Back to Journals » International Journal of Nanomedicine » Volume 20
Nanocomposites to Overcoming Sorafenib Resistance in Hepatocellular Carcinoma Therapy
Authors Liu J, Zhou S, Wei Y, Du J ![]() , Jia F, Dong W, Cao Y, Shi Z, Mu H, Chen L, Yu S
, Jia F, Dong W, Cao Y, Shi Z, Mu H, Chen L, Yu S ![]()
Received 19 April 2025
Accepted for publication 26 August 2025
Published 26 December 2025 Volume 2025:20 Pages 15723—15767
DOI https://doi.org/10.2147/IJN.S535455
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. RDK Misra
Jie Liu,1,* Shizhao Zhou,1,* Yingying Wei,2,3 Jinglei Du,2 Fan Jia,4 Wenhui Dong,1 Yangyi Cao,5 Zhenyang Shi,1 Hongkai Mu,1 Lin Chen,6 Shiping Yu2
1Medical Imaging Department, Shanxi Medical University, Taiyuan, Shanxi, 030001, People’s Republic of China; 2Department of Interventional Therapy, Shanxi Province Cancer Hospital, Shanxi Hospital Affiliated to Cancer Hospital, Chinese Academy of Medical Sciences, Cancer Hospital Affiliated to Shanxi Medical University, Taiyuan, Shanxi, 030013, People’s Republic of China; 3Affiliated Cancer Hospital, Shanxi Medical University, Taiyuan, Shanxi, 030001, People’s Republic of China; 4Department of Oncology and Vascular Intervention, First Hospital of Shanxi Medical University, Taiyuan, Shanxi, 030001, People’s Republic of China; 5Academy of Medical Sciences, Shanxi Medical University, Taiyuan, Shanxi, 030001, People’s Republic of China; 6Key Laboratory of Interface Science and Engineering in Advanced Materials, Ministry of Education, Taiyuan University of Technology, Taiyuan, Shanxi, 030024, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lin Chen, Email [email protected] Shiping Yu, Email [email protected]
Abstract: The emergence of drug resistance is the major obstacle to the clinical application of sorafenib (SOR), which often leads to disease progression, recurrence, and even death in hepatocellular carcinoma (HCC) patients. Nanocomposite-mediated drug delivery enhances targeting precision and therapeutic utilization efficiency. Nanocomposites constructed by nanoparticles (NPs) and various therapeutic components have emerged as effective approaches to enhance HCC therapeutic efficacy. Designing based on the mechanisms underlying SOR resistance, specially engineered nanocomposites can be designed to overcome SOR resistance. This review aims to highlight the advantages of nanocomposites in overcoming HCC SOR resistance. First, the various SOR resistance mechanisms that have been identified so far are briefly outlined. Second, the construction methods and characteristics of nanocomposites designed to overcome SOR resistance are summarized and categorized according to different types of NPs. Subsequently, the roles and therapeutic effects of nanocomposites in SOR-resistant HCC are analyzed, primarily including remodeling the tumor microenvironment (TME), restoring normal epigenetic regulation, improving drug metabolism, and inhibiting abnormally activated signaling molecules and pathways. Finally, the advantages and disadvantages of nanocomposites used to reverse drug resistance are discussed, and their development direction in future research is prospected, which provide new approaches for developing advanced nanocomposites to overcome SOR resistance.
Keywords: nanocomposites, nanoparticles, nanomedicine, hepatocellular carcinoma, sorafenib, drug resistance, drug delivery, combination therapies
Introduction
HCC is the most common histological subtype of liver cancer,1 accounting for approximately 90% of all cases. Owing to complex etiology, latent pathogenesis, difficult diagnosis, and rapid progression, in more than 50% of HCC patients are diagnosed at advanced stages (Barcelona clinic liver cancer stage C or higher), limiting treatment options to systemic therapy. Molecular targeting agents (MTAs), known for their “high efficacy and low toxicity”, are often the first choice for systemic treatment.2,3 Among them, multi-kinase inhibitors (MKIs) are widely used in various malignant tumors,4 including HCC. SOR, a typical MKI, has been approved by the US Food and Drug Administration (FDA) as a first-line MTA for advanced HCC. SOR, a clinically approved multi-kinase inhibitor, suppresses tumor progression by targeting key signaling pathways critical for cell proliferation and angiogenesis.5
The gradual development of resistance to SOR during HCC treatment often leads to tumor recurrence, progression, and ultimately patient death. Therefore, overcoming SOR resistance is crucial for prolonging the survival rate of HCC patients. To effectively tackle this resistance, it is imperative to understand its underlying causes. The mechanisms contributing to SOR resistance primarily involve the emergence of TME, abnormal alterations in epigenetic regulation, aberrant drug metabolism, epithelial–mesenchymal transition (EMT), and activation of abnormal signaling molecules associated with SOR resistance.6,7 Targeted inhibition of these pathways has been shown to resensitize tumors to SOR, significantly improving median survival in resistant patients. In recent years, researchers have developed various drugs, genes, and functional materials based on the mechanisms of SOR resistance. For instance, metapristone (Meta), hypoxia-inducible factor-1α (HIF1α) siRNA, and perfluorohexane (PFH) can overcome hypoxia-induced SOR resistance by inhibiting the expression of the CXCR4, inhibiting HIF1α mRNA expression, and delivering exogenous O2.8–10 Despite these advances, therapeutic efficacy is hindered by insufficient drug accumulation in tumor sites, highlighting the need for nanotechnology-based delivery systems to enhance targeting precision.
Advanced nanomedicine has offered a novel strategy for reversing SOR resistance. NPs can be used as carriers to construct nanocomposites loaded with drugs, genes, and functional materials. As multifunctional platforms, nanocomposites not only enhance SOR delivery to HCC tissues but also leverage unique physicochemical properties for microenvironment modulation.11 For example, manganese dioxide NPs (MnO2 NPs) catalytically decompose tumor-associated H2O2 into O2, effectively reversing hypoxia-induced SOR resistance.12 In recent years, NPs applied to overcome SOR resistance primarily include lipid NPs, polymeric NPs, inorganic NPs, and extracellular vesicles (EVs). The rational design of nanocomposites is of great significance for more effectively overcoming SOR resistance.
As the number of SOR-resistant HCC patients continues to rise, researchers have made significant strides in exploring solutions through nanomedicine in recent years. However, no comprehensive review has yet summarized the nanocomposites designed to overcome SOR resistance. A timely review of current research enables researchers to swiftly grasp the latest advancements in this field and guide them in designing nanocomposites to tackle SOR resistance and facilitating their translation into clinical applications. This review initially outlines the mechanisms underlying SOR resistance in HCC. Subsequently, the design principles of nanocomposites using organic NPs, inorganic NPs, and EVs as nanocarriers for overcoming SOR resistance are systematically elucidated. Furthermore, the applications and mechanisms of action of these nanocomposite particles in addressing different resistance mechanisms are analyzed. At last, the challenges and future opportunities of using nanocomposites to overcome SOR resistance in HCC are discussed and prospected, providing new insights and directions for extending the survival rate of patients with SOR and other MTA-resistant HCC in clinical practice. As schematized in Figure 1, nanocomposites precisely target distinct resistance mechanisms through multifunctional engineering.
 |
Figure 1 Nanocomposites for targeting inhibition of SOR resistance mechanisms to restore sensitivity of HCC to SOR. |
Mechanisms of SOR Resistance in HCC
SOR inhibits the proliferation of HCC by targeting the rat sarcoma virus gene (RAS)-rapidly accelerated fibrosarcoma (Raf)-mitogen-activated protein kinase kinase (MEK)-extracellular signal-regulated kinase (ERK) signaling pathway, mainly acting on Raf-1 and B-Raf kinases. Meanwhile, SOR also inhibits the tyrosine kinase activities of various receptors including (VEGFR)-2, VEGFR-3, platelet-derived growth factor receptor β (PDGF-β), and hepatocyte growth factor receptor. Under the combined effect of the dual mechanisms, tumor angiogenesis and tumor growth are inhibited by blocking the nutrient supply and metabolic reprogramming of HCC5 (Figure 2).
 |
Figure 2 The therapeutic mechanism of SOR in HCC. |
The development of SOR resistance in HCC is attributed to a multitude of mechanisms, including the remodeling of TME, abnormal alterations in epigenetic regulation, aberrant drug metabolism, EMT, and activation of abnormal signaling molecules associated with SOR resistance.6 Some of these mechanisms directly lead to mutations in SOR targets, preventing SOR from binding and exerting its effects, while others evade or diminish the lethal action of SOR, ultimately resulting in SOR resistance. This section provides a detailed review of the current mechanisms of SOR resistance, laying the foundation for the rational design of nanocomposites to overcome SOR resistance.
TME
The TME is a dynamic ecosystem composed of cellular components (such as CAFs and immunosuppressive cells) and non-cellular components (such as ECM and cytokines). When SOR is used to treat HCC, the high metabolic demand of HCC and the therapeutic effect of SOR interact with each other, causing the TME to be rapidly remodeling into a treatation-resistant microenvironment — characterized by hypoxia-induced metabolic reprogramming, enrichment of cancer stem cell properties, infiltration of immunosuppressive cells, and ferroptosis inhibition, which enhances SOR resistance6,13–15 (Figure 3).
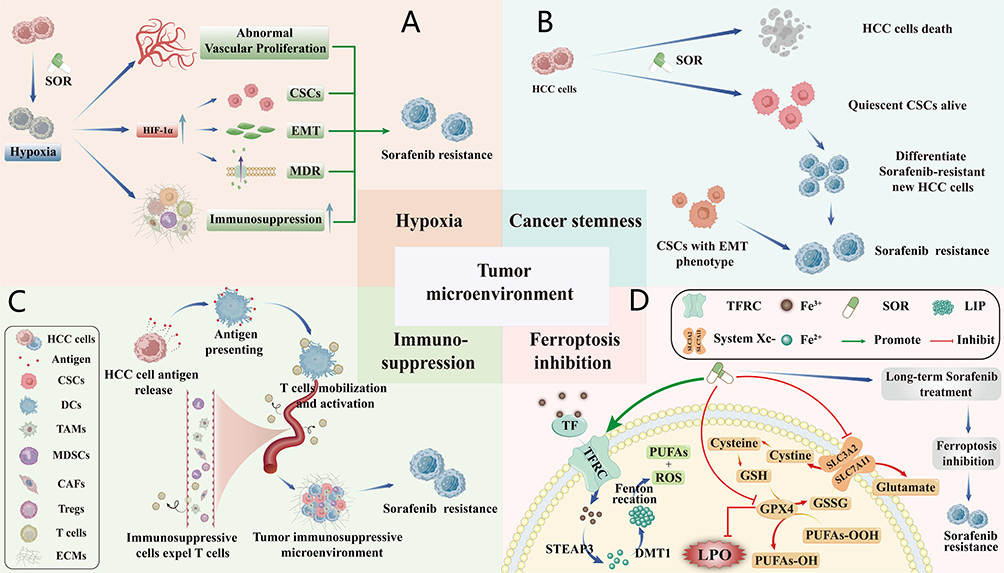 |
Figure 3 The related mechanisms of SOR resistance in TME. (A) Hypoxia. (B) Cancer stemness. (C) Immunosuppression. (D) Ferroptosis inhibition, upward arrows (↑) indicating upregulation or increased expression and downward arrows (↓) indicating downregulation or decreased expression. |
Hypoxia
SOR exerts an anti-angiogenic effect in HCC, but it also leads to the reduction in functional blood vessels and insufficient tumor blood perfusion, which in turn exacerbates hypoxia.16 Hypoxic conditions induce HIF-1α activation, which regulates glycolysis and tumor angiogenesis,17 allowing HCC to adapt to the anti-angiogenic effects of SOR and causing SOR resistance.18,19 Additionally, the upregulation of HIF-1 can activate the multidrug resistance-1 (MDR1) gene and upregulates P-glycoprotein (P-gp), promoting SOR efflux through ATP-dependent drug extrusion, thus inducing MDR and SOR resistance.20 Hypoxia drives immunosuppressive microenvironment formation, EMT, and extracellular acidosis, collectively establishing a resistance-permissive niche21,22 (Figure 3A). Therefore, targeting hypoxia signaling pathways represents a pivotal strategy for overcoming SOR resistance.23
Cancer Stemness
Cancer stemness refers to the self-renewal capacity and pluripotency of cancer stem cells (CSCs), which drive tumor initiation, metastasis, and therapy resistance. Nanocomposites inhibit CSC properties by suppressing key pathways. CSCs represent a unique subpopulation of cancer cells with self-renewal capabilities.24,25 Cytotoxic drugs such as SOR can inhibit the growth of non-quiescent cells. Although cytotoxic drugs like SOR inhibit non-quiescent cell proliferation, the majority of CSCs reside in a quiescent state, thereby evading SOR-induced cytotoxicity.26 Following treatment, quiescent CSCs can counteract SOR through DNA damage repair and activation of resistance-associated signaling pathways.27 These progenitor cells subsequently differentiate into SOR-resistant cancer populations (Figure 3B). In addition, a subset of CSCs located at the invasive front of tumors possess EMT-phenotype-related abilities, promoting HCC invasion and metastasis, thereby exacerbating SOR resistance through enhanced metastatic dissemination.28
Immunosuppression
In physiological conditions, the immune system effectively identifies and eliminates foreign pathogens as well as autologous cancer cells. However, immunosuppressive components within the TME—including regulatory T cells (Tregs), CAFs, myeloid-derived suppressor cells (MDSCs), and tumor-associated macrophages (TAMs)—promote irreversible T cell exhaustion by excluding effector T cells from tumor niches. This process drives immune evasion and compromises SOR efficacy.29–32 Furthermore, immunosuppressive cells, along with the remodeling of the ECM, secretion of immunosuppressive chemokines, and CSCs, collectively constitute the tumor immune microenvironment, which can contribute to decreased SOR efficacy by inducing immune tolerance in HCC cells, ultimately fostering the development of SOR resistance33,34 (Figure 3C).
Ferroptosis Inhibition
Ferroptosis, a type of programmed cell death driven by iron-dependent lipid peroxidation (LPO) accumulation35 is frequently suppressed in SOR resistance. This inhibition is mediated by GPX4 activation or direct LPO scavenging mechanisms. SOR promotes ferroptosis in HCC cells mainly through two pathways. First, SOR facilitates the recognition and uptake of the Fe3+-transferrin (TF) complex in the bloodstream into HCC cells via the transferrin receptor (TFRC). Upon entering the cell, Fe3+ is reduced to Fe2+ by the metal reductase six-transmembrane epithelial antigen of the prostate 3 (STEAP3) and mediated by divalent metal transporter 1 (DMT1) to store in labile iron pool (LIP) as the form of storage iron. Then, the Fe2+ reacts with H2O2 in TME through the Fenton reaction to generate reactive oxygen species (ROS). ROS can further react with polyunsaturated fatty acids (PUFAs) on the cell membrane and plasma membrane, causing their peroxidation to produce LPO, thereby triggering ferroptosis.36,37 Second, SOR reduces the concentration of intracellular glutathione (GSH) by inhibiting the transport GSH of by cystine/glutamate transporter system (System Xc-) and the activity of glutathione peroxidase 4 (GPX4), thus reducing the consumption of LPO by GSH.37 However, HCC cells develop resistance by disrupting ROS-PUFA interactions, reducing ROS generation, and elevating GSH synthesis38,39 (Figure 3D).
Abnormal Epigenetic Regulation
Epigenetic regulation is closely associated with the initiation and progression of cancer,40 including DNA methylation, histone modification, and non-coding RNA (ncRNA). Abnormal epigenetic regulation promotes SOR resistance by dysregulation in SOR therapeutic targets, which in turn affect drug transport, metabolism, cell proliferation, autophagy, apoptosis, cell cycle, TME remodeling, and EMT41 (Figure 4). This section summarizes the role of epigenetic regulation in SOR resistance.
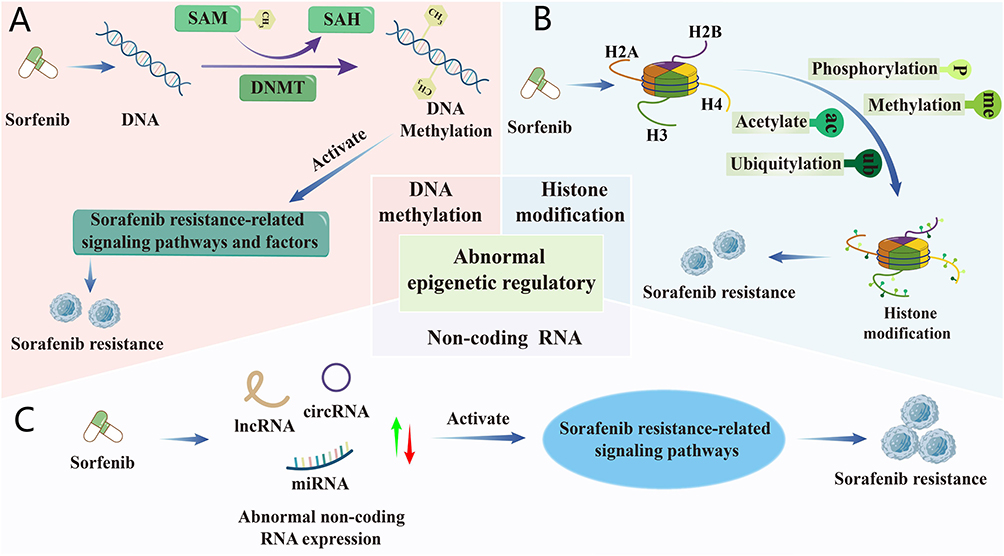 |
Figure 4 Mechanisms underlying SOR resistance in epigenetic regulation. (A) DNA methylation. (B) Histone modification. (C) Non-coding RNA, upward arrows (↑) indicating upregulation or increased expression and downward arrows (↓) indicating downregulation or decreased expression. |
DNA Methylation
DNA methylation involves the enzymatic transfer of a methyl group from S-adenosyl methionine (SAM) to cytosine at the CpG dinucleotide 5-position, forming 5-methylcytosine (5-mC) via DNA methyltransferases (DNMTs; eg, DNMT1, DNMT3A, DNMT3B). This catalysis simultaneously converts SAM to S-adenosyl-l-homocysteine (SAH). DNA methylation can suppress the expression of downstream genes by recruiting DNA-binding proteins and histone modifiers that inhibit transcription, particularly when located in promoter regions or at the transcription start site (TSS). For example, MORC2, a member of the microrchidia (MORC) protein family, can interact with DNMT3A and recruit it to the promoters of neurofibromatosis type 2 and KIBRA to facilitate their DNA methylation, thereby silencing the downstream Hippo signaling pathway and promoting HCC cancer stemness, ultimately leading to SOR resistance42 (Figure 4A). In addition, DNA methylation also induces HCC SOR resistance by inhibiting apoptosis of HCC cells, modulating the expression of genes related to SOR transport and metabolism, promoting EMT and causing immunosuppression.43,44
Histone Modification
Histones are highly conserved proteins consisting of five core types: H1, H3, H2A, H2B, and H4, which form the nucleosome with DNA. The amino-terminal tails of histones undergo post-translational modifications, including phosphorylation, methylation, acetylation, ubiquitination, glycosylation, ADP-ribosylation, deimination, sumoylation, and proline isomerization, leading to the activation of signaling molecules or pathways associated with SOR resistance (Figure 4B). For example, histone deacetylase 11 (HDAC11) reduces the expression of the serine/threonine protein kinase LKB1 by inhibiting histone acetylation in the promoter region of LKB1, thereby suppressing the activation of AMPK signaling pathway to enhance glycolysis, ultimately promoting both cancer stemness and SOR resistance in HCC.45 The methyltransferase SET domain containing 1A (SETD1A) facilitates the activation of yes-associated protein (YAP), a core transcriptional co-activator of the Hippo signaling pathway, and the downstream genes of YAP such as cysteine-rich angiogenic inducer 61 and connective tissue growth factor by reducing YAP phosphorylation, enabling HCC cells to adapt to hypoxia and thus induce SOR resistance.46,47 Furthermore, histone modifications can promote SOR resistance by influencing processes such as DNA repair, replication, cancer stemness, and cell state transitions.
MicroRNA
NcRNAs refer to functional RNAs that can be transcribed but not translated into proteins, primarily including microRNA (miRNA), long non-coding RNA (lncRNA), and circular RNA (circRNA). Their expression levels sometimes become aberrant during SOR treatment, leading to the activation of signaling molecules or pathways associated with SOR resistance (Figure 4C).48 This further induces hypoxia, EMT, cancer stemness, immunosuppression, and others, ultimately resulting in SOR resistance.49 Among them, miRNA is a type of single-stranded RNA about 21–25 nucleotides in length, which typically binds to the 3′ untranslated region (3’UTR) of target genes and acts as mRNA sponges to reduce the expression of relevant genes and signaling pathways, thereby exerting RNA interference (RNAi) and participating in SOR resistance. For instance, liver-specific miRNA-122 (miR-122) is the most abundant miRNA in normal liver, plays a pivotal role in hepatic homeostasis, metabolism, and differentiation.50 Studies have shown that the loss of miR-122 promotes the development of HCC and contributes to SOR resistance by enhancing cancer stemness, activating insulin-like growth factor 1 receptor and the RAS/RAF/ERK signaling pathway.49,51 Overexpression of miRNA-494 also induces SOR resistance by promoting cancer stemness and activating the PTEN/PI3K/Akt/mTOR signaling pathway.52 In addition, down-regulation of miR-145-5p can also contribute to SOR resistance by up-regulating HDAC11 expression.53 Evidently, miRNAs play a crucial role in SOR resistance.
Long Non-Coding RNA
lncRNAs, defined as non-coding transcripts exceeding 200 nucleotides in length, function not only as miRNA sponges to regulate miRNA expression but also interact with RNA-binding proteins (RBPs) to modulate drug resistance genes via DNA damage response pathways.54 Multifunctional lncRNA plays multifaceted roles in cancer development, including sustaining HCC cell proliferation, inhibiting HCC cell apoptosis, promoting angiogenesis, and facilitating HCC metastasis.55,56 For example, LncSNHG16 acts as an endogenous sponge for miRNA-140-5p, contributing to SOR resistance through enhancing the stemness of CSCs.57 The loss of Linc01056 leads to the transcriptional activation of peroxisome proliferator-activated receptor α, inducing the expression of fatty acid oxidation-related genes while suppressing glycolysis-related genes. This metabolic shift enhances intracellular energy production, counteracting SOR-induced cytotoxicity, and driving resistance.55 Additionally, lncRNAs promote SOR resistance through autophagy induction, EMT activation, AKT pathway stimulation, and apoptosis suppression.49,56
Circular RNA
CircRNA, a class of ncRNAs characterized by their closed-loop structure, contributes to SOR resistance in HCC through alterations in its expression patterns, as evidenced by relevant studies.58 For example, circRNA-SORE activates key downstream signaling pathways associated with SOR resistance, including AKT, Raf1, ERK, c-Myc, and transforming growth factor β1 (TGF-β1), and propagates this resistance between HCC cells via exosomal transfer.59 In addition, circFN1 enhances SOR resistance by regulating the miR-1205/E2F1 signaling pathway.60 circFOXM1 contributes to SOR resistance of HCC by regulating MECP2 as a sponge of miR-1324.61 Collectively, these findings demonstrate that circRNAs drive SOR resistance through diverse molecular mechanisms.
Other Mechanisms Related to SOR Resistance
Aberrant activation of signaling pathways or molecules associated with SOR resistance, EMT and insufficient effective SOR concentrations due to abnormal drug metabolism collectively drive SOR resistance in HCC (Figure 5). In-depth understanding of these mechanisms is crucial for developing effective strategies to overcome SOR resistance.
 |
Figure 5 Other mechanisms related to SOR resistance. A) Activation of signaling molecules and pathways associated with SOR resistance. (B) Epithelial-mesenchymal transtion. (C) Abnormal drug metabolism. |
Activation of Signaling Molecules and Pathways Associated with SOR Resistance
SOR was designed to treat HCC by inhibiting the aberrantly activated RAS/RAF/MEK/ERK pathway and its downstream signaling molecules. However, during treatment, compensatory activation of downstream signaling components allows tumor cells to bypass SOR-mediated inhibition. For instance, various genes or signaling molecules, including miRNA-494, DNA primase subunit 1, and FXYD domain containing ion transport regulator 5, can evade inhibitory effects of SOR effects by activating the PI3K/AKT/mTOR and RAS/Raf/MAPK pathways.52,62,63 Furthermore, activation of downstream signaling molecules can also diminish efficacy of SOR. Cofilin1 (CFL1) can facilitate Nrf2 nuclear translocation to eliminate excess ROS induced by SOR, contributing to SOR resistance.64 In summary, abnormal activation of downstream signaling pathways and factors in the RAS/RAF/MEK/ERK pathway is one of the factors contributing to SOR resistance (Figure 5A).
EMT
EMT refers to the biological process through which epithelial cells transform into cells with a mesenchymal phenotype via specific programs. Specifically, cancer cells develop type III EMT under the stimulation of intracellular and extracellular factors such as hypoxia and DNA methylation induced during SOR treatment.65 During this process, HCC cells gradually lose their epithelial phenotype and intercellular adhesive structures, acquire cancer stemness,66 thereby gaining self-renewal and drug resistance capabilities (Figure 5B). Various signaling pathways and factors, including Notch, TGF-β, Wnt, and Hedgehog, can promote the occurrence of EMT, ultimately inducing SOR resistance.67
Abnormal Drug Metabolism
Owing to its poor water solubility (~1.7 μg/mL), inability to selectively accumulate in HCC cells, and rapid in vivo metabolism. Consequently, SOR requires high-dose administration to achieve therapeutic efficacy,68 which may subsequently induce MDR. MDR is primarily mediated by P-gp overexpression. P-gp, an ATP-dependent efflux pump embedded in the cell membrane, exports SOR out of cells along a concentration gradient. Additionally, the weakly basic nature of SOR enables its acidic lysosomal sequestration via pH-driven diffusion or ABC transporter-mediated transport. These processes reduce SOR’s intracellular concentration below therapeutic levels, driving resistance development69 (Figure 5C).
As aforementioned, various mechanisms related to SOR resistance have been identified and extensively studied. However, how to overcome SOR resistance by regulating these resistance mechanisms remains a critical issue for improving the outcomes HCC patients.
Construction of SOR-Resistant Nanocomposites for Overcoming SOR Resistance in HCC
The emergence of nanomedicine has provided effective means to overcome SOR resistance. Researchers have designed numerous nanocomposites using NPs as carriers to target the mechanisms of SOR resistance and overcome SOR resistance. Currently, commonly used NPs include organic NPs, inorganic NPs, and EVs. The construction strategies, design methods, and their relevant properties of nanocomposites for overcoming SOR resistance are summarized (Table 1).
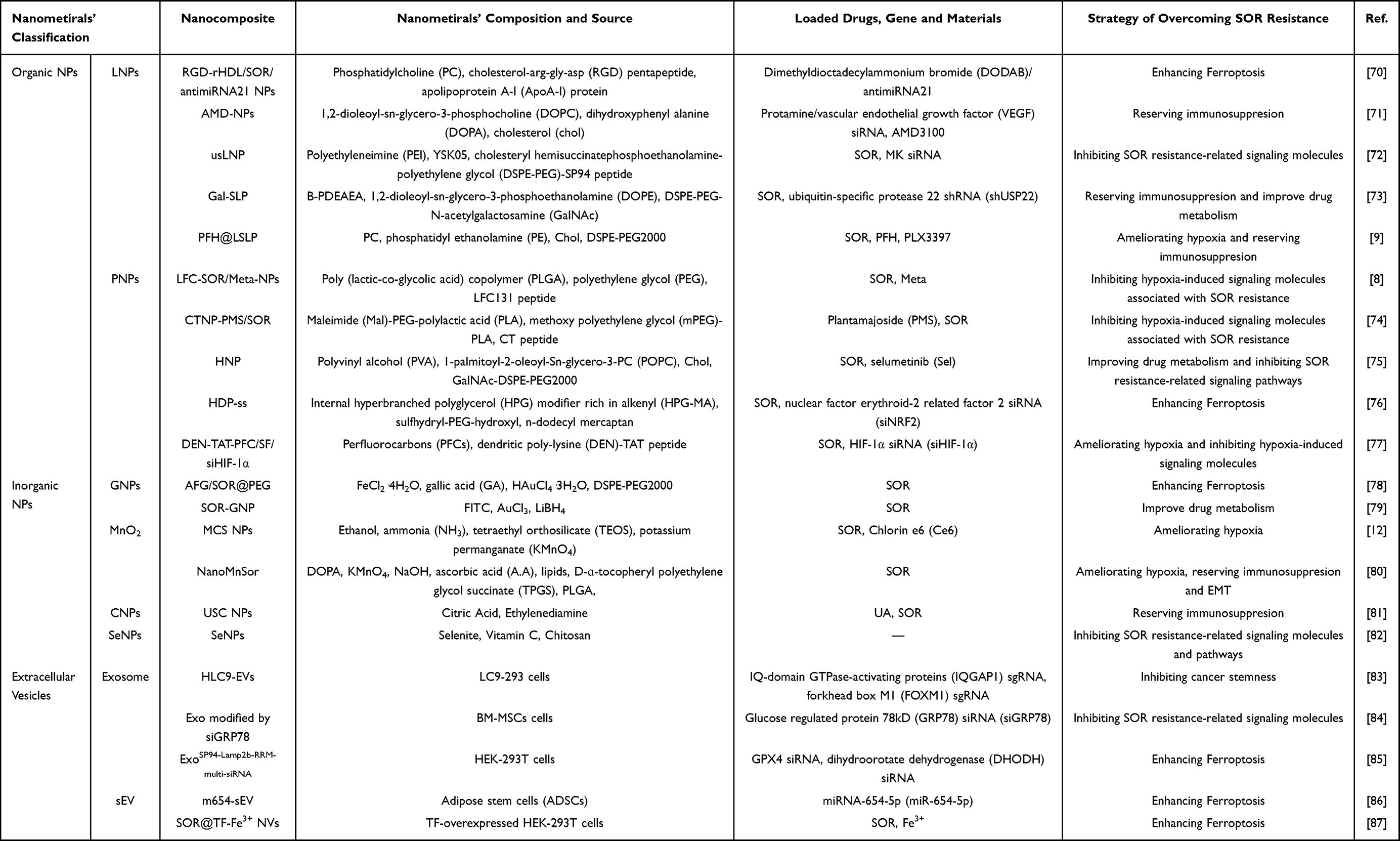 |
Table 1 Nanocomposites for Overcoming SOR Resistance |
Organic NPs
Organic NPs refer to NPs assembled from organic compounds such as small organic molecules and polymers. Organic NPs used to overcome SOR resistance mainly include lipid NPs (LNPs), polymer NPs (PNPs), and others.
LNPs
LNPs are lipid NPs (a subtype of lipid vesicles) with a homogeneous lipid core, which exhibit good biocompatibility because of their similar structure to the plasma membrane of biological cells and can be used as vector to deliver drugs, genes, or other functional materials to overcome SOR resistance.88,89 Furthermore, because the surface of LNPs is easy to modify, they can be loaded with specific proteins or ligands,90,91 which can increase the targeting ability of NPs to HCC and inhibit the expression of signaling molecules associated with SOR resistance.
LNPs are ideal vectors for gene delivery.92 Li et al70 condensed antimiRNA21 with dimethyldioctadecylammonium bromide (DODAB) to form the hydrophobic complex DODAB/antimiRNA21 by vortex and incubation methods, thereby enhancing the stability of antimiRNA21. Subsequently, PC, cholesterol-RGD pentapeptide and cholesteryl ester were dissolved in chloroform and mixed with SOR ethanol solution to form the oil phase. Then, the oil phase was evaporated under vacuum to obtain a lipid film, and the water phase (phosphate buffered saline) containing DODAB/antimiRNA21 complex was added to the lipid film to hydrate after thoroughly removing organic solvent. Finally, the final nanocomposite RGD-rHDL/So/antimiRNA21 was synthesized by co-incubation of the above products with ApoA-I protein by thin-film hydration method (Figure 6A). Among them, DODAB could firmly encapsulate antimiRNA21 in systemic circulation. ApoA-I and RGD can specifically bind to class B type 1 (SR-B1) and integrin αvβ3 overexpressed on the surface of HCC cells, respectively, which makes RGD-rHDL/So/antimiRNA21 have good targeting for delivering antimiRNA21 to HCC cells. These results illustrated RGD-rHDL/So/antimiRNA21 could stably and efficiently deliver antimiRNA21 to help it overcome SOR resistance.
 |
Figure 6 (A) Schematic diagram of RGD-rHDL/So/antimiRNA21 preparation process; (B) Schematic diagram of AMD NPs preparation process; (C) Schematic diagram of usLNPs preparation process; (D) Schematic diagram of Gal-SLPs preparation process. |
Liu et al71 mixed VEGF siRNA with protamine to prepare complex protamine/VEGF siRNA. Then, incorporating AMD3100 into protamine/VEGF siRNA formed smaller complexes protamine/VEGF siRNA/AMD3100. The surface of the protamine/VEGF siRNA/AMD3100 complex was coated with an anionic liposome composed of DOPC, DOPA, and chol to obtain nanocomposite AMD-NPs (Figure 6B). AMD3100, as a specific CXCR4 blocker, could both inhibit CXCR4 expression and serve as a ligand for the intracellular delivery of VEGF siRNA into HCC cells.93
Younis et al94 condensed PEI and midkine (MK) siRNA to form a polymer as the core of ultimate nanocomposite. Then, the researchers encapsulated this polymer core with the pH-sensitive cationic lipid YSK05 using the thin-film hydration method and loaded it with SOR to form a lipid nanocomposite.95 The surface of this nanocomposite was then modified with the SP94 peptide (SFSIIHTPILPL) to construct HCC-targeted nanocomposites SP94 NPs. Ultimately, the ultrasmall lipid nanocomposites usLNPs were synthesized using microfluidic technology, which allowed precise control over the particle size, resulting in an average diameter of 70–100 nm for the SP94 NPs72,96 (Figure 6C). HCC is characterized by a stroma-rich microenvironment with a dense ECM comprising collagen, hyaluronan, and fibroblasts, which acts as a physical barrier to nanocomposites with larger particle sizes.97 However, owing to their smaller size, usLNPs were successfully delivered into HCC tissues, reaching concentrations of approximately 15% injecting dose/g. Moreover, pharmacokinetic analyses demonstrated that usLNPs exhibited prolonged circulation times with minimal siRNA leakage. These findings confirmed that the accumulation of MK siRNA in HCC tissues is effectively facilitated by usLNPs, thus assisting in the overcoming SOR resistance.
Xu et al73 synthesized a polyplex through electrostatic interactions between cationic B-PDEAEA and anionic shUSP22. Then, this positively charged polyplex B-PDEAEA/shUSP22, along with SOR, was encapsulated within a fusogenic GalNAc-modified liposome composed of DOPE, cholesteryl hemisuccinate, and DSPE-PEG-GalNAc. This process resulted in the formation of the nanocomposite Gal-SLPs, which exhibited stealth properties, HCC-targeting ability, and membrane fusion characteristics (Figure 6D). Among them, B-PDEAEA, a ROS-responsive charge-reversal polymer, enabled Gal-SLPs to release drugs in response to specific stimuli that modify the surface charge.98 The GalNAc modification confers upon Gal-SLPs the capacity to target HCC by binding to asialoglycoprotein receptor (ASGPR) which is overexpressed on the surface of HCC cells.99 Gal-SLPs binding to ASGPR-induced ROS overproduction in Huh-7 cells, triggering shUSP22 release to overcome SOR resistance. LNPs also act as a versatile platform for the delivery of drugs and functional materials.
Wang et al9 prepared a lipid membrane incorporating PC, PE, Chol, DSPE-PEG2000, and LFC131 peptide through a rotary evaporation technique, then combined it with SOR, PFH, and PLX3397 via thin-film hydration to obtain PFH@LSLP (Figure 7A). Among them, LFC131 peptide, which could recognize and bind to the CXCR4 overexpressed on HCC cells, was decorated on the liposomal surface to endow the nanocomposites with HCC targeting capability. In vivo fluorescent images showed that fluorescence-labeled PFH@LSLP and PFH@LSL (without PLX3397) exhibited significantly higher accumulation in tumors compared with other organs and control groups devoid of the LFC131 peptide (Figure 7B). This finding substantiated the efficient delivery of SOR, PFH, and PLX3397 to HCC tissues by PFH@LSLP. Furthermore, the PFH encapsulated within the nanocomposite acted as an O2 reservoir, effectively mitigating tumor hypoxia and counteracting hypoxia-induced SOR resistance. O2 concentration curves revealed that the PFH-containing nanocomposite rapidly elevated dissolved O2 levels to approximately 8 mg/L within minutes, much more than that of the LS (without PFH) (Figure 7C). The release of O2 subsequently disrupted the nanocomposite’s integrity, facilitating the concurrent release of encapsulated SOR and PLX3397 (Figure 7D). Notably, PLX3397,100 a colony stimulating factor 1 (CSF1) immunosuppressant, further potentiated the reversal of immunosuppression-induced SOR resistance by activating tumor immunity.
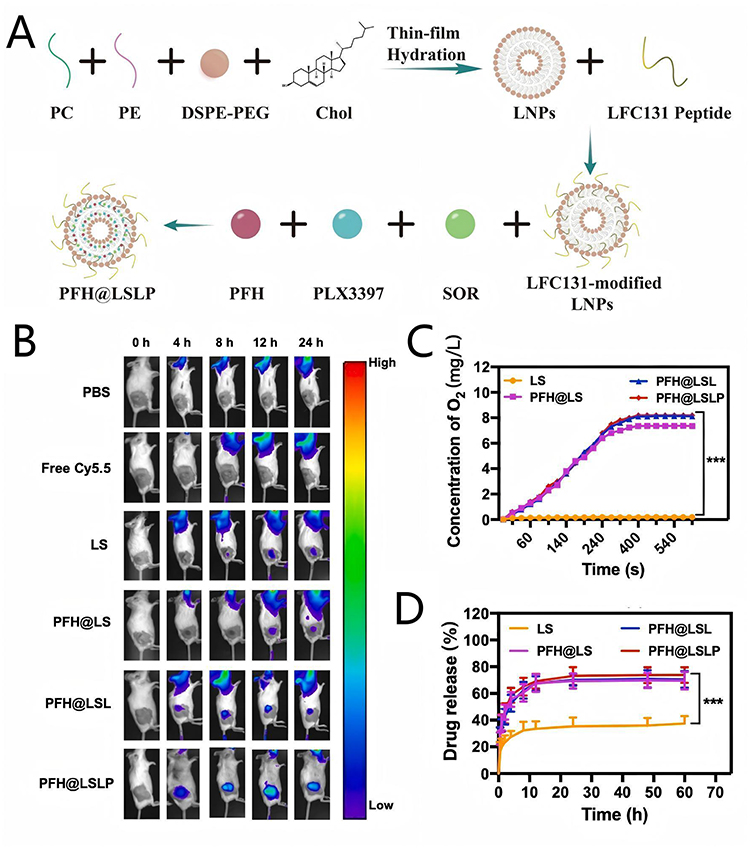 |
Figure 7 (A) Schematic diagram of PFH@LSLP preparation process; (B) In vivo fluorescence distribution of H22 tumor-bearing BALB/c mice after in vivo injection of various Cy5.5-labeled NPs at different time; (C) The O2 production curves after different treatments; (D) In vitro release rate of SOR in hypoxic conditions treat with different groups. Adapted from Wang Y, Wang Z, Jia F, et al. CXCR4-guided liposomes regulating hypoxic and immunosuppressive microenvironment for sorafenib-resistant tumor treatment. Bioact Mater. 2022;17:147–161. https://creativecommons.org/licenses/by-nc-nd/4.0/.9 |
Rent groups. Adapted with permission from ref.9 Copyright 2022 KeAi Communications Co.
Polymer NPs
PNPs generally refer to hydrophobic polymers-based NPs.101 Common polymers mainly include PEI, PLGA, PEG, and others. In contrast to alternative nano-delivery vectors, PNPs have the unique advantage: the precise tailoring of polymer composition to specific requirements, alongside modulation of size, shape, surface charge, and functional groups. This customizability renders them superior vectors for efficient drug and gene delivery, facilitating the overcoming of SOR challenges.102
The most prevalent functional modification is imparting HCC targeting to the nanocomposite by incorporating a targeting peptide.103,104 Zheng et al8 synthesized the copolymer PLGA-PEG-COOH by conjugating COOH-PEG-NH2 to COOH-PLGA. Subsequently, PLGA-PEG-COOH, SOR, and Meta were dissolved and stirred in tetrahydrofuran (TFH) to form the oil phase (SOR/Meta-NPs). This oil phase was then mixed with the LFC131 peptide at room temperature to obtain the final nanocomposite LFC-SOR/Meta-NPs (Figure 8A). The pharmacokinetic analysis revealed that free drug was rapidly cleared from the bloodstream, whereas LFC-SOR/Meta-NPs demonstrated prolonged circulation time, attributed to their resistance to degradation in the circulatory system. Furthermore, LFC-SOR/Meta-NPs exhibited significantly enhanced SOR/Meta delivery to tumor tissues compared with free drugs, which is facilitated by the binding of the LFC131 peptide to CXCR4 overexpressed on HCC cells. These findings suggested that LFC131-modified nanocomposite can effectively enhance tumor uptake of SOR and Meta, thereby aiding in the overcoming of SOR resistance.
 |
Figure 8 (A) Schematic diagram of LFC-SOR/Meta-NPs preparation process; (B) Schematic diagram of CTNP-PMS/SOR preparation process; (C) Schematic diagram of HNP preparation process. |
Zan et al74 dissolved PMS, SOR, Mal-PEG-PLA, and MPEG-PLA in dichloromethane and subsequently added sodium cholate solution. The mixture was then sonicated to form an oil-in-water emulsion, which was stirred and incubated to obtain drug-loaded NPs NP-PMS/SOR. Then, NP-PMS/SOR and CT peptide were resuspended in distilled water, and the resultant solution was subjected to high-speed centrifugation to obtain the final composite CTNP-PMS/SOR (Figure 8B). The CT peptide was prepared by conjugating tumor-homing peptide (CVNHPAFAC) to the fourth lysine of TAT via a pH-sensitive hydrazone bond, which could confer HCC targeting to CTNP-PMS/SOR and enhance the internalization of SOR and PMS by HCC cells and facilitates the overcoming of SOR resistance.
The addition of ligands on the surface of NPs that specifically bind to HCC cell surface receptors also endows NPs with targeting delivery to HCC. Farinha et al75 used PLGA and PVA to construct PLGA NPs as the core, and then loaded Sel within PLGA NPs. The surface of PLGA NPs was subsequently coated with a lipid membrane composed of Chol and POPC by thin-film hydration method, with GalNAc-DSPE-PEG2000 being incorporated prior to membrane formation, resulting in the final nanocomposite HNP (Figure 8C). HNP possessed spherical structure and external bilayer architecture, confirming the successful encapsulation of the lipid layer. The loading of GalNAc enabled efficient and specific targeting of HNP to HCC cells, facilitating Sel to inhibit the expression of pERK and thereby overcoming SOR resistance.105 PNPs can also serve as an efficient gene delivery vector. Tong et al76 introduced vinyl groups through a trans-esterification reaction between glycidyl methacrylate and HPG, subsequently synthesizing an alkenyl-rich internal HPG modifier (HPG-MA). Under nitrogen protection, HPG-MA was then reacted with sulfhydryl-PEG-hydroxyl and n-dodecyl mercaptan via UV light treatment, yielding the complex HDP through a thiol-ene click-chemistry approach.106 Subsequently, researchers loaded SOR and siNRF2 onto HDP using the thiol-ene click-chemistry method to construct the nanocomposite HDP-ss (Figure 9A). HDP-ss exhibited a size of approximately 200 nm, and its morphology and size remained relatively stable in serum for up to 12 hours. Confocal laser scanning microscope (CLSM) images revealed that compared with the control group, HCC cells treated with HDP-ss exhibited significantly more red fluorescence-labeled siNRF2, indicating that HDP enhances drug uptake through cell membrane endocytosis (Figure 9B). Furthermore, HDP-ss demonstrated sustained release and pH responsiveness, with siNRF2 and SOR being rapidly and smoothly released in the acidic environment of HCC cells (Figure 9C). These findings suggested that HDP-ss could efficiently deliver siNRF2 to help overcome SOR resistance mediated by NRF2 overexpression.
 |
Figure 9 (A) Schematic diagram of HDP-ss preparation process; (B) CLSM images of HCC cells treated by different group; (C) Release profiles of SOR from HDP at different pH. Adapted with permission from Tong R, Feng X, Sun J, et al. Co-delivery of siNRF2 and sorafenib by a “click” dual functioned hyperbranched nanocarrier for synergistically inducing ferroptosis in hepatocellular carcinoma. Small. 2024;20(21):e2307273.76 Copyright 2024 Wiley-VCH Verlag. (D) Schematic diagram of DEN-TAT-PFC/SF/siHIF-1α preparation process; (E) The corresponding O2 concentrations of various solutions using a portable dissolved oxygen meter. Adapted from Wan Y, Yang Y, Lai Q, Wang W, Wu M, Feng S. Fluorinated cell-penetrating peptide for co-delivering siHIF-1α and sorafenib to enhance in vitro anti-tumor efficacy. Pharmaceutics. 2023;15(12):2789. https://creativecommons.org/licenses/by/4.0/.77 Copyright 2023 MDPI (Basel, Switzerland). |
In addition, PNPs demonstrate efficient delivery of functional materials to HCC cells. Wan et al77 added triethylamine and pentadecafluorooctanoyl chloride to 2-(pyridinedithio)ethylamine hydrochloride dissolved in CH2Cl2 solution to synthesize the complex PFCs.107 PFCs not only encapsulate SOR through fluorine atom interactions but also possess unique O2 storage and transport capabilities, aiding in the amelioration of tumor hypoxia.108,109 Then, PFCs were linked to a DEN conjugated cell-penetrating peptide (TAT) through a disulfide bond, forming a fluorinated peptide (DEN-TAT-PFC). The final nanocomposite, DEN-TAT-PFC/SF/siHIF-1α, was obtained by mixing and standing DEN-TAT-PFC, SOR, and siHIF1-α (Figure 9D), which exhibited a spherical-like structure and a narrow size distribution with an average diameter of about 186 nm. As a nanocarrier, DEN-TAT-PFC exhibited excellent polar and non-polar phase separation properties, facilitating them through cellular lipid bilayers. In the presence of GSH, SOR was rapidly released (~68.43% at 1 h) from DEN-TAT-PFC/SF/siHIF-1α, which more than that in the absence of GSH (~28.1% at 10 h), indicating the GSH-triggered drug release characteristic of DEN-TAT-PFC/SF/siHIF-1α. The dissolved oxygen concentration of DEN-TAT-PFC after O2 saturation treatment increased by 5.70 mg/L within 120 s, while that of the DEN-TAT solution only increased by 2.73 mg/L (Figure 9E), indicating that DEN-TAT-PFC could be applied as an effective O2 delivery vehicle and also could slowly release O2 and could be utilized to assist in alleviating the tumor hypoxic microenvironment. Furthermore, siHIF-1α could be protected during transport by DEN-TAT-PFC/SF/siHIF-1α and internalized into HCC cells. Consequently, DEN-TAT-PFC/SF/siHIF-1α has the potential to overcome hypoxia-induced SOR resistance by delivering both O2 and siHIF-1α.
Inorganic NPs
Inorganic NPs, such as gold NPs (GNPs), manganese dioxide (MnO2), carbon NPs (CNPs), and selenium NPs (SeNPs), possess unique physicochemical properties, structural features, functional diversity, excellent biocompatibility, and the capability to traverse biological barriers at both cellular and tissue levels.110,111 Consequently, these inorganic NPs are extensively utilized in overcoming SOR resistance in cancer treatment.
GNPs
GNPs have various advantages such as a small volume ratio, large surface area, high biosafety, and easy surface modification, which can effectively deliver drugs/genes to overcome SOR resistance.112,113 Furthermore, GNPs exhibit excellent photothermal properties upon exposure to near-infrared laser light, functioning as a photosensitizer in photothermal therapy (PTT) to augment the lethal efficacy of the nanocomposite against SOR-resistant HCC.112,114
Wang et al78 prepared the complex Au/Fe-GA (AFG) by introducing aqueous solutions of FeCl2·4H2O and GA in the aqueous solution containing HAuCl4·3H2O and SH-PEG-COOH, leveraging the robust coordination interactions between the polyphenol groups of GA and Fe2+. Then, the dimethyl sulfoxide dissolved with SOR was added to the aqueous solution containing DSPE-PEG2000. Finally, AFG solution was added to the SOR solution to obtain nanocomposite AFG/SFB@PEG by stirring, condensing, and purifying (Figure 10A). Electron spin resonance (ESR) spectra presented the typical 1:2:2:1 characteristic peak of ·OH after the treatment of AFG/SFB@PEG and H2O2, proving that Fe2+ released from intracellular AFG/SFB@PEG could react with excessive H2O2 in HCC cells to generate highly cytotoxic ·OH (Figure 10B). Then, intracellular ·OH prompted the generation of LPO, augmenting the ability of AFG/SFB@PEG to overcome SOR resistance mediated by ferroptosis inhibition. Owing to the strong absorption throughout the visible and NIR region, AFG/SFB@PEG nanoreactors could act as a photothermal agent, and the incorporation of Au endowed AFG/SFB@PEG with exceptional photothermal conversion efficiency, enabling enhanced eradication of SOR-resistant HCC through PTT (Figure 10C and D).
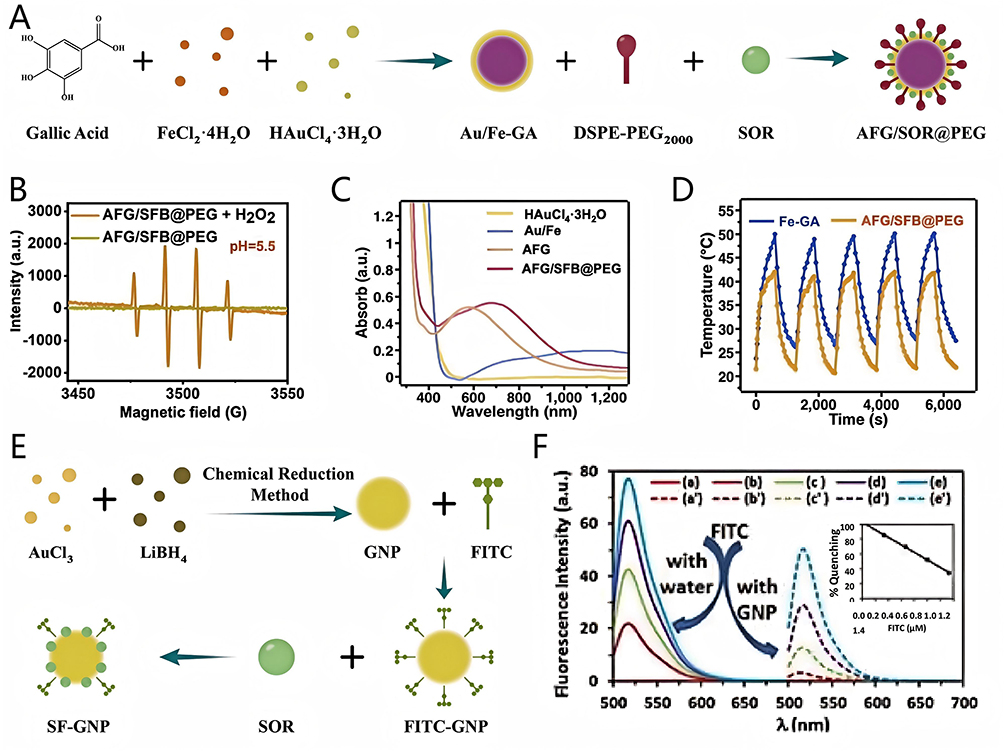 |
Figure 10 (A) Schematic diagram of AFG/SFB@PEG preparation process; (B) ESR spectra of AFG/SFB@PEG; (C) UV-Vis-NIR spectra of different group; (D) Photothermal cycles of AFG and AFG/SFB@PEG in water. Reprinted from Acta Biomaterialia: 159, Wang X, Zhao L, Wang C, et al. Potent nanoreactor-mediated ferroptosis-based strategy for the reversal of cancer chemoresistance to Sorafenib. 237–246, Copyright 2023 with permission from Elsevier.78 (E) Schematic diagram of SOR-GNP preparation process; (F) Quantification of FRET process between FITC and GNP at various concentration. Adapted from Vishwakarma S, Sharmila P, Bardia A, et al. Use of biocompatible sorafenib-gold nanoconjugates for reversal of drug resistance in human hepatoblatoma cells. Sci Rep. 2017;7(1):8539. https://creativecommons.org/licenses/by/4.0/.79 |
Vishwakarma et al79 used AuCl3 and LiBH4 to construct GNPs by chemical reduction. Then, FITC was added to a colloidal suspension of GNP to prepare FITC-functionalized GNP (FITC-GNP). Finally, SOR was loaded on GNP by self-assembly method to construct nanocomposite SF-GNP (Figure 10E). The characteristic surface plasmon resonance (SPR) peak of SF-GNP was observed at 524 nm in UV–Vis spectra. SF-GNP could achieve 80% SOR loading by quantification of fluorescence resonance energy transfer (FRET) (Figure 10F). This high drug loading capacity enables GNPs to further overcome SOR resistance induced by insufficient effective drug concentrations.
MnO2 NPs
MnO2 NPs possess the advantages of large specific surface areas, low toxicity, and excellent biodegradable metabolism.38 MnO2 NPs could catalyze excessive H2O2 in TME to produce O2 to relieve resistance caused by hypoxia.115,116 Moreover, Mn2+ produced by the degradation of MnO2 can be involved in regulating the immune response and overcoming immunosuppression-induced SOR resistance.117,118 Therefore, MnO2 NPs have emerged as a promising NPs for overcoming SOR resistance.
Xu et al12 synthesized solid SiO2 NPs (sSiO2 NPs) through a mixture of ethanol, ammonia, and tetraethyl orthosilicate (TEOS) by oil bath method. Subsequently, a KMnO4 solution, dissolved in deionized water, was added dropwise into the aqueous solution of sSiO2 NPs by ultrasonication and stirring to obtain sSiO2@MnO2. Then, the hollow MnO2 NPs H-MnO2 were prepared after dissolving the sSiO2@MnO2 with sodium bicarbonate (Na2CO3) solution. Finally, the mixed solution of SOR and Ce6 was added to H-MnO2 solution to construct the ultimate nanocomposite MSC NPs by stirring and centrifugation (Figure 11A). The specific surface area and average pore size of H-MnO2 are 276 m2/g and 4 nm, respectively, N2 adsorption/desorption isotherms and pore-size distribution curve, proved H-MnO2 had mesoporous structure (Figure 11B and C), which was beneficial for loading more SOR and Ce6 on MCS NPs. Additionally, MnO2 NPs could react with H2O2 in an acidic environment to produce O2 compared with the neutral environment.119 Responsive release is stimuli-triggered drug delivery. The release rate (58.24%) of SOR in MCS NPs at pH 6.5 after 24 h incubation with H2O2 is higher than at pH 7.4 without H2O2 (19.19%), suggesting responsive release capacity of MCS NPs under acidic conditions with H2O2 (Figure 11D). Furthermore, Ce6 has the ability of PTT under laser irradiation, and could help further kill SOR-resistant HCC. These results indicated that MCS NPs had high drug loading rate, responsive release capacity, and the capacity of producing endogenous O2, which promotes MCS NPs to overcome SOR resistance.
 |
Figure 11 (A) Schematic diagram of MCS NPs preparation process; (B) N2 adsorption/desorption isotherms of the H-MnO2; (C) The pore-size distribution curve of the H-MnO2; (D) The release rate of SOR from MCS NPs. Adapted with permission from Xu W, Yang M, Zhang W, Jia W, Zhang H, Zhang Y. Tumor microenvironment responsive nano-platform for overcoming sorafenib resistance of hepatocellular carcinoma. Mater Today Bio. 2024;24:100902.12 Copyright 2017 Elsevier. (E) Schematic diagram of NanoMnSor preparation process. |
Chang et al80 dispersed DOPA and KMnO4 into an oil phase to prepare the Mn7+-loaded microemulsions. Meanwhile, another microemulsion acting as a reducing agent was prepared by adding the mixture of A.A and NaOH into a separate oil phase. Then, the two microemulsions were mixed to reduce Mn7+ to Mn2+, and a stable DOPA-coated MnO2 core (DOPA-MnO2) was obtained by the binding of Mn2+ to phospholipid phosphate groups. Importantly, the MnO2 core exhibits TME-responsive decomposition: under acidic pH conditions in HCC tissues, MnO2 NPs release Mn2+ that act as T1-weighted MRI contrast agents, generating positive contrast enhancement for real-time tumor imaging. Subsequently, free lipids (consisted of Mal-DSPE-PEG2000, DSPE-PEG2000, DOPC, and chol), PLGA, TPGS, and DOPA-MnO2 were mixed construct NanoMn NPs by self-assembly method. Finally, SP94 peptide and SOR were loaded the surface and interior of the NanoMn NPs, respectively, to obtain the nanocomposite NanoMnSor (Figure 11E). NanoMnSor has dual functions of diagnosis and treatment, and it can be used for MRI-guided drug delivery and hypoxia modulation. Like MCS NPs, NanoMnSor could efficiently generate O2 and release SOR in TME. The SP94 peptide conferred active targeting to orthotopic HCA-1 HCC models, while the Mn2+-based MRI contrast allowed non-invasive monitoring of nanocomposite accumulation and hypoxia relief efficiency. These results demonstrate that NanoMnSor integrates diagnostic imaging with targeting therapy, effectively overcoming hypoxia-induced SOR resistance through precision feedback.
CNPs
CNPs, particularly carbon dots (CDs), represent a versatile class of theranostic nanomaterials for precision medicine. CDs inherently exhibit strong fluorescence for real-time imaging, and when manganese or gadolinium elements are doped into CDs, they have the dual-modal imaging function of MRI/fluorescence, thereby achieving synchronous monitoring of tumor localization and the treatment process.120
Lai et al81 used citric acid and ethylenediamine to construct CDs solution by hydrothermal method, meanwhile US NPs were synthesized using SOR and ursolic acid (UA) by self-assembly method. Then, nanocomposite USC NPs were constructed with CDs solution and US NPs by self-assembly method (Figure 12A). CLSM images showed that compared with the L-02 cells, more CDs were observed within HepG2 cells and SMMC 7721 cells after the treatment of CDs and USC NPs (Figure 12B), confirming the targeting accumulation and efficient cellular penetration of CDs and USC NPs in HCC cells. Intracellular USC NPs then could responsively release SOR and CDs in an acidic environment of TME, and Figure 12C reveals that more than 40% of SOR and nearly 26% of CDs were released from USC NPs at the pH=5.5 within 48 h, which might be beneficial for the efficacy (Figure 12C). Furthermore, owing to the better enhanced permeability and retention (EPR) effect of USC NPs, in vivo fluorescence quantitative curve showed that USC NPs could more be accumulated in tumor (Figure 12D). The above experiments demonstrated that USC NPs can effectively deliver SOR, UA, and CDs into HCC cells, thereby contributing to the overcoming of SOR resistance.
 |
Figure 12 (A) Schematic diagram of USC NPs preparation process; (B) CLSM image of the intracellular distribution of CDs and USC NPs equivalent concentration of CDs in three cell lines at 4 h post-incubation; (C) Cumulative drug release of NPs under different pH conditions; (D) The corresponding fluorescence quantification of different group in each organ. Reprinted from Journal of Colloid and Interface Science: 635, Lai C, Zhang B, Li D, et al. Rational design of a minimum nanoplatform for maximizing therapeutic potency: three birds with one stone. 441–455, Copyright 2023 with permission from Elsevier.81 |
SeNPs
SeNPs have high biocompatibility, low toxicity, in vivo degradability, significant X-ray radiosensitization effect, and pleiotropy, which can induce tumor cell apoptosis without affecting normal tissue cells.121 Furthermore, SeNps not only can target similar cellular pathways as SOR, but also target signaling molecules and pathways associated with SOR resistance.122 Therefore, combinatorial therapy incorporating SeNPs represents an efficacious strategy to reserve SOR resistance.
Tohada M. AL-Noshokaty et al82 dissolved H2SeO3 and VitC in double-distilled water, chitosan in acetic acid to construct SeNPs by stirring, dialysis, and freeze drying.123 Transmission electron microscopy (TEM) image showed that SeNPs was well-dispersed and spherical shaped with the size of 60 nm (Figure 13A). Researchers induced HCC in mice using thioacetamide (TAA). Owing to the antioxidant properties of Se, SeNPs can mitigate the adverse effects induced by SOR and TAA, thereby normalizing levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), and other liver function markers (Figure 13B–E), indicating that SeNPs can restore liver histology and function, alleviating SOR-related side effects, and potentially overcoming SOR resistance.
 |
Figure 13 (A) TEM images of SeNPs; (B–E) Effect of Se, SeNps, SOR and their combinations on ALT, AST, TP, GGT in mice. Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001) and “ns” represents no significance (P ≥ 0.05). Reprinted from Life Sciences: 303, Al-Noshokaty T, Mesbah N, Abo-Elmatty D, Abulsoud A, Abdel-Hamed A. Selenium nanoparticles overcomes sorafenib resistance in thioacetamide induced hepatocellular carcinoma in rats by modulation of mTOR, NF-κB pathways and LncRNA-AF085935/GPC3 axis. 303:120675, Copyright 2022 with permission from Elsevier.82 |
Extracellular Vesicles
EVs are phospholipid bilayer-coated NPs, which secreted by cell and serve as crucial mediators for intercellular communication and the transport of functional biomolecules.124 Owing to their immunogenicity and excellent biocompatibility, they can serve as carriers to evade clearance by the monocyte-macrophage system. Additionally, their tissue penetration ability and endocytosis pathway facilitate the delivery of SOR and other genes/drugs to HCC cells, providing an ideal drug delivery vehicle for overcoming SOR resistance.125,126
Exosome
Exosomes (Exos), a subset of EVs, encapsulate cellular constituents such as proteins, DNA, and RNA from their parent cells. They exhibit similarities in size, shape, and structure to LNPs, yet possess more intricate phospholipid bilayers.127 Researchers have utilized multiple techniques, including transfection, incubation, sonication, centrifugation, and electroporation, to load therapeutic molecules and functional materials onto the surface or into the interior of Exos in order to enhance the pharmacokinetics of SOR, inhibit genes and signaling molecule associated with SOR resistance, and ultimately overcome SOR resistance.128–130
He et al83 produced HN3-LAMP2-AcGFP/Cas9-expressing EV donor LC9-293 cells using lipid transfection and subsequently prepared the corresponding exosomes HLC9-EVs by centrifugation. As IQGAP1 can promote Akt activation to impede SOR action, and FOXM1 supports β-catenin translocation to the nucleus in HCC cells to aid cancer stemness maintenance and SOR resistance,131,132 single guide RNA (sgRNA) sgIF, which inhibits both IQGAP1 and FOXM1 expressions were loaded on HLC9-EVs using electroporation to obtain sgRNA-loaded HLC9-EVs (Figure 14A). HLC9-EVs were characterized with TEM, which showed that HLC9-EVs were membrane-surrounded nanovesicles, round, and with particle sizes of 100 to 400 nm (Figure 14B). Owing to the loading of HN3 which specifically targets the overexpressed Glypican-3 (GPC3) in HCC cells,133,134 HLC9-EVs possessed the quicker targeting ability, compared with LC9-EVs (without HN3). Consistent with in vitro cellular internalization results, significant accumulation of HLC9-EVs was observed at specific tumor sites in vivo, indicating efficient targeting of HCC for sgIF delivery.
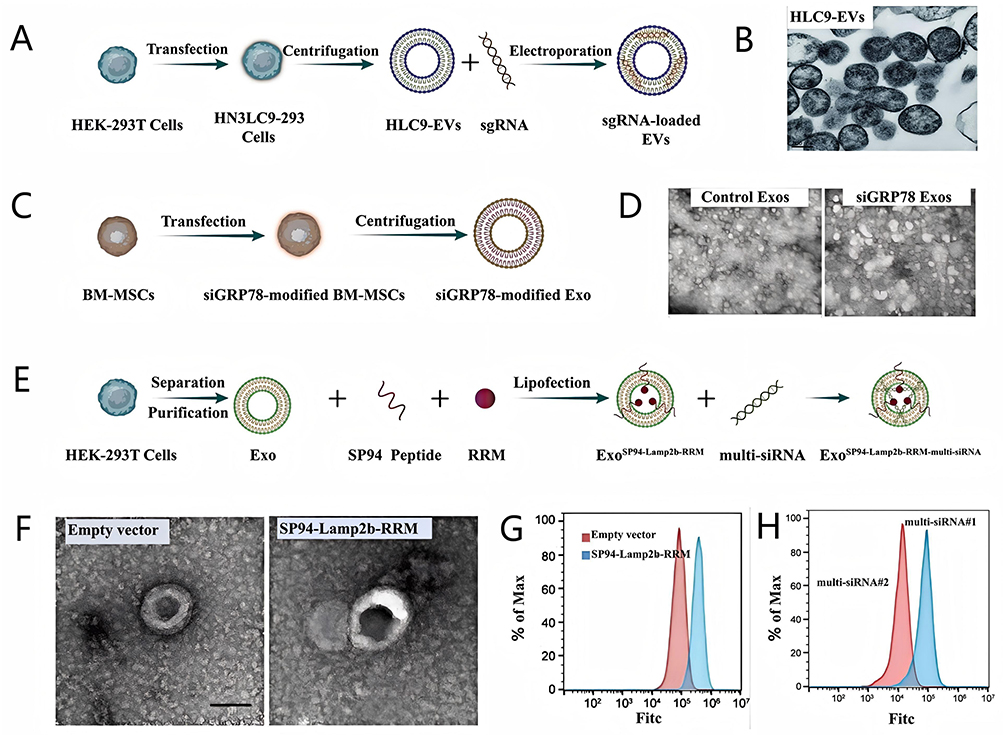 |
Figure 14 (A) Schematic diagram of sgRNA-loaded HLC9-EVs preparation process; (B) TEM image of HLC9-EVs. Adapted from He C, Jaffar Ali D, Qi Y, et al. Engineered extracellular vesicles mediated CRISPR-induced deficiency of IQGAP1/FOXM1 reverses sorafenib resistance in HCC by suppressing cancer stem cells. J Nanobiotechnology. 2023;21(1):154. https://creativecommons.org/licenses/by/4.0/.83 (C) Schematic diagram of siGRP78-modified Exo preparation process; (D) TEM image of BM-MSCs-derived Exos. Adapted from Li H, Yang C, Shi Y, Zhao L. Exosomes derived from siRNA against GRP78 modified bone-marrow-derived mesenchymal stem cells suppress Sorafenib resistance in hepatocellular carcinoma. J Nanobiotechnology. 2018;16(1):103. https://creativecommons.org/licenses/by/4.0/.84 (E) Schematic diagram of ExoSP94-Lamp2b-RRM-multi-siRNA#1 preparation process; (F) TEM image of empty Exo and ExoSP94-Lamp2b-RRM; (G) The binding ability of different Exo to HepG-2 cells; (H) The loading ability of different multi-siRNA to Exo. Adapted from Li X, Yu Q, Zhao R, et al. Designer exosomes for targeted delivery of a novel therapeutic cargo to enhance sorafenib-mediated ferroptosis in hepatocellular carcinoma. Front Oncol. 2022;12:898156. https://creativecommons.org/licenses/by/4.0/.85 |
Li et al84 isolated bone marrow mesenchymal stromal cells (BM-MSCs) and transfected these cells with siGRP78. Then, siGRP78-modified Exos were purified from the transfected BM-MSCs by high-speed centrifugation method (Figure 14C). TEM images presented that BM-MSCs-derived exosomes were about 50 to 130 nm in width and exhibited physical homogeneity (Figure 14D). Furthermore, the modification of siGRP78 did not influence the internalization of Exo, as CLSM images showed that siGRP78-modified Exos could be effectively internalized by SOR-resistant HepG2 (HepG2-SR) and PLC (PLC-SR) cells, respectively.
Li et al85 used an ingenious approach to increase the loading efficiency of miRNA by enabling the fusion of exosomal membrane proteins with specific RNA-binding protein. Specifically, the N-terminal RNA recognition motif (RRM) of U1-A was fused to the C-terminus of Lamp2b (an exosomal surface protein), and SP94 was fused to the N-terminus of Lamp2b, resulting in the generation of SP94-Lamp2b-RRM. Exos expressing SP94-Lamp2b-RRM (ExoSP94-Lamp2b-RRM) then were produced by isolating and purifying exosomes from HEK-293T cells transfected with the SP94-Lamp2b-RRM. Owing to the overexpression of GPX4 and DHODH can lead to SOR resistance by inhibiting ferroptosis,135 researchers designed a multi-siRNA#1 containing the sequences corresponding to GPX4 siRNA and DHODH siRNA and the sequence “AUUGCAC” was used as a linker to connect GPX4 siRNA and DHODH siRNA, and the multi-siRNA#2 as the control group did not contain “AUUGCAC” sequence. Finally, they used multi-siRNA#1 and ExoSP94-Lamp2b-RRM to construct the final nanocomposite ExoSP94-Lamp2b-RRM-multi-siRNA#1 by the high-binding ability of U1-A’s N-terminal RRM with the sequence “AUUGCAC” in multi-siRNA#1 (Figure 14E). TEM image revealed that ExoSP94-Lamp2b-RRM was a typical round-shaped vesicular morphology (Figure 14F). The loading of SP94 enhanced the HCC-targeting ability of the Exo, and flow cytometry analysis showed that ExoSP94-Lamp2b-RRM could bind strongly to HCC cells (Figure 14G). Furthermore, SP94-Lamp2b-RRM fusion protein more obviously promotes the exosomal loading of the “AUUGCAC” sequence linked to the multi-siRNA#1 compared with multi-siRNA#2 via RNA–protein interactions (Figure 14H). These functionalities are conducive to enhancing the efficacy of multi-siRNA#1 in overcoming SOR resistance.
Small Extracellular Vesicle
Small extracellular vesicles (sEVs), which are naturally occurring vesicles enveloped by a phospholipid bilayer membrane, can be secreted by a variety of cellular types, including tumor cells, endothelial cells, immune cells, and platelets. Like Exo, sEVs have been harnessed by researchers to protect encapsulated nucleic acids and proteins from RNase-mediated degradation. Furthermore, specific proteins with native structures have been engineered onto the sEV membrane to aid in overcoming SOR resistance.
Sun et al86 found miR-654-5p can target the ferroptosis negative regulator heat shock protein family B (small) member 1 (HSPB1) that can produce SOR resistance. ADSCs-derived sEVs were synthesized by ultracentrifugation and then the sEVs were used as delivery carriers to construct the miRNA-654-5p-loaded nanocomposite m654-sEVs by electroporation method for overcoming SOR resistance (Figure 15A). m654-sEVs and sEVs had similar spherical structures and their particle sizes were about 50–150 nm (Figure 15B). CLSM images revealed that fluorescence intensity of Cy3-labeled miR-654-5p inside SOR-resistant HUH7 and PLC/PRF/5 cells (HUH-7-R and PLC/PRF/5-R cells) increased with the concentration of m654-sEV (Figure 15C), indicating that miR-654-5p could be delivered to HCC cells through m654-sEV to aid in overcoming SOR resistance.
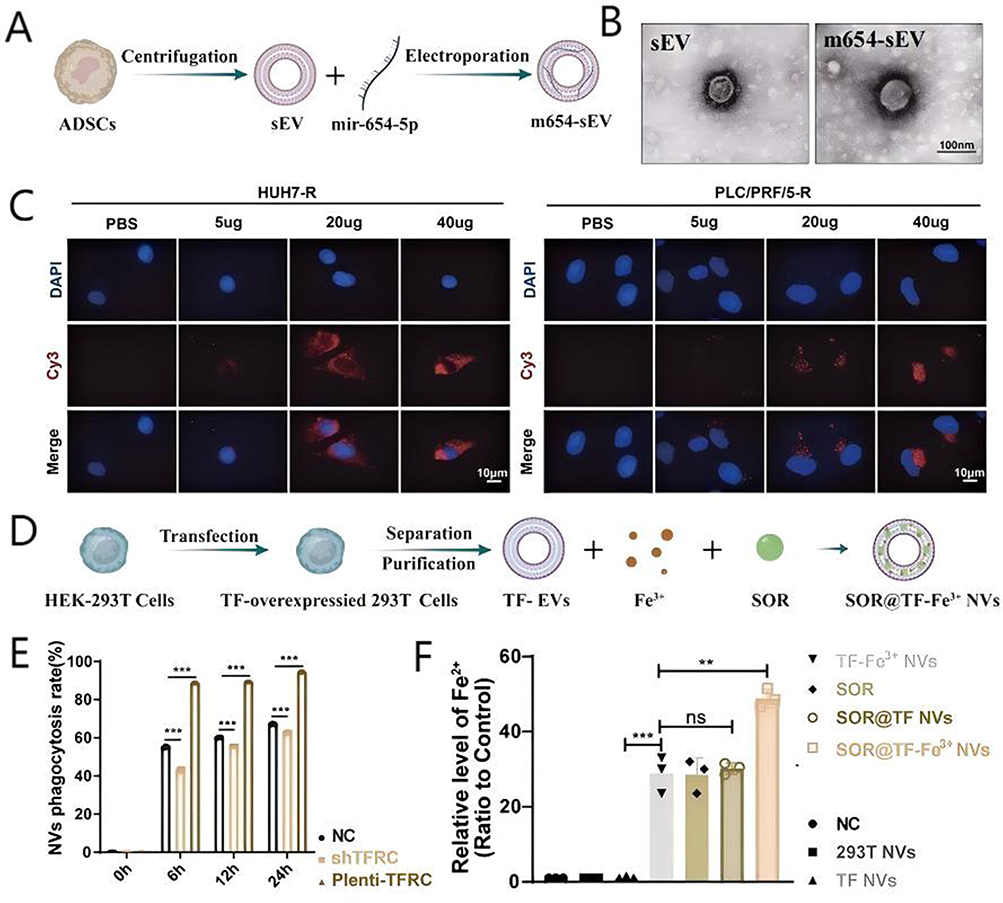 |
Figure 15 (A) Schematic diagram of m654-sEV preparation process; (B) TEM images of m654-sEV and sEV; (C) CLSM images HUH7-R and PLC/PRF/5-R cells after different treatment. Adapted from Jiao S, Qi L, Yanfang J, Zhihui C, Hui L, Huaiwen Z. Engineered small extracellular vesicles loaded with miR-654-5p promote ferroptosis by targeting HSPB1 to alleviate sorafenib resistance in hepatocellular carcinoma. Cell Death Discov. 2023;9(1):362. https://creativecommons.org/licenses/by/4.0/.86 (D) Schematic diagram of SOR@TF-Fe3+ NVs preparation process; (E) The binding rates of TF-Fe3+ NVs with different TFRC-expression HepG2 cells; (F) The Fe2+ content in cells treated with different groups. Adapted from Xiao Y, Xu Z, Cheng Y, et al. Fe-binding transferrin nanovesicles encapsulating sorafenib induce ferroptosis in hepatocellular carcinoma. Biomater Res. 2023;27(1):63. https://creativecommons.org/licenses/by/4.0/.87 Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001) and “ns” represents no significance (P ≥ 0.05). |
Xiao et al87 used lentivirus carrying a membrane-localized form of GFP-transferrin to infect HEK293T cells to make the cells overexpress TF. Then, the sEVs TF NVs were synthesized and purified from the HEK293T cells by differential centrifugation method. Finally, TF-Fe3+ NVs were prepared by co-incubating TF NVs and Fe3+ and further SOR was encapsulated on surface of TF-Fe3+ NVs to construct the ultimate nanocomposite SOR@TF-Fe3+ NVs through electro-transfection method (Figure 15D). Owing to the interaction of TF and TFRC, TF-expressing SOR@TF-Fe3+ NVs could be specifically phagocytosed (90%) by HepG2 cells with the exogenous expression of TFRC. Conversely, knockdown of TFRC reduced the phagocytosis efficiency of SOR@TF-Fe3+ NVs by HepG2 cells to 40% (Figure 15E). SOR@TF-Fe3+ NVs entered cells and released Fe3+, further the content of Fe2+ increased in HepG2 cells that more than free NVs and TF NVs groups (Figure 15F). In addition, nano-sized sEVs could passively accumulate in tumor, through EPR effect, indicating that SOR@TF-Fe3+ NVs could efficiently deliver Fe3+ to HCC, thereby aiding in the overcoming of SOR resistance mediated by ferroptosis inhibition.
Using NPs with unique structural characteristics and physicochemical properties as nanocarriers, nanocomposites are constructed to overcome SOR resistance by modifying NPs surfaces, loading drugs, genes, and others.136 For example, the phospholipid bilayer structure of LNPs and EVs makes nanocomposites have good biocompatibility. Therefore, LNPs and EVs usually are used to deliver gene for overcoming SOR resistance. The diverse raw materials for PNP preparation enable extensive functional modifications, facilitating the creation of highly HCC-targeted drug/gene delivery nanocomposites. Certain inorganic NPs have specific functions such as producing endogenous O2, heat or ROS after laser irradiation and inhibiting signaling molecules and pathways associated with SOR resistance. These functions allow inorganic NPs to play important roles in strategies of overcoming SOR resistance. Beyond conventional nanoformulations, emerging technologies offer novel strategies to overcome SOR resistance. Light-responsive systems enable precise, externally triggered drug release, improving tumor targeting and efficacy. Biomimetic nanocarriers enhance systemic circulation and biocompatibility for prolonged action. Additionally, gene co-delivery platforms enable synergistic therapy by combining SOR with genetic material to target critical resistance pathways. These advancements represent significant progress in developing next-generation nanotherapeutics for HCC.137–139 Hereby, the selection of NPs for constructing nanocomposites is of great importance in overcoming SOR resistance.
Application of Nanocomposite in Overcoming HCC SOR Resistance
In recent years, researchers had constructed nanocomposites utilizing NPs to overcome SOR resistance through the remodeling of the TME, restoration of normal epigenetic regulation, enhancement of drug metabolism, and inhibition of aberrant signaling molecules or pathways.
Remodelling TME
TME plays a crucial role in SOR resistance, enabling HCC cells to evade the cytotoxic effects of SOR. Remodeling TME by using nanocomposites is an effective method to overcome SOR resistance. Researchers used NPs as carriers to construct various nanocomposites with different functions, which reversed SOR resistance by ameliorating hypoxia, promoting ferroptosis, enhancing immunity, and inhibiting HCC cancer stemness. This section provides an overview of recent advancements in the utilization of nanocomplexes for TME remodeling to overcome SOR resistance.
Ameliorate Hypoxia and Inhibit Signaling Pathways Related to SOR Resistance Caused by Hypoxia
Hypoxia, or the elevated expression of hypoxia-induced signaling molecules and pathways such as HIF-1α and SDF-1/CXCR4 pathway, can lead to SOR resistance. Currently, there are three primary methods are employed by nanocomposites to overcome SOR resistance mediated by hypoxia: 1) generating endogenous O2 by reacting with H2O2 in tumor tissue; 2) delivering exogenous O2 by loading materials with oxygen-carrying function such as PFH; 3) delivering drugs or genes to inhibit hypoxia-induced signaling molecules and pathways associated with SOR resistance.
Xu et al12 constructed the nanocomposite MCS NPs which could efficiently deliver SOR and Ce6 to HCC cells and ameliorate hypoxia by generating endogenous O2, and further reduce HIF-1α expression in SOR-resistant HCC cells (Figure 16A and B). Researchers established SOR-resistant HCC cell lines (SMMC7721/S) to evaluate the efficacy of MCS NPs. The half-maximal inhibitory concentration (IC50) of SOR for SMMC7721/S cells was 18.42 μg/mL, which much higher than that of SMMC7721 cells (2.53 μg/mL) (Figure 16C), proving that SMMC7721/S cells have developed SOR resistance. MCS NPs could significantly inhibit the cell viability of SMMC7721/S cells compared with SOR treatment group, which was attributed to the amelioration of hypoxia. Moreover, the loading of Ce6 endowed MCS NPs with the ability of PDT and PTT which could more significantly kill SMMC7721/S cells under laser irradiation. In vivo experiments obtained the similar results that the tumor volume of SMMC7721-S cells hormonal nude mice increased to 900 mm2 after SOR alone treatment, while MCS NPs controlled the tumor volume to 500 mm2 by ameliorating hypoxia. Moreover, the tumor could be almost eliminated by the combination of MCS NPs with laser irradiation. NanoMnSor with MnO2 as the core constructed by Chang et al80 also could reverse hypoxia-induced SOR resistance by generating endogenous O2 for ameliorating hypoxic TME, which is similar to MCS NPs.
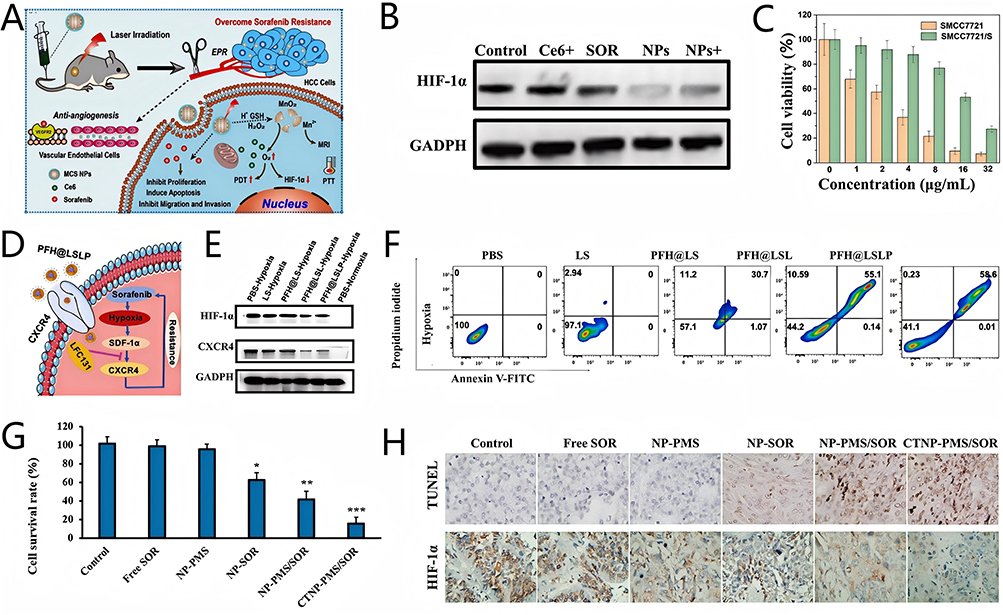 |
Figure 16 (A) Anticancer schematic illustration of TME-responsive MCS NPs; (B) The HIF-1α expression level after different treatment; (C) Cell survival rate of SMMC7721 and SMMC7721/S cells incubated by various concentrations of SOR. Adapted with permission from Xu W, Yang M, Zhang W, Jia W, Zhang H, Zhang Y. Tumor microenvironment responsive nano-platform for overcoming sorafenib resistance of hepatocellular carcinoma. Mater Today Bio. 2024;24:100902.12 Copyright 2017 Elsevier. (D) Schematic illustration of PFH@LSLP overcoming resistance to SOR through the SDF-1α/CXCR4 pathway under hypoxic conditions; (E) The HIF-1α and CXCR4 expression levels after different treatment; (F) Cell apoptosis rates of SOR-resistant HCC cells treated with different groups. Adapted from Wang Y, Wang Z, Jia F, et al. CXCR4-guided liposomes regulating hypoxic and immunosuppressive microenvironment for sorafenib-resistant tumor treatment. Bioact Mater. 2022;17:147–161. https://creativecommons.org/licenses/by/-nc-nd/4.0/.9 Copyright 2022 KeAi Communications Co. (G) Cell survival rate of SOR-resistant HCC cells treated with different groups; (H) Cell apoptosis and HIF-1α expression in SOR-resistant HepG2 cells (HepG2/SOR cells) tumor tissues after different treatment. Adapted from Wan Y, Yang Y, Lai Q, Wang W, Wu M, Feng S. Fluorinated cell-penetrating peptide for co-delivering siHIF-1α and sorafenib to enhance in vitro anti-tumor efficacy. Pharmaceutics. 2023;15(12):2789. https://creativecommons.org/licenses/by/4.0/.77 |
In addition, exogenous O2 can be delivered by loading materials with O2 carrying function to ameliorate hypoxia. Wang et al9 constructed a multifunctional nanocomposite PFH@LSL, which could ameliorate hypoxia by the O2 carrying capacity of PFH, while the encapsulated LFC131 peptide could inhibit HIF-1α expression (Figure 16D and E). Then, researchers used HCC cells under hypoxia conditions to simulate SOR-resistant HCC cells. FCM analysis showed that the percentage of apoptotic and necrosis region after incubation with PFH@LSL was up to 55.2%, which was higher than that of LS (0%) and PFH@LS (31.1%) treated SOR-resistant HCC cells (Figure 16F), indicating that PFH@LSL could reserve hypoxia-induced SOR resistance through the dual effects of PFH and LFC131.
The nanocomposite DEN-TAT-PFC/SOR/siHIF-1α constructed by Wan et al77 could efficiently deliver exogenous O2 and siHIF-1α which could reduce HIF-1α expression in HepG2-R cells. Then, researchers evaluated the anti-angiogenic capacity of different treatment groups by observing the tube formation of endothelial cells (HUVEC) in conditioned medium derived from SOR-resistant HCC cells. The results revealed that the DEN-TAT-PFC/SF/siHIF-1α treatment group has few microvascular networks and shortest HUVEC alignments compared with SOR alone treatment, indicating DEN-TAT-PFC/SF/siHIF-1α could restore the anti-angiogenic capacity of SOR in SOR-resistant HCC.
Other researchers used nanocomposites to deliver drugs or genes for inhibiting hypoxia-induced signaling molecules and pathways associated with SOR resistance. Zheng et al8 constructed the nanocomposite LFC-SOR/Meta-NPs with the ability of nice drug encapsulation, controlled drug release profile, persistent blood circulation, LFC131-receptor mediated recognition, and preferential accumulation in tumors, which could specifically and effectively co-deliver SOR and Meta into CXCR4-overexpressing HCC cells. Intracellular Meta could further inhibit the overexpression of CXCR4 related to SOR resistance in SOR-resistant SMMC-7721 (SMMC-7721-R) cells,140 and cell proliferation assays revealed a significant increase in the apoptosis rate of SMMC-7721-R cells to 42.56% following treatment with LFC-SOR/Meta-NPs, compared with the SOR group (only 9.87%).
Zan et al74 constructed the nanocomposite CTNP-PMS/SOR to efficiently deliver PMS to inhibit HIF-1α expression in drug-resistant cancer cells.141 Cell proliferation assays revealed that the IC50 value of CTNP-PMS/SOR to HepG2/SOR cells was 29.46 ng/mL, which is much lower than SOR treatment group (261.37 ng/mL) (Figure 16G). Similar results were obtained in vivo anti-tumor experiments. Immunostaining images showed that CTNP-PMS/SOR significantly down-regulated HIF-1α expression and promoted apoptosis in the tumor tissues of HepG2/SOR cancer-bearing mice (Figure 16H). These results proved that CTNP-PMS/SOR could effectively reverse the overexpression of HIF-1α induced by hypoxia to overcome SOR resistance.
Inhibition of Cancer Stemness
CSCs have self-renewal capacity and differentiation potential. During SOR treatment, the CSCs in quiescent phase can evade SOR-induced cytotoxicity by activating drug resistance-associated proteins and signaling pathways, ultimately leading to SOR resistance. Because of their excellent targeting, sustained release and good EPR effect, NPs can serve as a good mediator for RNAi therapy to deliver gene drugs that target to inhibit CSCs to overcome SOR resistance.142
Deubiquitinase USP22 not only activates the SIRT1/AKT/MRP1 pathway to promote MDR, but also promotes HCC cancer stemness by HIF1α/USP22 positive feedback loop, leading to SOR resistance.143,144 The lipid nanocomposite Gal-SLPs, loaded with shUSP22, were constructed by Xu et al73 to overcome SOR resistance (Figure 17A). Glycolysis stress assays demonstrated that SOR-induced overexpression of USP22 elevated the extracellular acidification rate (ECAR) related to cancer stemness, whereas Gal-SLPs effectively reversed this effect (Figure 17B). Then, researchers established USP22-knockdown (SH) and USP22-overexpressing (OE) HCC cells via lentiviral infection as SOR-sensitive and SOR-resistant HCC cells. Cytotoxicity experiments showed that Gal-SLPs inhibited more Huh-7 OE cell proliferation than the SOR treatment group (Figure 17C), suggesting Gal-SLPs could overcome SOR resistance by the inhibitory effect on HCC stemness. The amelioration of MDR by Gal-SLPs obtained to ideal results in vivo anti-tumor experiments, in which Gal-SLPs exhibited much better antitumor efficiency in the SOR-resistant HCC patient-derived xenograft (PDX) mice model and the tumor growth inhibition (TGI) rate was 82.7±4.9% at the end of treatment, significantly higher than that of SOR treatment group (53.4±15.5%) (Figure 17D and E), and immunostaining images showed that Gal-SLPs efficiently suppressed the expression of USP22 and Ki67 (Figure 17F) and further proved Gal-SLPs could overcome SOR resistance by effectively inhibiting USP22.
 |
Figure 17 (A) Schematic illustration of Gal-SLP for reversing SOR resistance; (B) ECAR of Huh-7-R cells treated with different groups; (C) Cytotoxicity of different treatment group in Huh-7 OE cells; (D) Images of the mice at the experimental endpoint; (E)The average weight of the excised tumor in each treatment group at the experimental endpoint, Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001); (F) Representative images of IHC staining (USP22 and Ki67) and H&E staining of tumors after different treatment group. Adapted from Xu S, Ling S, Shan Q, et al. Self-activated cascade-responsive sorafenib and USP22 shRNA co-delivery system for synergetic hepatocellular carcinoma therapy. Adv Sci. 2021;8(5):2003042. https://creativecommons.org/licenses/by/4.0/.73 (G) PI3K/Akt and MAPK/ERK signaling related protein expression level after different treatments. Adapted from He C, Jaffar Ali D, Qi Y, et al. Engineered extracellular vesicles mediated CRISPR-induced deficiency of IQGAP1/FOXM1 reverses sorafenib resistance in HCC by suppressing cancer stem cells. J Nanobiotechnology. 2023;21(1):154. https://creativecommons.org/licenses/by/4.0/.83 |
Nanocomposites can further overcome resistance SOR by jointly inhibiting other pathways associated with SOR resistance. He et al83 used EVs as a carrier to load sgIF which inhibiting IQGAP1 and FOXM1 to obtain the gene-loaded nanocomposite HLC9-EVs, and took CD133+Huh7 cells with high expression of CSCs-related markers as SOR-resistant cells. Western blot analysis revealed that the combined therapy of sgIF-loaded HLC9-EVs and SOR decreased the Akt, PI3K, mTOR, MEK, and also their downstream transcriptional molecules which associated with SOR resistance in CD133+Huh7 cells by the knock-out of IQGAP1/FOXM1 (Figure 17G). CCK-8 assay indicated that sgIF-loaded HLC9-EVs and SOR exhibited a noteworthy anti-proliferative effect (43.7% ± 3.0%) on CD133+Huh7 cells, compared with SOR treatment group (26.9% ± 3.6%).
Reversal of Immunosuppression
TAMs play a critical role in promoting immunosuppression and SOR resistance,145,146 therefore, many researchers regulate TAMs to overcome SOR resistance through NPs.147 At present, the strategies for regulating TAMs mainly include inhibiting the recruitment of TAMs in the TME and repolarizing M2-type TAMs to M1-type by modulating immunosuppression-related signaling molecules.
Hypoxia inhibits the polarization of TAMs towards the immunstimulatory M1 phenotype, resulting in a diminished count of cytotoxic CD8+ T cells and consequently attenuating SOR-induced immunotoxicity. Chang et al80 constructed the MnO2-containing nanocomposite NanoMnSor which could catalyze intracellular H2O2 to generate endogenous O2 and ·OH (Figure 18A). FCM analysis revealed NanoMnSor significantly increased the proportion of M1-like macrophages (F4/80 CD86) and decreased M2-like macrophages (F4/80 CD206) compared with untreated group and NanoSor group (Figure 18B and C), suggesting that NanoMnSor could promote the repolarization of TAMs to the M1 phenotype by the amelioration of hypoxia and further reverse the decline of cytotoxic CD8+ T cells caused by immunosuppression.80 In vivo anti-tumor experiments indicated that the tumor volume of SOR-resistant HCA-1-R cells hormonal mice treated with NanoMnSor was significantly inhibited, and that in NanoMnSor plus programmed death 1 (PD-1) treatment group was reduced to about 20 mm3 which much lower than the control group (320 mm3). These results demonstrate that incorporating MnO2 into SOR-loaded NPs could promote M1 TAM polarization and enhance CD8+ T cell infiltration by ameliorate hypoxia, which could affect further potentiated by PD-1 treatment. These results indicated that MnO2-containing NPs could reverse SOR resistance mediated by immunosuppression through alleviating hypoxia and further enhance the therapeutic efficacy of SOR against HCC when combined with PD-1 treatment.
Figure 18 Continued. Figure 18 (A) Schematic representation of the mechanism by which NanoMnSor can reverse SOR resistance; (B and C) The expressions of M1-like genes and M2-like genes after NanoMn treatment. Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001). Adapted with permission from Chang C, Dinh T, Lee Y, et al. Nanoparticle Delivery of MnO and Antiangiogenic Therapy to Overcome Hypoxia-Driven Tumor Escape and Suppress Hepatocellular Carcinoma. ACS Appl Mater Interfaces. 2020;12(40):44407–44419.80 Copyright 2020 American Chemical Society. (D) Schematic illustration of PFH@LSLP-mediated antitumor synergistic therapy by regulating hypoxic and immunosuppressive microenvironment for SOR-resistant tumor treatment; (E) FCM quantification of infiltration of CD8 and CD4 T cells in tumor tissues. Adapted from Wang Y, Wang Z, Jia F, et al. CXCR4-guided liposomes regulating hypoxic and immunosuppressive microenvironment for sorafenib-resistant tumor treatment. Bioact Mater. 2022;17:147–161. https://creativecommons.org/licenses/by/nc-nd/4.0/.9

The multifunctional nanocomposite PFH@LSLP, constructed by Wang et al9 was designed to ameliorate hypoxia and activate tumor immunity via PLX3397 for reversing SOR resistance (Figure 18D). CSF1, a key regulator of macrophage differentiation, promotes TAM polarization to the immunosuppressive T2 phenotype via CSF1R binding, whereas PLX3397 blocks CSF1/CSF1R pathway to reverse immunosuppression-induced SOR resistance.148,149 FCM analysis showed the proportion of CD8+T cells in tumor tissues was appreciably increased to 5.12% in PFH@LSL group by alleviating hypoxia and reduce CXCR4 expression, and further increased to 13.1% by the combination of the PLX3397 in PFH@LSLP100 (Figure 18E). Researchers investigated the therapeutic efficacy of PFH@LSLP on SOR-resistant PDX mice models, and the results revealed the tumor volume with fortnight treatment of PFH@LSLP was only about 600 mm3, which was much lower than SOR treatment group (1300 mm3).
Le et al150 constructed the cell-penetrating peptide-aptamer dual modified-nanocomposite (USILA NPs) consisting of SOR, UA, indocyanine green (ICG), and a cell-penetrating peptide and aptamer. UA has excellent immunomodulatory properties, which could stimulate the expression of relevant immune cytokines to promote the production of cytotoxic T cells (Figure 19A).151,152 FCM analysis revealed the proportion of CD8+T cells and CD4+T cells in tumor tissues treated with USILA NPs were 0.528% and 1.55%, respectively, which much higher than the SOR group (Figure 19B). Moreover, researchers combined USILA NPs with tumor-penetrating peptides to increase the effective concentration of the USILA NPs in tumor tissues. In vivo anti-tumor experiments revealed that the tumor volume inhibition (TVI) rate was 72.38% of H22 tumor-bearing mice treated with USILA NPs plus IRGD peptide treatment group, which significantly higher than SOR treatment group (35.15%) (Figure 19C), proving that USILA NPs could effectively deliver UA to overcome immunosuppression-induced SOR resistance by activating the generation of anti-tumor immune cells in HCC. The UA-loaded nanocomposite USC NPs constructed by Lai et al81 could overcome immunosuppression-induced SOR resistance, and further enhance the therapeutic efficacy by combining of laser irradiation.
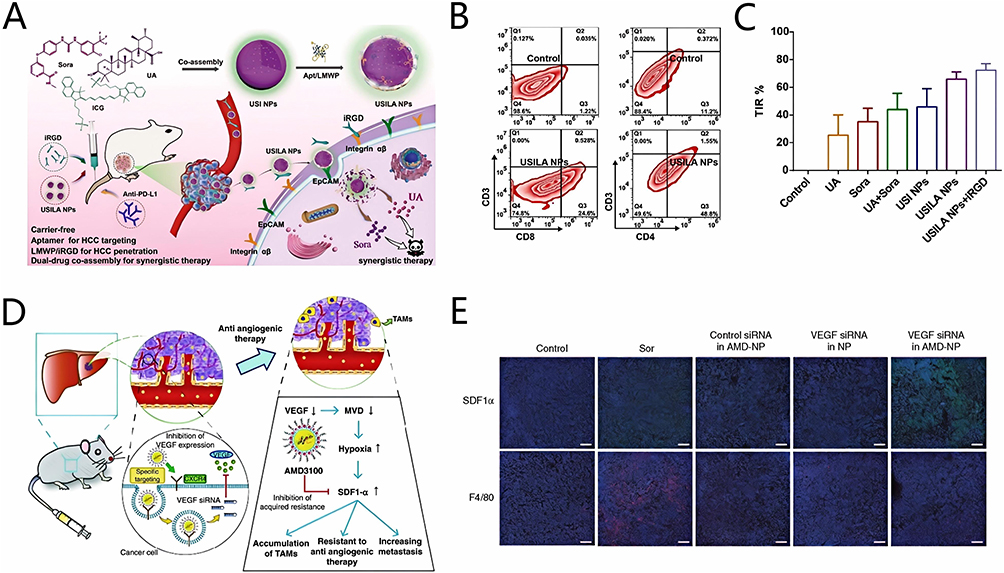 |
Figure 19 (A) Schematic illustration of mechanisms to overcome SOR resistance in HCC using USILA NPs. (B) the percentage of CD8+ T cells and CD4+ cells in tumor tissues after different treatment; (C) TIR of mice after different treatments. Reprinted from Journal of Colloid and Interface Science: 656, Le J, Song X, Tong L, et al. Dual-drug controllable co-assembly nanosystem for targeted and synergistic treatment of hepatocellular carcinoma. 177–188, Copyright 2024 with permission from Elsevier.150 Copyright 2024 Academic Press Inc. (D) Schematic illustration of mechanisms to overcome SOR resistance in HCC using CXCR4-targeted lipid-based NPs; (E) Immunostaining images of SDF1α and F4/80 treated with different groups. Reprinted from Liu J, Chiang T, Liu C, et al. Delivery of siRNA using CXCR4-targeted nanoparticles modulates tumor microenvironment and achieves a potent antitumor response in liver cancer. Mol Ther. 2015;23(11):1772–1782. Creative Commons. https://creativecommons.org/licenses/by/4.0/.71 |
Liu et al71 constructed nanocomposite AMD-NPs and AMD3100 within it could specifically inhibit CXCR4 expression93 (Figure 19D). Immunostaining images showed that AMD-NPs could reduce the expression of F4/80 (the marker of TAMs) in HCA-1 cells (simulated SOR-resistant HCC cells) which exposed to recombine SDF1α (Figure 19E), indicating that the inhibitory effect of AMD3100 on CRX4 reduced the recruitment of TAMs in HCC.153 In vivo anti-tumor experiments indicated that AMD-NPs carrying VEGF siRNA significantly suppressed the tumor growth of SOR-resistant mice models, which proved that AMD-NPs could overcome resistance by reversing immunosuppression and restoring the anti-angiogenesis effect of SOR.
Promotion of Ferroptosis
Long-term SOR treatment would cause changes such as the reduction of Fe2+ and ROS in HCC cells, which leading to SOR resistance. To overcome SOR resistance, researchers promoted ferroptosis in HCC cells by silencing genes related to ferroptosis inhibition, delivering exogenous Fe2+, and promoting ROS generation through NPs.
Some researchers used nanocomposites to overcome SOR resistance by inhibiting the expression of genes related to ferroptosis inhibition. Sun et al86 used ADSCs-derived sEVs as carriers for delivering miR-654-5p to overcome SOR resistance (Figure 20A). Researchers established HUH-7-R xenograft mice to appraise the therapeutic effect of m654-sEV combined with SOR. Western blotting analysis revealed that the HSPB1 expression in the HUH-7-R xenograft mouse models was significantly suppressed, while TFRC and COX2 levels were elevated by the combination of m654-sEV and SOR treatment (Figure 20B), resulting in increased Fe2+ and ROS levels and decreased GSH. The growth curve and appearance of the tumors showed that the combination of SOR and m654-sEV considerably suppressed tumor growth (the tumor volume was only 0.2 mm3 after 15-day treatment) compared with SOR treatment alone (0.7 mm3). The above experiments indicated that m654-sEV could target HSPB1 to upregulate SOR-induced ferroptosis and overcome SOR resistance caused by ferroptosis inhibition.
 |
Figure 20 (A) Schematic representation of the mechanism by which m654-sEV can enhance ferroptosis; (B) HSPB1, TFRC, and COX2 protein expression levels in each treatment group. Adapted from Jiao S, Qi L, Yanfang J, Zhihui C, Hui L, Huaiwen Z. Engineered small extracellular vesicles loaded with miR-654-5p promote ferroptosis by targeting HSPB1 to alleviate sorafenib resistance in hepatocellular carcinoma. Cell Death Discov. 2023;9(1):362. https://creativecommons.org/licenses/by/4.0/.86 (C) GPX4 and DHODH expressions in HepG2-R cells treated with different groups; (D) MDA levels in HepG2-R cells treated with different groups; (E) Viability of HepG2-R cells after different treatment. Adapted from Li X, Yu Q, Zhao R, et al. Designer exosomes for targeted delivery of a novel therapeutic cargo to enhance sorafenib-mediated ferroptosis in hepatocellular carcinoma. Front Oncol. 2022;12:898156. https://creativecommons.org/licenses/by/4.0/.85 (F) ROS and MDA production in HepG2-R cells treated with SOR@TF-Fe3+ NVs. (G) The change of ΔΨm after different treatment. Adapted from Xiao Y, Xu Z, Cheng Y, et al. Fe-binding transferrin nanovesicles encapsulating sorafenib induce ferroptosis in hepatocellular carcinoma. Biomater Res. 2023;27(1):63. https://creativecommons.org/licenses/by/4.0/.87 Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001) and “ns” represents no significance (P ≥ 0.05). (H) GPX4, SLC3A2, and TUNEL staining images for the resistant analysis of tumor tissues after different treatments. Reprinted from Acta Biomaterialia: 159, Wang X, Zhao L, Wang C, et al. Potent nanoreactor-mediated ferroptosis-based strategy for the reversal of cancer chemoresistance to Sorafenib. 237–246, Copyright 2023 with permission from Elsevier.78 |
Li et al85 designed a nanocomposite, ExoSP94-Lamp2b-RRM-multi-siRNA, for efficient delivery of multi-siRNA to downregulate the expression of GPX4 and DHODH (Figure 20C). The inhibition of GPX4 and DHODH further increased ROS and Malondialdehyde (MDA) levels in HepG2-R cells (Figure 20D). Cell proliferation assays revealed that ExoSP94-Lamp2b-RRM-multi-siRNA#1 could significantly enhance the effect of SOR and inhibit the growth of HepG2-R cells compared to other groups (Figure 20E), demonstrating that ExoSP94-Lamp2b-RRM-multi-siRNA restored the ferroptotic effect of SOR on HepG2-R cells by inhibiting the expressions of genes associated with ferroptosis inhibition.
HDP-ss constructed by Tong et al76 could effectively deliver siNRF2 to inhibit the overexpression of NRF2 in SOR-resistant HCC cells, and promote the production of LPO for reserving SOR resistance caused by ferroptosis inhibition.
Other researchers reversed ferroptosis inhibition-induced SOR resistance by delivering exogenous Fe3+ or producing endogenous ROS through nanocomposites. Xiao et al87 developed the sEVs-based nanocomposite SOR@TF-Fe3+ NVs, which coupled with Fe3+ and encapsulated SOR to increase intracellular Fe2+ content and SOR concentration. CLSM images showed that the obvious accumulation and generation of ROS and MDA in HepG2-R cells treated with SOR@TF-Fe3+ NVs (Figure 20F), and further resulting in cellular injury which evidenced by a substantial reduction (68%) in mitochondrial membrane potential (ΔΨm) (Figure 20G). Colony-formation assay showed that the number of colonies of HepG2-R cells after 7-day SOR@TF-Fe3+ NVs treatment was only 500 remaining, which significantly lower than SOR treatment group (about 1100), indicating that SOR@TF-Fe3+NVs explicitly induce ferroptotic growth inhibition of SOR-resistant HCC cells.
Wang et al78 constructed the nanocomposite AFG/SFB@PEG which could efficient inhibit the expressions of GPX4 and SLC3A2 in SMMC-7721-R tumor-bearing mice via increasing the effective concentration of SOR and generating extra LPO induced by AFG (Figure 20H). Furthermore, AFG endows the nanocomposite the ability of PDT and PTT treatment. Immunofluorescence images revealed that AFG/SFB@PEG plus laser irradiation significantly increased the terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) expression of tumor tissues of SMMC-7721-R tumor-bearing mice (Figure 20H), suggesting that AFG/SFB@PEG could reduce the expression of ferroptosis inhibitory protein to reverse SOR resistance and be combined with laser irradiation can further improve therapeutic effect.
Inhibiting Abnormal Activation of Signaling Molecules or Pathways
Abnormal activation of signaling molecules or pathways could lead to mutations in SOR therapeutic targets, which would directly induce SOR resistance. Various researchers delivered RNAi drugs or inhibitors by NPs to inhibit the expression of abnormal signaling molecules or pathways for overcoming SOR resistance.
MK is a heparin-binding cytokine that promotes SOR resistance when overexpressed.154 Mahmoud A. Younis et al72 constructed the nanocomposite usLNPs encapsulated with low doses of SOR and MK siRNA to efficiently overcome SOR resistance. Western blot analysis revealed that the MK expression was by 3-fold in the HepG2-R cells (Figure 21A). Notably, MK expression was silenced by approximately 70% in tumor tissues treated with usLNPs, with this effect persisting for three-day post-treatment cessation (Figure 21B). Furthermore, the combinational treatment with SOR plus MK siRNA mediated by usLNPs can restored SOR sensitive of HCC and tumor inhibition rate reached 70% (Figure 21C).
 |
Figure 21 (A) The MK expression in regular and resistant HepG2 cells; (B) MK expression in tumor tissues at the end of different treatment; (C) Representative photos for tumors at the end of different treatment. Reprinted from Journal of Controlled Release: 331, Younis MA, Khalil IA, Elewa YH, Kon Y, Harashima H. Ultra-small lipid nanoparticles encapsulating sorafenib and midkine-siRNA selectively-eradicate sorafenib-resistant hepatocellular carcinoma in vivo. 335–349, Copyright 2021 with permission from Elsevier.72 (D) The expression of various signaling molecules in PI3K/AKT/mTOR signaling pathway and Ras/Raf/MAPK signaling pathway. Adapted with permission from Wu B, Li A, Zhang Y, et al. Resistance of hepatocellular carcinoma to sorafenib can be overcome with co-delivery of PI3K/mTOR inhibitor BEZ235 and sorafenib in nanoparticles. Expert Opin Drug Deliv. 2020;17(4):573–587.155 Copyright 2020 Taylor and Francis Ltd. (E) The expression of CFL1 in MHCC-97L-R cells treated with NPs (siCFL1/Sor) which loaded different dose of CLF1; (F) Clone formation assays of MHCC-97L cells treated with different groups; (G) Expression of CFL1, Ki67, and TUNEL of the tumor tissues after different treatment. Adapted from Li S, Xu L, Wu G, et al. Remodeling serine synthesis and metabolism via nanoparticles (NPs)-mediated CFL1 silencing to enhance the sensitivity of hepatocellular carcinoma to sorafenib. Adv Sci. 2023;10(19):e2207118. https://creativecommons.org/licenses/by/4.0/. Copyright 2023 Wiley-VCH Verlag.64 Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001) and “ns” represents no significance (P ≥ 0.05). |
The nanocomposite SOR/BEZ235-NPs was constructed by Wu et al155 for the efficient co-delivery of SOR and BEZ235 to SOR-resistant HepG2 (HepG2R) cells. Intracellular BEZ235 inhibited the expression of signaling molecules in PI3K/AKT/mTOR and Ras/Raf/MAPK signaling pathways, which are implicated in SOR resistance156 (Figure 21D). In vivo anti-tumor experiments revealed that the TGI rate of SOR/BEZ235-NPs treatment group was about 82.6%±2.6%, which significantly higher than SOR treatment group (21.4%±5%).
Cofilin 1 (CFL1) could accelerate the production of antioxidant to scavenge SOR-induced excess ROS, which would reduce the efficacy of SOR and lead to SOR resistance. Li et al64 constructed nanocomposite NPs (siCFL1/Sor), utilizing polymeric NPs G0-C14 as the carrier to load CFL1 siRNA (siCFL1) and SOR. Western blot analysis demonstrated that the siCFL1/Sor NP treatment group inhibited CFL1 protein expression in SOR-resistant MHCC-97L cells (MHCC-97L-R) (Figure 21E). The number of colonies formed by MHCC-97L-R cells in the NPs (siCFL1/Sor) treatment group was significantly lower (30) compared to the SOR treatment group (approximately 170) (Figure 21F). Consistent results were observed in in vivo anti-tumor experiments displayed that NPs (siCFL1/Sor) significantly inhibited tumor growth in the SOR-resistant PDX mouse model. And histopathological images further confirmed that CFL1 expression in tumor tissues was lowest following NP treatment (Figure 21G), indicating that NPs (siCFL1/Sor) could overcome SOR resistance by efficiently inhibiting CFL1.
GRP78 play critical roles in HCC by activating EGFR signaling pathway, thereby facilitating resistance to SOR.157 siGRP78-modified Exos were sorted from BM-MSCs by Li et al84 could decrease the expression of GRP78 in HepG2-R and PLC-R cells and reverse SOR resistance (Figure 22A). The tumor volume of PLC-SR cells hormonal mice was obviously reduced in mice treated with siGRP78-modified Exos combined with SOR, which much smaller than that of the control group (scramble siRNA modified Exos) (Figure 22B). In short, siGRP78-modified Exo overcomed SOR resistance in HCC by inhibit GRP78 expression.
 |
Figure 22 (A) The expression of GRP78 in HCC cells treated with different groups; (B) Tumor volume of mice treated with different groups. Adapted from Li H, Yang C, Shi Y, Zhao L. Exosomes derived from siRNA against GRP78 modified bone-marrow-derived mesenchymal stem cells suppress Sorafenib resistance in hepatocellular carcinoma. J Nanobiotechnology. 2018;16(1):103. https://creativecommons.org/licenses/by/4.0/.84 Copyright 2018 BioMed Central. (C) The levels of ERK and MEK in HepG2-R and Hep3B-R cells after different treatment; (D and E) Representative phase contrast images and fluorescence images of 3D cells models at the end of 8-day treatment with different groups (i HNP without SOR and Sel; ii HNP; Diii free SOR and Sel). Adapted from Farinha D, Migawa M, Sarmento-Ribeiro A, Faneca H. A Combined antitumor strategy mediated by a new targeted nanosystem to hepatocellular carcinoma. Int J Nanomed. 2021;16:3385–3405. https://creativecommons.org/licenses/by/3.0/.75 |
Farinha et al75 constructed the nanocomposite HNP to effectively deliver Sel to HCC cells. Sel, as a potent and selective allosteric MEK1/2 inhibitor, could significantly inhibit the pERK expression by blocking the transduction of the signal by MEK-mediated phosphorylation of ERK105 (Figure 22C). Representative phase contrast images and fluorescence images of three-dimensional (3D) SOR-resistant HCC cell spheroids showed the spheroids treated with 8-day HNP did not present a spheroid structure but rather a structure of small aggregates (Figure 22D), which were composed with dead cells (Figure 22E).
Restoring Normal Epigenetic Regulation
Abnormal epigenetic regulation plays a primary role in SOR resistance.6 This section introduced the application of nanocomposites with different construction strategies for correcting abnormal epigenetic regulation.
The overexpression of miRNA21 in SOR-resistant HCC cells could up-regulate PTEN expression to reduce SOR efficacy by promoting hypoxic and vascular growth in tumor tissues.158 Li et al70 used the nanocomposite RGD-rHDL/So/antimiRNA21 to inhibit the expression of miRNA21 in SOR-resistant cells by stably and efficiently delivering antimiRNA21 (Figure 23A). Quantitative real-time polymerase chain reaction (qRT-PCR) analysis showed RGD-rHDL/So/antimiRNA21 significantly inhibit miRNA21 level with only 21% to the control treatment group in SOR-resistant HCC cells (Figure 23B). Then, researchers evaluated the anti-angiogenesis effect of each group by tube formation experiments on HUVEC cells, and the results showed that relatively complete tubular structures were observed in the control group, while RGD-rHDL/So/antimiRNA21 had the most suppression in tube formation with morphologies changing (Figure 23C), indicating that RGD-rHDL/So/antimiRNA21 could obviously restore the anti-angiogenesis effect of SOR in SOR-resistant HCC by the inhibition of miRNA21, and immunofluorescent staining microscopic images revealed that compared with SOR group (28.21%), the intratumoral CD31 inhibitory rate of RGD-rHDL/So/antimiRNA21 was 67.95% and further proved the above view (Figure 23D). Furthermore, RGD-rHDL/So/antimiRNA21 also induced the low-expression of HIF-1α and reduce the resistance of SOR showed in immunohistochemical staining microscopic images (Figure 23D). RGD-rHDL/So/antimiRNA21, as expected, achieved highest tumor inhibitory rate of 82.13%, which was far beyond any other group in vivo anti-tumor experiments, demonstrating that RGD-rHDL/So/antimiRNA21 could efficiently inhibit miRNA21 expression to overcome SOR resistance by enhancing the angiogenesis effect of SOR and down-regulating the expression of HIF-1α.
 |
Figure 23 (A) Schematic illustration of the proposed structure and anti-tumor mechanisms of RGD-rHDL/So/antimiRNA21 in HCC therapy; (B) miR-21 expression in HepG2 cells treated with different groups, asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001) and “ns” represents no significance (P ≥ 0.05); (C) Microscopic images of tubule formation of HUVEC after different treatment; (D) Representative immunofluorescent and immunohistochemical staining microscopic images of tumor sections from mice treated with different treatment group. Reprinted from Acta Biomaterialia: 75, Li M, Su Y, Zhang F, et al. A dual-targeting reconstituted high density lipoprotein leveraging the synergy of sorafenib and antimiRNA21 for enhanced hepatocellular carcinoma therapy. 413–426, Copyright 2018 with permission from Elsevier.70 |
lncRNA-AF085935 is implicated in the formation and proliferation of HCC through the modulation of multiple signaling pathways associated with resistance to SOR.159 AL-Noshokaty et al82 constructed the nanocomposite SeNPs for modulating resistance-related pathways to reverse SOR resistance (Figure 24A). Compared to other treatment groups, the combination of SOR and SeNPs significantly inhibited the expression of lncRNA-AF085935 (Figure 24B), which further suppressed the expression of downstream signaling molecules related to SOR resistance, such as NF-κB, mTOR (Figure 24C and D),160 thereby overcoming SOR resistance by inhibiting HCC cell proliferation and autophagy. SeNPs combined with SOR treatment significantly induced apoptosis in SOR-resistant HCC cells as seen in Western blot assays (Figure 24E).
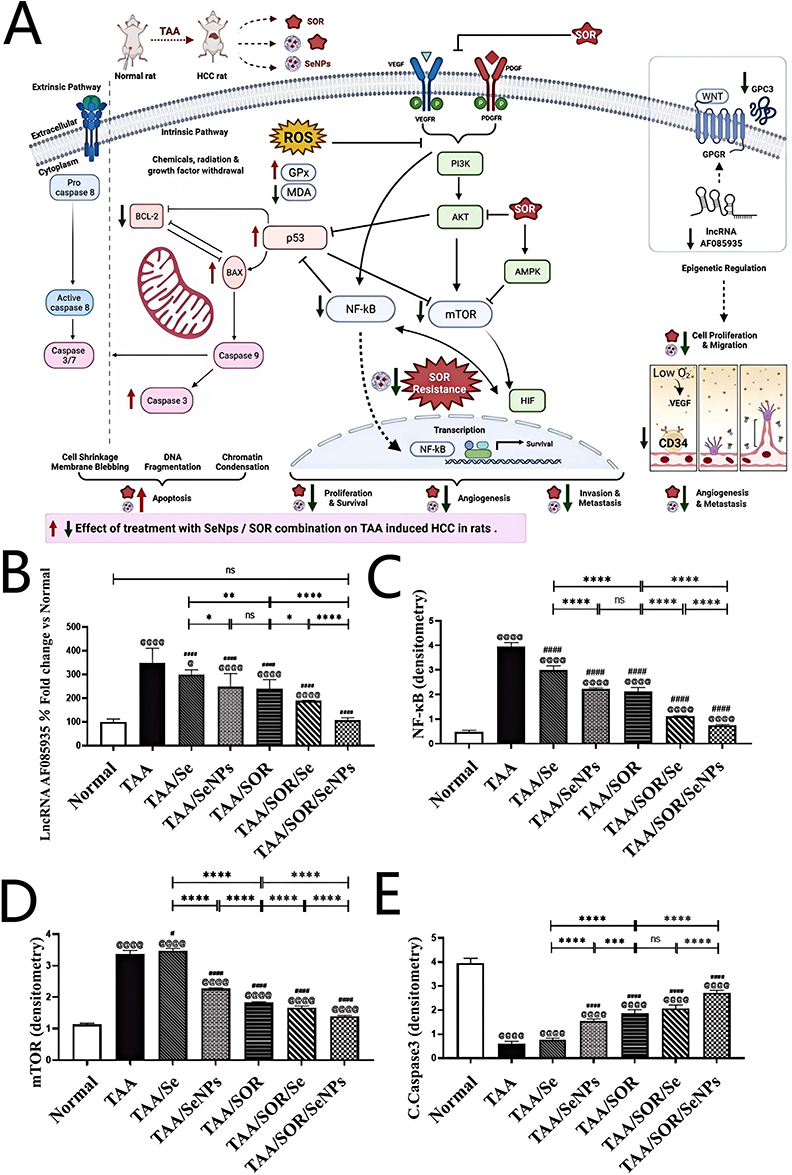 |
Figure 24 (A) Schematic illustration of SeNPs for reversing SOR resistance; (B–E) The expression of lncRNA-AF085935, NF-κB, mTOR and caspase-3 in HCC cells after different treatment, asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001), number signs (#) indicate trends (eg, #P < 0.1, ##P < 0.01) and “ns” represents no significance (P ≥ 0.05). Reprinted from Life Sciences: 303, Al-Noshokaty T, Mesbah N, Abo-Elmatty D, Abulsoud A, Abdel-Hamed A. Selenium nanoparticles overcomes sorafenib resistance in thioacetamide induced hepatocellular carcinoma in rats by modulation of mTOR, NF-κB pathways and LncRNA-AF085935/GPC3 axis. 303:120675, Copyright 2022 with permission from Elsevier.82 |
Reversing EMT
Hypoxia and DNA methylation caused by SOR treatment could stimulate and lead to EMT associated with SOR resistance. Therefore, SOR resistance can be reversed by removing EMT through nanocomposites.
Cheng et al80 constructed nanocomposites NanoMn and NanoMnSor with MnO2 as the core, which could catalyze excessive H2O2 in TME to generate O2. qRT-PCR analysis revealed that NanoMn effectively mitigated the hypoxia-induced upregulation of mRNA levels for EMT regulators (slug, snail, Zeb2, Twist1) and mesenchymal markers (vimentin, fibronectin, N-cadherin), as well as the downregulation of epithelial markers (E-cadherin, MTA3) in SOR-resistant HCA-1 (HCA-1-R) cells (Figure 25A), suggesting that NanoMn could reserve hypoxia-induced EMT by generating O2. The elevated incidence of invasive HCA-1 cells under hypoxic conditions was mitigated by NanoMnSor treatment (Figure 25B), which further proved the above results. In vivo anti-tumor experiments obtained similar result that the number of lung metastatic nodules in HAC-1-R cells hormonal mice with the treatment of NanoMnSor was significantly reduced compared to the SOR treatment group (Figure 25C), and NanoMnSor improved overall survival in hormonal mice (Figure 25D). These results proved that NanoMnSor reversed EMT by ameliorating hypoxia and overcoming SOR resistance.
 |
Figure 25 (A) The changes of different mRNA levels in HCA-1-R cells after different treatment; (B) Microscope images of HCA-1-R cells after different treatment; (C) The number of lung metastatic nodules in orthotopic HCC models treated with different groups; (D) The overall survival of orthotopic HCC models treated with different groups. Adapted with permission from ref.80 Copyright 2020 American Chemical Society. |
Improving Drug Metabolism
SOR resistance caused by insufficient effective drug concentration in HCC cells has become a major obstacle to the application of SOR, and MDR is the main problem to be overcome. P-gp, a membrane protein belonging to the ABC transporter proteins, which could reduce the concentration of SOR at the target site by pumping SOR out of HCC cells, which causing SOR resistance. Therefore, Reversing P-gp-mediated drug efflux to overcome MDR is a promising approach. Nanocomposites have many advantages such as targeting, slow controlled release, and excellent EPR effect, which can increase the effective SOR concentration in HCC cells and tissues, thus overcoming the SOR resistance.
Inhibition of MDR-related signaling pathway mediated by nanocomposites is an effective way to increase the intracellular concentration of effective drugs. Overexpression of USP22 significantly up-regulates MRP1 expression, which subsequently induces MDR and SOR resistance in HCC cells by facilitating the efflux of SOR from these cells.161 Xu et al73 constructed nanocomposite Gal-SLPs which could inhibit MPR1 expression by the efficient delivery of SOR and shUSP22 to HCC (Figure 26A). Western blot analysis demonstrated a significant downregulation of signaling molecules within the SIRT1/AKT/MRP1 signaling pathway in Huh-7-R cells with the treatment of Gal-SLPs (Figure 26B). High-performance liquid chromatography (HPLC) analysis revealed that intracellular SOR concentration was significantly increased through the delivery of Gal-SLPs (Figure 26C), indicating that Gal-SLPs effectively promote SOR accumulation in HCC cells by reserving MDR Researchers then used SOR-resistant HCC PDX model to evaluate the effect of Gal-SLPs and the results showed Gal-SLPs exhibited much better antitumor efficiency, resulting in the TIR of about 82.7%, which significantly higher than that of SOR group (53.4%) (Figure 26D). These results proved that Gal-SLP could increase the accumulation of SOR in HCC cells by reserving MDR, thus overcoming the SOR resistance caused by insufficient effective drug concentration of SOR.
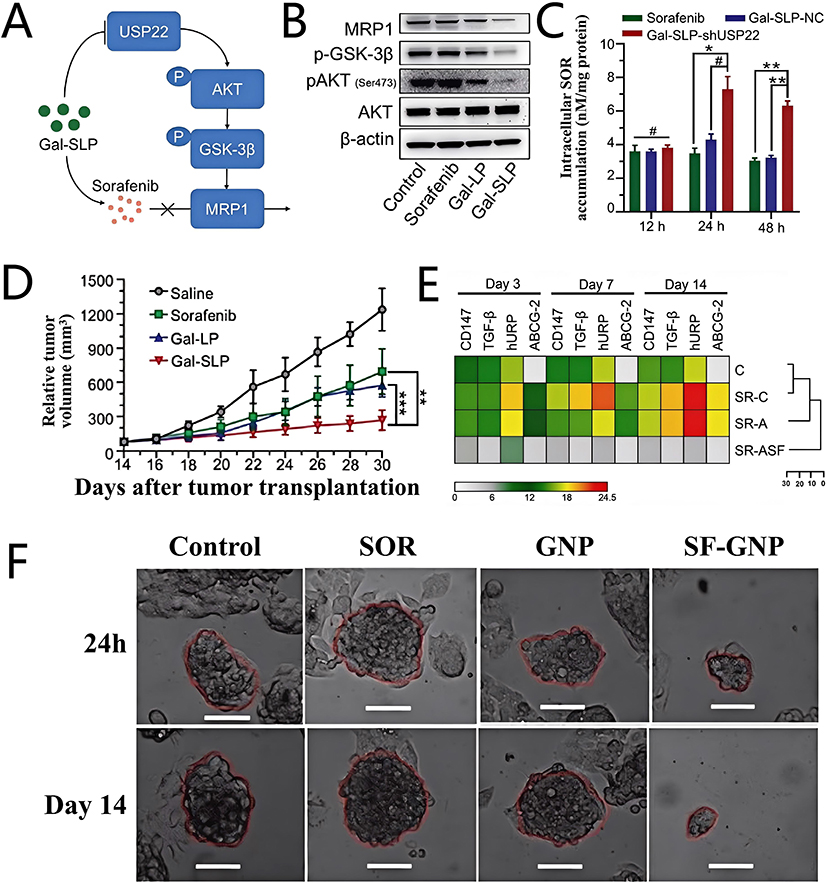 |
Figure 26 (A) Illustration of the mechanisms by which Gal-SLPs keeps HCC cells from pumping out SOR through MRP1; (B) The expression of signaling molecules in SIRT1/AKT/MRP1 signaling pathway in Huh-7-R cells after different treatment; (C) The SOR concentration in Huh-7-R cells after different treatment; (D) The tumor volume in mice tread with each group at different time. Adapted with permission from ref.73 Copyright 2021 Wiley-VCH Verlag. Asterisks (*) denote significance levels (eg, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001), number signs (#) indicate trends (eg, #P < 0.1, ##P < 0.01) and “ns” represents no significance (P ≥ 0.05). (E) The relative quantification of different signaling molecules after different treatment; (F) Microscope images of HepG2-R cell colonies after different treatment. Red color dotted circles outline the boundaries of developing HepG2 cell colonies to facilitate comparative observation of colony size changes across treatments and time. Adapted with permission from ref.79 Copyright 2017 Springer Nature. |
Vishwakarma et al79 constructed the nanocomposite SF-GNP with loading rate of 80% for SOR. The expression of ABCG-2 related MDR in HepG2-R cells treated with SF-GNP was significantly reduced displayed via the heat map162 (Figure 26E), proving that SF-GNP could help SOR evade the drug efflux mechanisms. Representative microscope images showed that the size of the 3D cell spheroids with the fortnight treatment of SF-GNP was reduced to approximately 14% of the initial size, which much lower than the SOR treatment group (60%) (Figure 26F), further demonstrating that SF-GNP could reverse SOR resistance by regulating drug efflux mechanisms and the expression of signaling molecules related to MDR.
Nanocomposites can overcome SOR resistance through different SOR resistance mechanisms. For example, to improve hypoxia, MnO2 NPs with oxygen-producing function or PFH with the function of delivering exogenous O2 can be selected. In order to reverse MDR, it is necessary to enhance the targeting ability of nanocarriers to HCC, thereby increasing the local drug concentration. However, an ideal reversal effect cannot be achieved merely through a single SOR resistance mechanism. Meanwhile, the components of the TME are complex and heterogeneous, which brings unpredictable problems for reversing SOR resistance and greatly affects the reversal effect. Therefore, the synergistic effects among various SOR resistance mechanisms can be comprehensively considered to design nanocomposites in order to improve the effect of reversing SOR resistance.
Conclusion and Outlook
The emergence of drug resistance is the primary reason limiting the clinical application of SOR. The mechanisms of SOR resistance are complex, including the development of the TME, aberrant epigenetic regulation, abnormal drug metabolism, and other factors. Moreover, drug resistance mechanisms exhibit considerable variability among individuals, making the effective overcoming of resistance a major challenge in the clinical use of SOR. The advent of nanomedicine offers an effective approach, and in recent years, researchers have developed novel nanocomposites using different NPs as carriers to efficiently overcome SOR resistance by inhibiting various resistance mechanisms. The advent of nanomedicine offers an effective approach, and in recent years, researchers have developed novel nanocomposites using different NPs as carriers to efficiently overcome SOR resistance by inhibiting various resistance mechanisms. Numerous types of nanocomposites are summarized in this review, including organic NPs, inorganic NPs, and EVs that have been developed to overcome cancer drug resistance, and some NPs are already in preclinical and clinical development stages. Current strategies for overcoming resistance using nanocomposites include: (a) delivering of drugs/genes to overcome SOR resistance, such as Meta, siHIF-1α, and LFC131 peptide; (b) selecting specific NPs or loading specific materials based on particular resistance mechanisms, such as using MnO2 NPs or loading PFH to overcome hypoxia, or using Fe-containing NPs to enhance ferroptosis; (c) modifying the exterior of NPs with specific peptides or groups to impart targeting ability, biocompatibility, and controlled drug release, such as LFC131 peptide and SP94 peptide; (d) combining two or more of the aforementioned methods.
Various types of NPs are designed to overcome SOR resistance, and each of them possesses unique advantages to effectively overcome different resistance mechanisms. However, many challenges remain to be addressed.
- Diversity of resistance mechanisms: Owing to the heterogeneity of SOR resistance mechanisms among patients, a single therapeutic strategy cannot overcome resistance in all individuals. Furthermore, SOR resistance mechanisms in HCC cells are diverse and dynamic, and multiple resistance mechanisms potentially coexisting within a single cell. Therefore, blindly constructing nanocomposites is unlikely to completely reverse resistance. Recent advancements in biotechnology, liquid biopsy, and next-generation sequencing have improved the detection of SOR resistance mutations. Analyses of tumor and cell-free DNA, immune markers, proteomics, metabolomics, and RNA can identify the biological characteristics of SOR-resistant tumors, aiding in a deeper understanding of resistance mechanisms and guiding the rational design of nanocomposites to overcome resistance.
- Rational selection of NPs: Although NPs share similar sizes, they differ in composition, structure, and properties such as fluorescence, immunogenicity, and biodegradability. Researchers need to select appropriate NPs based on the mechanisms of SOR resistance. For example, the problems that need to be overcome by NPs for RNAi therapy include low gene transfection efficiency, poor cell membrane penetration, and susceptibility to RNase degradation. EVs, as biocompatible vesicles, can evade immune surveillance and efficiently deliver RNAi genes into cancer cells. Inorganic NPs, such as CNPs and GNPs, possess photothermal properties that can be harnessed to design nanocomposites for synergistic PDT and PTT to further eliminate resistant cells. Organic NPs, such as liposomes and polymers, offer advantages like easy surface modification, allowing coupling with antibodies, peptides, aptamers, and small molecules to target HCC and inhibit SOR resistance pathways.
- Rational construction of nanocomposites: Most current nanocomposites are designed to overcome resistance based on MDR mechanisms. However, drug resistance is a multifactorial and complex phenomenon and delivering drugs efficiently to tumor cells is insufficient to overcome resistance. With increasingly studied resistance mechanisms and continuously developed NPs, the key to efficiently overcome resistance is to construct nanocomposites suitable for specific resistance mechanism. Some NPs, because of their inherent properties, can inhibit specific resistance mechanisms. For instance, MnO2 NPs can generate endogenous O2 to alleviate hypoxia-induced resistance, TF overexpression in EVs facilitates the delivery of exogenous Fe3+ to counteract resistance induced by ferroptosis inhibition, and SeNPs can overcome SOR resistance by targeting chemokines and signals related to SOR resistance. Thus, selecting appropriate NPs and rationally constructing nanocomposites based on resistance mechanisms are crucial for more effective resistance overcoming.
- Clinical application of nanocomposites: Researchers have developed a variety of nanocomposites for the treatment of HCC. However, they still face many problems in clinical translation applications, including the potential toxicity of long-term use of metal-based nanosystems, the difficulty in achieving standardized good manufacturing practice production of EVs as carriers, and the significant challenges in developing nanodelivery composites for regulating TME. For instance, researchers have designed hypoxia-responsive targeting nanocomposites to achieve precise drug release in hypoxic regions. However, TME has heterogeneity, resulting in poor therapeutic effects. Furthermore, patient-derived organoid platforms can be utilized to realistically simulate the in vivo therapeutic effects of nanocomposites in models that imitate the pathophysiology of human HCC.
- Current Technical Limitations and Knowledge Gaps: Current technologies and knowledge still remain insufficient to resolve the potential immunogenicity and toxicity of nanocomposites. The immune response triggered by nanocomposites may lead to serious complications such as allergies or thrombosis. When designing nanocomposites, it is essential to ensure low antigenicity and precise control of dosage. However, the TME of HCC is rich in a large number of immune cells (such as liver-specific Kupffer cells) and forms a dynamic adaptation network, which has compensatory resilience: inhibiting a single or multiple TME components often triggers unpredictable systemic effects through compensatory overexpression in other pathways, increasing the difficulty of constructing nanocomposites. Therefore, when designing nanocomposites, TME of HCC should be comprehensively analyzed and their long-term toxicity should be systematically investigated.
Future development and optimization of therapeutic strategies should focus on overcoming SOR resistance comprehensively. Additionally, a Phase III clinical trial in 2018 demonstrated that Lenvatinib was non-inferior to SOR, leading to FDA approval of Lenvatinib as the second first-line treatment for advanced HCC following SOR. However, resistance persists, and developing of nanocomposites to overcome SOR resistance also may provide new insights and directions for the clinical application of other TKIs, including Lenvatinib. To ensure safety, efficacy, and scalable industrial production for clinical application, nanomedicine researchers must pay sufficient attention to this field.
Acknowledgments
J. Liu and S. Zhou contributed equally to this work and should be considered as co-first authors. This work was supported by the National Natural Science Foundation of China (82172048, U21A20378), Shanxi Center of Technology Innovation for Controlled and Sustained Release of Nano-drugs (202104010911026), Foundational Research Project of Shanxi Province (202203021211159), Shanxi Scholarship Council of China (2024-058, 2022-039), Four “Batches” Innovation Project of Invigorating Medical through Science and Technology of Shanxi Province (2023XM012), The Science and Education Cultivation Fund of the National Cancer and Regional Medical Center of Shanxi Provincial Cancer Hospital (TD2023003, BD2023004).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Vogel A, Meyer T, Sapisochin G, Salem R, Saborowski A. Hepatocellular carcinoma. Lancet. 2022;400(10360):1345–1362. doi:10.1016/S0140-6736(22)01200-4
2. Llovet J, Montal R, Sia D, Finn R. Molecular therapies and precision medicine for hepatocellular carcinoma. Nat Rev Clin Oncol. 2018;15(10):599–616. doi:10.1038/s41571-018-0073-4
3. Wilhelm S, Carter C, Lynch M, et al. Discovery and development of sorafenib: a multikinase inhibitor for treating cancer. Nat Rev Drug Discov. 2006;5(10):835–844. doi:10.1038/nrd2130
4. Bahar M, Kim H, Kim D. Targeting the RAS/RAF/MAPK pathway for cancer therapy: from mechanism to clinical studies. Signal Transduct Target Ther. 2023;8(1):455. doi:10.1038/s41392-023-01705-z
5. Huang A, Yang X, Chung W, Dennison A, Zhou J. Targeted therapy for hepatocellular carcinoma. Signal Transduct Target Ther. 2020;5(1):146. doi:10.1038/s41392-020-00264-x
6. Weiwei T, Ziyi C, Wenling Z, et al. The mechanisms of sorafenib resistance in hepatocellular carcinoma: theoretical basis and therapeutic aspects. Signal Transduct Target Ther. 2020;5(1):87. doi:10.1038/s41392-020-0187-x
7. Zhang C, Jiang W, Ding J. New insights on sorafenib resistance in liver cancer with correlation of individualized therapy. BBA-Rev Cancer. 2020;1874(1):188382. doi:10.1016/j.bbcan.2020.188382
8. Zheng N, Liu W, Li B, et al. Co-delivery of sorafenib and metapristone encapsulated by CXCR4-targeted PLGA-PEG nanoparticles overcomes hepatocellular carcinoma resistance to sorafenib. J Exp Clin Cancer Res. 2019;38(1):232. doi:10.1186/s13046-019-1216-x
9. Wang Y, Wang Z, Jia F, et al. CXCR4-guided liposomes regulating hypoxic and immunosuppressive microenvironment for sorafenib-resistant tumor treatment. Bioact Mater. 2022;17:147–161. doi:10.1016/j.bioactmat.2022.01.003
10. Feng J, Dai W, Mao Y, et al. Simvastatin re-sensitizes hepatocellular carcinoma cells to sorafenib by inhibiting HIF-1α/PPAR-γ/PKM2-mediated glycolysis. J Exp Clin Cancer Res. 2020;39(1):24. doi:10.1186/s13046-020-1528-x
11. Kong F, Ye Q, Miao X, et al. Current status of sorafenib nanoparticle delivery systems in the treatment of hepatocellular carcinoma. Theranostics. 2021;11(11):5464–5490. doi:10.7150/thno.54822
12. Xu W, Yang M, Zhang W, Jia W, Zhang H, Zhang Y. Tumor microenvironment responsive nano-platform for overcoming sorafenib resistance of hepatocellular carcinoma. Mater Today Bio. 2024;24:100902. doi:10.1016/j.mtbio.2023.100902
13. Xia S, Pan Y, Liang Y, Xu J, Cai X. The microenvironmental and metabolic aspects of sorafenib resistance in hepatocellular carcinoma. EBioMedicine. 2020;51:102610. doi:10.1016/j.ebiom.2019.102610
14. Yang Y, Shuo L, Yujiao W, Yi Z, Qiu L. Protein tyrosine kinase inhibitor resistance in malignant tumors: molecular mechanisms and future perspective. Signal Transduct Target Ther. 2022;7(1):329. doi:10.1038/s41392-022-01168-8
15. Kyoko O, Asahiro M, Sae H, Koji F, Tsutomu M. The roles of epigenetic regulation and the tumor microenvironment in the mechanism of resistance to systemic therapy in hepatocellular carcinoma. Int J Mol Sci. 2023;24(3):2805. doi:10.3390/ijms24032805
16. Bhandari V, Hoey C, Liu LY, et al. Molecular landmarks of tumor hypoxia across cancer types. Nat Genet. 2019;51(2):308. doi:10.1038/s41588-018-0318-2
17. França GS, Baron M, King BR, et al. Cellular adaptation to cancer therapy along a resistance continuum. Nature. 2024;631(8022):876–883. doi:10.1038/s41586-024-07690-9
18. Méndez-Blanco C, Fondevila F, García-Palomo A, González-Gallego J, Mauriz JL. Sorafenib resistance in hepatocarcinoma: role of hypoxia-inducible factors. Exp Mol Med. 2018;50(10):1. doi:10.1038/s12276-018-0159-1
19. Lingyu Z, Long A, Zitao G, et al. Recombinant canstatin inhibits the progression of hepatocellular carcinoma by repressing the HIF-1α/VEGF signaling pathway. Biomed Pharmacother. 2024;179:117423. doi:10.1016/j.biopha.2024.117423
20. Comerford K, Wallace T, Karhausen J, Louis N, Montalto M, Colgan S. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 2002;62(12):3387–3394.
21. Won C, Kim B, Yi E, et al. Signal transducer and activator of transcription 3-mediated CD133 up-regulation contributes to promotion of hepatocellular carcinoma. Hepatology. 2015;62(4):1160–1173. doi:10.1002/hep.27968
22. Cooke V, LeBleu V, Keskin D, et al. Pericyte depletion results in hypoxia-associated epithelial-to-mesenchymal transition and metastasis mediated by met signaling pathway. Cancer Cell. 2012;21(1):66–81. doi:10.1016/j.ccr.2011.11.024
23. Liang Y, Zheng T, Song R, et al. Hypoxia-mediated sorafenib resistance can be overcome by EF24 through Von Hippel-Lindau tumor suppressor-dependent HIF-1α inhibition in hepatocellular carcinoma. Hepatology. 2013;57(5):1847–1857. doi:10.1002/hep.26224
24. Batlle E, Clevers H. Cancer stem cells revisited. Nat Med. 2017;23(10):1124–1134. doi:10.1038/nm.4409
25. Chu X, Tian W, Ning J, et al. Cancer stem cells: advances in knowledge and implications for cancer therapy. Signal Transduct Target Ther. 2024;9(1):170. doi:10.1038/s41392-024-01851-y
26. Wang H, Gong P, Chen T, et al. Colorectal cancer stem cell states uncovered by simultaneous single-cell analysis of transcriptome and telomeres. Adv Sci. 2021;8(8):2004320. doi:10.1002/advs.202004320
27. Andrea P-G, Kevin B, Cédric B. Cancer cell plasticity during tumor progression, metastasis and response to therapy. Nat Cancer. 2023;4(8):1063–1082. doi:10.1038/s43018-023-00595-y
28. den Hollander P, Maddela J, Mani S. Spatial and temporal relationship between epithelial-mesenchymal transition (EMT) and stem cells in cancer. Clin Chem. 2024;70(1):190–205. doi:10.1093/clinchem/hvad197
29. Ho P, Cahir-McFarland E, Fontenot J, et al. Harnessing regulatory T cells to establish immune tolerance. Sci Transl Med. 2024;16(738):eadm8859. doi:10.1126/scitranslmed.adm8859
30. Li K, Shi H, Zhang B, et al. Myeloid-derived suppressor cells as immunosuppressive regulators and therapeutic targets in cancer. Signal Transduct Target Ther. 2021;6(1):362. doi:10.1038/s41392-021-00670-9
31. Li M, Yang Y, Xiong L, Jiang P, Wang J, Li C. Metabolism, metabolites, and macrophages in cancer. J Hematol Oncol. 2023;16(1):80. doi:10.1186/s13045-023-01478-6
32. Caligiuri G, Tuveson D. Activated fibroblasts in cancer: perspectives and challenges. Cancer Cell. 2023;41(3):434–449. doi:10.1016/j.ccell.2023.02.015
33. van Weverwijk A, de Visser K. Mechanisms driving the immunoregulatory function of cancer cells. Nat Rev Cancer. 2023;23(4):193–215. doi:10.1038/s41568-022-00544-4
34. Kao K, Vilbois S, Tsai C, Ho P. Metabolic communication in the tumour-immune microenvironment. Nat Cell Biol. 2022;24(11):1574–1583. doi:10.1038/s41556-022-01002-x
35. Chen X, Kang R, Kroemer G, Tang D. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021;18(5):280–296. doi:10.1038/s41571-020-00462-0
36. Christophe L, Ingrid M, Hicham B, et al. The retinoblastoma (Rb) protein regulates ferroptosis induced by sorafenib in human hepatocellular carcinoma cells. Cancer Lett. 2014;356:971–977. doi:10.1016/j.canlet.2014.11.014
37. Yan L, Wenjuan Y, Yuanyuan Z, et al. Targeting fatty acid synthase modulates sensitivity of hepatocellular carcinoma to sorafenib via ferroptosis. J Exp Clin Cancer Res. 2023;42(1):6. doi:10.1186/s13046-022-02567-z
38. Kangnan Z, Qinghui Z, Rongrong J, Shihao X, Ling X. A comprehensive review of the relationship between autophagy and sorafenib-resistance in hepatocellular carcinoma: ferroptosis is noteworthy. Front Cell Dev Biol. 2023;11:1156383. doi:10.3389/fcell.2023.1156383
39. Zijian W, Chunyang Z, Yiming Z, et al. From synergy to resistance: navigating the complex relationship between sorafenib and ferroptosis in hepatocellular carcinoma. Biomed Pharmacother. 2023;170:116074. doi:10.1016/j.biopha.2023.116074
40. Yang J, Xu J, Wang W, Zhang B, Yu X, Shi S. Epigenetic regulation in the tumor microenvironment: molecular mechanisms and therapeutic targets. Signal Transduct Target Ther. 2023;8(1):210. doi:10.1038/s41392-023-01480-x
41. Elleithi Y, El-Gayar A, Amin MN. Autophagy modulation attenuates sorafenib resistance in HCC induced in rats. Cell Death Dis. 2024;15(8):595. doi:10.1038/s41419-024-06955-5
42. Xiong YX, Zhang XC, Zhu JH, et al. Collagen I-DDR1 signaling promotes hepatocellular carcinoma cell stemness via Hippo signaling repression. Cell Death Differ. 2023;30(7):1648–1665. doi:10.1038/s41418-023-01166-5
43. Liu A, Wu Q, Peng D, et al. A novel strategy for the diagnosis, prognosis, treatment, and chemoresistance of hepatocellular carcinoma: DNA methylation. Med Res Rev. 2020;40(5):1973–2018. doi:10.1002/med.21696
44. Galle E, Thienpont B, Cappuyns S, et al. DNA methylation-driven EMT is a common mechanism of resistance to various therapeutic agents in cancer. Clin Clin Epigenet. 2020;12(1):27. doi:10.1186/s13148-020-0821-z
45. Bi L, Ren Y, Feng M, et al. HDAC11 regulates glycolysis through the LKB1/AMPK signaling pathway to maintain hepatocellular carcinoma stemness. Cancer Res. 2021;81(8):2015–2028. doi:10.1158/0008-5472.CAN-20-3044
46. Wu J, Chai H, Li F, Ren Q, Gu Y. SETD1A augments sorafenib primary resistance via activating YAP in hepatocellular carcinoma. Life Sci. 2020;260:118406. doi:10.1016/j.lfs.2020.118406
47. Zhou T, Zhuang L, Hu Y, et al. Inactivation of hypoxia-induced YAP by statins overcomes hypoxic resistance tosorafenib in hepatocellular carcinoma cells. Sci Rep. 2016;6:30483. doi:10.1038/srep30483
48. Zhou K, Nguyen R, Qiao L, George J. Single cell RNA-seq analysis identifies a noncoding RNA mediating resistance to sorafenib treatment in HCC. Mol Cancer. 2022;21(1):6. doi:10.1186/s12943-021-01473-w
49. Hu X, Zhu H, Shen Y, Zhang X, He X, Xu X. The role of non-Coding RNAs in the sorafenib resistance of hepatocellular carcinoma. Front Oncol. 2021;11:696705. doi:10.3389/fonc.2021.696705
50. Kunden R, Khan J, Ghezelbash S, Wilson J. The role of the liver-specific microRNA, miRNA-122 in the HCV replication cycle. Int J Mol Sci. 2020;21(16):5677. doi:10.3390/ijms21165677
51. Xu Y, Huang J, Ma L, et al. MicroRNA-122 confers sorafenib resistance to hepatocellular carcinoma cells by targeting IGF-1R to regulate RAS/RAF/ERK signaling pathways. Cancer Lett. 2016;371(2):171–181. doi:10.1016/j.canlet.2015.11.034
52. Pollutri D, Patrizi C, Marinelli S, et al. The epigenetically regulated miR-494 associates with stem-cell phenotype and induces sorafenib resistance in hepatocellular carcinoma. Cell Death Dis. 2018;9(1):4. doi:10.1038/s41419-017-0076-6
53. Wang W, Ding B, Lou W, Lin S. Promoter Hypomethylation and miR-145-5p downregulation- mediated HDAC11 overexpression promotes sorafenib resistance and metastasis of hepatocellular carcinoma cells. Front Cell Dev Biol. 2020;8:724. doi:10.3389/fcell.2020.00724
54. Yao Z, Yang Y, Sun M, et al. New insights into the interplay between long non-coding RNAs and RNA-binding proteins in cancer. Cancer Commun. 2022;42(2):117–140. doi:10.1002/cac2.12254
55. Chan Y, Wu J, Lu Y, et al. Loss of lncRNA LINC01056 leads to sorafenib resistance in HCC. Mol Cancer. 2024;23(1):74. doi:10.1186/s12943-024-01988-y
56. Wei L, Wang X, Lv L, et al. The emerging role of microRNAs and long noncoding RNAs in drug resistance of hepatocellular carcinoma. Mol Cancer. 2019;18(1):147. doi:10.1186/s12943-019-1086-z
57. Ghafouri-Fard S, Bahroudi Z, Shoorei H, Abak A, Ahin M, Taheri M. microRNA-140: a miRNA with diverse roles in human diseases. Biomed Pharmacother. 2021;135:111256. doi:10.1016/j.biopha.2021.111256
58. Wang M, Yu F, Chen X, Li P, Wang K. The underlying mechanisms of noncoding RNAs in the chemoresistance of hepatocellular carcinoma. Mol Ther Nucleic Acids. 2020;21:13–27. doi:10.1016/j.omtn.2020.05.011
59. Xu J, Ji L, Liang Y, et al. CircRNA-SORE mediates sorafenib resistance in hepatocellular carcinoma by stabilizing YBX1. Signal Transduct Target Ther. 2020;5(1):298. doi:10.1038/s41392-020-00375-5
60. Yang C, Dong Z, Hong H, et al. circFN1 mediates sorafenib resistance of hepatocellular carcinoma cells by sponging miR-1205 and regulating E2F1 expression. Mol Ther Nucleic Acids. 2020;22:421–433. doi:10.1016/j.omtn.2020.08.039
61. Weng H, Zeng L, Cao L, et al. circFOXM1 contributes to sorafenib resistance of hepatocellular carcinoma cells by regulating MECP2 via miR-1324. Mol Ther Nucleic Acids. 2021;23:811–820. doi:10.1016/j.omtn.2020.12.019
62. Zhu M, Wu M, Bian S, et al. DNA primase subunit 1 deteriorated progression of hepatocellular carcinoma by activating AKT/mTOR signaling and UBE2C-mediated P53 ubiquitination. Cell Biosci. 2021;11(1):42. doi:10.1186/s13578-021-00555-y
63. Tan X, Xiong B, Zhang Y, Wang S, Zuo Q, Li J. FXYD5 promotes sorafenib resistance through the Akt/mTOR signaling pathway in hepatocellular carcinoma. Eur J Pharmacol. 2022;931:175186. doi:10.1016/j.ejphar.2022.175186
64. Li S, Xu L, Wu G, et al. Remodeling serine synthesis and metabolism via nanoparticles (NPs)-mediated CFL1 silencing to enhance the sensitivity of hepatocellular carcinoma to sorafenib. Adv Sci. 2023;10(19):e2207118. doi:10.1002/advs.202207118
65. Erin N, Grahovac J, Brozovic A, Efferth T. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist Updat. 2020;53:100715. doi:10.1016/j.drup.2020.100715
66. Mani S, Guo W, Liao M, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704–715. doi:10.1016/j.cell.2008.03.027
67. Du B, Shim J. Targeting epithelial-mesenchymal transition (EMT) to overcome drug resistance in cancer. Molecules. 2016;21(7):965. doi:10.3390/molecules21070965
68. Chen F, Fang Y, Chen X, Deng R, Zhang Y, Shao J. Recent advances of sorafenib nanoformulations for cancer therapy: smart nanosystem and combination therapy. Asian J Pharm Sci. 2021;16(3):318–336. doi:10.1016/j.ajps.2020.07.003
69. Zhou T, Liang X, Wang P, et al. A hepatocellular carcinoma targeting nanostrategy with hypoxia-ameliorating and photothermal abilities that, combined with immunotherapy, inhibits metastasis and recurrence. ACS Nano. 2020;14(10):12679–12696. doi:10.1021/acsnano.0c01453
70. Li M, Su Y, Zhang F, et al. A dual-targeting reconstituted high density lipoprotein leveraging the synergy of sorafenib and antimiRNA21 for enhanced hepatocellular carcinoma therapy. Acta Biomater. 2018;75:413–426. doi:10.1016/j.actbio.2018.05.049
71. Liu J, Chiang T, Liu C, et al. Delivery of siRNA using CXCR4-targeted nanoparticles modulates tumor microenvironment and achieves a potent antitumor response in liver cancer. Mol Ther. 2015;23(11):1772–1782. doi:10.1038/mt.2015.147
72. Younis MA, Khalil IA, Elewa YH, Kon Y, Harashima H. Ultra-small lipid nanoparticles encapsulating sorafenib and midkine-siRNA selectively-eradicate sorafenib-resistant hepatocellular carcinoma in vivo. J Control Release. 2021;331:335–349. doi:10.1016/j.jconrel.2021.01.021
73. Xu S, Ling S, Shan Q, et al. Self-activated cascade-responsive sorafenib and USP22 shRNA co-delivery system for synergetic hepatocellular carcinoma therapy. Adv Sci. 2021;8(5):2003042. doi:10.1002/advs.202003042
74. Zan Y, Dai Z, Liang L, Deng Y, Dong L. Co-delivery of plantamajoside and sorafenib by a multi-functional nanoparticle to combat the drug resistance of hepatocellular carcinoma through reprograming the tumor hypoxic microenvironment. Drug Deliv. 2019;26(1):1080–1091. doi:10.1080/10717544.2019.1654040
75. Farinha D, Migawa M, Sarmento-Ribeiro A, Faneca H. A Combined antitumor strategy mediated by a new targeted nanosystem to hepatocellular carcinoma. Int J Nanomed. 2021;16:3385–3405. doi:10.2147/IJN.S302288
76. Tong R, Feng X, Sun J, et al. Co-delivery of siNRF2 and sorafenib by a “click” dual functioned hyperbranched nanocarrier for synergistically inducing ferroptosis in hepatocellular carcinoma. Small. 2024;20(21):e2307273. doi:10.1002/smll.202307273
77. Wan Y, Yang Y, Lai Q, Wang W, Wu M, Feng S. Fluorinated cell-penetrating peptide for co-delivering siHIF-1α and sorafenib to enhance in vitro anti-tumor efficacy. Pharmaceutics. 2023;15(12):2789. doi:10.3390/pharmaceutics15122789
78. Wang X, Zhao L, Wang C, et al. Potent nanoreactor-mediated ferroptosis-based strategy for the reversal of cancer chemoresistance to Sorafenib. Acta Biomater. 2023;159:237–246. doi:10.1016/j.actbio.2023.01.053
79. Vishwakarma S, Sharmila P, Bardia A, et al. Use of biocompatible sorafenib-gold nanoconjugates for reversal of drug resistance in human hepatoblatoma cells. Sci Rep. 2017;7(1):8539. doi:10.1038/s41598-017-08878-y
80. Chang C, Dinh T, Lee Y, et al. Nanoparticle Delivery of MnO and Antiangiogenic Therapy to Overcome Hypoxia-Driven Tumor Escape and Suppress Hepatocellular Carcinoma. ACS Appl Mater Interfaces. 2020;12(40):44407–44419. doi:10.1021/acsami.0c08473
81. Lai C, Zhang B, Li D, et al. Rational design of a minimum nanoplatform for maximizing therapeutic potency: three birds with one stone. J Colloid Interface Sci. 2023;635:441–455. doi:10.1016/j.jcis.2022.12.157
82. Al-Noshokaty T, Mesbah N, Abo-Elmatty D, Abulsoud A, Abdel-Hamed A. Selenium nanoparticles overcomes sorafenib resistance in thioacetamide induced hepatocellular carcinoma in rats by modulation of mTOR, NF-κB pathways and LncRNA-AF085935/GPC3 axis. Life Sci. 2022;303:120675. doi:10.1016/j.lfs.2022.120675
83. He C, Jaffar Ali D, Qi Y, et al. Engineered extracellular vesicles mediated CRISPR-induced deficiency of IQGAP1/FOXM1 reverses sorafenib resistance in HCC by suppressing cancer stem cells. J Nanobiotechnology. 2023;21(1):154. doi:10.1186/s12951-023-01902-6
84. Li H, Yang C, Shi Y, Zhao L. Exosomes derived from siRNA against GRP78 modified bone-marrow-derived mesenchymal stem cells suppress Sorafenib resistance in hepatocellular carcinoma. J Nanobiotechnology. 2018;16(1):103. doi:10.1186/s12951-018-0429-z
85. Li X, Yu Q, Zhao R, et al. Designer exosomes for targeted delivery of a novel therapeutic cargo to enhance sorafenib-mediated ferroptosis in hepatocellular carcinoma. Front Oncol. 2022;12:898156. doi:10.3389/fonc.2022.898156
86. Jiao S, Qi L, Yanfang J, Zhihui C, Hui L, Huaiwen Z. Engineered small extracellular vesicles loaded with miR-654-5p promote ferroptosis by targeting HSPB1 to alleviate sorafenib resistance in hepatocellular carcinoma. Cell Death Discov. 2023;9(1):362. doi:10.1038/s41420-023-01660-2
87. Xiao Y, Xu Z, Cheng Y, et al. Fe-binding transferrin nanovesicles encapsulating sorafenib induce ferroptosis in hepatocellular carcinoma. Biomater Res. 2023;27(1):63. doi:10.1186/s40824-023-00401-x
88. Gasco M. Lipid nanoparticles: perspectives and challenges. Adv Drug Deliv Rev. 2007;59(6):377–378. doi:10.1016/j.addr.2007.05.004
89. Xiao Y, Shi J. Lipids and the emerging RNA medicines. Chem Rev. 2021;121(20):12109–12111. doi:10.1021/acs.chemrev.1c00778
90. Yamin L, Zhongfeng Y, Hanyi Y, Qiaobing X. Tailoring combinatorial lipid nanoparticles for intracellular delivery of nucleic acids, proteins, and drugs. Acta Pharm Sin B. 2022;12(6):2624–2639. doi:10.1016/j.apsb.2022.04.013
91. Sonju JJ, Dahal A, Singh SS, Jois SD. Peptide-functionalized liposomes as therapeutic and diagnostic tools for cancer treatment. J Control Release. 2020;329:624–644. doi:10.1016/j.jconrel.2020.09.055
92. Zhang Y, Sun C, Wang C, Jankovic K, Dong Y. Lipids and Lipid Derivatives for RNA Delivery. Chem Rev. 2021;121(20):12181–12277. doi:10.1021/acs.chemrev.1c00244
93. Wang J, Tannous BA, Poznansky MC, Chen H. CXCR4 antagonist AMD3100 (plerixafor): from an impurity to a therapeutic agent. Pharmacol Res. 2020;159:105010. doi:10.1016/j.phrs.2020.105010
94. Younis M, Khalil I, Abd Elwakil M, Harashima H. A Multifunctional Lipid-based nanodevice for the highly specific codelivery of sorafenib and midkine siRNA to hepatic cancer cells. Mol Pharm. 2019;16(9):4031–4044. doi:10.1021/acs.molpharmaceut.9b00738
95. Sato Y, Hatakeyama H, Sakurai Y, Hyodo M, Akita H, Harashima H. A pH-sensitive cationic lipid facilitates the delivery of liposomal siRNA and gene silencing activity in vitro and in vivo. J Control Release. 2012;163(3):267–276. doi:10.1016/j.jconrel.2012.09.009
96. Gyan P, Ahmed S, Niels W, Showkeen Muzamil B, Su Ryon S, Shabir H. Microfluidic fabrication of lipid nanoparticles for the delivery of nucleic acids. Adv Drug Deliv Rev. 2022;184:114197. doi:10.1016/j.addr.2022.114197
97. Sun Y, Ma W, Yang Y, et al. Cancer nanotechnology: enhancing tumor cell response to chemotherapy for hepatocellular carcinoma therapy. Asian J Pharm Sci. 2019;14(6):581–594. doi:10.1016/j.ajps.2019.04.005
98. Chen X, Liu L, Jiang C. Charge-reversal nanoparticles: novel targeted drug delivery carriers. Acta Pharm Sin B. 2016;6(4):261–267. doi:10.1016/j.apsb.2016.05.011
99. Prakash T, Yu J, Migawa M, et al. Comprehensive structure-activity relationship of triantennary N-Acetylgalactosamine conjugated antisense oligonucleotides for targeted delivery to hepatocytes. J Med Chem. 2016;59(6):2718–2733. doi:10.1021/acs.jmedchem.5b01948
100. Wen J, Wang S, Guo R, Liu D. CSF1R inhibitors are emerging immunotherapeutic drugs for cancer treatment. Eur J Med Chem. 2023;245:114884. doi:10.1016/j.ejmech.2022.114884
101. Banik B, Fattahi P, Brown J. Polymeric nanoparticles: the future of nanomedicine. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2016;8(2):271–299. doi:10.1002/wnan.1364
102. Kamaly N, Xiao Z, Valencia P, Radovic-Moreno A, Farokhzad O. Targeted polymeric therapeutic nanoparticles: design, development and clinical translation. Chem Soc Rev. 2012;41(7):2971–3010. doi:10.1039/c2cs15344k
103. Zhu Y, Tang K, Lv J. Peptide-drug conjugate-based novel molecular drug delivery system in cancer. Trends Pharmacol Sci. 2021;42(10):857–869. doi:10.1016/j.tips.2021.07.001
104. Su H, Rong G, Li L, Cheng Y. Subcellular targeting strategies for protein and peptide delivery. Adv Drug Deliv Rev. 2024;212:115387. doi:10.1016/j.addr.2024.115387
105. Yeh T, Marsh V, Bernat B, et al. Biological characterization of ARRY-142886 (AZD6244), a potent, highly selective mitogen-activated protein kinase kinase 1/2 inhibitor. Clin Cancer Res. 2007;13(5):1576–1583. doi:10.1158/1078-0432.CCR-06-1150
106. Wang Q, Cui H, Wang X, et al. Exceptional light sensitivity by thiol-ene click lithography. J Am Chem Soc. 2023;145(5):3064–3074. doi:10.1021/jacs.2c11887
107. Wan Y, Yang Y, Wu M, Feng S. Fluorinated vectors for gene delivery. Expert Opin Drug Deliv. 2022;19(11):1435–1448. doi:10.1080/17425247.2022.2131769
108. Wu P, Luo X, Wu H, Zhang Q, Dai Y, Sun M. Efficient and targeted chemo-gene delivery with self-assembled fluoro-nanoparticles for liver fibrosis therapy and recurrence. Biomaterials. 2020;261:120311. doi:10.1016/j.biomaterials.2020.120311
109. Krafft M, Riess J. Therapeutic oxygen delivery by perfluorocarbon-based colloids. Adv Colloid Interface Sci. 2021;294:102407. doi:10.1016/j.cis.2021.102407
110. Pugazhendhi A, Edison T, Karuppusamy I, Kathirvel B. Inorganic nanoparticles: a potential cancer therapy for human welfare. Int J Pharm. 2018;539(1–2):104–111. doi:10.1016/j.ijpharm.2018.01.034
111. Tee J, Yip L, Tan E, et al. Nanoparticles’ interactions with vasculature in diseases. Chem Soc Rev. 2019;48(21):5381–5407. doi:10.1039/c9cs00309f
112. Sharifi M, Attar F, Saboury A, et al. Plasmonic gold nanoparticles: optical manipulation, imaging, drug delivery and therapy. J Control Release. 2019;311:170–189. doi:10.1016/j.jconrel.2019.08.032
113. Pasparakis G. Recent developments in the use of gold and silver nanoparticles in biomedicine. Wiley Interdiscip Rev Nanobiotechnol. 2022;14(5):e1817. doi:10.1002/wnan.1817
114. Kuo W, Chang Y, Cho K, et al. Gold nanomaterials conjugated with indocyanine green for dual-modality photodynamic and photothermal therapy. Biomaterials. 2012;33(11):3270–3278. doi:10.1016/j.biomaterials.2012.01.035
115. Xu X, Duan J, Liu Y, et al. Multi-stimuli responsive hollow MnO-based drug delivery system for magnetic resonance imaging and combined chemo-chemodynamic cancer therapy. Acta Biomater. 2021;126:445–462. doi:10.1016/j.actbio.2021.03.048
116. Liang L, Jia M, Zhao M, et al. Progress of nanomaterials based on manganese dioxide in the field of tumor diagnosis and therapy. Int J Nanomed. 2024;19:8883–8900. doi:10.2147/IJN.S477026
117. Song M, Liu T, Shi C, Zhang X, Chen X. Bioconjugated manganese dioxide nanoparticles enhance chemotherapy response by priming tumor-associated macrophages toward M1-like phenotype and attenuating tumor hypoxia. ACS nano. 2016;10(1):633–647. doi:10.1021/acsnano.5b06779
118. Huang Y, Ruan Y, Ma Y, et al. Immunomodulatory activity of manganese dioxide nanoparticles: promising for novel vaccines and immunotherapeutics. Front Immunol. 2023;14:1128840. doi:10.3389/fimmu.2023.1128840
119. Tang Q, Cheng Z, Yang N, et al. Hydrangea-structured tumor microenvironment responsive degradable nanoplatform for hypoxic tumor multimodal imaging and therapy. Biomaterials. 2019;205:1–10. doi:10.1016/j.biomaterials.2019.03.005
120. Jia F, Zhou S, Liu J, et al. Metal-modified carbon dots: synthesis, properties, and applications in cancer diagnosis and treatment. Appl Mater Today. 2024;37:102133. doi:10.1016/j.apmt.2024.102133
121. Varlamova E, Turovsky E, Blinova E. Therapeutic potential and main methods of obtaining selenium nanoparticles. Int J Mol Sci. 2021;22(19). doi:10.3390/ijms221910808
122. Liu T, Zeng L, Jiang W, Fu Y, Zheng W, Chen T. Rational design of cancer-targeted selenium nanoparticles to antagonize multidrug resistance in cancer cells. Nanomedicine. 2015;11(4):947–958. doi:10.1016/j.nano.2015.01.009
123. Zhang C, Zhai X, Zhao G, Ren F, Leng X. Synthesis, characterization, and controlled release of selenium nanoparticles stabilized by chitosan of different molecular weights. Carbohydr Polym. 2015;134:158–166. doi:10.1016/j.carbpol.2015.07.065
124. Qiuyue L, Jiahui Z, Zhongjian C, Wei H, Wei W. Current research trends of nanomedicines. Acta Pharm Sin B. 2023;13(11):4391–4416. doi:10.1016/j.apsb.2023.05.018
125. Chen M-K, Chen Z-X, Cai M-P, Chen H, Chen Z-F, Zhao S-C. Engineered extracellular vesicles: a new approach for targeted therapy of tumors and overcoming drug resistance. Cancer Commun. 2023;44(2):205–225. doi:10.1002/cac2.12518
126. Qiurong Y, Jing X, Jianmei G, et al. Extracellular vesicles in cancer drug resistance: roles, mechanisms, and implications. Adv Sci. 2022;9(34):202201609. doi:10.1002/advs.202201609
127. Ramos CC, Pires J, Gonzalez E, et al. Extracellular vesicles in tumor-adipose tissue crosstalk: key drivers and therapeutic targets in cancer cachexia. Extracell Vesicles Circ Nucl Acids. 2024;5(3):371–396. doi:10.20517/evcna.2024.36
128. Chen W, Li Z, Yu N, et al. Bone-targeting exosome nanoparticles activate Keap1 / Nrf2 / GPX4 signaling pathway to induce ferroptosis in osteosarcoma cells. J Nanobiotechnology. 2023;21(1):355. doi:10.1186/s12951-023-02129-1
129. Khosravi N, Pishavar E, Baradaran B, Oroojalian F, Mokhtarzadeh A. Stem cell membrane, stem cell-derived exosomes and hybrid stem cell camouflaged nanoparticles: a promising biomimetic nanoplatforms for cancer theranostics. J Control Release. 2022;348:706–722. doi:10.1016/j.jconrel.2022.06.026
130. Chen Q, Li Q, Liang Y, et al. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer ROS generation and microbiota modulation. Acta Pharm Sin B. 2022;12(2):907–923. doi:10.1016/j.apsb.2021.08.016
131. Wilson TR, Fridlyand J, Yan Y, et al. Widespread potential for growth-factor-driven resistance to anticancer kinase inhibitors. Nature. 2012;487(7408):505–509. doi:10.1038/nature11249
132. Zhang N, Wei P, Gong A, et al. FoxM1 promotes β-catenin nuclear localization and controls Wnt target-gene expression and glioma tumorigenesis. Cancer Cell. 2011;20(4):427–442. doi:10.1016/j.ccr.2011.08.016
133. Wu Y, Liu H, Ding H. GPC-3 in hepatocellular carcinoma: current perspectives. J Hepatocell Carcinoma. 2016;3:63–67. doi:10.2147/JHC.S116513
134. Zhou F, Shang W, Yu X, Tian J. Glypican-3: a promising biomarker for hepatocellular carcinoma diagnosis and treatment. Med Res Rev. 2018;38(2):741–767. doi:10.1002/med.21455
135. Wang F, Min J. DHODH tangoing with GPX4 on the ferroptotic stage. Signal Transduct Target Ther. 2021;6(1):244. doi:10.1038/s41392-021-00656-7
136. Sun L, Liu H, Ye Y, et al. Smart nanoparticles for cancer therapy. Signal Transduct Target Ther. 2023;8(1):418. doi:10.1038/s41392-023-01642-x
137. Yanan C, Zhonghao J, Zhuo Y, et al. A codelivery system loaded with PDL1-siRNA and sorafenib enhances the therapeutic effect of sorafenib on hepatocellular carcinoma via TAT-poly-SS-lysine modified chitosan. Int J Biol Macromol. 2024;263:130162. doi:10.1016/j.ijbiomac.2024.130162
138. Xiuli Y, Qiang Z, Zhifei D. Near-infrared light-activatable polymeric nanoformulations for combined therapy and imaging of cancer. Adv Drug Deliv Rev. 2017;115:155–170. doi:10.1016/j.addr.2017.04.007
139. Jun D, Zhaojun C, Shixuan W, Fan X, Xiaoding L. Erythrocyte membrane-camouflaged nanoparticles as effective and biocompatible platform: either autologous or allogeneic erythrocyte-derived. Mater Today Bio. 2022;15:100279. doi:10.1016/j.mtbio.2022.100279
140. Zheng N, Chen J, Li T, et al. Abortifacient metapristone (RU486 derivative) interrupts CXCL12/CXCR4 axis for ovarian metastatic chemoprevention. Mol Carcinog. 2017;56(8):1896–1908. doi:10.1002/mc.22645
141. Pei S, Yang X, Wang H, et al. Plantamajoside, a potential anti-tumor herbal medicine inhibits breast cancer growth and pulmonary metastasis by decreasing the activity of matrix metalloproteinase-9 and −2. BMC Cancer. 2015;15:965. doi:10.1186/s12885-015-1960-z
142. Wang WD, Guo YY, Yang ZL, Su GL, Sun ZJ. Sniping Cancer Stem Cells with Nanomaterials. ACS Nano. 2023;17(23):23262–23298. doi:10.1021/acsnano.3c07828
143. Ling S, Shan Q, Zhan Q, et al. USP22 promotes hypoxia-induced hepatocellular carcinoma stemness by a HIF1α/USP22 positive feedback loop upon TP53 inactivation. Gut. 2020;69(7):1322–1334. doi:10.1136/gutjnl-2019-319616
144. Cortez J, Montauti E, Shifrut E, et al. CRISPR screen in regulatory T cells reveals modulators of Foxp3. Nature. 2020;582(7812):416–420. doi:10.1038/s41586-020-2246-4
145. Wang H, Haung L, Wang C, et al. Tumor-associated macrophages promote resistance of hepatocellular carcinoma cells against sorafenib by activating CXCR2 signaling. J Biomed Sci. 2022;29(1):99. doi:10.1186/s12929-022-00881-4
146. Li W, Zhao B, Wang Q, Lu J, Wu X, Chen X. M2 macrophage exosomes promote resistance to sorafenib in hepatocellular carcinoma cells via miR-200c-3p. Int Immunopharmacol. 2024;139:112807. doi:10.1016/j.intimp.2024.112807
147. Huang L, Xu R, Li W, et al. Repolarization of macrophages to improve sorafenib sensitivity for combination cancer therapy. Acta biomater. 2023;162:98–109. doi:10.1016/j.actbio.2023.03.014
148. Buechler M, Fu W, Turley S. Fibroblast-macrophage reciprocal interactions in health, fibrosis, and cancer. Immunity. 2021;54(5):903–915. doi:10.1016/j.immuni.2021.04.021
149. Hume D, MacDonald K. Therapeutic applications of macrophage colony-stimulating factor-1 (CSF-1) and antagonists of CSF-1 receptor (CSF-1R) signaling. Blood. 2012;119(8):1810–1820. doi:10.1182/blood-2011-09-379214
150. Le J, Song X, Tong L, et al. Dual-drug controllable co-assembly nanosystem for targeted and synergistic treatment of hepatocellular carcinoma. J Colloid Interface Sci. 2024;656:177–188. doi:10.1016/j.jcis.2023.11.109
151. Zhang B, Jiang J, Wu P, et al. A smart dual-drug nanosystem based on co-assembly of plant and food-derived natural products for synergistic HCC immunotherapy. Acta Pharm Sin B. 2021;11(1):246–257. doi:10.1016/j.apsb.2020.07.026
152. Zhao R, Li T, Zheng G, Jiang K, Fan L, Shao J. Simultaneous inhibition of growth and metastasis of hepatocellular carcinoma by co-delivery of ursolic acid and sorafenib using lactobionic acid modified and pH-sensitive chitosan-conjugated mesoporous silica nanocomplex. Biomaterials. 2017;143:1–16. doi:10.1016/j.biomaterials.2017.07.030
153. Chen J, Feng W, Sun M, et al. TGF-β1-induced SOX18 elevation promotes hepatocellular carcinoma progression and metastasis through transcriptionally upregulating PD-L1 and CXCL12. Gastroenterology. 2024;167(2):264–280. doi:10.1053/j.gastro.2024.02.025
154. Christou C, Stylianou A, Gkretsi V. Midkine (MDK) in hepatocellular carcinoma: more than a biomarker. Cells. 2024;13(2):136. doi:10.3390/cells13020136
155. Wu B, Li A, Zhang Y, et al. Resistance of hepatocellular carcinoma to sorafenib can be overcome with co-delivery of PI3K/mTOR inhibitor BEZ235 and sorafenib in nanoparticles. Expert Opin Drug Deliv. 2020;17(4):573–587. doi:10.1080/17425247.2020.1730809
156. Roulin D, Waselle L, Dormond-Meuwly A, Dufour M, Demartines N, Dormond O. Targeting renal cell carcinoma with NVP-BEZ235, a dual PI3K/mTOR inhibitor, in combination with sorafenib. Mol Cancer. 2011;10:90. doi:10.1186/1476-4598-10-90
157. Chiou J, Tai C, Huang M, et al. Glucose-regulated protein 78 is a novel contributor to acquisition of resistance to sorafenib in hepatocellular carcinoma. Ann Surg Oncol. 2010;17(2):603–612. doi:10.1245/s10434-009-0718-8
158. Wang W, Li J, Zhu W, et al. MicroRNA-21 and the clinical outcomes of various carcinomas: a systematic review and meta-analysis. BMC Cancer. 2014;14:819. doi:10.1186/1471-2407-14-819
159. Sherif I, Al-Mutabagani L, Sabry D, Elsherbiny N. Antineoplastic activity of chrysin against human hepatocellular carcinoma: new insight on GPC3/SULF2 axis and lncRNA-AF085935 expression. Int J Mol Sci. 2020;21(20):7642. doi:10.3390/ijms21207642
160. Song R, Ma S, Xu J, et al. A novel polypeptide encoded by the circular RNA ZKSCAN1 suppresses HCC via degradation of mTOR. Mol Cancer. 2023;22(1):16. doi:10.1186/s12943-023-01719-9
161. Chang YS, Su CW, Chen SC, Chen YY, Liang YJ, Wu JC. Upregulation of USP22 and ABCC1 during sorafenib treatment of hepatocellular carcinoma contribute to development of resistance. Cells. 2022;11(4):634. doi:10.3390/cells11040634
162. Wu C, Hsiao S, Wu Y. Perspectives on drug repurposing to overcome cancer multidrug resistance mediated by ABCB1 and ABCG2. Drug Resist Updat. 2023;71:101011. doi:10.1016/j.drup.2023.101011
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Potential Treatment Strategies for Hepatocellular Carcinoma Cell Sensitization to Sorafenib
Jiang Z, Dai C
Journal of Hepatocellular Carcinoma 2023, 10:257-266
Published Date: 15 February 2023
Development of Cytolytic Iridium-Complexed Octaarginine Peptide Albumin Nanomedicine for Hepatocellular Carcinoma Treatment
Sun X, Wang D, Chang S, Yin L, Zhang H, Ji S, Fei H, Jin Y
International Journal of Nanomedicine 2025, 20:2395-2409
Published Date: 25 February 2025
Bosentan Delivery via Nano Metal-Organic Framework nanoMIL-89 Restores Vascular Homeostasis in Pulmonary Arterial Hypertension

Al-Badr MA, Abunada HH, Gill R, Fayed HS, Al Haj Zen A, Al-Ghouti MA, Rahman MM, Mohamed NA, Abou-Saleh H
International Journal of Nanomedicine 2025, 20:11045-11060
Published Date: 10 September 2025
Intelligent Nanomedicine Systems Utilizing Diverse Nanoparticles for Osteosarcoma Therapy: A Review
Cai J, Yang H, He Z, Wu C
International Journal of Nanomedicine 2025, 20:14115-14130
Published Date: 25 November 2025
Decoy Nanoparticles and Protein Corona Modulation: A Novel Frontier in Sepsis Treatment
Denisova ER, Volkova DS, Streltsova VV, Kopytov SA, Krivosheev AS, Panfilova EV, Yan N, Kostyushev D, Ganjalikhani-Hakemi M, Pokrovsky VS, Lopus M, Golovin AV, Zamyatnin Jr AA, Celia C, Parodi A
International Journal of Nanomedicine 2026, 21:593748
Published Date: 27 April 2026