Back to Journals » International Journal of Nanomedicine » Volume 21
Nanocarrier-Enabled Melittin Therapy: Mechanistic Advances, Therapeutic Applications, and Translational Challenges
Authors Xiong Y ![]() , Cai H
, Cai H ![]() , Jia S, Gong B, Hou K, Wu H, Chen H
, Jia S, Gong B, Hou K, Wu H, Chen H ![]()
Received 29 January 2026
Accepted for publication 17 May 2026
Published 26 May 2026 Volume 2026:21 599827
DOI https://doi.org/10.2147/IJN.S599827
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Jie Huang
Yuezhihong Xiong,1,* Huili Cai,2,* Siyu Jia,1,2,* Boya Gong,2 Kairong Hou,2 Hongyan Wu,3 Haidan Chen1
1Department of Spine Surgery, Yichang Central People’s Hospital, Yichang, Hubei, People’s Republic of China; 2Department of Hematology, Yichang Central People’s Hospital, Yichang, Hubei, People’s Republic of China; 3College of Basic Medicine, China Three Gorges University, Yichang, Hubei, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Haidan Chen, Department of Spine Surgery, Yichang Central People’s Hospital, No. 183 Yilin Road, Yichang, Hubei, 443003, People’s Republic of China, Email [email protected] Hongyan Wu, College of Basic Medicine, China Three Gorges University, No. 8 Daxue Road, Xiling District, Yichang, Hubei, 443002, People’s Republic of China, Email [email protected]
Abstract: Melittin, a 26-amino-acid amphipathic peptide derived from bee venom, has attracted sustained interest in nanomedicine because of its broad anticancer, antiviral, and anti-inflammatory activities. These effects arise not only from direct membrane disruption, but also from secondary intracellular stress signaling and context-dependent immunomodulatory responses. The same mechanism, however, also underlies melittin’s major limitation: potent biological activity is closely coupled to hemolysis, nonspecific cytotoxicity, and poor systemic tolerability. For this reason, the central question in melittin nanomedicine is no longer whether the peptide is bioactive, but how that activity can be spatially, temporally, and pharmacologically constrained in vivo. Nanocarrier engineering has therefore become integral to melittin development, with lipidic, polymeric, inorganic, and hybrid systems being explored to reduce premature toxicity, improve lesion-site accumulation, regulate peptide release, and enable active or stimulus-responsive targeting. In this review, we synthesize current progress from three linked perspectives: the mechanistic basis of melittin action, the disease-specific logic of its reported applications, and the translational criteria that distinguish proof-of-concept platforms from development-relevant ones. This review was prepared by searching peer-reviewed literature published primarily between 2020 and early 2026 in PubMed, Web of Science, and Scopus, with priority given to studies addressing melittin biological activity, nanocarrier formulation design, toxicity modulation, and translational considerations. Across the current literature, the most decisive issues are not efficacy alone, but the extent to which toxicity attenuation, exposure control, PK/PD interpretability, and formulation tractability can be achieved within the same platform. The available evidence suggests that melittin is most plausibly advanced in delivery-controlled settings, particularly localized, route-constrained, or selected combination-oriented applications, whereas broader systemic use still requires stronger evidence in long-term safety, pharmacological standardization, and product-level reproducibility.
Keywords: nanocarriers, melittin, multi-target therapy, clinical translation, toxicity modulation
Introduction
Melittin is the major bioactive peptide component of bee venom and consists of 26 amino acids arranged in an amphipathic sequence that readily interacts with lipid membranes.1,2 This structural feature underlies a wide range of reported biological effects, including anticancer, antiviral, antimicrobial, and anti-inflammatory activities.3 At the same time, it also explains why melittin remains difficult to translate: the same membrane activity that enables therapeutic action can also produce hemolysis, nonspecific cytotoxicity, vascular irritation, and a narrow tolerability window after systemic exposure.4,5 In melittin research, potency and liability are therefore mechanistically inseparable rather than sequentially distinct problems.
Compared with other well-characterized antimicrobial peptides such as defensins and magainins, melittin is distinguished by a short linear amphipathic α-helical architecture and pronounced cationic membrane affinity, which support rapid membrane insertion and pore-forming or membrane-destabilizing activity.2,6 These structural and biophysical properties help explain why melittin has been investigated beyond antimicrobial use, including anticancer, antiviral, and anti-inflammatory contexts, but they also intensify the need for delivery systems that restrict nonspecific membrane injury.3,7
The therapeutic interest in melittin persists because it can act across disease settings that differ markedly in biology yet share one common feature: membrane vulnerability or membrane-dependent signaling remains relevant to pathology. In cancer, melittin can directly injure tumor cells, induce secondary stress responses, and in some settings promote immunogenic or microenvironmental changes that reshape antitumor immunity.8,9 In antiviral applications, its most consistent activity arises at structurally accessible stages of infection, particularly where virion integrity or early entry processes remain susceptible to peptide-mediated disruption.10,11 In inflammatory disease, melittin has been shown to modulate macrophage- and cytokine-associated pathways that contribute to tissue injury.7 These observations explain why melittin has continued to attract attention despite its formulation challenges, but they do not by themselves resolve the central translational problem: therapeutic plausibility depends on whether these effects can be expressed selectively in pathological tissue rather than indiscriminately across normal blood and organs.12
Nanocarrier engineering has therefore become central to the field rather than adjunct to it. Liposomes, micelles, lipid disks, lipid-coated polymeric particles, inorganic or magnetic carriers, hydrogels, peptide-based assemblies, and multifunctional bioinspired systems have all been used to alter how melittin is loaded, shielded, distributed, activated, or released.13,14 In this context, delivery design does more than improve convenience or stability; it determines whether membrane-active exposure occurs in circulation, at lesion sites, or after intracellular entry, and thus whether melittin behaves as a broadly lytic toxin or as a controllable therapeutic agent.2,5 The significance of nanocarriers in melittin research should therefore be understood at the mechanistic level as well as at the formulation level.
Much of the literature has emphasized individual formulation successes, often by showing reduced hemolysis, increased uptake, improved retention, or stronger biological efficacy in selected models.15–17 However, the field now requires a more integrated framework for comparison. The key questions are not simply which platforms produce biological effects, but which ones do so in a manner that is pharmacologically interpretable and developmentally credible. In practical terms, four issues are especially important across the literature: whether a platform attenuates toxicity without abolishing activity, whether it restricts exposure to relevant tissues or cell populations, whether it provides a controllable and interpretable pharmacological profile, and whether its composition and behavior are sufficiently defined to support product-oriented development. These questions connect mechanistic understanding to disease application and ultimately to translational prioritization.18–20
To define the scope of this review, peer-reviewed studies published primarily between 2020 and early 2026 were searched in PubMed, Web of Science, and Scopus, with emphasis on melittin biological activity, nanocarrier-enabled delivery, toxicity modulation, PK/PD considerations, CMC issues, and translational readiness. Accordingly, this review examines melittin nanomedicine through three linked levels of analysis. First, we revisit the mechanistic basis of melittin action, beginning with membrane disruption and extending to intracellular stress and microenvironmental regulation. Second, we evaluate the current literature across anticancer, antiviral, and anti-inflammatory applications by asking how route, selectivity, and delivery control shape disease-specific plausibility. Third, we compare major nanocarrier classes from a translational perspective, focusing on formulation clarity, PK/PD interpretability, long-term safety maturity, and manufacturing burden. With this structure, this review aims to clarify how the mechanistic activity of melittin can be controllably regulated through delivery design and which carrier classes currently hold the most promising prospects for clinical translation.
Mechanistic Basis of Melittin and the Role of Nanocarriers
Melittin should not be understood as a single-function cytolytic peptide. Its biological effects are better viewed as a cascade that begins with membrane interaction and extends into secondary intracellular responses and broader changes in inflammatory or immune signaling.7,13 This sequence is important because it explains both the therapeutic versatility of melittin and the difficulty of controlling it in vivo. Primary membrane disruption provides the initiating event, but the downstream consequence depends strongly on cell type, tissue context, exposure level, and whether delivery systems restrict when and where active peptide is released.6,21 The mechanistic basis of melittin activity and the delivery-control functions of major nanocarrier classes are summarized in Figure 1.
 |
Figure 1 Mechanistic basis of melittin activity and nanocarrier-enabled delivery control. Melittin activity involves membrane disruption, intracellular stress responses, inflammatory signaling regulation, macrophage polarization, and tumor microenvironment remodeling. Nanocarriers can reduce premature membrane interaction, attenuate nonspecific toxicity, improve lesion-site accumulation, and enable controlled or stimulus-responsive release. |
Primary Membrane-Disruptive Activity
The first and most fundamental event in melittin action is its direct interaction with lipid bilayers.2 Owing to its amphipathic and cationic structure, melittin can associate with cell membranes, insert into the phospholipid bilayer, and assemble into pore-forming or membrane-destabilizing structures.22 At sufficiently high local concentrations, this process leads to loss of membrane integrity, ion imbalance, leakage of intracellular components, and cell death.8 In cancer and antiviral studies, this membrane-active behavior is often the most immediate source of efficacy; in toxicity studies, it is also the clearest explanation for hemolysis and nonspecific injury.5,10 The key implication is that therapeutic use depends not on suppressing membrane activity altogether, but on restricting where and when that activity is expressed.
This point is especially important because membrane disruption is only partly disease-selective in its native form.23 Tumor cells, infected cells, activated immune cells, and erythrocytes can all be affected if exposure is not spatially controlled.24 As a result, free melittin often shows strong bioactivity in vitro but a limited safety margin in vivo.4 Many nanocarrier strategies can be understood as attempts to solve precisely this problem by shielding the peptide during circulation, delaying membrane-active exposure until lesion sites are reached, or confining interaction to cells defined by route, ligand recognition, or microenvironmental triggers.25,26 In mechanistic terms, nanocarriers are therefore not merely passive containers for melittin; they are regulators of the initiating event that determines whether downstream pharmacology becomes therapeutically useful or systemically harmful.
Secondary Intracellular Responses
Although membrane disruption initiates melittin activity, many reported effects cannot be explained by lysis alone.13 Once membrane permeability is altered, cells may undergo a broader range of intracellular responses that include calcium dysregulation,27 mitochondrial dysfunction, oxidative stress, caspase activation, and changes in survival-related signaling pathways.28–30 Depending on exposure conditions and cell context, these events may culminate in apoptosis, necrosis, immunogenic stress, or mixed death phenotypes.31,32 In tumor cells, such secondary responses help explain why melittin can remain pharmacologically effective even when membrane damage is only partial or transient.
These downstream effects are also important because they create the bridge between direct cytotoxicity and disease-specific therapeutic logic. In oncology, intracellular stress and apoptosis-related cascades support the use of melittin not only as a lytic peptide but also as a trigger of immunogenic or combination-sensitive tumor killing.33,34 In antiviral studies, membrane perturbation may be followed by inhibition of entry-related processes or other early infection events.35 In inflammatory settings, intracellular signaling changes in activated macrophages or related effector cells can reduce the production of proinflammatory mediators without requiring unrestricted tissue damage.36 Seen in this way, melittin’s therapeutic profile is not determined by membrane activity alone, but by how that activity is converted into interpretable intracellular consequences. This is one reason why delivery systems that control local concentration, uptake, or intracellular release can materially reshape melittin pharmacology even when the peptide sequence itself remains unchanged.
Immunomodulatory and Microenvironmental Regulation
Beyond direct membrane and intracellular effects, melittin can also act as a regulator of the surrounding biological environment.37 Reported studies have shown that melittin or melittin-based formulations can influence inflammatory cytokines, dendritic cell maturation, macrophage polarization, and the immune composition of tumor or inflamed tissue.38,39 In some cancer platforms, these effects are linked to features consistent with immunogenic cell death, antigen presentation, or relief of local immunosuppression.32,40 In inflammatory disease, the same peptide has been associated with downregulation of TNF-α, IL-1β, IL-6, NO, or related mediators in activated macrophages and injured tissue.41,42 These observations indicate that melittin should be understood not only as a membrane-lytic peptide, but also as a context-dependent modulator of microenvironmental and immune processes that shape therapeutic outcome.
This microenvironmental regulatory activity is critical for understanding melittin’s therapeutic value. Anticancer efficacy is not explained solely by tumor-cell lysis if treatment also changes dendritic-cell activation or suppressive immune populations. Antiviral plausibility is not simply a matter of in vitro virucidal activity if tolerability depends on restricting exposure to infection-relevant compartments. Anti-inflammatory benefit is not reducible to general cytotoxicity if the dominant effect is reduction of activated macrophage signaling in a route-defined tissue context. In each case, the same mechanistic breadth that makes melittin attractive also makes indiscriminate exposure harder to justify. Thus, the more melittin is recognized as a modulator of microenvironmental biology, the more necessary delivery design becomes for separating useful immunopharmacology from unwanted tissue injury.
Mechanistic Significance of Nanocarrier-Enabled Delivery
For melittin, delivery design is inseparable from mechanism. Encapsulation, conjugation, surface engineering, and route-specific administration do more than improve formulation stability; they reshape when membrane-active exposure occurs, how much peptide reaches pathological tissue, whether intracellular delivery is favored, and whether secondary immunological effects emerge in a restricted rather than systemic manner.12,43,44 Lipidic, polymeric, inorganic, and hybrid carriers may achieve this through passive retention, receptor-mediated targeting, trigger-responsive release, or local retention after topical, intratumoral, lymphatic, or transdermal administration.14,17,31 The resulting biological profile of melittin in vivo is therefore determined as much by carrier behavior as by peptide potency itself.
Once delivery is recognized as a mechanistic regulator rather than a formulation afterthought, differences among disease settings become easier to understand. Some applications benefit from route-defined exposure and therefore require less structural complexity. Others depend on highly selective or responsive delivery to make melittin usable at all. Likewise, translational comparison among carrier classes can no longer be based only on whether they produce efficacy, but on whether they provide a credible degree of control over the mechanisms outlined above. Melittin’s clinical feasibility lies not in avoiding its membrane activity, but in controlling it via delivery design.
Applications of Melittin
Anticancer Activity
Oncology remains the setting in which melittin has been explored most extensively, largely because membrane vulnerability, apoptotic susceptibility, and microenvironmental dysregulation create several potential points of intervention at once.9,13,14,21 The central challenge is not whether melittin can damage tumor cells, but whether that activity can be directed toward malignant tissue without reproducing the same membrane injury in blood and healthy organs.4,5,12 As summarized in Table 1, currently reported anticancer studies span free melittin, targeted micelles, lymphatic delivery systems, pH-responsive mesoporous silica platforms, bioinspired multicomponent nanovehicles, and other formulation classes, with evidence distributed across orthotopic, xenograft, metastatic, and in vitro-resistant tumor models.32,37,43,45–51
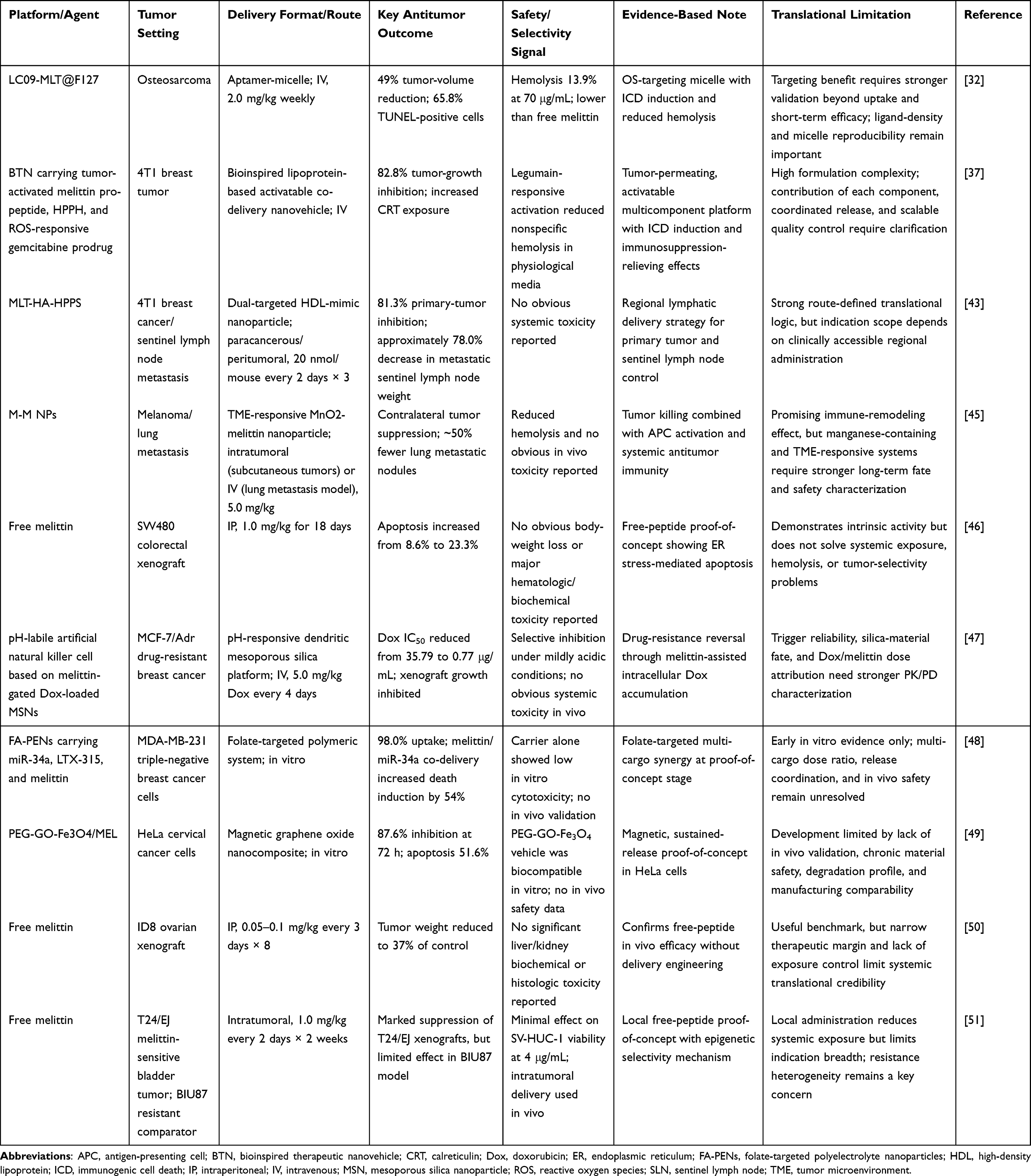 |
Table 1 Nanocarrier-Enabled Melittin Strategies for Cancer Therapy |
Table 1 supports a consistent pattern across these systems: antitumor activity is relatively easy to demonstrate, whereas selectivity, exposure control, and formulation tractability are harder to achieve simultaneously. Targeted platforms such as LC09-MLT@F127 and MLT-HA-HPPS show how lesion-oriented delivery can preserve melittin activity while improving hemocompatibility or regional tolerability, particularly in osteosarcoma and sentinel lymph node–associated disease settings.32,43 Multicomponent systems, including the bioinspired BTN platform and the pH-labile artificial natural killer cell construct, further illustrate that melittin can be integrated into activatable or resistance-reversal strategies with strong biological effect, but at the cost of greater formulation complexity.37,47 By contrast, free-melittin studies in colorectal, ovarian, and bladder tumor models remain useful as benchmarks for intrinsic peptide activity and mechanistic plausibility, yet they also underscore that efficacy without delivery engineering does not by itself resolve the exposure problem.46,50,51 Taken together, the evidence in Table 1 suggests that the most credible oncologic role of melittin lies in tumor-directed, delivery-controlled, or combination-oriented strategies rather than unrestricted systemic use.12,32,37,43,45–51
Antiviral Activity
Antiviral applications of melittin are conceptually attractive for a different reason from oncology.10,35,52 Rather than relying on a single intracellular oncogenic pathway, antiviral activity can arise from structurally vulnerable stages of infection, particularly where virion integrity, envelope stability, or early attachment processes remain accessible to peptide intervention.10,35,52 As summarized in Table 2, the current antiviral literature spans herpes simplex virus, SARS-CoV-2, HIV-1, vesicular stomatitis virus, and enteroviruses, and is dominated by free-peptide studies, with only limited evidence from formulation-based systems such as the SIT-MEL nanoformula.35,52–56 Across these reports, the most consistent signal is not broad post-entry suppression, but direct virucidal or early-stage interference under controlled in vitro conditions.35,52,53,55,56
 |
Table 2 Antiviral Activities of Melittin and Melittin-Based Formulations |
Table 2 supports a clear hierarchy within the available antiviral evidence.35,52–56 The strongest and most reproducible findings concern direct inactivation of viral particles or interference with viral attachment, as illustrated by the anti-HSV studies and the broad virucidal effects reported against VSV and EV-71.35,52 For SARS-CoV-2, both free melittin and the SIT-MEL nanoformula show measurable in vitro activity, but these data still primarily establish proof of concept rather than development-ready antiviral positioning.53,54 The literature on HIV-1 further suggests that melittin-related antiviral effects may extend beyond simple membrane disruption to suppression of viral gene expression, yet these observations also remain preclinical.55,56 Taken together, the antiviral evidence indicates that melittin is best understood at present as a potent but exposure-sensitive antiviral candidate whose most plausible applications are likely to be localized, exposure-limited, or adjunctive rather than broadly systemic.10,35,52–56
Anti-Inflammatory Activity
In inflammatory disease, melittin is most relevant where pathology is spatially concentrated and immune dysregulation can be modulated without requiring broad systemic exposure.7,36,57–60 This is why the anti-inflammatory literature tends to favor topical, regional, organ-directed, or otherwise route-defined interventions rather than unrestricted delivery.36,39,57–60 As summarized in Table 3, currently reported evidence spans gastrointestinal inflammation, acute liver injury, wound-associated inflammation, allergic skin disease, vascular inflammatory remodeling, arthritis, lumbar spinal stenosis, and cerebral ischemia-associated neuroinflammation.28,29,36,38,41,42,57–60 Across these settings, both free melittin and formulation-based systems have shown the ability to reduce inflammatory mediators, limit tissue injury, and improve functional or histologic outcomes, but the most interpretable studies are generally those in which route of administration itself helps define tissue selectivity.28,29,36,39,41,42,57–60
 |
Table 3 Melittin-Based Strategies for Inflammatory Diseases |
Table 3 supports a consistent distinction within the anti-inflammatory literature.7,28,29,36,38,39,41,42,57–60 In localized or organ-confined settings, such as ulcerative colitis, wound repair, allergic dermatitis, and microneedle-assisted arthritis therapy, melittin-based treatment can be linked more directly to reduced cytokine burden, improved histology, and better local tolerability.36,42,57,58,60 By contrast, studies using systemic administration in acute liver failure, lumbar spinal stenosis, cerebral ischemia, or vascular inflammatory remodeling remain valuable, but they depend more heavily on mechanistic interpretation and controlled dosing to separate therapeutic benefit from the intrinsic exposure liability of free melittin.28,29,38,41 A second pattern is that formulation engineering does not simply enhance efficacy; in several models, it also improves interpretability by lowering hemolysis, reducing local irritation, or concentrating activity in inflammatory macrophages or lymphoid compartments.36,39,41,60 Taken together, the evidence in Table 3 indicates that melittin is most plausibly developed for anti-inflammatory use in settings where route, lesion localization, and formulation control can be aligned within the same therapeutic strategy.7,36,39,42,57–60
Translational Considerations for Melittin Nanocarriers
At this stage, the central issue is no longer whether nanocarrier incorporation can improve the apparent in vivo performance of melittin in isolated preclinical models. That has been shown repeatedly across lipidic, polymeric, inorganic, targeted, responsive, and multifunctional systems.16,61–63 The more difficult question is which formulation classes can carry that improvement into a development setting where product definition, reproducibility, long-term safety, pharmacological interpretability, and manufacturing control matter as much as biological activity.18,64 In this sense, translational evaluation of melittin nanomedicine depends less on the number of available platforms than on whether the evidence supporting each platform can be read as product-relevant rather than proof-of-concept alone.20,65 A stepwise translational evaluation framework for development-relevant melittin nanocarriers is presented in Figure 2.
 |
Figure 2 Stepwise translational evaluation framework for melittin nanocarriers. The framework summarizes key development criteria for melittin nanocarriers, including preclinical improvement, translational interpretability, nanoformulation definition, chemistry, manufacturing, and controls (CMC) feasibility, and nonclinical readiness. |
Nonclinical and Preclinical Evidence
Preclinical evidence already supports a clear conclusion: nanocarrier incorporation can materially alter how melittin is distributed, tolerated, and administered in vivo.5,16,39 Across current studies, formulation design has repeatedly been associated with reduced hemolysis, improved lesion-site accumulation, prolonged retention, route-specific delivery, or more favorable short-term tolerability compared with free melittin.12,44,63 At the same time, the field is characterized by marked heterogeneity in model selection, administration route, formulation architecture, and readout depth, which weakens direct comparison across studies and limits how confidently one can rank platforms on the basis of efficacy alone.18,65
This issue is especially relevant for melittin because the same membrane activity that underlies therapeutic benefit also defines the main source of risk.4,23 A formulation that shows tumor suppression, antiviral activity, or attenuation of inflammatory injury is not necessarily the one that offers the clearest development path.5,39,66 The relevant distinction is whether efficacy is accompanied by interpretable exposure control, evidence of tolerability, and a formulation structure that can plausibly support reproducible nonclinical evaluation.63,67 For this reason, the most informative next studies are not simply additional proof-of-concept reports, but better-standardized studies that connect formulation attributes, tissue exposure, and biological effect within the same framework.18,65
Regulatory Grounding for Peptide Nanotherapeutics
Melittin enters development first as a peptide drug substance and only then as part of a nanocarrier product.68,69 Retained biological activity is therefore necessary but insufficient; the formulation must also be definable, reproducible, and interpretable as a product.70,71 This principle becomes especially important in melittin systems because relatively small changes in loading stability, release behavior, or surface structure may alter both efficacy and membrane-associated toxicity.4,12,44 In practical terms, regulatory readiness in this field is inseparable from scientific maturity.18
The current literature already reflects a gradual shift from exploratory carrier design toward more product-oriented thinking.18,19 Simpler lipidic or composition-defined systems often make this transition more readily because their material identity and quality attributes are easier to define.72–74 By contrast, highly layered targeted, responsive, or multifunctional systems may show stronger biological breadth but usually demand more extensive analytical characterization before their preclinical results can be interpreted as development-relevant.64,75,76 Regulatory considerations in melittin nanomedicine should therefore not be viewed as external constraints imposed after discovery, but as part of the criteria by which platform credibility is recognized from the outset.18,71,72
CMC and Manufacturability Considerations
Once melittin nanocarriers are considered as candidate products rather than experimental constructs, CMC becomes part of the core scientific question.70,71 A platform is valuable not only for what it does biologically, but for whether its composition, critical attributes, and manufacturing behavior can be defined with sufficient consistency to support development.18,64 In melittin systems, this matters acutely because changes in formulation behavior may alter both the degree of target exposure and the extent to which hemolytic or nonspecific cytolytic activity remains restrained.4,12
Source and Material Control
The reliability of any melittin nanocarrier begins with the definition of the peptide input itself.68–78 Purity, impurity profile, degradation tendency, and source-related reproducibility all influence downstream loading, release, and batch comparability.79–81 This issue is often understated when the formulation focus dominates the discussion, but for melittin it is particularly relevant because peptide integrity directly affects membrane activity and therefore the biological behavior of the final product.2,4 Source definition should thus be treated as part of the formulation quality framework rather than as a separate upstream consideration.71,82
Particle Reproducibility and Formulation Robustness
Particle reproducibility is the next defining layer of formulation control.72,77 Early-stage reports often describe successful nanoparticle formation through particle size, zeta potential, morphology, or loading efficiency measured in a single prepared batch.12,16,17 These readouts remain important, but translationally the relevant issue is whether they behave as controlled quality attributes rather than descriptive outputs.70,71 This distinction is especially important in melittin systems because small shifts in particle structure or loading stability may change release behavior and safety margins.12,44 The problem is therefore not successful encapsulation per se, but whether those particle features remain stable across batches, media, and preparation conditions.64,77
This burden increases as platform architecture becomes more sophisticated.20,64 Ligand-modified systems require additional control of surface density and binding performance.16,32,75 Responsive systems depend on trigger reliability under biologically realistic conditions.31,47,75 Co-delivery platforms must preserve coordinated loading and release of more than one active component.33,83 These demands do not diminish the scientific value of such systems, but they do mean that formulation robustness, comparability, and process windows become integral to how their biological results should be interpreted.64,70,71
Endotoxin, Sterility, and Injectable Suitability
For parenteral melittin nanocarriers, endotoxin control, sterility assurance, and injectable suitability are not ancillary technical details.72,84,85 They are part of mechanistic credibility. Where cytokine modulation, inflammatory injury, immune activation, or macrophage behavior are central readouts, uncertainty in formulation cleanliness can blur the distinction between peptide effect, carrier effect, and product-related artifact.39,84,86 This is particularly relevant in melittin research, where many of the most interesting biological outcomes overlap with inflammatory and immunological pathways.2,8
Scale-up Feasibility
Scale-up feasibility is where formulation elegance is tested against product reality.64,87 Favorable loading, toxicity attenuation, or lesion-targeting under optimized laboratory conditions does not automatically translate into scalable product behavior.19,20 The relevant question is whether those same features remain intact when manufacturing conditions broaden, batch numbers increase, and control over composition and release must be maintained prospectively rather than retrospectively.70,72 Simpler lipidic, liposomal, or composition-defined systems generally offer a clearer path in this regard, whereas highly layered bioinspired, multifunctional, or hybrid systems impose a heavier development burden as process complexity rises.73,88,89 In melittin formulations, that increase matters because structural inconsistency may directly affect the very toxicity profile that nanocarrier design is meant to improve.4,77
Nonclinical Evaluation: Long-Term Safety and PK/PD Characterization
Nanocarrier engineering has most consistently improved the short-term tolerability profile of melittin in vivo.5,12,60 Reduced hemolysis, preserved body weight, limited acute organ injury, lower local irritation, and improved lesion-site restriction are all repeatedly reported across several formulation classes.12,39,44 These findings are important, but short-term tolerability is not equivalent to nonclinical safety maturity.90,91 For melittin, that distinction is central because the mechanism of benefit and the mechanism of harm remain closely linked.2,4
A development-relevant safety package requires more than acute tolerability signals.91,92 Repeated-dose toxicity, delayed organ injury, recovery-phase reversibility, complement-related effects, vascular compatibility, hypersensitivity risk, and the fate of carrier materials become especially important once melittin is evaluated as a formulation-dependent therapeutic rather than as an isolated proof-of-concept peptide.86,93,94 This issue is less pressing in localized or route-defined applications, where exposure is partly constrained by administration strategy itself, but becomes much more important when the intended use involves intravenous delivery, complex multifunctional carriers, or prolonged circulation.67,73,95
PK/PD characterization remains less mature than formulation innovation.63,67,96 Many studies still infer in vivo behavior from imaging-based biodistribution, lesion accumulation, or stimulus-responsive release-related observations.16,41,43 These approaches are useful for showing that a system reaches the intended site, but they do not by themselves establish a comparable dose–exposure–response framework across platforms.63,69,97 What is often still missing is a direct connection between administered dose, tissue-level exposure, released peptide, and pharmacological consequence.67,98 This gap becomes particularly important when platform classes differ substantially in structural complexity, trigger dependence, or local versus systemic administration, because the same efficacy signal can arise from very different exposure conditions.64,76,83
At this point, additional proof-of-concept studies are less informative than better-standardized ones.18,65 The field stands to gain more from comparable nonclinical evaluation than from continued expansion in platform diversity alone.19,64 The formulations most worth prioritizing are those that can link long-term safety, PK/PD behavior, and therapeutic effect within one coherent framework.63,90,91 That requirement does not exclude targeted, responsive, or multifunctional systems, but it does raise the evidentiary threshold they must meet before they can be regarded as product-relevant rather than conceptually promising.75,76,83
Translational Prioritization and Future Outlook
Not all melittin nanocarriers occupy the same developmental position, even when several produce clear biological effects in preclinical models.5,39,99 What separates them is no longer efficacy alone, but the balance they achieve among formulation simplicity, exposure control, pharmacological interpretability, and manufacturing burden.18,64 As summarized in Table 4, the most development-relevant systems at present are generally those with clearer material architecture and more tractable quality attributes, particularly simple lipid or composition-defined platforms and route-constrained local or regional delivery systems.73,88,95 These classes are not intrinsically superior in all biological settings, but they are more readily interpreted as candidate products rather than as single-study constructs.19,20
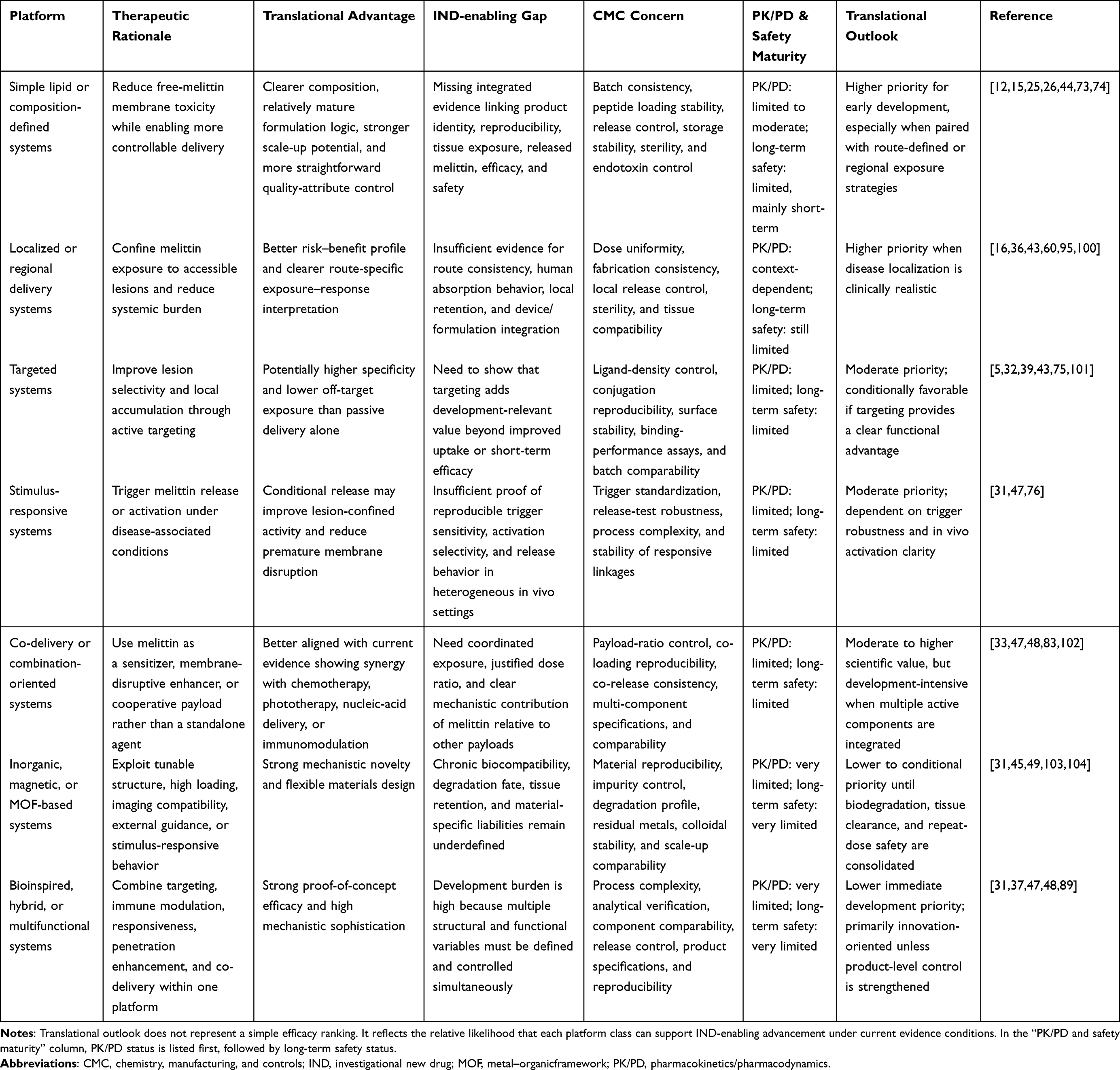 |
Table 4 Translational Outlook of Melittin Nanocarrier Platforms |
Table 4 also clarifies why several more sophisticated platform classes occupy a more conditional position.64,75,76 Targeted systems can improve lesion-site binding or cell-class selectivity, as shown by aptamer-, folate-, estrone-, or integrin-directed melittin carriers, but they also introduce additional demands in ligand control, surface reproducibility, and cross-study comparability.5,32,101 Stimulus-responsive systems, including pH- or microenvironment-activated formulations, are mechanistically attractive because they seek to confine melittin activity to tumor-relevant conditions.31,47,76 In both cases, the central question is not whether the concept is compelling, but whether the added functional sophistication yields a sufficiently clear advantage to justify the extra analytical and manufacturing burden.64,70
A similar distinction applies to co-delivery, inorganic, and bioinspired multifunctional platforms.33,89,100 These systems have generated some of the most conceptually ambitious results in the current literature, including improved intracellular uptake, immune remodeling, photodynamic or photothermal synergy, and resistance-reversal effects.33,47,100 Yet their interpretive burden is correspondingly heavier: multiple active components, layered structures, or hybrid materials make it harder to isolate the contribution of each element, define critical quality attributes, and project scalable product behavior.64,83,104 For that reason, these platforms remain highly informative as innovation engines for the field, but they are not automatically the most immediate translational candidates simply because they are biologically broader.18,19 In melittin nanomedicine, greater functionality often improves mechanistic reach while simultaneously raising the threshold for reproducibility, PK/PD interpretation, and CMC maturity.64,70
Taken together, the comparison in Table 4 suggests a practical near-term principle for the field.18,64 Melittin becomes more clinically credible when its potency is made governable through delivery design rather than merely amplified through structural complexity. In that sense, the most plausible early-entry applications are those in which route, formulation clarity, and exposure restriction can be aligned within the same therapeutic strategy.67,73,95 More elaborate targeted, responsive, and multifunctional systems remain important, but their progression depends on stronger evidence that biological sophistication is matched by product-level control.75,76,83 The near-term future of melittin nanomedicine is therefore likely to favor platforms that simplify interpretation while still restraining peptide toxicity, rather than those that maximize functional layering without equivalent gains in development readiness.18,64
Conclusion
Melittin remains a compelling but intrinsically difficult therapeutic molecule. Its appeal lies in the unusual breadth of its biological activity: the peptide can act directly on membranes, trigger downstream intracellular stress responses, and reshape inflammatory or immune signaling in ways that are relevant across cancer, infection, and inflammatory disease. Yet this same breadth also defines its central limitation. Melittin is not a conventional drug whose efficacy and toxicity can be cleanly separated at the molecular level; rather, its therapeutic effect and its liability arise from the same membrane-active foundation. For that reason, the central problem in melittin nanomedicine is not simply how to enhance activity, but how to govern it.
The literature reviewed here suggests that nanocarrier design should be understood in exactly that sense: not as a secondary formulation refinement, but as the primary means by which melittin becomes pharmacologically usable in vivo. Across diverse platforms, the most meaningful advances have come from restricting premature membrane exposure, improving tissue or cellular selectivity, and creating conditions under which melittin can act within a more interpretable therapeutic window. This is why the field has gradually moved beyond asking whether melittin is active and toward asking what kind of delivery architecture is required to make that activity spatially, temporally, and biologically controllable.
Viewed across application settings, melittin does not occupy a single therapeutic role. In oncology, it is most persuasive when incorporated into delivery-controlled strategies that balance tumoricidal potency with selectivity and tolerability. In antiviral research, the evidence is strongest where structurally accessible stages of infection remain vulnerable to direct peptide action, but broader systemic development remains less mature. In inflammatory disease, the therapeutic logic is often clearest when route of administration, lesion localization, and immune modulation can be aligned within the same intervention. These differences matter because they show that the value of melittin is not fixed; it depends on how disease context interacts with exposure pattern and formulation behavior.
This review also makes clear that preclinical promise is not evenly translatable across all nanocarrier classes. Platforms that appear highly sophisticated biologically are not always those best positioned for development, whereas simpler or more route-defined systems may offer a clearer path because their behavior is easier to reproduce, interpret, and control. The decisive distinctions at this stage therefore lie less in whether a platform can generate efficacy in a favorable model, and more in whether it can support a coherent product logic that includes formulation robustness, PK/PD interpretability, long-term safety relevance, and manageable manufacturing burden. In melittin nanomedicine, translational credibility begins where biological performance and product-level discipline start to converge.
Taken together, the field no longer lacks proof that melittin can be therapeutically useful. What it still lacks, in many cases, is a sufficiently mature framework for deciding when that usefulness is developmentally credible. The most plausible near-term future for melittin nanomedicine is therefore not unlimited expansion into ever more complex systems, but the disciplined advancement of platforms that make melittin’s potency governable, interpretable, and clinically defensible. Under those conditions, melittin is best viewed not as a broadly deployable peptide drug in its native form, but as a high-potential therapeutic component whose real clinical value emerges only when delivery design successfully converts intrinsic activity into controlled pharmacology.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Khalil A, Elesawy BH, Ali TM, Ahmed OM. Bee venom: from venom to drug. Molecules. 2021;26(16):4941. doi:10.3390/molecules26164941
2. Guha S, Ferrie RP, Ghimire J, et al. Applications and evolution of melittin, the quintessential membrane active peptide. Biochem Pharmacol. 2021;193:114769. doi:10.1016/j.bcp.2021.114769
3. Zhang S, Liu Y, Ye Y, et al. Bee venom therapy: potential mechanisms and therapeutic applications. Toxicon. 2018;148:64–21. doi:10.1016/j.toxicon.2018.04.012
4. DeGrado WF, Musso GF, Lieber M, Kaiser ET, Kézdy FJ. Kinetics and mechanism of hemolysis induced by melittin and by a synthetic melittin analogue. Biophys J. 1982;37(1):329–338. doi:10.1016/S0006-3495(82)84681-X
5. Soman NR, Baldwin SL, Hu G, et al. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J Clin Invest. 2009;119(9):2830–2842. doi:10.1172/JCI38842
6. Bechinger B, Lohner K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim Biophys Acta. 2006;1758(9):1529–1539. doi:10.1016/j.bbamem.2006.07.001
7. Memariani H, Memariani M, Moravvej H, Shahidi-Dadras M. Melittin: a venom-derived peptide with promising anti-viral properties. Eur J Clin Microbiol Infect Dis. 2020;39(1):5–17. doi:10.1007/s10096-019-03674-0
8. Zhang H-Q, Sun C, Xu N, Liu W. The current landscape of the antimicrobial peptide melittin and its therapeutic potential. Front Immunol. 2024;15:1326033. doi:10.3389/fimmu.2024.1326033
9. Pandey P, Khan F, Khan MA, Kumar R, Upadhyay TK. An updated review summarizing the anticancer efficacy of melittin from bee venom in several models of human cancers. Nutrients. 2023;15(14):3111. doi:10.3390/nu15143111
10. Chianese A, Zannella C, Palma F, et al. Melittin-related peptides interfere with sandfly fever naples virus infection by interacting with heparan sulphate. Microorganisms. 2023;11(10):2446. doi:10.3390/microorganisms11102446
11. Lee G, Bae H. Anti-inflammatory applications of melittin, a major component of bee venom: detailed mechanism of action and adverse effects. Molecules. 2016;21(5):616. doi:10.3390/molecules21050616
12. Ye R, Zheng Y, Chen Y, et al. Stable loading and delivery of melittin with lipid-coated polymeric nanoparticles for effective tumor therapy with negligible systemic toxicity. ACS Appl Mater Interfaces. 2021;13(47):55902–55912. doi:10.1021/acsami.1c17618
13. Rizkallah J, Charbel N, Yassine A, et al. Melittin-based nanoparticles for cancer therapy: mechanisms, applications, and future perspectives. Pharmaceutics. 2025;17(8):1019. doi:10.3390/pharmaceutics17081019
14. Yu X, Jia S, Yu S, et al. Recent advances in melittin-based nanoparticles for antitumor treatment: from mechanisms to targeted delivery strategies. J Nanobiotechnol. 2023;21(1):454. doi:10.1186/s12951-023-02223-4
15. Mao J, Liu S, Ai M, et al. A novel melittin nano-liposome exerted excellent anti-hepatocellular carcinoma efficacy with better biological safety. J Hematol Oncol. 2017;10(1):71. doi:10.1186/s13045-017-0442-y
16. Yu X, Dai Y, Zhao Y, et al. Melittin-lipid nanoparticles target to lymph nodes and elicit a systemic anti-tumor immune response. Nat Commun. 2020;11(1):1110. doi:10.1038/s41467-020-14906-9
17. Alshememry AK, Kalam MA, Alshehri AA, et al. Enhanced delivery of Melittin via PEGylated PLGA nanoparticles: formulation, characterization, and in vitro evaluation. J Drug Deliv Sci Technol. 2025;110:107101. doi:10.1016/j.jddst.2025.107101
18. Joyce P, Allen CJ, Alonso MJ, et al. A translational framework to DELIVER nanomedicines to the clinic. Nat Nanotechnol. 2024;19(11):1597–1611. doi:10.1038/s41565-024-01754-7
19. Younis MA, Tawfeek HM, Abdellatif AAH, Abdel-Aleem JA, Harashima H. Clinical translation of nanomedicines: challenges, opportunities, and keys. Adv Drug Deliv Rev. 2022;181:114083. doi:10.1016/j.addr.2021.114083
20. Đorđević S, Gonzalez MM, Conejos-Sánchez I, et al. Current hurdles to the translation of nanomedicines from bench to the clinic. Drug Deliv Transl Res. 2022;12(3):500–525. doi:10.1007/s13346-021-01024-2
21. Li X, Li Z, Meng Y-Q, et al. Melittin kills A549 cells by targeting mitochondria and blocking mitophagy flux. Redox Rep. 2023;28(1):2284517. doi:10.1080/13510002.2023.2284517
22. Klocek G, Schulthess T, Shai Y, Seelig J. Thermodynamics of melittin binding to lipid bilayers. Aggregation and pore formation. Biochemistry. 2009;48(12):2586–2596. doi:10.1021/bi802127h
23. Askari P, Namaei MH, Ghazvini K, Hosseini M. In vitro and in vivo toxicity and antibacterial efficacy of melittin against clinical extensively drug-resistant bacteria. BMC Pharmacol Toxicol. 2021;22(1):42. doi:10.1186/s40360-021-00503-z
24. Kim H, Hong JY, Jeon W-J, Baek SH, Ha I-H. Bee venom melittin protects against cisplatin-induced acute kidney injury in mice via the regulation of M2 macrophage activation. Toxins. 2020;12(9):574. doi:10.3390/toxins12090574
25. Zetterberg MM, Reijmar K, Pränting M, Engström Å, Andersson DI, Edwards K. PEG-stabilized lipid disks as carriers for amphiphilic antimicrobial peptides. J Control Release. 2011;156(3):323–328. doi:10.1016/j.jconrel.2011.08.029
26. Gao J, Xie C, Zhang M, et al. RGD-modified lipid disks as drug carriers for tumor targeted drug delivery. Nanoscale. 2016;8(13):7209–7216. doi:10.1039/C5NR05577F
27. Mau SE, Vilhardt H. Cross talk between substance P and melittin-activated cellular signaling pathways in rat lactotroph-enriched cell cultures. J Neurochem. 1997;69(2):762–772. doi:10.1046/j.1471-4159.1997.69020762.x
28. Fan X-G, Pei S-Y, Zhou D, et al. Melittin ameliorates inflammation in mouse acute liver failure via inhibition of PKM2-mediated Warburg effect. Acta Pharmacol Sin. 2021;42(8):1256–1266. doi:10.1038/s41401-020-00516-0
29. Xing X, Zhang X, Fan J, et al. Neuroprotective effects of melittin against cerebral ischemia and inflammatory injury via upregulation of MCPIP1 to Suppress NF-κB activation In Vivo and In Vitro. Neurochem Res. 2024;49(2):348–362. doi:10.1007/s11064-023-04030-7
30. Wan L, Zhang D, Zhang J, Ren L. TT-1, an analog of melittin, triggers apoptosis in human thyroid cancer TT cells via regulating caspase, Bcl-2 and Bax. Oncol Lett. 2018;15(1):1271–1278. doi:10.3892/ol.2017.7366
31. Zhang D, Wang T, Zhang X, et al. Synchronously delivering melittin and evoking ferroptosis via tumor microenvironment-triggered self-destructive metal–organic frameworks to boost cancer immunotherapy. Adv Healthc Mater. 2025;14(11):e2500003. doi:10.1002/adhm.202500003
32. Jia S, Chen Y, Zhuo C, et al. Aptamer-modified melittin micelles efficiently inhibit osteosarcoma deterioration by inducing immunogenic cell death. Colloids Surf B Biointerfaces. 2025;249:114512. doi:10.1016/j.colsurfb.2025.114512
33. Liu H, Hu Y, Sun Y, et al. Co-delivery of bee venom melittin and a photosensitizer with an organic–inorganic hybrid nanocarrier for photodynamic therapy and immunotherapy. ACS Nano. 2019;13(11):12638–12652. doi:10.1021/acsnano.9b04181
34. Kong G-M, Tao W-H, Diao Y-L, et al. Melittin induces human gastric cancer cell apoptosis via activation of mitochondrial pathway. World J Gastroenterol. 2016;22(11):3186–3195. doi:10.3748/wjg.v22.i11.3186
35. Praphawilai P, Kaewkod T, Suriyaprom S, Panya A, Disayathanoowat T, Tragoolpua Y. Anti-herpes simplex virus and anti-inflammatory activities of the melittin peptides derived from Apis mellifera and apis florea venom. Insects. 2024;15(2):109. doi:10.3390/insects15020109
36. Wang J, Zhao F, Zhang Y, et al. Melittin-loaded nanoparticle microneedles targeting M1 macrophage for arthritis treatment. Mol Pharm. 2025;22(7):3679–3692. doi:10.1021/acs.molpharmaceut.4c01383
37. Wang H, Li J, Wang Z, et al. Tumor-permeated bioinspired theranostic nanovehicle remodels tumor immunosuppression for cancer therapy. Biomaterials. 2021;269:120609. doi:10.1016/j.biomaterials.2020.120609
38. Kim H, Hong JY, Jeon W-J, Lee J, Lee YJ, Ha I-H. Melittin regulates iron homeostasis and mediates macrophage polarization in rats with lumbar spinal stenosis. Biomed Pharmacother. 2022;156:113776. doi:10.1016/j.biopha.2022.113776
39. Zheng Y, Ye N, Yang Y, et al. Targeted counteracting of overactive macrophages by melittin stable-loaded solid lipid nanoparticles alleviates cytokine storm and acute inflammatory injury. Biomed Pharmacother. 2024;179:117371. doi:10.1016/j.biopha.2024.117371
40. Shen W, Li Y, Yang Z, et al. Tumor microenvironment reprogramming combined with immunogenic enhancement by nanoemulsions potentiates immunotherapy. J Nanobiotechnol. 2024;22(1):154. doi:10.1186/s12951-024-02401-y
41. Vu HD, Huynh PT, Ryu J, et al. Melittin-loaded iron oxide nanoparticles prevent intracranial arterial dolichoectasia development through inhibition of macrophage-mediated inflammation. Int J Biol Sci. 2021;17(14):3818–3836. doi:10.7150/ijbs.60588
42. Ahmedy OA, Ibrahim SM, Salem HH, Kandil EA. Antiulcerogenic effect of melittin via mitigating TLR4/TRAF6 mediated NF-κB and p38MAPK pathways in acetic acid-induced ulcerative colitis in mice. Chem Biol Interact. 2020;331:109276. doi:10.1016/j.cbi.2020.109276
43. Dai Y, Yu X, Leng Y, et al. Effective treatment of metastatic sentinel lymph nodes by dual-targeting melittin nanoparticles. J Nanobiotechnol. 2023;21(1):245. doi:10.1186/s12951-023-02026-7
44. Li Y, Ruan S, Wang Z, Feng N, Zhang Y. Hyaluronic acid coating reduces the leakage of melittin encapsulated in liposomes and increases targeted delivery to melanoma cells. Pharmaceutics. 2021;13(8):1235. doi:10.3390/pharmaceutics13081235
45. Tang S, Zhou L, He H, et al. MnO2-melittin nanoparticles serve as an effective anti-tumor immunotherapy by enhancing systemic immune response. Biomaterials. 2022;288:121706. doi:10.1016/j.biomaterials.2022.121706
46. Luo Y, Xu C-M, Luo B, Liang G, Zhang Q. Melittin treatment prevents colorectal cancer from progressing in mice through ER stress-mediated apoptosis. J Pharm Pharmacol. 2023;75(5):645–654. doi:10.1093/jpp/rgad008
47. Li Q, Shi Z, Ou M, et al. pH-labile artificial natural killer cells for overcoming tumor drug resistance. J Control Release. 2022;352:450–458. doi:10.1016/j.jconrel.2022.10.042
48. Motiei M, Aboutalebi F, Forouzanfar M, Dormiani K, Nasr-Esfahani MH, Mirahmadi-Zare SZ. Smart co-delivery of miR-34a and cytotoxic peptides (LTX-315 and melittin) by chitosan based polyelectrolyte nanocarriers for specific cancer cell death induction. Mater Sci Eng C Mater Biol Appl. 2021;128:112258. doi:10.1016/j.msec.2021.112258
49. Qi J, Chen Y, Xue T, et al. Graphene oxide-based magnetic nanocomposites for the delivery of melittin to cervical cancer HeLa cells. Nanotechnology. 2020;31(6):065102. doi:10.1088/1361-6528/ab5084
50. Wu R, Li N, Huang W, et al. Melittin suppresses ovarian cancer growth by regulating SREBP1-mediated lipid metabolism. Phytomedicine. 2025;137:156367. doi:10.1016/j.phymed.2025.156367
51. Yan R, Dai W, Wu R, Huang H, Shu M. Therapeutic targeting m6A-guided miR-146a-5p signaling contributes to the melittin-induced selective suppression of bladder cancer. Cancer Lett. 2022;534:215615. doi:10.1016/j.canlet.2022.215615
52. Uddin MB, Lee B-H, Nikapitiya C, et al. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J Microbiol. 2016;54(12):853–866. doi:10.1007/s12275-016-6376-1
53. Enayathullah MG, Parekh Y, Banu S, et al. Gramicidin S and melittin: potential anti-viral therapeutic peptides to treat SARS-CoV-2 infection. Sci Rep. 2022;12(1):3446. doi:10.1038/s41598-022-07341-x
54. Al-Rabia MW, Alhakamy NA, Ahmed OAA, et al. Repurposing of sitagliptin- melittin optimized nanoformula against SARS-CoV-2; antiviral screening and molecular docking studies. Pharmaceutics. 2021;13(3):307. doi:10.3390/pharmaceutics13030307
55. Wachinger M, Saermark T, Erfle V. Influence of amphipathic peptides on the HIV-1 production in persistently infected T lymphoma cells. FEBS Lett. 1992;309(3):235–241. doi:10.1016/0014-5793(92)80780-K
56. Brack-Werner R, Erfle V, von Pechmann N, et al. Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J Gen Virol. 1998;79(4):731–740. doi:10.1099/0022-1317-79-4-731
57. Yaghoubi A, Amel Jamehdar S, Reza Akbari Eidgahi M, Ghazvini K. Evaluation of the therapeutic effect of melittin peptide on the ulcerative colitis mouse model. Int Immunopharmacol. 2022;108:108810. doi:10.1016/j.intimp.2022.108810
58. Eid BG, Alhakamy NA, Fahmy UA, et al. Melittin and diclofenac synergistically promote wound healing in a pathway involving TGF-β1. Pharmacol Res. 2022;175:105993. doi:10.1016/j.phrs.2021.105993
59. Pareek A, Mehlawat K, Tripathi K, et al. Melittin as a therapeutic agent for rheumatoid arthritis: mechanistic insights, advanced delivery systems, and future perspectives. Front Immunol. 2024;15:1510693. doi:10.3389/fimmu.2024.1510693
60. Liu Z, Fan Z, Liu J, et al. Melittin-carrying nanoparticle suppress T cell-driven immunity in a murine allergic dermatitis model. Adv Sci. 2023;10(7):e2204184. doi:10.1002/advs.202204184
61. Dhakne P, Pillai M, Mishra S, Chatterjee B, Tekade RK, Sengupta P. Refinement of safety and efficacy of anti-cancer chemotherapeutics by tailoring their site-specific intracellular bioavailability through transporter modulation. Biochim Biophys Acta Rev Cancer. 2023;1878(4):188906. doi:10.1016/j.bbcan.2023.188906
62. Liu M, Wang H, Liu L, Wang B, Sun G. Melittin-MIL-2 fusion protein as a candidate for cancer immunotherapy. J Transl Med. 2016;14(1):155. doi:10.1186/s12967-016-0910-0
63. Kumar M, Kulkarni P, Liu S, Chemuturi N, Shah DK. Nanoparticle biodistribution coefficients: a quantitative approach for understanding the tissue distribution of nanoparticles. Adv Drug Deliv Rev. 2023;194:114708. doi:10.1016/j.addr.2023.114708
64. Zhang X, Chan HW, Shao Z, Wang Q, Chow S, Chow SF. Navigating translational research in nanomedicine: a strategic guide to formulation and manufacturing. Int J Pharm. 2025;671:125202. doi:10.1016/j.ijpharm.2025.125202
65. Leong HS, Butler KS, Brinker CJ, et al. On the issue of transparency and reproducibility in nanomedicine. Nat Nanotechnol. 2019;14(7):629–635. doi:10.1038/s41565-019-0496-9
66. Hood JL, Jallouk AP, Campbell N, Ratner L, Wickline SA. Cytolytic nanoparticles attenuate HIV-1 infectivity. Antivir Ther. 2013;18(1):95–103. doi:10.3851/IMP2346
67. Cabral H, Li J, Miyata K, et al. Controlling the biodistribution and clearance of nanomedicines. Nat Rev Bioeng. 2024;2:214–232.
68. Kuril AK, Saravanan K, Subbappa PK. Analytical considerations for characterization of generic peptide product: a regulatory insight. Anal Biochem. 2024;694:115633. doi:10.1016/j.ab.2024.115633
69. U.S. Food and Drug Administration. Clinical pharmacology considerations for peptide drug products. Guidance for Industry. 2023.
70. Birla D, Khandale N, Bashir B, et al. Application of quality by design in optimization of nanoformulations: principle, perspectives and practices. Drug Deliv Transl Res. 2025;15(3):798–830. doi:10.1007/s13346-024-01681-z
71. International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use. Q8(R2) Pharmaceutical Development; 2009.
72. U.S. Food and Drug Administration. Liposome drug products: chemistry, manufacturing, and controls; human pharmacokinetics and bioavailability; and labeling documentation. Guidance Industry. 2018.
73. Chen J, Hu S, Sun M, et al. Recent advances and clinical translation of liposomal delivery systems in cancer therapy. Eur J Pharm Sci. 2024;193:106688. doi:10.1016/j.ejps.2023.106688
74. Senjab RM, AlSawaftah N, Abuwatfa WH, Husseini GA. Advances in liposomal nanotechnology: from concept to clinics. RSC Pharmaceutics. 2024;1(5):928–948. doi:10.1039/D4PM00176A
75. Veiga N, Diesendruck Y, Peer D. Targeted nanomedicine: lessons learned and future directions. J Control Release. 2023;355:446–457. doi:10.1016/j.jconrel.2023.02.010
76. Ochioni AC, Vieira IRS, Conte-Junior CA. Advances in stimuli-responsive nanoparticles for cancer therapy: mechanisms, functional materials, and emerging technologies. J Drug Deliv Sci Technol. 2025;114:107519. doi:10.1016/j.jddst.2025.107519
77. Mülhopt S, Diabaté S, Dilger M, et al. Characterization of nanoparticle batch-to-batch variability. Nanomaterials. 2018;8(5):311. doi:10.3390/nano8050311
78. Teoh ACL, Ryu K-H, Lee EG. One-step purification of melittin derived from Apis mellifera bee venom. J Microbiol Biotechnol. 2017;27(1):84–91. doi:10.4014/jmb.1608.08042
79. Patel S, Vyas VK, Mehta PJ. A review on forced degradation strategies to establish the stability of therapeutic peptide formulations. Int J Pept Res Ther. 2023;29(2):22. doi:10.1007/s10989-023-10492-8
80. Tabesh A, De Luca C, Faraji Shovey A, et al. Expanding the use of green solvents for the isolation of melittin from honeybee venom. J Chromatogr A. 2025;1763:466460. doi:10.1016/j.chroma.2025.466460
81. Fatahian F, Rezadoost H, Golestan SMJS, et al. Analytical characterization and stability evaluation of high-purity melittin isolated from crude bee venom via one-step, scalable reversed-phase liquid chromatography. J Chromatogr A. 2026;1766:466605. doi:10.1016/j.chroma.2025.466605
82. U.S. Food and Drug Administration. ANDAs for certain highly purified synthetic peptide drug products that refer to listed drugs of rDNA origin. Guidance Industry. 2021.
83. Mu Q, Gong T, Behl M, et al. Translation of combination nanodrugs into nanomedicines: lessons learned and future outlook. J Drug Target. 2018;26(5–6):435–447. doi:10.1080/1061186X.2017.1419363
84. Hannon G, Heaton BJ, Plant-Hately A, et al. Interlaboratory comparison of endotoxin contamination assessment of nanomaterials. Nanoscale. 2024;16(45):21011–21020. doi:10.1039/D4NR02821J
85. Neun BW, Dobrovolskaia MA. Current considerations and practical solutions for overcoming nanoparticle interference with LAL assays and minimizing endotoxin contamination. Methods Mol Biol. 2024;2789:87–99.
86. La-Beck NM, Islam MR, Markiewski MM. Nanoparticle-induced complement activation: implications for cancer nanomedicine. Front Immunol. 2021;11:603039. doi:10.3389/fimmu.2020.603039
87. de Souza Cardoso Delfino C, de Paula Pereira MC, Dos Santos Oliveira M, et al. Scaling nanopharmaceutical production for personalized medicine: challenges and strategies. J Nanopart Res. 2025;27(4):108. doi:10.1007/s11051-025-06293-3
88. Sivadasan D, Ramakrishnan K, Mahendran J, Ranganathan H, Karuppaiah A, Rahman H. Solid lipid nanoparticles: applications and prospects in cancer treatment. Int J Mol Sci. 2023;24(7):6199. doi:10.3390/ijms24076199
89. Aalhate M, Mahajan S, Dhuri A, et al. Biohybrid nano-platforms manifesting effective cancer therapy: fabrication, characterization, challenges and clinical perspective. Adv Colloid Interface Sci. 2025;335:103331. doi:10.1016/j.cis.2024.103331
90. Kumar R, Kumar A, Bhardwaj S, et al. Nanotoxicity unveiled: evaluating exposure risks and assessing the impact of nanoparticles on human health. J Trace Elem Miner. 2025;13:100252. doi:10.1016/j.jtemin.2025.100252
91. Food and Drug Administration, HHS. International conference on harmonisation; guidance on M3(R2) nonclinical safety studies for the conduct of human clinical trials and marketing authorization for pharmaceuticals; availability. Notice. Fed Regist. 2010;75(13):3471–3472.
92. U.S. Food and Drug Administration. Immunogenicity assessment for therapeutic protein products. Guidance Industry. 2014.
93. Moghimi SM, Haroon HB, Yaghmur A, et al. Perspectives on complement and phagocytic cell responses to nanoparticles: from fundamentals to adverse reactions. J Control Release. 2023;356:115–129. doi:10.1016/j.jconrel.2023.02.022
94. Ilinskaya A, Shah A, Van Dusen A, Dobrovolskaia MA. Detection of intracellular complement activation by nanoparticles in human T lymphocytes. Methods Mol Biol. 2024;2789:109–120.
95. Zheng M, Sheng T, Yu J, et al. Microneedle biomedical devices. Nat Rev Bioeng. 2023;2(4):324–342. doi:10.1038/s44222-023-00141-6
96. Paramshetti S, Angolkar M, Talath S, et al. Unravelling the in vivo dynamics of liposomes: insights into biodistribution and cellular membrane interactions. Life Sci. 2024;346:122616. doi:10.1016/j.lfs.2024.122616
97. Fletcher EP, Sahre M, Hon YY, et al. Impact of organ impairment on the pharmacokinetics of therapeutic peptides and proteins. AAPS J. 2023;25(4):54. doi:10.1208/s12248-023-00819-0
98. Peralta-Cuevas E, Degollado-Hernández NY, Martínez-Ortiz IC, et al. How do nanoparticle properties shape pharmacokinetics and pharmacodynamics? A mechanistic review. Front Pharmacol. 2026;16:1704814. doi:10.3389/fphar.2025.1704814
99. Huang C, Jin H, Qian Y, et al. Hybrid melittin cytolytic peptide-driven ultrasmall lipid nanoparticles block melanoma growth in vivo. ACS Nano. 2013;7(7):5791–5800. doi:10.1021/nn400683s
100. Jin H, Zhao G, Hu J, et al. Melittin-containing hybrid peptide hydrogels for enhanced photothermal therapy of glioblastoma. ACS Appl Mater Interfaces. 2017;9(31):25755–25766. doi:10.1021/acsami.7b06431
101. Raveendran R, Chen F, Kent B, Stenzel MH. Estrone-decorated polyion complex micelles for targeted melittin delivery to hormone-responsive breast cancer cells. Biomacromolecules. 2020;21(3):1222–1233. doi:10.1021/acs.biomac.9b01681
102. Hematyar M, Soleimani M, Es-Haghi A, Rezaei Mokarram A. Synergistic co-delivery of doxorubicin and melittin using functionalized magnetic nanoparticles for cancer treatment: loading and in vitro release study by LC–MS/MS. Artif Cells Nanomed Biotechnol. 2018;46(sup3):S1226–S1235. doi:10.1080/21691401.2018.1536063
103. Choi M, Ryu J, Vu HD, et al. Transferrin-conjugated melittin-loaded L-arginine-coated iron oxide nanoparticles for mitigating beta-amyloid pathology of the 5XFAD mouse brain. Int J Mol Sci. 2023;24(19):14954. doi:10.3390/ijms241914954
104. Wiśniewska P, Haponiuk J, Saeb MR, Rabiee N, Bencherif SA. Mitigating Metal-Organic Framework (MOF) toxicity for biomedical applications. Chem Eng J. 2023;471:144400. doi:10.1016/j.cej.2023.144400
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Extracellular Vesicle-Based Therapeutic Cargo Delivery for Cancer Therapy
Huang Z, Cheng J, Deng Z, Liu C, Huang T, Lin W
International Journal of Nanomedicine 2025, 20:13007-13037
Published Date: 27 October 2025