Back to Journals » ImmunoTargets and Therapy » Volume 15
Near Infrared Photoimmunotherapy for Lung Cancer: Recent Development and Perspective
Authors Okada R ![]() , Yamada M, Sato M, Sato K
, Yamada M, Sato M, Sato K ![]()
Received 12 March 2026
Accepted for publication 29 June 2026
Published 17 July 2026 Volume 2026:15 608901
DOI https://doi.org/10.2147/ITT.S608901
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Jadwiga Jablonska
Ryu Okada,1– 3,* Mizuki Yamada,1,3,4,* Mitsuo Sato,1,4 Kazuhide Sato1,3,5
1Graduate School of Medicine, Nagoya University, Nagoya, Japan; 2Research Unit for Life Sciences with Light Modulation, B3 Unit, Institute for Advanced Research, Nagoya University, Nagoya, Japan; 3Advanced Analytical and Diagnostic Imaging Center (AADIC) / Medical Engineering Unit (MEU), B3 Unit Frontier, Institute for Advanced Research, Nagoya University, Nagoya, Japan; 4Division of Host Defense Sciences, Department of Integrated Health Sciences, Graduate School of Medicine, Nagoya University, Nagoya, Japan; 5FOREST-Souhatsu, Japan Science and Technology Agency, Tokyo, Japan
*These authors contributed equally to this work
Correspondence: Kazuhide Sato, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, 466-8550, Japan, Tel +81-052-744-2167, Fax +81-052-744-2176, Email [email protected]
Background: Lung cancer is the leading cause of cancer mortality worldwide, and despite advances in surgery, chemotherapy, radiotherapy, and immunotherapy, challenges persist due to off-target toxicity and limited specificity. Photodynamic therapy (PDT), a minimally invasive treatment using photosensitizers and light to induce ROS-mediated cell death, has been explored for early-stage lung cancer; however, its clinical utility remains limited by poor tissue penetration, nonspecific photosensitizer uptake, and oxygen dependence.
Discussion: Near-infrared photoimmunotherapy (NIR-PIT) is a highly specific cancer treatment that overcomes many limitations of conventional PDT. It employs antibody–photoabsorber conjugates, such as monoclonal antibodies linked to IRDye700DX, which bind selectively to cancer cell surface antigens and are activated by 690-nm NIR light. This illumination induces a unique, non-ROS-dependent cell death, "photochemosis”, marked by immediate physical disruption of cancer cell membranes. Beyond direct cytotoxicity, NIR-PIT elicits robust immunogenic cell death, promotes systemic antitumor immunity, and permits repeated treatment cycles without compromising safety. The air-rich lung environment and deep penetration of NIR light make lung cancer an ideal NIR-PIT candidate. Advances in light-delivery devices—including endoscopic catheters, flexible LED systems, and endovascular therapy–based technologies—have greatly improved the feasibility of targeting deep pulmonary lesions for clinical implementation. Moreover, the modular design of NIR-PIT allows adaptation to diverse tumor targets, including EGFR and components of the tumor microenvironment, broadening its applicability.
Conclusion: In this review, we provide a comprehensive overview of the mechanisms, advantages, and recent technological innovations of NIR-PIT, with a focus on its potential as a transformative therapeutic approach for lung cancer.
Keywords: near-infrared photoimmunotherapy, IR700, photo-chemical reaction, lung cancer, immunotherapy
Introduction
Lung cancer is the leading cause of cancer-related deaths worldwide.1,2 Conventional treatments for lung cancer include surgery, anti-cancer drugs, radiation, and cancer immunotherapy; however, they are associated with challenges such as non-specificity, cytotoxicity against normal cells, and systemic side effects.
Phototherapy has been used as an alternative treatment for lung cancer. Conventional phototherapy, also known as photodynamic therapy (PDT), has been used to treat patients with minimally invasive lung cancer, especially those with early-stage central lung cancers.3 Although the relative prevalence of PDT as a treatment for lung cancer is low, many clinical studies have shown it to be an effective cancer treatment strategy that improves the quality of life and survival of patients with inoperable cancer.4
In this review, we summarize recent developments in near-infrared photoimmunotherapy (NIR-PIT) and discuss its potential application to lung cancer treatment. First, we outline the principles and limitations of conventional PDT, with particular attention to issues such as limited tumor selectivity, oxygen dependence, and insufficient light delivery to deep pulmonary lesions. We then describe the basic mechanism of NIR-PIT, including the design of antibody–photoabsorber conjugates, target-specific binding to cancer cell surface antigens, and NIR light–induced cancer cell death. In particular, we focus on the unique cell death mechanism of NIR-PIT, which differs from conventional ROS-dependent PDT and is characterized by rapid physical disruption of the plasma membrane.
We also review candidate molecular targets for NIR-PIT in lung cancer, including tumor-associated antigens such as EGFR, HER2, CEA, and other emerging targets, as well as components of the tumor microenvironment. Furthermore, we discuss the immunological consequences of NIR-PIT, including immunogenic cell death, activation of antitumor immunity, and the potential for combination with immune checkpoint inhibitors and other systemic therapies. Finally, we highlight recent advances in light-delivery technologies, including endoscopic, interstitial, flexible LED-based, and endovascular approaches, which may expand the clinical feasibility of NIR-PIT for both superficial and deep-seated lung tumors. Through this review, we aim to provide an integrated perspective on how NIR-PIT may overcome the limitations of existing phototherapies and contribute to the development of a more selective and effective therapeutic strategy for lung cancer.
Current Status and Limitations of Phototherapy for Lung Cancer (Conventional Photodynamic Therapy)
PDT is a noninvasive form of modern non-ionizing radiation therapy. This treatment is based on the local or systemic administration of a photosensitizing compound (PS), which preferentially accumulates in cancerous cells and tissues through active or passive mechanisms,5 in combination with activation by light of a single wavelength. Upon absorbing light of an appropriate wavelength, PS molecules initiate activation processes that generate reactive oxygen species (ROS), leading to the selective destruction of cancer cells.5,6 Free radicals are generated when a ground-state PS absorbs a photon and transitions into an excited singlet state. This singlet state subsequently undergoes intersystem crossing to form a triplet state that can interact with the surrounding molecules. The PS may abstract an electron from nearby reducing molecules to produce ROS or interact directly with molecular oxygen to generate singlet oxygen.7 In this two-step process, the otherwise harmless PS is activated exclusively by light exposure, causing localized tissue destruction and significantly minimizing side effects. Organelles and cellular membranes can be directly damaged by PDT, depending on the type, concentration, and intracellular localization of the PS, as well as light fluence and spatial distribution of the radicals (Figure 1).8
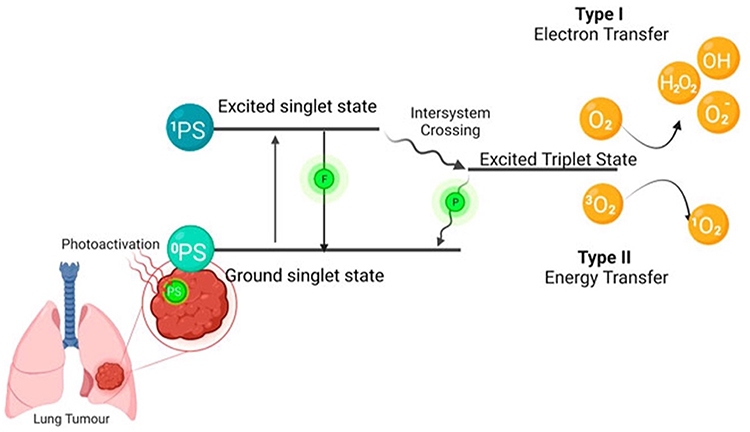 |
Figure 1 The mechanism of photodynamic therapy (PDT) The photosensitizer (PS) is absorbed when the PS is in its ground state. It goes into its first excited singlet state because of photoactivation. This state can be broken down by emitting fluorescence, or it can cross over to the more stable excited triplet state. Type I is when the PS in its excited triplet state reacts with biomolecules (such as lipids, proteins, and nucleic acids), and the radical mechanism is used to transfer hydrogen atoms. It generates free radicals and radical ions (the type of radical varies depending on the target molecule, such as lipids, proteins, or nucleic acids), which react with oxygen to produce reactive oxygen species. Type II reactions are based on a phenomenon known as triplet–triplet annihilation. In these reactions, the PS in its excited triplet state reacts with oxygen in its triplet ground state. This results in the formation of highly reactive and cytotoxic singlet oxygen. Reproduced from Crous and Abrahamse, 2022,8 Frontiers in Pharmacology, 13:932098, under the terms of the Creative Commons Attribution License (CC BY). No changes were made. |
While PDT has advantages over other therapies, such as being minimally invasive and having less impact on surrounding normal tissues owing to its target specificity and cancer tissue visualization, it has limitations when immediate results are required because of the need for a certain amount of time after drug administration, and the nonspecific uptake of PS into normal cells may cause undesirable side effects. It may also mediate the production of ROS, which may damage not only target cancer cells but also normal cells.6,9
Therefore, phototherapy based on mechanisms other than ROS production is highly desirable.
NIR-PIT as a Novel Cancer Therapy
Conventional PDT uses a hydrophobic photosensitizer to pass through the lipid cell membrane, limiting its ability to selectively target and biodistribute drugs at the target site.10–12 Conventional PDT drugs conjugated to a target moiety have limited biodistribution because the target moiety is usually hydrophilic. Therefore, the PDT antibody conjugates may be less biocompatible. To overcome this problem, a hydrophilic photosensitizer is required. Moreover, absorption wavelengths ranging from 650 to 900 nm (ie, the biological window) would be ideal for penetrating biological tissues.13
In 2011, Hisataka Kobayashi et al developed NIR-PIT as a new phototherapy to overcome the challenges associated with conventional PDT.14 NIR-PIT, a novel cancer therapy, is a localized cancer therapy with dual selectivity for antibodies and light. The therapeutic agent consists of an antibody-photoabsorber conjugate (APC) that binds to the cell surface markers of cancer. In other words, this therapy can selectively kill cancer cells because of the cancer specificity of the monoclonal antibody and the photoabsorber IRDye700DX (IR700) bound to the antibody, which is activated with local NIR-light irradiation (Figure 2).15
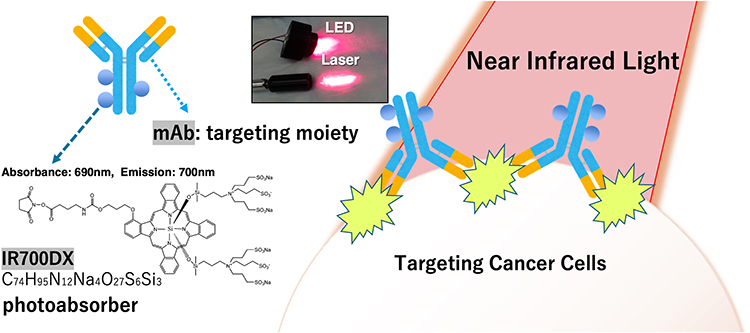 |
Figure 2 Scheme of NIR-PIT NIR-PIT is composed of antibody and photoabsorber, IR700DX. Monoclonal antibodies (mAbs) can target specific antigen on targeting cancer cells. IR700DX absorbs light with a peak wavelength of 690 nm and emits fluorescence at 700 nm. mAb-IR700 is a type of antibody-drug conjugate (ADC) and can be considered a photo-activated antibody-drug conjugate. It enables dual targeting with light and antibodies, making it a highly selective cancer therapy. NIR-PIT is a molecularly photo-targeted cancer therapy, which represents a next-generation anti-cancer treatment modality. |
Antibodies play a role in the target specificity of this therapy because they can bind specifically and stably to cell membrane surface molecules. More than 100 antibody drugs are currently approved in Japan, the US, and Europe. Recently, the development of various modified antibody drugs and antibody drugs for new therapeutic targets has been progressing in order to improve efficacy and safety.
Regarding light, which contributes to the selectivity of the treatment, NIR light at 690 nm was chosen because, unlike ultraviolet light, it efficiently induces photochemical reactions that damage the target cells through physical and chemical mechanisms without causing DNA damage. Furthermore, NIR light avoids the absorption spectrum of hemoglobin, which is abundant in the body, and allows sufficient light to penetrate relatively deep tissues. The silicon-phthalocyanine derivative IR700 was employed as a photosensitizer that absorbs NIR light, generates energy, and selectively damages cells bound by antibodies. In contrast, conventional a PS typically absorbs only visible light and exhibits limited tissue penetration.14,16,17
PS is a crucial component that confers target specificity in phototherapy, and IR700, used in NIR-PIT, is a next-generation dye with numerous properties that are well suited for cancer therapy.16 IR700, as well as other phthalocyanine PS, has a large molar extinction coefficient, moderate quantum yield, and relatively good photochemical stability under natural light.14
A key advantage of IR700 is its hydrophilic properties resulting from specific chemical modifications. Unlike conventional hydrophobic PSs, IR700’s hydrophilicity prevents aggregation or precipitation in buffered physiological solutions, even at high dye-to-protein ratios, making it more suitable for therapeutic drug formulations and applications. Additionally, while traditional hydrophobic PS-antibody conjugates tend to aggregate and accumulate in the liver for metabolism, hydrophilic IR700 remains evenly dispersed in the bloodstream and does not impair the tumor-targeting ability of the antibody or alter tissue distribution following intravenous administration. Furthermore, the mechanism of cell death in NIR-PIT is involved protein aggregation of the conjugates and cancer cell surface antigens, along with physical damage to the cell membrane caused by the photochemical reactions—referred to as “photochemosis”—that are triggered by NIR light exposure.18,19 This photochemosis-induced disruption of the plasma membrane, unlike the ROS-dependent apoptosis or delayed cell death observed in conventional PDT, is accompanied by a rapid collapse of osmotic balance and induce necrotic cell death within only a few minutes.20 Therefore, there is no need for a therapeutic agent that is internalized or dependent on ROS. Thus, it is expected to overcome the challenges of conventional hydrophobic PS that hinder target selectivity and anti-tumor efficacy. The fact that IR700 is a hydrophilic photopigment without phototoxicity or biotoxicity also allows IR700 to dissociate from APCs by photoirradiation and be safely and rapidly excreted via urine.21
Third, IR700 can emit NIR fluorescence, facilitating fluorescent imaging of target tissues. This feature could facilitate more specific tumor irradiation and real-time monitoring of treatment progress by attenuating the IR700 fluorescence signals.22
Thus, NIR-PIT facilitates highly precise cancer treatment with minimal damage to normal tissues by combining target-specific APCs with light irradiation limited to tumors.
A further important point is that, unlike conventional cancer therapies, photochemosis by NIR-PIT activates diverse tumor-specific immune responses without compromising host immunity against cancer. When cancer-specific antigens and danger signals at the time of membrane damage are rapidly released extracellularly by NIR-PIT, local dendritic cells are activated. The activated dendritic cells then activate and educate cancer-specific naïve T cells, which in turn cause the proliferation of newly primed CD8+ T cells and cell-mediated cancer cell death. This process is known as immunogenic cell death (ICD), and NIR-PIT could be an ideal method for inducing anti-tumor immunity.12,23 Furthermore, the number of NIR-PIT repeat treatments is not limited; therefore, multiple NIR-PIT sessions can help overcome these challenges.
An international Phase III clinical trial for locoregional recurrent head and neck squamous cell carcinoma (HNSCC) (LUZERA-301, NCT03769506) was initiated in 2018 and is ongoing. The trial was approved by the US Food and Drug Administration (FDA) and the Japan Pharmaceuticals and Medical Devices Agency (PMDA) under Fast Track and SAKIGAKE, respectively.24
Although clinical studies in head and neck cancer have provided important proof of concept for the feasibility of NIR-PIT in humans, these findings should be distinguished from the current status of NIR-PIT in lung cancer. At present, direct clinical evidence for NIR-PIT in lung cancer remains limited, and its potential application to lung cancer is supported mainly by preclinical studies and ongoing advances in light-delivery technologies. Further investigation is required to establish its efficacy and safety in lung cancer.
Mechanism of NIR-PIT
Oxidative stress, the primary mechanism of PDT, is thought to be cellular damage caused by oxidative stress agents such as free radicals and singlet oxygen (type I and type II).25,26 The energy difference between the photoexcitation and photoemission of PSs acts on mitochondria and other structures, mainly inside cells, leading to cell apoptosis.27 This oxidative stress affects not only the target cells but also non-targeted adjacent cells.
By contrast, the mechanism of cell death induced by NIR-PIT is unique, termed “photochemosis” (Figure 3).18 When an antibody-photoabsorber (IR700) conjugate bound to a cancer cell surface antigen undergoes a photochemical reaction upon local near-infrared light irradiation, protein aggregation of the conjugates and the cancer cell surface antigen occurs, which causes physical cell membrane damage. An influx of water into the cell causes an osmotic difference, which ruptures the cell and induces cell death, a mechanism that is very different from that of existing cancer therapies.19 This therapy shows rapid therapeutic effects with few side effects owing to its high target specificity.12
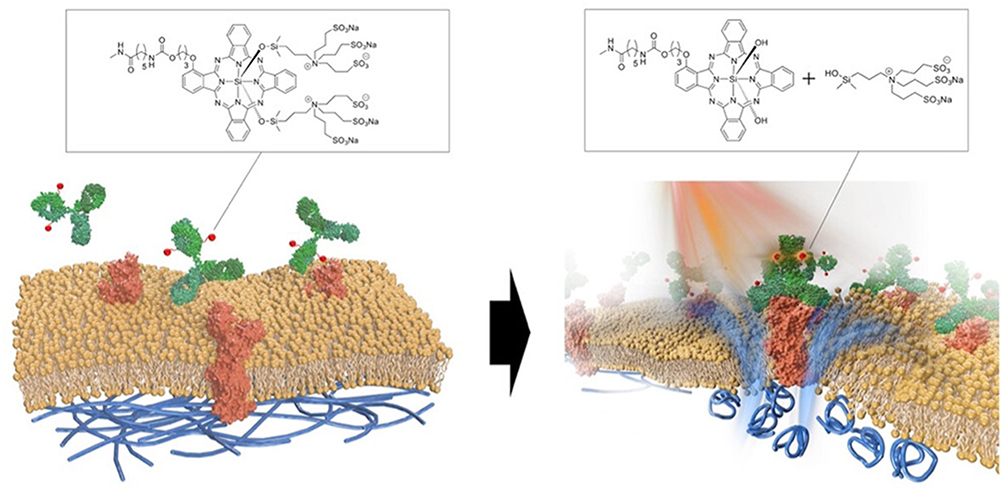 |
Figure 3 Scheme indicates the proposed mechanism of NIR-PIT An antibody-IR700-antigen complex is formed on the cell membrane. With NIR light irradiation, the ligands of IR700 were released form the antibody-IR700-antigen complex. The physical changes in aggregation and solubility of the antibody-IR700-antigen complex may produce physical stress on the membrane locally impairing cellular membrane function for maintaining membrane pressure, and then the water outside of the cell was flown into the cell to burst the cell. Reproduced from the graphical abstract of Sato et al, ACS Nano, 2025,18 under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND 4.0). No changes were made. |
In NIR-PIT, the chemical properties of IR700 (hydrophilic) are rapidly changed to hydrophobic by the release of the silanol side chain under NIR light. The hydrophobic IR700 aggregates in an aqueous solution, resulting in the loss of IR700 fluorescence from the solution, which is a series of photochemical reactions. This hypothesis was proven using analytical chemistry and nano-in-liquid imaging.19
Furthermore, after PIT, damage-associated molecular patterns (DAMPs) are rapidly released to activate an anti-tumor immune response,23 known as ICD, which makes NIR-PIT a promising approach for the treatment of distant metastases and heterogeneous tumors.
This mechanism can be proposed as a new light-induced cell-death mechanism that does not depend on oxygen and is completely different from the oxidative stress proposed in conventional phototherapy, establishing a theory that NIR-PIT has advantages as a new anti-cancer modality that is different from previous cell-death concepts.19
NIR-PIT as a Treatment for Lung Cancer (Tissue Characteristics of the Lung and Physical Properties of NIR)
The NIR light used in NIR-PIT is easily transmitted through air-filled tissues. Therefore, lungs with their high air content are suitable organs for NIR-PIT. In addition, NIR light in the 650–900 nm wavelength range is not readily absorbed by water or hemoglobin and exhibits maximum tissue penetration.28 Therefore, the 690 nm wavelength used in NIR-PIT is considered suitable for human therapy and can be used for treatment without harming the body.
In addition, the lung can be said to be a relatively easy organ to access internally due to their high air content, but there have been challenges in delivering sufficient light to deep cancer cells. However, recent developments in NIR light delivery devices, such as needle catheters that can be inserted into deep tumors,29 endoscopy, optical fiber-based techniques,30,31 and endovascular therapy-based catheter systems (endovascular therapy-based light illumination technology, ET-BLIT),32 are expected to be effective for deep cancer approaches.
Promising Therapeutic Targets for NIR-PIT in Lung Cancer
NIR-PIT can be used to alter the treatment pathway by creating mAb-IR700 using antibodies that target different ligands. Here, we summarize the promising therapeutic targets with a focus on NIR-PIT for lung cancer (Table 1).
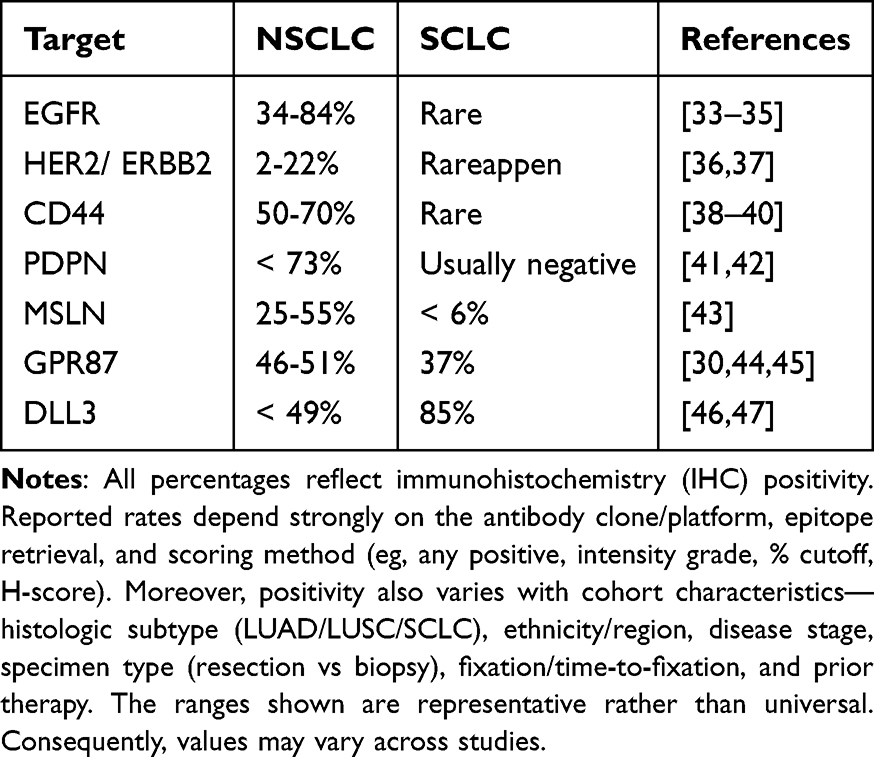 |
Table 1 Potential Molecular Targets for NIR-PIT in Lung Cancer |
Epidermal Growth Factor Receptor
Epidermal growth factor receptor (EGFR) is a transmembrane tyrosine kinase receptor belonging to the erythroblastosis oncogene B (ErbB) family.48,49 EGFR overexpression has been observed in a variety of solid tumors, including lung cancers, especially squamous cell carcinoma,50,51 and is associated with poor prognosis in several tumor types.52
Several preclinical studies have confirmed the therapeutic efficacy of EGFR-targeted NIR-PIT in several types of cancers, including lung cancer.53–56 Cetuximab-IR700, a chimeric IgG1 monoclonal antibody against EGFR, was first conditionally registered and conditionally approved for clinical use in recurrent head and neck cancer (HNC) clinical use in Japan in 2020.57
HER2
HER2 is a membrane tyrosine kinase receptor and a member of the ErbB family along with EGFR.58 Overexpression of Her2 is found in a variety of cancer cells, including breast,59,60 gastric,61 and esophageal cancers.62
Her2-targeted NIR-PIT, including trastuzumab-IR700, has demonstrated therapeutic efficacy in several cancers, including lung metastases63,64 and non-small cell lung cancer (NSCLC) models.65 Recent studies have shown that Her2-targeted NIR-PIT is more effective in CDDP chemoresistant small-cell lung cancer because chemotherapeutic agents, especially cisplatin-resistant (SBC-3/CDDP) cell lines, upregulate HER2 expression.66
Moreover, a previous study demonstrated the induction of a cytotoxic photo-bystander effect resulting from the photo-triggered release of the drug from IR700-conjugated trastuzumab emtansine (T-DM1), T-DM1–IR700.67
CD44
CD44 is a non-kinase transmembrane glycoprotein and a cancer stem cell marker that promotes tumor initiation and progression.68,69 CD44 overexpression is a poor prognostic factor in various types of cancer, including lung cancer.70
Anti-CD44 monoclonal antibodies (mAbs) can play crucial roles in the regulation of cancer cell growth and progression. For instance, in acute myeloid leukemia (AML), a malignant clonal disorder characterized by the abnormal proliferation of immature myeloid cells, monoclonal antibodies such as H90 and A3D8 have been shown to induce cell differentiation by targeting and inhibiting the CD44 antigen. In addition to promoting differentiation of AML cell lines, these mAbs can inhibit cell proliferation and induce apoptosis.71 Moreover, CD44-targeting antibodies have been used to block lymphocyte migration, lymphohematopoiesis, and tumor metastasis.72 In a different study focused on lung diseases, treatment with anti-CD44 antibodies disrupted fibroblast adhesion to the substratum, thereby increasing the apoptosis of fibroblast cells.73
A study using CD44-positive oral squamous cell carcinoma and breast cancer models confirmed the benefits of NIR-PIT.74,75 In addition, the combination of type 1 cytokines (short-term IL-15 administration) was shown to further enhance CD44-targeted NIR-PIT coupled with immune activation compared with monotherapy for lung tumors.76
Podoplanin
Podoplanin (PDPN) is a mucin-type transmembrane glycoprotein expressed on the cell surface that plays an essential role in tumor progression and normal development of the lungs, kidneys, and lymphatic vasculature.77
PDPN expression has been documented in a wide range of malignancies,78,79 including squamous cell carcinomas of the head and neck, lungs, uterus, oral cavity, and esophagus, as well as in malignant gliomas,80,81 mesotheliomas,82 bladder cancer,83 osteosarcoma,84 ovarian cancer,85 and testicular tumors.86
PDPN is expressed not only in tumor cells but also within the tumor stroma, including cancer-associated fibroblasts (CAFs). Increased PDPN expression in CAFs derived from lung,87–89 breast,90 and pancreatic91 tumors is associated with greater tumor aggressiveness and poorer patient prognosis.77
Several studies using malignant pleural mesothelioma (MPM) and pleural disseminated lung cancer models have demonstrated that NZ-1 antibody-based PDPN-targeted NIR-PIT has demonstrated therapeutic efficacy.92
Mesothelin
Mesothelin (MSLN) is a glycoprotein that attaches to the plasma membrane via a glycophosphatidylinositol (GPI) anchor. The Mesothelin gene encodes a 69-kDa precursor protein, which is cleaved into two parts: a 31-kDa secreted fragment known as megakaryocyte potentiating factor and a 40-kDa membrane-bound form referred to as MSLN.93,94 MSLN is largely absent in normal tissues and its expression is limited to the mesothelial cells of the peritoneum, pleura, and pericardium.95 In contrast, it is highly expressed in a wide range of cancers, including nearly all cases of mesothelioma, and in a significant proportion of solid tumors, such as lung cancer (60–70%), pancreatic cancer (80–85%), cholangiocarcinoma (60–65%), ovarian cancer (60–65%), gastric cancer (50–55%), colon cancer (40–45%), breast cancer (25–30%), and endometrial cancer (20–25%).96 Owing to its high prevalence in tumors, MSLN has emerged as a promising target for immunotherapy, and its soluble fragment is under investigation as a potential biomarker for cancer diagnosis.96,97
Recently, a humanized non-region I antibody (hYP218) with a high binding affinity for the C-terminal region (residues 487–581) of mesothelin was developed. Promising results have been reported regarding the mesothelin-specific cytotoxic activity.98
The hYP218-IR700 conjugate demonstrated strong efficacy in treating mesothelin-expressing tumors using NIR-PIT. In vitro, NIR-PIT with hYP218-IR700 induced rapid cell death, whereas in vivo, it significantly reduced tumor growth and improved survival outcomes. These findings suggest that hYP218-IR700 could serve as an effective platform for NIR-PIT targeting mesothelin-expressing tumors.99
GPR87
GPR87 belongs to the G protein-coupled receptor (GPCRs).100 GPR87 is generally expressed at very low levels in most normal human tissues, except in the prostate, placenta, and head and neck regions.78,79,101,102 However, markedly elevated levels of GPR87 have been observed in various malignancies, including squamous cell carcinomas of the lung, urinary bladder, cervix, testis, skin, and head and neck, as well as in lung adenocarcinomas and transitional cell carcinomas of the urinary bladder. Moreover, GPR87 expression is associated with clinical stage, disease recurrence, resistance to immunotherapy and chemotherapy, and reduced overall and progression-free survival in patients with lung cancer.102–106
Studies in mouse models of transplanted lung adenocarcinoma, small cell lung cancer (SCLC), and MPM cell lines have confirmed the therapeutic efficacy of GPR87-targeted NIR-PIT.107
Delta-Like Canonical NOTCH Ligand 3
Delta-like canonical Notch ligand 3 (DLL3), a single-pass transmembrane protein and member of the Notch ligand family, plays a role in cell signaling.108–110 Although it is predominantly expressed in neural and neuroendocrine tissues during embryonic development, its expression is minimal or absent in normal adult tissues. However, it is found on the surface of tumor cells.109,110 DLL3 expression is markedly elevated in glioblastoma, lower-grade glioma, skin cutaneous melanoma, tenosynovial giant cell tumors, and uterine carcinosarcoma compared to that in normal tissues. Additionally, high DLL3 expression has been linked to more aggressive diseases, indicating its potential role in promoting malignant phenotypes.46,111 DLL3 has also been detected in circulating tumor cells and blood of patients with cancer.112–115 Based on these findings, DLL3 is under investigation as a biomarker for cancer diagnosis and prognosis, as well as a therapeutic target.114,116 Its minimal expression in normal tissues relative to tumors makes it a promising candidate for targeted therapies designed to reduce off-target effects.
DLL3, a ligand for the Notch receptor, is a promising therapeutic target for SCLC and other neuroendocrine tumors. Rovalpitumab tesirin, the first antibody targeting DLL3, was discontinued in August 2019 after the TAHOE (NCT03061812) and MERU (NCT03033511) clinical trials yielded negative results.109 However, a study using an SCLC xenograft model showed that rovalpituzumab demonstrated therapeutic efficacy against DLL3-targeted NIR-PIT.117,118
Current Clinical Status of NIR-PIT in Lung Cancer
At present, NIR-PIT has not yet been established as a standard clinical treatment for lung cancer, and large-scale clinical trials specifically targeting lung cancer have not been completed. The most advanced clinical experience with NIR-PIT has been obtained in recurrent or unresectable head and neck cancer using cetuximab sarotalocan sodium, an EGFR-targeted antibody–photoabsorber conjugate. Therefore, the clinical evidence from head and neck cancer provides important proof of concept for the safety and feasibility of NIR-PIT in humans, but it should be clearly distinguished from the current developmental status of NIR-PIT in lung cancer. Although direct clinical evidence in lung cancer remains limited, accumulating animal model studies and advances in light-delivery devices provide a strong rationale for future clinical investigation. Further studies are required to identify optimal target antigens, select appropriate patient populations, establish safe and effective light-delivery methods, and evaluate combination strategies with chemotherapy, molecular targeted therapy, radiotherapy, or immune checkpoint inhibitors.
Therapeutic Targets of Non-Tumor Cells in the Tumor Microenvironment
In the TME, T and NK cells that recognize cancer cells are abundant; however, the presence of neighboring immunosuppressive cells serves as a mechanism to circumvent cytotoxic functions.119 Immune checkpoint molecules inhibit anti-tumor immune responses. Therefore, modulation of immunosuppressor cells or immune checkpoint molecules in the TME is an important step in enhancing the anti-tumor immune response and inhibiting tumor growth.120 Here, we summarize the promising therapeutic targets for non-tumor cells in the TME (Table 2).
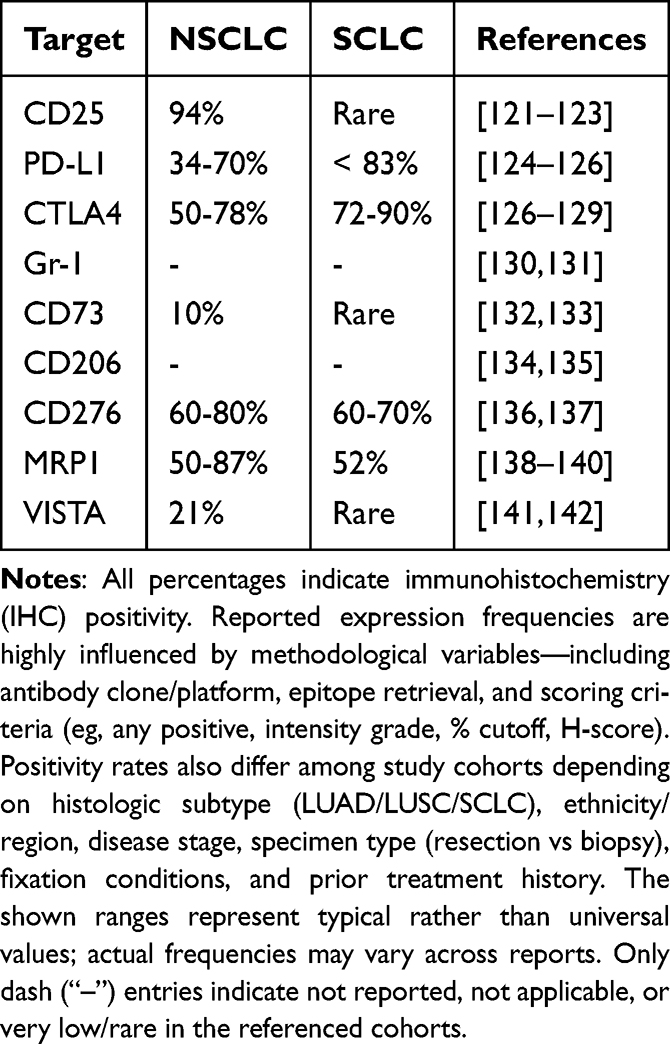 |
Table 2 Potential Molecular Targets for NIR-PIT in the Lung Cancer Tumor Microenvironment |
CD25
CD25 is an IL-2 receptor expressed on the surface of Tregs and is associated with the immunosuppression of effector T and NK cells at the tumor site. A high percentage of Tregs among tumor-infiltrating lymphocytes is associated with poor prognosis. In HNSCC, the extent of TME Treg infiltration is correlated with prognosis.143,144
CD25-targeted NIR-PIT selectively eradicated Tregs in the TME without eliminating local effector T cells or Tregs present in other organs, and induced tumor regression, resulting in the rapid activation of CD8+ T cells and NK cells. Furthermore, this effect was observed in non-irradiated, distant, and untreated tumors, confirming the abscopal effect.145
In addition, repeated depletion of Tregs, anti-CD25-F(ab’)2NIR-PIT, which is characterized by rapid clearance while avoiding Fc-mediated ADCC and complement-dependent cytotoxicity (CDC),146 and CD25-targeted NIR-PIT in combination with CD44- or hEGFR-targeted NIR-PIT147 showed better therapeutic efficacy.
Programmed Death Ligand 1
Programmed death ligand 1 (PD-L1) is a transmembrane protein and an immune checkpoint protein that binds to PD-1 expressed on tumor-infiltrating lymphocytes and induces immune tolerance in tumor cells by reducing immune response cytokines.148,149 Overexpression in many cancer cells, including lung cancer cells, is associated with poor prognosis.150–153
Two anti-PD-L1 antibodies, atezolizumab and durvalumab, have been approved by the European Medicines Agency (EMA) and the US FDA for the treatment of advanced NSCLC.154
PD-L1-targeted NIR-PIT promotes rupture of PD-L1-positive cells. Furthermore, by inhibiting tumor immunosuppressive pathways, it activates CD8+ T cells and NK cells in the TME, resulting in enhanced anti-tumor immunity.155,156 Enhanced anti-tumor effects have been observed in various major types of models, including mouse models of lung cancer, whereas in vitro studies on cytotoxic effects are limited.157,158
Cytotoxic T-Lymphocyte-Associated Protein 4
Cytotoxic T-lymphocyte-associated protein 4 (CTLA4) is an immune checkpoint molecule primarily expressed on activated T lymphocytes and regulatory T cells (Tregs), where it inhibits T cell activation.159–162 Ipilimumab, an anti-CTLA4 antibody, was among the first immune checkpoint inhibitors developed for clinical use; it binds to CTLA4 on T cells, blocking its inhibitory function, and thereby enhancing T cell activation and anti-tumor responses.163,164
Local depletion of CTLA4-expressing Tregs by NIR-PIT is a promising anti-tumor therapy that promotes CD8+ T cell activation and infiltration in the TME. Furthermore, the combination with cancer cell-targeted NIR-PIT showed better therapeutic efficacy.165,166
Gr-1
Gr-1, together with CD11b, is a widely characterized target of murine myeloid-derived suppressor cells (MDSCs), with two major cell subsets: granulocytic MDSCs (PMN-MDSCs, CD11b+Ly6ClowLy6G+) and monocytic MDSCs (M-MDSCs, CD11b+Ly 6ChighLy6G-).167,168 MDSCs, such as Tregs and tumor-associated macrophages (TAMs), are immunosuppressive cells that act as a suppressive time component to attenuate the immune response169,170 and are of increasing interest because of their role in the prognosis, development, and therapy of lung cancer.171–173
Gr-1-targeted NIR-PIT has been used to eliminate splenic MDSCs, suggesting the potential of eliminating MDSCs from tumors to reduce immunosuppression.
CD73
CD73, also known as extracellular 5′-nucleotidase, is a ribonucleotidase encoded by the NT5E gene. Predominantly anchored to the cell membrane via GPI, CD73 catalyzes the conversion of extracellular adenosine monophosphate (AMP) into adenosine and inorganic phosphate.174,175 A soluble form of CD73 also exists that retains functional properties partially similar to those of its membrane-bound counterpart.176–178
Overexpression of CD73 in cancer cells leads to increased adenosine levels, whereas tumor-driven proliferation creates a hypoxic environment that causes normal cell lysis and ATP release, which is subsequently converted to adenosine via the CD39/CD73 axis, promoting immune evasion and facilitating cancer metastasis, persistence, and proliferation.179
In lung adenocarcinoma and NSCLC, CD73 expression is associated with increased PD-L1 levels and increased infiltration of tumor-associated immune cells, thereby promoting an immunosuppressive TME.180–186
CD73 has emerged as a promising therapeutic target for various cancers, including breast and lung cancers.187–189 In breast cancer, CD73 promotes local invasion via the epidermal growth factor (EGF)/EGF receptor signaling pathway.190–192 Moreover, studies on glioblastoma have shown that, although treatment can elevate CD73 expression, its depletion significantly improves survival,193–196 offering valuable insights into future anti-CD73 therapeutic strategies.
CD73-targeted NIR-PIT simultaneously eliminates tumor cells and key immunosuppressor cells.16,197
CD206
CD206 is a highly specific marker of M2 macrophages that lack intracellular signaling domains198 and is primarily regarded as an endocytic receptor involved in the internalization of extracellular substances for clearance or antigen presentation.199 Recent studies have suggested that CD206 also plays an active role in the immune response by directly modulating the activity of other immune cells,200 although the relationship between CD206 expression and tumor metastasis remains controversial owing to conflicting findings.201,202
M2 macrophage polarization promotes metastasis in various malignancies, including colorectal, pancreatic, and gastric cancers.203–205 In NSCLC, M2 TAMs facilitate the proliferation and migration of cancer cells.206,207 An increased number of M2 TAMs is associated with metastasis and poor prognosis in patients with SCLC.208 Moreover, macrophages in the lungs of the SCLC mouse transplant models predominantly exhibited the M2 phenotype, indicating their potential immunosuppressive role in facilitating SCLC invasion.208
TAM-targeted NIR-PIT using anti-CD206 antibodies demonstrated therapeutic efficacy and inhibited lung metastasis in a mouse model of breast cancer.209
CD276
CD276 is a type I transmembrane protein expressed in many tissues and cell types and has been implicated in the regulation of T cell-mediated immune responses by functioning as both a T cell co-stimulator and co-inhibitor.210 It is overexpressed in a wide range of human solid tumors, has been implicated in cancer progression and metastasis, and has been investigated as a potential target for cancer immunotherapy.211,212
NIR-PIT with IR700-αCD276/Fab significantly inhibited subcutaneous 4T1 tumor growth and lung metastasis. In addition, it caused in vivo anti-tumor immunity. Combined with an anti-PD-1/PD-L1 blockade, it significantly inhibited tumor growth and prevented lung metastasis by recruiting CD8+ T cells for tumor infiltration.213 It could be argued that local and systemic anti-tumor responses, both by CD276 targeted therapy for local immunomodulation and combination therapy with PD-L1/PD-1 axis inhibition, may be promising strategies for eliminating primary tumors and disseminated metastases in NIR-PIT.
Multidrug Resistance-Associated Protein 1
Multidrug resistance-associated protein 1 (MRP1) encodes plasma membrane proteins that perform distinct and essential physiological transport functions and actively export various xenobiotics and their metabolites (including both hydrophobic and hydrophilic antineoplastic agents) against their concentration gradients (drug threshold). MRP1 can transport a wide range of substrates.214,215 MRP1 is overexpressed in leukemia, esophageal cancer, and NSCLC.216
MRP1 contributes to the resistance to several chemotherapeutic agents, including anthracyclines (eg, doxorubicin), vinca alkaloids, podophyllotoxins, mitoxantrone, methotrexate, and alkylating agents.217 It also recognizes and effluxes newer targeted anti-cancer agents that alter tumor growth, proliferation, and metastatic signaling pathways. One such targeted agent is vandetanib, a small-molecule TKI that blocks the ATP-binding domains of ABC drug transporters. However, the exact mechanism remains unclear.218 Other targeted therapies targeting MRP1 include geldanamycin, the farnesyl protein transferase inhibitor lonafarnib, and nutlin-3.218–221 Additionally, MRP1 has been implicated in various physiological and pathophysiological processes, such as inflammatory responses, oxidative defense mechanisms, and the pathogenesis and prognosis of cancer.222,223 Leukotriene C4 (LTC4), a pro-inflammatory product of arachidonic acid involved in asthmatic and allergic reactions as well as smooth muscle constriction, is a key physiological substrate of MRP1.224,225
Recent studies by Li et al have demonstrated that Mab-IR700, an anti-MRP1 antibody, exerts a strong modulatory effect on H69AR cells. NIR-PIT with an anti-MRP1 antibody (Mab)-IR700 conjugate (Mab-IR700) exhibited therapeutic efficacy, suggesting its potential of MRP1 as a new target for the treatment of drug-resistant SCLC.226
V-Domain Immunoglobulin Suppresses T Cell Activation
The V-domain immunoglobulin suppressor of T cell activation (VISTA),227 also referred to as Dies1,228 GI24,229 PD-1H,230 or DD1alpha,231 is a newly identified member of the B7 family that has gained attention as a promising target for combination cancer immunotherapy.232 Initially, VISTA was described by two research groups in the context of embryonic development and tumor invasion.228,229 Subsequent studies have primarily focused on the role of VISTA in the regulation of immune responses.230,233–235 VISTA, a protein encoded by the B7/CD28 immunomodulatory gene family, is expressed on antigen-presenting cells and exerts an inhibitory effect on T cell activity. In mouse models, the blockade of VISTA exacerbated the progression of T cell-mediated autoimmune diseases, whereas the agonistic antibody MH5A, targeting VISTA (PD-1H), was shown to suppress graft-versus-host disease (GvHD).227,230
In addition to its role as an immune receptor on T cells, VISTA can function as a ligand on antigen-presenting cells or tumor cells, triggering immune regulatory activities. Several studies on human cancers have shown that VISTA is expressed in melanoma, NSCLC, ovarian cancer, endometrial cancer, malignant pleural epithelial mesothelioma, and various other tumors or tumor cell lines.236–239
In a study on NSCLC, the VISTA protein was found in most samples, with expression observed in 21% of tumor cells and 98% of stromal cells.141 The proteins displayed membrane and cytoplasmic staining. Similarly, other studies have demonstrated that VISTA is highly expressed in the lymphocytes of lung squamous cell carcinoma and adenocarcinoma.240–242 VISTA expression positively correlated with the PD-1 axis, CD8+ T cells, and CD68+ macrophages. In lung adenocarcinoma, an increase in VISTA expression within stromal cells was significantly associated with the absence of EGFR mutations, whereas higher VISTA expression in tumor areas correlated with improved 5-year survival rates.141 Conversely, other studies have reported that high VISTA expression in tumor-associated immune cells is associated with poor survival outcomes and increased mortality.241,242
Animal model studies have demonstrated that dual inhibition of VISTA and PD-L1 significantly suppresses the growth of B16BL6 melanoma tumors, whereas treatment with a single monoclonal antibody is largely ineffective.243
VISTA-targeted NIR-PIT using two mouse tumor models, MC38-luc (colon cancer) and LL2-luc (lung adenoma), showed that the TME effectively treated tumors by locally reducing the number of VISTA-expressing immunosuppressor cells.244
NIR Light Delivery Device for Pulmonary NIR-PIT
For effective NIR-PIT, irradiation of the target tissue with NIR light is critical; the depth of penetration of NIR light into the tissue is approximately 2–3 cm from the surface and attenuates significantly with increasing distance. Accurate irradiation with NIR light is necessary to produce effects on therapeutic targets without adversely affecting normal tissues, even in deep areas, such as the lungs. Here, we summarize the information on light irradiation to enable localized NIR-PIT for lung cancer and highlight the development of a variety of new devices.
The first is a needle catheter that allows the introduction of a tissue irradiation diffuser into the tissue for intra-tissue therapy. Fiber-optic diffusers placed in needle catheters can be inserted into deep tumors; this technique has been used in clinical practice.29
Another option is to administer NIR light endoscopically. In respiratory diseases, optical fibers are utilized through bronchoscopes to deliver NIR light to critical areas. Modern bronchoscopic techniques deliver NIR light to any intrapulmonary lesion. This bronchoscopic technique can be navigated using a 3D navigation system based on previous CT images to precisely guide the fiber to the tumor in the lung, thereby minimizing damage to the normal lung field.30
More recently, a new catheter with LEDs was developed to remedy the problems of conventional devices, which risk twisting during deep insertion. The new catheter with LEDs mounted on a flexible printed circuit board is flexible, does not twist, and allows for deep insertion and irradiation using an endoscope or guidewire. It also has a temperature sensor to prevent thermal injury. Tests on mice with cholangiocarcinoma xenografts demonstrate the effectiveness of this new device.31
However, attention should be paid to the risk of thermal injury caused by light irradiation. According to the report by Okuyama et al, thermal damage with skin burns was observed in NIR-PIT when the power density of light exceeded 600 mW/cm2. In contrast, it was demonstrated that cooling the irradiation surface with air, or lowering the power density while extending the irradiation time to maintain the total energy, could prevent thermal injury while preserving therapeutic efficacy.245 This finding provides important implications for the safe design of various devices, including LED-mounted catheters and wireless LEDs.
Other developments include ET-BLIT, which delivers light deep into the body. The ET-BLIT catheter system consists of a catheter tip with a transparent distal end attached to a thermocouple head that can fit multiple vessels and a distal portion with a light-emitting function, which consists of two distinctive components: (i) a tip-partially transparent catheter with a transparent distal end connected to a thermocouple head, with an outer diameter of approximately 1–5 mm to accommodate multiple vessels, and (ii) a light diffuser equipped with a luminescent function at its distal portion. Once the catheter is navigated to the target lesion using the guidewire, the light diffuser is introduced into the lumen of the tip-partially transparent catheter. To confirm light emission, the catheter allows manual adjustment by rotating the light diffuser toward the lesion. The basic procedure of this system is comparable to that of existing endovascular therapies and is a promising way to bring light anywhere in the body for clinical use. In addition, tests in animal models have proven the effectiveness of this system, with NIR light penetrating the walls of blood vessels and reaching the liver and kidneys without causing temperature increase, vascular damage, or changes in blood composition in the big artery.32 This technology can produce better therapeutic effects than single-treatment repetitive therapy,246,247 which would be less invasive.
An implantable, wireless-driven LED has also been developed to limit the invasive nature of repetitive treatments in deep tissues while maintaining efficacy. The power supplied to the embedded LED is replenished by electromagnetic induction from an external transmitter coil coupled to the LED to an embedded receiver coil. In vitro and implanted tumor-bearing mouse tests revealed that this device may be a possible solution for treating tumors in deep areas of the human body, in vitro, and in implanted tumor-bearing mice.32
Thus, additional light irradiation devices are being developed to enable localized NIR-PIT. The use of a variety of advanced devices combined with other treatment modalities will make it possible to select the appropriate local NIR-PIT for a patient’s disease.
Challenges and Limitations of Pulmonary NIR-PIT
Although NIR-PIT has several theoretical and experimental advantages for lung cancer treatment, there are still important challenges that must be overcome before its broad clinical application to pulmonary lesions.12,248 The most critical issue is light delivery. Unlike superficial tumors or lesions directly accessible from the body surface, many lung cancers are located deep within the thoracic cavity and are surrounded by complex anatomical structures, including bronchi, pulmonary vessels, pleura, ribs, and mediastinal organs. Therefore, sufficient and homogeneous irradiation of the entire tumor volume can be technically difficult, particularly for peripheral lesions or tumors located far from the central airway. To address this limitation, several light-delivery strategies, including endoscopic, interstitial, and endovascular approaches, have been investigated as potential methods to expand the applicability of NIR-PIT to deep-seated tumors.30,32
Another important limitation is the treatment of multiple pulmonary lesions. Lung cancer frequently presents with multifocal disease, intrapulmonary metastases, pleural dissemination, or microscopic residual lesions. Because NIR-PIT requires both selective accumulation of antibody–photoabsorber conjugates and local light irradiation, untreated lesions outside the illuminated field may remain viable. Thus, pulmonary NIR-PIT may initially be more suitable for selected local lesions, such as centrally located tumors, endobronchial tumors, pleural dissemination, or tumors accessible by endoscopic, interstitial, or endovascular light-delivery approaches. Previous preclinical studies have demonstrated the feasibility of NIR-PIT in lung metastasis or thoracic cancer models, but these findings also underscore the importance of defining which lesion types are technically accessible to light irradiation.56,63 For multifocal lung disease, strategies such as repeated irradiation, multiple light-delivery routes, image-guided illumination, or combination with systemic therapies may be required.
Respiratory motion is another specific challenge in lung cancer treatment. During irradiation, the target lesion can move due to breathing, cardiac motion, or patient positioning. This displacement may reduce the accuracy of light delivery and cause insufficient irradiation of the tumor margin. In addition, excessive irradiation of surrounding normal tissues should be avoided, especially in fragile pulmonary structures. In radiotherapy for lung cancer, respiratory motion management, four-dimensional imaging, respiratory gating, and image-guided techniques have been developed to improve dose accuracy and reduce exposure to normal tissues.249,250 Similar concepts may be important for pulmonary NIR-PIT. Therefore, respiratory motion management, real-time imaging guidance, respiratory gating, and flexible or catheter-based light-delivery devices may be necessary to improve the precision and reproducibility of pulmonary NIR-PIT.
Tumor heterogeneity also represents a major biological limitation. The efficacy of NIR-PIT depends on the expression of target antigens on the tumor cell surface. However, lung cancers are highly heterogeneous, and antigen expression may differ among histological subtypes, metastatic sites, individual lesions, and even within the same tumor. Large-scale multiregion sequencing studies, including TRACERx, have demonstrated substantial intratumor heterogeneity and branched evolutionary patterns in non-small cell lung cancer, which can contribute to treatment resistance and disease progression.251,252 Incomplete antigen expression can result in residual antigen-negative tumor cells after treatment. Therefore, careful selection of target antigens, assessment of antigen expression before treatment, and the development of multi-target or sequential NIR-PIT strategies may be important for improving therapeutic efficacy.
The immune response induced by NIR-PIT should also be considered in the pulmonary tumor microenvironment. NIR-PIT can induce immunogenic cell death and may enhance antitumor immunity; however, the lung tumor microenvironment often contains immunosuppressive cells and stromal barriers that may limit durable immune responses.23,253 In this context, antibody-mediated immune mechanisms may be relevant. Gu et al recently highlighted the importance of antibody-dependent cellular phagocytosis in cancer immunotherapy, emphasizing the role of macrophages and Fc receptor-mediated immune responses.254 These findings suggest that, in addition to direct phototoxicity, the immunological properties of antibody-based therapeutics may influence the therapeutic outcome of NIR-PIT. Future studies should clarify how NIR-PIT interacts with macrophages, dendritic cells, and other immune components in lung cancer.
Another emerging strategy to improve the selectivity and flexibility of NIR-PIT is pre-targeting. Conventional NIR-PIT requires direct conjugation of IR700 to a target-specific antibody, which may limit the flexibility of target selection and pharmacokinetic optimization. Mohiuddin et al reported a synthetic zipper-mediated pre-targeting system for NIR-PIT, in which an IR700-conjugated Zip1-SNAP protein was combined with scFv-ZIP2 fusion proteins targeting tumor-associated molecules.255 Such pre-targeting approaches may help separate tumor targeting from photoabsorber delivery, potentially improving tumor selectivity, reducing off-target exposure, and expanding the range of applicable target molecules. Although further validation in lung cancer models is required, this concept may be useful for addressing antigen heterogeneity and optimizing the therapeutic window of pulmonary NIR-PIT.
Overall, pulmonary NIR-PIT should not be regarded as a universally applicable treatment for all lung cancer lesions at the present stage. Its clinical translation will require optimization of target selection, antibody–photoabsorber conjugates, light-delivery devices, imaging guidance, respiratory motion management, and combination strategies. Addressing these technical and biological challenges will be essential for establishing NIR-PIT as a practical therapeutic option for selected patients with lung cancer.
Conclusion
In this review, we summarized the recent development of NIR-PIT and its potential application to lung cancer. Compared with conventional PDT, NIR-PIT offers several advantages, including target-specific cytotoxicity mediated by antibody–photoabsorber conjugates, rapid light-induced membrane damage, and the potential to induce antitumor immune responses.12,248 Preclinical studies using lung cancer and thoracic malignancy models have suggested that several molecular targets, including EGFR, HER2, PDPN, MSLN, GPR87, DLL3, and MUC1, may be applicable to NIR-PIT-based strategies.24,56,64,107,256,257
However, the clinical translation of NIR-PIT for lung cancer remains at an early stage. Unlike head and neck cancer, in which clinical experience has already provided proof of concept for NIR-PIT in humans, direct clinical evidence in lung cancer is still limited.258,259 Several challenges specific to pulmonary application must be addressed, including light delivery to deep or multiple lung lesions, respiratory motion during irradiation, anatomical complexity of the thoracic cavity, heterogeneous antigen expression, and the need for image-guided treatment strategies.32,249,251 In addition, the interaction between NIR-PIT and the lung tumor microenvironment, including antibody-mediated immune mechanisms such as antibody-dependent cellular phagocytosis, should be further investigated.254
Future advances in target selection, antibody engineering, pre-targeting systems, light-delivery devices, and respiratory motion management may expand the therapeutic potential of NIR-PIT for selected lung cancer patients.30,32,251,255 In particular, novel approaches such as synthetic zipper-mediated pre-targeting systems may provide additional flexibility for optimizing tumor selectivity and photoabsorber delivery.255 Therefore, while pulmonary NIR-PIT is a promising therapeutic candidate, further preclinical and clinical studies are required to establish its efficacy, safety, and optimal clinical indications in lung cancer.
Abbreviations
NIR-PIT, Near-infrared photoimmunotherapy; ROS, Reactive oxygen species; PDT, Photodynamic therapy; LED, Light emitting diode; EGFR, Epidermal growth factor receptor; NIR, Near-infrared; APC, Antibody-photoabsorber conjugate; IR700, IRDye700DX; PS, Photosensitizer; DNA, Deoxyribonucleic acid; ICD, Immunogenic cell death; ADC, Antibody-drug conjugate; HNSCC, Head and neck squamous cell carcinoma; FDA, Food and Drug Administration; PMDA, Pharmaceuticals and Medical Devices Agency; DAMPs, Damage-associated molecular patterns; ET-BLIT, Endovascular therapy-based light illumination technology; mAb, Monoclonal antibody; ErbB, Erythroblastosis oncogene B; HNC, Head and neck cancer; HER2, Human epidermal growth factor receptor 2; TKI, Tyrosine kinase inhibitor; ADCC, Antibody-dependent cell-mediated cytotoxicity; NSCLC, Non-small cell lung cancer; CDDP, Cis-diamminedichloro-platinum(II); T-DM1, Trastuzumab emtansine; AML, Acute myeloid leukemia; PD-1, Programmed death 1; CTLA4, Cytotoxic T-lymphocyte-associated protein 4; CRC, Colorectal cancer; RIT, Radioimmunotherapy; NK, Natural killer; PDPN, Podoplanin; CAF, Cancer-associated fibroblast; MPM, Malignant pleural mesothelioma; MSLN, Mesothelin; GPI, Glycophosphatidylinositol; GPCR, G protein-coupled receptor; SCLC, Small cell lung cancer; DLL3, Delta-like canonical Notch ligand 3; TME, Tumor microenvironment; CDC, Complement-dependent cytotoxicity; PD-L1, Programmed death ligand 1; EMA, European Medicines Agency; MDSC, Murine myeloid-derived suppressor cell; TAM, Tumor-associated macrophage; AMP, Adenosine monophosphate; MRP1, Multidrug resistance-associated protein 1; LTC4, Leukotriene C4; VISTA, V-domain immunoglobulin suppressor of T cell activation; GvHD, Graft-versus-host disease.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this research.
Funding
The research was supported by the Advanced Research Infrastructure for Materials and Nanotechnology in Japan (ARIM JAPAN of MEXT), KAKEN; 21K07217, 25K03451, 25K22916 (JSPS), FOREST-Souhatsu (JPMJFR2017, JST), AMED SeedsA (26ym0126807j0005, A-226, AMED), JST GAP FUND TONGALI, and research scholarships from Toyota Physical and Chemical Research Institute, Sumitomo Electric Group CSR Foundation, Takeda Science Foundation, SGH Foundation, Japanese Society for Thermal Medicine, Research Grant from The Chemo-Sero-Therapeutic Research Institute, Suzuki-kenzo scholarship, the Hori Science & Arts Foundation, Casio foundation, the Naito Science & Engineering Fondation, Iketani Foundation and TERUMO Science Foundation. Funders provided only financial support and had no role in the study design, data collection, data analysis, interpretation, or writing of the reports.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Forde PM, Spicer J, Lu S, et al. Neoadjuvant nivolumab plus chemotherapy in resectable lung cancer. N Engl J Med. 2022;386(21):1973–24. doi:10.1056/NEJMoa2202170
2. Kordiak J, Bielec F, Jabłoński S, Pastuszak-Lewandoska D. Role of beta-carotene in lung cancer primary chemoprevention: a systematic review with meta-analysis and meta-regression. Nutrients. 2022;14(7):1361. doi:10.3390/NU14071361
3. Hayata Y, Kato H, Konaka C, Ono J, Takizawa N. Hematoporphyrin derivative and laser photoradiation in the treatment of lung cancer. Chest. 1982;81(3):269–277. doi:10.1378/CHEST.81.3.269
4. Wang K, Yu B, Pathak JL. An update in clinical utilization of photodynamic therapy for lung cancer. J Cancer. 2021;12(4):1154–1160. doi:10.7150/JCA.51537
5. Castano AP, Demidova TN, Hamblin MR. Mechanisms in photodynamic therapy: part one—photosensitizers, photochemistry and cellular localization. Photodiagnosis Photodyn Ther. 2004;1(4):279–293. doi:10.1016/S1572-1000(05)00007-4
6. Kwiatkowski S, Knap B, Przystupski D, et al. Photodynamic therapy – mechanisms, photosensitizers and combinations. Biomed Pharmacother. 2018;106:1098–1107. doi:10.1016/j.biopha.2018.07.049
7. Benov L. Photodynamic therapy: current status and future directions. Med Princ Pract. 2015;24(1):14–28. doi:10.1159/000362416
8. Crous A, Abrahamse H. Photodynamic therapy of lung cancer, where are we? Front Pharmacol. 2022;13:932098. doi:10.3389/fphar.2022.932098
9. van Straten D, Mashayekhi V, de Bruijn HS, Oliveira S, Robinson DJ. Oncologic photodynamic therapy: basic principles, current clinical status and future directions. Cancers. 2017;9(2):19. doi:10.3390/CANCERS9020019
10. Sato K, Gorka AP, Nagaya T, et al. Effect of charge localization on the in vivo optical imaging properties of near-infrared cyanine dye/monoclonal antibody conjugates. Mol Biosyst. 2016;12(10):3046–3056. doi:10.1039/C6MB00371K
11. Watanabe R, Sato K, Hanaoka H, et al. Minibody-indocyanine green based activatable optical imaging probes: the role of short polyethylene glycol linkers. ACS Med Chem Lett. 2014;5(4):411–415. doi:10.1021/jacsau.2c00243
12. Kobayashi H, Choyke PL. Near-Infrared photoimmunotherapy of cancer. Acc Chem Res. 2019;52(8):2332–2339. doi:10.1021/acs.accounts.9b00273
13. Hemmer E, Benayas A, Légaré F, Vetrone F. Exploiting the biological windows: current perspectives on fluorescent bioprobes emitting above 1000 nm. Nanoscale Horiz. 2016;1(3):168–184. doi:10.1039/C5NH00073D
14. Mitsunaga M, Ogawa M, Kosaka N, Rosenblum LT, Choyke PL, Kobayashi H. Cancer cell-selective in vivo near infrared photoimmunotherapy targeting specific membrane molecules. Nat Med. 2011;17(12):1685–1691. doi:10.1038/nm.2554
15. Sato K, Nakajima T, Choyke PL, Kobayashi H. Selective cell elimination in vitro and in vivo from tissues and tumors using antibodies conjugated with a near infrared phthalocyanine. RSC Adv. 2015;5(32):25105–25114. doi:10.1039/C4RA13835J
16. Wei D, Qi J, Hamblin MR, Wen X, Jiang X, Yang H. Near-infrared photoimmunotherapy: design and potential applications for cancer treatment and beyond. Theranostics. 2022;12(16):7108–7131. doi:10.7150/THNO.74820
17. Shirasu N, Yamada H, Shibaguchi H, Kuroki M, Kuroki M. Potent and specific antitumor effect of CEA-targeted photoimmunotherapy. Int, J, Cancer. 2014;135(11):2697–2710. doi:10.1002/IJC.28907
18. Sato K, Okada T, Okada R, et al. Photoinduced actin aggregation involves cell death: a mechanism of cancer cell cytotoxicity after near-infrared photoimmunotherapy. ACS Nano. 2025;19(8):8338–8356. doi:10.1021/acsnano.5c00104
19. Sato K, Ando K, Okuyama S, et al. Photoinduced ligand release from a silicon phthalocyanine dye conjugated with monoclonal antibodies: a mechanism of cancer cell cytotoxicity after near-infrared photoimmunotherapy. ACS Cent Sci. 2018;4(11):1559–1569. doi:10.1021/acscentsci.8b00565
20. Maruoka Y, Nagaya T, Nakamura Y, et al. Evaluation of early therapeutic effects after Near-Infrared Photoimmunotherapy (NIR-PIT) using luciferase-luciferin photon-counting and fluorescence imaging. Mol Pharm. 2017;14(12):4628–4635. doi:10.1021/acs.molpharmaceut.7b00731
21. Okuyama S, Fujimura D, Furusawa A, et al. Urinary IR700 ligand as an early biomarker of therapeutic efficacy of near-infrared photoimmunotherapy. ACS Omega. 2025;10:6991. doi:10.1021/ACSOMEGA.4C09850
22. Inagaki FF, Fujimura D, Furusawa A, et al. Diagnostic imaging in near-infrared photoimmunotherapy using a commercially available camera for indocyanine green. Cancer Sci. 2021;112(3):1326–1330. doi:10.1111/cas.14809
23. Ogawa M, Tomita Y, Nakamura Y, et al. Immunogenic cancer cell death selectively induced by near infrared photoimmunotherapy initiates host tumor immunity. Oncotarget. 2017;8(6):10425–10436. doi:10.18632/ONCOTARGET.14425
24. Matsuoka K, Yamada M, Sato M, Sato K. Near-Infrared photoimmunotherapy for thoracic cancers: a translational perspective. Biomedicines. 2022;10(7):1662. doi:10.3390/BIOMEDICINES10071662
25. Shigemitsu H, Sato K, Hagio S, et al. Amphiphilic rhodamine nano-assembly as a type i supramolecular photosensitizer for photodynamic therapy. ACS Appl Nano Mater. 2022;5(10):14954–14960. doi:10.1021/acsanm.2c03192
26. Shigemitsu H, Ohkubo K, Sato K, et al. Fluorescein-Based type i supramolecular photosensitizer via induction of charge separation by self-assembly. JACS Au. 2022;2(6):1472–1478. doi:10.1021/jacsau.2c00243
27. Agostinis P, Berg K, Cengel KA, et al. Photodynamic therapy of cancer: an update. CA Cancer J Clin. 2011;61(4):250–281. doi:10.3322/caac.20114
28. Ash C, Dubec M, Donne K, Bashford T. Effect of wavelength and beam width on penetration in light-tissue interaction using computational methods. Lasers Med Sci. 2017;32(8):1909–1918. doi:10.1007/S10103-017-2317-4
29. Cognetti DM, Johnson JM, Curry JM, et al. Phase 1/2a, open-label, multicenter study of RM-1929 photoimmunotherapy in patients with locoregional, recurrent head and neck squamous cell carcinoma. Head Neck. 2021;43(12):3875–3887. doi:10.1002/HED.26885
30. Furumoto H, Kato T, Wakiyama H, Furusawa A, Choyke PL, Kobayashi H. Endoscopic Applications of Near-Infrared Photoimmunotherapy (NIR-PIT) in cancers of the digestive and respiratory tracts. Biomedicines. 2022;10(4):846. doi:10.3390/BIOMEDICINES10040846
31. Hirata H, Kuwatani M, Nakajima K, et al. Near-infrared photoimmunotherapy (NIR-PIT) on cholangiocarcinoma using a novel catheter device with light emitting diodes. Cancer Sci. 2021;112(2):828–838. doi:10.1111/cas.14780
32. Tsukamoto T, Fujita Y, Shimogami M, et al. Inside-the-body light delivery system using endovascular therapy-based light illumination technology. EBioMedicine. 2022:85. doi:10.1016/j.ebiom.2022.104289.
33. Gaber R, Watermann I, Kugler C, et al. Correlation of EGFR expression, gene copy number and clinicopathological status in NSCLC. Diagn Pathol. 2014;9(1):1–15. doi:10.1186/s13000-014-0165-0
34. Yokouchi H, Nishihara H, Harada T, et al. Immunohistochemical profiling of receptor tyrosine kinases, MED12, and TGF-βRII of surgically resected small cell lung cancer, and the potential of c-kit as a prognostic marker. Oncotarget. 2016;8(24):39711–39726. doi:10.18632/oncotarget.14410
35. Niederst MJ, Sequist LV, Poirier JT, et al. RB loss in resistant EGFR mutant lung adenocarcinomas that transform to small-cell lung cancer. Nat Commun. 2015;6(1):1–10. doi:10.1038/ncomms7377
36. Ren S, Wang J, Ying J, et al. Consensus for HER2 alterations testing in non-small-cell lung cancer. ESMO Open. 2022;7(1). doi:10.1016/j.esmoop.2022.100395
37. Lashkarizadeh M, Lashkarizadeh M, Nikian M, Kouhestani Parizi M. The expression of HER2/neu in patients with lung cancer and its associated factors. Clin Respir J. 2023;17(2):90. doi:10.1111/CRJ.13572
38. Leung ELH, Fiscus RR, Tung JW, et al. Non-Small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties. PLoS One. 2010;5(11):e14062. doi:10.1371/journal.pone.0014062
39. Hu B, Ma Y, Yang Y, Zhang L, Han H, Chen J. CD44 promotes cell proliferation in non-small cell lung cancer. Oncol Lett. 2018;15(4):5627. doi:10.3892/OL.2018.8051
40. Ariza A, Mate JL, Isamat M, et al. Standard and variant CD44 isoforms are commonly expressed in lung cancer of the non-small cell type but not of the small cell type. J Pathol. 1995;177(4):363–368. doi:10.1002/PATH.1711770406
41. Li J, Chen H, Li X, et al. Co-expression of podoplanin and fibroblast growth factor 1 predicts poor prognosis in patients with lung squamous cell carcinoma. Mol Med Rep. 2017;16(2):1643–1652. doi:10.3892/mmr.2017.6830
42. Terada T. Cytokeratin-negative small cell lung carcinoma. Rare Tumors. 2011;3(4):e38. doi:10.4081/RT.2011.E38
43. Weidemann S, Gagelmann P, Gorbokon N, et al. Mesothelin expression in human tumors: a tissue microarray study on 12,679 tumors. Biomedicines. 2021;9(4):397. doi:10.3390/biomedicines9040397
44. Nii K, Tokunaga Y, Liu D, et al. Overexpression of G protein-coupled receptor 87 correlates with poorer tumor differentiation and higher tumor proliferation in non-small-cell lung cancer. Mol Clin Oncol. 2014;2(4):539. doi:10.3892/mco.2014.292
45. Nii K, Liu D, Tokunaga Y, et al. Association of tumor-specific GPR87 overexpression with poor prognosis in non-small cell lung cancer. J Clin Oncol. 2013;31(15):e22108–e22108. doi:10.1200/jco.2013.31.15_suppl.e22108
46. Alì G, Di Stefano I, Poma AM, et al. Prevalence of delta-like protein 3 in a consecutive series of surgically resected lung neuroendocrine neoplasms. Front Oncol. 2021;11:729765. doi:10.3389/FONC.2021.729765/BIBTEX
47. Rojo F, Corassa M, Mavroudis D, et al. International real-world study of DLL3 expression in patients with small cell lung cancer. Lung Cancer. 2020;147:237–243. doi:10.1016/j.lungcan.2020.07.026
48. Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001;2(2):127–137. doi:10.1038/35052073
49. Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103(2):211–225. doi:10.1016/S0092-8674(00)00114-8
50. Ishitoya J, Toriyama M, Oguchi N, et al. Gene amplification and overexpression of EGF receptor in squamous cell carcinomas of the head and neck. Br J Cancer. 1989;59(4):559–562. doi:10.1038/BJC.1989.113
51. Kearsley JH, Furlong KL, Cooke RA, Waters MJ. An immunohistochemical assessment of cellular proliferation markers in head and neck squamous cell cancers. Br J Cancer. 1990;61(6):821–827. doi:10.1038/BJC.1990.184
52. Nicholson RI, Gee JMW, Harper ME. EGFR and cancer prognosis. Eur J Cancer. 2001;37(4):9–15. doi:10.1016/S0959-8049(01)00231-3
53. Okada R, Furusawa A, Vermeer DW, et al. Near-infrared photoimmunotherapy targeting human-EGFR in a mouse tumor model simulating current and future clinical trials. EBioMedicine. 2021:67. doi:10.1016/j.ebiom.2021.103345.
54. Fukushima H, Turkbey B, Pinto PA, Furusawa A, Choyke PL, Kobayashi H. Near-Infrared Photoimmunotherapy (NIR-PIT) in urologic cancers. Cancers. 2022;14(12):2996. doi:10.3390/CANCERS14122996
55. Ciardiello F, De Vita F, Orditura M, Tortora G. The role of EGFR inhibitors in nonsmall cell lung cancer. Curr Opin Oncol. 2004;16(2):130–135. doi:10.1097/00001622-200403000-00008
56. Nakamura Y, Ohler ZW, Householder D, et al. Near infrared photoimmunotherapy in a transgenic mouse model of spontaneous epidermal growth factor receptor (EGFR)-expressing lung cancer. Mol Cancer Ther. 2017;16(2):408–414. doi:10.1158/1535-7163.MCT-16-0663
57. Study details | study of RM-1929 and photoimmunotherapy in patients with recurrent head and neck cancer | clinicalTrials.gov. Available from: https://clinicaltrials.gov/study/NCT02422979.
58. Roskoski R. The ErbB/HER family of protein-tyrosine kinases and cancer. Pharmacol Res. 2014;79:34–74. doi:10.1016/J.PHRS.2013.11.002
59. Berman JD, Leishmaniasis IN, Chang KP, et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science. 1989;244(4905):707–712. doi:10.1126/SCIENCE.2470152
60. Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A, McGuire WL. Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science. 1987;235(4785):182–191. doi:10.1126/SCIENCE.3798106
61. Boku N. HER2-positive gastric cancer. Gastric Cancer. 2014;17(1):1–12. doi:10.1007/S10120-013-0252-Z
62. Gerson JN, Skariah S, Denlinger CS, Astsaturov I. Perspectives of HER2-targeting in gastric and esophageal cancer. Expert Opin Investig Drugs. 2017;26(5):531–540. doi:10.1080/13543784.2017.1315406
63. Sato K, Nagaya T, Nakamura Y, et al. Near infrared photoimmunotherapy prevents lung cancer metastases in a murine model. Oncotarget. 2015;6(23):19747–19758. doi:10.18632/ONCOTARGET.3850
64. Sato K, Nagaya T, Mitsunaga M, Choyke PL, Kobayashi H. Near infrared photoimmunotherapy for lung metastases. Cancer Lett. 2015;365(1):112–121. doi:10.1016/J.CANLET.2015.05.018
65. Sato K, Nagaya T, Choyke PL, Kobayashi H. Near infrared photoimmunotherapy in the treatment of pleural disseminated NSCLC: preclinical experience. Theranostics. 2015;5(7):698. doi:10.7150/THNO.11559
66. Takahashi K, Taki S, Yasui H, et al. HER2 targeting near-infrared photoimmunotherapy for a CDDP-resistant small-cell lung cancer. Cancer Med. 2021;10(24):8808–8819. doi:10.1002/CAM4.4381
67. Takahashi K, Yasui H, Taki S, et al. Near-infrared-induced drug release from antibody–drug double conjugates exerts a cytotoxic photo-bystander effect. Bioeng Transl Med. 2022;7(3):e10388. doi:10.1002/BTM2.10388
68. Gronthos S, Franklin DM, Leddy HA, Robey PG, Storms RW, Gimble JM. Surface protein characterization of human adipose tissue-derived stromal cells. J Cell Physiol. 2001;189(1):54–63. doi:10.1002/JCP.1138
69. Domev H, Amit M, Laevsky I, Dar A, Itskovitz-Eldor J. Efficient engineering of vascularized ectopic bone from human embryonic stem cell-derived mesenchymal stem cells. Tissue Eng Part A. 2012;18(21–22):2290–2302. doi:10.1089/ten.TEA.2011.0371
70. Thapa R, Wilson GD. The Importance of CD44 as a stem cell biomarker and therapeutic target in cancer. Stem Cells Int. 2016;2016(1):2087204. doi:10.1155/2016/2087204
71. Gadhoum Z, Delaunay J, Maquarre E, et al. The effect of anti-CD44 monoclonal antibodies on differentiation and proliferation of human acute myeloid leukemia cells. Leuk Lymphoma. 2004;45(8):1501–1510. doi:10.1080/1042819042000206687
72. Zheng Z, Katoh S, He Q, et al. Monoclonal antibodies to CD44 and their influence on hyaluronan recognition. J Cell Biol. 1995;130(2):485–495. doi:10.1083/JCB.130.2.485
73. Henke C, Bitterman P, Roongta U, Ingbar D, Polunovsky V. Induction of fibroblast apoptosis by anti-CD44 antibody: implications for the treatment of fibroproliferative lung disease. Am J Pathol. 1996;149(5):1639.
74. Cui Y, Xu Y, Li Y, et al. Antibody Drug Conjugates of Near-Infrared Photoimmunotherapy (NIR-PIT) in breast cancers. Technol Cancer Res Treat. 2023:22. doi:10.1177/15330338221145992.
75. Nagaya T, Nakamura Y, Okuyama S, et al. Syngeneic mouse models of oral cancer are effectively targeted by anti-cd44-based NIR-PIT. Mol Cancer Res. 2017;15(12):1667–1677. doi:10.1158/1541-7786.MCR-17-0333
76. Maruoka Y, Furusawa A, Okada R, et al. Interleukin-15 after Near-Infrared Photoimmunotherapy (NIR-PIT) enhances T cell response against syngeneic mouse tumors. Cancers. 2020;12(9):2575. doi:10.3390/cancers12092575
77. Suzuki H, Kaneko MK, Kato Y. Roles of podoplanin in malignant progression of tumor. Cells. 2022;11(3):575. doi:10.3390/CELLS11030575
78. Glatt S, Halbauer D, Heindl S, et al. hGPR87 contributes to viability of human tumor cells. Int, J, Cancer. 2008;122(9):2008–2016. doi:10.1002/ijc.23349
79. Niss Arfelt K, Fares S, Sparre-Ulrich AH, et al. Signaling via G proteins mediates tumorigenic effects of GPR87. Cell Signal. 2017;30:9–18. doi:10.1016/j.cellsig.2016.11.009
80. Mishima K, Kato Y, Kaneko MK, et al. Podoplanin expression in primary central nervous system germ cell tumors: a useful histological marker for the diagnosis of germinoma. Acta Neuropathol. 2006;111(6):563–568. doi:10.1007/s00401-006-0033-4
81. Mishima K, Kato Y, Kaneko MK, Nishikawa R, Hirose T, Matsutani M. Increased expression of podoplanin in malignant astrocytic tumors as a novel molecular marker of malignant progression. Acta Neuropathol. 2006;111(5):483–488. doi:10.1007/s00401-006-0063-y
82. Abe S, Morita Y, Kaneko MK, et al. A novel targeting therapy of malignant mesothelioma using anti-podoplanin antibody. J Immunol. 2013;190(12):6239–6249. doi:10.4049/JIMMUNOL.1300448
83. Takagi S, Oh-Hara T, Sato S, Gong B, Takami M, Fujita N. Expression of Aggrus/podoplanin in bladder cancer and its role in pulmonary metastasis. Int, J, Cancer. 2014;134(11):2605–2614. doi:10.1002/ijc.28602
84. Ariizumi T, Ogose A, Kawashima H, et al. Expression of podoplanin in human bone and bone tumors: new marker of osteogenic and chondrogenic bone tumors. Pathol Int. 2010;60(3):193–202. doi:10.1111/j.1440-1827.2009.02510.x
85. Chang MC, Vargas SO, Hornick JL, Hirsch MS, Crum CP, Nucci MR. Embryonic stem cell transcription factors and D2-40 (podoplanin) as diagnostic immunohistochemical markers in ovarian germ cell tumors. Int J Gynecol Pathol. 2009;28(4):347–355. doi:10.1097/PGP.0b013e318195da86
86. Kato Y, Sasagawa I, Kaneko M, Osawa M, Fujita N, Tsuruo T. Aggrus: a diagnostic marker that distinguishes seminoma from embryonal carcinoma in testicular germ cell tumors. Oncogene. 2004;23(52):8552–8556. doi:10.1038/sj.onc.1207869
87. Sasaki K, Sugai T, Ishida K, et al. Analysis of cancer-associated fibroblasts and the epithelial-mesenchymal transition in cutaneous basal cell carcinoma, squamous cell carcinoma, and malignant melanoma. Hum Pathol. 2018;79:1–8. doi:10.1016/J.HUMPATH.2018.03.006
88. Hoshino A, Ishii G, Ito T, et al. Podoplanin-positive fibroblasts enhance lung adenocarcinoma tumor formation: podoplanin in fibroblast functions for tumor progression. Cancer Res. 2011;71(14):4769–4779. doi:10.1158/0008-5472.CAN-10-3228
89. Suzuki J, Aokage K, Neri S, et al. Relationship between podoplanin-expressing cancer-associated fibroblasts and the immune microenvironment of early lung squamous cell carcinoma. Lung Cancer. 2021;153:1–10. doi:10.1016/J.LUNGCAN.2020.12.020
90. Pula B, Jethon A, Piotrowska A, et al. Podoplanin expression by cancer-associated fibroblasts predicts poor outcome in invasive ductal breast carcinoma. Histopathology. 2011;59(6):1249–1260. doi:10.1111/j.1365-2559.2011.04060.x
91. Shindo K, Aishima S, Ohuchida K, et al. Podoplanin expression in cancer-associated fibroblasts enhances tumor progression of invasive ductal carcinoma of the pancreas. Mol Cancer. 2013;12(1):1–16. doi:10.1186/1476-4598-12-168
92. Nishinaga Y, Sato K, Yasui H, et al. Targeted phototherapy for malignant pleural mesothelioma: near-infrared photoimmunotherapy targeting podoplanin. Cells. 2020;9(4):1019. doi:10.3390/CELLS9041019
93. Scholler N, Fu N, Yang Y, et al. Soluble member(s) of the mesothelin/megakaryocyte potentiating factor family are detectable in sera from patients with ovarian carcinoma. Proc Natl Acad Sci U S A. 1999;96(20):11531–11536. doi:10.1073/pnas.96.20.11531
94. Chang K, Pastan I. Molecular cloning of mesothelin, a differentiation antigen present on mesothelium, mesotheliomas, and ovarian cancers. Proc Natl Acad Sci. 1996;93(1):136–140. doi:10.1073/PNAS.93.1.136
95. He X, Wang L, Riedel H, et al. Mesothelin promotes epithelial-to-mesenchymal transition and tumorigenicity of human lung cancer and mesothelioma cells. Mol Cancer. 2017;16(1):1–13. doi:10.1186/S12943-017-0633-8
96. Morello A, Sadelain M, Adusumilli PS. Mesothelin-targeted CARs: driving T cells to solid tumors. Cancer Discov. 2016;6(2):133–146. doi:10.1158/2159-8290.CD-15-0583
97. Somers EB, O’Shannessy DJ. Folate receptor alpha, mesothelin and megakaryocyte potentiating factor as potential serum markers of chronic kidney disease. Biomark Insights. 2014;9:S15245. doi:10.4137/BMI.S15245
98. Zhang YF, Phung Y, Gao W, et al. New high affinity monoclonal antibodies recognize non-overlapping epitopes on mesothelin for monitoring and treating mesothelioma. Sci Rep. 2015;5(1):1–14. doi:10.1038/srep09928
99. Nagaya T, Nakamura Y, Sato K, et al. Near infrared photoimmunotherapy with an anti-mesothelin antibody. Oncotarget. 2016;7(17):23361. doi:10.18632/ONCOTARGET.8025
100. Sak K. The path of GPR87: from a P2Y-like receptor to its role in cancer progression. Naunyn Schmiedebergs Arch Pharmacol. 2024;398(5):4803–4815. doi:10.1007/S00210-024-03684-6
101. Zhang Y, Qian Y, Lu W, Chen X. The G protein-coupled receptor 87 is necessary for p53-dependent cell survival in response to genotoxic stress. Cancer Res. 2009;69(15):6049–6056. doi:10.1158/0008-5472.CAN-09-0621
102. Kazuhito N, Yoshimasa T, Dage L, et al. Overexpression of G protein-coupled receptor 87 correlates with poorer tumor differentiation and higher tumor proliferation in non-small-cell lung cancer. Mol Clin Oncol. 2014;2(4):539–544. doi:10.3892/mco.2014.292
103. Park SM, Choi EY, Bae M, et al. Histone variant H3F3A promotes lung cancer cell migration through intronic regulation. Nat Commun. 2016;7(1):1–14. doi:10.1038/ncomms12914
104. Cohen AS, Khalil FK, Welsh EA, et al. Cell-surface marker discovery for lung cancer. Oncotarget. 2017;8(69):113373. doi:10.18632/ONCOTARGET.23009
105. Ahn HM, Choi EY, Kim YJ. Gpr87 promotes metastasis through the akt-enos-no axis in lung adenocarcinoma. Cancers. 2022;14(1):19. doi:10.3390/CANCERS14010019/S1
106. Bai R, Zhang J, He F, et al. GPR87 promotes tumor cell invasion and mediates the immunogenomic landscape of lung adenocarcinoma. Commun Biol. 2022;5(1):1–12. doi:10.1038/s42003-022-03506-6
107. Yasui H, Nishinaga Y, Taki S, et al. Near-infrared photoimmunotherapy targeting GPR87: development of a humanised anti-GPR87 mAb and therapeutic efficacy on a lung cancer mouse model. EBioMedicine. 2021;67:103372. doi:10.1016/j.ebiom.2021.103372
108. Wang H, Zheng T, Xu D, Sun C, Huang D, Liu X. Targeting DLL3: innovative strategies for tumor treatment. Pharmaceutics. 2025;17(4):520. doi:10.3390/PHARMACEUTICS17040520
109. Saunders LR, Bankovich AJ, Anderson WC, et al. A DLL3-targeted antibody-drug conjugate eradicates high-grade pulmonary neuroendocrine tumor-initiating cells in vivo. Sci Transl Med. 2015;7(302).
110. Ranallo N, Bocchini M, Menis J, et al. Delta-like ligand 3 (DLL3): an attractive actionable target in tumors with neuroendocrine origin. Expert Rev Anticancer Ther. 2022;22(6):597–603. doi:10.1080/14737140.2022.2071703
111. Puca L, Gavyert K, Sailer V, et al. Delta-like protein 3 expression and therapeutic targeting in neuroendocrine prostate cancer. Sci Transl Med. 2019;11(484):891. doi:10.1126/scitranslmed.aav0891
112. Shrestha P, Kao S, Cheung VK, et al. Circulating tumor cells: advancing personalized therapy in small cell lung cancer patients. Mol Oncol. 2024. doi:10.1002/1878-0261.13696
113. Obermayr E, Agreiter C, Schuster E, et al. Molecular characterization of circulating tumor cells enriched by a microfluidic platform in patients with small-cell lung cancer. Cells. 2019;8(8):880. doi:10.3390/CELLS8080880
114. Messaritakis I, Nikolaou M, Koinis F, et al. Characterization of DLL3-positive circulating tumor cells (CTCs) in patients with small cell lung cancer (SCLC) and evaluation of their clinical relevance during front-line treatment. Lung Cancer. 2019;135:33–39. doi:10.1016/j.lungcan.2019.06.025
115. Welsch E, Holzer B, Schuster E, et al. Prognostic significance of circulating tumor cells and tumor related transcripts in small cell lung cancer: a step further to clinical implementation. Int, J, Cancer. 2024;154(12):2189–2199. doi:10.1002/IJC.34886
116. Su PL, Chakravarthy K, Furuya N, et al. DLL3-guided therapies in small-cell lung cancer: from antibody-drug conjugate to precision immunotherapy and radioimmunotherapy. Mol Cancer. 2024;23(1):1–5. doi:10.1186/s12943-024-02012-z
117. Isobe Y, Sato K, Nishinaga Y, et al. Near infrared photoimmunotherapy targeting DLL3 for small cell lung cancer. EBioMedicine. 2020;52:139–140. doi:10.1016/j.ebiom.2020.102632
118. Rudin CM, Pietanza MC, Bauer TM, et al. Rovalpituzumab tesirine, a DLL3-targeted antibody-drug conjugate, in recurrent small-cell lung cancer: a first-in-human, first-in-class, open-label, phase 1 study. Lancet Oncol. 2017;18(1):42–51. doi:10.1016/S1470-2045(16)30565-4
119. Haist M, Stege H, Grabbe S, Bros M. The functional crosstalk between myeloid-derived suppressor cells and regulatory T cells within the immunosuppressive tumor microenvironment. Cancers. 2021;13(2):210. doi:10.3390/CANCERS13020210
120. Webb ES, Liu P, Baleeiro R, Lemoine NR, Yuan M, Wang Y. Immune checkpoint inhibitors in cancer therapy. J Biomed Res. 2017;32(5):317. doi:10.7555/JBR.31.20160168
121. Kasprzak A, Olejniczak K, Przybyszewska W, Zabel M. Cellular expression of interleukin 2 (IL-2) and its receptor (IL-2R, CD25) in lung tumours. Folia Morphol. 2007;66(3):159–166.
122. Yano T, Fukuyama Y, Yokoyama H, et al. Interleukin-2 receptors in pulmonary adenocarcinoma tissue. Lung Cancer. 1996;16(1):13–19. doi:10.1016/S0169-5002(96)00608-3
123. Meazza R, Marciano S, Sforzini S, et al. Analysis of IL-2 receptor expression and of the biological effects of IL-2 gene transfection in small-cell lung cancer. Br J Cancer. 1996;74(5):788. doi:10.1038/BJC.1996.437
124. Dietel M, Savelov N, Salanova R, et al. Real-world prevalence of programmed death ligand 1 expression in locally advanced or metastatic non–small-cell lung cancer: the global, multicenter EXPRESS study. Lung Cancer. 2019;134:174–179. doi:10.1016/J.LUNGCAN.2019.06.012
125. Acheampong E, Abed A, Morici M, et al. Tumour PD-L1 expression in small-cell lung cancer: a systematic review and meta-analysis. Cells. 2020;9(11):2393. doi:10.3390/CELLS9112393
126. Amer BS, Amer RS, Awad RA, Darwish SA, Bedeer AE, Saed Amer B. Association of CTLA4 and PD-L1 immunohistchemical expression with various clinicopathological parameters and survival in non–small-cell lung cancer. Egyptian J Cancer Biomed Res. 2022;6(4):1–12. doi:10.21608/JCBR.2022.168536.1282
127. Paulsen EE, Kilvaer TK, Rakaee M, et al. CTLA-4 expression in the non-small cell lung cancer patient tumor microenvironment: diverging prognostic impact in primary tumors and lymph node metastases. Cancer Immunol Immunother. 2017;66(11):1449. doi:10.1007/S00262-017-2039-2
128. Regzedmaa O, Li Y, Li Y, et al. Prevalence of DLL3, CTLA-4 and MSTN expression in patients with small cell lung cancer. Onco Targets Ther. 2019;12:10043–10055. doi:10.2147/OTT.S216362
129. Dutta R, Rathor A, Sharma HP, et al. Mapping the immune landscape in small cell lung cancer by analysing expression of immuno-modulators in tissue biopsies and paired blood samples. Sci Rep. 2023;13(1):1–10. doi:10.1038/s41598-023-30841-3
130. Bergenfelz C, Leandersson K. The generation and identity of human myeloid-derived suppressor cells. Front Oncol. 2020;10:514774. doi:10.3389/FONC.2020.00109/FULL
131. Condamine T, Dominguez GA, Youn JI, et al. Lectin-type oxidized LDL receptor-1 distinguishes population of human polymorphonuclear myeloid-derived suppressor cells in cancer patients. Sci Immunol. 2016;1(2). doi:10.1126/sciimmunol.aaf8943
132. Inoue Y, Yoshimura K, Kurabe N, et al. Prognostic impact of CD73 and A2A adenosine receptor expression in non-small-cell lung cancer. Oncotarget. 2017;8(5):8738–8751. doi:10.18632/ONCOTARGET.14434
133. Kowash RR, Akbay EA. Tumor intrinsic and extrinsic functions of CD73 and the adenosine pathway in lung cancer. Front Immunol. 2023;14:1130358. doi:10.3389/FIMMU.2023.1130358
134. Xu G, Mo Y, Li J, Wei Q, Zhou F, Chen J. Two tripartite classification systems of CD86+ and CD206+ macrophages are significantly associated with tumor recurrence in stage II-III colorectal cancer. Front Immunol. 2023;14:1136875. doi:10.3389/fimmu.2023.1136875
135. Xu F, Wei Y, Tang Z, Liu B, Dong J. Tumor-associated macrophages in lung cancer: friend or foe? Mol Med Rep. 2020;22(5):4107–4115. doi:10.3892/mmr.2020.11518
136. Yonesaka K, Haratani K, Takamura S, et al. B7-H3 negatively modulates CTL-Mediated cancer immunity. Clin Cancer Res. 2018;24(11):2653–2664. doi:10.1158/1078-0432.CCR-17-2852
137. Qiu MJ, Xia Q, Chen YB, et al. The expression of three negative co-stimulatory B7 family molecules in small cell lung cancer and their effect on prognosis. Front Oncol. 2021;11:600238. doi:10.3389/fonc.2021.600238
138. Abd El-Rehim DM, Abd-Elghany MI, Nazmy MH. Integrin-Linked kinase, snail and multidrug resistance protein 1: three concordant players in the progression of non-small cell lung cancer. J Egypt Natl Canc Inst. 2015;27(3):129–137. doi:10.1016/J.JNCI.2015.04.004
139. Wright SR, Boag AH, Valdimarsson G, et al. Immunohistochemical detection of multidrug resistance protein in human lung cancer and normal lung1. 1998.
140. Liu H, Huang J, Peng J, et al. Upregulation of the inwardly rectifying potassium channel Kir2.1 (KCNJ2) modulates multidrug resistance of small-cell lung cancer under the regulation of miR-7 and the Ras/MAPK pathway. Mol Cancer. 2015;14(1):1–19. doi:10.1186/S12943-015-0298-0
141. Villarroel-Espindola F, Yu X, Datar I, et al. Spatially resolved and quantitative analysis of Vista/pd-1h as a novel immunotherapy target in human non–small cell lung cancer. Clin Cancer Res. 2018;24(7):1562–1573. doi:10.1158/1078-0432.CCR-17-2542
142. Ferencz B, Török K, Pipek O, et al. Expression patterns of novel immunotherapy targets in intermediate- and high-grade lung neuroendocrine neoplasms. Cancer Immunol Immunother. 2024;73(6):1–15. doi:10.1007/S00262-024-03704-7
143. Shan F, Somasundaram A, Bruno TC, Workman CJ, Vignali DAA. Therapeutic targeting of regulatory T cells in cancer. Trends Cancer. 2022;8(11):944–961. doi:10.1016/J.TRECAN.2022.06.008
144. Cho JH, Lim YC. Prognostic impact of regulatory T cell in head and neck squamous cell carcinoma: a systematic review and meta-analysis. Oral Oncol. 2021;112:105084. doi:10.1016/J.ORALONCOLOGY.2020.105084
145. Sato K, Sato N, Xu B, et al. Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy. Sci Transl Med. 2016;8(352). doi:10.1126/scitranslmed.aaf6843
146. Okada R, Maruoka Y, Furusawa A, et al. The effect of antibody fragments on CD25 targeted regulatory T cell near-infrared photoimmunotherapy. Bioconjug Chem. 2019;30(10):2624–2633. doi:10.1021/acs.bioconjchem.9b00547
147. Maruoka Y, Furusawa A, Okada R, et al. Combined CD44- And CD25-targeted near-infrared photoimmunotherapy selectively kills cancer and regulatory T cells in syngeneic mouse cancer models. Cancer Immunol Res. 2020;8(3):345–355. doi:10.1158/2326-6066.CIR-19-0517
148. Sun C, Mezzadra R, Schumacher TN. Regulation and Function of the PD-L1 Checkpoint. Immunity. 2018;48(3):434–452. doi:10.1016/j.immuni.2018.03.014
149. Sanmamed MF, Chen L. Inducible expression of B7-H1 (PD-L1) and its selective role in tumor site immune modulation. Cancer J. 2014;20(4):256–261. doi:10.1097/PPO.0000000000000061
150. Herbst RS, Soria JC, Kowanetz M, et al. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature. 2014;515(7528):515(7528):563–567. doi:10.1038/nature14011
151. Gao Q, Wang XY, Qiu SJ, et al. Overexpression of PD-L1 significantly associates with tumor aggressiveness and postoperative recurrence in human hepatocellular carcinoma. Clin Cancer Res. 2009;15(3):971–979. doi:10.1158/1078-0432.CCR-08-1608
152. Hamanishi J, Mandai M, Iwasaki M, et al. Programmed cell death 1 ligand 1 and tumor-infiltrating CD8+ T lymphocytes are prognostic factors of human ovarian cancer. Proc Natl Acad Sci U S A. 2007;104(9):3360–3365. doi:10.1073/pnas.0611533104
153. Mu CY, Huang JA, Chen Y, Chen C, Zhang XG. High expression of PD-L1 in lung cancer may contribute to poor prognosis and tumor cells immune escape through suppressing tumor infiltrating dendritic cells maturation. Med Oncol. 2011;28(3):682–688. doi:10.1007/S12032-010-9515-2
154. Qu J, Mei Q, Liu L, et al. The progress and challenge of anti-PD-1/PD-L1 immunotherapy in treating non-small cell lung cancer. Ther Adv Med Oncol. 2021:13. doi:10.1177/1758835921992968.
155. Taki S, Matsuoka K, Nishinaga Y, et al. Spatiotemporal depletion of tumor-associated immune checkpoint PD-L1 with near-infrared photoimmunotherapy promotes antitumor immunity. J Immunother Cancer. 2021;9(11):e003036. doi:10.1136/JITC-2021-003036
156. Nagaya T, Nakamura Y, Sato K, et al. Near infrared photoimmunotherapy with avelumab, an anti-programmed death-ligand 1 (PD-L1) antibody. Oncotarget. 2016;8(5):8807. doi:10.18632/ONCOTARGET.12410
157. Jin J, Sivakumar I, Mironchik Y, et al. PD-L1 near infrared photoimmunotherapy of ovarian cancer model. Cancers. 2022;14(3):619. doi:10.3390/CANCERS14030619/S1
158. Inagaki FF, Kano M, Furusawa A, et al. Near-infrared photoimmunotherapy targeting PD-L1: improved efficacy by preconditioning the tumor microenvironment. Cancer Sci. 2024;115(7):2396–2409. doi:10.1111/CAS.16195
159. Ling V, Wu PW, Finnerty HF, Sharpe AH, Gray GS, Collins M. Complete sequence determination of the mouse and human CTLA4 gene loci: cross-species DNA sequence similarity beyond exon borders. Genomics. 1999;60(3):341–355. doi:10.1006/GENO.1999.5930
160. Klocke K, Sakaguchi S, Holmdahl R, Wing K. Induction of autoimmune disease by deletion of CTLA-4 in mice in adulthood. Proc Natl Acad Sci U S A. 2016;113(17):E2383–E2392. doi:10.1073/pnas.1603892113
161. Paterson AM, Lovitch SB, Sage PT, et al. Deletion of CTLA-4 on regulatory T cells during adulthood leads to resistance to autoimmunity. J Exp Med. 2015;212(10):1603–1621. doi:10.1084/JEM.20141030
162. Read S, Malmström V, Powrie F. Cytotoxic T lymphocyte–associated antigen 4 plays an essential role in the function of Cd25+Cd4+ regulatory cells that control intestinal inflammation. J Exp Med. 2000;192(2):295–302. doi:10.1084/JEM.192.2.295
163. Darvin P, Toor SM, Sasidharan Nair V, Elkord E. Immune checkpoint inhibitors: recent progress and potential biomarkers. Exp Mol Med. 2018;50(12):1–11. doi:10.1038/s12276-018-0191-1
164. Kvistborg P, Philips D, Kelderman S, et al. Anti-CTLA-4 therapy broadens the melanoma-reactive CD8+ T cell response. Sci Transl Med. 2014;6(254). doi:10.1126/scitranslmed.3008918
165. Kato T, Okada R, Furusawa A, et al. Simultaneously combined cancer cell- and CTLA4-targeted NIR-PIT causes a synergistic treatment effect in syngeneic mouse models. Mol Cancer Ther. 2021;20:2262–2273. doi:10.1158/1535-7163.MCT-21-0470
166. Okada R, Kato T, Furusawa A, et al. Local depletion of immune checkpoint ligand CTLA4 expressing cells in tumor beds enhances antitumor host immunity. Adv Ther. 2021;4(5). doi:10.1002/adtp.202000269
167. Hegde S, Leader AM, Merad M. MDSC: markers, development, states, and unaddressed complexity. Immunity. 2021;54(5):875–884. doi:10.1016/j.immuni.2021.04.004
168. Dorhoi A, Glaría E, Garcia-Tellez T, et al. MDSCs in infectious diseases: regulation, roles, and readjustment. Cancer Immunol Immunother. 2019;68(4):673–685. doi:10.1007/s00262-018-2277-y
169. Peranzoni E, Zilio S, Marigo I, et al. Myeloid-derived suppressor cell heterogeneity and subset definition. Curr Opin Immunol. 2010;22(2):238–244. doi:10.1016/J.COI.2010.01.021
170. Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9(3):162–174. doi:10.1038/nri2506
171. Liu T, Xie C, Ma H, et al. Gr-1+CD11b+ cells facilitate Lewis lung cancer recurrence by enhancing neovasculature after local irradiation. Sci Rep. 2014;4(1):1–9. doi:10.1038/srep04833
172. Ma J, Xu H, Wang S. Immunosuppressive role of myeloid-derived suppressor cells and therapeutic targeting in lung cancer. J Immunol Res. 2018;2018(1):6319649. doi:10.1155/2018/6319649
173. Ortiz ML, Lu L, Ramachandran I, Gabrilovich DI. Myeloid-derived suppressor cells in the development of lung cancer. Cancer Immunol Res. 2014;2(1):50–58. doi:10.1158/2326-6066.CIR-13-0129
174. Zimmermann H. 5’-Nucleotidase: molecular structure and functional aspects. Biochem. J. 1992;285(Pt 2):345. doi:10.1042/BJ2850345
175. Robson SC, Sévigny J, Zimmermann H. The E-NTPDase family of ectonucleotidases: structure function relationships and pathophysiological significance. Purinergic Sig. 2006;2(2):409–430. doi:10.1007/S11302-006-9003-5
176. Yegutkin G, Bodin P, Burnstock G. Effect of shear stress on the release of soluble ecto-enzymes ATPase and 5′-nucleotidase along with endogenous ATP from vascular endothelial cells. Br J Pharmacol. 2000;129(5):921–926. doi:10.1038/SJ.BJP.0703136
177. Fini C, Talamo F, Cherri S, et al. Biochemical and mass spectrometric characterization of soluble ecto-5’-nucleotidase from bull seminal plasma. Biochem. J. 2003;372(2):443–451. doi:10.1042/BJ20021687
178. Thomson LF, Ruedi JM, Glass A, et al. Production and characterization of monoclonal antibodies to the glycosyl phosphatidylinositol-anchored lymphocyte differentiation antigen ecto-5′-nucleotidase (CD73). Tissue Antigens. 1990;35(1):9–19. doi:10.1111/j.1399-0039.1990.tb01750.x
179. Linden J, Koch-Nolte F, Dahl GPR. Metabolism, and signaling in the inflammatory response. Annu Rev Immunol. 2019;37(37):2019):325–347. doi:10.1146/annurev-immunol-051116-052406
180. Su AL, Tian CQ, Ou YJ, et al. Proteasome inhibitors reduce CD73 expression partly via decreasing p-ERK in NSCLC cells. Life Sci. 2023;332:122129. doi:10.1016/J.LFS.2023.122129
181. Ishii H, Azuma K, Kawahara A, et al. Predictive value of CD73 expression for the efficacy of immune checkpoint inhibitors in NSCLC. Thorac Cancer. 2020;11(4):950–955. doi:10.1111/1759-7714.13346
182. Tu E, McGlinchey K, Wang J, et al. Anti–PD-L1 and anti-CD73 combination therapy promotes T cell response to EGFR-mutated NSCLC. JCI Insight. 2022;7(3):e142843. doi:10.1172/JCI.INSIGHT.142843
183. Rocha P, Salazar R, Zhang J, et al. CD73 expression defines immune, molecular, and clinicopathological subgroups of lung adenocarcinoma. Cancer Immunol Immunother. 2021;70(7):1965–1976. doi:10.1007/s00262-020-02820-4
184. Han Y, Lee T, He Y, et al. The regulation of CD73 in non-small cell lung cancer. Eur J Cancer. 2022;170:91–102. doi:10.1016/J.EJCA.2022.04.025
185. Giatromanolaki A, Kouroupi M, Pouliliou S, et al. Ectonucleotidase CD73 and CD39 expression in non-small cell lung cancer relates to hypoxia and immunosuppressive pathways. Life Sci. 2020;259:118389. doi:10.1016/J.LFS.2020.118389
186. Zhang H, Cao Y, Tang J, Wang R. Retracted: CD73 (NT5E) promotes the proliferation and metastasis of lung adenocarcinoma through the EGFR/AKT/mTOR pathway. Biomed Res Int. 2023;2023(1):9875750. doi:10.1155/2023/9875750
187. Petruk N, Tuominen S, Åkerfelt M, et al. CD73 facilitates EMT progression and promotes lung metastases in triple-negative breast cancer. Sci Rep. 2021;11(1):1–13. doi:10.1038/s41598-021-85379-z
188. Petruk N, Siddiqui A, Tadayon S, et al. CD73 regulates zoledronate-induced lymphocyte infiltration in triple-negative breast cancer tumors and lung metastases. Front Immunol. 2023;14:1179022. doi:10.3389/fimmu.2023.1179022
189. Yoshida R, Saigi M, Tani T, et al. MET-Induced CD73 restrains STING-Mediated immunogenicity of EGFR-Mutant lung cancer. Cancer Res. 2022;82(21):4079–4092. doi:10.1158/0008-5472.CAN-22-0770
190. Hasmim M, Xiao M, Van Moer K, et al. SNAI1-dependent upregulation of CD73 increases extracellular adenosine release to mediate immune suppression in TNBC. Front Immunol. 2022;13:982821. doi:10.3389/fimmu.2022.982821
191. De Leve S, Wirsdörfer F, Jendrossek V. Targeting the immunomodulatory CD73/adenosine system to improve the therapeutic gain of radiotherapy. Front Immunol. 2019;10:434045. doi:10.3389/fimmu.2019.00698
192. Shi E, Wu Z, Karaoglan BS, et al. 5ʹ-Ectonucleotidase CD73/NT5E supports EGFR-mediated invasion of HPV-negative head and neck carcinoma cells. J Biomed Sci. 2023;30(1):1–20. doi:10.1186/S12929-023-00968-6
193. Yan A, Joachims ML, Thompson LF, Miller AD, Canoll PD, Bynoe MS. CD73 promotes glioblastoma pathogenesis and enhances its chemoresistance via A2B adenosine receptor signaling. J Neurosci. 2019;39(22):4387–4402. doi:10.1523/JNEUROSCI.1118-18.2019
194. Goswami S, Walle T, Cornish AE, et al. Immune profiling of human tumors identifies CD73 as a combinatorial target in glioblastoma. Nat Med. 2020;26(1):39–46. doi:10.1038/s41591-019-0694-x
195. Lopes DV, de Fraga Dias A, Silva LFL, et al. Influence of NSAIDs and methotrexate on CD73 expression and glioma cell growth. Purinergic Sig. 2021;17(2):273–284. doi:10.1007/S11302-021-09775-W
196. Figueiro F, De Oliveira CP, Rockenbach L, et al. Pharmacological improvement and preclinical evaluation of methotrexate-loaded lipid-core nanocapsules in a glioblastoma model. J Biomed Nanotechnol. 2015;11(10):1808–1818. doi:10.1166/JBN.2015.2125
197. Xue G, Wang Z, Zheng N, et al. Elimination of acquired resistance to PD-1 blockade via the concurrent depletion of tumour cells and immunosuppressive cells. Nat Biomed Eng. 2021;5(11):1306–1319. doi:10.1038/s41551-021-00799-6
198. Xu ZJ, Gu Y, Wang CZ, et al. The M2 macrophage marker CD206: a novel prognostic indicator for acute myeloid leukemia. Oncoimmunology. 2020;9(1). doi:10.1080/2162402X.2019.1683347
199. van der Zande HJP, Nitsche D, Schlautmann L, Guigas B, Burgdorf S. The mannose receptor: from endocytic receptor and biomarker to regulator of (Meta)inflammation. Front Immunol. 2021;12:765034. doi:10.3389/fimmu.2021.765034
200. Schuette V, Embgenbroich M, Ulas T, et al. Mannose receptor induces T-cell tolerance via inhibition of CD45 and up-regulation of CTLA-4. Proc Natl Acad Sci U S A. 2016;113(38):10649–10654. doi:10.1073/pnas.1605885113
201. Jaynes JM, Sable R, Ronzetti M, et al. Mannose receptor (CD206) activation in tumor-associated macrophages enhances adaptive and innate antitumor immune responses. Sci Transl Med. 2020;12(530). doi:10.1126/scitranslmed.aax6337
202. Strack E, Rolfe PA, Fink AF, et al. Identification of tumor-associated macrophage subsets that are associated with breast cancer prognosis. Clin Transl Med. 2020;10(8):e239. doi:10.1002/CTM2.239
203. Zhao S, Mi Y, Guan B, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13(1):1–19. doi:10.1186/S13045-020-00991-2
204. Wang X, Luo G, Zhang K, et al. Hypoxic tumor-derived exosomal miR-301a mediates M2 macrophage polarization via PTEN/PI3Kg to promote pancreatic cancer metastasis. Cancer Res. 2018;78(16):4586–4598. doi:10.1158/0008-5472.CAN-17-3841
205. Li W, Zhang X, Wu F, et al. Gastric cancer-derived mesenchymal stromal cells trigger M2 macrophage polarization that promotes metastasis and EMT in gastric cancer. Cell Death Dis. 2019;10(12):1–16. doi:10.1038/s41419-019-2131-y
206. Lu CS, Shiau AL, Su BH, et al. Oct4 promotes M2 macrophage polarization through upregulation of macrophage colony-stimulating factor in lung cancer. J Hematol Oncol. 2020;13(1):1–16. doi:10.1186/S13045-020-00887-1
207. Zhang J, Li H, Wu Q, et al. Tumoral NOX4 recruits M2 tumor-associated macrophages via ROS/PI3K signaling-dependent various cytokine production to promote NSCLC growth. Redox Biol. 2019;22:101116. doi:10.1016/j.redox.2019.101116
208. Rao X, Zhou X, Wang G, et al. NLRP6 is required for cancer-derived exosome-modified macrophage M2 polarization and promotes metastasis in small cell lung cancer. Cell Death Dis. 2022;13(10):1–12. doi:10.1038/s41419-022-05336-0
209. Zhang C, Gao L, Cai Y, et al. Inhibition of tumor growth and metastasis by photoimmunotherapy targeting tumor-associated macrophage in a sorafenib-resistant tumor model. Biomaterials. 2016;84:1–12. doi:10.1016/j.biomaterials.2016.01.027
210. Guo X, Chang M, Wang Y, Xing B, Ma W. B7-H3 in brain malignancies: immunology and immunotherapy. Int J Biol Sci. 2023;19(12):3762. doi:10.7150/IJBS.85813
211. Wang L, Kang FB, Shan BE. B7-H3-mediated tumor immunology: friend or foe? Int, J, Cancer. 2014;134(12):2764–2771. doi:10.1002/IJC.28474
212. Picarda E, Ohaegbulam KC, Zang X. Molecular pathways: targeting B7-H3 (CD276) for human cancer immunotherapy. Clin Cancer Res. 2016;22(14):3425–3431. doi:10.1158/1078-0432.CCR-15-2428
213. Bao R, Wang Y, Lai J, et al. Enhancing Anti-PD-1/PD-L1 immune checkpoint inhibitory cancer therapy by CD276-targeted photodynamic ablation of tumor cells and tumor vasculature. Mol Pharm. 2019;16(1):339–348. doi:10.1021/acs.molpharmaceut.8b00997
214. Keppler D. Multidrug resistance proteins (MRPs, ABCCs): importance for pathophysiology and drug therapy. Handb Exp Pharmacol. 2011;201:299–323. doi:10.1007/978-3-642-14541-4_8
215. Sodani K, Patel A, Kathawala RJ, Chen ZS. Multidrug resistance associated proteins in multidrug resistance. Chin J Cancer. 2012;31(2):58. doi:10.5732/CJC.011.10329
216. Nooter K, Westerman AM, Flens MJ, et al. Expression of the multidrug resistance-associated protein (MRP) gene in human cancers. Clin Cancer Res. 1995;1(11):1301–1310.
217. Elfadadny A, El-Husseiny HM, Abugomaa A, et al. Role of multidrug resistance-associated proteins in cancer therapeutics: past, present, and future perspectives. Environ. Sci. Pollut. Res. 2021;28(36):49447–49466. doi:10.1007/S11356-021-15759-5
218. Brózik A, Hegedüs C, Erdei Z, et al. Tyrosine kinase inhibitors as modulators of ATP binding cassette multidrug transporters: substrates, chemosensitizers or inducers of acquired multidrug resistance? Expert Opin Drug Metab Toxicol. 2011;7(5):623–642. doi:10.1517/17425255.2011.562892
219. Michaelis M, Rothweiler F, Klassert D, et al. Reversal of P-glycoprotein–Mediated multidrug resistance by the murine double minute 2 antagonist Nutlin-3. Cancer Res. 2009;69(2):416–421. doi:10.1158/0008-5472.CAN-08-1856
220. Pham AN, Wang J, Fang J, et al. Pharmacogenomics approach reveals MRP1 (ABCC1)-mediated resistance to geldanamycins. Pharm Res. 2009;26(4):936–945. doi:10.1007/S11095-008-9796-8
221. Wang E, Johnson WW. The farnesyl protein transferase inhibitor lonafarnib (SCH66336) Is an inhibitor of multidrug resistance proteins 1 and 2. Chemotherapy. 2003;49(6):303–308. doi:10.1159/000074531
222. Cole SPC. Targeting multidrug resistance protein 1 (MRP1, ABCC1): past, present, and future. Annu Rev Pharmacol Toxicol. 2014;54(54):2014):95–117. doi:10.1146/annurev-pharmtox-011613-135959
223. Yang J, Song P, Zhou G. A study on the correlations of MRP-1 expression with the pathogenesis and prognosis of colorectal cancer. JBUON. 2019;24(1):84–90.
224. Yonetomi Y, Sekioka T, Kadode M, et al. Leukotriene C4 induces bronchoconstriction and airway vascular hyperpermeability via the cysteinyl leukotriene receptor 2 in S-hexyl glutathione-treated Guinea pigs. Eur J Pharmacol. 2015;754:98–104. doi:10.1016/J.EJPHAR.2015.02.014
225. Leier I, Jedlitschky G, Buchholz U, Cole SPC, Deeley RG, Keppler D. The MRP gene encodes an ATP-dependent export pump for leukotriene C4 and structurally related conjugates. J Biol Chem. 1994;269(45):27807–27810. doi:10.1016/S0021-9258(18)46856-1
226. Li F, Mao C, Yeh S, et al. MRP1-targeted near infrared photoimmunotherapy for drug resistant small cell lung cancer. Int J Pharm. 2021;604:120760. doi:10.1016/J.IJPHARM.2021.120760
227. Wang L, Rubinstein R, Lines JL, et al. Vista, a novel mouse Ig superfamily ligand that negatively regulates T cell responses. J Exp Med. 2011;208(3):577–592. doi:10.1084/JEM.20100619
228. Aloia L, Parisi S, Fusco L, Pastore L, Russo T. Differentiation of embryonic stem cells 1 (Dies1) is a component of bone morphogenetic protein 4 (BMP4) signaling pathway required for proper differentiation of mouse embryonic stem cells. J Biol Chem. 2010;285(10):7776–7783. doi:10.1074/jbc.M109.077156
229. Sakr MA, Takino T, Domoto T, et al. GI24 enhances tumor invasiveness by regulating cell surface membrane-type 1 matrix metalloproteinase. Cancer Sci. 2010;101(11):2368–2374. doi:10.1111/j.1349-7006.2010.01675.x
230. Flies DB, Wang S, Xu H, Chen L. Cutting edge: a monoclonal antibody specific for the programmed death-1 homolog prevents graft-versus-host disease in mouse models. J Immunol. 2011;187(4):1537–1541. doi:10.4049/JIMMUNOL.1100660
231. Yoon KW, Byun S, Kwon E, et al. Control of signaling-mediated clearance of apoptotic cells by the tumor suppressor p53. Science. 2015;349(6247). doi:10.1126/science.1261669
232. Yuan L, Tatineni J, Mahoney KM, Freeman GJ. Vista: a mediator of quiescence and a promising target in cancer immunotherapy. Trends Immunol. 2021;42(3):209–227. doi:10.1016/j.it.2020.12.008
233. Gao Y, He Y, Tang Y, Chen ZS, Qu M. Vista: a novel checkpoint for cancer immunotherapy. Drug Discov Today. 2024;29(7):104045. doi:10.1016/J.DRUDIS.2024.104045
234. Flies DB, Han X, Higuchi T, et al. Coinhibitory receptor PD-1H preferentially suppresses CD4+ T cell–mediated immunity. J Clin Invest. 2014;124(5):1966–1975. doi:10.1172/JCI74589
235. Lines JL, Pantazi E, Mak J, et al. Vista is an immune checkpoint molecule for human T cells. Cancer Res. 2014;74(7):1924–1932. doi:10.1158/0008-5472.CAN-13-1504
236. Rosenbaum SR, Knecht M, Mollaee M, et al. FOXD3 regulates vista expression in melanoma. Cell Rep. 2020;30(2):510–524.e6. doi:10.1016/j.celrep.2019.12.036
237. Nastase A, Dima SO, Lupo A, et al. Molecular markers for long-term survival in stage IIIA (N2) NSCLC patients. Cancer Genomics Proteomics. 2022;19(1):94–104. doi:10.21873/CGP.20306
238. Muller S, Victoria Lai W, Adusumilli PS, et al. V-domain Ig-containing suppressor of T-cell activation (Vista), a potentially targetable immune checkpoint molecule, is highly expressed in epithelioid malignant pleural mesothelioma. Mod Pathol. 2020;33(2):303–311. doi:10.1038/S41379-019-0364-Z
239. Mulati K, Hamanishi J, Matsumura N, et al. Vista expressed in tumour cells regulates T cell function. Br J Cancer. 2019;120(1):115–127. doi:10.1038/s41416-018-0313-5
240. Brcic L, Stanzer S, Krenbek D, et al. Immune cell landscape in therapy-naïve squamous cell and adenocarcinomas of the lung. Virchows Archiv. 2018;472(4):589–598. doi:10.1007/S00428-018-2326-0
241. Long L, Zhang L, Yang Y, Zhou Y, Chen H. Different clinical significance of novel B7 family checkpoints vista and HHLA2 in human lung adenocarcinoma. Immunotherapy. 2022;14(6):419–431. doi:10.2217/IMT-2021-0093
242. Ma S, Qin L, Wang X, et al. The expression of Vista on CD4+ T cells is associated with poor prognosis and immune status in non-small cell lung cancer patients. Bosn J Basic Med Sci. 2022;22(5):707. doi:10.17305/BJBMS.2021.6531
243. Liu J, Yuan Y, Chen W, et al. Immune-checkpoint proteins Vista and PD-1 nonredundantly regulate murine T-cell responses. Proc Natl Acad Sci U S A. 2015;112(21):6682–6687. doi:10.1073/pnas.1420370112
244. Wakiyama H, Furusawa A, Okada R, et al. Opening up new VISTAs: v-domain immunoglobulin suppressor of T cell activation (Vista) targeted near-infrared photoimmunotherapy (NIR-PIT) for enhancing host immunity against cancers. Cancer Immunol Immunother. 2022;71(12):2869–2879. doi:10.1007/s00262-022-03205-5
245. Okuyama S, Nagaya T, Ogata F, et al. Avoiding thermal injury during near-infrared photoimmunotherapy (NIR-PIT): the importance of NIR light power density. Oncotarget. 2017;8(68):113194–113201. doi:10.18632/ONCOTARGET.20179
246. Ogata F, Nagaya T, Nakamura Y, et al. Near-infrared photoimmunotherapy: a comparison of light dosing schedules. Oncotarget. 2017;8(21):35069–35075. doi:10.18632/ONCOTARGET.17047
247. Mitsunaga M, Nakajima T, Sano K, Choyke PL, Kobayashi H. Near-infrared theranostic photoimmunotherapy (PIT): repeated exposure of light enhances the effect of immunoconjugate. Bioconjug Chem. 2012;23(3):604–609. doi:10.1021/bc200648m
248. Maruoka Y, Wakiyama H, Choyke PL, Kobayashi H. Near infrared photoimmunotherapy for cancers: a translational perspective. EBioMedicine. 2021;70:103501. doi:10.1016/j.ebiom.2021.103501
249. Keall PJ, Mageras GS, Balter JM, et al. The management of respiratory motion in radiation oncology report of AAPM task group 76. Med Phys. 2006;33(10):3874–3900. doi:10.1118/1.2349696
250. Korreman SS. Image-guided radiotherapy and motion management in lung cancer. British J Radiol. 2015;88(1051). doi:10.1259/BJR.20150100/7445560
251. Jamal-Hanjani M, Wilson GA, McGranahan N, et al. Tracking the evolution of non–small-cell lung cancer. New Engl J Med. 2017;376(22):2109–2121. doi:10.1056/NEJMoa1616288
252. Al Bakir M, Huebner A, Martínez-Ruiz C, et al. The evolution of non-small cell lung cancer metastases in TRACERx. Nature. 2023;616(7957):534–542. doi:10.1038/s41586-023-05729-x
253. Nagaya T, Friedman J, Maruoka Y, et al. Host immunity following near-infrared photoimmunotherapy is enhanced with PD-1 checkpoint blockade to eradicate established antigenic tumors. Cancer Immunol Res. 2019;7(3):401–413. doi:10.1158/2326-6066.CIR-18-0546
254. Gu Q, Li S, Hou S, et al. Antibody-dependent cellular phagocytosis in cancer immunotherapy: research and perspectives. J Translat Med. 2026;24(1):601. doi:10.1186/S12967-026-08020-5
255. Mohiuddin TM, Zhang C, Sheng W, et al. Synthetic zipper mediated pre-targeting system for near-infrared photoimmunotherapy. iScience. 2026;29(2):114558. doi:10.1016/J.ISCI.2025.114558
256. Sato K, Nagaya T, Choyke PL, Kobayashi H. Near infrared photoimmunotherapy in the treatment of pleural disseminated NSCLC: preclinical experience. Theranostics. 2015;5(7):698–709. doi:10.7150/THNO.11559
257. Mohiuddin TM, Zhang C, Sheng W, et al. Near infrared photoimmunotherapy: a review of recent progress and their target molecules for cancer therapy. Int J Mol Sci. 2023;24(3):2655. doi:10.3390/IJMS24032655
258. Biel MA, Gillenwater AM, Cognetti DM, Johnson JM, Argiris A, Tahara M. A global phase III multicenter, randomized, double-arm, open label trial of ASP-1929 photoimmunotherapy versus physician’s choice standard of care for the treatment of patients with locoregional, recurrent head and neck squamous cell carcinoma (rHNSCC). J Clin Oncol. 2019;37(15):TPS6094–TPS6094. doi:10.1200/JCO.2019.37.15_suppl.TPS6094
259. Cognetti DM, Curry JM, Johnson J, et al. Safety and efficacy findings from a phase Ib/II study of ASP-1929 photoimmunotherapy with pembrolizumab in recurrent and/or metastatic head and neck squamous cell carcinoma. Head Neck. 2026;48(1):160–174. doi:10.1002/hed.70014
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
GABAA Receptor Modulators as Emerging Therapeutic Agents for Depressive Disorders
Guan BY, Li Y
Drug Design, Development and Therapy 2026, 20:608834
Published Date: 8 June 2026