dentate gyrus (DG) region exhibit migration towards the granule cell layer. Conversely, stem cells residing in the SVZ demonstrate migration towards the olfactory bulb, following a predetermined pathway within the anterior horn of the lateral ventricle. During migration, these cells align to form a contiguous band resembling a chain enveloped by astrocytes.49 Notably, the migration process occurs within the astrocyte-formed sheath.
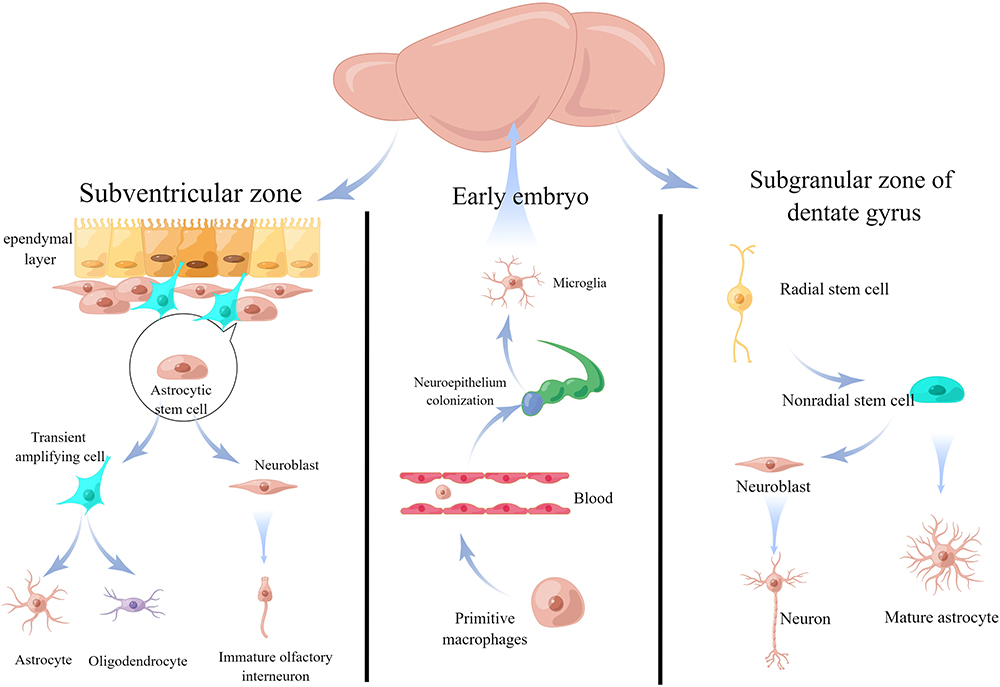 |
Figure 2 Adult neurogenesis/gliogenesis. In adults, neural stem cells are found in the subgranular zone of the dentate gyrus (SGZ/DG) in the hippocampus and the subventricular zone (SVZ) around the lateral ventricles. In the SVZ, astrocytic stem cells differentiate into early neurons and glial cells, allowing them to enter the olfactory bulb via the rostral migratory stream. Radial glial cells, or radial stem or progenitor cells, are the primary source of early neurons and glial cells in the SGZ/DG area. By Figdraw. |
The primary components of neural stem cell therapy for ischemic stroke encompass two fundamental features.50 Firstly, cell replacement involves the activation of endogenous NSCs within the body, typically found in a quiescent condition. In brain tissue damage cases, endogenous NSCs’ activation occurs, leading to their migration and localization at the injury site. Subsequently, these NSCs differentiate into neurons and glial cells, facilitating neural regeneration and repair. This reparative mechanism involves restoring damaged tissue and containing future expansion of the lesion.51 Nevertheless, the quantity of activated endogenous NSCs within the organism is constrained. Furthermore, many of these endogenous NSCs tend to develop into glial cells due to adverse microenvironmental conditions. This phenomenon contributes to the diminished capacity of the body to undergo self-repair following a stroke. Transplanting exogenous stem cells into the brain can fill the shortage of endogenous NSCs and stimulate nerve regeneration, rebuilding or repairing the brain-damaged area’s neural network.52 Secondly, the paracrine effect can be observed in the context of stem cell transplantation in the brain.
Since nerve fibers and blood vessels in the brain often travel concomitantly, transplanted stem cells can migrate along the blood vessels to the ischemia-associated areas and stimulate endogenous repair by secreting cytokines. Monocyte chemotactic protein 1 and chemokine stromal cell-derived factor 1 were found to be essential molecules for the homing of stem cells to ischemic regions.53,54 Transplanted stem cells can induce angiogenesis by releasing vascular endothelial growth factor, eliciting anti-inflammatory responses and other benefits.55 The comprehensive array of ways NSCs operate in managing ischemic stroke indicates that the restoration and healing of neural networks in the brain following such an event are facilitated through various means. Because of the low survival rate of transplanted stem cells, stem cell therapy acts primarily through time-limited paracrine mechanisms. Interestingly, Khan found that stem cell-derived sEVs provide beneficial therapeutic effects similar to those of stem cells and overcome the limitations of stem cell therapy.56
NSC-sEVs for Ischemic Stroke
sEVs range in size from 30–200 nm and are exocytotic vesicles secreted by most cells.57 Initially, they were believed to be waste particles released during the stripping of the plasma membrane.58 Both sEVs and microvesicles are vesicles surrounded by a lipid bilayer but do not have the same membrane composition due to their different origins.59 sEVs, crucial mediators in storing and transmitting cellular signals (Figure 3),60 can transfer their cargo to target cells through fusion with the recipient cell membrane. This process, therefore, bestows novel characteristics upon the recipient cells.
 |
Figure 3 Biogenesis and biological characteristics of sEVs. sEVs secretes and form a lipid bilayer that contains a range of transmembrane proteins like CD9, CD63, and CD81, as well as heat shock proteins like HSP90 and HSP70, all of which can perform various immunological activities. By Figdraw. |
Multiple studies have confirmed that sEVs significantly modulate cell connections within the neurovascular unit following an ischemic stroke.61 The neurovascular unit comprises several components, such as the BBB, glial cells, neurons, and extracellular matrix. The involvement of sEVs generated from neurovascular unit cells during ischemic events is of considerable importance in the modulation of neurological function impairment and subsequent repair processes. Stem cell-derived sEVs can also mediate communication by delivering nucleic acids, lipids and proteins between cells and tissues and influence neurovascular regeneration,62 inflammatory responses,63 and neuronal remodeling after cerebral ischemia by controlling gene expression and signaling pathways in recipient cells.64 Many studies have also confirmed the potential functions of NSC-sEVs.65,66 As illustrated in Figure 4,67–69 the administration of NSC-sEVs has been shown to facilitate the process of angiogenesis, neurogenesis, and axonal development while simultaneously suppressing inflammatory reactions. Consequently, this intervention has the potential to effectively reduce the restoration of standard functionality following an ischemic stroke. Extracting NSC-sEVs from cell cultures can pose considerable difficulties as neuroscience researchers navigate complex neural pathways. Common challenges encountered when extracting NSC-sEVs from cell culture include contamination (caused by the medium used for cell cultivation containing lipids, proteins, and other substances which may also interact with NSC-sEVs).70 Also, without standardized procedures for NSC-sEVs extraction from cell culture, discrepancies may arise between studies and compromise consistency.71 Further investigation is required to tackle the challenges associated with isolating the NSC-sEVs.
 |
Figure 4 NSC-sEVs for treating ischemic stroke. The specific processes encompass neuroprotection, inflammatory response suppression, endogenous nerve regeneration facilitation, and vascular regeneration stimulation. By Figdraw. |
Neuroprotective Effects of NSC-sEVs
Neurons play a crucial role in the brain’s functioning, and it has been shown that adults possess a restricted capacity for neuronal replacement.72 Numerous studies have shown that the severity of ischemic stroke depends mainly on the number of neurons dying in the region of brain injury.73,74 Furthermore, the demise of neurons within the ischemic core results in the release of pro-inflammatory and pro-apoptotic substances into the adjacent brain parenchyma, inducing neuronal death in the ischemic penumbra region. The point of view emphasizes the importance of cells’ functional and structural integrity in the penumbra region, as it is vital in limiting the size and extent of infarction. Therefore, it is essential for restoring damaged neurons.
Studies have shown that the lengths of dendrites and axons in neurons were reduced after experiencing oxygen-glucose deprivation/reperfusion. Additionally, there was a considerable reduction in the density of dendritic spines. However, it was observed that sEVs effectively counteracted these effects, reversing the trends mentioned above.75–78 The study conducted by Wang et al demonstrated that the administration of NSC-sEVs resulted in a considerable reduction in the presence of TUNEL-positive neurons subjected to oxidative stress.75 The Western blot analysis revealed a significant upregulation of Bcl-2 expression in the sEVs group, while the expression of Bax was observed to be downregulated in the same group. The findings suggest NSC-sEVs can impede neuronal death induced by oxidative stress damage and facilitate neuronal remodeling.
NSC-sEVs Inhibits the Activation of Glial Cells
Astrocytes represent the predominant glial cell population within the CNS. They fulfil various essential functions, including regulating extracellular ion concentrations, mitigating excitotoxicity, clearing amino acid neurotransmitters, releasing neurotransmitters, and facilitating synaptic development. These roles are critical for maintaining the integrity of the BBB and supporting neural activity.79,80 When the CNS is stimulated by injury, astrocytes will be in an activated state. Their number, morphology and biological function may change from resting astrocytes to reactive astrocytes. The activation of astrocytes into A1-responsive astrocytes, characterized by their capacity to generate significant quantities of inflammatory factors and neurotoxins, can be triggered by several stimuli, including inflammation and other factors. This transformation renders these astrocytes detrimental to surrounding nerve cells, leading to their demise. Promoting neuronal survival is facilitated by inhibiting A1-responsive astrocyte activation and suppressing inflammatory factor release.81 The study revealed that mouse NSC-sEVs exhibited a notable decrease in lactate dehydrogenase release from astrocytes when exposed to hypoxic settings. Additionally, NSC-sEVs reduced neuronal apoptosis and cerebral infarct volume.82
Microglia, a type of macrophage in the CNS, is crucial in maintaining the balance between cell growth and cell death by engulfing and eliminating dying neurons. This process is essential for the correct development of the CNS.83 Microglia activation is promptly initiated upon brain tissue injury, releasing substantial quantities of pro-inflammatory chemicals and cytotoxic substances. Consequently, microglia assume a significant role in the progression of secondary injury after neurotrauma and neuroinflammation. In their study, Gao et al found that NSC-sEVs can selectively accumulate in cerebral ischemia regions following intravenous treatment. Furthermore, they observed a significant reduction in the inflammatory response of microglia with exposure to NSC-sEVs, leading to decreased production of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6.84 Through high-throughput sequencing, it was discovered that seven microRNAs (miRNAs) present in NSC-sEVs can impede microglia activation. This inhibition is achieved by effectively regulating the mitogen-activated protein kinase (MAPK) pathway.
NSC-sEVs Facilitate NSC Differentiation
Neuronal death and lack of regeneration capacity are leading causes of limited recovery after cerebral ischemia. The critical determinant for influencing the restoration of nerve function after ischemic events is the proportion of neurons present within the differentiated cells derived from endogenous NSCs.85,86 Nevertheless, oxidative stress, hypoxia, and inflammation induce programmed cell death in endogenous NSCs and influence the transformation of NSCs into astrocytes.87 Consequently, this impacts the reorganization of the neural network following an episode of ischemia.88 The study conducted by Chen et al revealed that sEVs derived from NSCs subjected to modification by IFN-γ (IFN-γ-NSC-sEVs) exhibited the ability to enhance the differentiation of endogenous NSCs into neurons in rats with cerebral ischemia. Furthermore, these sEVs demonstrated the capacity to improve the survival of NSCs in the presence of oxidative stress while mitigating the inflammatory response.89 However, a comprehensive investigation of the mechanism of action of IFN-γ-NSC-sEVs has not been conducted.
NSC-sEVs Promotes Angiogenesis
The treatment of ischemic stroke requires both the regeneration of brain tissue and the activation of angiogenesis.90 The stress-induced damage of vascular endothelial cells has been observed to cause a decrease in blood supply, the cessation of local angiogenesis, and the disruption of nutrition supply, ultimately resulting in an unfavorable prognosis. The organism employs angiogenesis as a means to alleviate ischemic hemidiaphragm injury. Research has indicated a positive association between the quantity of neovascularization and patient survival. The blood vessel development and reconstruction process necessitates the involvement of endothelial cells, perivascular cells, and signaling factors produced by smooth muscle cells.91 The study determined that NSC-sEVs had a pro-angiogenic effect during cerebral ischemia, but the underlying mechanism remained inconclusive.68,92 In their study, Hu et al discovered that NSC-sEVs can enhance the migration and proliferation of vascular endothelial cells in mice with spinal cord injuries. Furthermore, the researchers revealed that VEGF-A facilitated this pro-angiogenic impact within the sEVs.93 VEGF, a highly selective pro-vascular endothelial cell growth factor, can enhance vascular permeability, degrade the extracellular matrix, facilitate migration and proliferation, and induce neovascularization in vascular endothelial cells. Subsequent experimental investigations revealed that the angiogenic effects of NSC-sEVs were reversed mainly by the down-regulation of VEGF-A.93
NSC-sEVs Enhances BBB Integrity
The BBB is a crucial component of cerebral microcirculation, composed of endothelial cells of the capillary wall, astrocyte ends wrapping capillaries, and pericytes embedded in the capillary basement membrane.94 Under normal conditions, the BBB isolates the CNS from peripheral blood circulation, effectively restricting and regulating molecular, ionic, and cellular exchanges between the blood and the CNS, making it difficult for most molecules, viruses, inflammatory factors, and other harmful components to pass through the BBB, thus effectively maintaining CNS homeostasis.95,96 As a critical component of the BBB, brain endothelial cells have specific transport mechanisms for luminal/extraluminal polarization, tight junctions, attachment of adhesion molecules, and restriction of polar substances, which efficiently maintain the stability of the BBB. ABCB1 belongs to the ATP-binding cassette family of transporter proteins, which are responsible for actively expelling substrates from cells upon ATP energy stimulation.97 The expression of ABCB1 is observed to be increased in brain endothelial cells three hours following the generation of stroke. This upregulation may be a normal reaction to removing harmful compounds within the cells. Nevertheless, unfortunately, this response also leads to the transfer of neuroprotective molecules and inflammatory chemokines.98 Prior research has indicated that the suppression of ABCB1 leads to a decrease in the size of infarcts in mice subjected to experimental stroke scenarios.99 The study conducted by Thorsten et al showed that NSC-sEVs effectively reduced the overexpression of ABCB1 by blocking the activation of the NF-κB pathway in endothelial cells.100 Astrocytes that have been activated following a stroke play a significant role in the production of matrix metalloproteinases, which subsequently result in the degradation of the basement membrane and the breakdown of the BBB. However, the administration of NSC-sEVs demonstrated a notable suppression of MMP-9 activation, a protein that exhibited widespread upregulation in several brain areas 24 hours post-stroke. This effect ultimately resulted in the reinforcement of BBB integrity in a rodent stroke model.100
Using Engineered sEVs for Treating Ischemic Stroke
As research progresses, the limitations of utilizing natural sEVs for treating ischemic stroke are becoming increasingly evident, specifically in their inadequate targeting capabilities and content loading.101,102 Several approaches have been devised to rectify these limitations and enhance the targeting, stability, and loading capacity of NSC-sEVs. These modifications together fall under the category of engineered NSC-sEVs.103
Targeting
The effective delivery of medications to brain tissue for treating ischemic stroke is still a challenging task despite the advancements made in this area. One of the fundamental difficulties is in reaching optimal drug concentration in the brain while also possessing qualities like the ability to pass the BBB and low toxicity. The BBB presents a significant impediment to drug therapy, as the ability of medications to effectively traverse this barrier and exert their desired therapeutic effects is limited.104 sEVs can traverse the BBB and exhibit potential as nanoscale carriers for neurological medication administration, characterized by their low toxicity and good biocompatibility.105 Currently, the primary method of delivering natural sEVs is through intravenous injection. Nevertheless, following intravenous delivery, the primary distribution of endogenous sEVs occurs in peripheral organs, including the liver, spleen, kidney, lungs, and gastrointestinal system. This distribution pattern ultimately diminishes the effectiveness of the treatment.106,107 Furthermore, empirical research has demonstrated that sEVs possess specific signaling molecules capable of binding to certain cellular targets. The interplay between receptors and ligands facilitates target cells’ selective internalization of sEVs, promoting cellular uptake with a high specificity.108 There is a pressing demand to engineer sEVs to achieve maximum efficiency and efficacy in drug delivery targeting.
The specificity of sEVs targeting can be achieved by employing various technical methods for extracellular vesicle surface engineering, primarily through genetic and chemical modification. Genetic engineering involves using plasmid vectors and other encoded proteins or peptides to bind to extracellular vesicle membrane proteins, such as glycophosphatidylinositol and lysosome-associated membrane protein.109,110 Chemical modification mainly relies on the bioconjugation of targeted ligands to surface proteins to modify the surface of sEVs (Figure 5).111 An instance of this can be observed in the case of integrin αvβ3, which is absent in the vasculature of normal tissues but exhibits significant expression on endothelial cells during instances of pathological angiogenesis, such as vascular regeneration following ischemic tissues and the neovascularization of tumors.112,113 Consequently, this integrin can serve as a viable target for delivering therapeutic agents. The Arg-Gly-Asp (RGD) peptide functions as a ligand that specifically binds to the integrin αvβ3 receptor with high affinity.114 In their study, Gao et al modified the RGD peptide on the membrane of NSC-sEVs and injected it into cerebral ischemic rats via the tail vein.107 The study revealed that the sEVs treated with peptides exhibited notable accumulation in the area of ischemic brain lesions, indicating that this targeting capability is attributed to the high affinity between the RGD peptide and integrin αvβ3.84
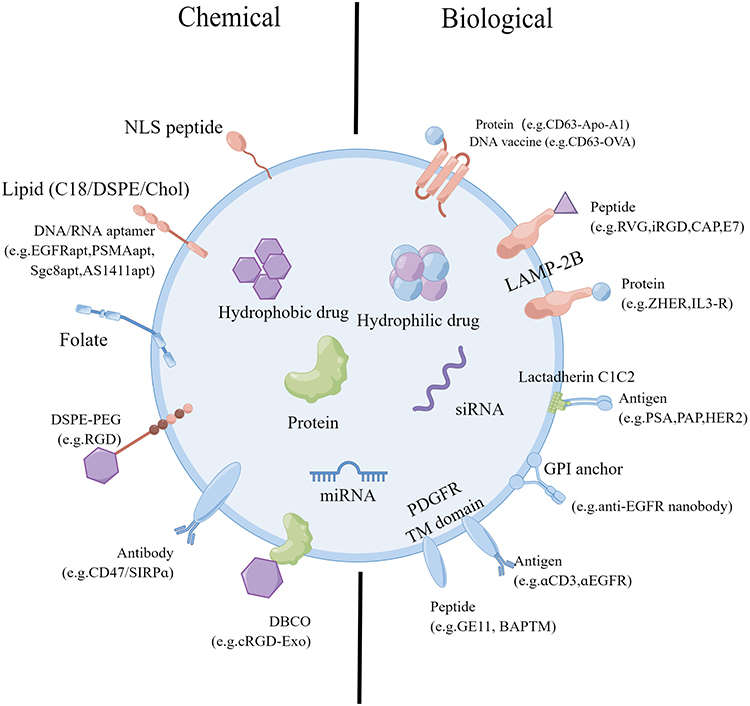 |
Figure 5 The genetic/biological or chemical manipulation of sEVs for surface engineering. Chemical modifications using lipids, membrane-bound proteins, or lipid-lipid interactions introduce peptides, proteins, small molecules, lipids, and polymers. The biological fusion of membrane-bound proteins introduces targeting elements like peptides and proteins. By Figdraw. |
Content Carrying
Pharmacological intervention constitutes a crucial component of the therapeutic regimen for ischemic stroke.115,116 Nevertheless, traditional pharmacological treatment frequently encounters challenges such as limited cellular penetration, inadequate aqueous solubility, and dispersion within the organism. These issues may result in an escalation in drug dosage, constraining the effectiveness of the medication and augmenting the probability of adverse reactions. Therefore, alongside the further advancement of pharmaceuticals, implementing a rational drug delivery system holds the potential to facilitate the efficient and secure transportation of medications to the specific site of injury. sEVs can serve as a novel drug delivery vehicle cohort due to their diminished immunogenicity, elevated biocompatibility, and sustained circulation within the vascular system.117 To obtain engineered sEVs containing target genes, drugs, and other substances, drug loading procedures are mainly achieved by electroporation, incubation methods, and chemical conjugation methods (Figure 6).118
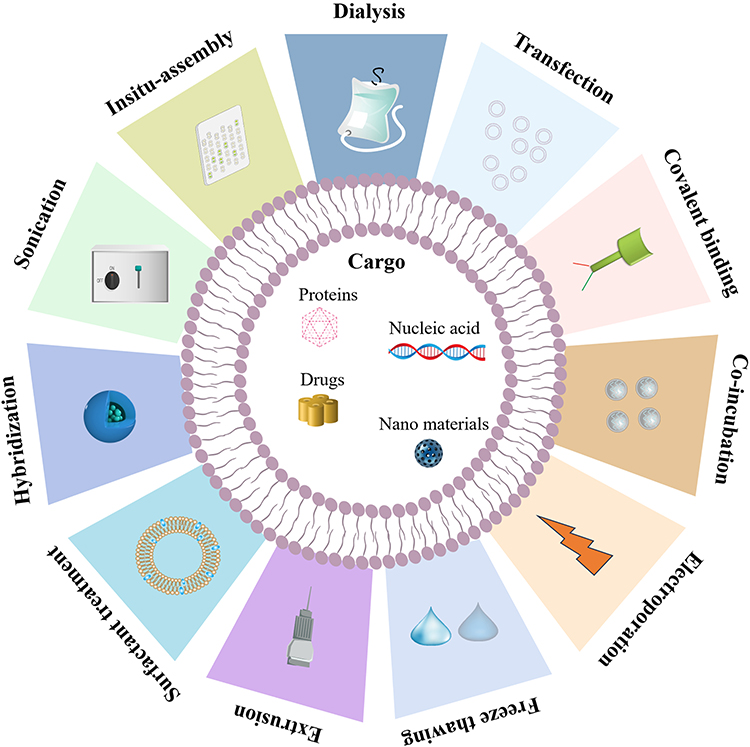 |
Figure 6 The utilization of physical treatment modalities for enhancing the therapeutic effectiveness of sEVs. Physical treatments load cargo into sEVs. Using surfactant, sonication, and electroporation to create exosomal membrane pores enhances this process. Extrusion, freeze-thaw treatment, and dialysis increase cargo loading in membrane recombination. By Figdraw. |
Neurotrophic factors are a group of proteins essential for the nervous system’s development, growth, survival, and differentiation.119 BDNF, the most abundant and widely distributed neurotrophic factor in the nervous system, promotes cell differentiation, synaptogenesis, and neurogenesis in the nervous system through stimulation of its pro-myosin receptor kinase B and exhibits many neuroprotective properties in post-ischemic and traumatic brain injury.120 It has been shown that BDNF promotes the proliferation of endogenous NSCs and regeneration of neural axons in spinal cord injured rats. However, the effectiveness of administering these neurotrophic factors from an external source is constrained by their relatively brief duration of action, limited capacity to cross the blood-brain barrier, and rapid degradation.121,122 Chen et al successfully generated BDNF-NSC-sEVs by co-incubation and transfection to introduce BDNF into NSC-sEVs. Their investigation revealed that BDNF-NSC-sEVs can enhance the differentiation of hypoxic NSCs into neurons. Additionally, BDNF-NSC-sEVs showed the capacity to establish a suitable immune microenvironment for neural regeneration by decreasing inflammatory responses.67 This approach effectively produces a synergistic therapeutic outcome by augmentation of neurotrophic factors’ bioavailability and consolidating Exo’s inherent therapeutic impact.
Benefits of Imaging-Based NSC-sEVs Tracking
Tracking the behavior of NSC-sEVs in living organisms presents a considerable challenge. One reason is the small size of sEVs, which allows them to disperse quickly in body fluids. Additionally, their composition is similar to that of body cells, making it difficult to distinguish them using imaging techniques.123 To gain a deeper understanding of the functionality, viability, and trafficking of these vesicles within the body, it is essential to have a dependable and noninvasive NSC-sEVs in vivo imaging technique.124 Various imaging technologies are currently available, each with its unique capabilities. These include optical (bioluminescent imaging (BLI) and fluorescence imaging are its primary forms), tomographical (CT and MRI), nuclear imaging modalities (Radioactive substances), and photoacoustic imaging.125,126 Imaging technologies provide numerous advantages, including high sensitivity, perfect spatiotemporal precision, the ability to penetrate deep into tissues, superior resolution, and remarkable tissue contrasts.127 NSC-sEVs have a wide range of therapeutic potential, and numerous tracking methods have been designed to investigate how they work. Further research will likely give a more comprehensive understanding of their capabilities and limitations.
The Future Prospective
Numerous studies have substantiated the efficacy of NSC-sEVs in the treatment of ischemic stroke. Its primary mechanisms involve the inhibition of neuroinflammation, mitigation of apoptosis, and facilitation of neurovascular regeneration. Consequently, this therapeutic approach presents a novel avenue for managing neurological disorders, including stroke. However, it is imperative to acknowledge specific possible challenges associated with NSC-sEVs-based therapeutic approaches that necessitate careful consideration and resolution.
sEVs possess distinct proteins and genetic material exclusive to the originating cell or tissue, indicating their cellular origin and physiological condition.128,129 Therefore, sEVs released by distinct cellular entities exhibit distinct attributes and functionalities. The generation, surface protein expression and content composition of sEVs are influenced by cell development conditions and the culture medium formulation.130 Maintaining consistent cell growth ensures extracellular vesicle stability, uniformity, and efficacy. Furthermore, it should be noted that storage conditions might influence the size distribution of sEVs and the stability and biodistribution of cells. Previous research has demonstrated that sEVs, when stored in individual doses at −80°C, maintain their functional activity for seven months.131 The variables above possess the potential to exert a significant influence on the efficacy of the clinical-grade NSC-sEVs and will serve as crucial determinants in their production.
The composition and function of exosomal subtypes exhibit variability, which has implications for various applications. One notable challenge is the inability to effectively label and identify specific exosomal subtypes. To conduct thorough biochemical investigations, such as identifying bioactive compounds and therapeutic screening, it is imperative to employ stringent purification and grading procedures for purified sEVs. In addition, dosing remains a significant challenge when delivering NSC-sEVs-based therapies, with factors such as dose selection, assessment, and administration (route, frequency, time window) influencing the outcome. There is a need to create potency assays that utilize appropriate functional endpoints to determine in vitro and in vivo efficacy accurately. The efficacy assessments of sEVs rely on standardizing extracellular vesicle samples and the specific disease under investigation.
In treating ischemic stroke, stem cell-derived sEVs, especially NSC-sEVs, can establish an immune microenvironment conducive to recovery, suppress neuronal apoptosis and neuroinflammation, and actively engage in the endogenous tissue repair mechanism. Consequently, these sEVs are crucial in restoring neurological function after a stroke. Despite the existing limitations and challenges in the current implementation of NSC-sEVs, their potential for use in refractory neurological conditions such as ischemic stroke remains promising. Hence, it is anticipated that novel therapeutic approaches utilizing NSC-sEVs will be crucial for future clinical implementation.
Abbreviations
NSCs, Neural stem cells; NSC-sEVs, Neural stem cell-derived small extracellular vesicles; sEVs, Small extracellular vesicles; BBB, Blood-brain barrier; tPA, Tissue-type plasminogen activator; MSCs, Mesenchymal stem cells; BDNF, Brain-derived neurotrophic factor; VEGF, Vascular endothelial growth factor; SVZ, Subventricular zone; SGZ, Subgranular zone; DG, Dentate gyrus; CNS, Central nervous system; miRNAs, microRNAs; MAPK, Mitogen-activated protein kinase; RGD, Arg-Gly-Asp.
Acknowledgments
We thank Fig.Draw (https://www.figdraw.com/) for editing figures.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was funded by the National Key R&D Program of China (2022YFA1104900 & 2022YFA1104904), the National Natural Science Foundation of China (82171459), the Department of Education of Guangdong Province (2021ZDZX2011).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Wang Z, Zhao Y, Hou Y, et al. A thrombin-activated peptide-templated nanozyme for remedying ischemic stroke via thrombolytic and neuroprotective actions. Adv Mater. 2024;36(10):e2210144. doi:10.1002/adma.202210144
2. Zhang GL, Zhu ZH, Wang YZ. Neural stem cell transplantation therapy for brain ischemic stroke: review and perspectives. World J Stem Cells. 2019;11(10):817–830. doi:10.4252/wjsc.v11.i10.817
3. Tang YN, Zhang GF, Chen HL, et al. Selective brain hypothermia-induced neuroprotection against focal cerebral ischemia/reperfusion injury is associated with Fis1 inhibition. Neural Regen Res. 2020;15(5):903–911. doi:10.4103/1673-5374.268973
4. Fukuta T, Asai T, Yanagida Y, et al. Combination therapy with liposomal neuroprotectants and tissue plasminogen activator for treatment of ischemic stroke. FASEB j. 2017;31(5):1879–1890. doi:10.1096/fj.201601209R
5. Yang Y, Fan Y, Zhang H, et al. Small molecules combined with collagen hydrogel direct neurogenesis and migration of neural stem cells after spinal cord injury. Biomaterials. 2021;269:120479. doi:10.1016/j.biomaterials.2020.120479
6. Hosseini SM, Borys B, Karimi-Abdolrezaee S. Neural stem cell therapies for spinal cord injury repair: an update on recent preclinical and clinical advances. Brain. 2024;147(3):766–793. doi:10.1093/brain/awad392
7. Luo J, Feng Y, Hong Z, et al. High-frequency repetitive transcranial magnetic stimulation promotes neural stem cell proliferation after ischemic stroke. Neural Regen Res. 2024;19(8):1772–1780. doi:10.4103/1673-5374.389303
8. Wang X, Zhou R, Xiong Y, et al. Sequential fate-switches in stem-like cells drive the tumorigenic trajectory from human neural stem cells to malignant glioma. Cell Res. 2021;31(6):684–702. doi:10.1038/s41422-020-00451-z
9. Hejrati N, Wong R, Khazaei M, et al. How can clinical safety and efficacy concerns in stem cell therapy for spinal cord injury be overcome? Expert Opin Biol Ther. 2023;23:1–17. doi:10.1080/14712598.2023.2245321
10. Qin T, Li C, Xu Y, et al. Local delivery of EGFR(+)NSCs-derived exosomes promotes neural regeneration post spinal cord injury via miR-34a-5p/HDAC6 pathway. Bioact Mater. 2024;33:424–443. doi:10.1016/j.bioactmat.2023.11.013
11. Ahmed W, Huang S, Chen L. Engineered exosomes derived from stem cells: a new brain-targeted strategy. Expert Opin Drug Deliv. 2024;21(1):91–110. doi:10.1080/17425247.2024.2306877
12. Zhou X, Jia Y, Mao C, et al. Small extracellular vesicles: non-negligible vesicles in tumor progression, diagnosis, and therapy. Cancer Lett. 2024;580:216481. doi:10.1016/j.canlet.2023.216481
13. Arya SB, Collie SP, Parent CA. The ins-and-outs of exosome biogenesis, secretion, and internalization. Trends Cell Biol. 2024;34(2):90–108. doi:10.1016/j.tcb.2023.06.006
14. Spellicy SE, Kaiser EE, Bowler MM, et al. Neural stem cell extracellular vesicles disrupt midline shift predictive outcomes in porcine ischemic stroke model. Transl Stroke Res. 2020;11(4):776–788. doi:10.1007/s12975-019-00753-4
15. Ceulemans A, Spronk HMH, Ten Cate H, et al. Current and potentially novel antithrombotic treatment in acute ischemic stroke. Thromb Res. 2024;236:74–84. doi:10.1016/j.thromres.2024.02.009
16. Fitzmaurice C, Abate D, Abbasi N, et al. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 29 cancer groups, 1990 to 2017: a systematic analysis for the global burden of disease study. JAMA Oncol. 2019;5(12):1749–1768. doi:10.1001/jamaoncol.2019.2996
17. Yang S, Qin C, Chen M, et al. TREM2-IGF1 Mediated Glucometabolic Enhancement Underlies Microglial Neuroprotective Properties During Ischemic Stroke. Adv Sci. 2024;11(10):e2305614. doi:10.1002/advs.202305614
18. Wang Z, Zhang S, Cheng R, et al. Knockdown of RGMA improves ischemic stroke via Reprogramming of Neuronal Metabolism. Free Radic Biol Med. 2024:41–56. doi:10.1016/j.freeradbiomed.2024.03.020
19. Calabrese B, Jones SL, Shiraishi-Yamaguchi Y, et al. INF2-mediated actin filament reorganization confers intrinsic resilience to neuronal ischemic injury. Nat Commun. 2022;13(1):6037. doi:10.1038/s41467-022-33268-y
20. Zong P, Feng J, Yue Z, et al. Functional coupling of TRPM2 and extrasynaptic NMDARs exacerbates excitotoxicity in ischemic brain injury. Neuron. 2022;110(12):1944–58.e8. doi:10.1016/j.neuron.2022.03.021
21. Howell JA, Gaouette N, Lopez M, et al. Elastin-like polypeptide delivery of anti-inflammatory peptides to the brain following ischemic stroke. FASEB j. 2023;37(8):e23077. doi:10.1096/fj.202300309RR
22. Yang L, Qian J, Yang B, et al. Challenges and improvements of novel therapies for ischemic stroke. Front Pharmacol. 2021;12:721156. doi:10.3389/fphar.2021.721156
23. Arkelius K, Wendt TS, Andersson H, et al. LOX-1 and MMP-9 inhibition attenuates the detrimental effects of delayed rt-PA therapy and improves outcomes after acute ischemic stroke. Circ Res. 2024;134:954–969. doi:10.1161/circresaha.123.323371
24. Tsivgoulis G, Katsanos AH, Sandset EC, et al. Thrombolysis for acute ischaemic stroke: current status and future perspectives. Lancet Neurol. 2023;22(5):418–429. doi:10.1016/s1474-4422(22)00519-1
25. Haupt M, Gerner ST, Bähr M, et al. Neuroprotective Strategies for Ischemic Stroke-Future Perspectives. Int J Mol Sci. 2023;24(5):4334. doi:10.3390/ijms24054334
26. Huang G, Zang J, He L, et al. Bioactive nanoenzyme reverses oxidative damage and endoplasmic reticulum stress in neurons under ischemic stroke. ACS Nano. 2022;16(1):431–452. doi:10.1021/acsnano.1c07205
27. Dergunova LV, Filippenkov IB, Limborska SA, et al. Neuroprotective peptides and new strategies for ischemic stroke drug discoveries. Genes. 2023;14(5):953. doi:10.3390/genes14050953
28. Ding S, Chen Q, Chen H, et al. The neuroprotective role of neuroserpin in ischemic and hemorrhagic stroke. Curr Neuropharmacol. 2021;19(8):1367–1378. doi:10.2174/1570159x18666201008113052
29. Eren F, Yilmaz SE. Neuroprotective approach in acute ischemic stroke: a systematic review of clinical and experimental studies. Brain Circ. 2022;8(4):172–179. doi:10.4103/bc.bc_52_22
30. Guo Y, Peng Y, Zeng H, et al. Progress in mesenchymal stem cell therapy for ischemic stroke. Stem Cells Int. 2021;2021:9923566. doi:10.1155/2021/9923566
31. Lu X, Lv C, Zhao Y, et al. TSG-6 released from adipose stem cells-derived small extracellular vesicle protects against spinal cord ischemia reperfusion injury by inhibiting endoplasmic reticulum stress. Stem Cell Res Ther. 2022;13(1):291. doi:10.1186/s13287-022-02963-4
32. Wang M, Zhao N, Wang C, et al. Immunomodulatory properties of mesenchymal stem cells: a potential therapeutic strategy for allergic rhinitis. Allergy. 2023;78(6):1425–1440. doi:10.1111/all.15729
33. Zipser CM, Cragg JJ, Guest JD, et al. Cell-based and stem-cell-based treatments for spinal cord injury: evidence from clinical trials. Lancet Neurol. 2022;21(7):659–670. doi:10.1016/s1474-4422(21)00464-6
34. Kawada-Horitani E, Kita S, Okita T, et al. Human adipose-derived mesenchymal stem cells prevent type 1 diabetes induced by immune checkpoint blockade. Diabetologia. 2022;65(7):1185–1197. doi:10.1007/s00125-022-05708-3
35. Lian M, Qiao Z, Qiao S, et al. Nerve growth factor-preconditioned mesenchymal stem cell-derived exosome-functionalized 3D-printed hierarchical porous scaffolds with neuro-promotive properties for enhancing innervated bone regeneration. ACS Nano. 2024;18(10):7504–7520. doi:10.1021/acsnano.3c11890
36. Lin Z, Wu Y, Xu Y, et al. Mesenchymal stem cell-derived exosomes in cancer therapy resistance: recent advances and therapeutic potential. Mol Cancer. 2022;21(1):179. doi:10.1186/s12943-022-01650-5
37. Lan T, Luo M, Wei X. Mesenchymal stem/stromal cells in cancer therapy. J Hematol Oncol. 2021;14(1):195. doi:10.1186/s13045-021-01208-w
38. Li Z, Dong X, Tian M, et al. Stem cell-based therapies for ischemic stroke: a systematic review and meta-analysis of clinical trials. Stem Cell Res Ther. 2020;11(1):252. doi:10.1186/s13287-020-01762-z
39. Toman NG, Grande AW, Low WC. Neural repair in stroke. Cell Transplant. 2019;28(9–10):1123–1126. doi:10.1177/0963689719863784
40. Jiang F, Zhou H, Cheng Y, et al. Various detailed characteristics of a new enhanced neurotrophic factor secreting rat derived bone marrow mesenchymal stem cells and its preliminary application in rat models of ischemic stroke. Exp Cell Res. 2022;416(1):113140. doi:10.1016/j.yexcr.2022.113140
41. Brown J, Park YJ, Lee JY, et al. Bone marrow-derived NCS-01 cells advance a novel cell-based therapy for stroke. Int J Mol Sci. 2020;21(8):2845. doi:10.3390/ijms21082845
42. Rao J, Li H, Zhang H, et al. Periplaneta Americana (L.) extract activates the ERK/CREB/BDNF pathway to promote post-stroke neuroregeneration and recovery of neurological functions in rats. J Ethnopharmacol. 2024;321:117400. doi:10.1016/j.jep.2023.117400
43. Liao Y, Ming J, Song W, et al. Mitochondrial transplantation and immune response of human bone marrow mesenchymal stem cells for the therapeutic of ischemic stroke. Curr Stem Cell Res Ther. 2024;19(5):678–687. doi:10.2174/1574888x18666230505103407
44. Wang S, He Q, Qu Y, et al. Emerging strategies for nerve repair and regeneration in ischemic stroke: neural stem cell therapy. Neural Regen Res. 2024;19(11):2430–2443. doi:10.4103/1673-5374.391313
45. Li MY, Yang XL, Chung CC, et al. TRIP6 promotes neural stem cell maintenance through YAP-mediated Sonic Hedgehog activation. FASEB j. 2024;38(5):e23501. doi:10.1096/fj.202301805RRR
46. Reynolds BA, Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science. 1992;255(5052):1707–1710. doi:10.1126/science.1553558
47. Woo K, Fraser SE. Specification of the zebrafish nervous system by nonaxial signals. Science. 1997;277(5323):254–257. doi:10.1126/science.277.5323.254
48. Su P, Zhang J, Zhao F, et al. The interaction between microglia and neural stem/precursor cells. Brain Res Bull. 2014;109:32–38. doi:10.1016/j.brainresbull.2014.09.005
49. Lu J, Manaenko A, Hu Q. Targeting adult neurogenesis for poststroke therapy. Stem Cells Int. 2017;2017:5868632. doi:10.1155/2017/5868632
50. Huang L, Zhang L. Neural stem cell therapies and hypoxic-ischemic brain injury. Prog Neurobiol. 2019;173:1–17. doi:10.1016/j.pneurobio.2018.05.004
51. Deshpande SS, Malik SC, Conforti P, et al. P75 neurotrophin receptor controls subventricular zone neural stem cell migration after stroke. Cell Tissue Res. 2022;387(3):415–431. doi:10.1007/s00441-021-03539-z
52. Ha GH, Kim EJ, Park JS, et al. JAK2/STAT3 pathway mediates neuroprotective and pro-angiogenic treatment effects of adult human neural stem cells in middle cerebral artery occlusion stroke animal models. Aging (Albany NY). 2022;14(22):8944–8969. doi:10.18632/aging.204410
53. Wilson KL, Joseph NI, Onweller LA, et al. SDF-1 bound heparin nanoparticles recruit progenitor cells for their differentiation and promotion of angiogenesis after stroke. Adv Healthc Mater. 2023:e2302081. doi:10.1002/adhm.202302081
54. Zheng H, Haroon K, Liu M, et al. Monomeric CXCL12-engineered adipose-derived stem cells transplantation for the treatment of ischemic stroke. Int J Mol Sci. 2024;25(2):792. doi:10.3390/ijms25020792
55. Tian H, Yang X, Zhao J, et al. Hypoxia-preconditioned bone marrow mesenchymal stem cells improved cerebral collateral circulation and stroke outcome in mice. Arterioscler Thromb Vasc Biol. 2023;43(7):1281–1294. doi:10.1161/atvbaha.122.318559
56. Khan M, Nickoloff E, Abramova T, et al. Embryonic stem cell-derived exosomes promote endogenous repair mechanisms and enhance cardiac function following myocardial infarction. Circ Res. 2015;117(1):52–64. doi:10.1161/circresaha.117.305990
57. Ghanam J, Chetty VK, Zhu X, et al. Single molecule localization microscopy for studying small extracellular vesicles. Small. 2023;19(12):e2205030. doi:10.1002/smll.202205030
58. Couch Y, Buzàs EI, Di Vizio D, et al. A brief history of nearly EV-erything - The rise and rise of extracellular vesicles. J Extracell Vesicles. 2021;10(14):e12144. doi:10.1002/jev2.12144
59. Ramalhete L, Araújo R, Ferreira A, et al. Exosomes and microvesicles in kidney transplantation: the long road from trash to gold. Pathology. 2024;56(1):1–10. doi:10.1016/j.pathol.2023.10.004
60. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
61. Zagrean AM, Hermann DM, Opris I, et al. Multicellular crosstalk between exosomes and the neurovascular unit after cerebral ischemia. Therap Implicati Front Neurosci. 2018;12:811. doi:10.3389/fnins.2018.00811
62. Hu H, Hu X, Li L, et al. Exosomes derived from bone marrow mesenchymal stem cells promote angiogenesis in ischemic stroke mice via upregulation of MiR-21-5p. Biomolecules. 2022;12(7):883. doi:10.3390/biom12070883
63. Zhang Z, Zou X, Zhang R, et al. Human umbilical cord mesenchymal stem cell-derived exosomal miR-146a-5p reduces microglial-mediated neuroinflammation via suppression of the IRAK1/TRAF6 signaling pathway after ischemic stroke. Aging (Albany NY). 2021;13(2):3060–3079. doi:10.18632/aging.202466
64. Dumbrava DA, Surugiu R, Börger V, et al. Mesenchymal stromal cell-derived small extracellular vesicles promote neurological recovery and brain remodeling after distal middle cerebral artery occlusion in aged rats. Geroscience. 2022;44(1):293–310. doi:10.1007/s11357-021-00483-2
65. Zhong L, Wang J, Wang P, et al. Neural stem cell-derived exosomes and regeneration: cell-free therapeutic strategies for traumatic brain injury. Stem Cell Res Ther. 2023;14(1):198. doi:10.1186/s13287-023-03409-1
66. Vogel AD, Upadhya R, Shetty AK. Neural stem cell derived extracellular vesicles: attributes and prospects for treating neurodegenerative disorders. EBioMedicine. 2018;38:273–282. doi:10.1016/j.ebiom.2018.11.026
67. Zhu ZH, Jia F, Ahmed W, et al. Neural stem cell-derived exosome as a nano-sized carrier for BDNF delivery to a rat model of ischemic stroke. Neural Regen Res. 2023;18(2):404–409. doi:10.4103/1673-5374.346466
68. Gu C, Li Y, Liu J, et al. Neural stem cell-derived exosomes-loaded adhesive hydrogel controlled-release promotes cerebral angiogenesis and neurological function in ischemic stroke. Exp Neurol. 2023;370:114547. doi:10.1016/j.expneurol.2023.114547
69. Li Z, Chen Z, Peng J. Neural stem cell-derived exosomal FTO protects neuron from microglial inflammatory injury by inhibiting microglia NRF2 mRNA m6A modification. J Neurog. 2023;37(3):103–114. doi:10.1080/01677063.2023.2259995
70. Haroon K, Ruan H, Zheng H, et al. Bio-clickable, small extracellular vesicles-COCKTAIL therapy for ischemic stroke. J Control Release. 2023;363:585–596. doi:10.1016/j.jconrel.2023.10.003
71. Haroon K, Zheng H, Wu S, et al. Engineered exosomes mediated targeted delivery of neuroprotective peptide NR2B9c for the treatment of traumatic brain injury. Int J Pharm. 2024;649:123656. doi:10.1016/j.ijpharm.2023.123656
72. Zhao J, Liu S, Xiang X, et al. Versatile strategies for adult neurogenesis: avenues to repair the injured brain. Neural Regen Res. 2024;19(4):774–780. doi:10.4103/1673-5374.382224
73. Jiang J, Qi T, Li L, et al. MRPS9-mediated regulation of the PI3K/Akt/mTOR pathway inhibits neuron apoptosis and protects ischemic stroke. J Mol Neurosci. 2024;74(1):23. doi:10.1007/s12031-024-02197-4
74. Long J, Sun Y, Liu S, et al. Ginsenoside Rg1 treats ischemic stroke by regulating CKLF1/CCR5 axis-induced neuronal cell pyroptosis. Phytomedicine. 2024;123:155238. doi:10.1016/j.phymed.2023.155238
75. Liu Q, Tan Y, Qu T, et al. Therapeutic mechanism of human neural stem cell-derived extracellular vesicles against hypoxia-reperfusion injury in vitro. Life Sci. 2020;254:117772. doi:10.1016/j.lfs.2020.117772
76. Peng J, He J, Lin L, et al. Neural stem cell extracellular vesicles carrying YBX1 inhibited neuronal pyroptosis through increasing m6A-modified GPR30 stability and expression in ischemic stroke. Transl Stroke Res. 2023. doi:10.1007/s12975-023-01210-z
77. Mahdavipour M, Hassanzadeh G, Seifali E, et al. Effects of neural stem cell-derived extracellular vesicles on neuronal protection and functional recovery in the rat model of middle cerebral artery occlusion. Cell Biochem Funct. 2020;38(4):373–383. doi:10.1002/cbf.3484
78. Xu X, Zhang H, Li J, et al. Combination of EPC-EXs and NPC-EXs with miR-126 and miR-210 overexpression produces better therapeutic effects on ischemic stroke by protecting neurons through the Nox2/ROS and BDNF/TrkB pathways. Exp Neurol. 2023;359:114235. doi:10.1016/j.expneurol.2022.114235
79. Zhou X, Zhu Y, Gao D, et al. Matrilin-3 supports neuroprotection in ischemic stroke by suppressing astrocyte-mediated neuroinflammation. Cell Rep. 2024;43(4):113980. doi:10.1016/j.celrep.2024.113980
80. Lu W, Wen J. Crosstalk among glial cells in the blood-brain barrier injury after ischemic stroke. Mol Neurobiol. 2024. doi:10.1007/s12035-024-03939-6
81. Liu W, Wang Y, Gong F, et al. Exosomes derived from bone mesenchymal stem cells repair traumatic spinal cord injury by suppressing the activation of A1 neurotoxic reactive astrocytes. J Neurotrauma. 2019;36(3):469–484. doi:10.1089/neu.2018.5835
82. Sun X, Jung JH, Arvola O, et al. Stem cell-derived exosomes protect astrocyte cultures from in vitro ischemia and decrease injury as post-stroke intravenous therapy. Front Cell Neurosci. 2019;13:394. doi:10.3389/fncel.2019.00394
83. Planas AM. Role of microglia in stroke. Glia. 2024;72:1016–1053. doi:10.1002/glia.24501
84. Tian T, Cao L, He C, et al. Targeted delivery of neural progenitor cell-derived extracellular vesicles for anti-inflammation after cerebral ischemia. Theranostics. 2021;11(13):6507–6521. doi:10.7150/thno.56367
85. Tang H, Li Y, Tang W, et al. Endogenous neural stem cell-induced neurogenesis after ischemic stroke: processes for brain repair and perspectives. Transl Stroke Res. 2023;14(3):297–303. doi:10.1007/s12975-022-01078-5
86. Duan H, Li S, Hao P, et al. Activation of endogenous neurogenesis and angiogenesis by basic fibroblast growth factor-chitosan gel in an adult rat model of ischemic stroke. Neural Regen Res. 2024;19(2):409–415. doi:10.4103/1673-5374.375344
87. Gan L, Liao S, Tong Y, et al. Long noncoding RNA H19 mediates neural stem/progenitor cells proliferation, differentiation and apoptosis through the p53 signaling pathway after ischemic stroke. Biochem Biophys Res Commun. 2022;597:8–15. doi:10.1016/j.bbrc.2022.01.095
88. Zhang K, Yang Y, Ge H, et al. Neurogenesis and proliferation of neural stem/progenitor cells conferred by artesunate via FOXO3a/p27Kip1 axis in mouse stroke model. Mol Neurobiol. 2022;59(8):4718–4729. doi:10.1007/s12035-021-02710-5
89. Zhang G, Zhu Z, Wang H, et al. Exosomes derived from human neural stem cells stimulated by interferon gamma improve therapeutic ability in ischemic stroke model. J Adv Res. 2020;24:435–445. doi:10.1016/j.jare.2020.05.017
90. Fang J, Wang Z, Miao CY. Angiogenesis after ischemic stroke. Acta Pharmacol Sin. 2023;44(7):1305–1321. doi:10.1038/s41401-023-01061-2
91. Hatakeyama M, Ninomiya I, Kanazawa M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen Res. 2020;15(1):16–19. doi:10.4103/1673-5374.264442
92. Li R, Duan W, Feng T, et al. Lycium barbarum polysaccharide inhibits ischemia-induced autophagy by promoting the biogenesis of neural stem cells-derived extracellular vesicles to enhance the delivery of miR-133a-3p. Chin Med. 2023;18(1):117. doi:10.1186/s13020-023-00831-8
93. Zhong D, Cao Y, Li CJ, et al. Neural stem cell-derived exosomes facilitate spinal cord functional recovery after injury by promoting angiogenesis. Exp Biol Med. 2020;245(1):54–65. doi:10.1177/1535370219895491
94. Candelario-Jalil E, Dijkhuizen RM, Magnus T. Neuroinflammation, stroke, blood-brain barrier dysfunction, and imaging modalities. Stroke. 2022;53(5):1473–1486. doi:10.1161/strokeaha.122.036946
95. Xu N, Jiang X, Zhang W, et al. Endothelial peroxiredoxin-4 is indispensable for blood-brain barrier integrity and long-term functional recovery after ischemic stroke. Proc Natl Acad Sci U S A. 2024;121(11):e2400272121. doi:10.1073/pnas.2400272121
96. Gao M, Li Y, Ho W, et al. Targeted mRNA nanoparticles ameliorate blood-brain barrier disruption postischemic stroke by modulating microglia polarization. ACS Nano. 2024;18(4):3260–3275. doi:10.1021/acsnano.3c09817
97. Schulz JA, Hartz AMS, Bauer B. ABCB1 and ABCG2 regulation at the blood-brain barrier: potential new targets to improve brain drug delivery. Pharmacol Rev. 2023;75(5):815–853. doi:10.1124/pharmrev.120.000025
98. Spudich A, Kilic E, Xing H, et al. Inhibition of multidrug resistance transporter-1 facilitates neuroprotective therapies after focal cerebral ischemia. Nat Neurosci. 2006;9(4):487–488. doi:10.1038/nn1676
99. Murozono M, Matsumoto S, Okada S, et al. Reduction of brain infarction induced by a transient brain ischemia in mdr1a knockout mice. Neurochem Res. 2009;34(9):1555–1561. doi:10.1007/s11064-009-9943-6
100. Zhang L, Graf I, Kuang Y, et al. Neural progenitor cell-derived extracellular vesicles enhance blood-brain barrier integrity by NF-κB (Nuclear Factor-κB)-dependent regulation of ABCB1 (ATP-binding cassette transporter B1) in stroke mice. Arterioscler Thromb Vasc Biol. 2021;41(3):1127–1145. doi:10.1161/atvbaha.120.315031
101. Xie Q, Hao Y, Li N, et al. Cellular uptake of engineered extracellular vesicles: biomechanisms, engineered strategies, and disease treatment. Adv Healthc Mater. 2024;13(2):e2302280. doi:10.1002/adhm.202302280
102. Chen Z, Xiong M, Tian J, et al. Encapsulation and assessment of therapeutic cargo in engineered exosomes: a systematic review. J Nanobiotechnology. 2024;22(1):18. doi:10.1186/s12951-023-02259-6
103. Chen J, Zhang C, Li S, et al. Exosomes derived from nerve stem cells loaded with FTY720 promote the recovery after spinal cord injury in rats by PTEN/AKT signal pathway. J Immunol Res. 2021;2021:8100298. doi:10.1155/2021/8100298
104. Correia AC, Monteiro AR, Silva R, et al. Lipid nanoparticles strategies to modify pharmacokinetics of central nervous system targeting drugs: crossing or circumventing the blood-brain barrier (BBB) to manage neurological disorders. Adv Drug Deliv Rev. 2022;189:114485. doi:10.1016/j.addr.2022.114485
105. Shetgaonkar GG, Marques SM, DC CEM, et al. Exosomes as cell-derivative carriers in the diagnosis and treatment of central nervous system diseases. Drug Deliv Transl Res. 2022;12(5):1047–1079. doi:10.1007/s13346-021-01026-0
106. Song Y, Li Z, He T, et al. M2 microglia-derived exosomes protect the mouse brain from ischemia-reperfusion injury via exosomal miR-124. Theranostics. 2019;9(10):2910–2923. doi:10.7150/thno.30879
107. Tian T, Zhang HX, He CP, et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials. 2018;150:137–149. doi:10.1016/j.biomaterials.2017.10.012
108. Purushothaman A, Bandari SK, Liu J, et al. Fibronectin on the surface of myeloma cell-derived exosomes mediates exosome-cell interactions. J Biol Chem. 2016;291(4):1652–1663. doi:10.1074/jbc.M115.686295
109. Kim H, Yun N, Mun D, et al. Cardiac-specific delivery by cardiac tissue-targeting peptide-expressing exosomes. Biochem Biophys Res Commun. 2018;499(4):803–808. doi:10.1016/j.bbrc.2018.03.227
110. Mentkowski KI, Snitzer JD, Rusnak S, et al. Therapeutic potential of engineered extracellular vesicles. Aaps j. 2018;20(3):50. doi:10.1208/s12248-018-0211-z
111. Liang Y, Duan L, Lu J, et al. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11(7):3183–3195. doi:10.7150/thno.52570
112. Li L, Liu F, Welser-Alves JV, et al. Upregulation of fibronectin and the α5β1 and αvβ3 integrins on blood vessels within the cerebral ischemic penumbra. Exp Neurol. 2012;233(1):283–291. doi:10.1016/j.expneurol.2011.10.017
113. Lv X, Song X, Long Y, et al. Preclinical evaluation of a dual-receptor targeted tracer [(68)Ga]Ga-HX01 in 10 different subcutaneous and orthotopic tumor models. Eur J Nucl Med Mol Imaging. 2023;51:54–67. doi:10.1007/s00259-023-06412-z
114. Deng C, Zhang Q, He P, et al. Targeted apoptosis of macrophages and osteoclasts in arthritic joints is effective against advanced inflammatory arthritis. Nat Commun. 2021;12(1):2174. doi:10.1038/s41467-021-22454-z
115. Cen J, Zhang R, Zhao T, et al. A water-soluble quercetin conjugate with triple targeting exerts neuron-protective effect on cerebral ischemia by mitophagy activation. Adv Healthc Mater. 2022;11(22):e2200817. doi:10.1002/adhm.202200817
116. Cho S, Szeto HH, Kim E, et al. A novel cell-permeable antioxidant peptide, SS31, attenuates ischemic brain injury by down-regulating CD36. J Biol Chem. 2007;282(7):4634–4642. doi:10.1074/jbc.M609388200
117. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
118. Sadeghi S, Tehrani FR, Tahmasebi S, et al. Exosome engineering in cell therapy and drug delivery. Inflammopharmacology. 2023;31(1):145–169. doi:10.1007/s10787-022-01115-7
119. Kim J, He MJ, Widmann AK, et al. The role of neurotrophic factors in novel, rapid psychiatric treatments. Neuropsychopharmacology. 2024;49(1):227–245. doi:10.1038/s41386-023-01717-x
120. Tuwar MN, Chen WH, Yeh HL, et al. Association between brain-derived neurotrophic factor and lipid profiles in acute ischemic stroke patients. Int J Mol Sci. 2024;25(4):2380. doi:10.3390/ijms25042380
121. Hassanzadeh P, Arbabi E, Atyabi F, et al. Nerve growth factor-carbon nanotube complex exerts prolonged protective effects in an in vitro model of ischemic stroke. Life Sci. 2017;179:15–22. doi:10.1016/j.lfs.2016.11.029
122. Di Raimondo D, Rizzo G, Musiari G, et al. Role of regular physical activity in neuroprotection against acute ischemia. Int J Mol Sci. 2020;21(23):9086. doi:10.3390/ijms21239086
123. Hyenne V, Lefebvre O, Goetz JG. Going live with tumor exosomes and microvesicles. Cell Adh Migr. 2017;11(2):173–186. doi:10.1080/19336918.2016.1276694
124. Khan AA, Man F, Faruqu FN, et al. PET imaging of small extracellular vesicles via [(89)Zr]Zr(oxinate)(4) direct radiolabeling. Bioconjug Chem. 2022;33(3):473–485. doi:10.1021/acs.bioconjchem.1c00597
125. Gao Y, Chu C, Jablonska A, et al. Imaging as a tool to accelerate the translation of extracellular vesicle-based therapies for central nervous system diseases. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2021;13(3):e1688. doi:10.1002/wnan.1688
126. Han Z, Liu S, Pei Y, et al. Highly efficient magnetic labelling allows MRI tracking of the homing of stem cell-derived extracellular vesicles following systemic delivery. J Extracell Vesicles. 2021;10(3):e12054. doi:10.1002/jev2.12054
127. Betzer O, Barnoy E, Sadan T, et al. Advances in imaging strategies for in vivo tracking of exosomes. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2020;12(2):e1594. doi:10.1002/wnan.1594
128. Hánělová K, Raudenská M, Masařík M, et al. Protein cargo in extracellular vesicles as the key mediator in the progression of cancer. Cell Commun Signal. 2024;22(1):25. doi:10.1186/s12964-023-01408-6
129. Xu X, Iqbal Z, Xu L, et al. Brain-derived extracellular vesicles: potential diagnostic biomarkers for central nervous system diseases. Psychiatry Clin Neurosci. 2024;78(2):83–96. doi:10.1111/pcn.13610
130. Ma CY, Zhai Y, Li CT, et al. Translating mesenchymal stem cell and their exosome research into GMP compliant advanced therapy products: promises, problems and prospects. Med Res Rev. 2024;44(3):919–938. doi:10.1002/med.22002
131. Zhou H, Yuen PS, Pisitkun T, et al. Collection, storage, preservation, and normalization of human urinary exosomes for biomarker discovery. Kidney Int. 2006;69(8):1471–1476. doi:10.1038/sj.ki.5000273
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.