Back to Journals » Drug Design, Development and Therapy » Volume 20
Natural G-Quadruplex Stabilizers: A Targeted Strategy for Breast Cancer Therapy
Received 21 March 2026
Accepted for publication 14 May 2026
Published 26 May 2026 Volume 2026:20 611085
DOI https://doi.org/10.2147/DDDT.S611085
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Chengjian Cao,1 Chaoxiang Lv,2 Ali ElFar2
1Zigong Academy of Medical Sciences, Zigong First People’s Hospital, Zigong, Sichuan, People’s Republic of China; 2Key Laboratory of Epigenetics and Oncology, The Research Center for Preclinical Medicine, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China
Correspondence: Ali ElFar, Key Laboratory of Epigenetics and Oncology, The Research Center for Preclinical Medicine, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China, Email [email protected] Chaoxiang Lv, Key Laboratory of Epigenetics and Oncology, The Research Center for Preclinical Medicine, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China, Email [email protected]
Abstract: G-quadruplexes (G4s) are non-canonical DNA structures that are important in gene regulation, telomere repair, and cancer. The G4s in breast cancer (BC) are enriched in the promoters of important oncogenes, such as c-MYC, KRAS, BCL2, HER2, and ESR1, which are good therapeutic targets. Although synthetic G4 stabilizers are well-researched, natural products are a poorly investigated source of selective ligands with good biocompatibility and multi-target properties. This review systematically collects and analyzes natural compounds, such as flavonoids, alkaloids, polyphenols, terpenoids, and organosulfur compounds, which exhibit G4-stabilizing effects, particularly their suitability in BC. We provide a comprehensive landscape approach to define interactions between G4-ligands, discuss how these interactions are stabilized (via π-π stacking, electrostatic interactions, and groove binding), and map these compounds to genomic regions associated with BC G4-formation. We also identify key research gaps that are essential, including limited in vivo validation, subtype-specific efficacy, and pharmacokinetic issues. The available evidence suggests that natural G4 stabilizers have potential as multi-target and biocompatible agents for BC therapy. However, their effectiveness is currently constrained by a shortage of thorough in vivo studies and subtype-specific data. Lastly, to incorporate in silico docking, mechanistic studies, and translational strategies, we present a prospective roadmap for drug discovery to bring natural G4 stabilizers targeting BC. The diagram illustrates the relationship between natural products, mechanisms, breast cancer oncogenes and the S cubed framework. Natural products include flavonoids, alkaloids, polyphenols, terpenoids and organosulfur. These are linked to mechanisms such as pi-pi stacking, electrostatic, hydrogen bonding and groove binding. These mechanisms connect to breast cancer oncogenes, including c-MYC, KRAS, BCL2, HER2, ESR1, VEGF and hTERT. The goal is to identify the best natural products for breast cancer G4s stabilization. The S cubed framework, described as a fail-early strategy, focuses on selectivity, stability and solubility.Diagram linking natural products, mechanisms, breast cancer oncogenes and S cubed framework for G4 stabilization.
Keywords: G-quadruplexes, breast cancer, G4 stabilizers, natural products, drug discovery, future roadmap
Introduction
G-quadruplexes (G4s) consist of a G-tetrad, the square-planar structure of four guanine bases, forming quartets,1 stabilized by Hoogsteen hydrogen bonding, which allows the tetrad to stack with other two or more tetrads to create G4s, and stabilized by the presence of cations, such as K+, Na+, and Li+.2 Those guanine bases that are connected by hydrogen bonds between the N1, N7, O6, and N2 atoms of the guanine molecules form a G-tetrad between adjacent bases,3 as shown in Figure 1. G4s occur in great numbers in both prokaryotic and eukaryotic genomes and are vital for telomere maintenance, nucleic acid metabolism, and genomic integrity.4 G4s within genes are regulatory factors involved in cellular processes and pathogenesis, including cancer. They are therefore considered therapeutic targets in cancer treatment. G4s are essential for gene expression and DNA replication. The first G4s were reported in a study in 1962, and G4 tetrameric structures had been reported to form with their unique composition.5 They have, since that time, served as a model of research on their structural features and functional role in health processes and diseases.3
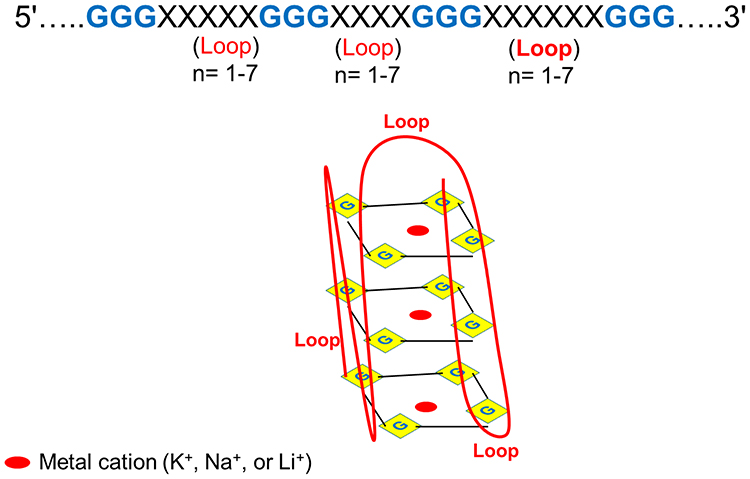 |
Figure 1 Structure of G-quadruplex. n = 1–7 means that the number of nucleotides in the loop is from 1 to 7. |
The importance of targeting G4s in breast cancer (BC) is profound and exhibits unique features that distinguish it from other cancer types. BC is still the top cause of cancer death among women globally, and resistance to treatment—especially in triple-negative breast cancer (TNBC)—remains a major obstacle to improving patient outcomes.6–9 Traditional targeted therapies, such as endocrine treatments and HER2-focused drugs, face limitations due to acquired resistance and subtype specificity.10 Genome-wide mapping of 22 BC patient-derived tumor xenograft models shows BC-G4s mainly in gene promoters and first introns, near transcription start sites. These G4s serve as hubs for transcription factors and relate to cell cycle genes.11 G4 dynamics have subtype-specific roles: stabilizing an rG4 in BCL2 pre-mRNA affects splicing differently in TNBC vs. non-TNBC. In TNBC, it boosts m6A, lowers BCL2 mRNA, and promotes BCL2β; in non-TNBC, it raises BCL2α.12 G4s at the MAPK12 promoter influence a switch affecting stemness in TNBC, driving cancer stem cell growth and poorer survival due to metastasis.13 These findings highlight that G4s are more than passive structural features; they are active, subtype-specific regulators involved in BC pathogenesis.
Possible G4 stabilizers are natural products derived from plants, fungi, and marine organisms that interact with G4 structures. Their stabilization mechanism can be understood to aid in improving and clarifying the biological functions of G4. Many researchers are placing more emphasis on natural products as G4 stabilizers, which influence gene expression, DNA replication, and telomere maintenance. Investigation of the action and biological impact of these substances will help identify new anticancer candidate drugs.14 Isolated natural compounds (and in particular alkaloids and flavonoids) have been shown to have G4-stabilizing properties in detail by numerous reviews, but, from a chemical-biology perspective, these compounds have been studied either in partially purified form or as extracts.15 The main benefits of natural products in G4 stabilization offer strong reasons to explore them over synthetic options. While synthetic G4 stabilizers like pyridostatin and CX-5461 show promise in preclinical studies, they face significant hurdles for clinical use, such as off-target toxicity, poor bioavailability, and high development costs.16 In contrast, natural products have distinct advantages: they are inherently biocompatible, structurally diverse, capable of targeting multiple pathways, and have a long history of safe use in humans.17–19 Natural products offer a wide variety of bioactive scaffolds for drug development due to their structural diversity. They can target multiple pathways simultaneously by binding to G4 structures at various promoters, which is especially beneficial for overcoming pathway redundancy, a common driver of therapy resistance. Additionally, they are readily accessible and have a long record of safe human use.17,20 Dietary polyphenols, such as epigallocatechin gallate (EGCG), quercetin, and curcumin, have demonstrated the ability to bind G4 structures and serve as epigenetic modifiers with anticancer properties, underscoring the translational potential of natural compounds as G4-targeted ligands.20 Nonetheless, the field lacks a comprehensive, critical overview that links specific natural compounds to G4 targets relevant in BC.
Despite natural products being recognized as a broad-spectrum G4 stabilizer in oncology, there remains a significant gap in translating this knowledge into practice in the BC therapeutic field. This gap becomes especially urgent due to the significant unmet clinical needs in BC treatment. Traditional targeted therapies, such as endocrine treatments and HER2-specific drugs, are limited by acquired resistance and by their focus on specific subtypes.10 TNBC, which comprises 10–20% of BCs and lacks estrogen receptor, progesterone receptor, and HER2, is a severe challenge. Its absence of typical targets renders it unresponsive to conventional therapies, leaving cytotoxic chemotherapy as the main systemic option despite high resistance rates.21 Resistance develops rapidly via drug efflux, enhanced DNA repair, apoptosis evasion, epithelial-to-mesenchymal transition, and microenvironment defenses, limiting treatment success.22 G4 stabilization offers a promising alternative by targeting DNA to inhibit multiple cancer pathways, potentially overcoming resistance from pathway redundancy.14,23 Exploring natural G4 stabilizers is an urgent priority to address therapeutic gaps for aggressive BC subtypes.
There is no standard body of literature that compiles the known natural G4 ligands against the well-known oncogenic drivers of BC progression, including c-MYC, KRAS, BCL2, HER2, and ESR1. Moreover, the empirical evidence for these compounds is not supported by a logical structure-based forecast of their competence in treating BC-selective G4 targets. This review distinctly fills these gaps by: (i) systematically classifying natural G4 stabilizers based on their molecular mechanisms, such as π-π stacking, electrostatic interactions, and groove binding; (ii) critically assessing the evidence linking each compound to G4-forming genes relevant to BC; and (iii) outlining a practical roadmap for future validation and development, incorporating in silico, biophysical, and in vivo methods.24,25 By doing so, we go beyond mere cataloging to establish a critical framework that highlights compounds with the highest therapeutic potential for specific BC subtypes. This gap in the literature will be addressed by not only listing the natural G4 stabilizers that have potential applications in BC but also by developing a roadmap for future experimental validation and drug development in phytotherapy.
Techniques of Characterization of G-Quadruplex and G-Quadruplex-Ligand Interactions
G4s and their interactions with small-molecule ligands have become of great interest within the scope of the study due to their potential as therapeutic agents in cancer and hereditary diseases. A range of experimental and computational tools has been developed to best describe these structures and their binding events and include spectroscopic, biophysical, computational, cell-based, and specialized methods (Table 1). This report is a systematically organized compilation of these techniques, including the description of one of the methods, offering researchers the opportunity to have a detailed overview of the principles of such techniques, their applications, and the existing sources, which are vital in the further development of the G4-related studies and discovery of ligands.
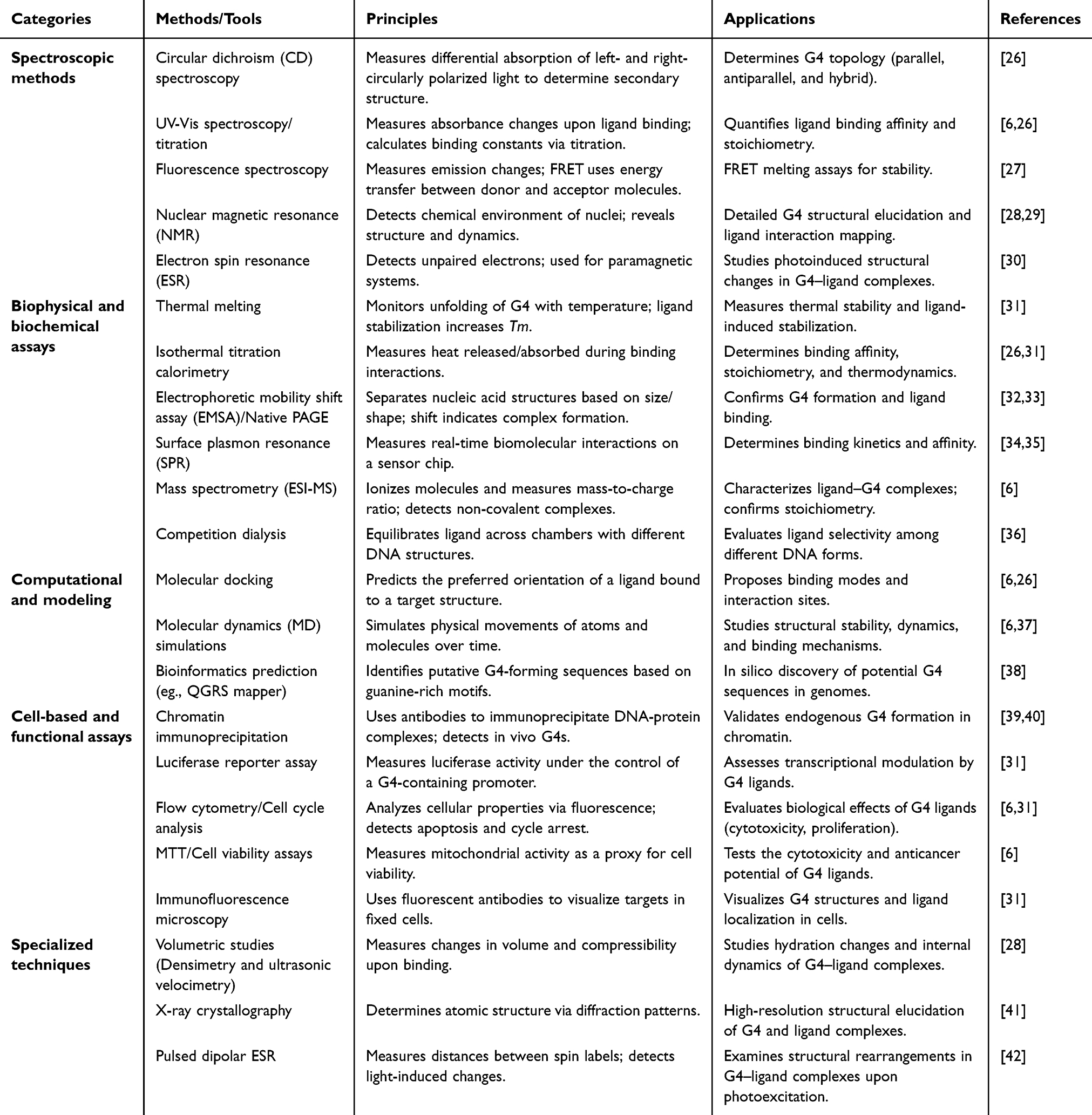 |
Table 1 G-Quadruplexes Characterization Tools |
The light-matter interaction is essential for characterizing G4 structures and their interactions with ligands using spectroscopic methods. Circular dichroism (CD) spectroscopy is a method for determining G4 topology (parallel, antiparallel, and hybrid) by detecting differences in the absorption of circularly polarized light.26 The binding affinity and stoichiometry between the ligand and the targeted protein are quantified by UV-Vis spectroscopy, a technique that can be performed by titration and measured by monitored changes in absorbance.6,26 Fluorescence spectroscopy and fluorescence resonance energy transfer (FRET)-based techniques are used to measure the G4 stability, ligand selectivity, and intrinsic fluorescence.27 Nuclear magnetic resonance (NMR) provides structural and dynamic information by identifying the chemical surroundings of nuclei.28,29 Paramagnetic systems are studied by electron spin resonance (ESR) to detect photoinduced structural changes in G4-ligand complexes.30
Quantitative and functional details of interactions between G4-ligands can be derived by biophysical and biochemical assays. G4 folding with temperature is used in thermodynamic studies of thermal stability and ligand-induced stabilization with thermal melting assays to monitor folding.31 It is the method that directly measures the thermodynamics of binding, such as the affinity, stoichiometry, and enthalpy changes.26,31 Complex formation is measured by electrophoretic mobility shift assays (EMSA) or native PAGE because of mobility shifts.32,33 Surface plasmon resonance (SPR) can be used to dynamically measure binding kinetics and affinity on sensor chips.34,35 The mass spectrometry (ESI-MS/HR-MS) identifies non-covalent complexes and stoichiometry.6 Competition dialysis determines the selectivity of ligands to the various DNA structures.36
Mechanistic and predictive in silico studies of G4 systems are possible by computational methods. Molecular docking is an approach that predicts the preferred orientations of the ligand and identifies interaction sites of G4 structures.6,26 Atomic motions can be tracked over time to investigate stability, dynamics, and binding by modifying atomic positions using molecular dynamics (MD) simulations.6,37 Bioinformatics applications, such as QGRS mapper, identify G4-forming sequences in a genome, which helps identify possible G4 motifs.38
Cell-based assays are used to determine G4 biology and the effects of ligands in cellular systems, thereby bridging the gap between cellular and in vitro studies. Endogenous G4 formation and protein interaction in chromatin is confirmed by chromatin immunoprecipitation (ChIP).39,40 The transcriptional modulation by G4 ligands under G4-containing promoters is measured using luciferase reporter assays.31 The principles of cytotoxicity, apoptosis, and proliferation effects are assessed by flow cytometry and cell cycle analysis.6,31 Mitochondrial activity is the measure of cytotoxicity and anticancer potential that is tested by MTT and cell viability assays.6 G4 structures and ligand localization in fixed cells can be visualized using immunofluorescence microscopy.31
The specialized methods provide high-resolution/special information about G4-ligand complexes using sophisticated physical and structural procedures. Volumetric measurements (densimetry and ultrasonic velocimetry) assess changes in volume and compressibility in the study of hydration and internal dynamics.28 X-ray crystallography provides atomic-resolution structures of G4 and ligand complexes from diffraction patterns.41 Pulsed dipolar ESR is a distance measurement technique used to study light-induced structural rearrangements between spin labels.42 Single-molecule experiments monitor individual molecules continuously to investigate folding dynamics, mechanical stability, and the impact of ligands.36
There is a significant methodological gap in standardizing G4-ligand characterization assays. Although techniques, such as CD spectroscopy and FRET melting, are prevalent, cross-study comparisons are difficult due to differences in buffer conditions (K⁺ vs. Na⁺), oligonucleotide sequences, and ligand concentrations. For instance, binding affinity estimates for berberine to c-MYC G4 vary depending on the method.43–45 Most in vitro assays use isolated oligonucleotides in non-physiological buffers that do not mimic the complex chromatin environment, competing nucleic acids, and molecular crowding within the nucleus. Consequently, many compounds showing promise in vitro fail in cellular or in vivo models—contributing to the high failure rate in G4‑targeted drug development. We suggest that future research adopt a minimum reporting standard that includes: (i) cation type and concentration, (ii) G4 topology confirmed by CD, and (iii) both thermal (ΔTm) and thermodynamic (ΔG and ΔH) parameters when possible. Additionally, we recommend incorporating cellular thermal shift assays (CETSA) or ChIP as standard procedures to verify target engagement within a physiologically relevant setting.
In summary, the overall toolkit for characterizing G4 and its interactions with ligands encompasses several fields of science, including spectroscopic and biophysical experiments, computational modeling, cell-based experiments, and other specialized structural methodologies. The different categories provide complementary information; spectroscopy can be used to determine structural and binding properties, biophysical assays can be used to measure thermodynamic and kinetic parameters, computational tools can be used to predict and simulate interactions, cell-based experiments can be used to establish biological relevance, and special techniques can be used to provide high-resolution or dynamic information. All these strategies can facilitate the development of a comprehensive approach to the study of G4 biology and ligand targeting, thereby advancing drug discovery, genomic studies, and the development of therapeutics. Combinations of these approaches are key to clarifying the multifaceted roles of G4 in cellular biology and disease.
Synthetic G-Quadruplex Stabilizers
Synthetic compounds with a wide variety of chemical structures that have been used as G4 stabilizers are listed in Table 2. Among them, platinum complexes (including Pt(II) squares and Pt(II) complexes),46–51 and heterocyclic derivatives (such as quinoxaline-based and carbazole-benzoindolium ligands).52,53 These anticancer ligands repress the transcription of oncogenes, including c-MYC,54 block telomerase activity,55 regulate immune response by cGAS-STING activation,56 and induce synthetic lethality in cells with defects in DNA repair.57 Furthermore, fluorescent probes such as thioflavin T can be used to image G4 structures in live cells.58 Although several platinum-based complexes exhibit strong binding to G4 structures, none have moved beyond preclinical research, primarily due to worries about metal toxicity and DNA adduct formation. In contrast, natural products such as berberine and quercetin have well-established safety profiles from centuries of traditional use but do not achieve the nanomolar potency seen in synthetic agents. As a result, the field is caught in a “potency-selectivity-safety triangle,” in which no current compound effectively balances all three aspects.
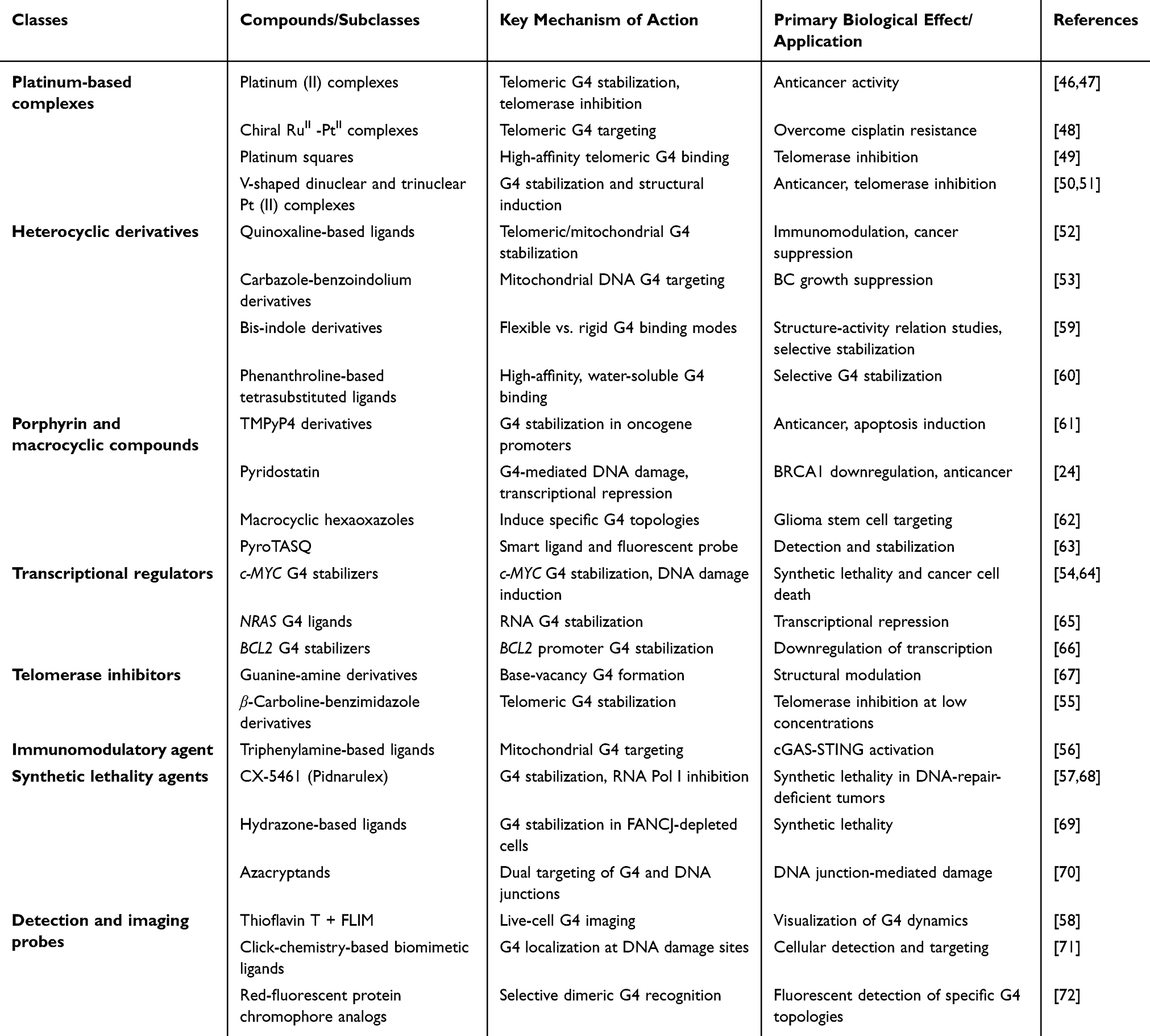 |
Table 2 Synthetic G-Quadruplex Stabilizers |
Although the signs in preclinical studies were encouraging, there is still work to be done to overcome barriers such as topological selectivity, improved cell delivery, and further development of candidates, eg., CX‑5461 (also known as pidnarulex), for clinical application. G4 stabilizers are synthetic agents that target the G4 structure, which can be used to treat cancer. They target G4 structures within oncogene promoters, silencing gene expression, reducing tumor progression, and increasing the effect of chemotherapy. CX‑5461 is an advanced G4 stabilizer in development with FDA Fast Track, initially targeting RNA polymerase I‑driven rDNA transcription. It stabilizes G4 structures and shows toxicity to topoisomerase II.73 Besides, oncogenes with G4-forming sequences are particularly sensitive to pyridostatin, which is a specific drug that binds to G4 structures in promoters of these oncogenes and reduces the growth of cancer cells by inducing apoptosis.24 Thus, it is a potential candidate drug in the treatment of tumors with G4-forming sequences. Although CX-5461 exhibits synthetic lethality in DNA-repair-deficient tumors.68 Its clinical development is hindered by dose-limiting toxicities, such as thrombocytopenia74 and an induced DNA damage response in primary vascular smooth muscle cells,75 as well as by side effects rarely reported with natural products. Likewise, pyridostatin effectively stabilizes G4 structures in vitro, but struggles with poor cellular uptake and causes off-target DNA damage in neurons.76 This underscores a fundamental challenge: balancing potency and selectivity remains unresolved in synthetic chemistry.
The other G4 stabilizer, BRACO-19, targets telomeric G4 structures, stabilizing them and leading to telomere shortening. This causes senescence or death of cancer cells. G4 stabilization and telomerase activity inhibition (excessively expressed in cancers to sustain telomeres) make BRACO-19 induce telomere dysfunction and inhibit cancer cell growth.77 Cationic porphyrins, including TMPyP3 and TMPyP4, can be designed to exhibit increased binding affinity for G4 structures, thereby providing high stabilization capacity for targeted cancer treatment. They are designed to intercalate into G4s, thereby stabilizing them through electrostatic and π-π stacking, and the positive charges on the molecules enable them to bind the negatively charged G4s.61,78 However, TMPyP4 binds non-selectively to duplex DNA, RNA, and G4 structures. A crystallographic study shows that it prefers to stack with loop nucleotides rather than G-tetrads.79 Interestingly, although it is often described as a G4 stabilizer, TMPyP4 can also destabilize certain G4 topologies.80
Furthermore, derivatives of temozolomide increase the stability of G4, thereby intensifying the anticancer activity of G4-targeted compounds on high-grade glioma by targeting both G4 structures and normal DNA. Such modifications enhance binding affinity through chemical changes, thereby interfering with key processes, such as telomere maintenance and oncogenic signaling, limiting the growth of cancer cells.81 Such synthetic stabilizers, such as quindoline analogs with a fused bicyclic structure, bind to G4, thereby mediating cancer cell growth by stabilizing G4 structures and destabilizing oncogenic pathways. Such stabilization has significant potential to influence gene expression and tumor progression, which is why it is a promising cancer treatment method.82 Moreover, the G4s are stabilized by phenazine derivatives and preferentially bind, thereby increasing their stability and affecting gene expression and oncogenic signaling, which can inhibit cancer growth. The compounds have the potential to prevent tumor growth and improve the effectiveness of the treatment.83
Synthetic G4 stabilizers have potential but have many challenges. One of them is selectivity: many compounds have been shown to act on multiple DNA structures, leading to off-target effects and making their therapeutic application more difficult. For example, the clinical development of CX‑3543 (quarfloxin), an initial G4‑targeting compound, was halted after Phase II trials because of bioavailability challenges and a limited therapeutic window.84,85 This illustrates that even compounds with promising preclinical results may fail during later development stages.
Although attempts have been made to minimize toxicity, there are still some stabilizers that lead to cytotoxicity at therapeutic doses, and this raises concerns about the safety of long-term treatment, as in the case of cancer. Numerous synthetic agents are also characterized by inadequate solubility and bioavailability and diminish clinical effectiveness. Poor solubility reduces the ability to achieve suitable concentrations in target tissues, which influences treatment success.16 The formation of G4 requires cellular conditions, such as ion levels and other nucleic acids, which synthetic stabilizers are not always able to handle, thereby affecting their biological activity.86 DNA repair modifications, whether through alterations in DNA repair proteins or through alternative survival pathways, lead to the development of cancer cells. The creation of new compounds is expensive and technologically sophisticated, which limits access to high-technology treatments.87 These complex interactions between synthetic stabilizers and G4 are causing problems with the attempts to maximize their design and performance. In addition, regulatory approval is a lengthy process that requires substantial safety and efficacy data to approve new therapies. To recap, synthetic G4 stabilizers are not entirely negative, but it is important to overcome these obstacles to their successful development and clinical application.
Natural Products with Proven G-Quadruplex Stabilizing Activity
Natural products stabilize G4 structures through a series of specific, non-covalent interactions, thereby allowing them to target and regulate important gene expression as represented in Table 3. The structures of natural product G4 stabilizers are illustrated in Figure 2. The main action is π-π stacking, in which planar aromatic parts are stacked over the guanine tetrads on the terminal to inhibit the process of unfolding and influence the gene promoters. For example, flavonoid products, including quercetin and kaempferol, exert this effect by stabilizing G4s within the promoters of oncogenes, such as c-MYC and VEGF, thereby repressing their transcription.88–91 On the same note, luteolin is preferred for intercalation via π-π stacking to stabilize G4s and suppress c-MYC expression.92 A similar mechanism also applies to polyphenols such as curcumin and EGCG that stabilize G4s at c-MYC.7,93 Terpenoids, like thymoquinone, also use π-π stacking to bind c-MYC,88 as do anthraquinones, like the binders of G4s in c-KIT and c-MYC promoters, emodin and aloe-emodin.94,95
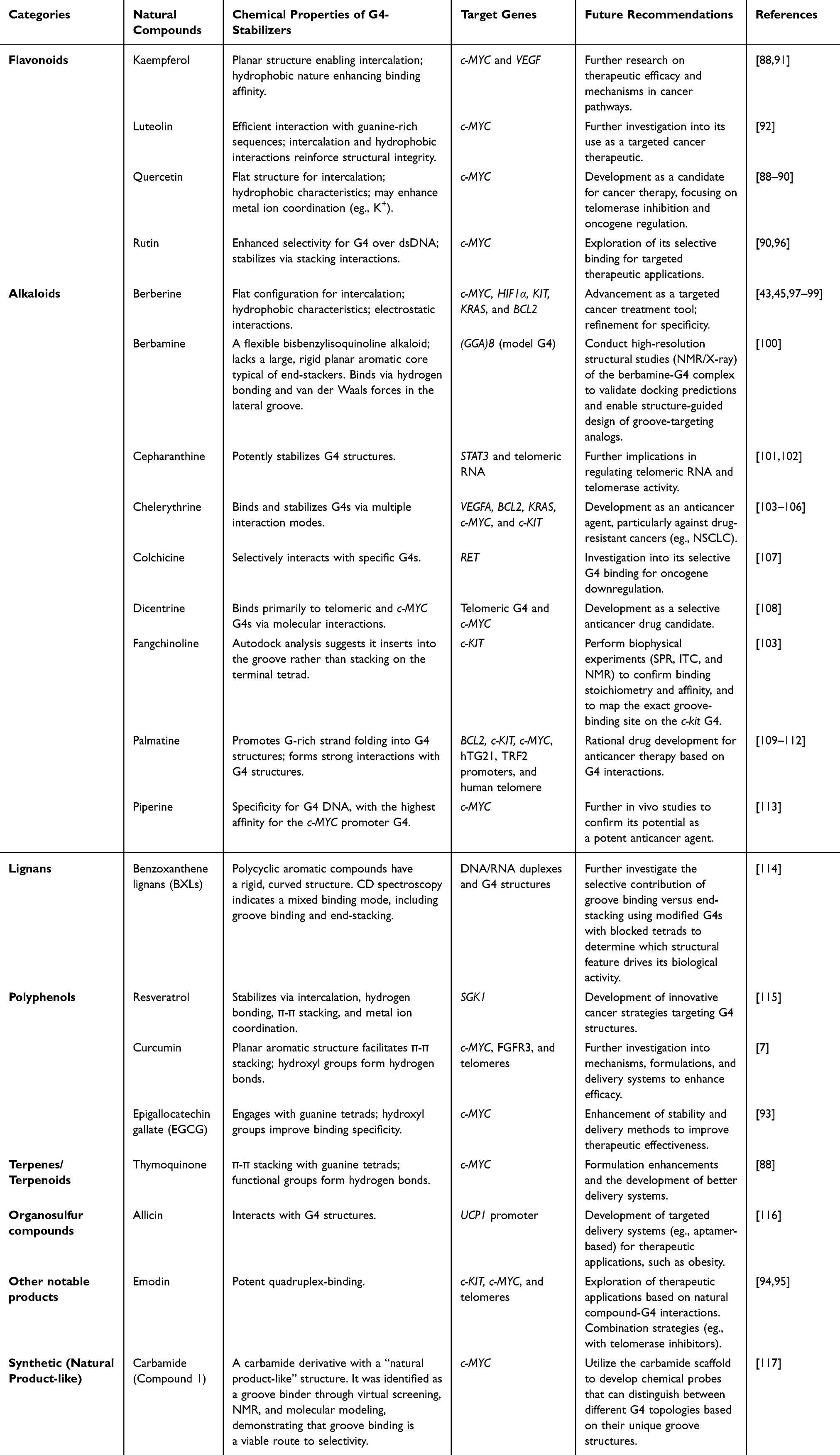 |
Table 3 Natural Compounds as G4-Stabilizers |
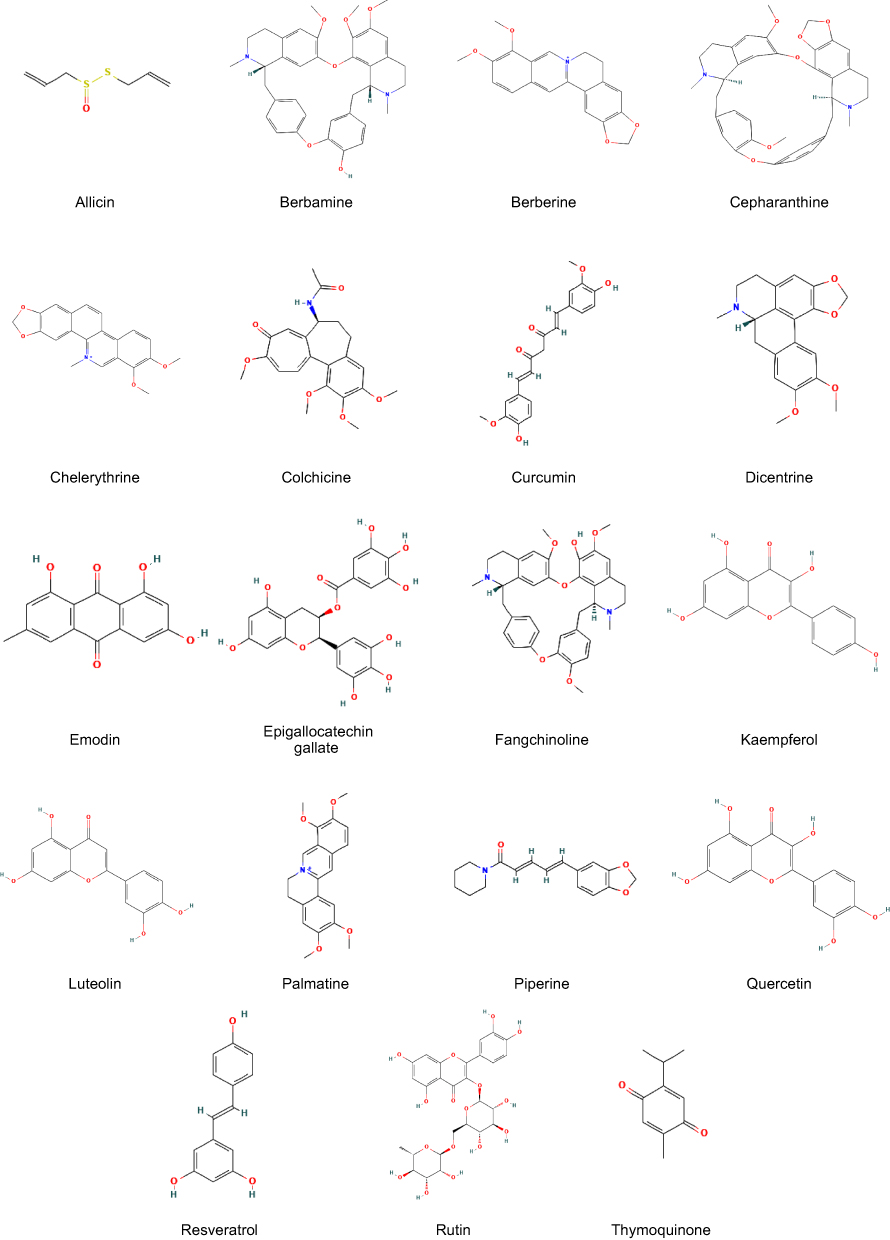 |
Figure 2 Chemical structures of natural product G4 stabilizers. |
A second important mechanism is electrostatic interaction, often accompanied by π-π stacking, particularly when using cationic alkaloids, which act on G4 promoters. Sanguinarine has a positive charge, which promotes its affinity to the phosphate backbone to stabilize G4s and suppress the activity of oncogenes such as human telomeric DNA, c-MYC, BCL2, KRAS, and VEGF.97,109,118,119 This dual mechanism is used by berberine to stabilize G4s, decrease the expression of c-MYC, HIF1α, KIT, and KRAS.43,45,97–99 Other alkaloids, such as chelerythrine and epiberberine, utilize π-π and electrostatic forces to act on G4s in the promoter of VEGFA, BCL2, KRAS, c-MYC, and c-KIT.103–106 Multiple studies consistently show that cationic alkaloids such as sanguinarine, berberine, and chelerythrine generally target various G4-forming promoters, including c-MYC, KRAS, BCL2, and VEGF, but with limited specificity. Additionally, piperine exhibits high selectivity for binding to c-MYC promoter G4.113 The structural reasons behind piperine’s selectivity—particularly its distinctive aliphatic chain and methylenedioxy ring—remain unexplored, offering a promising avenue for the rational design of isoform-specific G4 ligands.
A third very selective mode of action is hydrogen bonding and groove binding. Groove binding, hydrogen bonding, and interactions with loop nucleotides are all used by quercetin and rutin to specifically stabilize the c-MYC promoter G4.90,96 A comparative analysis of flavonoid G4 interactions uncovers a notable difference. While quercetin stabilizes c-MYC G4s through π-π stacking,90 while rutin—a glycoside of quercetin—shows significantly greater selectivity for G4 DNA over duplex DNA.96 This indicates that glycosylation could be an important but underrecognized factor influencing selectivity. However, no comprehensive structure-activity relationship study has explored how the position, number, or type of sugar moieties affect discrimination between G4 structures and duplex DNA.
Resveratrol stabilizes G4 structures in the SGK1 promoter by simultaneously forming hydrogen bonds and intercalating.115 Furthermore, several natural and synthetic compounds demonstrate this novel mechanism of action. For example, berbamine,100 a flexible bisbenzylisoquinoline alkaloid lacking a rigid aromatic backbone, binds to the (GGA)8 model G4 via hydrogen bonds and Van der Waals forces in the lateral groove, suggesting a need to revisit structural studies for more precise design. Similarly, Autodock predicts that fangchinoline binds within the c-KIT G4 groove instead of stacking on the terminal tetrad.103 However, these predictions lack experimental validation through NMR, SPR, or X-ray crystallography. This is a crucial gap because groove binding is thought to offer greater topological selectivity than end-stacking, but direct evidence has yet to be provided.
Benzoxanthene lignans are characterized by their rigid, curved structure, exhibit a mixed binding mode involving both groove binding and end-stacking with DNA/RNA duplexes and G4 structures, warranting further study with modified G4s to clarify which structural features drive their biological activity.114 Lastly, a synthetic carbamide analog (Compound 1) was designed to resemble a natural product and to be a groove binder through virtual screening and NMR; this suggests targeting unique groove topologies as an alternative strategy for selectivity and provides a foundation for developing chemical probes that differentiate various G4 structures.117
Other natural products exploit particular genes by stabilizing G4 structures. Cepharanthine is useful for stabilizing G4 structure in the STAT3 gene and telomeric RNA.101,102 Colchicine also specifically binds to the G4 of the RET oncogene,107 and dicentrine regulates G4s in telomeric and c-MYC regions.108 Also, allicin can bind to a G4 motif in the mitochondrial uncoupling protein-1 (UCP1) promoter.116 Generally, natural products may specifically stabilize G4s, particularly in the regulatory regions of essential genes, through mechanisms such as π-π stacking, electrostatic attraction, and hydrogen bonding, and thus have considerable potential as targeted therapies for cancer treatment.
Limitations
Natural products have a promising future; nevertheless, they face significant challenges for use as G4 stabilizers in therapeutic uses. Their inability to be selective and specific is the main problem. One such example is flavonoids such as quercetin and kaempferol, and alkaloids such as berberine and sanguinarine, which are generally broad-acting, capable of binding to numerous nucleic acid structures, including duplex DNA and RNA, as well as unrelated cellular proteins, causing off-target effects.17,120 This low selectivity makes it difficult to have a direct correlation between G4 stabilization and certain phenotypic outcomes. There is also a tendency toward poor pharmacokinetic profiles, including low solubility, high membrane permeability, rapid metabolic stability, and systemic instability. Examples of such compounds include curcumin, EGCG, and thymoquinone, whose bioavailability is low, and they break down easily in the gastrointestinal tract, making their use in the body challenging.121–123 Different studies rely on simplified in vitro systems (eg., synthetic oligonucleotides in non-physiological buffers) that are not representative of the chromatin context, competing biomolecules, or intracellular ion concentration, making it difficult to predict actual in vivo efficacy.
Research Gaps
Various knowledge gaps in the development of natural G4 stabilizers are barriers between the basic research and clinical practice. To begin with, mechanistic knowledge at the cellular and organismal levels is lacking significantly. Despite biophysical evidence of G4 binding and stabilization observed in vitro (eg., with chelerythrine), in this case, it is not always clear that a particular therapeutic effect in complex cellular systems has been primarily mediated by G4 interaction, rather than by other targets.104,105 Secondly, 21 natural compounds are listed in Table 3; only 4 (berberine, quercetin, curcumin, and sanguinarine) have been evaluated in more than three independent studies. The remaining compounds have single-study reports, raising concerns about reproducibility and the potential for publication bias. We strongly recommend that future studies include positive controls (eg., pyridostatin or TMPyP4) to enable cross-laboratory comparisons. Thirdly, the structure-activity relation is not sufficiently analyzed. In addition, there is limited systematic medicinal chemistry research aimed at improving natural product leads to achieve even better potency, specificity, and drug-like properties.
Future Recommendations
Developing promising natural G4 stabilizers into effective therapies will require a multi-pronged approach. To begin with, it is important to develop advanced chemical probes, including clickable, fluorescent, or biotinylated derivatives of berberine and sanguinarine. Such probes will be used to support accurate target engagement research in live cells using techniques such as pull-down assays, CETSA, and imaging, directly relating G4 binding to phenotypic changes. Meanwhile, target validation should be further than correlations, involving genetic manipulation of G4-forming sequences or G4 helicases with the help of CRISPR to confirm causality – eg., by demonstrating the relationship between G4 stabilization and c-MYC inhibition by quercetin.90 Rational design and semi-synthesis should also be embraced in the field. Based on the structural data of NMR and crystallographies of the ligands-G4 complexes, the authors design the base modifications using the natural scaffolds, either modifying the glycoside component of rutin to display a higher level of selectivity96 or modifying berberine to enhance π-π stacking and electrostatic interactions.124 The solution to poor pharmacokinetics involves cutting-edge delivery methods, such as nanoparticle encapsulation and aptamer-based targeting, for allicin.116 Lastly, systems-wide G4-ome investigations of treated cells using transcriptomics and proteomics will be useful for mapping cellular responses, novel G4-regulated pathways, and identifying biomarkers to stratify patients in future clinical trials.
G-Quadruplex-Containing Genes Involved in Breast Cancer
The central drivers of BC pathogenesis are the oncogenes c-MYC, KRAS, VEGF, BCL2, HER2, ESR1, MDM2, EGFR, and c-KIT that control a broader spectrum of malignant phenotypes, such as uncontrolled cell growth and proliferation, resistance to apoptosis, metastasis, and angiogenesis, and their inhibition is a major therapeutic objective.125–128 Equally, KRAS is a highly conserved critical signaling pathway switchboard that facilitates cell survival and proliferation, and its expression is frequently triggered by G4 stabilization in its promoter.129 VEGF is a key angiogenic protein; its expression drives tumor vascularization and metastasis, and its transcription is also regulated by G4-forming sequences, making it a direct target for intervention.130,131 BCL2 is a strong anti-apoptotic protein that prevents programmed cell death in cancer cells, and its expression can be intricately regulated by G4 structures and alternative splicing, making it an important therapeutic target.12 HER2 is a tyrosine kinase receptor overexpressed in a substantial proportion of BCs and causes aggressive tumor growth, making it an ideal target for therapies that interfere with its expression or dimerization.10 The estrogen receptor-α (ESR1) contributes to hormone-sensitive BCs, and its translation is regulated by G4 structures in its transcript, providing a new avenue for inhibiting its activity beyond conventional hormone therapy.132 The overexpression of MDM2 is a negative regulator of the tumor suppressor p53, and it has been effectively shown that MDM2 overexpression renders a potent anticancer barrier inactive; inhibition of MDM2 transcription through promoter G4s can promote p53 activity.133 Lastly, c-KIT is a receptor tyrosine kinase, the aberrant expression of which is associated with poor prognosis and whose transcription is tightly regulated by G4 structures that control transcription factor recruitment.134 The significance of targeting these genes is incontrovertible, as dysregulation of their combination underlies the BC hallmarks.
One particularly important target is the c-MYC oncogene, whose promoter G4 is a key regulatory element. Transcription and cell growth can be prevented by stabilizing this G4.6,7 G4s in BRCA1 are useful for coordinating epigenetic regulation by linking DNA structure to gene silencing by methylation.24 Besides, angiogenesis is controlled by G4 structures in the VEGF promoter.130 Other notable genes include hTERT, which encodes telomerase;8 KRAS, which modulates signaling pathways,127 and EGFR,129 which has a unique G4 topology. On the whole, these studies demonstrate that gene-specific G4s are major controllers of cancer-related networks and can serve as targets for BC therapy.
Table 4 provides an overview of the research gaps and prospects of G4-targeted therapy in BC. The major lapses include the absence of in vivo validation. To highlight, BRCA1 is associated with G4-induced methylation and tumor growth in mouse models, but its functional role in human BC and the mechanistic link between G4 structures and methylation remain unclear.24 For EGFR and MDM2, the lack of selective ligands has been studied, or their metastatic and subtype-specific biological role are understudied.129,133
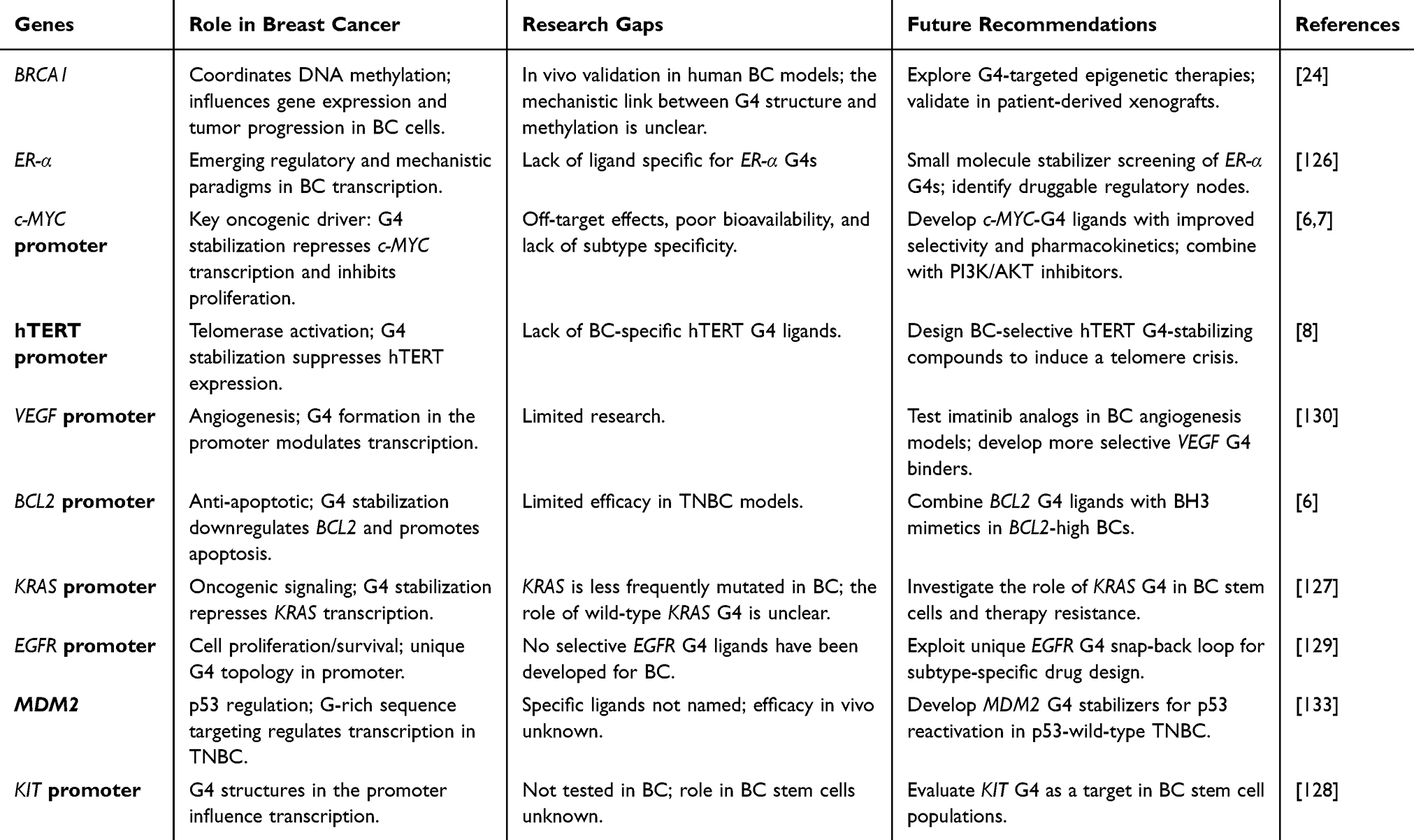 |
Table 4 G4-Targeted Therapy in Breast Cancer |
Concerns remain about off-target effects, low bioavailability, and low efficacy in aggressive subtypes such as TNBC, even in the presence of ligands, eg., c-MYC and BCL2.6,7 Finally, the reference to the clinical relevance of global G4 landscape mapping is speculative, and no direct association with the patient outcomes has been demonstrated.25 Recent research finds that FDA-approved drugs, such as azelastine, belotecan, and irinotecan, are effective G4 binders with antiproliferative effects in BC cells. Belotecan and irinotecan showed synergistic activity by stabilizing G4s and inhibiting topoisomerase I, increasing cytotoxicity. This confirms G4 stabilization’s potential in BC treatment and suggests combining it with other drugs could improve outcomes.135
The following recommendations can address future knowledge gaps about the findings and investigate the background mechanisms. A lot of work is needed to develop and optimize improved G4 ligands with improved selectivity and pharmacokinetics, particularly against high-value proteins such as c-MYC and hTERT.6,8 Strategy combinations are also highlighted, such as using the BCL2-G4 ligands with BH3 mimetics in BCL2-high cancer or using the c-MYC-G4 ligands in combination with PI3K/AKT inhibitors to enhance response and overcome resistance.6,7 For less-studied targets, future studies should include functional screening using CRISPR to verify the functions of conserved non-coding elements and G4s, and determine the capabilities of ER-α promoter G4s in hormone-resistant BC.126 Research on the application of G4s in BC-targeting KIT and KRAS may provide novel treatment methods.127,128 Finally, to move forward to clinical use, it is necessary to formulate prognostic signatures using G4 mapping of tumor biopsies and develop methods to target tumors, that is, a system of nanoparticles, to minimize risks associated with genomic instability.25 Such focused initiatives can help G4 biology transform from a regulatory phenomenon to a provider of specific therapies for BC.
Roadmap: Discovery of Natural Products as G-Quadruplexes Stabilizers for Breast Cancer Therapy
Research on designing specific BC treatments requires creative approaches that would go to the source of oncogenic expression. Attention can be given to DNA G4s, non-standard nucleic acid structures that occur in the promoter regions of key oncogenes such as c-MYC, KRAS, and BCL2, which are attractive targets of treatment. Oncogene expression, tumor growth, and cancer cell apoptosis can be selectively inhibited by stabilizing these structures, thereby interfering with tumor progression. Nevertheless, the transformation of G4-targeted agents into useful clinical therapies is a multi-step process that includes the identification and biophysical characterization of targets as well as the development of cellular and animal models. This is a detailed plan to identify and develop natural product-based G4 stabilizers in BC, integrating computational predictions, experimental biophysics, mechanistic simulations, functional genomics, and preclinical testing (Figure 3). Based on sophisticated methods such as AI screening, single-molecule biophysics, and multi-omics, the approach will generate selective, effective G4 ligands with strong potential for clinical use.
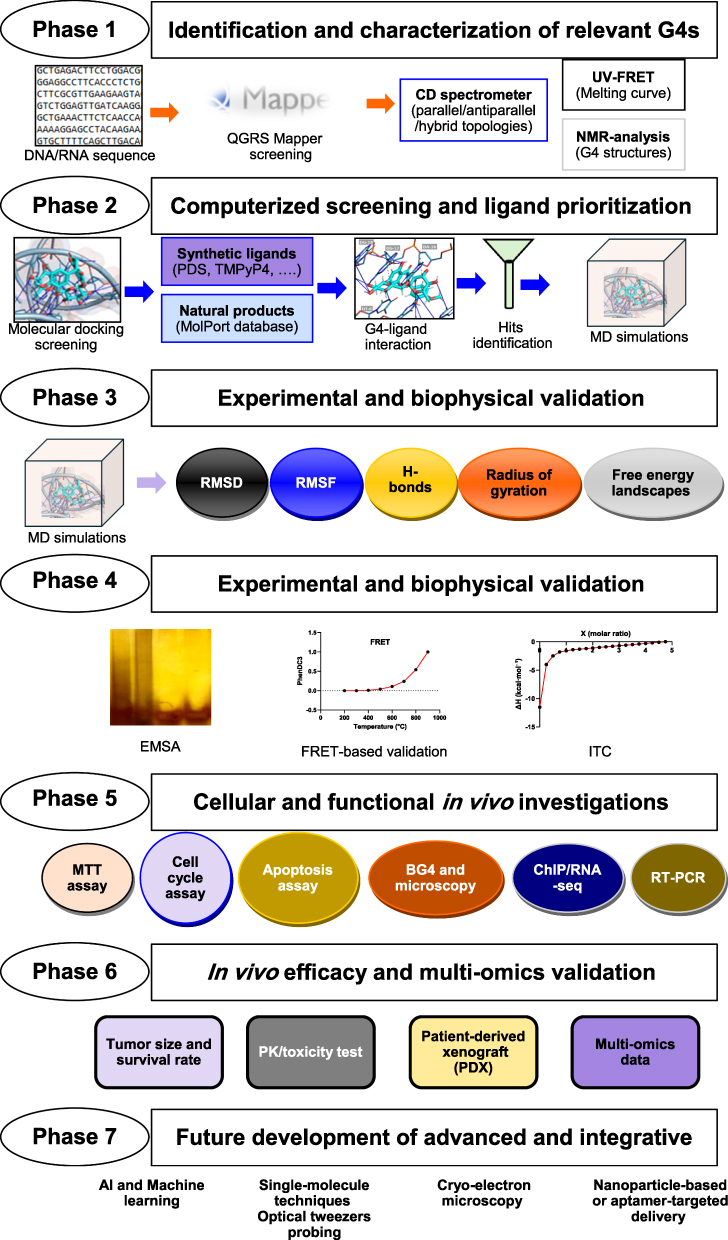 |
Figure 3 Future roadmap for the discovery of natural products as G-quadruplexes stabilizers for breast cancer therapy. |
Phase 1: Identification and Characterization of Relevant G4s in Breast Cancer
The first phase aims to properly identify and provide a biophysical description of G4-forming sequences in the main oncogenes and regulatory elements that participate in the BC progression. A key in this phase is the development of a “G4 priority score” that combines: (i) in silico prediction quality (G-score from QGRS Mapper), (ii) BC subtype-specific expression levels of the downstream gene (from TCGA), (iii) evidence of natural product binding from literature, and (iv) ligandability, indicated by the presence of solvent-accessible G4 tetrads. This scoring system aims to identify targets most likely to respond effectively to natural product intervention. Recent genome-wide G4 mapping in BC shows BC-G4s mainly in promoters and first introns near transcription start site, providing high-confidence targets for prioritization.11 Furthermore, the experimental validation is necessary after the prediction. The key technique for studying the topology (parallel, antiparallel, and hybrid) of the G4 structures is CD spectroscopy.26 Using UV or FRET, thermal melting assays will be used to measure the stability (Tm) of these G4s and serve as a reference point to assess the effects of ligand stabilization.27 For high-value targets, delicate examination methods such as NMR may be used to provide detailed atomic structures, which are crucial in rational drug design.28 This stage leads to the approval of G4 BC-specific targets for subsequent screening.
Phase 2: Computerized Screening and Ligand Prioritization
This step involves a layerwise computational method for discovering promising G4 stabilizers. To measure their binding poses, affinity scores, and interaction profiles, molecular docking will first be performed with known synthetic G4 stabilizers (such as pyridostatin, TMPyP4, and CX-5461) to assess these properties. The docking experiments will then incorporate natural products reported to be G4 stabilizers, such as berberine, quercetin, curcumin, and sanguinarine, to determine how they bind their BC targets and identify the most promising scaffolds. Finally, a virtual screen of commercially accessible natural product collections (eg., MolPort) will also be performed to discover new chemotypes. The process of docking scores, interaction analysis (such as π-π stacking with G-tetrads and groove binding), and initial drug-likeness screening will be used to narrow down to a final shortlist of 20–30 best hits for laboratory testing.26 The best candidates can subsequently be subjected to MD simulations to test binding stability and behavior.6
In this phase, we address the “synthetic bias” in current docking algorithms, as most docking scoring functions (AutoDock Vina or Glide) are parameterized using protein-ligand and duplex DNA-ligand complexes, rather than G4-ligand structures. This introduces systematic errors in predicting π-π stacking with G-tetrads.
Phase 3: Mechanistic Modeling with Simulations of Molecular Dynamics
For the most tested compounds, all-atom MD simulations will be conducted to gain deeper insight into the binding mechanism. Such simulations of the 100–200-ns-long ligand-G4 complex will evaluate the stability of the binding pose in terms of root-mean-square deviation (RMSD), the flexibility of the residues via root-mean-square fluctuation (RMSF), and the dynamics of the interaction network.37 We recommend that future MD simulations also report residence time (kinetics) and not just RMSD (stability), as residence time may be a better predictor of cellular potency than binding affinity. Finally, be aware of cation coordination when performing MD simulations of G4‑ligand complexes, as incorrect parameters can destabilize the G4 structure. Perform simulations of at least 500 ns to adequately sample conformational space and identify metastable intermediates in the binding process, which help to map binding pathways.
The main stabilizing forces, ie., exact π-π stacking with guanine quartets, hydrogen bonds with loops or grooves, and hydration effects, can be identified by this analysis. MD can also be used to understand the efficacy disparities between analogous compounds and to design superior derivatives by identifying key interaction sites.6
Phase 4: Experimental and Biophysical Validation
The main test is the FRET-based melting test, which measures the increase in the G4 melting temperature upon ligand binding, reflecting the strength of stabilization.27 Isothermal titration calorimetry will be used to measure binding affinity and stoichiometry and to provide detailed thermodynamic information.26 Competition dialysis is used to assess ligand binding selectively to G4 versus duplex DNA to determine how the ligand is distributed in various forms of DNA.36 Also, the formation of stable ligand-G4 complexes will be confirmed graphically by EMSA or native PAGE.32,33 Such mixed procedures will confirm and prioritize the G4-stabilizing property and selectivity of the natural product candidates. Furthermore, we strongly advise conducting all biophysical validation experiments in at least two separate laboratories, using standardized buffer conditions (such as 10 mM Tris‑HCl and 100 mM KCl, pH 7.4). Additionally, all data, including negative results, should be uploaded to a public repository.
Phase 5: Cellular and Functional in vivo Investigations of Breast Cancer Models
G4 stabilization effects will be analyzed in BC cell lines. At first, cell viability and anti-proliferative effects will be measured using the MTT test or other similar tests.6 Flow cytometry will help analyze cell cycle phases and apoptosis induction. To confirm the involvement of targets, immunofluorescence microscopy will be used to assess changes in nuclear G4 foci after treatment using the G4-specific BG4 antibody. Suppression of oncogenes, such as c-MYC and KRAS, will be quantified by the RT-PCR method as well as by the luminescence reporter assays forced by G4-containing promoters. To obtain epigenetic and transcriptomic data, the ChIP will validate in vivo G4 formation and the histone modifications. Also, RNA-seq can provide a comprehensive view of gene expression, potentially revealing synthetic lethals or pathways affected by G4 stabilization.31 Lastly, in cellular studies, controls such as G4-mutant promoter constructs (with point mutations in the G4-forming region) should be used. This ensures the transcriptional repression is indeed G4-specific and not simply due to toxicity.
Phase 6: In vivo Efficacy and Multi-Omics Validation
The last preclinical trial is an in vivo preclinical efficacy trial. The most popular natural product will be used to treat BC patient-derived xenograft models, particularly those with aggressive types of disease, such as TNBC.9 Growth and metastasis of the tumor, as well as survival rates, will be observed. Pharmacokinetic and toxicity tests shall be done. Multi-omic analysis of the treatment will be performed by collecting tumors to assess genomic stability, RNA-seq transcriptomics to confirm oncogene silencing and identify response biomarkers, and proteomic analysis to assess the effect of the downstream pathway.25 This combined approach links G4-targeted therapy with phenotypic outcome and provides a holistic set of data to comprehend the mechanism of action and potential resistance.
Given recent findings that sustained G4 stabilization can cause lasting transcriptomic changes and trigger mitophagy in healthy cells, all in vivo efficacy studies should include thorough toxicological assessments of key organs (liver, kidney, heart, and bone marrow) and long-term monitoring to detect delayed toxic effects.
Phase 7: Future Development of Advanced and Integrative Tools
To make this pipeline even better, several sophisticated tools should be used. AI and machine learning models can be trained with screening data to increase the accuracy of virtual screening and detect new G4-binding chemotypes.136 To obtain the structure of natural product-G4 complexes of difficult targets, cryo-electron microscopy can be applied. To address the bioavailability issues that natural products may have, it may be considered to investigate methods of nanoparticle-based or aptamer-targeted delivery systems to enhance tumor-targeted delivery and performance.116 In addition, recent nanotechnology formulations, such as curcumin/piperine nanoemulsions and polysaccharide-coated mPEG-PLA nanoparticles for thymoquinone, improve stability and targeted delivery, but still need safety validation.137,138
This comprehensive research proposal is a systematic and stringent step towards transforming the G4 biology into novel BC treatments. Its power lies in its rational, multi-stage approach, from the identification of the computational target up to the in vivo testing. It provides a feedback mechanism in which theories of behavior predict experiments and biophysics discovers ways to optimize compounds. The strategy leverages natural products with diverse bioactive chemical structures and seeks to overcome pharmacokinetic challenges by using advanced formulation technologies. The strategies integrate various new technologies, including AI screens, single-molecule biophysics, patient-derived xenografts, and multi-omics validation. This ensures the effectiveness of potential drugs as potent G4 stabilizers, selective, active, and with a clear mechanism of action. Overall, the strategy helps bridge the knowledge gap between nucleic acid structure and therapy, and offers an integrated strategy to generate targeted transcription inhibitors as part of cancer therapy.
Finally, we suggest a “fail-early” approach to reduce the high failure rate in natural product development. Using the S3 framework, natural products should be simultaneously screened for: (i) G4 selectivity (competition dialysis), (ii) stability in mouse/human plasma (37°C, 24 h), and (iii) solubility in PBS (pH 7.4). Those that do not meet these criteria should be deprioritized or modified to make semisynthetic derivatives before in vivo testing. Adopting this strategy may improve the current attrition rate, which exceeds 90% in natural products discovery for cancer therapy.
Conclusion
G4s are highly attractive non-canonical nucleic acid structures that are now being considered key regulatory factors in BC biology. Their overexpression in the promoters of key oncogenes, such as c-MYC, KRAS, BCL2, HER2, and ESR1, makes them a promising therapeutic target. Despite progress in synthetic G4 stabilizers to understand G4 biology and demonstrate preclinical potential, the natural products remain an underutilized but promising source of selective, biocompatible, and multitarget ligands. The review critically examines a wide range of natural compounds, including flavonoids, alkaloids, polyphenols, terpenoids, and organosulfur compounds, with G4-stabilizing activity and relevance to BC pathways.
This review clearly shows that natural G4 stabilizers primarily act through three non-covalent mechanisms: π-π stacking with terminal tetrads, electrostatic interactions with the phosphate backbone (notably in cationic alkaloids, such as berberine and sanguinarine), and groove binding. An important insight from this study is the evident structure-activity divergence: many flavonoids and alkaloids act as broad-spectrum stabilizers targeting various oncogenic promoters (c-MYC, VEGF, and KRAS), whereas compounds like piperine and glycosides such as rutin display more selective activity, which remains underused. Additionally, a consistent gap exists between “potency, selectivity, and safety”: while synthetic stabilizers reach nanomolar potency, natural products tend to be more biocompatible and target multiple genes but fall short in specificity and pharmacokinetics.
Beyond mechanistic insights, this review highlights three key practical implications addressing critical barriers in the field. First, our systematic mapping of binding mechanisms to specific BC-relevant G4 targets (c-MYC, KRAS, BCL2, HER2, ESR1, VEGF, and hTERT) provides a logical framework for guiding natural product screening and structure refinement. For example, the increased G4 selectivity of rutin compared to its aglycone quercetin suggests that glycosylation is an underexplored structural modification for enhanced target discrimination—immediately relevant for medicinal chemistry. Second, the proposed seven-phase translational roadmap aims to solve the clinical translation bottleneck common to both synthetic and natural G4 ligands. Third, this work directly addresses unmet clinical needs in BC, especially the difficult-to-treat TNBC and drug resistance across subtypes.
The significance of these findings for BC therapy is twofold. First, natural G4 ligands have a multi-targeting ability that aligns well with the need to overcome pathway redundancy, which causes resistance in aggressive BC subtypes like TNBC. Simultaneous targeting of c-MYC, BCL2, and KRAS by a natural product could address multiple cancer hallmarks. Second, natural products like quercetin and curcumin are safe, offering a potential edge over synthetic G4 ligands, which tend to have dose-limiting toxicities. Despite these exciting results, there is a critical gap in the research: a direct demonstration of G4 engagement and, ultimately, efficacy in biological systems.
This gap can only be filled by extending the research from biophysical (CD and FRET) to cellular-level studies to confirm target engagement with chemical tools. In order to address the gaps, we strongly recommend concentrating on the future directions: 1. Medicinal Chemistry: Focus on semi-synthetic modifications of specific scaffolds, such as glycosylation of rutin or tailoring of the aliphatic chain in piperine to dissociate G4 stabilization with unintentional duplex DNA binding. 2. Improved Delivery Methods: Employ nanotechnology or aptamer-based strategies to overcome problems of low solubility and rapid metabolism in promising candidates like curcumin, EGCG, and allicin. 3. Testing on BC Subtypes: Conduct extensive in vivo studies using patient-derived xenograft models of different BC subtypes (triple negative, HER2 overexpressing, or estrogen receptor positive) to determine the most effective natural G4 ligands for each particular genetic context. 4. Employ the S3 Framework: With a “fail-early” approach in drug discovery, which evaluates Selectivity (G4 vs. duplex), Stability (plasma half-life), and Solubility simultaneously, discover lead molecules with the potential for successful translation.
In conclusion, a multidisciplinary approach combining traditional pharmacognosy and modern drug discovery technologies, along with the rich chemical diversity of natural products, will enable the successful development of natural G4 stabilizers into valuable BC drugs. Shifting from merely inventorying natural products to a critical-thinking, mechanism-based approach that highlights target engagement and subtype specificity will unlock the potential of G4-targeted phytotherapy to develop more targeted, efficient, and personalized BC treatment strategies.
Abbreviations
CD, Circular dichroism; EGCG, Epigallocatechin gallate; EMSA, Electrophoretic mobility shift assays; ESR, Electron spin resonance; G4, G-quadruplex; MD, Molecular dynamics; NMR, Nuclear magnetic resonance; SPR, Surface plasmon resonance; UCP1, Mitochondrial uncoupling protein-1.
Data Sharing Statement
All data are available in the manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was funded by the General Program of Natural Science Foundation of Sichuan Province (Grant No.:2026NSFSC0562), Zigong Municipal Health Commission 2025 High-level Talent Science and Technology Incubation Project (Grant No.: WJW-GCCRC009), and the Zigong Public Hospital Reform and High-Quality Development Demonstration Project in 2023 (Grant No.: ZG-KY-2023-013).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Shiekh S, Kodikara SG, Balci H. Structure, topology, and stability of multiple G-quadruplexes in long telomeric overhangs. J Mol Biol. 2024;436:168205.
2. Barzegar A, Tohidifar N. Stabilization of Hoogsteen H-bonds in G-quartet sheets by coordinated K+ ion for enhanced efficiency in guanine-rich DNA nanomotor. Bioimpacts. 2025;15:30596. doi:10.34172/bi.30596
3. Fracchioni G, Vailati S, Grazioli M, Pirota V. Structural unfolding of G-quadruplexes: from small molecules to antisense strategies. Molecules. 2024;29(15):3488. doi:10.3390/molecules29153488
4. Mizumoto A, Yokoyama Y, Miyoshi T, et al. DHX36 maintains genomic integrity by unwinding G-quadruplexes. Genes Cells. 2023;28(10):694–23. doi:10.1111/gtc.13061
5. Gellert M, Lipsett MN, Davies DR. Helix formation by guanylic acid. Proc Natl Acad Sci. 1962;48:2013–2018. doi:10.1073/pnas.48.12.2013
6. Gao Z, Wang X, Li D, et al. Design, synthesis and evaluation of styrylquinolinium derivatives targeting c-MYC/BCL2 G-quadruplexes against breast cancer. Bioorg Chem. 2025;163:108665. doi:10.1016/j.bioorg.2025.108665
7. Roy A, Chatterjee O, Banerjee N, et al. Curcumin arrests G-quadruplex in the nuclear hyper-sensitive III 1 element of c-MYC oncogene leading to apoptosis in metastatic breast cancer cells. J Biomol Struct Dyn. 2022;40(20):10203–10219. doi:10.1080/07391102.2021.1940284
8. Mock PY, Chua EW, Stevens A, Salim F, Lam KW. Exploration of chromenone-based ligands as DNA G-quadruplex targets in triple-negative breast cancer. Bioorg Chem. 2025;166:109150. doi:10.1016/j.bioorg.2025.109150
9. Majocha MR, Jackson DE, Ha N-H, et al. Resf1 is a compound G4 quadruplex-associated tumor suppressor for triple negative breast cancer. PLoS Genet. 2024;20(5):e1011236. doi:10.1371/journal.pgen.1011236
10. Gao Y, Wang R, Dou X, et al. Patient-derived cell lines unveil COL1A2 as a predictor of docetaxel resistance in breast cancer. Front Oncol. 2025;15:1737405. doi:10.3389/fonc.2025.1737405
11. Shu H, Xiao K, Zhu W, et al. The relevance of G-Quadruplexes in gene promoters and the first introns associated with transcriptional regulation in breast cancer. Int J Mol Sci. 2025;26(14):6874. doi:10.3390/ijms26146874
12. Yang C, Wu S, Wang H, et al. G-quadruplex/m6A-regulated BCL2 pre-mRNA alternative splicing and comparative characterization of its two isoforms. Int J Biol Macromol. 2026;338:149670. doi:10.1016/j.ijbiomac.2025.149670
13. Sengupta P, Dutta A, Suseela YV, et al. G-quadruplex structural dynamics at MAPK12 promoter dictates transcriptional switch to determine stemness in breast cancer. Cell Mol Life Sci. 2024;81(1):33. doi:10.1007/s00018-023-05046-6
14. Wang K-B, Wang Y, Dickerhoff J, Yang D. DNA G-Quadruplexes as targets for natural product drug discovery. Engineering. 2024;38:39–51. doi:10.1016/j.eng.2024.03.015
15. Shan C, Tan J-H, Ou T-M, Huang Z-S. Natural products and their derivatives as G-quadruplex binding ligands. Sci China Chem. 2013;56:1351–1363. doi:10.1007/s11426-013-4920-y
16. Awadasseid A, Ma X, Wu Y, Zhang W. G-quadruplex stabilization via small-molecules as a potential anti-cancer strategy. Biomed Pharmacother. 2021;139:111550. doi:10.1016/j.biopha.2021.111550
17. Wali AF, Talath S, Babiker R, et al. Natural products as kinase inhibitors in lung cancer: molecular mechanisms, therapeutic potential, and clinical trials. Front Pharmacol. 2026;17:1764550. doi:10.3389/fphar.2026.1764550
18. El-Shehawy AA, Elmetwalli A, El-Far AH, et al. Thymoquinone, piperine, and sorafenib combinations attenuate liver and breast cancers progression: epigenetic and molecular docking approaches. BMC Complement Med Ther. 2023;23(1):69. doi:10.1186/s12906-023-03872-6
19. El-Far A, Liu X, Xiao T, et al. TQFL19, a novel derivative of thymoquinone (TQ), plays an essential role by inhibiting cell growth, metastasis, and invasion in triple-negative breast cancer. Molecules. 2025;30(4):773. doi:10.3390/molecules30040773
20. Meraz-Rodriguez MA, Cháirez-Ramírez MH, De la cruz-lópez KG, González-Laredo RF, García-Carrancá A. Polyphenols bind G4-Quadruplex structures and act as epigenetic modifiers with anti-cancer effects. EXCLI J. 2025;24:957–991. doi:10.17179/excli2025-8507
21. Raman R, Debata S, Govindarajan T, Kumar P. Targeting triple-negative breast cancer: resistance mechanisms and therapeutic advancements. Cancer Med. 2025;14(9):e70803. doi:10.1002/cam4.70803
22. Crintea A, Bocșan CI, Jianu EM, et al. Overcoming chemotherapy resistance in triple-negative breast cancer with nanocarrier-delivered siRNA therapeutics. J Clin Med. 2026;15(6):2311. doi:10.3390/jcm15062311
23. Figueiredo J, Mergny J-L, Cruz C. G-quadruplex ligands in cancer therapy: progress, challenges, and clinical perspectives. Life Sci. 2024;340:122481. doi:10.1016/j.lfs.2024.122481
24. Zhou X, Jin Z, Zhan X, et al. Expression of G-quadruplex coordinates BRCA1, CDH1, and RASSF1 via DNA methylation in mouse breast cancer cells. Epigenomics. 2025;17(18):1335–1346. doi:10.1080/17501911.2025.2577003
25. Hänsel-Hertsch R, Simeone A, Shea A, et al. Landscape of G-quadruplex DNA structural regions in breast cancer. Nat Genet. 2020;52(9):878–883. doi:10.1038/s41588-020-0672-8
26. Kumari P, Pandav K, Panwar A, et al. Elucidating the binding mechanism of 1,4-bis[(3-Piperidino)propanamido]anthracen-9,10-dione (1,4-BPAQ) to human telomeric G-quadruplexes: a multi-technique approach including spectroscopic, calorimetric and molecular docking insights. J Comput Aided Mol Des. 2025;39(1):37. doi:10.1007/s10822-025-00615-2
27. Çelik Ç, Kakusho N, Xu T, et al. Water-soluble cationic porphyrins with enhanced phototoxicity to cancer cell lines for G4-targeting photodynamic therapy. RSC Med Chem. 2026;17(1):225–235. doi:10.1039/D5MD00706B
28. Li X, Cevec M, Dubins DN, Plavec J, Chalikian TV. Binding of tetrapentylammonium to a c-MYC G-quadruplex depicted through NMR and volumetric assessment. Chemistry. 2025;31(67):e02670. doi:10.1002/chem.202502670
29. Banerjee N, Roy L, Panda S, Roychowdhury T, Chatterjee S. In silico-designed G-Quadruplex targeting peptide attenuates VEGF-A expression, preventing angiogenesis in cancer cells. Chem Biol Drug Des. 2024;104(6):e70018. doi:10.1111/cbdd.70018
30. Okamoto C, Kotani H, Kojima T, Yamamoto Y, Momotake A. Mechanistic insights into oxygen-independent DNA photodegradation by pheophorbide a: implications for future photodynamic therapy applications. Chembiochem. 2025;26:e202400781. doi:10.1002/cbic.202400781
31. Soriano-Lerma A, Sánchez-Martín V, Murciano-Calles J, et al. In-depth analysis of the mode of action of resveratrol: genome-wide characterization of G-quadruplex binding properties. Cell Mol Biol Lett. 2025;30(1):74. doi:10.1186/s11658-025-00747-1
32. Verma SP, Das P. G-quadruplex structure at intron 2 of TFE3 and its role in Xp11.2 translocation and splicing. Biochim Biophys Acta Gen Subj. 2018;1862:630–636. doi:10.1016/j.bbagen.2017.11.011
33. Ye H, Zhang H, Chen H, et al. Structural and functional insights into targeting hTERT G-quadruplex by levo-Tetrahydropalmatine in the non-small cell lung cancer. Bioorg Med Chem. 2025;131:118411. doi:10.1016/j.bmc.2025.118411
34. Krasitskaya VV, Shchugoreva IA, Rogova AV, et al. Structure-affinity balance of anti-cardiac troponin I aptamer: effects of sequence truncation. Int J Biol Macromol. 2026;341:150299. doi:10.1016/j.ijbiomac.2026.150299
35. Lago S, Nadai M, Rossetto M, Richter SN. Surface plasmon resonance kinetic analysis of the interaction between G-quadruplex nucleic acids and an anti-G-quadruplex monoclonal antibody. Biochim Biophys Acta Gen Subj. 2018;1862:1276–1282. doi:10.1016/j.bbagen.2018.03.002
36. Yıldız U, Gökçek Ö. A comparison of human telomeric G-quadruplex-double stranded DNA interactions of newly synthesized Pt(II) complex of benzimidazole based ligand. Nucleosides Nucleotides Nucleic Acids. 2025;1–15. doi:10.1080/15257770.2025.2586132
37. Vinnarasi S, Radhika R, Vijayakumar S, Shankar R. Structural insights into the anti-cancer activity of quercetin on G-tetrad, mixed G-tetrad, and G-quadruplex DNA using quantum chemical and molecular dynamics simulations. J Biomol Struct Dyn. 2020;38:317–339. doi:10.1080/07391102.2019.1574239
38. Panova VV, Dolinnaya NG, Novoselov KA, et al. Conserved G-Quadruplex-Forming sequences in mammalian TERT promoters and their effect on mutation frequency. Life. 2023;13(7):1478. doi:10.3390/life13071478
39. Hänsel-Hertsch R, Spiegel J, Marsico G, Tannahill D, Balasubramanian S. Genome-wide mapping of endogenous G-quadruplex DNA structures by chromatin immunoprecipitation and high-throughput sequencing. Nat Protoc. 2018;13:551–564. doi:10.1038/nprot.2017.150
40. He X, Yuan J, Gao Z, Wang Y. Promoter R-Loops recruit U2AF1 to modulate its phase separation and RNA splicing. J Am Chem Soc. 2023;145:21646–21660. doi:10.1021/jacs.3c08204
41. Lu X, Passalacqua LM, Nodwell M, et al. Symmetry breaking of fluorophore binding to a G-quadruplex generates an RNA aptamer with picomolar KD. Nucleic Acids Res. 2024;52(14):8039–8051. doi:10.1093/nar/gkae493
42. Sannikova NE, Kolokolov MI, Khlynova TA, et al. Revealing light-induced structural shifts in G-quadruplex-porphyrin complexes: a pulsed dipolar EPR study. Phys Chem Chem Phys. 2023;25(33):22455–22466. doi:10.1039/D3CP01775C
43. Wen L, Han Z, Li J, Du Y. c-MYC and HIF1α promoter G-quadruplexes dependent metabolic regulation mechanism of berberine in colon cancer. J Gastrointest Oncol. 2022;13:1152–1168. doi:10.21037/jgo-22-389
44. Pradhan S, Campanile M, Sharma S, Oliva R, Patra S. Mechanistic Insights into the c-MYC G-Quadruplex and berberine binding inside an aqueous two-phase system mimicking biomolecular condensates. J Phys Chem Lett. 2024;15:8706–8714. doi:10.1021/acs.jpclett.4c01806
45. Dickerhoff J, Brundridge N, McLuckey SA, Yang D. Berberine molecular recognition of the parallel MYC G-Quadruplex in solution. J Med Chem. 2021;64:16205–16212. doi:10.1021/acs.jmedchem.1c01508
46. Wang J-T, Li Y, Tan J-H, Ji L-N, Mao Z-W. Platinum(II)-triarylpyridines complexes with electropositive pendants as efficient G-quadruplex binders. Dalton Trans. 2011;40:564–566. doi:10.1039/C0DT01161D
47. Wang J-T, Zheng X-H, Xia Q, et al. 1,10-Phenanthroline platinum(II) complex: a simple molecule for efficient G-quadruplex stabilization. Dalton Trans. 2010;39(31):7214–7216. doi:10.1039/c0dt00211a
48. Xiong K, Ouyang C, Liu J, et al. Chiral Ru II -Pt II complexes inducing telomere dysfunction against cisplatin-resistant cancer cells. Angew Chem Int Ed Engl. 2022;61(33):e202204866. doi:10.1002/anie.202204866
49. Zheng X-H, Chen H-Y, Tong M-L, Ji L-N, Mao Z-W. Platinum squares with high selectivity and affinity for human telomeric G-quadruplexes. Chem Commun. 2012;48:7607–7609. doi:10.1039/c2cc32942e
50. Xu C-X, Zheng Y-X, Zheng X-H, et al. V-shaped dinuclear Pt(II) complexes: selective interaction with human telomeric G-quadruplex and significant inhibition towards telomerase. Sci Rep. 2013;3(1):2060. doi:10.1038/srep02060
51. Xu C-X, Shen Y, Hu Q, et al. Stabilization of human telomeric G-Quadruplex and inhibition of telomerase activity by propeller-shaped trinuclear Pt II complexes. Chem Asian J. 2014;9(9):2519–2526. doi:10.1002/asia.201402258
52. Wang X-D, Li Z-M, Liu X-Z, Hu M-H. Flexible long-chain substituted quinoxalines as G-quadruplex binders exert antitumor effects via cGAS-STING immunomodulatory pathway. Int J Biol Macromol. 2025;321:146352. doi:10.1016/j.ijbiomac.2025.146352
53. Wang Z, Zhou J, Lin L, Hu M-H. Discovery of a far-red carbazole-benzoindolium fluorescent ligand that selectively targets mitochondrial DNA and suppresses breast cancer growth. Eur J Med Chem. 2024;264:116046. doi:10.1016/j.ejmech.2023.116046
54. Yan J, Bu L, Wang Y, et al. Pyridine-bis(benzimidazole) induces DNA damage at G-quadruplex loci and promotes synthetic lethality with DNA repair inhibition. Nucleic Acids Res. 2025;53(12):gkaf543. doi:10.1093/nar/gkaf543
55. Yadav K, Meka PNR, Sadhu S, et al. Telomerase inhibition and human telomeric G-Quadruplex DNA stabilization by a β-Carboline-Benzimidazole derivative at low concentrations. Biochemistry. 2017;56(33):4392–4404. doi:10.1021/acs.biochem.7b00008
56. Wang X-D, Liu Y-S, Chen M-D, Hu M-H. Discovery of a triphenylamine-based ligand that targets mitochondrial DNA G-quadruplexes and activates the cGAS-STING immunomodulatory pathway. Eur J Med Chem. 2024;269:116361. doi:10.1016/j.ejmech.2024.116361
57. Hilton J, Gelmon K, Bedard PL, et al. Results of the phase I CCTG IND.231 trial of CX-5461 in patients with advanced solid tumors enriched for DNA-repair deficiencies. Nat Commun. 2022;13(1):3607. doi:10.1038/s41467-022-31199-2
58. Bradford T, Summers PA, Majid A, et al. Imaging G-Quadruplex nucleic acids in live cells using thioflavin T and fluorescence lifetime imaging microscopy. Analy Chem. 2024;96(51):20223–20229. doi:10.1021/acs.analchem.4c04207
59. Prasad B, Jamroskovic J, Bhowmik S, et al. Flexible versus rigid G-Quadruplex DNA ligands: synthesis of two series of bis-indole derivatives and comparison of their interactions with G-Quadruplex DNA. Chemistry. 2018;24(31):7926–7938. doi:10.1002/chem.201800078
60. Larsen AF, Nielsen MC, Ulven T. Tetrasubstituted phenanthrolines as highly potent, water-soluble, and selective G-quadruplex ligands. Chemistry. 2012;18(35):10892–10902. doi:10.1002/chem.201200081
61. Chilakamarthi U, Koteshwar D, Jinka S, et al. Novel amphiphilic G-Quadruplex binding synthetic derivative of TMPyP4 and its effect on cancer cell proliferation and apoptosis induction. Biochemistry. 2018;57(46):6514–6527. doi:10.1021/acs.biochem.8b00843
62. Nakamura T, Okabe S, Yoshida H, et al. Targeting glioma stem cells in vivo by a G-quadruplex-stabilizing synthetic macrocyclic hexaoxazole. Sci Rep. 2017;7(1):3605. doi:10.1038/s41598-017-03785-8
63. Laguerre A, Stefan L, Larrouy M, et al. A twice-as-smart synthetic G-Quartet: pyroTASQ is both a smart quadruplex ligand and a smart fluorescent probe. J Am Chem Soc. 2014;136(35):12406–12414. doi:10.1021/ja506331x
64. Wang X-D, Wang J-X, Yu B-Y, Zhang S-Q, Hu M-H. Non-fused imidazole-biphenyl analogs repress triple-negative breast cancer growth by mainly stabilizing the c-MYC G-quadruplex via a multi-site binding mode. Bioorg Med Chem. 2023;88-89:117336. doi:10.1016/j.bmc.2023.117336
65. Peng W, Sun Z-Y, Zhang Q, et al. Design, synthesis, and evaluation of novel p -(Methylthio)styryl substituted quindoline derivatives as neuroblastoma RAS (NRAS) repressors via specific stabilizing the RNA G-Quadruplex. J Med Chem. 2018;61(15):6629–6646. doi:10.1021/acs.jmedchem.8b00257
66. Pandya N, Rani R, Kumar V, Kumar A. Discovery of a potent guanidine derivative that selectively binds and stabilizes the human BCL-2 G-quadruplex DNA and downregulates the transcription. Gene. 2023;851:146975. doi:10.1016/j.gene.2022.146975
67. Liang B, Zhu J, Yu H, et al. Guanine amine derivatives promote forming a Base-Vacancy-G quadruplex structure. ACS Omega. 2025;10(4):3820–3827. doi:10.1021/acsomega.4c09242
68. Masud T, Soong C, Xu H, et al. Ubiquitin-mediated DNA damage response is synthetic lethal with G-quadruplex stabilizer CX-5461. Sci Rep. 2021;11(1):9812. doi:10.1038/s41598-021-88988-w
69. Germoglio M, D’Aria F, Cortone G, et al. Effects of hydrazone-based G-quadruplex ligands on FANCJ/BRIP1 -depleted cancer cells and a caenorhabditis elegans dog-1−/− strain. NAR Cancer. 2025;7(1):zcaf004. doi:10.1093/narcan/zcaf004
70. Zell J, Duskova K, Chouh L, et al. Dual targeting of higher-order DNA structures by azacryptands induces DNA junction-mediated DNA damage in cancer cells. Nucleic Acids Res. 2021;49(18):10275–10288. doi:10.1093/nar/gkab796
71. Rota Sperti F, Dupouy B, Mitteaux J, et al. Click-Chemistry-Based biomimetic ligands efficiently capture G-quadruplexes in vitro and help localize them at DNA damage sites in human cells. JACS Au. 2022;2(7):1588–1595. doi:10.1021/jacsau.2c00082
72. Choudhary NK, Gupta S, Das G, et al. Selective recognition of the dimeric NG16 parallel G-Quadruplex structure using synthetic turn-on red fluorescent protein chromophore. Biochemistry. 2024;63(21):2842–2854. doi:10.1021/acs.biochem.4c00407
73. Li H-X, He Y-M, Fei J, et al. The G-quadruplex ligand CX-5461: an innovative candidate for disease treatment. J Transl Med. 2025;23(1):457. doi:10.1186/s12967-025-06473-8
74. Bigot T, Gabinaud E, Hannouche L, et al. Single-cell analysis of megakaryopoiesis in peripheral CD34+ cells: insights into ETV6-related thrombocytopenia. J Thromb Haemost. 2023;21(9):2528–2544. doi:10.1016/j.jtha.2023.04.007
75. Liu T, Pan G, Zhang J, et al. Molecular basis of CX-5461-induced DNA damage response in primary vascular smooth muscle cells. Heliyon. 2024;10(17):e37227. doi:10.1016/j.heliyon.2024.e37227
76. Moruno-Manchon JF, Koellhoffer EC, Gopakumar J, et al. The G-quadruplex DNA stabilizing drug pyridostatin promotes DNA damage and downregulates transcription of Brca1 in neurons. Aging. 2017;9(9):1957–1970. doi:10.18632/aging.101282
77. Libera V, Fasolato C, Ripanti F, et al. Molecular mechanisms behind BRACO19 and human telomeric G-quadruplex interaction. Spectrochim Acta Part A. 2024;322:124684. doi:10.1016/j.saa.2024.124684
78. Singh A, Joshi S, Kukreti S. Cationic porphyrins as destabilizer of a G-quadruplex located at the promoter of human MYH7 β gene. J Biomol Struct Dyn. 2020;38:4801–4816. doi:10.1080/07391102.2019.1689850
79. Parkinson GN, Ghosh R, Neidle S. Structural basis for binding of porphyrin to human telomeres. Biochemistry. 2007;46:2390–2397. doi:10.1021/bi062244n
80. Morris MJ, Wingate KL, Silwal J, Leeper TC, Basu S. The porphyrin TmPyP4 unfolds the extremely stable G-quadruplex in MT3-MMP mRNA and alleviates its repressive effect to enhance translation in eukaryotic cells. Nucleic Acids Res. 2012;40:4137–4145. doi:10.1093/nar/gkr1308
81. Singh P, Sahel DK, Jatyan R, et al. Enhancing temozolomide in vivo stability and efficacy through hybrid nanoconjugate approach for improved glioblastoma multiforme treatment. Asian J Pharm Sci. 2025;20(3):101022. doi:10.1016/j.ajps.2025.101022
82. Sun J-W, Zou J, Zheng Y, et al. Design, synthesis, and evaluation of novel quindoline derivatives with fork-shaped side chains as RNA G-quadruplex stabilizers for repressing oncogene NRAS translation. Eur J Med Chem. 2024;271:116406. doi:10.1016/j.ejmech.2024.116406
83. Park JH, Lee HS, Jang MD, et al. Enantioselective light switch effect of Δ- and Λ-[Ru(phenanthroline) 2 dipyrido[3,2-a:2′, 3′-c]phenazine] 2+ bound to G-quadruplex DNA. J Biomol Struct Dyn. 2018;36(8):1948–1957. doi:10.1080/07391102.2017.1345324
84. Xu H, Hurley LH. A first-in-class clinical G-quadruplex-targeting drug. The bench-to-bedside translation of the fluoroquinolone QQ58 to CX-5461 (Pidnarulex). Bioorg Med Chem Lett. 2022;77:129016. doi:10.1016/j.bmcl.2022.129016
85. Jin M, Hurley LH, Xu H. A synthetic lethal approach to drug targeting of G-quadruplexes based on CX-5461. Bioorg Med Chem Lett. 2023;91:129384. doi:10.1016/j.bmcl.2023.129384
86. Jonchhe S, Lahiri S, Rothenberg E. DNA G-quadruplexes: structural and functional insights. DNA Repair. 2025;156:103910. doi:10.1016/j.dnarep.2025.103910
87. Tyagi M. DNA repair in cancers: why is there an alternative? Med Oncol. 2025;42:66. doi:10.1007/s12032-024-02581-5
88. Zenkov RG, Kirsanov KI, Ogloblina AM, et al. Effects of G-Quadruplex-Binding plant secondary metabolites on c-MYC expression. Int J Mol Sci. 2022;23(16):9209. doi:10.3390/ijms23169209
89. Bhattacharjee S, Sengupta PK, Bhowmik S. Exploring the preferential interaction of quercetin with VEGF promoter G-quadruplex DNA and construction of a pH-dependent DNA-based logic gate. RSC Adv. 2017;7:37230–37240. doi:10.1039/C7RA05930B
90. Tawani A, Mishra SK, Kumar A. Structural insight for the recognition of G-quadruplex structure at human c-myc promoter sequence by flavonoid quercetin. Sci Rep. 2017;7:3600. doi:10.1038/s41598-017-03906-3
91. Bhattacharjee S, Chakraborty S, Chorell E, Sengupta PK, Bhowmik S. Importance of the hydroxyl substituents in the B-ring of plant flavonols on their preferential binding interactions with VEGF G-quadruplex DNA: multi-spectroscopic and molecular modeling studies. Int J Biol Macromol. 2018;118:629–639. doi:10.1016/j.ijbiomac.2018.06.115
92. Tawani A, Kumar A. Structural insight into the interaction of flavonoids with human telomeric sequence. Sci Rep. 2015;5:17574. doi:10.1038/srep17574
93. Tang J. Using UV–Vis Titration to Elucidate Novel Epigallocatechin Gallate (EGCG)-Induced binding of the c-MYC G-Quadruplex. Pharmaceuticals. 2025;18:719. doi:10.3390/ph18050719
94. Das A, Dutta S. Binding studies of aloe-active compounds with G-Quadruplex sequences. ACS Omega. 2021;6:18344–18351. doi:10.1021/acsomega.1c02207
95. Das A, Chakraborty J, Luikham S, et al. Targeting aloe active compounds to c-KIT promoter G-quadruplex and comparative study of their anti proliferative property. J Biomol Struct Dyn. 2023;41(19):9686–9694. doi:10.1080/07391102.2022.2145370
96. Ribaudo G, Oselladore E, Ongaro A, et al. Enhanced G-quadruplex selectivity of flavonoid glycoside rutin over quercetin. Nat Prod Res. 2020;36:1–5. doi:10.1080/14786419.2020.1859505
97. Wen L-N, Xie M-X. Spectroscopic investigation of the interaction between G-quadruplex of KRAS promoter sequence and three isoquinoline alkaloids. Spectrochim Acta A Mol Biomol Spectrosc. 2017;171:287–296. doi:10.1016/j.saa.2016.08.013
98. Soundarapandian S, Alexander A, Sumohan Pillai A, et al. Differential interaction of Fluorescein-β-cyclodextrin conjugate to quadruplex kit22 DNA: inclusion of berberine and modulation of binding. J Biomol Struct Dyn. 2023;41(9):3791–3799. doi:10.1080/07391102.2022.2056508
99. Wang K-B, Liu Y, Li J, et al. Structural insight into the bulge-containing KRAS oncogene promoter G-quadruplex bound to berberine and coptisine. Nat Commun. 2022;13(1):6016. doi:10.1038/s41467-022-33761-4
100. Tan W, Zhou J, Yuan G. Electrospray ionization mass spectrometry probing of binding affinity of berbamine, a flexible cyclic alkaloid from traditional Chinese medicine, with G-quadruplex DNA. Rapid Commun Mass Spectrom. 2014;28:143–147. doi:10.1002/rcm.6763
101. Lin S, Li S, Chen Z, et al. Formation, recognition and bioactivities of a novel G-quadruplex in the STAT3 gene. Bioorg Med Chem Lett. 2011;21(19):5987–5991. doi:10.1016/j.bmcl.2011.07.121
102. Cui X, Lin S, Zhou J, Yuan G. Investigation of non-covalent interaction of natural flexible cyclic molecules with telomeric RNA G-quadruplexes by electrospray ionization mass spectrometry. Rapid Commun Mass Spectrom. 2012;26:1803–1809. doi:10.1002/rcm.6295
103. Cui X, Lin S, Yuan G. Spectroscopic probing of recognition of the G-quadruplex in c-kit promoter by small-molecule natural products. Int J Biol Macromol. 2012;50:996–1001. doi:10.1016/j.ijbiomac.2012.02.029
104. Ghosh S, Dasgupta D. Quadruplex forming promoter region of c-myc oncogene as a potential target for a telomerase inhibitory plant alkaloid, chelerythrine. Biochem Biophys Res Commun. 2015;459:75–80. doi:10.1016/j.bbrc.2015.02.072
105. Jana J, Mondal S, Bhattacharjee P, et al. Chelerythrine down regulates expression of VEGFA, BCL2 and KRAS by arresting G-Quadruplex structures at their promoter regions. Sci Rep. 2017;7(1):40706. doi:10.1038/srep40706
106. Zhou Z-L, Hu M-H. Chelerythrine may bind to KRAS mRNA G-quadruplex in the cisplatin-resistant non-small cell lung cancer cells. Bioorg Chem. 2025;163:108692. doi:10.1016/j.bioorg.2025.108692
107. Wang F, Wang C, Liu Y, et al. Colchicine selective interaction with oncogene RET G-quadruplex revealed by NMR. Chem Commun. 2020;56(14):2099–2102. doi:10.1039/D0CC00221F
108. Platella C, Ghirga F, Musumeci D, et al. Selective targeting of Cancer-Related G-Quadruplex structures by the natural compound dicentrine. Int J Mol Sci. 2023;24:4070.
109. Ji X, Sun H, Zhou H, et al. The interaction of telomeric DNA and C-myc22 G-quadruplex with 11 natural alkaloids. Nucleic Acid Ther. 2012;22(2):127–136. doi:10.1089/nat.2012.0342
110. Ruiz N, Jarosova P, Taborsky P, Gargallo R. Study of the interaction of the palmatine alkaloid with hybrid G-quadruplex/duplex and i-motif/duplex DNA structures. Biophys Chem. 2022;281:106715. doi:10.1016/j.bpc.2021.106715
111. Bhadra K, Kumar GS. Interaction of berberine, palmatine, coralyne, and sanguinarine to quadruplex DNA: a comparative spectroscopic and calorimetric study. Biochim Biophys Acta. 2011;1810:485–496. doi:10.1016/j.bbagen.2011.01.011
112. Fazelifar P, Cucchiarini A, Khoshbin Z, Mergny J-L, Kazemi Noureini S. Strong and selective interactions of palmatine with G-rich sequences in TRF2 promoter; experimental and computational studies. J Biomol Struct Dyn. 2025;43:1674–1688. doi:10.1080/07391102.2023.2292793
113. Pandya N, Kumar A. Piperine analogs arrest c-myc gene leading to downregulation of transcription for targeting cancer. Sci Rep. 2021;11:22909. doi:10.1038/s41598-021-01529-3
114. Tumir L-M, Zonjić I, Žuna K, et al. Synthesis, DNA/RNA-interaction and biological activity of benzo[k,l]xanthene lignans. Bioorg Chem. 2020;104:104190. doi:10.1016/j.bioorg.2020.104190
115. Chen M, Chen Q, Li Y, et al. A G-quadruplex nanoswitch in the SGK1 promoter regulates isoform expression by K+/Na+ balance and resveratrol binding. Biochim Biophys Acta Gen Subj. 2021;1865(2):129778. doi:10.1016/j.bbagen.2020.129778
116. Chen X, He X, Gao R, et al. Aptamer-Functionalized Binary-Drug delivery system for synergetic obesity therapy. ACS Nano. 2022;16:1036–1050.
117. Ma D-L, Chan DS-H, Fu W-C, et al. Discovery of a natural product-like c-myc G-quadruplex DNA groove-binder by molecular docking. PLoS One. 2012;7(8):e43278. doi:10.1371/journal.pone.0043278
118. Zhong L-T, Yuan J-M, Fu W-L, et al. Identification of sanguinarine as c-MYC transcription inhibitor through enhancing the G-quadruplex-NM23-H2 interactions. Bioorg Chem. 2024;153:107842. doi:10.1016/j.bioorg.2024.107842
119. Yang S, Xiang J, Yang Q, et al. Distinct G-quadruplex structures of human telomeric DNA formed by the induction of sanguinarine and nitidine under salt-deficient condition. Fitoterapia. 2010;81(8):1026–1032. doi:10.1016/j.fitote.2010.06.024
120. Białek W, et al. Plant flavonoid effects on the non-apoptotic mechanisms of regulated cell death. Phytochem Rev. 2026;25:597–641.
121. Ma Z, Lv J, Zhang S, et al. ε-Polylysine/Sodium alginate bilayer-modified nanoliposomes enhancing the stability and in vitro bioavailability of epigallocatechin gallate. Foods. 2026;15(5):818. doi:10.3390/foods15050818
122. Nurohman I, Suhandi C, Chaerunisaa A, et al. Design evolution of curcumin-loaded nanostructured lipid carriers: formulation strategies, functional modifications, and mechanistic-translational perspectives. Int J Nanomed. 2026;21:586727. doi:10.2147/IJN.S586727
123. Khan SH, Younus H, Khan MA. Polysaccharide-coated mPEG-PLA nanoparticles enhance thymoquinone delivery and therapeutic efficacy against colorectal cancer. J Drug Target. 2025;1–13. doi:10.1080/1061186X.2025.2593461
124. Mostafavi M, Hassani L, Khoshkam M. Interaction of berberine with different forms of DNA in human telomeric region. J Struct Biol. 2025;217:108175.
125. Wang X, Liu Y, Gao Z, et al. Design and synthesis of novel styrylquinolinium derivatives for the treatment of breast Cancer: targeting the c-Myc G-quadruplex. Eur J Med Chem. 2025;291:117663. doi:10.1016/j.ejmech.2025.117663
126. Endoh T, Kawasaki Y, Sugimoto N. Stability of RNA quadruplex in open reading frame determines proteolysis of human estrogen receptor α. Nucleic Acids Res. 2013;41:6222–6231.
127. Qian J, Liu R, Liu N, et al. Arene Ru(II) complexes acted as potential KRAS G-Quadruplex DNA stabilizer induced DNA damage mediated apoptosis to inhibit breast cancer progress. Molecules. 2022;27(10):3046. doi:10.3390/molecules27103046
128. Lizunova SA, Tsvetkov VB, Skvortsov DA, et al. Anticancer activity of G4-targeting phenoxazine derivatives in vitro. Biochimie. 2022;201:43–54. doi:10.1016/j.biochi.2022.07.001
129. Horchani M, Della Sala G, Caso A, et al. Molecular docking and biophysical studies for antiproliferative assessment of synthetic pyrazolo-pyrimidinones tethered with hydrazide-hydrazones. Int J Mol Sci. 2021;22(5):2742. doi:10.3390/ijms22052742
130. Morrissey KL, DeWitt D, Shah N, et al. Comparison of protein capture from a human cancer cell line by genomic G-quadruplex DNA sequences toward aptamer discovery. Anal Bioanal Chem. 2021;413(14):3775–3788. doi:10.1007/s00216-021-03328-1
131. Zeng L, Wu Q, Wang T, et al. Selective stabilization of multiple promoter G-quadruplex DNA by using 2-phenyl-1H-imidazole-based tanshinone IIA derivatives and their potential suppressing function in the metastatic breast cancer. Bioorg Chem. 2021;106:104433.
132. Balkwill GD, Derecka K, Garner TP, et al. Repression of translation of human estrogen receptor alpha by G-quadruplex formation. Biochemistry. 2009;48(48):11487–11495. doi:10.1021/bi901420k
133. Feng Y, Xuan X, Hu Y, et al. Targeting G-rich sequence to regulate the transcription of murine double minute (MDM) genes in triple-negative breast cancers. Eur J Med Chem. 2024;267:116156. doi:10.1016/j.ejmech.2024.116156
134. Da Ros S, Nicoletto G, Rigo R, et al. G-Quadruplex modulation of SP1 functional binding sites at the KIT proximal promoter. Int J Mol Sci. 2020;22(1):329. doi:10.3390/ijms22010329
135. Moraca F, Arciuolo V, Marzano S, et al. Repurposing FDA-approved drugs to target G-quadruplexes in breast cancer. Eur J Med Chem. 2025;285:117245.
136. Esposito D, Locatelli A, Morigi R. Molecular tools for precision targeting and detection of G-Quadruplex structures. Molecules. 2025;30:4099.
137. El-Far AH, Al Jaouni SK, Li W, Mousa SA. Protective roles of thymoquinone nanoformulations: potential nanonutraceuticals in human diseases. Nutrients. 2018;10(10):1369. doi:10.3390/nu10101369
138. Ashrafpour S, Ashrafpour M. The double-edged sword of nutraceuticals: comprehensive review of protective agents and their hidden risks. Front Nutr. 2025;12:1524627. doi:10.3389/fnut.2025.1524627
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Natural Source-Derived Compounds with Antifungal Activity Against Medically Relevant Fungi
Gómez-Gaviria M, Baruch-Martínez DA, Mora-Montes HM
Infection and Drug Resistance 2025, 18:6389-6406
Published Date: 6 December 2025
Recent Advances and Emerging Directions in Machine Learning-Based Breast Cancer Drug Discovery: A Comprehensive Review
Cheng Y, Kong J, Liu X, Li S
Breast Cancer: Targets and Therapy 2026, 18:586786
Published Date: 9 March 2026