Back to Journals » International Journal of Nanomedicine » Volume 20
Nanomaterial-Driven Regulated Cell Death: Mechanistic Insights and Novel Strategies for Tumor Therapy
Authors Luo Y, Lin X, Su C, Zheng Z, Li Y ![]() , Guo W, Feng X
, Guo W, Feng X
Received 4 August 2025
Accepted for publication 21 October 2025
Published 30 October 2025 Volume 2025:20 Pages 13183—13208
DOI https://doi.org/10.2147/IJN.S558103
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Lijie Grace Zhang
Yiheng Luo,1,2,* Xundong Lin,1,2,* Chuyu Su,1 Zesen Zheng,1 Yuezi Li,1 Weihong Guo,2 Xiaoli Feng1
1Stomatological Hospital, School of Stomatology, Southern Medical University, Guangzhou, 510280, People’s Republic of China; 2Department of General Surgery, Nanfang Hospital, Southern Medical University, Guangzhou, 510515, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Weihong Guo, Email [email protected] Xiaoli Feng, Email [email protected]
Abstract: The rapid development of nanomaterials (NMs) and nanotechnology has profoundly impacted the biomedical field. Due to their unique physicochemical properties, NMs can interact with biological molecules, including proteins and DNA, to induce regulated cell death (RCD). Recent studies have demonstrated that NMs trigger RCD by promoting reactive oxygen species generation, activating signaling proteins, and disrupting intracellular ion homeostasis. Among the various forms of RCD, necroptosis, pyroptosis, ferroptosis, and cuproptosis have garnered significant research attention. Increasing evidence suggests that these emerging RCD pathways are crucial for tumor proliferation and migration, tumor microenvironment (TME) remodeling, and drug resistance mitigation. Accordingly, leveraging NM-driven RCD’s effects in designing multifunctional drug delivery systems holds great promise for cancer therapy. NMs can also serve as versatile drug carriers in synergistic treatments with targeting agents, photothermal/photodynamic therapy, and immunomodulators, thereby overcoming the limitations of conventional treatments. Collectively, this review systematically summarizes recent advancements in NM-induced RCD in cancer cells and discusses the diverse roles of RCD in tumor pathology. We highlight the innovative potential of NMs in inducing RCD through precise targeting and synergistic therapy, offering novel strategies to overcome tumor resistance and reshape the immunosuppressive TME.
Keywords: nanomaterials, regulated cell death, cuproptosis, tumor drug resistance, tumor microenvironment
Introduction
Regulated cell death (RCD) refers to forms of cell death induced by activating one or more signal transduction modules.1 Currently, the primary known types of RCD include autophagic cell death, apoptosis, necroptosis, pyroptosis, ferroptosis, and cuproptosis.2 RCD may be initiated when cells fail to adapt to microenvironmental disturbances.3 In recent years, necroptosis, ferroptosis, cuproptosis, and pyroptosis have become research hotspots considering their important role in tumor proliferation and migration,2 tumor microenvironment (TME) remodeling,4 and tumor drug resistance mechanisms.5,6 For example, tumor-associated macrophages (TAMs) in the TME have been reported to transfer ceruloplasmin mRNA to human fibrosarcoma HT1080 cells via extracellular vesicles, leading to reduced intracellular iron content and lipid peroxidation, and thereby inhibiting ferroptosis.7 In addition, induction of copper metabolic dysregulation and disruption of Fe–S cluster proteins in cervical cancer cells has been shown to trigger cuproptosis, thereby overcoming cisplatin resistance.8
Nanomaterials (NMs) are materials with at least one nanometer-sized dimension (1–100 nm), with a high specific surface area and adjustable optical, electronic, and surface modification capabilities.9 By modifying the physical and chemical properties of NMs, such as their shape, structure, and adsorption capacity, researchers can design functional materials with important applications in energy, the environment, and medicine.10 Traditional cancer treatments like surgery, radiotherapy, and chemotherapy remain challenged by insufficient specificity, systemic toxicity, and drug resistance. To overcome these limitations, nanomaterial-guided therapeutic strategies have emerged as a promising approach for precise and low-toxicity interventions.11,12 As Nanomaterials can precisely target tumors and enhance drug accumulation, thereby reducing damage to healthy tissues.13,14 Moreover, by delivering immune stimulatory factors (eg, antigens, adjuvants, and immune checkpoint inhibitors), nanomaterials can directly modulate immune cells, thus enhancing anti-tumor immune responses and reducing systemic toxicity.15
Notably, a growing body of evidence indicates a close association between nanomaterials and RCD.16,17 For example, Si nanoparticles (NPs) induce ferroptosis through two mechanisms: by promoting the accumulation of iron ions, triggering the generation of reactive oxygen species (ROS) and lipid peroxidation; and by depleting glutathione (GSH) and inhibiting glutathione peroxidase-4 (GPX4) activity, which aggravate lipid peroxidation and eventually lead to cell death.18 ZnO NPs have been reported to increase the levels of necroptosis-related proteins (RIP3, MLKL) and pyroptosis-related proteins (GSDMD, caspase-1) by upregulating Z-DNA-binding protein 1 (ZBP1) expression, thereby triggering necroptosis and pyroptosis simultaneously.19 These findings suggest that nanomaterial-guided RCD represents a novel direction and strategy for tumor therapy. Indeed, a significant body of research has confirmed this. For example, biomimetic copper-doped polydopamine nanoparticles (PC NPs) can synergize with radiotherapy to inhibit tumor growth. PC NPs can release Cu2+ in acidic microenvironments to trigger ferroptosis. Moreover, when combined with radiotherapy, the key ferroptosis protein FDX1 is downregulated most significantly.20 Gao et al designed an Fe-MOF@Erastin@Herceptin nanoplatform that could target HER2-positive breast cancer through trastuzumab modification. By releasing Fe3+ and the ferroptosis inducer Erastin, lipid peroxidation and ferroptosis are triggered, resulting in precise ferroptosis therapy for HER2-positive breast cancer.21
This review systematically synthesizes the advancements in research concerning NM-induced novel RCD in tumors, with a primary focus on elucidating the key molecular mechanisms involved in the regulation of necroptosis, pyroptosis, cuproptosis, and ferroptosis. Furthermore, the article provides an in-depth analysis of the pivotal role that novel RCD plays in modulating tumor cell growth, invasion, and migration, as well as in reshaping the tumor microenvironment and counteracting tumor drug resistance. Notably, while the therapeutic strategy of integrating NMs with RCD demonstrates considerable potential, challenges persist regarding the molecular mechanisms of action, pharmacokinetics, and standardized production processes. This review aspires to furnish a theoretical foundation and novel insights for anti-tumor therapies based on NMs, facilitating their more reliable clinical translation.
Nanomaterial-Induced RCD in Tumor Cells: Mechanistic Insights
Necroptosis
Necroptosis is characterized by increased cell size, organelle swelling, and perforation of cell membranes. The most important molecular feature is the formation of necrosomes, which are complexes of receptor-interacting protease 1 (RIP1), receptor-interacting protease 3 (RIP3), and mixed lineage kinase domain-like (MLKL). Many signaling molecules, such as tumor necrosis factor (TNF), lipopolysaccharide, and interferon-gamma (IFN-γ), can induce necroptosis.22 Among them, the most widely reported mechanism is triggered when TNF binds to tumor necrosis factor receptor 1 to form tumor necrosis factor receptor type 1-associated death domain protein (TRADD), which then recruits FAS-associated death domain protein (FADD) and eventually forms RIPK3-MLKL necrosomes to induce necroptosis.23
NMs have been reported to activate various signaling molecules to induce cell necroptosis. Ag NPs induced necroptosis of human pancreatic ductal adenocarcinoma cells (PANC-1 cells) by increasing the protein levels of RIP1, RIP3, and MLKL.24 Zn NPs induced necroptosis in human breast cancer cells (MCF-7 cells) through a similar mechanism. Moreover, the expression of RIPK1, RIPK3, and MLKL was further enhanced after autophagy inhibitor 3-MA treatment.25 In another study, graphene oxychloroquine nanoconjugates increased the expression of autophagy receptor protein p62 by inducing autophagosome accumulation in human lung adenocarcinoma cells (A549 cells). In addition, the nanoconjugates recruited RIPK1 and promoted the assembly of the RIPK1-RIPK3-MLKL complex on autophagosomes to initiate necroptosis.26 These studies suggest that the level of autophagy may regulate cancer cell necroptosis. An increased level of ZBP1 is also one of the hallmarks of necroptosis. In one study, ZBP1 promoted RIPK3-mediated activation of MLKL in the nucleus, disrupting the nuclear membrane and DNA leakage into the cytoplasm.27 Ag NPs were reported to induce necroptosis in human breast cancer cells (MDA-MB-231 cells) by upregulating ZBP1 expression.28 NMs can also induce cell death by slowing the degradation of necrotizing proteins. A trastuzumab/graphene oxide complex reduced RIPK3 and RIPK1 cleavage by inhibiting the expression of caspase-8 in Osteosarcoma cells. The complex also promoted the formation of complex II containing FADD, TRADD, and RIPK1 by downregulating the cellular inhibitor of apoptosis (cIAP) protein. Together, these two pathways led to necroptosis in cells.29 Similarly, tracheal perfusion of C57 mice with 2 mg/kg Si NPs activated FLICE inhibitory protein in testicular tissue, which competitively bound to caspase-8 and inhibited its cleavage of RIPK3 and RIPK1, ultimately triggering necroptosis.30 We summarized the signaling pathways involved in the induction of necroptosis by NMs and presented them in Figure 1.
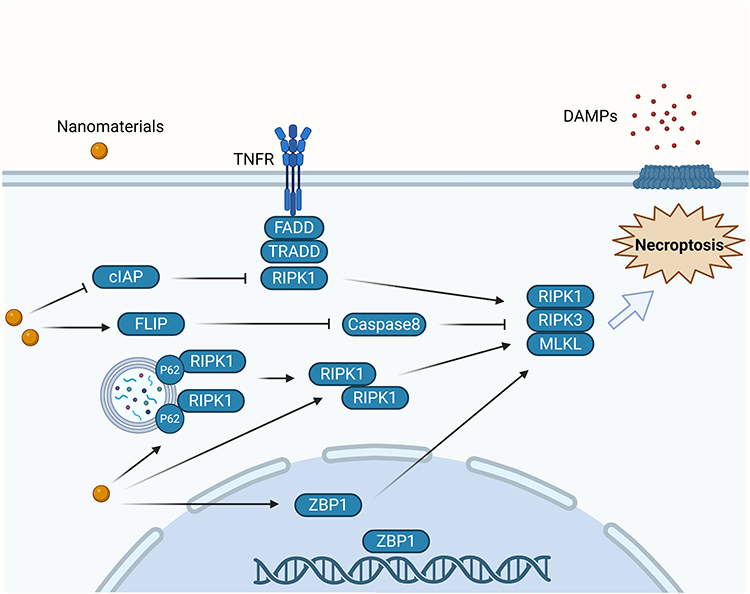 |
Figure 1 Molecular mechanism of nanomaterial-induced Necroptosis. Nanomaterials downregulate cIAPs and inhibit caspase-8, promoting necroptosis through the formation of complex II (TRADD–FADD–RIPK1) and stabilization of RIPK1/RIPK3. Additionally, nanomaterials upregulate FLICE inhibitory protein (FLIP), which interferes with caspase-8 activation and further enhances necrosome assembly. Nanomaterials also increase p62 expression and autophagosome accumulation, facilitating the recruitment of RIPK1–RIPK3–MLKL complexes to autophagosomes. In the nucleus, nanomaterials elevate ZBP1 levels, which activate RIPK3 and MLKL, ultimately leading to necroptosis. |
Pyroptosis
The classic pyroptosis pathway is mediated by inflammasomes that recognize pathogen-associated molecular patterns and host-derived danger-associated molecular patterns (DAMPs). This leads to the recruitment and activation of caspase-1, which then cleaves gasdermin-D (GSDMD) to form pores in the cell membrane, causing the cell to swell and leak its cellular contents.31 An atypical pathway involves caspases-11/4/5, which directly cleave GSDMD in response to lipopolysaccharide and trigger pyroptosis.32 Studies have increasingly suggested that NMs and NP-associated molecular patterns act as novel danger signals of the NOD-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome.33,34 A novel drug delivery system using the homing effect of cancer cell membranes to deliver the composite drug Ca@GOx to mitochondria induced mitochondrial Ca2+ overload and generated ROS, thus triggering pyroptosis in tumor cells.35 Other caspases can also initiate pyroptosis. In cancer cells with high expression of gasdermin-E (GSDME), such as SH-SY5Y neuroblastoma cells and MeWo melanoma cells, chemotherapeutic agents such as cisplatin and paclitaxel activate caspase-3 to cleave GSDME, releasing the N-terminal domain, which forms pores in the membrane.36 In melanoma cells, iron ions act as redox enzyme sensitizers, enhancing ROS signaling initiated by chemotherapeutic agents such as sulfasalazine. This promotes oxidation of the mitochondrial outer membrane protein Tom20, which then releases cytochrome c and activates caspase-3 to cleave GSDME, inducing pyroptosis.37 GSDME levels largely constrain the efficiency of caspase-3-mediated pyroptosis. A biomimetic NP loaded with decitabine increased the cytochrome c content in the cytoplasm after internalization in the cell, thereby activating caspase-3. Decitabine inhibited DNA methylation, upregulated GSDME expression, enhanced caspase-3 cleavage of GSDME, and promoted pyroptosis in cancer cells.38 We summarized the signaling pathways involved in the induction of pyroptosis by NMs and presented them in Figure 2.
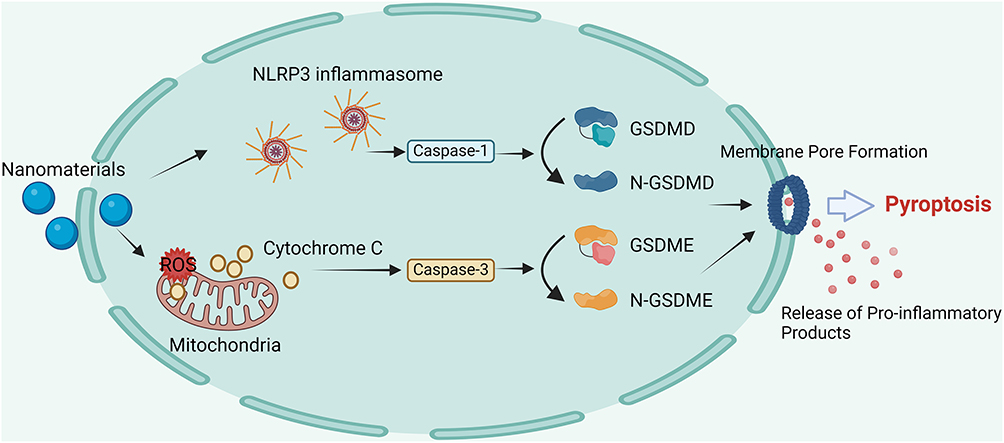 |
Figure 2 Molecular mechanism of nanomaterial-induced pyroptosis. Nanomaterials activate the NLRP3 inflammasome, leading to caspase-1 activation and cleavage of GSDMD into N-GSDMD, which forms membrane pores and promotes the release of pro-inflammatory products. Additionally, nanomaterials induce mitochondrial ROS accumulation and damage, triggering Cytochrome C release and caspase-3 activation, which cleaves GSDME into N-GSDME. These processes collectively result in pyroptosis through membrane pore formation and the release of pro-inflammatory products. |
Ferroptosis
The morphological characteristics of the mitochondria are important indicators by which ferroptosis is distinguished from other forms of RCD. Electron microscopy revealed mitochondrial shrinkage, degradation, and loss of cristae, as well as increased membrane density after ferroptosis was induced in cells. The accumulation of ROS and lipid peroxides (LPOs) is widely considered the primary cause of ferroptosis. NMs generate toxic hydroxyl radicals (•OH) via Fenton reactions involving iron ions, which produce high levels of ROS that cause lipid peroxidation of the cell membrane and ultimately induce ferroptosis.39 For example, body-centered cubic Fe NPs efficiently catalyzed Fenton reactions under acidic conditions (pH 5.5), generated ROS, and induced LPO accumulation in HepG2 liver cancer cells, leading to ferroptosis (shown in Figure 3).40 GSH reduces toxic lipid peroxides (L-OOH) to alcohols (L-OH). Thus, inhibiting this process is an effective means of inducing ferroptosis.41 Zhang et al encapsulated imidazole ketone erastin (a system Xc− inhibitor) in polyethylene glycol NPs to reduce GSH synthesis by inhibiting the amino acid antiporter system Xc− and successfully induced ferroptosis.42 Similarly, Gao et al used nanopolymer micelles to deliver RSL3 (a GPX4 inhibitor) to cancer cells. This reduced the activity of GPX4, weakened the GSH reduction reaction, and induced ferroptosis.43
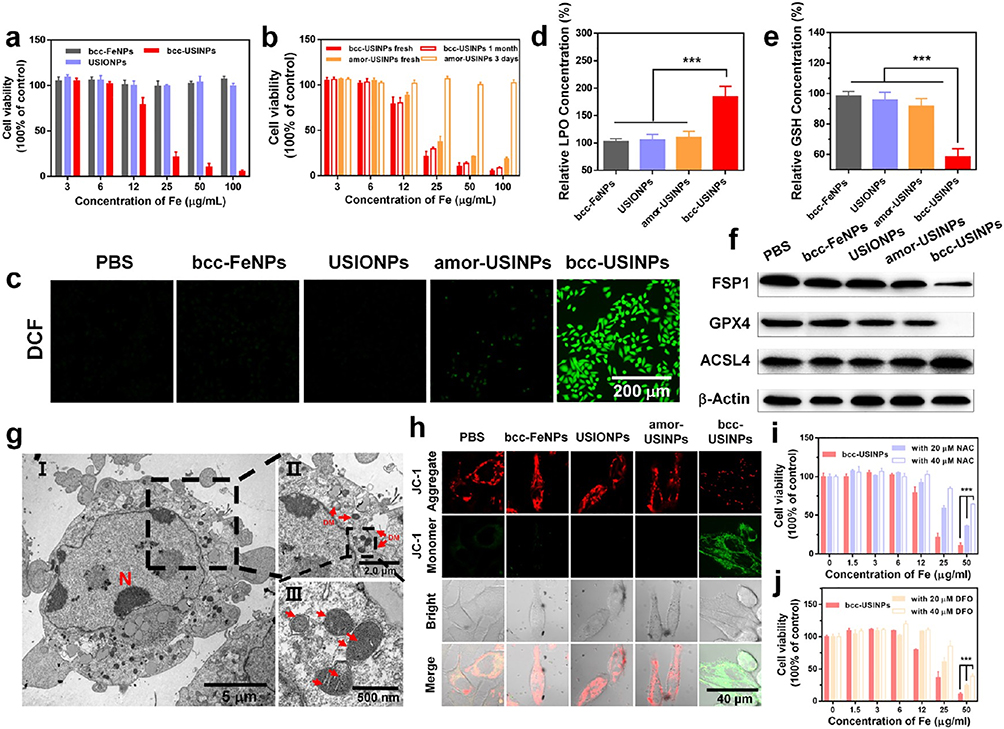 |
Figure 3 In vitro assessment of the cytotoxicity and ferroptosis of bcc-USINPs. (a) Relative cell viability of HepG2 cells after being incubated with 15 nm bcc-Fe NPs, USIONPs, and bcc-USINPs for 24 h. (b) Comparative cytotoxicity of fresh prepared bcc-USINPs, 1-month air-exposed bcc-USINPs, fresh prepared amor-USINPs, and 3-day air-exposed amor-USINPs for HepG2 cells after 24 h incubation. (c) Fluorescence microscope images of HepG2 cells after incubation with PBS, bcc-Fe NPs, USIONPs, amor-USINPs, and bcc-USINPs at a concentration of 20 μg/mL for 4 h and subsequently stained with ROS fluorescence probe DCFH-DA. Scale bar: 200 μm. (d and e) Relative (d) LPO content and (e) GSH content of HepG2 cells treated with different nanoparticles (20 μg/mL). (f) Expression of FSP1, GPX4, and ACSL4 in HepG2 cells after being incubated with bcc-Fe NPs, USIONPs, amor-USINPs, or bcc-USINPs at a concentration of 20 μg/mL for 12 h. (g) Bio-TEM image of the cross section of HepG2 cell (I) and its local view (II, III) after being incubated with bcc-USINPs for 4 h. N indicates nucleus, DM indicates damaged mitochondrion. (h) CLSM images of HepG2 cells after incubation with different nanoparticles and stained with JC-1. Green fluorescence was emitted by cytosolic JC-1 monomers, which indicates Δψm dissipation. The JC-1 aggregates in healthy mitochondria emitted red fluorescence. Scale bar: 40 μm. (i and j) Relative cell viability of HepG2 cells after being incubated with (i) bcc-USINPs + NAC and (j) bcc-USINPs + DFO. ***P < 0.001 indicates statistical significance.40 Copyright © 2021, American Chemical Society. |
Cuproptosis
The morphological features of cuproptosis include cell membrane rupture, mitochondrial shrinkage, and chromatin fragmentation. Ferredoxin 1 (FDX1), lipoic acid synthase (LIAS), and dihydrolipoamide S-acetyltransferase (DLAT) are key proteins involved in cuproptosis. FDX1 is a crucial upstream regulator of cuproptosis. FDX1 reduces Cu2+ to Cu+ and regulates protein lipoylation. Lipoic acid, synthesized by LIAS, is a necessary substrate for protein lipoylation. After Cu+ accumulates within a cell, it binds to lipoylated proteins (particularly DLAT) and induces their oligomerization, which also lowers the levels of Fe-S cluster proteins in the mitochondria. Together, these effects lead to protein toxicity stress and ultimately cuproptosis.44 Su et al discovered that CuO NPs increased Cu2+ levels in HTR8/SVneo human trophoblast cells, leading to DLAT oligomerization that triggered cuproptosis.45 Moreover, CuO NPs were shown to activate the p53 pathway and deplete GSH, causing apoptosis and ferroptosis. Further research has indicated that GSH depletion is a common regulatory mechanism for both ferroptosis and cuproptosis.46 Supplementing tumor cells with GSH can simultaneously inhibit ferroptosis and cuproptosis through the following mechanisms: (1) as an antioxidant, GSH prevents LPO, and (2) as a copper chaperone, GSH binds to copper, reducing the aggregation of lipoylated proteins.47 Additionally, p53 regulates cuproptosis by upregulating the expression of ferredoxin reductase and influencing electron transfer in the mitochondrial respiratory chain, thereby affecting two key steps in the cuproptosis pathway, Fe-S clustering and GSH biosynthesis.48 These findings indicate crosstalk between different programmed cell death mechanisms, and regulating a single signaling molecule can affect multiple cell death pathways. The mechanism of ferroptosis and cuproptosis caused by NMs is demonstrated in Figure 4. A detailed summary of NM-induced RCD, including NMs’ characteristics, experimental conditions, and molecular mechanisms, is provided in Table 1.
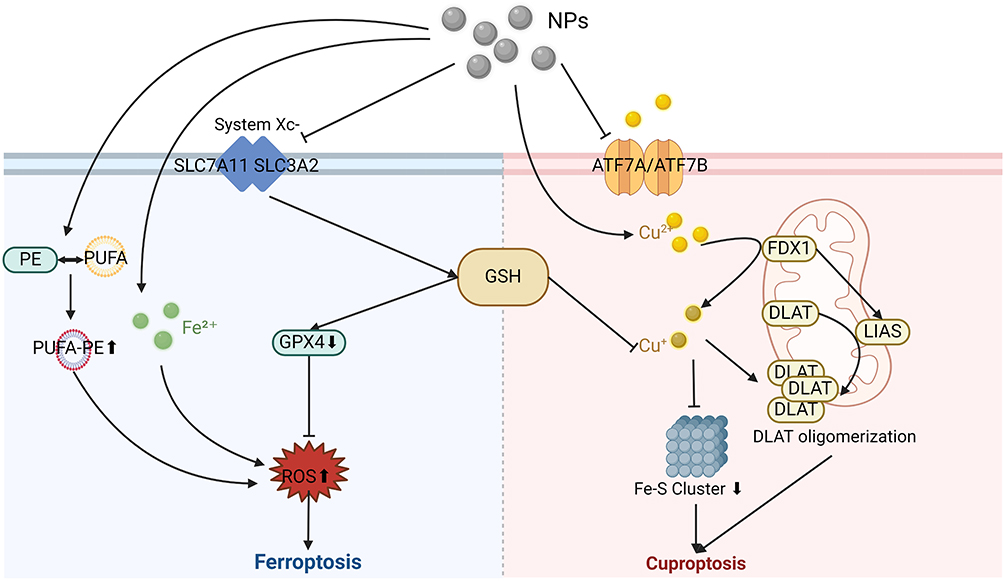 |
Figure 4 Molecular mechanism of nanomaterial-induced ferroptosis and cuproptosis. Nanomaterials deliver Fe2+ into the cell, which generates toxic •OH via the Fenton reaction, producing large amounts of ROS and causing Ferroptosis. By inhibiting system Xc− conveying cystine into cells, nanomaterials also down-regulate GSH and GPX4 levels to generate ROS; this result can also be achieved by inducing the binding of PUFA to PE. On the other side, nanomaterials can deliver excess Cu2+ to the cell, which is converted by FDX to the more toxic Cu+. This ultimately leads to Cuproptosis by promoting DLAT oligomerization and reducing the synthesis of the Fe-S Cluster. During this process, Cu2+ overload can be facilitated by inhibiting the Cu2+ transporter ATPases ATP7A and ATP7B or down-regulating GSH level. |
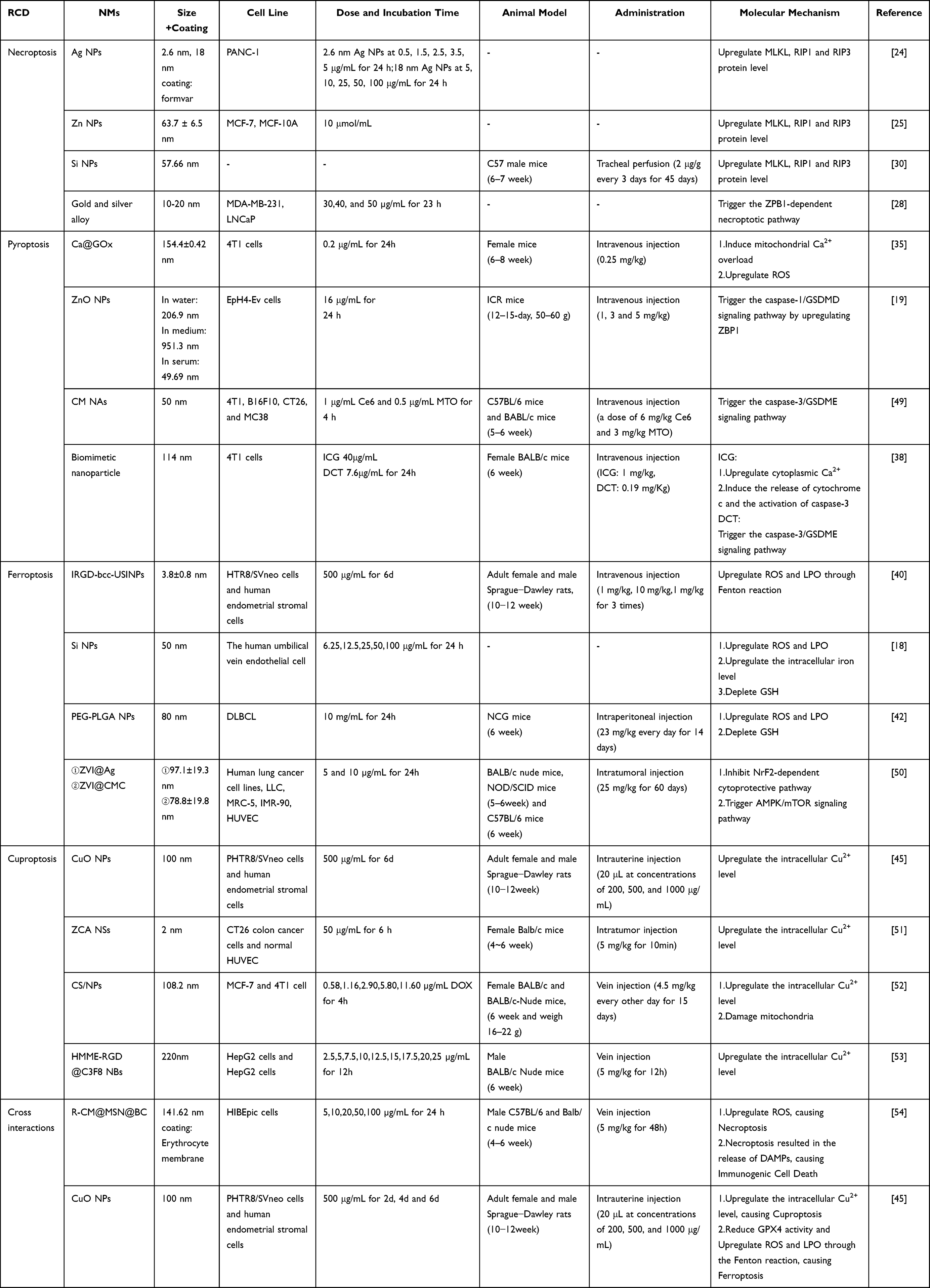 |
Table 1 Nanomaterial-Induced Regulated Cell Death Under in vitro and in vivo Conditions |
Integrating RCD Into Tumor Biology
Tumor Cell Proliferation, Invasion, and Metastasis
RIPK1-RIPK3-MLKL Pathway
Necroptosis is closely associated with malignant tumor phenotypes. Necroptosis defects and the low expression of related molecules often contribute to tumor initiation and progression.55 In colorectal cancer (CRC) cells, tumor necrosis factor receptor-associated factor 6 promoted the polyubiquitination and subsequent degradation of RIPK1, consequently suppressing the assembly of the RIPK1-RIPK3-MLKL necroptotic complex and facilitating tumor cell proliferation.56 Erigeron breviscapus enhanced the phosphorylation of RIPK3 and MLKL in CRC, leading to ROS generation and necrosome formation, which significantly suppressed tumor proliferation.57 RIP3 expression levels were shown to determine CRC tumor cell sensitivity to necroptosis.58 Studies have confirmed that abnormal DNA methylation leads to RIP3 deficiency and necroptosis defects in many tumor cell types, thereby enhancing malignant proliferation and tumor cell survival.55 Interestingly, Vucur et al reported that chronic activation of necroptosis in hepatocytes promoted hepatocarcinogenesis in mice, potentially by increasing the levels of cytokines and chemokines released by cells.59 Liu et al summarized the role of necroptosis in tumors and proposed that low-level, chronic necroptosis enhanced the infiltration of immunosuppressive cells in the tumor microenvironment by continuously releasing immunosuppressive molecules such as CXCL1, thereby promoting tumor growth. In contrast, high levels of necroptosis induced the release of immunostimulatory cytokines, which exerted anti-tumor effects.60 Tumor necrosis is often associated with tumor invasiveness and metastasis, and necroptosis cells are frequently found near necrotic areas in solid tumors.61 In one study, compared with pancreatic ductal adenocarcinoma (PDAC) patients without metastasis, those with early liver metastasis exhibited increased MLKL expression and enhanced necroptotic signaling. Specifically, MLKL-driven necroptosis recruited macrophages, strengthened the tumor’s CD47 “don’t eat me” signal, and induced macrophages to form extracellular traps, which facilitated the epithelial-mesenchymal transition (EMT) and extracellular matrix degradation and ultimately promoted PDAC liver metastasis.62 Additionally, tumor cells can express amyloid precursor protein, which was shown to interact with death receptor 6 to induce necroptosis in endothelial cells, creating channels that allowed tumor cells to extravasate and migrate.63
NLRP3/GSDMD Pathway
The NLRP3 inflammasome is a key molecule that recognizes pyroptosis signals, and it has a double-edged role in tumor cell invasion, proliferation, and metastasis.64 In hepatocellular carcinoma (HCC), significant downregulation or loss of NLRP3 expression promoted tumor progression.65 He et al also reported that the virulence factor CagA produced by Helicobacter pylori enhanced gastric cancer cell invasion and migration by activating the NLRP3 inflammasome and inducing ROS production.66 However, in some instances, NLRP3 activation instead promotes cancer growth. IL-1β secretion induced by NLRP3 activation was shown to be crucial in the proliferation, invasion, and migration of breast cancer cells and gastric cancer cells.67,68 In an animal study, tumor growth and metastatic capacity were lower in NLRP3-knockout mice injected with breast tumor cells (PyT8) than in wild-type mice.69 Another group of pyroptosis effector proteins, the gasdermin family, has also gained attention for its role in tumor progression.70 Xu et al found that higher GSDMD expression in non-small cell lung cancer was closely associated with increased tumor size and accelerated late-stage metastasis.71 The EGFR/Akt signaling pathway is involved in tumor cell proliferation, migration, and angiogenesis.72 In non-small cell lung cancer, GSDMD downregulation inhibited tumor proliferation by promoting apoptosis and suppressing EGFR/Akt signaling.73 In contrast, in both in vitro and in vivo models of gastric cancer, GSDMD downregulation activated the STAT3 and PI3K/PKB signaling pathways, which facilitated the S/G2 phase transition and significantly promoted tumor proliferation.74 Similarly, high expression of GSDMB and GSDME was shown to promote the proliferation and invasion of breast cancer cells, bladder cancer cells, and colorectal cancer cells.75,76
SLC Family Signaling
A growing body of evidence suggests that activating ferroptosis suppresses tumor cell proliferation. The expression of tumor suppressor genes such as P53 and BRCA1-associated protein 1 induces ferroptosis by inhibiting SLC7A11 (the light chain of system Xc−) transcription and by deubiquitination, respectively, thereby restraining tumor cell proliferation.77,78 Consequently, tumors have developed mechanisms for evading ferroptosis, including reducing ROS levels and decreasing lipid peroxidation, which then promotes proliferation.79–81 SLC7A11 and SLC3A2 form system Xc−, a crucial signaling complex in the ferroptosis pathway, which primarily regulates ferroptosis by modulating glutamate-cysteine exchange. Upregulated integrin ITGB3 increased SLC7A11 expression in platelet-derived extracellular vesicles from nasopharyngeal carcinoma patients, inhibiting ferroptosis and promoting invasion and distant metastasis of nasopharyngeal carcinoma cells, whereas SLC7A11 knockdown significantly reduced nasopharyngeal carcinoma cell adhesion, colony formation, and migration.82 In gastric cancer, ubiquitin-mediated degradation of stearoyl-CoA desaturase, a lipid metabolism enzyme, increased lipid peroxidation, induced ferroptosis, and suppressed tumor migration.83 The EMT is a key process in cancer metastasis that enhances tumor cell motility and invasiveness by downregulating epithelial genes and upregulating mesenchymal genes.84 There are multiple interactions between ferroptosis-related molecular signals and the EMT.85 For example, in pancreatic ductal adenocarcinoma, SMAD4 increased ferroptosis susceptibility by downregulating GPX4 expression and also promoted cancer cell invasion and migration by facilitating the EMT through the TGF-β1 signaling pathway.86 Additionally, Bi et al reported that metadherin increased ferroptosis susceptibility by inhibiting GPX4 and SLC3A2 and by positively regulating the EMT, thereby promoting invasion and migration in triple-negative breast cancer (TNBC) and endometrial cancer.87
FDX1/DLAT Signaling
Several cuproptosis-related genes (CRGs) have been shown to be closely associated with tumor progression and metastasis. For example, FDX1, a key gene regulating cuproptosis, suppressed colorectal cancer growth and invasion by delaying the EMT progression.88 Wang et al also demonstrated that FDX1 overexpression reduced HCC proliferation, and in vivo experiments showed a decrease in pulmonary metastases.89 Moreover, FDX1 downregulation activated the PI3K/AKT signaling pathway, inducing mitophagy of damaged mitochondria to maintain metabolic stability, and ultimately promoted HCC progression. DLAT is an upstream regulator of cuproptosis. Peng et al found that silencing DLAT caused gastric cancer cells to arrest in the G0/G1 phase, effectively inhibiting their proliferation.90 Notably, increasingly more research has focused on the association between CRGs and tumor development, suggesting that targeting CRGs such as LIAS, CTR1, ATP7A, and ATP7B using gene therapy holds promise for cancer treatment.91
Tumor Microenvironment
The TME is a multifaceted ecosystem comprising tumor cells, immune cells, stromal cells, and other components. These elements interact in a complex manner to collectively influence tumor growth and progression. Immune cells are critical in the TME. CD8+ T cells and regulatory T cells primarily exhibit anti-tumor effects, while myeloid-derived suppressor cells (MDSCs) exert immunosuppressive functions, promoting tumor development. Additionally, CD4+ T cells, B cells, neutrophils, macrophages, and monocytes often play dual roles in tumor progression and immunity.92
Dual Role of RIPK3/RIP3
Necroptosis leads to the release of intracellular contents, generating DAMPs that alter cell communication, cell fate decisions, and immune responses in the TME.93,94 The level of the necroptosis-related molecule RIPK3 is closely associated with anti-tumor immunity. In HT-29 colorectal cancer cells and SNU-620 gastric cancer cells, necroptosis led to upregulated RIPK3 expression, which activated the NF-κB signaling pathway and promoted the secretion of inflammatory cytokines such as IL-1β and CCL4. This mechanism enhanced the activity of dendritic cells (DCs) and CD8+ T cells in the TME and ultimately promoted tumor antigen presentation and tumor-killing effects.95 Moreover, RIPK3 was shown to promote IFN-β release, which facilitated CD4+ T cell activation and infiltration into tumor sites, further shaping an anti-tumor TME.96 Conversely, RIPK3 downregulation activated the PPAR pathway, promoting the polarization of TAMs toward the M2 phenotype and ultimately fostering tumor growth.97 Notably, the role of necroptosis signaling molecules in the TME is more complex than previously thought. In PDAC and colitis-associated colorectal cancer, high RIP3 expression activated CXCL1 signaling, which recruited MDSCs and TAMs for a pro-tumorigenic TME.98,99
Immune Cells and Signals of Ferroptosis
Immune cells in the TME regulate tumor ferroptosis by releasing various signaling molecules.100 For example, activated CD8+ T cells release IFN-γ, which downregulates SLC3A2 and SLC7A11 expression in system Xc−; this reduces cystine uptake in tumor cells and ultimately promotes lipid peroxidation and ferroptosis.101 In contrast, TAMs in the TME inhibited ferroptosis by transferring ceruloplasmin mRNA via extracellular vesicles to human fibrosarcoma HT1080 cells, which reduced intracellular iron levels and lipid peroxidation.7 Ferroptosis also occurs in immune cells, which affects tumor progression. Ma et al found that CD8+ T cells upregulated CD36 expression, increasing arachidonic acid uptake, which in turn triggered ferroptosis and reduced the number of CD8+ T cells in the TME, ultimately promoting tumor progression.102 Similarly, ferroptosis in MDSCs led to the production of prostaglandin E2 and oxidized arachidonic acid-phosphatidylethanolamine, thereby suppressing CD8+ T cells and promoting a tumor-supportive TME.103 Moreover, cancer-associated fibroblasts (CAFs) in the TME could release exosomes containing lncRNA FTX, which, upon being taken up by oral squamous cell carcinoma (OSCC) cells, led to the demethylation of the flap endonuclease-1 promoter, thereby inhibiting ferroptosis and facilitating tumor invasion and migration.104 Interestingly, ferroptosis also activates anti-tumor immunity. In TAMs, APOC1 downregulation increased intracellular iron levels and ROS, which induced ferroptosis and subsequently promoted the polarization of other TAMs into the M1 phenotype, thereby inhibiting hepatocellular carcinoma growth.105
Immune Cells and Signals of Pyroptosis
Pyroptotic tumor cells release tumor-associated antigens,106 which activate anti-tumor immune responses. Some pyroptosis-derived factors also act as DAMPs,107,108 which either promote inflammation or suppress immune responses. For example, IL-1β and TNF-α released from pyroptotic cells induced IL-8 production in an NF-κB-dependent manner, recruiting MDSCs to the TME and suppressing tumor immunity.109 Blocking IL-1β converted an immunosuppressive TME into an anti-tumor immune environment by increasing CD8+ T-cell infiltration and inhibiting neutrophils and MDSCs.110 The gasdermin family is also important in tumor immunity.34,111 It was found that IFN-γ in the TME induced GSDMB expression in tumor cells; granzyme A produced by NK (Natural Killer) cells and cytotoxic T lymphocytes killed the GSDMB-positive tumor cells via the cell pyroptosis pathway.112 In addition, granzyme B secreted by NK cells was shown to induce tumor cell pyroptosis and enhance anti-tumor immunity by directly cleaving GSDME.113 TAMs are abundant in the TME, and because they primarily exhibit an immunosuppressive M2 phenotype, the TME is usually in an immunosuppressed state. However, inflammatory factors released by pyroptotic tumor cells were shown to promote the release of INF-γ from NK cells and T cells, which in turn promoted the polarization of TAMs toward the immunoreactive M1 phenotype.114 TAMs also increased the number of CD8+ T cells, CD4+ T cells, and NK cells in the TME, while significantly decreasing the number of neutrophils, MDSCs, and monocytes.115 These changes in immune cell populations ultimately remodel the immunosuppressive TME into an immunogenic TME with anti-tumor effects.116,117
Interactions Between CRGs, Immune Cells, or CAFs
Cuproptosis influences immune cell infiltration into the TME. In hepatocellular carcinoma, increased expression of CRGs such as DLAT, GLS, and CDKN2A correlated with the activation of the TGF-β-mediated immunosuppressive pathway, whereas their downregulation was associated with increased numbers of T memory and macrophage FCN1+ cells, suggesting that downregulation of these genes enhanced anti-tumor immunity.118 Similarly, in bladder cancer, high cuproptosis activity correlated with reduced immune cell infiltration, including decreased levels of CD4+ T cells, CD8+ T cells, DCs, and NK cells.119 Notably, different cuproptosis signals generated the opposite immune response. For example, in colorectal cancer, upregulation of the CRG DLAT increased the number of CD8+ GZMK+ T cells, activating anti-tumor immunity. However, COX17 expression promoted tumor growth by increasing the level of exhausted CD8+ PDCD1+ T cells and generating immunosuppressive CD4+ CXCL13+ T cells. Studies on non-immune cells in the TME have focused on cancer-associated fibroblasts (CAFs), which are implicated in angiogenesis, tumor metabolism, and connective tissue proliferation and contribute to cancer immune evasion.120 In studies of colon adenocarcinomas, high expression of FDX1 inhibited tumor growth and metastasis, which was closely related to the low level of infiltrating CAFs.121 In contrast, Li et al found that extensive proliferation of CAFs in oral squamous cell carcinoma significantly increased the expression of the CRGs GLS and MTF1, which in turn affected tumor cell copper homeostasis.122 Moreover, CAFs released exosomes containing downregulated miR-148b-3p, resulting in the upregulation of ATP7A expression in OSCC, ultimately inhibiting cuproptosis and promoting the malignant phenotype of the tumor.123
Treatment Resistance
Although drug therapy has made significant progress in slowing tumor development, progression, and metastasis, cancer drug resistance seriously reduces the effectiveness of drug therapy.4 It has been demonstrated that novel types of RCD and their molecular signaling influence tumor drug resistance through both direct and indirect effects. Here, we examine this finding with respect to chemotherapy and immunotherapy.
Chemotherapy Resistance
One mechanism by which tamoxifen kills breast cancer cells is ferroptosis. Downregulation of the long non-coding RNA LINC00152 inhibited the expression of the ferroptosis signals GPX4 and SLC7A11, increased intracellular iron ions, and reversed tamoxifen resistance in breast cancer.124 Similarly, reduced androgen receptor expression was found to inhibit GPX4 expression, increasing the intracellular redox imbalance and causing ferroptosis, which increased the sensitivity of glioblastoma to temozolomide.125 Cuproptosis was found to alter prostate cancer resistance to docetaxel. Wen et al induced DLAT expression in prostate cancer, which led to mTOR phosphorylation and inhibition of cellular autophagy; this, in turn, caused cancer cells to remain in the G2/M phase, which enhanced the killing effect of docetaxel on tumor cells and effectively reversed tumor resistance.126 Under specific circumstances, RCD may also promote tumor resistance. For example, inflammatory factors secreted by ovarian cancer cells, such as IL-6 and IL-8, induced pyroptosis of omental adipocytes. The free fatty acids released during pyroptosis of omental adipocytes were then taken up by ovarian cancer cells to activate the STAT3/CPT1B pathway, which increased phospholipid biosynthesis and the mitochondrial membrane potential, thus resisting apoptosis induced by chemotherapeutic agents.127 Under hypoxic conditions, HSP90α inhibited MLKL activation by binding to necrosomes. This attenuated necroptosis was triggered by sorafenib, which ultimately led to the emergence of tumor resistance.128 Notably, RCD also acts on other TME components to indirectly affect tumor resistance. For example, cisplatin and paclitaxel were shown to promote the secretion of miR-522 by CAFs. MiR-522 entered gastric cancer cells and reduced the accumulation of lipid peroxides and ferroptosis by inhibiting the expression of ALOX15, enhancing acquired chemoresistance in gastric cancer cells.129
Immunotherapy Resistance
Increasingly more studies are focusing on the effects of RCD on tumor immune resistance. Ferroptosis is an important mechanism by which CD8+ T cells achieve anti-tumor effects. CD8+ T cells inhibited the expression of system Xc− in tumor cells by secreting IFN-γ, reducing the ability of the cells to scavenge lipid peroxides; IFN-γ then upregulated the expression of intracellular ACSL4, which accelerated the formation of lipid peroxidation, further promoting ferroptosis.130 Carnitine palmitoyl transferase 1A is a key enzyme in lipid metabolism and forms a positive feedback loop with the transcription factor c-Myc, which was shown to activate the NRF2/GPX4 antioxidant system and enhance the tolerance of cancer stem cells to ferroptosis. Because this tolerance impaired the killing effect of ferroptosis induced by CD8+ T cells, the efficacy of immunotherapy was reduced.131 Similarly, overexpression of the tyrosine kinase receptor TYRO3 in TNBC inhibited CD8+ T cell-induced ferroptosis. At the same time, TYRO3 overexpression also upregulated the vascular endothelial growth factor signaling pathway and promoted the polarization of macrophages from the M1-type to the M2-type, further inhibiting anti-tumor immune responses.132 In contrast, activation of necroptosis and cuproptosis reverses tumor immune resistance by promoting infiltration of CD8+ T cells. In the TME, ZBP1-dependent fibroblast necroptosis promoted CD8+ T cell recruitment, which enhanced anti-tumor immunity and, therefore, is a potential target for reversing drug resistance.133 Microsatellite-stable colon cancer has been considered resistant to immunotherapy. However, recent studies have reported that inducing cuproptosis in Microsatellite-stable colon cancer inhibited WNT signaling and thus downregulated the expression of the immunosuppressive receptor PD-L1 by tumor cells, enhancing immune attack by CD8+ T cells and effectively slowing tumor progression.134
Anti-Tumor Design Strategies Using Nanomaterials
Due to the intrinsic or acquired resistance of cancer cells, conventional anti-tumor drugs often yield unsatisfactory clinical outcomes. The introduction of NMs has opened new possibilities for improving these outcomes. Because of their unique physicochemical properties and biological behaviors, NMs can directly induce RCD in cancer cells. Moreover, as drug carriers or conjugated to tumor-targeting molecules, they can further enhance therapeutic efficacy.135
Suppressing Tumor Proliferation Combined with Photothermal or Photodynamic Therapy
NMs have shown great potential for inhibiting tumor cell proliferation and invasion. For example, researchers developed a disulfide-crosslinked polypeptide nanogel system, STP-NG/SHK, targeting the VIM protein on a tumor surface, which significantly enhanced tumor specificity. The system induced RIP1- and RIP3-dependent necroptosis, effectively suppressing osteosarcoma cell proliferation and reducing pulmonary metastasis.136 Ferroptosis has also demonstrated strong tumor-suppressive effects. Li et al designed a nanoplatform, MPDA/Fe/RSL3@M-gy1, carrying both iron ions and the GPX4 inhibitor RSL3, and coated it with macrophage membranes to enhance targeting.137 The system significantly induced ferroptosis in castration-resistant prostate cancer cells, which effectively suppressed their proliferation under low androgen conditions. Similarly, diethyl dithiocarbamate-ferrous oxide NPs increased intracellular ROS levels and inhibited the GSH-GPX4 antioxidant system, triggering ferroptosis in cisplatin-resistant A549 cancer cells, thereby successfully suppressing cancer cell proliferation.138 Research on pyroptosis has further expanded the scope of therapeutic strategies for cancer. Elisa et al developed the CXCR4-targeted self-assembling nanomicelles T22-PE24-H6 and T22-DITOX-H6 that activated the caspase-3/GSDME-dependent pyroptotic pathway, thereby inhibiting head and neck squamous cell carcinoma proliferation while minimizing systemic toxicity due to their high targeting efficiency.139 In another study, a self-assembled nanocomposite system comprising the photosensitizer chlorin e6 and the photochemical drug mitoxantrone induced pyroptosis through combined photothermal and photodynamic therapy, which effectively suppressed tumor growth while avoiding damage to normal tissues.49 In recent years, nanotherapeutic strategies targeting cuproptosis have advanced rapidly. To overcome the suppression of sonodynamic therapy (SDT) efficacy by GSH in the TME, researchers combined HMME-RGD@C3F8 NBs, nanobubbles loaded with the sonosensitizer HMME and the tumor-targeting peptide RGD, with the cuproptosis inducer Elesclomol-Cu. This approach not only directly induced cuproptosis in tumor cells but also enhanced SDT efficacy by increasing Cu²+ consumption to deplete GSH.53 All these NMs leveraged precise targeting and activation of specific RCD pathways to suppress tumor cell proliferation and invasion, suggesting high clinical potential.
Activating Anti-Tumor Immune Responses by Releasing DAMPs
NMs modulate different aspects of the immune microenvironment through multiple mechanisms, including the release of DAMPs, polarization of immune cells, and activation of CD8+ T cells. These mechanisms collectively establish a multilayered immune regulatory network, providing various approaches for tumor therapy. For example, an R-CM@MSN@BC nanocomposite induced necroptosis in cholangiocarcinoma cells, releasing DAMPs that enhanced antigen presentation, which promoted M1 macrophage infiltration and reduced the activity of immunosuppressive cells, ultimately boosting anti-tumor immune responses.54 Similarly, metabolism-aiming Cu2-xS nanoagents induced ICD in 4T1 cells via cuproptosis, releasing DAMPs that promoted the conversion of M2 macrophages to the M1 phenotype, significantly reducing the tumor size and inhibiting metastasis.140 Another research focus is antigen uptake enhancement to promote DC maturation and activate anti-tumor immunity. The CaO2-core-driven nanoplatform CTFAP released oxygen synergistically with sonodynamic therapy and Fenton reactions to induce ferroptosis, which significantly enhanced DC antigen uptake and strengthened anti-tumor immunity.141 Similarly, Zhong et al developed the nanoliposome GM@LR that co-delivered a GSDME-expressing plasmid and manganese carbonate into TNBC cells.142 The treatment upregulated GSDME expression while increasing DC infiltration and antigen uptake within tumors. Recent studies highlighted interactions between CD8+ T cells and novel RCD pathways.130 For example, zero-valent iron NPs inhibited NRF2-mediated cytoprotective mechanisms and activated the AMPK/mTOR signaling pathway to induce ferroptosis, significantly enhancing CD8+ T cell cytotoxicity and promoting M1 macrophage polarization for dual modulation of the tumor immune microenvironment.50 Similarly, Cu-substituted ZnAl ternary layered double hydroxide nanosheets in combination with sonodynamic therapy induced cuproptosis by activating ICD, promoting DC maturation, and enhancing CD8+ T cell activity, thereby reversing immune suppression and inhibiting tumor growth and metastasis.51 The detailed mechanisms are illustrated in Figure 5.
 |
Figure 5 Activation of Anti-tumor Immune Responses by Release of DAMPs in Nano-Integrated Therapy. Nano-integrated therapy inhibits glutamine metabolism to reverse T cell exhaustion and restore anti-tumor function. It also induces necroptosis, leading to the release of HMGB1 and calreticulin (CRT), which promotes macrophage polarization toward the M1 phenotype. Cuproptosis-based and ferroptosis-based strategies trigger immunogenic cell death, accompanied by the release of HMGB1, CRT, and ATP, thereby enhancing antigen uptake and presentation. Moreover, nanomaterials upregulate the cGAS–STING signaling pathway through cGAMP accumulation, promoting DC maturation and CD8+ T cell activation; pyroptosis can further amplify anti-tumor immunity by facilitating DAMPs release and remodeling the immune microenvironment. |
Overcoming Tumor Drug Resistance by Modulating Key Signals
Chemoresistance is a major challenge in conventional cancer treatment. The use of NMs can improve drug delivery efficiency and alter tumor cell responses to anticancer drugs to reverse chemoresistance. For example, a near-infrared-activated photothermal therapy system combining fluorescent carbon nanodots with irinotecan successfully overcame breast cancer apoptosis resistance. The CD-PEG-BT/IT complex, upon near-infrared activation, locally released heat and drugs, which induced necroptosis through the TNF-α and RIPK1 pathways, upregulated pro-apoptotic genes and downregulated anti-apoptotic genes, thus promoting apoptosis.143 Both cuproptosis and ferroptosis have shown promise in overcoming resistance. Tsai et al designed iron-platinum alloy NPs that generated high levels of ROS via the Fenton reaction, inducing cell death and inhibiting the proliferation and migration of tyrosine kinase inhibitor-resistant lung cancer cells.144 PAB@MIL-53 NPs enhanced drug efficacy by increasing ROS production, depleting GSH, and reducing P-gp expression, thereby preventing drug efflux and reversing multidrug resistance.145 Similarly, Lu et al synthesized an ellagic acid-copper nanoplatform combined with doxorubicin to form CS/NPs, which increased intracellular copper concentrations, reduced ATP-dependent P-gp expression, and overcame doxorubicin resistance in breast cancer cells.52 In response to radiotherapy resistance, nanocapsules synthesized by Liao et al released copper ions under controlled ionizing radiation, inducing cancer cell death and thus reversing tumor radioresistance.146 The NM-based strategies for inducing various forms of RCD in tumor therapy are summarized in Table 2.
 |
Table 2 Nanomaterial-Based Strategies for Inducing Regulated Cell Death in Tumor Therapy |
Anti-Tumor Activity Influenced by Physicochemical Properties of NMs
A significant focus in NM-based anti-tumor therapy is the meticulous design of their physicochemical properties, as these attributes profoundly affect cytotoxicity, circulation, biodistribution, and the initiation of various forms of RCD.
Particle size is particularly influential. Numerous studies have indicated that smaller NMs tend to exhibit enhanced cytotoxicity and RCD induction, attributed to their more efficient cellular penetration, nuclear localization, and ROS generation.147–149 Nonetheless, accumulating evidence indicates that the relationship between particle size and biological outcomes is highly context-dependent, varying according to the type of NMs and the tumor cell environment. For example, research on Au NPs has shown that particles of intermediate size (~50 nm) achieve the highest cellular uptake, outperforming both smaller and larger particles.150–152 This observation is believed to result from a balance between membrane wrapping energy and receptor-ligand binding kinetics.153–156 Importantly, nanoparticles of varying sizes can activate distinct RCD pathways. For instance, larger graphene oxide particles have been reported to activate phospholipase C signaling, leading to pyroptosis.157 Similarly, Si NPs with diameters less than 20 nm are capable of directly penetrating the cell membrane, leading to necroptosis. In contrast, particles measuring 50 nm are internalized via endocytosis and predominantly trigger caspase-mediated apoptosis.158
The shape of NMs is another critical factor, as it affects not only the rate of internalization but also the type of RCD that occurs.159 Rod-shaped NMs are internalized more rapidly and exhibit higher uptake compared to their spherical counterparts, likely due to the increased contact area provided by their longitudinal axis with the cell membrane.159,160 Additionally, surface modification of NMs can enhance their functionality through various strategies. For instance, ligand modification can improve tumor targeting and facilitate cellular uptake, thereby augmenting cytotoxicity and the induction of RCD.161–164 The formation of a protein corona can sometimes mitigate toxicity by decelerating internalization or neutralizing reactive surface groups.165 Hydrophilic modifications, such as PEG or hyaluronic acid, are known to extend blood circulation, enhance biodistribution, and improve the biocompatibility of NMs.166–168 The surface charge of NMs is also a critical determinant of their biological interactions. Cationic NMs typically exhibit greater cytotoxicity, which is likely attributable to their interactions with lysosomal membranes, resulting in a “proton sponge effect” and facilitating endosomal escape.169,170 Studies involving gold AuNPs have demonstrated that cationic particles can induce apoptosis at low concentrations (10 µg/mL) via mitochondrial stress and activation of the p53/caspase-3 pathway. Conversely, neutral particles necessitate higher concentrations (≥ 25 µg/mL) to primarily induce necrotic cell death.171
Challenges and Future Directions
By triggering RCD pathways, NMs unlock novel therapeutic possibilities in oncology.172 Organic NMs (eg, lipid nanoparticles, metal–organic frameworks) exhibit excellent biocompatibility, degradability, and stimulus-responsive properties. They enable precision delivery of drugs or immunomodulators while inducing ferroptosis or immunogenic cell death ICD within the TME, thereby enhancing anti-tumor immune responses.173,174 Inorganic NMs (eg, metal oxides) leverage photothermal, photodynamic, or magnetothermal effects to achieve precise tumor targeting while synergizing with RCD to amplify cytotoxic effects against tumor cells.175–177 Despite their potential, challenges remain in understanding the molecular mechanisms, pharmacokinetics, and standardization required for clinical translation.
Currently, the effects of physicochemical properties of NMs (eg, particle size, shape, surface modifications) on RCD induction remain insufficiently understood.158 Variations in NM properties, such as size and shape, critically determine their interactions with cellular membranes, internalization pathways, and activation of RCD signaling cascades. This intrinsic heterogeneity poses a fundamental constraint on the precision engineering of NMs for therapeutic use. In addition, the interplay between RCD and the TME remains unclear.4 The induction of RCD does not necessarily lead to the formation of a tumor-suppressive TME. For example, downregulating NRF2-related genes can enhance ferroptosis, but under certain conditions, this may paradoxically promote tumor progression.178,179 These findings highlight the need for more precise regulation of RCD pathways in therapy design. Current studies primarily focus on the activation of DCs and promotion of CD8+ T cells following DAMP release during NM-induced RCD. However, the effects on immunosuppressive cells, such as Tregs and MDSCs, have not been systematically evaluated. Likewise, the role of non-immune stromal cells in the TME, such as CAFs, remains poorly understood. The lack of studies in these areas indicates that therapeutic strategies based on NM-driven RCD still face uncertainties in both efficacy assessment and safety risks.
Equally important, the biodistribution, circulation, and clearance of NMs are critical determinants of the efficiency and safety of RCD induction. Different administration routes can lead to entirely distinct pharmacokinetic profiles and toxicity responses.180–182 The clinical translation of NMs remains hindered by insufficient data on their long-term toxicity, immunological side effects, and in vivo accumulation.180,182 Future research should focus on establishing standardized evaluation models and optimizing NM designs (eg, TME-responsive nanoplatforms) to enhance targeting specificity and long-term biosafety, thereby promoting their potential for clinical translation. Beyond these limitations, relying on a single RCD pathway is inadequate for complete tumor eradication and often leads to therapeutic resistance. Consequently, multimodal combination strategies represent a critical direction for future research. For example, photothermal and photodynamic therapies can directly kill tumor cells through localized heating or ROS generation, while simultaneously promoting necroptosis. The combined activation of ferroptosis and cuproptosis can induce severe oxidative stress and metabolic disruption, thereby amplifying cytotoxic effects. Moreover, RCD induction can upregulate immune checkpoint molecules such as PD-L1 on tumor cells, and combining this approach with immune checkpoint inhibitors can significantly enhance anti-tumor immune responses.183,184 Overall, therapeutic strategies integrating NMs and RCD remain in the early stages of exploration. The number of clinical studies demonstrating positive outcomes remains very limited,172,185,186 underscoring the urgent need for substantial research efforts to achieve successful clinical translation.
Conclusion
This review summarizes recent advances in using NMs to induce novel forms of RCD in tumors, including necroptosis, pyroptosis, ferroptosis, and cuproptosis. NMs regulate key signaling molecules and cellular metabolism to trigger these RCD types. For example, the RIPK1-RIPK3-MLKL complex mediates necroptosis, while NLRP3 inflammasome activation and GSDMD/GSDME cleavage induce pyroptosis. Excessive ROS and Fe³+ or Cu²+ accumulation drive ferroptosis and cuproptosis. RCD plays dual roles in tumor progression. On the one hand, it releases DAMPs to activate anti-tumor immunity, promoting DC maturation, CD8+ T cell recruitment, and M1 macrophage polarization, thereby inhibiting tumor growth and metastasis. On the other hand, specific cytokines and immunosuppressive signals induced by RCD may facilitate immune evasion and tumor progression. Understanding the context-dependent effects of RCD in specific tumor types and microenvironments is essential for precision therapy. Although NMs exhibit potential in cancer therapy, their in vivo distribution, metabolism, and long-term biological safety remain inadequately characterized. The impact of NMs on immunosuppressive and stromal cells within the TME is also insufficiently understood. Furthermore, the physicochemical properties of NMs, such as particle size, morphology, and surface functionalization, play a crucial role in determining their interactions with cell membranes, cellular uptake mechanisms, and the activation of RCD signaling pathways. However, inconsistencies and variability in current research findings present significant obstacles to the rational design of NMs and NM-based composites. Future research should prioritize optimizing the physicochemical properties and delivery strategies of NMs, as well as elucidating interactions among RCD, the TME, and metabolic signaling, to achieve precise therapeutic outcomes and facilitate clinical translation.
Abbreviations
NMs, Nanomaterials; RCD, regulated cell death; TME, tumor microenvironment; TAMs, tumor-associated macrophages; ROS, reactive oxygen species; GPX4, glutathione peroxidase-4; GSH, glutathione; ZBP1, Z-DNA-binding protein 1; ICD, immunogenic cell death; RIP1, receptor-interacting protease 1; RIP3, receptor-interacting protease 3; MLKL, mixed lineage kinase domain-like protein; TNF, tumor necrosis factor; IFN-γ, interferon gamma; TRADD, tumor necrosis factor receptor type 1-associated death domain protein; FADD, FAS-associated death domain protein; DAMPs, danger-associated molecular patterns; NLRP3, NOD-like receptor family pyrin domain-containing 3; GSDMD, gasdermin-D; GSDME, gasdermin-E; LPO, lipid peroxides; FDX1, ferredoxin 1; LIAS, lipoic acid synthase; DLAT, dihydrolipoamide S-acetyltransferase; PDAC, pancreatic ductal adenocarcinoma; HCC, hepatocellular carcinoma; EMT, epithelial-mesenchymal transition; TNBC, triple-negative breast cancer; CRGs, cuproptosis-related genes; MDSCs, myeloid-derived suppressor cells; CAFs, cancer-associated fibroblasts; NK, natural killer; DCs, dendritic cells; SDT, sonodynamic therapy; OSCC, oral squamous cell carcinoma; PEG, polyethylene glycol; CRT, calreticulin.
Consent for Publication
All authors read and agreed to submit the manuscript.
Acknowledgments
We thank LetPub (www.letpub.com.cn) for its linguistic assistance during the preparation of this manuscript. The graphic abstract, Figures 1–5 were created using biorender (www.biorender.com).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82001298), Nature Science Foundation of Guangdong Province (2024A1515030269), Characteristic and Innovative Projects of Common Colleges in Guangdong Province (2024KTSCX071), Science research cultivation program of stomatological hospital, Southern medical university (PY2023020), Outstanding Youths Development Scheme of Nanfang Hospital, Southern Medical University (2022J004).
Disclosure
The authors declare that they have no competing interests.
References
1. Liu W, Jin W, Zhu S, Chen Y, Liu B. Targeting regulated cell death (RCD) with small-molecule compounds in cancer therapy: a revisited review of apoptosis, autophagy-dependent cell death and necroptosis. Drug Discov Today. 2022;27(2):612–625. doi:10.1016/j.drudis.2021.10.011
2. Peng F, Liao M, Qin R, et al. Regulated cell death (RCD) in cancer: key pathways and targeted therapies. Signal Transduct Target Ther. 2022;7(1):286. doi:10.1038/s41392-022-01110-y
3. Zeng Q, Ma X, Song Y, Chen Q, Jiao Q, Zhou L. Targeting regulated cell death in tumor nanomedicines. Theranostics. 2022;12(2):817–841. doi:10.7150/thno.67932
4. Tong X, Tang R, Xiao M, et al. Targeting cell death pathways for cancer therapy: recent developments in necroptosis, pyroptosis, ferroptosis, and cuproptosis research. J Hematol Oncol. 2022;15(1):174. doi:10.1186/s13045-022-01392-3
5. Zhu X, Ferroptosis LS. Necroptosis, and pyroptosis in gastrointestinal cancers: the chief culprits of tumor progression and drug resistance. Adv Sci. 2023;10(26):e2300824. doi:10.1002/advs.202300824
6. Zhang C, Liu X, Jin S, Chen Y, Guo R. Ferroptosis in cancer therapy: a novel approach to reversing drug resistance. Mol Cancer. 2022;21(1):47. doi:10.1186/s12943-022-01530-y
7. Schwantes A, Wickert A, Becker S, et al. Tumor associated macrophages transfer ceruloplasmin mRNA to fibrosarcoma cells and protect them from ferroptosis. Redox Biol. 2024;71:103093. doi:10.1016/j.redox.2024.103093
8. Jin Y, Wu Q, Pan S, et al. Baicalein enhances cisplatin sensitivity in cervical cancer cells by promoting cuproptosis through the Akt pathway. Biomed Pharmacother. 2024;179:117415. doi:10.1016/j.biopha.2024.117415
9. Mohammadinejad R, Moosavi MA, Tavakol S, et al. Necrotic, apoptotic and autophagic cell fates triggered by nanoparticles. Autophagy. 2019;15(1):4–33. doi:10.1080/15548627.2018.1509171
10. Bonilla-Represa V, Abalos-Labruzzi C, Herrera-Martinez M, Guerrero-Pérez MO. Nanomaterials in dentistry: state of the art and future challenges. Nanomaterials. 2020;10(9):1770. doi:10.3390/nano10091770
11. García-Domínguez DJ, López-Enríquez S, Alba G, et al. Cancer nano-immunotherapy: the novel and promising weapon to fight cancer. IJMS. 2024;25(2):1195. doi:10.3390/ijms25021195
12. Cheng Z, Li M, Dey R, Chen Y. Nanomaterials for cancer therapy: current progress and perspectives. J Hematol Oncol. 2021;14(1):85. doi:10.1186/s13045-021-01096-0
13. Ma X, Cheng Z, Jin Y, et al. SM5-1-conjugated PLA nanoparticles loaded with 5-fluorouracil for targeted hepatocellular carcinoma imaging and therapy. Biomaterials. 2014;35(9):2878–2889. doi:10.1016/j.biomaterials.2013.12.045
14. Gao F, Wu Y, Wang R, et al. Precise nano-system-based drug delivery and synergistic therapy against androgen receptor-positive triple-negative breast cancer. Acta Pharmaceutica Sinica B. 2024;14(6):2685–2697. doi:10.1016/j.apsb.2024.03.012
15. Huang P, Wang X, Liang X, et al. Nano-, micro-, and macroscale drug delivery systems for cancer immunotherapy. Acta Biomater. 2019;85:1–26. doi:10.1016/j.actbio.2018.12.028
16. Zheng H, Jiang J, Xu S, et al. Nanoparticle-induced ferroptosis: detection methods, mechanisms and applications. Nanoscale. 2021;13(4):2266–2285. doi:10.1039/d0nr08478f
17. Wang Z, Wang M, Zeng X, Yue X, Wei P. Nanomaterial-induced pyroptosis: a cell type-specific perspective. Front Cell Dev Biol. 2023;11:1322305. doi:10.3389/fcell.2023.1322305
18. Li Z, Wang Y, Xu J, et al. Silica nanoparticles induce ferroptosis of HUVECs by triggering NCOA4-mediated ferritinophagy. Ecotoxicol Environ Saf. 2024;270:115889. doi:10.1016/j.ecoenv.2023.115889
19. Zhu Z, Zhang Y, Wang R, Dong Y, Wu J, Shao L. Zinc oxide nanoparticles disrupt the mammary epithelial barrier via Z-DNA binding protein 1-triggered PANoptosis. Ecotoxicol Environ Saf. 2024;283:116777. doi:10.1016/j.ecoenv.2024.116777
20. Xin L, Ning S, Wang H, Shi R. Tumor microenvironment responsive and platelet membrane coated polydopamine nanoparticles for cancer radiosensitization by inducing cuproptosis. IJN. 2025;Volume 20:3643–3652. doi:10.2147/IJN.S504148
21. Gao J, Ye T, Miao H, et al. Antibody-functionalized iron-based nanoplatform for ferroptosis-augmented targeted therapy of HER2-positive breast cancer. Bioact Mater. 2025;52:702–718. doi:10.1016/j.bioactmat.2025.06.034
22. Galluzzi L, Kepp O, Chan FKM, Kroemer G. Necroptosis: mechanisms and Relevance to Disease. Annu Rev Pathol. 2017;12:103–130. doi:10.1146/annurev-pathol-052016-100247
23. Sharifi M, Hosseinali SH, Saboury AA, Szegezdi E, Falahati M. Involvement of planned cell death of necroptosis in cancer treatment by nanomaterials: recent advances and future perspectives. J Control Release. 2019;299:121–137. doi:10.1016/j.jconrel.2019.02.007
24. Zielinska E, Zauszkiewicz-Pawlak A, Wojcik M, Inkielewicz-Stepniak I. Silver nanoparticles of different sizes induce a mixed type of programmed cell death in human pancreatic ductal adenocarcinoma. Oncotarget. 2018;9(4):4675–4697. doi:10.18632/oncotarget.22563
25. Farasat M, Niazvand F, Khorsandi L. Zinc oxide nanoparticles induce necroptosis and inhibit autophagy in MCF-7 human breast cancer cells. Biologia. 2020;75(1):161–174. doi:10.2478/s11756-019-00325-9
26. Arya BD, Mittal S, Joshi P, Pandey AK, Ramirez-Vick JE, Singh SP. Graphene oxide-chloroquine nanoconjugate induce necroptotic death in A549 cancer cells through autophagy modulation. Nanomedicine. 2018;13(18):2261–2282. doi:10.2217/nnm-2018-0086
27. Jiao H, Wachsmuth L, Kumari S, et al. Z-nucleic-acid sensing triggers ZBP1-dependent necroptosis and inflammation. Nature. 2020;580(7803):391–395. doi:10.1038/s41586-020-2129-8
28. Katifelis H, Nikou MP, Mukha I, et al. Ag/Au bimetallic nanoparticles trigger different cell death pathways and affect damage associated molecular pattern release in human cell lines. Cancers. 2022;14(6):1546. doi:10.3390/cancers14061546
29. Xiao H, Jensen PE, Chen X. Elimination of osteosarcoma by necroptosis with graphene oxide-associated anti-HER2 antibodies. Int J Mol Sci. 2019;20(18):4360. doi:10.3390/ijms20184360
30. Zhou XQ, Ren L, Zhang J, et al. Silica nanoparticles induce reversible damage of spermatogenic cells via RIPK1 signal pathways in C57 mice. IJN. 2016:2251. doi:10.2147/IJN.S102268
31. Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Sig Transduct Target Ther. 2021;6(1):128. doi:10.1038/s41392-021-00507-5
32. Burdette BE, Esparza AN, Zhu H, Wang S. Gasdermin D in pyroptosis. Acta Pharm Sin B. 2021;11(9):2768–2782. doi:10.1016/j.apsb.2021.02.006
33. Shirasuna K, Karasawa T, Takahashi M. Exogenous nanoparticles and endogenous crystalline molecules as danger signals for the NLRP3 inflammasomes. J Cell Physiol. 2019;234(5):5436–5450. doi:10.1002/jcp.27475
34. Hsu SK, Li CY, Lin IL, et al. Inflammation-related pyroptosis, a novel programmed cell death pathway, and its crosstalk with immune therapy in cancer treatment. Theranostics. 2021;11(18):8813–8835. doi:10.7150/thno.62521
35. Deng Y, Jia F, Jiang P, et al. Biomimetic nanoparticle synchronizing pyroptosis induction and mitophagy inhibition for anti-tumor therapy. Biomaterials. 2023;301:122293. doi:10.1016/j.biomaterials.2023.122293
36. Wang Y, Gao W, Shi X, et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature. 2017;547(7661):99–103. doi:10.1038/nature22393
37. Zhou B, Zhang JY, Liu XS, et al. Tom20 senses iron-activated ROS signaling to promote melanoma cell pyroptosis. Cell Res. 2018;28(12):1171–1185. doi:10.1038/s41422-018-0090-y
38. Zhao P, Wang M, Chen M, et al. Programming cell pyroptosis with biomimetic nanoparticles for solid tumor immunotherapy. Biomaterials. 2020;254:120142. doi:10.1016/j.biomaterials.2020.120142
39. Liu J, Kang R, Tang D. Signaling pathways and defense mechanisms of ferroptosis. FEBS J. 2022;289(22):7038–7050. doi:10.1111/febs.16059
40. Liang H, Wu X, Zhao G, Feng K, Ni K, Sun X. Renal clearable ultrasmall single-crystal fe nanoparticles for highly selective and effective ferroptosis therapy and immunotherapy. J Am Chem Soc. 2021;143(38):15812–15823. doi:10.1021/jacs.1c07471
41. Li J, Cao F, Yin HL, et al. Ferroptosis: past, present and future. Cell Death Dis. 2020;11(2):88. doi:10.1038/s41419-020-2298-2
42. Zhang Y, Tan H, Daniels JD, et al. Imidazole ketone erastin induces ferroptosis and slows tumor growth in a mouse lymphoma model. Cell Chem Biol. 2019;26(5):623–633.e9. doi:10.1016/j.chembiol.2019.01.008
43. Gao M, Deng J, Liu F, et al. Triggered ferroptotic polymer micelles for reversing multidrug resistance to chemotherapy. Biomaterials. 2019;223:119486. doi:10.1016/j.biomaterials.2019.119486
44. Tsvetkov P, Coy S, Petrova B, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. 2022;375(6586):1254–1261. doi:10.1126/science.abf0529
45. Su Z, Yao C, Tipper J, et al. Nanostrategy of targeting at embryonic trophoblast cells using cuo nanoparticles for female contraception. ACS Nano. 2023;17(24):25185–25204. doi:10.1021/acsnano.3c08267
46. Liu N, Chen M. Crosstalk between ferroptosis and cuproptosis: from mechanism to potential clinical application. Biomed Pharmacother. 2024;171:116115. doi:10.1016/j.biopha.2023.116115
47. Wang W, Lu K, Jiang X, et al. Ferroptosis inducers enhanced cuproptosis induced by copper ionophores in primary liver cancer. J Exp Clin Cancer Res. 2023;42(1):142. doi:10.1186/s13046-023-02720-2
48. Xiong C, Ling H, Hao Q, Zhou X. Cuproptosis: p53-regulated metabolic cell death? Cell Death Differ. 2023;30(4):876–884. doi:10.1038/s41418-023-01125-0
49. Su Q, Wang Z, Zhou H, et al. Eradication of large tumors by nanoscale drug self-assembly. Adv Mater. 2024;36(49):e2410536. doi:10.1002/adma.202410536
50. Hsieh CH, Hsieh HC, Shih FS, et al. An innovative NRF2 nano-modulator induces lung cancer ferroptosis and elicits an immunostimulatory tumor microenvironment. Theranostics. 2021;11(14):7072–7091. doi:10.7150/thno.57803
51. Tang W, Wu J, Wang L, et al. Bioactive layered double hydroxides for synergistic sonodynamic/cuproptosis anticancer therapy with elicitation of the immune response. ACS Nano. 2024;18(15):10495–10508. doi:10.1021/acsnano.3c11818
52. Lu S, Tian H, Li B, et al. An ellagic acid coordinated copper-based nanoplatform for efficiently overcoming cancer chemoresistance by cuproptosis and synergistic inhibition of cancer cell stemness. Small. 2024;20(17):e2309215. doi:10.1002/smll.202309215
53. Sun Y, Chen Y, Wu B, et al. Synergistic SDT/cuproptosis therapy for liver hepatocellular carcinoma: enhanced antitumor efficacy and specific mechanisms. J Nanobiotechnol. 2024;22(1):762. doi:10.1186/s12951-024-02995-3
54. Zheng Q, Zou T, Wang W, et al. Necroptosis-mediated synergistic photodynamic and glutamine-metabolic therapy enabled by a biomimetic targeting nanosystem for cholangiocarcinoma. Adv Sci. 2024;11(29):e2309203. doi:10.1002/advs.202309203
55. Koo GB, Morgan MJ, Lee DG, et al. Methylation-dependent loss of RIP3 expression in cancer represses programmed necrosis in response to chemotherapeutics. Cell Res. 2015;25(6):707–725. doi:10.1038/cr.2015.56
56. Lin P, Lin C, He R, et al. TRAF6 regulates the abundance of RIPK1 and inhibits the RIPK1/RIPK3/MLKL necroptosis signaling pathway and affects the progression of colorectal cancer. Cell Death Dis. 2023;14(1):6. doi:10.1038/s41419-022-05524-y
57. Guo X, Li R, Cui J, et al. Induction of RIPK3/MLKL-mediated necroptosis by Erigeron breviscapus injection exhibits potent antitumor effect. Front Pharmacol. 2023;14:1219362. doi:10.3389/fphar.2023.1219362
58. Yang C, Li J, Yu L, et al. Regulation of RIP3 by the transcription factor Sp1 and the epigenetic regulator UHRF1 modulates cancer cell necroptosis. Cell Death Dis. 2017;8(10):e3084. doi:10.1038/cddis.2017.483
59. Vucur M, Ghallab A, Schneider AT, et al. Sublethal necroptosis signaling promotes inflammation and liver cancer. Immunity. 2023;56(7):1578–1595.e8. doi:10.1016/j.immuni.2023.05.017
60. Liu ZG, Jiao D. Necroptosis, tumor necrosis and tumorigenesis. Cell Stress. 2019;4(1):1–8. doi:10.15698/cst2020.01.208
61. Jiao D, Cai Z, Choksi S, et al. Necroptosis of tumor cells leads to tumor necrosis and promotes tumor metastasis. Cell Res. 2018;28(8):868–870. doi:10.1038/s41422-018-0058-y
62. Liao CY, Li G, Kang FP, et al. Necroptosis enhances “don’t eat me” signal and induces macrophage extracellular traps to promote pancreatic cancer liver metastasis. Nat Commun. 2024;15(1):6043. doi:10.1038/s41467-024-50450-6
63. Strilic B, Yang L, Albarrán-Juárez J, et al. Tumour-cell-induced endothelial cell necroptosis via death receptor 6 promotes metastasis. Nature. 2016;536(7615):215–218. doi:10.1038/nature19076
64. Tang YL, Tao Y, Zhu L, Shen JL, Cheng H. Role of NLRP3 inflammasome in hepatocellular carcinoma: a double-edged sword. Int Immunopharmacol. 2023;118:110107. doi:10.1016/j.intimp.2023.110107
65. Wei Q, Mu K, Li T, et al. Deregulation of the NLRP3 inflammasome in hepatic parenchymal cells during liver cancer progression. Lab Invest. 2014;94(1):52–62. doi:10.1038/labinvest.2013.126
66. Zhang X, Li C, Chen D, et al. H. pylori CagA activates the NLRP3 inflammasome to promote gastric cancer cell migration and invasion. Inflamm Res. 2022;71(1):141–155. doi:10.1007/s00011-021-01522-6
67. Li S, Liang X, Ma L, et al. MiR-22 sustains NLRP3 expression and attenuates H. pylori-induced gastric carcinogenesis. Oncogene. 2018;37(7):884–896. doi:10.1038/onc.2017.381
68. Voronov E, Shouval DS, Krelin Y, et al. IL-1 is required for tumor invasiveness and angiogenesis. Proc Natl Acad Sci U S A. 2003;100(5):2645–2650. doi:10.1073/pnas.0437939100
69. Guo B, Fu S, Zhang J, Liu B, Li Z. Targeting inflammasome/IL-1 pathways for cancer immunotherapy. Sci Rep. 2016;6:36107. doi:10.1038/srep36107
70. Ding J, Wang K, Liu W, et al. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature. 2016;535(7610):111–116. doi:10.1038/nature18590
71. Xu C, Ma H, Chen J, et al. Prognostic, immunity, stemness, and anticancer drug sensitivity characterization of pyroptosis related genes in non-small cell lung cancer. Thorac Cancer. 2024;15(3):215–226. doi:10.1111/1759-7714.15180
72. Zhang P, Guo X, Li J, et al. Immunoglobulin-like transcript 4 promotes tumor progression and metastasis and up-regulates VEGF-C expression via ERK signaling pathway in non-small cell lung cancer. Oncotarget. 2015;6(15):13550–13563. doi:10.18632/oncotarget.3624
73. Gao J, Qiu X, Xi G, et al. Downregulation of GSDMD attenuates tumor proliferation via the intrinsic mitochondrial apoptotic pathway and inhibition of EGFR/Akt signaling and predicts a good prognosis in non‑small cell lung cancer. Oncol Rep. 2018;40(4):1971–1984. doi:10.3892/or.2018.6634
74. Wang WJ, Chen D, Jiang MZ, et al. Downregulation of gasdermin D promotes gastric cancer proliferation by regulating cell cycle-related proteins. J Dig Dis. 2018;19(2):74–83. doi:10.1111/1751-2980.12576
75. Tan G, Huang C, Chen J, Zhi F. HMGB1 released from GSDME-mediated pyroptotic epithelial cells participates in the tumorigenesis of colitis-associated colorectal cancer through the ERK1/2 pathway. J Hematol Oncol. 2020;13(1):149. doi:10.1186/s13045-020-00985-0
76. Hergueta-Redondo M, Sarrió D, Molina-Crespo Á, et al. Gasdermin-B promotes invasion and metastasis in breast cancer cells. PLoS One. 2014;9(3):e90099. doi:10.1371/journal.pone.0090099
77. Jiang L, Kon N, Li T, et al. Ferroptosis as a p53-mediated activity during tumour suppression. Nature. 2015;520(7545):57–62. doi:10.1038/nature14344
78. Zhang Y, Shi J, Liu X, et al. BAP1 links metabolic regulation of ferroptosis to tumour suppression. Nat Cell Biol. 2018;20(10):1181–1192. doi:10.1038/s41556-018-0178-0
79. Luo M, Bao L, Chen Y, et al. ZMYND8 is a master regulator of 27-hydroxycholesterol that promotes tumorigenicity of breast cancer stem cells. Sci Adv. 2022;8(28):eabn5295. doi:10.1126/sciadv.abn5295
80. Luo M, Bao L, Xue Y, et al. ZMYND8 protects breast cancer stem cells against oxidative stress and ferroptosis through activation of NRF2. J Clin Invest. 2024;134(6):e171166. doi:10.1172/JCI171166
81. Li Z, Hu Y, Zheng H, et al. LPCAT1-mediated membrane phospholipid remodelling promotes ferroptosis evasion and tumour growth. Nat Cell Biol. 2024;26(5):811–824. doi:10.1038/s41556-024-01405-y
82. Li F, Xu T, Chen P, et al. Platelet-derived extracellular vesicles inhibit ferroptosis and promote distant metastasis of nasopharyngeal carcinoma by upregulating ITGB3. Int J Biol Sci. 2022;18(15):5858–5872. doi:10.7150/ijbs.76162
83. Guan X, Wang Y, Yu W, et al. Blocking Ubiquitin-Specific protease 7 induces ferroptosis in gastric cancer via targeting Stearoyl-CoA desaturase. Adv Sci. 2024;11(18):2307899. doi:10.1002/advs.202307899
84. Dongre A, Weinberg RA. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat Rev Mol Cell Biol. 2019;20(2):69–84. doi:10.1038/s41580-018-0080-4
85. Ren Y, Mao X, Xu H, et al. Ferroptosis and EMT: key targets for combating cancer progression and therapy resistance. Cell Mol Life Sci. 2023;80(9):263. doi:10.1007/s00018-023-04907-4
86. di CH, Ye Z, feng HH, et al. SMAD4 endows TGF-β1-induced highly invasive tumor cells with ferroptosis vulnerability in pancreatic cancer. Acta Pharmacol Sin. 2024;45(4):844–856. doi:10.1038/s41401-023-01199-z
87. Bi J, Yang S, Li L, et al. Metadherin enhances vulnerability of cancer cells to ferroptosis. Cell Death Dis. 2019;10(10):682. doi:10.1038/s41419-019-1897-2
88. Wang C, Guo J, Zhang Y, Zhou S, Jiang B. Cuproptosis-related gene FDX1 suppresses the growth and progression of colorectal cancer by retarding EMT progress. Biochem Genet. 2024. doi:10.1007/s10528-024-10784-8
89. Sun B, Ding P, Song Y, et al. FDX1 downregulation activates mitophagy and the PI3K/AKT signaling pathway to promote hepatocellular carcinoma progression by inducing ROS production. Redox Biol. 2024;75:103302. doi:10.1016/j.redox.2024.103302
90. Peng Y, Shi R, Yang S, Zhu J. Cuproptosis‐related gene
91. Zhao Q, Qi T. The implications and prospect of cuproptosis-related genes and copper transporters in cancer progression. Front Oncol. 2023;13:1117164. doi:10.3389/fonc.2023.1117164
92. de Visser KE, Joyce JA. The evolving tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41(3):374–403. doi:10.1016/j.ccell.2023.02.016
93. Khan N, Lawlor KE, Murphy JM, Vince JE. More to life than death: molecular determinants of necroptotic and non-necroptotic RIP3 kinase signaling. Curr Opin Immunol. 2014;26:76–89. doi:10.1016/j.coi.2013.10.017
94. Zhang L, He Y, Jiang Y, et al. PRMT1 reverts the immune escape of necroptotic colon cancer through RIP3 methylation. Cell Death Dis. 2023;14(4):233. doi:10.1038/s41419-023-05752-w
95. Park HH, Kim HR, Park SY, et al. RIPK3 activation induces TRIM28 derepression in cancer cells and enhances the anti-tumor microenvironment. Mol Cancer. 2021;20(1):107. doi:10.1186/s12943-021-01399-3
96. Chan F, Rucker AJ, Park C, Li QJ, Moseman EA. Necroptosis stimulates interferon-mediated protective anti-tumor immunity. Res Sq. 2023;rs.3.rs–3713558. doi:10.21203/rs.3.rs-3713558/v1
97. Wu L, Zhang X, Zheng L, et al. RIPK3 orchestrates fatty acid metabolism in tumor-associated macrophages and hepatocarcinogenesis. Cancer Immunol Res. 2020;8(5):710–721. doi:10.1158/2326-6066.CIR-19-0261
98. Seifert L, Werba G, Tiwari S, et al. The necrosome promotes pancreatic oncogenesis via CXCL1 and mincle-induced immune suppression. Nature. 2016;532(7598):245–249. doi:10.1038/nature17403
99. Liu ZY, Zheng M, Li YM, et al. RIP3 promotes colitis-associated colorectal cancer by controlling tumor cell proliferation and CXCL1-induced immune suppression. Theranostics. 2019;9(12):3659–3673. doi:10.7150/thno.32126
100. Lei G, Zhuang L, Gan B. The roles of ferroptosis in cancer: tumor suppression, tumor microenvironment, and therapeutic interventions. Cancer Cell. 2024;42(4):513–534. doi:10.1016/j.ccell.2024.03.011
101. Wang W, Green M, Choi JE, et al. CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. Nature. 2019;569(7755):270–274. doi:10.1038/s41586-019-1170-y
102. Ma X, Xiao L, Liu L, et al. CD36-mediated ferroptosis dampens intratumoral CD8+ T cell effector function and impairs their antitumor ability. Cell Metab. 2021;33(5):1001–1012.e5. doi:10.1016/j.cmet.2021.02.015
103. Kim R, Hashimoto A, Markosyan N, et al. Ferroptosis of tumour neutrophils causes immune suppression in cancer. Nature. 2022;612(7939):338–346. doi:10.1038/s41586-022-05443-0
104. Li Y, Ma Z, Li W, et al. PDPN+ CAFs facilitate the motility of OSCC cells by inhibiting ferroptosis via transferring exosomal lncRNA FTX. Cell Death Dis. 2023;14(11):759. doi:10.1038/s41419-023-06280-3
105. Hao X, Zheng Z, Liu H, et al. Inhibition of APOC1 promotes the transformation of M2 into M1 macrophages via the ferroptosis pathway and enhances anti-PD1 immunotherapy in hepatocellular carcinoma based on single-cell RNA sequencing. Redox Biol. 2022;56:102463. doi:10.1016/j.redox.2022.102463
106. Ma X, Su W, Ye M, et al. Endogenous/exogenous stimulies inspired polyprodrug nano-inducer switches pyroptosis path for promoting antitumor immunity. Nano Today. 2023;48:101727. doi:10.1016/j.nantod.2022.101727
107. Wang B, Wang T, Jiang T, et al. Circulating immunotherapy strategy based on pyroptosis and STING pathway: mn-loaded paclitaxel prodrug nanoplatform against tumor progression and metastasis. Biomaterials. 2024;306:122472. doi:10.1016/j.biomaterials.2024.122472
108. Ma M, Jiang W, Zhou R. DAMPs and DAMP-sensing receptors in inflammation and diseases. Immunity. 2024;57(4):752–771. doi:10.1016/j.immuni.2024.03.002
109. Olivera I, Sanz-Pamplona R, Bolaños E, et al. A therapeutically actionable protumoral axis of cytokines involving IL-8, TNFα, and IL-1β. Cancer Discov. 2022;12(9):2140–2157. doi:10.1158/2159-8290.CD-21-1115
110. Yuan B, Clowers MJ, Velasco WV, et al. Targeting IL-1β as an immunopreventive and therapeutic modality for K-ras–mutant lung cancer. JCI Insight. 2022;7(11):e157788. doi:10.1172/jci.insight.157788
111. Meybodi SM, Ejlalidiz M, Manshadi MR, et al. Crosstalk between hypoxia-induced pyroptosis and immune escape in cancer: from mechanisms to therapy. Crit rev oncol/hematol. 2024;197:104340. doi:10.1016/j.critrevonc.2024.104340
112. Zhou Z, He H, Wang K, et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science. 2020;368(6494):eaaz7548. doi:10.1126/science.aaz7548
113. Zhang Z, Zhang Y, Xia S, et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature. 2020;579(7799):415–420. doi:10.1038/s41586-020-2071-9
114. Qiao L, Zhu G, Jiang T, et al. Self-destructive copper carriers induce pyroptosis and cuproptosis for efficient tumor immunotherapy against dormant and recurrent tumors. Adv Mater. 2024;36(8):e2308241. doi:10.1002/adma.202308241
115. Wang Q, Wang Y, Ding J, et al. A bioorthogonal system reveals antitumour immune function of pyroptosis. Nature. 2020;579(7799):421–426. doi:10.1038/s41586-020-2079-1
116. Denk D, Greten FR. Inflammation: the incubator of the tumor microenvironment. Trends Cancer. 2022;8(11):901–914. doi:10.1016/j.trecan.2022.07.002
117. Zhang RN, Jing ZQ, Zhang L, Sun ZJ. Epigenetic regulation of pyroptosis in cancer: molecular pathogenesis and targeting strategies. Cancer Lett. 2023;575:216413. doi:10.1016/j.canlet.2023.216413
118. Wang Y, Mang X, Guo X, Pu J. Distinct cuproptosis patterns in hepatocellular carcinoma patients correlate with unique immune microenvironment characteristics and cell-cell communication, contributing to varied overall survival outcomes. Front Immunol. 2024;15:1379690. doi:10.3389/fimmu.2024.1379690
119. Li H, Zu X, Hu J, et al. Cuproptosis depicts tumor microenvironment phenotypes and predicts precision immunotherapy and prognosis in bladder carcinoma. Front Immunol. 2022;13:964393. doi:10.3389/fimmu.2022.964393
120. Rimal R, Desai P, Daware R, et al. Cancer-associated fibroblasts: origin, function, imaging, and therapeutic targeting. Adv Drug Deliv Rev. 2022;189:114504. doi:10.1016/j.addr.2022.114504
121. Wang L, Cao Y, Guo W, Xu J. High expression of cuproptosis-related gene FDX1 in relation to good prognosis and immune cells infiltration in colon adenocarcinoma (COAD). J Cancer Res Clin Oncol. 2023;149(1):15–24. doi:10.1007/s00432-022-04382-7
122. Li J, Chen S, Liao Y, Wang H, Zhou D, Zhang B. Arecoline is associated with inhibition of cuproptosis and proliferation of cancer-associated fibroblasts in oral squamous cell carcinoma: a potential mechanism for tumor metastasis. Front Oncol. 2022;12:925743. doi:10.3389/fonc.2022.925743
123. Zhang S, Liu X, Zhang J, et al. Cancer-associated fibroblasts promote oral squamous cell carcinoma progression by targeting ATP7A via exosome-mediated paracrine miR-148b-3p. Cell Signalling. 2025;128:111631. doi:10.1016/j.cellsig.2025.111631
124. Saatci O, Alam R, Huynh-Dam KT, et al. Targeting LINC00152 activates cAMP/Ca2+/ferroptosis axis and overcomes tamoxifen resistance in ER+ breast cancer. Cell Death Dis. 2024;15(6):418. doi:10.1038/s41419-024-06814-3
125. Chen TC, Chuang JY, Ko CY, et al. AR ubiquitination induced by the curcumin analog suppresses growth of temozolomide-resistant glioblastoma through disrupting GPX4-Mediated redox homeostasis. Redox Biol. 2020;30:101413. doi:10.1016/j.redox.2019.101413
126. Wen H, Qu C, Wang Z, et al. Cuproptosis enhances docetaxel chemosensitivity by inhibiting autophagy via the DLAT/mTOR pathway in prostate cancer. FASEB J. 2023;37(9):e23145. doi:10.1096/fj.202300980R
127. Lin CN, Liang YL, Tsai HF, et al. Adipocyte pyroptosis occurs in omental tumor microenvironment and is associated with chemoresistance of ovarian cancer. J Biomed Sci. 2024;31(1):62. doi:10.1186/s12929-024-01051-4
128. Liao Y, Yang Y, sPan D, et al. HSP90α mediates sorafenib resistance in human hepatocellular carcinoma by necroptosis inhibition under hypoxia. Cancers. 2021;13(2):243. doi:10.3390/cancers13020243
129. Zhang H, Deng T, Liu R, et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol Cancer. 2020;19(1):43. doi:10.1186/s12943-020-01168-8
130. Zhang X, Tang B, Luo J, et al. Cuproptosis, ferroptosis and PANoptosis in tumor immune microenvironment remodeling and immunotherapy: culprits or new hope. Mol Cancer. 2024;23(1):255. doi:10.1186/s12943-024-02130-8
131. Ma L, Chen C, Zhao C, et al. Targeting carnitine palmitoyl transferase 1A (CPT1A) induces ferroptosis and synergizes with immunotherapy in lung cancer. Sig Transduct Target Ther. 2024;9(1):64. doi:10.1038/s41392-024-01772-w
132. Jiang Z, Lim SO, Yan M, et al. TYRO3 induces anti-PD-1/PD-L1 therapy resistance by limiting innate immunity and tumoral ferroptosis. J Clin Invest. 2021;131(8):e139434,139434. doi:10.1172/JCI139434
133. Zhang T, Yin C, Fedorov A, et al. ADAR1 masks the cancer immunotherapeutic promise of ZBP1-driven necroptosis. Nature. 2022;606(7914):594–602. doi:10.1038/s41586-022-04753-7
134. Zeng J, Chen H, Liu X, et al. Cuproptosis in microsatellite stable colon cancer cells affects the cytotoxicity of CD8+ T through the WNT signaling pathway. Chem Biol Interact. 2024;403:111239. doi:10.1016/j.cbi.2024.111239
135. Li J, Yi X, Liu L, Wang X, Ai J. Advances in tumor nanotechnology: theragnostic implications in tumors via targeting regulated cell death. Apoptosis. 2023;28(7–8):1198–1215. doi:10.1007/s10495-023-01851-3
136. Li S, Zhang T, Xu W, et al. Sarcoma-targeting peptide-decorated polypeptide nanogel intracellularly delivers shikonin for upregulated osteosarcoma necroptosis and diminished pulmonary metastasis. Theranostics. 2018;8(5):1361–1375. doi:10.7150/thno.18299
137. Li Y, Li H, Zhang K, et al. Genetically engineered membrane-coated nanoparticles for enhanced prostate-specific membrane antigen targeting and ferroptosis treatment of castration-resistant prostate cancer. Adv Sci. 2024;11(33):e2401095. doi:10.1002/advs.202401095
138. Abu-Serie MM, Osuka S, Heikal LA, Teleb M, Barakat A, Dudeja V. Diethyldithiocarbamate-ferrous oxide nanoparticles inhibit human and mouse glioblastoma stemness: aldehyde dehydrogenase 1A1 suppression and ferroptosis induction. Front Pharmacol. 2024;15:1363511. doi:10.3389/fphar.2024.1363511
139. Rioja-Blanco E, Arroyo-Solera I, Álamo P, et al. CXCR4-targeted nanotoxins induce GSDME-dependent pyroptosis in head and neck squamous cell carcinoma. J Exp Clin Cancer Res. 2022;41(1):49. doi:10.1186/s13046-022-02267-8
140. Zu H, Wu Y, Meng H, et al. Tumor metabolism aiming Cu2-xS nanoagents mediate photothermal-derived cuproptosis and immune activation. ACS Nano. 2024;18(35):23941–23957. doi:10.1021/acsnano.3c10588
141. Jiang C, Li W, Yan J, et al. A Robust ROS generation and ferroptotic lipid modulation nanosystem for mutual reinforcement of ferroptosis and cancer immunotherapy. Adv Healthc Mater. 2024;13(30):e2401502. doi:10.1002/adhm.202401502
142. Zhong H, Chen G, Li T, et al. Nanodrug augmenting antitumor immunity for enhanced tnbc therapy via pyroptosis and cGAS-STING activation. Nano Lett. 2023;23(11):5083–5091. doi:10.1021/acs.nanolett.3c01008
143. Nicosia A, Cavallaro G, Costa S, et al. Carbon nanodots for on demand chemophotothermal therapy combination to elicit necroptosis: overcoming apoptosis resistance in breast cancer cell lines. Cancers. 2020;12(11):3114. doi:10.3390/cancers12113114
144. Tsai T, Wu S, Lai Y, et al. CD44-hyaluronan mediating endocytosis of iron-platinum alloy nanoparticles induces ferroptotic cell death in mesenchymal-state lung cancer cells with tyrosine kinase inhibitor resistance. Acta Biomater. 2024;186:396–410. doi:10.1016/j.actbio.2024.07.020
145. Peng H, Zhang X, Yang P, et al. Defect self-assembly of metal-organic framework triggers ferroptosis to overcome resistance. Bioact Mater. 2023;19:1–11. doi:10.1016/j.bioactmat.2021.12.018
146. Liao Y, Wang D, Gu C, et al. A cuproptosis nanocapsule for cancer radiotherapy. Nat Nanotechnol. 2024;19(12):1892–1902. doi:10.1038/s41565-024-01784-1
147. Kim TH, Kim M, Park HS, Shin US, Gong MS, Kim HW. Size-dependent cellular toxicity of silver nanoparticles. J Biomed Mater Res A. 2012;100(4):1033–1043. doi:10.1002/jbm.a.34053
148. Ye Y, Liu J, Xu J, Sun L, Chen M, Lan M. Nano-SiO2 induces apoptosis via activation of p53 and Bax mediated by oxidative stress in human hepatic cell line. Toxicol In Vitro. 2010;24(3):751–758. doi:10.1016/j.tiv.2010.01.001
149. L K, L J, K M, et al. Two distinct cellular pathways leading to endothelial cell cytotoxicity by silica nanoparticle size. J Nanobiotechnol. 2019;17(1). doi:10.1186/s12951-019-0456-4
150. Chithrani BD, Ghazani AA, Chan WCW. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006;6(4):662–668. doi:10.1021/nl052396o
151. Yah CS. The toxicity of gold nanoparticles in relation to their physiochemical properties. Biomed Res. 2013;24(3):400–413.
152. Fratoddi I, Venditti I, Cametti C, Russo MV. How toxic are gold nanoparticles? The state-of-the-art. Nano Res. 2015;8(6):1771–1799. doi:10.1007/s12274-014-0697-3
153. Tzlil S, Deserno M, Gelbart WM, Ben-Shaul A. A statistical-thermodynamic model of viral budding. Biophys J. 2004;86(4):2037–2048. doi:10.1016/S0006-3495(04)74265-4
154. Gao H, Shi W, Freund LB. Mechanics of receptor-mediated endocytosis. Proc Natl Acad Sci U S A. 2005;102(27):9469–9474. doi:10.1073/pnas.0503879102
155. Chithrani BD, Chan WCW. Elucidating the mechanism of cellular uptake and removal of protein-coated gold nanoparticles of different sizes and shapes. Nano Lett. 2007;7(6):1542–1550. doi:10.1021/nl070363y
156. Chakraborty A, Das A, Raha S, Barui A. Size-dependent apoptotic activity of gold nanoparticles on osteosarcoma cells correlated with SERS signal. J Photochem Photobiol B Biol. 2020;203:111778. doi:10.1016/j.jphotobiol.2020.111778
157. Li J, Wang X, Mei KC, et al. Lateral size of graphene oxide determines differential cellular uptake and cell death pathways in Kupffer cells, LSECs, and hepatocytes. Nano Today. 2021;37:101061. doi:10.1016/j.nantod.2020.101061
158. Kim W, Kim WK, Lee K, et al. A reliable approach for assessing size-dependent effects of silica nanoparticles on cellular internalization behavior and cytotoxic mechanisms. IJN. 2019;Volume 14:7375–7387. doi:10.2147/IJN.S224183
159. Huang X, Teng X, Chen D, Tang F, He J. The effect of the shape of mesoporous silica nanoparticles on cellular uptake and cell function. Biomaterials. 2010;31(3):438–448. doi:10.1016/j.biomaterials.2009.09.060
160. Bhamidipati M, Fabris L. Multiparametric assessment of gold nanoparticle cytotoxicity in cancerous and healthy cells: the role of size, shape, and surface chemistry. Bioconjugate Chem. 2017;28(2):449–460. doi:10.1021/acs.bioconjchem.6b00605
161. Wu G, Jiang C, Zhang T. FcγRIIB receptor-mediated apoptosis in macrophages through interplay of cadmium sulfide nanomaterials and protein Corona. Ecotoxicol Environ Saf. 2018;164:140–148. doi:10.1016/j.ecoenv.2018.08.025
162. Ren Y, Yuan B, Hou S, et al. Delivery of RGD-modified liposome as a targeted colorectal carcinoma therapy and its autophagy mechanism. J Drug Target. 2021;29(8):863–874. doi:10.1080/1061186X.2021.1882469
163. Rompicharla SVK, Kumari P, Ghosh B, Biswas S. Octa-arginine modified poly(amidoamine) dendrimers for improved delivery and cytotoxic effect of paclitaxel in cancer. Artif Cells Nanomed Biotechnol. 2018;46(sup2):847–859. doi:10.1080/21691401.2018.1470527
164. De K. Decapeptide modified doxorubicin loaded solid lipid nanoparticles as targeted drug delivery system against prostate cancer. Langmuir. 2021;37(45):13194–13207. doi:10.1021/acs.langmuir.1c01370
165. Ji Y, Wang Y, Shen D, Kang Q, Chen L. Mucin Corona delays intracellular trafficking and alleviates cytotoxicity of nanoplastic-benzopyrene combined contaminant. J Hazard Mater. 2021;406:124306. doi:10.1016/j.jhazmat.2020.124306
166. Ying N, Lin X, Xie M, Zeng D. Effect of surface ligand modification on the properties of anti-tumor nanocarrier. Colloids Surf B Biointerfaces. 2022;220:112944. doi:10.1016/j.colsurfb.2022.112944
167. Ji P, Wang L, Chen Y, Wang S, Wu Z, Qi X. Hyaluronic acid hydrophilic surface rehabilitating curcumin nanocrystals for targeted breast cancer treatment with prolonged biodistribution. Biomater Sci. 2020;8(1):462–472. doi:10.1039/c9bm01605h
168. Shaik AS, Shaik AP, Bammidi VK, Al Faraj A. Effect of polyethylene glycol surface charge functionalization of SWCNT on the in vitro and in vivo nanotoxicity and biodistribution monitored noninvasively using MRI. Toxicol Mech Methods. 2019;29(4):233–243. doi:10.1080/15376516.2018.1540674
169. Ruenraroengsak P, Tetley TD. Differential bioreactivity of neutral, cationic and anionic polystyrene nanoparticles with cells from the human alveolar compartment: robust response of alveolar type 1 epithelial cells. Part Fibre Toxicol. 2015;12(1):19. doi:10.1186/s12989-015-0091-7
170. Cupic KI, Rennick JJ, Johnston AP, Such GK. Controlling endosomal escape using nanoparticle composition: current progress and future perspectives. Nanomedicine. 2019;14(2):215–223. doi:10.2217/nnm-2018-0326
171. Schaeublin NM, Braydich-Stolle LK, Schrand AM, et al. Surface charge of gold nanoparticles mediates mechanism of toxicity. Nanoscale. 2011;3(2):410–420. doi:10.1039/c0nr00478b
172. Zhu X, Li S. Nanomaterials in tumor immunotherapy: new strategies and challenges. Mol Cancer. 2023;22(1):94. doi:10.1186/s12943-023-01797-9
173. Chen J, Ye Z, Huang C, et al. Lipid nanoparticle-mediated lymph node-targeting delivery of mRNA cancer vaccine elicits robust CD8(+) T cell response. Proc Natl Acad Sci U S A. 2022;119(34):e2207841119. doi:10.1073/pnas.2207841119
174. Yang C, Ming H, Li B, et al. A pH and glutathione-responsive carbon monoxide-driven nano-herb delivery system for enhanced immunotherapy in colorectal cancer. J Control Release. 2024;376:659–677. doi:10.1016/j.jconrel.2024.10.043
175. Wang J, Wu X, Shen P, et al. Applications of inorganic nanomaterials in photothermal therapy based on combinational cancer treatment. Int J Nanomed. 2020;15:1903–1914. doi:10.2147/IJN.S239751
176. Ailioaie LM, Ailioaie C, Litscher G. Fighting cancer with photodynamic therapy and nanotechnologies: current challenges and future directions. Int J Mol Sci. 2025;26(7):2969. doi:10.3390/ijms26072969
177. Wang Y, Chang L, Gao H, Yu C, Gao Y, Peng Q. Nanomaterials-based advanced systems for photothermal / photodynamic therapy of oral cancer. Eur J Med Chem. 2024;272:116508. doi:10.1016/j.ejmech.2024.116508
178. DeNicola GM, Karreth FA, Humpton TJ, et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature. 2011;475(7354):106–109. doi:10.1038/nature10189
179. Jiang M, Qiao M, Zhao C, Deng J, Li X, Zhou C. Targeting ferroptosis for cancer therapy: exploring novel strategies from its mechanisms and role in cancers. Transl Lung Cancer Res. 2020;9(4):1569–1584. doi:10.21037/tlcr-20-341
180. Guo X, Mei N. Assessment of the toxic potential of graphene family nanomaterials. J Food Drug Anal. 2014;22(1):105–115. doi:10.1016/j.jfda.2014.01.009
181. Xu M, Qi Y, Liu G, Song Y, Jiang X, Du B. Size-dependent in vivo transport of nanoparticles: implications for delivery, targeting, and clearance. ACS Nano. 2023;17(21):20825–20849. doi:10.1021/acsnano.3c05853
182. Qi Y, Liu Y, Xia T, Xu A, Liu S, Chen W. The biotransformation of graphene oxide in lung fluids significantly alters its inherent properties and bioactivities toward immune cells. NPG Asia Materials. 2018;10:385–396. doi:10.1038/s41427-018-0039-0
183. Gao S, Ge H, Gao L, et al. Silk fibroin nanoparticles for enhanced cuproptosis and immunotherapy in pancreatic cancer treatment. Adv Sci. 2025;12(18):2417676. doi:10.1002/advs.202417676
184. Kim KS, Choi B, Choi H, Ko MJ, Kim DH, Kim DH. Enhanced natural killer cell anti-tumor activity with nanoparticles mediated ferroptosis and potential therapeutic application in prostate cancer. J Nanobiotechnol. 2022;20(1):428. doi:10.1186/s12951-022-01635-y
185. Zuckerman JE, Gritli I, Tolcher A, et al. Correlating animal and human phase Ia/Ib clinical data with CALAA-01, a targeted, polymer-based nanoparticle containing siRNA. Proc Natl Acad Sci USA. 2014;111(31):11449–11454. doi:10.1073/pnas.1411393111
186. Ishikawa T, Kageyama S, Miyahara Y, et al. Safety and antibody immune response of CHP-NY-ESO-1 vaccine combined with poly-ICLC in advanced or recurrent esophageal cancer patients. Cancer Immunol Immunother. 2021;70(11):3081–3091. doi:10.1007/s00262-021-02892-w
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Nanotechnology-Driven Strategies in Osteosarcoma Advances in Treatment: Immunotherapy and Drug Delivery
Su W, Li Y, Yang G, Zhao Y, Zhou X, Liu G, Huang X, Sohail MF, Hussain I, Liu Q, Chen F
International Journal of Nanomedicine 2025, 20:12913-12937
Published Date: 24 October 2025
Tumor Microenvironment Reprogramming via Copper-Enriched Black Phosphorus Nanoplatform for Cuproptosis-Sensitized Low-Dose Radioimmunotherapy
Chen Y, Wang J, Zheng D, Zhang W
International Journal of Nanomedicine 2025, 20:14551-14569
Published Date: 5 December 2025
Targeting Lactylation Offers Therapy to Reverse Cell Death Resistance
Wang Y, Chen J, Wang Y, Cao Y, Li Y, Zhu Y, Zhang Z, Wu S, Wang H
Drug Design, Development and Therapy 2025, 19:11371-11394
Published Date: 20 December 2025
From Barrier to Gateway: Nanomaterials Reshaping the Tumor Microenvironment for Therapy
Xie M, Sun W, Hu D, Liu L, Han C, Liu W, Liu L, Bao X, Zhang W, Hao X, Zhou Y
International Journal of Nanomedicine 2026, 21:573490
Published Date: 10 March 2026