Back to Journals » Journal of Inflammation Research » Volume 14
N-Acetylcysteine as Adjuvant Therapy for COVID-19 – A Perspective on the Current State of the Evidence
Authors Wong KK, Lee SWH ![]() , Kua KP
, Kua KP ![]()
Received 24 February 2021
Accepted for publication 26 May 2021
Published 6 July 2021 Volume 2021:14 Pages 2993—3013
DOI https://doi.org/10.2147/JIR.S306849
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Editor who approved publication: Professor Ning Quan
Kon Ken Wong,1,2 Shaun Wen Huey Lee,3– 6 Kok Pim Kua7
1Department of Microbiology and Immunology, Hospital Canselor Tuanku Muhriz UKM, Cheras, Kuala Lumpur, Malaysia; 2Faculty of Medicine, The National University of Malaysia, Cheras, Kuala Lumpur, Malaysia; 3School of Pharmacy, Monash University, Bandar Sunway, Selangor, Malaysia; 4Asian Centre for Evidence Synthesis in Population, Implementation, and Clinical Outcomes (PICO), Health and Well-being Cluster, Global Asia in the 21st Century (GA21) Platform, Monash University, Bandar Sunway, Selangor, Malaysia; 5Gerontechnology Laboratory, Global Asia in the 21st Century (GA21) Platform, Monash University, Bandar Sunway, Selangor, Malaysia; 6Faculty of Health and Medical Sciences, Taylor’s University, Bandar Sunway, Selangor, Malaysia; 7Puchong Health Clinic, Petaling District Health Office, Ministry of Health Malaysia, Petaling, Selangor, Malaysia
Correspondence: Kok Pim Kua Email [email protected]
Abstract: The looming severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has caused a long-lasting pandemic of coronavirus disease 2019 (COVID-19) around the globe with substantial morbidity and mortality. N-acetylcysteine, being a nutraceutical precursor of an important antioxidant glutathione, can perform several biological functions in mammals and microbes. It has consequently garnered a growing interest as a potential adjunctive therapy for coronavirus disease. Here, we review evidence concerning the effects of N-acetylcysteine in respiratory viral infections based on currently available in vitro, in vivo, and human clinical investigations. The repurposing of a known drug such as N-acetylcysteine may significantly hasten the deployment of a novel approach for COVID-19. Since the drug candidate has already been translated into the clinic for several decades, its established pharmacological properties and safety and side-effect profiles expedite preclinical and clinical assessment for the treatment of COVID-19. In vitro data have depicted that N-acetylcysteine increases antioxidant capacity, interferes with virus replication, and suppresses expression of pro-inflammatory cytokines in cells infected with influenza viruses or respiratory syncytial virus. Furthermore, findings from in vivo studies have displayed that, by virtue of immune modulation and anti-inflammatory mechanism, N-acetylcysteine reduces the mortality rate in influenza-infected mice animal models. The promising in vitro and in vivo results have prompted the initiation of human subject research for the treatment of COVID-19, including severe pneumonia and acute respiratory distress syndrome. Albeit some evidence of benefits has been observed in clinical outcomes of patients, precision nanoparticle design of N-acetylcysteine may allow for greater therapeutic efficacy.
Keywords: N-acetylcysteine, SARS-CoV-2; COVID-19, coronavirus, repurposing approved drugs, engineering nanoparticles, virus infected cells, respiratory viral diseases, antioxidant, glutathione, T lymphocytes, immune modulating activity, anti-inflammatory response, antiviral effect, clinical translation
Introduction
The acute respiratory disease COVID-19 caused by the novel coronavirus, SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2), has spawned a global pandemic with untold morbidity and mortality, accompanied by devastating disruption to all facets of society, economy, and health care system.1,2 SARS-CoV-2 is a single-stranded, positive-sense RNA virus that was initially identified in Wuhan city in China in December 2019 from an outbreak of pneumonia cases in connection with Huanan Seafood Wholesale Market.3 It is closely related to other tremendously pathogenic beta-coronaviruses that have emanated in this century, namely severe acute respiratory syndrome coronavirus 1 (SARS-CoV-1) and Middle East respiratory syndrome coronavirus (MERS-CoV).4 Unlike SARS-CoV-1 and MERS-CoV that exhibit only limited human-to-human transmissions,5 a person being infected with SARS-CoV-2 who is just mildly ill or even asymptomatic can spread the disease to an average of two or three others, resulting in an exponential rate of increase in infection cases. SARS-CoV-2 virus has engendered 10-fold higher in the number of cases than the 2003 SARS epidemic in a quarter of the time.6 The rapid transmission of this highly pathogenic virus has warranted a pressing global need for the instantaneous development and deployment of therapeutic approaches and preventive measures against the disease.7
Common hematological manifestations of COVID-19 infection include lymphocytopenia associated with intensification of the inflammatory process and direct infection of lymphocytes and destruction of lymphoid organs, increased ferritin levels owing to inflammation, and a higher rate of erythrocyte sedimentation in severe disease. Blood group A and males are more likely to become infected. Male sex, older age, and the presence of comorbidities are correlated with increased risk of COVID-19-related mortality.8,9 The development of thrombocytopenia may occur in severe disease due to reduced platelet production and increased destruction or consumption of platelets. Elevated D-dimer levels concomitant with high levels of fibrin degradation products and low antithrombin activity render COVID-19 patients to be at risk of hypercoagulability and thrombotic complications. Neutrophil-to-lymphocyte ratio and plasma D-dimer concentrations are relatively easy to quantify and possess clinical value for disease prognosis.10 High levels of proinflammatory cytokines and chemokines, including interleukin (IL)-6, IL-2, IL-2 receptor, IL-10, tumor necrosis factor alpha (TNF-α), and interferon gamma (IFN-γ) may cause multiorgan damage as well as cardiovascular complications.11 Cardiac arrhythmia has also been reported and is associated with a cytokine storm-triggered systemic hyperinflammatory state and immune response that may cause injury to cardiac monocytes, resulting in myocardial dysfunction and the ensuing development of arrhythmia. Similarly, infection of alveolar pneumocytes cells by SARS-CoV-2 virus triggers the initiation of systemic inflammation and elevated immunoreactivity that potentiate T-cell and macrophage activation infiltrating infected myocardial tissues, leading to cardiovascular damage and myocarditis. Additionally, atherosclerotic plaques can be destabilized by systemic inflammatory response which happens simultaneously with pro-inflammatory and pro-oxidative effects of SARS-CoV-2, thereby giving rise to acute coronary syndrome and ischemic heart disease among COVID-19 patients.12
Previous research with coronaviruses using both in vitro and in vivo experimental designs has contributed to a valuable guiding foundation for elucidating therapeutic strategies for the treatment of COVID-19. On top of understanding the microbial pathogenesis and the molecular and cellular mechanisms of disease biology, activity of a therapeutic agent in a translational research, be it either in vitro or in vivo, is critical to proffer advancement to first-in-human clinical trials based on laboratory findings of pharmacology, toxicology, and immunology.13 For COVID-19 repurposing of an existing clinically approved drug, it is pivotal to demonstrate its antiviral, anti-inflammatory, and related effects against SARS-CoV-2 in cell-based systems in vitro. The assessment of drug potency may be impacted by the type of the virus (eg, full-length wildtype, reporter viruses, or sub-genomic replicons, etc.) and the cell culture utilized (eg, Vero E6, Huh7, FRhK, or human airway epithelial cells), thus, necessitating high quality and standardized cellular assays, or at least with robust and universally accepted control groups.14
Recent randomized controlled trials depict that repurposed antiviral drugs such as remdesivir, lopinavir, and interferon beta-1α regimens have small or null effect on hospitalized patients with COVID-19, as determined by outcomes such as overall mortality, initiation of mechanical ventilation, and duration of hospitalization.15,16 Combination treatment of remdesivir with anti-inflammatory drug baricitinib is associated with shorter time to recovery and accelerated improvement in clinical status, notably among those receiving high-flow oxygen or non-invasive ventilation.17 Treatment with dexamethasone reduces 28-day mortality in patients who are receiving invasive mechanical ventilation or oxygen without invasive mechanical ventilation, however, no discernible benefit and the possibility of causing harm have been found among those who are not receiving respiratory support.18 Neutralizing antibody bamlanivimab results in fewer hospitalizations and a lower symptom burden.19 Safe and effective vaccines that can confer significant protection against COVID-19 infection in real-world settings encompass BNT162b2 [Pfizer-BioNTech],20 mRNA-1273 [Moderna],21 NVX-CoV2373 [Novavax],22 CoV2 preS dTM-AS03 [Sanofi Pasteur-GlaxoSmithKline],23 Ad26.COV2.S [Johnson & Johnson/Janssen],24 and ChAdOx1 nCoV-19 (AZD1222) [Oxford-AstraZeneca].25
The overwhelming impact of the COVID-19 crisis has driven the push for reimagining and repositioning of previously approved medical treatments for other indications to speed up the discovery and development of safe and efficacious agents to enlarge the alternatives for adjunctive treatment or prevention of progression into severe COVID-19 illness.14 From the clinical front, it is presently worrisome to have no effective antimicrobial agents to treat the infected individuals and, optimally, eliminate viral shedding and the ensuing transmission cascades.
N-acetylcysteine is a mucolytic drug which exhibits antioxidant and anti-inflammatory effects.26 The compound has been available in clinical practice for several decades to treat various medical conditions, including bronchitis, acute respiratory distress syndrome, paracetamol intoxication, chemotherapy-related toxicity, doxorubicin cardiotoxicity, heavy metal intoxication, ischemia-reperfusion cardiac injury, human immunodeficiency virus infection or acquired immunodeficiency syndrome, and neuropsychiatric disorders. N-acetylcysteine is also marketed as a dietary supplement that is suggested to possess antioxidant and hepatic-protecting effects. The antioxidant characteristic of N-acetylcysteine has been ascribable to its reactivity with •OH, CO3•−, •NO2, and thiyl radicals, the ability to repair oxidative damaged key cellular molecules, and activity as a precursor for biosynthesis of glutathione.27 There is a growing body of evidence that highlights the intrinsic antimicrobial and antibiofilm activities in many respiratory pathogens, including Escherichia, Pseudomonas, Staphylococcus, Acinetobacter, Haemophilus, and Klebsiella.26,28–30 High concentrations of N-acetylcysteine do not carry the risk of adverse interactions with most commonly used antibiotics and can exert intrinsic antimicrobial activity against Haemophilus influenza.26 Being a precursor for glutathione biosynthesis which is a crucial determinant of antimicrobial activity against bacteria, N-acetylcysteine is often prescribed as a mucolytic agent in conjunction with antibiotic treatment in respiratory tract infections to improve the outcomes of the course of therapy.31
N-acetylcysteine has recently been suggested as an adjunctive therapy to the standard care for SARS-CoV-2 infection considering the favorable risk and benefit ratio and its effects on synthesizing glutathione, improving immune function, and modulating inflammatory response.32–34 It achieves the therapeutic effects through two main activities: 1) mucolytic action conferred by the free sulfhydryl group which reduces disulfide bonds in the cross-linked mucus glycoproteins matrix, thus decreasing the viscosity of mucus; 2) antioxidative action attributable to a direct interaction with free radicals, an indirect effect as a precursor to cysteine which is required for glutathione biosynthesis, and a replenishment of thiol pools that is central to redox regulation and control.35 In light of these properties, we hypothesize that N-acetylcysteine plays a role in the treatment of COVID-19 infection by the following postulated mechanisms of action (Figure 1):
- Envelope (E) and spike (S) proteins have a triple cysteine structural motif located directly after the E protein’s transmembrane domain (NH2- … L-Cys-A-Y-Cys-Cys-N … -COOH) and a similar motif located in the carboxy terminus of the S protein (NH2- … S-Cys-G-S-Cys-Cys-K … -COOH). The position, orientation, and composition of these two motifs may serve as a center for the structural link between the E and S proteins which is mediated by the formation of disulfide bonds between the corresponding cysteine residues.36 Previous studies have indicated that the entry of viral glycoprotein is affected by thiol-disulfide balance within the viral surface and the cell-surface of the host.37,38 Any perturbations in the thiol-disulfide interchange equilibrium would deter the entry of the virus into host cells.39,40 Cleavage of disulfide bridges by N-acetylcysteine disrupts the structural components of the interacting proteins, thereby impairing receptor binding affinity and infectivity.
- N-acetylcysteine is a chemical reducer of disulfide bonds via its free sulfhydryl groups may interact with the extracellular disulfide bridges of angiotensin II receptor, alter its tertiary structure, and inhibit the binding of angiotensin II to its surface receptors (AT1a receptors) with subsequent attenuation of signal transduction and cell action. The AT1a receptors possess two sets of disulfide bridges at the extracellular domain of the receptors: C18-C274 and C101-C180. N-acetylcysteine can reduce the disulfide bonds in a dose-dependent manner,41,42 decreasing angiotensin II and increasing angiotensin 1–7 (a biologically active peptide exerting many opposing actions to angiotensin II), thus protecting against lung inflammation and fibrosis.43
- The sulfhydryl group of N-acetylcysteine inhibits angiotensin converting enzyme, reducing production of angiotensin II.44 In human lungs, angiotensin converting enzyme is expressed in lower lungs on type I and II alveolar epithelial cells. Following infection, viral entry begins with the attachment of spike (S) protein expressed on the viral envelope to angiotensin converting enzyme on the alveolar surface. Hence, N-acetylcysteine may prevent viral entry by limiting viral protein angiotensin converting enzyme interaction and internalization of the receptor-ligand complex.45 It also protects against oxidative stress and prevents glycosylation of proteins which may confer protection against respiratory disease syndrome and lung failure.
- The antioxidant effect of N-acetylcysteine ameliorates oxidative stress and inflammatory response in COVID-19.46 It amplifies the signaling functions of toll-like receptor 7 protein and mitochondrial antiviral-signaling protein for boosting type 1 interferon production.47 Type I interferon functions to induce expression of various interferon-stimulated genes that exert antiviral activities to host cells.48
- The receptor for advanced glycation end products (RAGE) and its ligands have a crucial role in the pathogenesis of COVID-19 pneumonia and acute respiratory distress syndrome as well as lung inflammation. Circulating levels of soluble RAGE (sRAGE, a decoy receptor) are positively associated with acute respiratory distress syndrome severity and mortality risk, whereas reduction in circulating levels of sRAGE drop results in disease resolution.49 Advanced glycation end products are formed by a reaction of the dicarbonyl compounds methylglyoxal and glyoxal with amino acids in proteins during glycolysis. Methylglyoxal and methylglyoxal-derived AGE can further activate inflammatory cells by binding to RAGE.50 N-acetylcysteine induces endogenous glutathione and hydrogen sulfide synthesis, thus attenuating methylglyoxal-induced protein glycation and additional glycosylation events in SARS-CoV-2 which may then inhibit the virus’s infectivity and associated pathologies.51
- N-acetylcysteine inhibits NF-κB activation by suppressing TNF-induced IκB kinases, followed by impediment of proteasome-dependent degradation.52 This prevents translocation of NF-κB from cytoplasm to the nucleus and block expression of pro-inflammatory cytokines and chemokines which have been correlated with severity and lethality in various acute respiratory viral infections, including Influenza A H5N1, highly pathogenic H1N1, SARS-CoV, MERS-CoV, and SARS-CoV-2.53
 |
Figure 1 Schematic representation of the possible effects of N-acetylcysteine on SARS-CoV-2 infection. N-acetylcysteine may inhibit envelope (E) protein and spike (S) protein of the virus, decrease angiotensin II receptor binding (AT2R), inhibit angiotensin converting enzyme (ACE), induce endogenous glutathione (GSH) synthesis which is associated with increased antioxidant effect and decreased glycation of intracellular proteins, and prevent nuclear translocation of NF-κB which suppresses the production of pro-inflammatory mediators and cytokines. |
Experimental in vitro Models
In vitro studies show that N-acetylcysteine holds therapeutic promise in numerous viral respiratory tract infections.
Influenza Viruses
In H5N1-infected A549 cells, N-acetylcysteine was found to attenuate H5N1-induced cytopathogenic effects, apoptosis, and virus yields. In addition, it decreased pro-inflammatory cytokine levels of CXCL8, CXCL10, CCL5, and interleukin-6. It was posited that the antiviral and anti-inflammatory effects of N-acetylcysteine were mediated by the inhibition of the oxidative metabolic pathway including transcription factor NF-κB and mitogen activated protein kinase p38.54 In another study, N-acetylcysteine was demonstrated to have immune‐modulating properties by increasing influenza virus-specific cytotoxic T‐lymphocyte clones and interferon-γ generation.55
In an in vitro A549 model infected with influenza (strains A and B), N-acetylcysteine was shown to inhibit expression and release of MUC5AC, IL6, and TNF-α, reduce intracellular hydrogen peroxide level, restore intracellular pool of total thiols, diminish NF-κB translocation to the cell nucleus, attenuate activation of mitogen activated protein kinase p38, and prevent replication of the viruses.56 N-acetylcysteine also prevented apoptosis in H1N1-infected cell lines.57
In Vero cells (ATCC CCL-81) infected with porcine H1N1 strain, N-acetylcysteine was reported to exert a dose-dependent inhibition on virus replication. However, the inhibitory effect of N-acetylcysteine was significantly less than that observed with H5N1, suggesting the susceptibility is strain-dependent.58
Respiratory Syncytial Virus
In in vitro modeling of RSV infection carried out in cultures of primary normal human bronchial epithelial cells, N-acetylcysteine significantly inhibited viral infection, re-established the number of beating cells, restored the virus-induced decrease in expression of β-tubulin and genes involved in ciliagenesis such as DNAI2 and FOXJ1, inhibited IL-13, MUC5AC, and GOB5 upregulation, inhibited ICAM1 expression, increased heme-oxygenase 1 expression which correlated with the restoration of cellular antioxidant capacity, and enhanced intracellular hydrogen peroxide and glutathione concentrations.59
In another experiment, N-acetylcysteine significantly reduced MUC5AC expression in RSV-infected A549 cells. It also diminished induction of TNF-α and IL-6, decreased NF-κB nuclear translocation and p38 mitogen activated protein kinase phosphorylation, abolished intracellular hydrogen peroxide production, restored intracellular thiol levels, and reduced virus titre.56
Likewise, RSV infection of A549 epithelial cell lines showed the inhibitory effects of N-acetylcysteine on virus-induced chemokine expression and DNA-binding activity of NF-κB subunits of p50 and p65.60
Experimental in vivo Models
In vivo studies depict some beneficial effects of N-acetylcysteine in numerous viral respiratory tract infections.
Influenza Viruses
In mice following intranasal infection with a lethal dose of influenza A virus A/PR/8, the use of N-acetylcysteine increased survival rate from 58% (ribavirin monotherapy) to 92% (N-acetylcysteine and ribavirin), suggesting the antioxidant’s role in improving host defense mechanism and preventing pathogenesis of lung inflammation.61,62 Further experiment depicted that survival rates of mice were 20% in the N-acetylcysteine group, 60% in the oseltamivir group, and 100% in combination treatment of N-acetylcysteine and oseltamivir.63 In these three models, the mice were infected with 2–3 LD50 of influenza virus and N-acetylcysteine was administered as a single daily dose of 1,000 mg/kg.61–63 N-acetylcysteine increased survival by contributing to host defense mechanisms or by direct antioxidant effect against oxidative stress associated with viral infection.63 On the contrary, mice infected intranasally with a deadly dose of porcine A/swine/Iowa/4 (H1N1) influenza strain showed no significant difference in percent survival and mean survival time between N-acetylcysteine and control groups, showing that different virus strains resulted in different susceptibilities to N-acetylcysteine.58 The porcine A/swine/Iowa/4 (H1N1) strain was found to be more resistant to N-acetylcysteine than the human strain A/PR/8 (H1N1) deployed in other animal models.61–63 The lack of treatment effect was possibly due to the virus phenotypic susceptibility to N-acetylcysteine and the viral pathotype.64
Findings from H9N2 swine influenza virus-infected murine model indicated that N-acetylcysteine ameliorated pulmonary edema and inflammatory response, reduced myeloperoxidase activity in the lungs, decreased total and differential cell counts, neutrophils, macrophages, IL-6, IL-1β, TNF-α, and CXCL-10 in the bronchoalveolar lavage fluid, and inhibited protein expression of TLR4 and its mRNA in the lungs.65
In an experiment of H3N2-infected mice, N-acetylcysteine decreased pulmonary responsiveness and increased immune cytokine expression of IFN-γ in comparison to saline controls, with no effect on virus titers and expression of IL-4, IL-13, and IL-12p40.66
Clinical Trials
Currently, there have been several clinical studies to measure the effects of N-acetylcysteine on viral respiratory tract infections. A randomized controlled trial demonstrated long-term N-acetylcysteine regimen resulted in significantly lower episodes of influenza and influenza-like illness, disease severity, and duration of confinement to bed due to respiratory diseases. Only 25% of the virus-infected subjects in the N-acetylcysteine arm developed symptoms compared to 79% receiving placebo.67 Benefits were also documented in patients diagnosed with community acquired pneumonia whose plasma levels of malondialdehyde (oxidative stress parameter) and TNF-α (inflammatory mediator) were decreased and total antioxidant capacity was increased significantly among those treated with N-acetylcysteine.68 Moreover, N-acetylcysteine resulted in a lower rate of ventilator-associated-pneumonia compared with placebo. Patients treated with N-acetylcysteine had significantly shorter length of stay in the ICU, reduced time to hospital discharge, and a higher rate of complete recovery.69 A clinical case study revealed that combination therapy of oseltamivir and N-acetylcysteine improved clinical outcomes in a woman with H1N1 influenza pneumonia, with clearance of pulmonary infiltrates, decreased requirement for oxygen supplementation, and lower plasma level of C-reactive protein.70
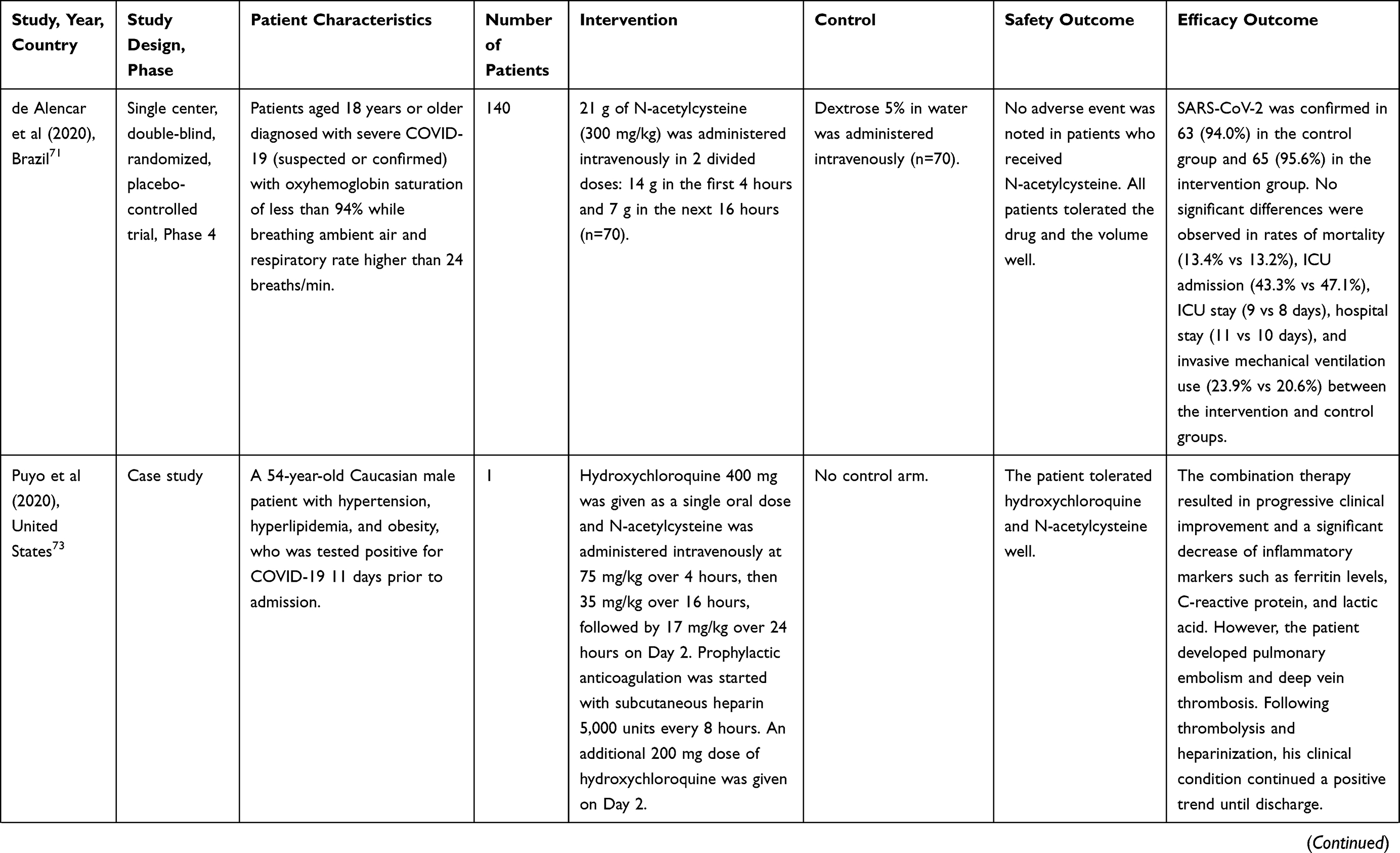
Nonetheless, the majority of human studies utilizing N-acetylcysteine in COVID-19 infection have been small in scale, non-randomized, and lack appropriate comparison groups (Table 1). In a single-center, randomized, double-blind, placebo-controlled trial conducted at the Emergency Department of a hospital in Brazil, 140 patients with severe COVID-19 infection and oxyhemoglobin saturation of less than 94% or respiratory rate higher than 24 breaths/minute were randomized to receive N-acetylcysteine 21,000 mg (approximately 300 mg/kg) for 20 hours or dextrose 5% (placebo). Of the intention-to-treat population comprising 135 patients, 16 patients (24%) receiving placebo required endotracheal intubation and invasive mechanical ventilation compared to 14 patients (21%) in the N-acetylcysteine group (P=0.675). No differences were noted in the duration of mechanical ventilation, death rate, rate of ICU admission, duration of stay in the ICU, and hospital stay.71
 |
 |
 |
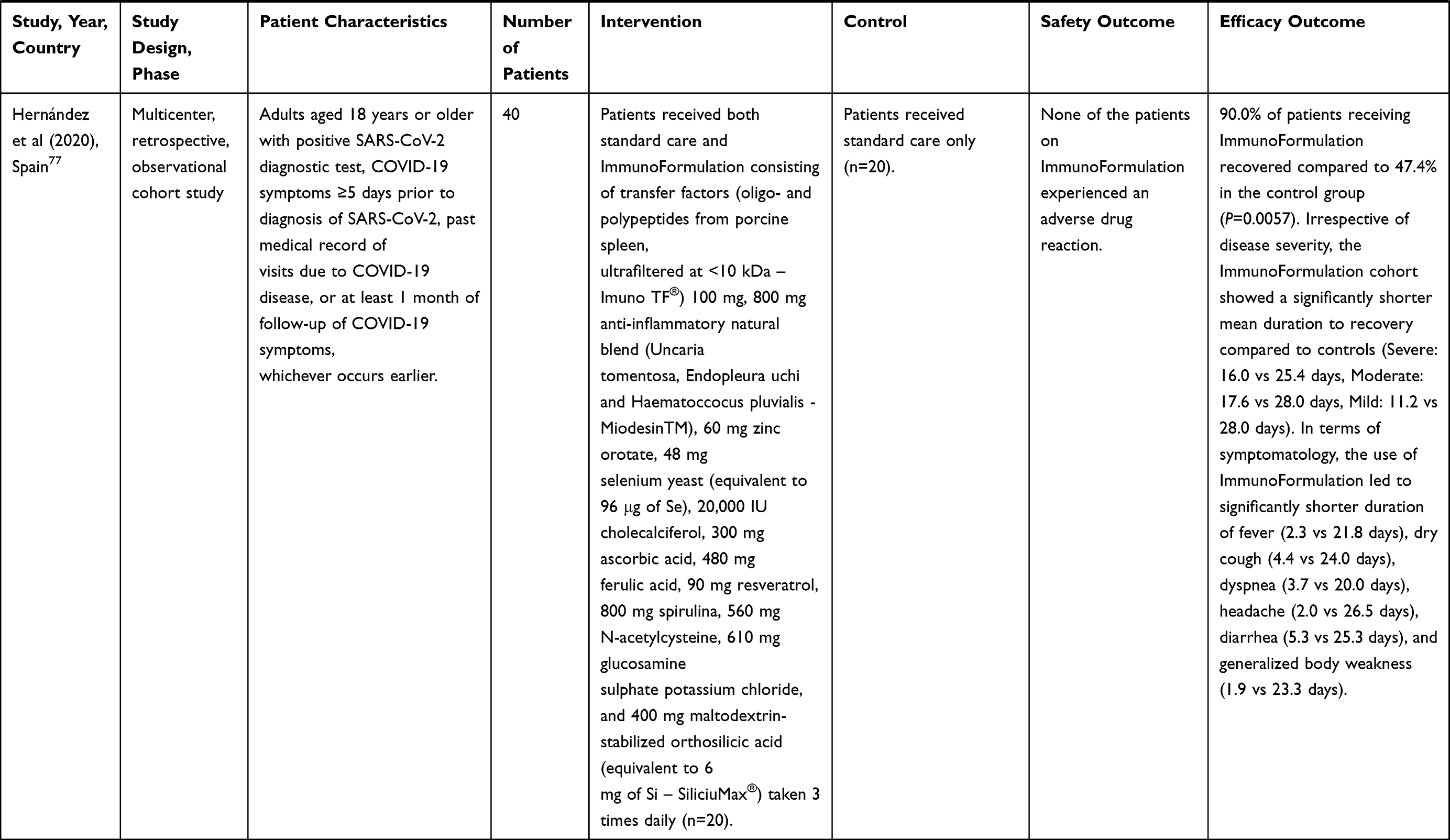 |
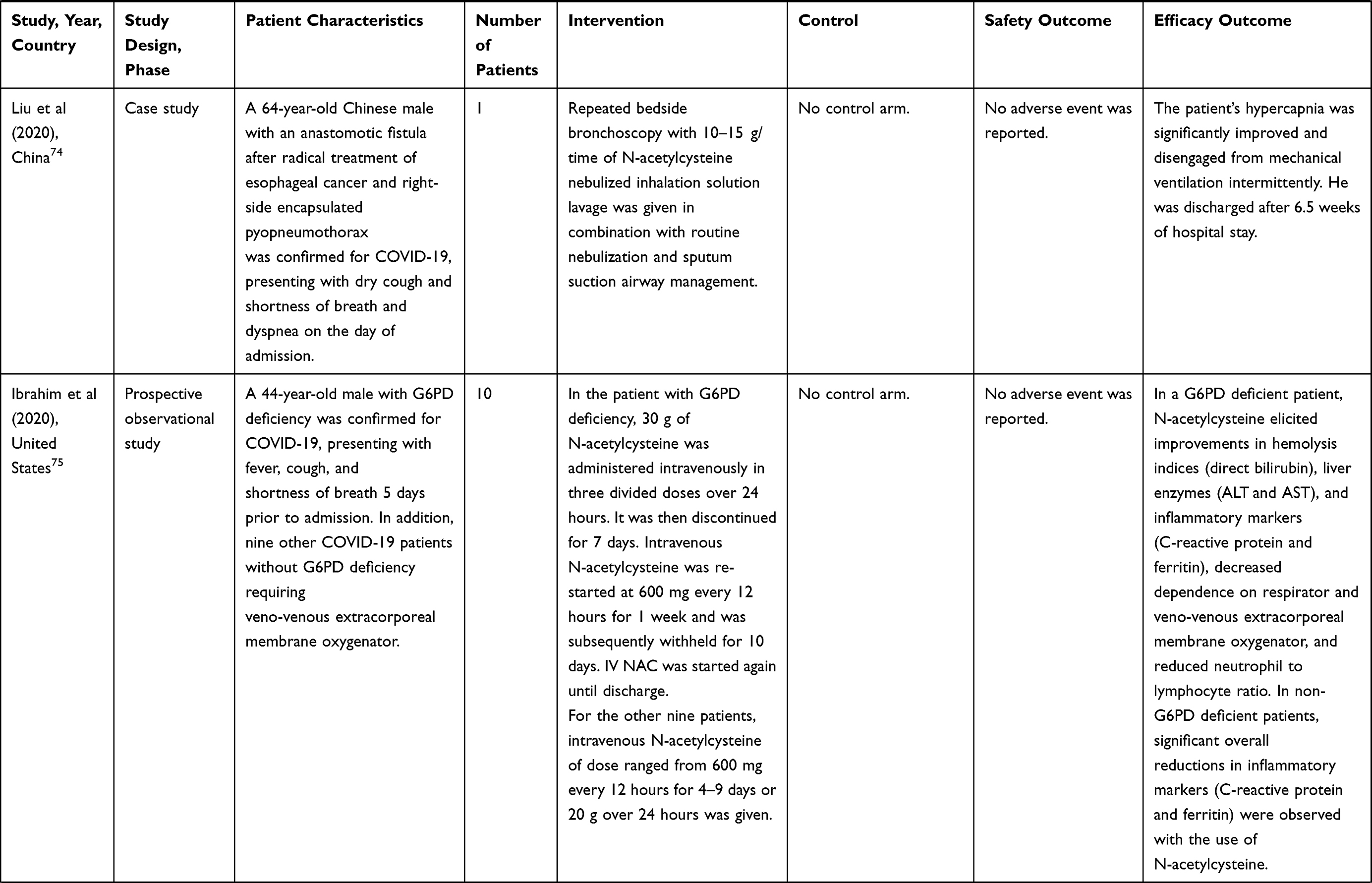
Table 1 Results from Human Trials of N-Acetylcysteine in COVID-19 Infection |
A single-center, randomized, double-blind, placebo-controlled trial evaluating a combined metabolic cofactors supplementation encompassing L-serine, N-acetylcysteine, nicotinamide riboside, and L-carnitine tartrate in 309 adult patients with laboratory-confirmed COVID-19 infection reported that mean recovery time in the intervention group was significantly shorter compared to the placebo group (5.7 vs 9.2 days, P<0.0001). In the intervention group, serum alanine aminotransferase (P=0.032), lactate dehydrogenase (P<0.0001), and creatinine levels (P<0.0001) were significantly lower on Day 14 compared to Day 0. Moreover, patients receiving the metabolic cofactors supplementation showed a greater magnitude of decrease in plasma level of downregulated cytokines compared with placebo group on Day 14. The plasma levels of cytokines associated with inflammatory reactions (CSF-1, IL-15RA, IL18, MCP-1, and TNF) were significantly downregulated in the intervention group compared to the placebo group on Day 14. Only two patients in the intervention group (2.8%) reported a similar adverse event of mild rash on the upper part of the body.72
A case report of a 54-year-old male patient with pre-existing hypertension, hyperlipidemia, and obesity who was tested positive for COVID-19 11 days prior to admission to Holy Family Hospital in Massachusetts showed that combination therapy of hydroxychloroquine and N-acetylcysteine led to a progressive clinical improvement and a decrease in inflammatory markers for inflammation, for instance, C-reactive protein and lactic acid. The patient was mechanically ventilated for only 3 days, was then transferred from the ICU to the general ward on Day 7, and was finally discharged from the hospital on Day 12 with stable vital signs and normalized laboratory results.73
In another case report, a 64-year-old man presented with acute respiratory distress syndrome due to COVID-19 severe pneumonia and secondary bacterial lung infection was admitted to Taihe Hospital in Shiyan, China and was given repeated bedside bronchoscopy at intervals of 1 day or 2 days with a large dose of 10,000–15,000 mg/time of N-acetylcysteine nebulized inhalation solution lavage combined with routine nebulization and sputum suction airway management. The patient’s refractory hypercapnia was gradually improved and was cured and discharged after 46 days of hospitalization.74
Similar clinical benefit was noted in a case study of a 44-year-old man presented to the emergency department of NYU Langone, New York, with a previously diagnosed G6PD deficiency and a current COVID-19 infection with a redisposed risk of hemolysis. The use of intravenous N-acetylcysteine at 30,000 mg in three divided doses over 24 hours was associated with an immediate improvement in hemolysis indices (direct bilirubin levels), liver enzyme levels (alanine aminotransferase and aspartate aminotransferase), resolution of hemolysis as quantified by a sustained reduction in total and direct bilirubin and an elevated haptoglobin, and a sustained decrease in neutrophil to lymphocyte ratio which was an inflammatory predictor. Furthermore, patient’s oxygenation continued to improve and his veno-venous extracorporeal membrane oxygenator was discontinued after 2 weeks. It was observed that the combination of corticosteroids and intravenous N-acetylcysteine resulted in a marked reduction in inflammatory markers (C-reactive protein and ferritin) and the patient was finally discharged home after 41 days of hospital stay. Following these encouraging outcomes, a cohort of nine COVID-19 patients without G6PD deficiency were prescribed with intravenous N-acetylcysteine and demonstrated a significant overall reduction in inflammatory markers (C-reactive protein and ferritin), whereas a rebound inflammation was noted in six patients following discontinuation of the therapy. The median C-reactive protein level during IV N-acetylcysteine administration was 55 mg/dL which was considerably lower compared to the time before administration (143 mg/dL) or after N-acetylcysteine discontinuation (69 mg/dL). N-acetylcysteine was found to mitigate COVID-19-associated cytokine storm, elicit progressive clinical improvement, and facilitate hospital discharge readiness.75
A retrospective case series study in the inpatient department of Medical College Kolkata, India revealed that two out of 148 patients with pre-existing comorbidities and laboratory-confirmed COVID-19 infection were treated with triple combination therapy of Ivermectin, Atorvastatin, and N-acetylcysteine, and the regimen was found to be a useful adjunct to standard of care (oxygenation, restricted fluid therapy, anticoagulation, and corticosteroid). It had no apparent adverse effects and could potentially reduce mortality in patients with COVID-19.76
A multicenter, retrospective, cohort study in Clinic Bascoy and Clínica Arvila Magna, Spain assessed the effectiveness of an ImmunoFormulation containing N-acetylcysteine versus standard care in 40 patients with laboratory-confirmed COVID-19 infection. Ninety percent of patients in the ImmunoFormulation cohort recovered from the infection as compared to 47% in the control cohort (P=0.006). The mean recovery time for the ImmunoFormulation cohort was significantly shorter in comparison to the control cohort for patients with mild symptoms (11.22 vs 28.00 days), moderate symptoms (17.57 vs 28.00 days), and severe symptoms (16.00 vs 25.42 days). The duration of symptoms was also significantly shorter in the ImmunoFormulation cohort compared to the control group, notably for fever (2.25 vs 21.78 days), dry cough (4.38 vs 24.00 days), dyspnea (3.67 vs 20.00 days), headache (2.00 vs 26.50 days), diarrhea (5.25 vs 25.25 days), and weakness (1.92 vs 23.30 days) (all P<0.05). The use of ImmunoFormulation was associated with resolution of all clinical symptoms within 2–5 days, except for the loss of taste or smell (19.73 days) that was recognized as a long-term complication of COVID-19. No patient experienced any adverse drug reactions. The ImmunoFormulation containing 560 mg N-acetylcysteine daily was an effective adjuvant therapy on evolution of symptomatology in COVID-19 patients.77
Notwithstanding the inconsistencies on the degree of efficacy of N-acetylcysteine as an adjunct in COVID-19 infection, all reported studies support the apparent safety profile of N-acetylcysteine irrespective of the dose and route of administration. There are many more clinical studies that are currently underway to generate the necessary evidence of its efficacy to inform future patient care and clinical action (Table 2).78–83
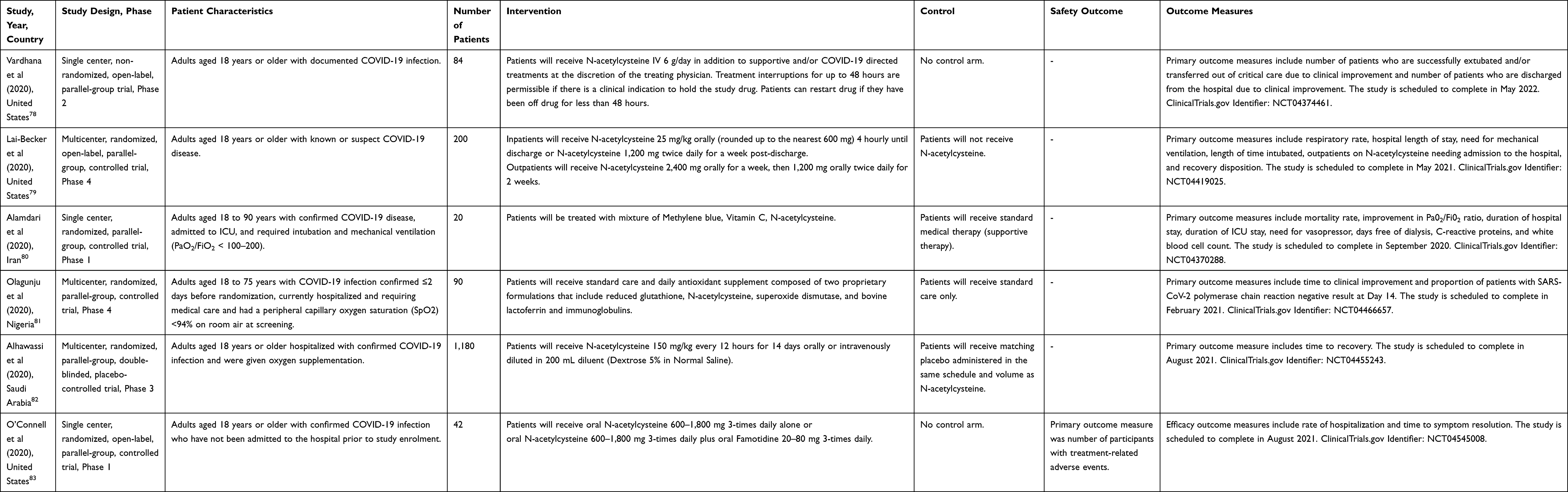 |
Table 2 Protocol Information of Currently Ongoing Human Trials of N-Acetylcysteine in COVID-19 Infection |
Discussion
There has been no in vitro or in vivo research that specifically examines the effect of N-acetylcysteine on COVID-19 infection. The potential use of N-acetylcysteine in COVID-19 is largely inferred from previous research on other viruses such as influenza and respiratory syncytial virus. Promising results have been noted in the ability of N-acetylcysteine to synthesize glutathione, improve T lymphocyte proliferative response, and modulate the inflammatory pathway. Glutathione precursors such as N-acetylcysteine are suggested as a potential therapeutic approach for blocking NF-κB activation and addressing cytokine storm syndrome and respiratory distress in patients suffering from COVID-19 pneumonia.84 In plasma, N-acetylcysteine reacts with cystine, reducing it to cysteine and yielding diacetylcystine and N-acetylcysteine-cysteine via redox exchange reactions, and subsequently enters human erythrocytes and sustains glutathione synthesis.85 Additional research has illuminated the action of N-acetylcysteine on T-cell proliferation and IL-2 secretion which implies intracellular thiols regulate selective signaling pathways for a novel target of immunoregulation.86 Furthermore, in vitro treatment of murine T-cells with N-acetylcysteine induces reactive oxygen species scavenging and initiates NFAT expression and nuclear translocation. Addition of N-acetylcysteine to Cd4cre-Gclc T-cells increases Myc expression and CD98 production. Indeed, intrinsic Gclc expression in T-cells is necessary for antigen-specific immunity to virus infections. The antioxidative glutathione pathway is found to be central to metabolic integration and reprogramming in inflammatory responses mediated by T-cells.87 In this regard, high-dose N-acetylcysteine has been exemplified to replenish depleted pulmonary glutathione concentrations and yield concomitant favorable effects on lung function.88 The therapy also increases glutathione levels in peripheral blood T lymphocytes and disrupts the mammalian target of rapamycin activation in chronic inflammatory disease.89 Given that severe COVID-19 possesses shareable dominant risk factors with idiopathic pulmonary fibrosis, the effectiveness of N-acetylcysteine for slowing the rate of deterioration of vital capacity and single-breath lung diffusion capacity for carbon monoxide presents a rational option for treating patients infected with COVID-19.88,90 Therefore, it is conjectured that high dose N-acetylcysteine can enhance innate and adaptive immunity by elevating stores of glutathione levels in T lymphocytes, along with modulating immune-system responses to alleviate the degree of severity of COVID-19 infection and thus improve patient outcomes. Future in vitro studies to test candidate therapeutic compound in COVID-19 should deploy standardized assays, for example, Vero cells that can competently replicate and isolate the virus readily, whereas in vivo studies should utilize transgenic mice and Syrian hamsters, or cats and ferrets if opting for larger experimental animals.91
Of note, one of the important regulators of inflammation in COVID-19 is IL-6.92 Elevations in serum IL-6 concentrations have been identified under critical conditions such as sepsis, acute respiratory distress syndrome, and COVID-19.93,94 IL-6 is crucial for innate and adaptive immunity, efficient pathogen clearance, and physiological functions such as regulation of acute-phase response, hematopoiesis, lipid homeostasis, metabolism, and neural development.95 In essence, anti-inflammatory, pro-resolution, and antimicrobial activities of IL-6 are facilitated by classical signalling, whilst pro-inflammatory activities of IL-6 are facilitated by trans-signalling.96 Targeting the pathological effects of IL-6-mediated inflammation should avoid the unintentional concurrent abolition of its anti-inflammatory and pro-resolution functions.93 Blockade of IL-6 and the consequent reduction of downstream effects on inflammation and the innate immune response may have beneficial effects on clinical outcomes in patients with COVID-19, including those with acute hypoxemic respiratory failure.97 Contemporary in vitro and in vivo evidence indicating a potential effect of N-acetylcysteine in IL-6 inhibition may stimulate further research to understand how the drug affects disease outcomes and maximize its benefits with concomitant pharmacotherapy to selectively inhibit the trans-signalling pathway through which IL-6 signals.
Oral and intravenous N-acetylcysteine regimens are associated with adverse events such as diarrhea, nausea, vomiting, and mild-to-moderate hypersensitivity reactions such as urticaria, rash, angioedema, and wheezing.98,99 Such adverse events are dose-related and antihistamines are effective for treating and preventing recurrence of the anaphylactic reactions. Urticaria should be treated with antihistamine and N-acetylcysteine can be continued after symptom resolution. Angioedema and respiratory symptoms require the administration of antihistamine, corticosteroid, or symptomatic management.100 In such cases, N-acetylcysteine should be discontinued, and if necessary, it can be re-started 1 hour after the administration of antihistamine in the absence of signs and symptoms of anaphylaxis.101,102 Patients with a history of atopy (asthma and allergy) are recommended to receive prophylactic antihistamine 15 minutes prior to N-acetylcysteine administration to eliminate the excess risk of adverse reactions.102,103
At present, there are six registered clinical trials evaluating the potential therapeutic value of N-acetylcysteine against COVID-19.78–83 The findings will build on in vitro and early in vivo evidence for the use of N-acetylcysteine as an adjuvant therapy for COVID-19. Similar to clinical trials on other infectious respiratory illnesses, study endpoints should appropriately encompass changes in SARS-CoV-2 associated blood cytokine levels, change in peripheral-blood lymphocyte count and activation, improvement in clinical status, hospital length of stay, time to recovery, safety outcomes, and overall mortality. While SARS-CoV-2 variants appear to occur sporadically in different geographical locations across the globe, they do not culminate in more severe diseases than the ancestral strain, albeit altered virus virulence, pathogenesis, and transmissibility have been documented.104,105 Future trials in patients infected with mutated SARS-CoV-2 strains may help to confirm or refute the possibility of a treatment benefit with N-acetylcysteine.
It is noteworthy that molecular attraction forces between chitin found on the exoskeleton of mites on human skin (Demodecidae or Pyemotidae species) and lipids present on the viral envelope cause interactions between arthropod and coronavirus which play a major role in the transmission of SARS-CoV-2.106 Natural small molecules, namely cyclodextrins and phytosterols, may hinder viral lipid‐dependent attachment to host cells and thus reduce infectivity of the virus.107 Nowadays, these natural compounds exist in various forms of dietary supplements.107 Their potential to exhibit antiviral effects invigorates additional research to create novel biomedical applications for the treatment and prevention of COVID-19. This also implies the possibility that N-acetylcysteine has a role in inhibiting arthropod‐coronavirus interaction and producing antimicrobial effect on endosymbionts of Demodex folliculorum.108 Ivermectin is a broad-spectrum anti-parasitic drug which elicits anti-inflammatory and acaricidal actions against Demodex mites,109,110 in addition to antiviral activity against a range of viruses.111 Possible mechanisms include downregulation of the expression of pro-inflammatory genes, including those of IL-8, TNF-α, and cathelicidin LL-37 and downregulation of the ACE-2 receptor and viral entry into the cells of the respiratory epithelium and olfactory bulb.112 As a free radical scavenger, N-acetylcysteine can prevent ivermectin-induced cell death due to reactive oxygen species generation,113 suggestive of potential benefits with their combined use. However, current clinical findings do not support the use of ivermectin for the treatment of COVID-19 as it does not significantly improve the time to resolution of symptoms.114
Patients with COVID-19 pneumonia can present with blood coagulation abnormalities, commonly manifested by elevated levels of fibrinogen and D-dimer in tandem with mild thrombocytopenia.115,116 Rise in D-dimer levels has been linked to poorer prognosis and higher mortality rate.117,118 D-dimer levels, lung inflammation, and pulmonary hemorrhage are influenced by neutrophil elastase activity.119,120 As such, suppression of elastase and neutrophil activation may be helpful in hemorrhagic or thrombotic complications associated with COVID-19.121 N-acetylcysteine has been found to inhibit release of elastase and modulate neutrophil activity when used at high concentrations.122 In neutrophilic airway inflammation in cystic fibrosis, high-dose N-acetylcysteine decreases the neutrophil burden in airways and the number of airway neutrophils actively releasing elastase-rich granules.123 Treatment with N-acetylcysteine can also attenuate lung emphysema induced by elastase as depicted in amelioration of airspace enlargements, partial recovery of expiratory flows, and normalization of lung collagen content.124 This evidence sheds light on the possible role of N-acetylcysteine on mediating inflammation-mediated lung injury and abnormalities of blood coagulation in severe COVID-19.
Novel formulations have recently been discovered to overcome the low bioavailability and short plasma half-life of N-acetylcysteine.125 Loading of N-acetylcysteine into lipid-based and polymer-based nanoparticles can improve muco-penetrating properties in pulmonary and oral delivery and hence the drug efficacy.126,127 A detailed in vivo and in vitro investigation is warranted to derive their applicability and efficacy in clinical settings.
As the most commonly approved nanomedicines by the US FDA,128 lipid-based nanoparticles afford many advantages such as relatively simple formulation, self-assembled structure, biocompatibility, drug load capacity, and adjustable physicochemical properties to suit biological characteristics.129 However, the lipid nanoparticle system is limited by low physicochemical stability and low-to-moderate encapsulation efficiency for N-acetylcysteine.130 A liposomal formulation of co-encapsulated azithromycin and N-acetylcysteine has been demonstrated to have a synergistic effect against bacterial strains.131 The high cellular membrane permeability and targeting accuracy associated with such a delivery system illuminates a potential implication for the co-formulation strategy of N-acetylcysteine with antiviral drug.
Recent evidence shows that polymer-based nanoparticles like N-acetylcysteine-loaded poly(lactic-co-glycolic acid) have desirable efficacy in preventing acute lung injury by mitigating the effects of reactive oxygen species and inflammation. The formulation delivers N-acetylcysteine directly to the lungs with increased pulmonary deposition and higher pulmonary concentrations with a lower dose of N-acetylcysteine.132 Polymer-based nanoparticle enhances pharmacokinetic and pharmacodynamics properties of encapsulated drug via prolonged drug release and particle retention.133,134 While poly(lactic-co-glycolic acid) is one of the most commonly used biodegradable polymers, its poor drug loading requires higher polymer load to facilitate dose delivery.135 Polymer-based nanoparticles are biodegradable, water soluble, biocompatible, biomimetic, and stable during storage. The surfaces can be easily modified for additional targeting. Disadvantages comprise higher risk of particle aggregation and toxicity.127
To date, N-acetylcysteine has been used intranasally for the treatment of non-allergic chronic rhinitis with goblet cell metaplasia in which significant reduction in neutrophils, lymphocytes, goblet cells, bacterial count, turbinate hypertrophy, nasal symptoms, and rhinorrhea have been observed. The benefits are attributed to N-acetylcysteine’s mucolytic activity, the ability to restore surface ciliary activity at the nasal epithelium, and modulation of inflammatory response which is central to the immune defence mechanism of nasal mucosa.136 Likewise, acute recurrent rhinosinusitis treated with nasal douche consisting of flunisolide (corticosteroid) and N-acetylcysteine has been associated with a less stuffy nose, reduction in severity of symptoms and rhinosinus signs as assessed by endoscopy, improved cytological grading of neutrophil and eosinophil cells, less exacerbations, and increased mucociliary motility.137 In addition, topical intranasal drug combining tuaminoheptane sulphate (vasoconstrictor) and N-acetylcysteine exhibits a rapid decongestant effect with a significant decrease of resistance and increase of inspiratory flow.138 Topical delivery of N-acetylcysteine in the nostrils has also been investigated in ragweed sensitive patients and discovered that late phase allergic response mediated nasal symptoms can be reduced.139 In vivo studies have demonstrated nasal application of N-acetylcysteine in rats can reduce goblet cell loss and inflammation as well as promote wound healing of nasal mucosa,140 and combination of N-acetylcysteine and non-ionic surfactant (polyoxyethylene (C25) lauryl ether) can improve nasal bioavailability as a result of mucolytic activity of N-acetylcysteine in decreasing mucus viscosity.141 Accruing evidence suggests nasal route drug administration may be a potentially attractive strategy for N-acetylcysteine on top of the highly vascularized nasal cavity and large surface area for drug absorption.142
Conclusions
The COVID-19 pandemic has highlighted the critical need for new drugs to complement existing therapies. In view of the widespread recognition of the safety and efficacy of N-acetylcysteine in numerous diseases over several decades, the aroused scientific interest has prompted the evaluation of its efficacy in COVID-19 clinical trials. As such, it opens a window for drug discovery with additional advantages, comprising the known pharmacological and human safety profiles. Advances in nanoparticle design are foreseen to have an impact on optimizing drug delivery and targeted activity of N-acetylcysteine in coronavirus disease. Currently, there is some evidence supporting the use of N-acetylcysteine as an adjunctive therapy for COVID-19. Further studies are warranted to design a formulation with increased bioavailability or target-oriented delivery, ascertain optimal dosage and route of administration, and conduct well-controlled, adequately powered, randomized clinical trials to determine the safety and efficacy of N-acetylcysteine in patients with COVID-19.
Funding
The authors received no specific funding for this work.
Disclosure
The authors declared that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Blumenthal D, Fowler EJ, Abrams M, Collins SR. Covid-19 — implications for the Health Care System. N Eng J Med. 2020;383(15):1483–1488. doi:10.1056/NEJMsb2021088
2. Gandhi RT, Lynch JB, Del Rio C. Mild or Moderate Covid-19. N Eng J Med. 2020;383(18):1757–1766. doi:10.1056/NEJMcp2009249
3. Li Q, Guan X, Wu P, et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N Eng J Med. 2020;382(13):1199–1207. doi:10.1056/NEJMoa2001316
4. Hassan AO, Case JB, Winkler ES, et al. A SARS-CoV-2 Infection Model in Mice Demonstrates Protection by Neutralizing Antibodies. Cell. 2020;182(3):744–753.e4. doi:10.1016/j.cell.2020.06.011
5. Sariol A, Perlman S. Lessons for COVID-19 Immunity from Other Coronavirus Infections. Immunity. 2020;53(2):248–263. doi:10.1016/j.immuni.2020.07.005
6. Gates B. Responding to Covid-19 — a Once-in-a-Century Pandemic? N Eng J Med. 2020;382(18):1677–1679. doi:10.1056/NEJMp2003762
7. Siemieniuk RA, Bartoszko JJ, Ge L, et al. Drug treatments for covid-19: living systematic review and network meta-analysis. BMJ. 2020;370:m2980. doi:10.1136/bmj.m2980
8. Rabi FA, Al Zoubi MS, Kasasbeh GA, Salameh DM. SARS-CoV-2 and Coronavirus Disease 2019: what We Know So Far. Pathogens. 2020;9(3):231. doi:10.3390/pathogens9030231
9. Holman N, Knighton P, Kar P, et al. Risk factors for COVID-19-related mortality in people with type 1 and type 2 diabetes in England: a population-based cohort study. Lancet Diabetes Endocrinol. 2020;8(10):823–833. doi:10.1016/S2213-8587(20)30271-0
10. Słomka A, Kowalewski M, Żekanowska E. Coronavirus Disease 2019 (COVID-19): a Short Review on Hematological Manifestations. Pathogens. 2020;9(6):493. doi:10.3390/pathogens9060493
11. Gencer S, Lacy M, Atzler D, van der Vorst EPC, Döring Y, Weber C. Immunoinflammatory, Thrombohaemostatic, and Cardiovascular Mechanisms in COVID-19. Thromb Haemost. 2020;120(12):1629–1641. doi:10.1055/s-0040-1718735
12. Kalra RS, Tomar D, Meena AS, Kandimalla R. SARS-CoV-2, ACE2, and Hydroxychloroquine: cardiovascular Complications, Therapeutics, and Clinical Readouts in the Current Settings. Pathogens. 2020;9(7):546. doi:10.3390/pathogens9070546
13. Hewitt JA, Lutz C, Florence WC, et al. ACTIVating Resources for the COVID-19 Pandemic: in Vivo Models for Vaccines and Therapeutics. Cell Host Microbe. 2020;28(5):646–659. doi:10.1016/j.chom.2020.09.016
14. Grobler JA, Anderson AS, Fernandes P, et al. Accelerated Preclinical Paths to Support Rapid Development of COVID-19 Therapeutics. Cell Host Microbe. 2020;28(5):638–645. doi:10.1016/j.chom.2020.09.017
15. Pan H, Peto R, Henao-Restrepo AM, et al. Repurposed Antiviral Drugs for Covid-19 - Interim WHO Solidarity Trial Results. N Engl J Med. 2020;384(6):497–511. doi:10.1056/NEJMoa2023184
16. Teoh SL, Lim YH, Lai NM, Lee SWH. Directly Acting Antivirals for COVID-19: where Do We Stand? Front Microbiol. 2020;11:1857. doi:10.3389/fmicb.2020.01857
17. Kalil AC, Patterson TF, Mehta AK, et al. Baricitinib plus Remdesivir for Hospitalized Adults with Covid-19. N Eng J Med. 2021;384(9):795-807. doi:10.1056/NEJMoa2031994
18. The RECOVERY Collaborative Group. Dexamethasone in Hospitalized Patients with Covid-19. N Eng J Med. 2021;384(8):693-704. doi:10.1056/NEJMoa2021436
19. Chen P, Nirula A, Heller B, et al. SARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19. N Eng J Med. 2021;384(3):229-237. doi:10.1056/NEJMoa2029849
20. Polack FP, Thomas SJ, Kitchin N, et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Eng J Med. 2020;383(27):2603–2615. doi:10.1056/NEJMoa2034577
21. Baden LR, El Sahly HM, Essink B, et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Eng J Med. 2021;384(5):403–416. doi:10.1056/NEJMoa2035389
22. Heath PT, Galiza EP, Baxter DN, et al. Safety and Efficacy of NVX-CoV2373 Covid-19 Vaccine. N Eng J Med. 2021. doi:10.1056/NEJMoa2107659
23. Goepfert PA, Fu B, Chabanon A-L, et al. Safety and immunogenicity of SARS-CoV-2 recombinant protein vaccine formulations in healthy adults: interim results of a randomised, placebo-controlled, phase 1-2, dose-ranging study. Lancet Infect Dis. 2021;S1473-3099(21)00147-X. doi:10.1016/S1473-3099(21)00147-X
24. Sadoff S, Gray G, Vandebosch A, et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N Eng J Med. 2021;384(23):2187-2201. doi:10.1056/NEJMoa2101544
25. Voysey M, Clemens SAC, Madhi SA, et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet. 2021;397(10269):99–111. doi:10.1016/S0140-6736(20)32661-1
26. Landini G, Di Maggio T, Sergio F, Docquier J-D, Rossolini GM, Pallecchi L. Effect of High N-Acetylcysteine Concentrations on Antibiotic Activity against a Large Collection of Respiratory Pathogens. Antimicrob Agents Chemother. 2016;60(12):7513–7517. doi:10.1128/aac.01334-16
27. Samuni Y, Goldstein S, Dean OM, Berk M. The chemistry and biological activities of N-acetylcysteine. Biochim Biophys Acta. 2013;1830(8):4117–4129. doi:10.1016/j.bbagen.2013.04.016
28. Rodríguez-Beltrán J, Cabot G, Valencia EY, et al. N-Acetylcysteine Selectively Antagonizes the Activity of Imipenem in Pseudomonas aeruginosa by an OprD-Mediated Mechanism. Antimicrob Agents Chemother. 2015;59(6):3246–3251. doi:10.1128/aac.00017-15
29. Goswami M, Jawali N. Acetylcysteine-Mediated Modulation of Bacterial Antibiotic Susceptibility. Antimicrob Agents Chemother. 2010;54(8):3529–3530. doi:10.1128/aac.00710-10
30. Domenech M, García E. N-Acetyl-L-Cysteine and Cysteamine as New Strategies against Mixed Biofilms of Nonencapsulated Streptococcus pneumoniae and Nontypeable Haemophilus influenzae. Antimicrob Agents Chemother. 2017;61(2):e01992–16. doi:10.1128/aac.01992-16
31. Blasi F, Page C, Rossolini GM, et al. The effect of N-acetylcysteine on biofilms: Implications for the treatment of respiratory tract infections. Respir Med. 2016;117:190‒197. doi:10.1016/j.rmed.2016.06.015
32. Jorge-Aarón R-M, Rosa-Ester M-P. N-acetylcysteine as a potential treatment for COVID-19. Future Microbiol. 2020;15:959–962. doi:10.2217/fmb-2020-0074
33. Poe FL, Corn J. N-Acetylcysteine: a potential therapeutic agent for SARS-CoV-2. Med Hypotheses. 2020;143:109862. doi:10.1016/j.mehy.2020.109862
34. Der Kinderen DJ, Koten JW. Rapid response: N-acetylcysteine as a possible adjuvant in the treatment of SARS-CoV-2. BMJ. 2020;371:m3862. Available from: https://www.bmj.com/content/371/bmj.m3862/rr-8. Accessed May 21, 2021.
35. Aldini G, Altomare A, Baron G, et al. N-Acetylcysteine as an antioxidant and disulphide breaking agent: the reasons why. Free Radic Res. 2018;52(7):751–762. doi:10.1080/10715762.2018.1468564
36. Schoeman D, Fielding BC. Coronavirus envelope protein: current knowledge. Virol J. 2019;16(1):69. doi:10.1186/s12985-019-1182-0
37. Suhail S, Zajac J, Fossum C, et al. Role of Oxidative Stress on SARS-CoV (SARS) and SARS-CoV-2 (COVID-19) Infection: a Review. Protein J. 2020;39(6):644–656. doi:10.1007/s10930-020-09935-8
38. Fenouillet E, Barbouche R, Jones IM. Cell entry by enveloped viruses: redox considerations for HIV and SARS-coronavirus. Antioxid Redox Signal. 2007;9(8):1009–1034. doi:10.1089/ars.2007.1639
39. Ryser HJ, Levy EM, Mandel R, DiSciullo GJ. Inhibition of human immunodeficiency virus infection by agents that interfere with thiol-disulfide interchange upon virus-receptor interaction. Proc Natl Acad Sci U S A. 1994;91(10):4559–4563. doi:10.1073/pnas.91.10.4559
40. Hati S, Bhattacharyya S. Impact of Thiol–Disulfide Balance on the Binding of Covid-19 Spike Protein with Angiotensin-Converting Enzyme 2 Receptor. ACS Omega. 2020;5(26):16292–16298. doi:10.1021/acsomega.0c02125
41. Ullian ME, Gelasco AK, Fitzgibbon WR, Beck CN. N-Acetylcysteine Decreases Angiotensin II Receptor Binding in Vascular Smooth Muscle Cells. J Am Soc Nephrol. 2005;16(8):2346–2353. doi:10.1681/asn.2004060458
42. Altomare A, Baron G, Brioschi M, et al. N-Acetyl-Cysteine Regenerates Albumin Cys34 by a Thiol-Disulfide Breaking Mechanism: an Explanation of Its Extracellular Antioxidant Activity. Antioxidants. 2020;9(5):367. doi:10.3390/antiox9050367
43. Peiró C, Moncada S. Substituting Angiotensin-(1-7) to Prevent Lung Damage in SARS-CoV-2 Infection? Circulation. 2020;141(21):1665–1666. doi:10.1161/CIRCULATIONAHA.120.047297
44. Fishbane S. N-Acetylcysteine in the Prevention of Contrast-Induced Nephropathy. Clin J Am Soc Nephrol. 2008;3(1):281–287. doi:10.2215/cjn.02590607
45. Yehualashet AS, Belachew TF. ACEIs and ARBs and Their Correlation with COVID-19: a Review. Infect Drug Resist. 2020;13:3217–3224. doi:10.2147/IDR.S264882
46. Mahmoud Abd El Hafiz A, Mohammed El Wakeel L, Mohammed El Hady H, Mourad AER. High dose N-acetyl cysteine improves inflammatory response and outcome in patients with COPD exacerbations. Egyptian J Chest Dis Tuberculosis. 2013;62(1):51‒57. doi:10.1016/j.ejcdt.2013.02.012
47. McCarty MF, DiNicolantonio JJ. Nutraceuticals have potential for boosting the type 1 interferon response to RNA viruses including influenza and coronavirus. Prog Cardiovasc Dis. 2020;63(3):383–385. doi:10.1016/j.pcad.2020.02.007
48. Lee JS, Shin E-C. The type I interferon response in COVID-19: implications for treatment. Nat Rev Immunol. 2020;20(10):585–586. doi:10.1038/s41577-020-00429-3
49. De Francesco EM, Vella V, Belfiore A. COVID-19 and Diabetes: the Importance of Controlling RAGE. Opinion. Front Endocrinol (Lausanne). 2020;11:526. doi:10.3389/fendo.2020.00526
50. Wetzels S, Wouters K, Schalkwijk CG, Vanmierlo T, Hendriks JJA. Methylglyoxal-Derived Advanced Glycation Endproducts in Multiple Sclerosis. Int J Mol Sci. 2017;18(2):421. doi:10.3390/ijms18020421
51. Bourgonje AR, Offringa AK, van Eijk LE, et al. N-Acetylcysteine and Hydrogen Sulfide in Coronavirus Disease 2019. Antioxid Redox Signal. 2021. doi:10.1089/ars.2020.8247
52. Oka S, Kamata H, Kamata K, Yagisawa H, Hirata H. N-Acetylcysteine suppresses TNF-induced NF-κB activation through inhibition of IκB kinases. FEBS Lett. 2000;472(2):196–202. doi:10.1016/S0014-5793(00)01464-2
53. Kircheis R, Haasbach E, Lueftenegger D, Heyken WT, Ocker M, Planz O. NF-κB Pathway as a Potential Target for Treatment of Critical Stage COVID-19 Patients. Hypothesis and Theory. Front Immunol. 2020;11:598444. doi:10.3389/fimmu.2020.598444
54. Geiler J, Michaelis M, Naczk P, et al. N-acetyl-L-cysteine (NAC) inhibits virus replication and expression of pro-inflammatory molecules in A549 cells infected with highly pathogenic H5N1 influenza A virus. Biochem Pharmacol. 2010;79(3):413–420. doi:10.1016/j.bcp.2009.08.025
55. Boon AC, Vos AP, Graus YM, Rimmelzwaan GF, Osterhaus AD. In vitro effect of bioactive compounds on influenza virus specific B- and T-cell responses. Scand J Immunol. 2002;55(1):24–32. doi:10.1046/j.1365-3083.2002.01014.x
56. Mata M, Morcillo E, Gimeno C, Cortijo J. N-acetyl-L-cysteine (NAC) inhibit mucin synthesis and pro-inflammatory mediators in alveolar type II epithelial cells infected with influenza virus A and B and with respiratory syncytial virus (RSV). Biochem Pharmacol. 2011;82(5):548–555. doi:10.1016/j.bcp.2011.05.014
57. Wu H, Song W, Gao X, et al. Proteomics study of N-acetylcysteine response in H1N1-infected cells by using mass spectrometry. Rapid Commun Mass Spectrom. 2014;28(7):741–749. doi:10.1002/rcm.6840
58. Garigliany MMO, Desmecht DJ. N-acetylcysteine lacks universal inhibitory activity against influenza A viruses. J Negat Results Biomed. 2011;10:5. doi:10.1186/1477-5751-10-5
59. Mata M, Sarrion I, Armengot M, et al. Respiratory syncytial virus inhibits ciliagenesis in differentiated normal human bronchial epithelial cells: effectiveness of N-acetylcysteine. PLoS One. 2012;7(10):e48037–e48037. doi:10.1371/journal.pone.0048037
60. Carpenter LR, Moy JN, Roebuck KA. Respiratory syncytial virus and TNFalpha induction of chemokine gene expression involves differential activation of Rel A and NF-kappaB1. BMC Infect Dis. 2002;2(1):5. doi:10.1186/1471-2334-2-5
61. Ghezzi P, Ungheri D. Synergistic combination of N-acetylcysteine and ribavirin to protect from lethal influenza viral infection in a mouse model. Int J Immunopathol Pharmacol. 2004;17(1):99–102. doi:10.1177/039463200401700114
62. Ungheri D, Pisani C, Sanson G, et al. Protective effect of n-acetylcysteine in a model of influenza infection in mice. Int J Immunopathol Pharmacol. 2000;13(3):123–128.
63. Garozzo A, Tempera G, Ungheri D, Timpanaro R, Castro A. N-acetylcysteine synergizes with oseltamivir in protecting mice from lethal influenza infection. Int J Immunopathol Pharmacol. 2007;20(2):349–354. doi:10.1177/039463200702000215
64. Garigliany MM, Habyarimana A, Lambrecht B, et al. Influenza A strain-dependent pathogenesis in fatal H1N1 and H5N1 subtype infections of mice. Emerg Infect Dis. 2010;16(4):595–603. doi:10.3201/eid1604.091061
65. Zhang RH, Li CH, Wang CL, et al. N-acetyl-l-cystine (NAC) protects against H9N2 swine influenza virus-induced acute lung injury. Int Immunopharmacol. 2014;22(1):1–8. doi:10.1016/j.intimp.2014.06.013
66. Gowdy KM, Krantz QT, King C, et al. Role of oxidative stress on diesel-enhanced influenza infection in mice. Part Fibre Toxicol. 2010;7:34. doi:10.1186/1743-8977-7-34
67. De Flora S, Grassi C, Carati L. Attenuation of influenza-like symptomatology and improvement of cell-mediated immunity with long-term N-acetylcysteine treatment. Eur Respir J. 1997;10(7):1535–1541. doi:10.1183/09031936.97.10071535
68. Zhang Q, Ju Y, Ma Y, Wang T. N-acetylcysteine improves oxidative stress and inflammatory response in patients with community acquired pneumonia: a randomized controlled trial. Medicine. 2018;97(45):e13087–e13087. doi:10.1097/MD.0000000000013087
69. Sharafkhah M, Abdolrazaghnejad A, Zarinfar N, Mohammadbeigi A, Massoudifar A, Abaszadeh S. Safety and efficacy of N-acetyl-cysteine for prophylaxis of ventilator-associated pneumonia: a randomized, double blind, placebo-controlled clinical trial. Med Gas Res. 2018;8(1):19–23. doi:10.4103/2045-9912.229599
70. Lai KY, Ng WY, Osburga Chan PK, Wong KF, Cheng F. High-dose N-acetylcysteine therapy for novel H1N1 influenza pneumonia. Ann Intern Med. 2010;152(10):687–688. doi:10.7326/0003-4819-152-10-201005180-00017
71. de Alencar JCG, Moreira CDL, Müller AD, et al. Double-blind, Randomized, Placebo-controlled Trial With N-acetylcysteine for Treatment of Severe Acute Respiratory Syndrome Caused by Coronavirus Disease 2019 (COVID-19). Clin Infect Dis. 2021;72(11):e736-e741. doi:10.1093/cid/ciaa1443
72. Altay O, Yang H, Aydin M, et al. Combined metabolic cofactor supplementation accelerates recovery in mild-to-moderate COVID-19. medRxiv. 2020. doi:10.1101/2020.10.02.20202614
73. Puyo C, Kreig D, Saddi V, Ansari E, Prince O. Case Report: use of hydroxychloroquine and N-acetylcysteine for treatment of a COVID-19 patient [version 2; peer review: 2 not approved]. F1000Research. 2020;9. doi:10.12688/f1000research.23995.2
74. Liu Y, Wang M, Luo G, et al. Experience of N-acetylcysteine airway management in the successful treatment of one case of critical condition with COVID-19: a case report. Medicine. 2020;99(42):e22577–e22577. doi:10.1097/MD.0000000000022577
75. Ibrahim H, Perl A, Smith D, et al. Therapeutic blockade of inflammation in severe COVID-19 infection with intravenous N-acetylcysteine. Clin Immunol. 2020;219:108544. doi:10.1016/j.clim.2020.108544
76. Bhattacharya R, Ghosh R, Kulshrestha M, Chowdhury S, Mukherjee R, Ray I. Observational Study on Clinical Features, Treatment and Outcome of COVID 19 in a tertiary care Centre in India- a retrospective case series. medRxiv. 2020. doi:10.1101/2020.08.12.20170282
77. Hernández MD, Urrea J, Bascoy L. Evolution of COVID-19 patients treated with ImmunoFormulation, a combination of nutraceuticals to reduce symptomatology and improve prognosis: a multi-centred, retrospective cohort study. medRxiv. 2020. doi:10.1101/2020.12.11.20246561
78. Vardhana S, Wolchok J. Memorial Sloan Kettering Cancer Center. A study of N-acetylcysteine in patients with COVID-19 infection. Available from: https://clinicaltrials.gov/ct2/show/NCT04374461. NLM identifier: NCT04374461. Accessed December 15, 2020.
79. Lai-Becker M. Cambridge Health Alliance. Efficacy of N-Acetylcysteine (NAC) in preventing COVID-19 from progressing to severe disease. Available from: https://clinicaltrials.gov/ct2/show/NCT04419025. NLM identifier: NCT04419025. Accessed December 15, 2020.
80. Alamdari DH, Yarahmadi A. Mashhad University of Medical Sciences. Clinical application of methylene blue for treatment of COVID-19 patients (COVID-19). Available from: https://clinicaltrials.gov/ct2/show/NCT04370288. NLM identifier: NCT04370288. Accessed December 20, 2020.
81. Olagunju A. Obafemi Awolowo University. Antioxidant therapy for COVID-19 study (GSHSOD-COVID). Available from: https://clinicaltrials.gov/ct2/show/NCT04466657. NLM identifier: NCT04466657. Accessed December 20, 2020.
82. Alhawassi T, Alabdulbaqi B. King Saud University. Inflammatory regulation effect of NAC on COVID-19 treatment (INFECT-19). Available from: https://clinicaltrials.gov/ct2/show/NCT04455243. NLM identifier: NCT04455243. Accessed December 20, 2020.
83. O'Connell JJ, Martin J. Prisma Health-Upstate. Trial of famotidine & N-acetyl cysteine for outpatients with COVID-19. Available from: https://clinicaltrials.gov/ct2/show/NCT04545008. NLM identifier: NCT04545008. Accessed December 20, 2020.
84. Horowitz RI, Freeman PR, Bruzzese J. Efficacy of glutathione therapy in relieving dyspnea associated with COVID-19 pneumonia: a report of 2 cases. Respiratory Medicine Case Rep. 2020;30:101063. doi:10.1016/j.rmcr.2020.101063
85. Whillier S, Raftos JE, Chapman B, Kuchel PW. Role of N-acetylcysteine and cystine in glutathione synthesis in human erythrocytes. Redox Rep. 2009;14(3):115–124. doi:10.1179/135100009x392539
86. Hadzic T, Li L, Cheng N, Walsh SA, Spitz DR, Knudson CM. The role of low molecular weight thiols in T lymphocyte proliferation and IL-2 secretion. J Immunol. 2005;175(12):7965–7972. doi:10.4049/jimmunol.175.12.7965
87. Mak TW, Grusdat M, Duncan GS, et al. Glutathione Primes T Cell Metabolism for Inflammation. Immunity. 2017;46(4):675–689. doi:10.1016/j.immuni.2017.03.019
88. Demedts M, Behr J, Buhl R, et al. High-Dose Acetylcysteine in Idiopathic Pulmonary Fibrosis. N Eng J Med. 2005;353(21):2229–2242. doi:10.1056/NEJMoa042976
89. Lai ZW, Hanczko R, Bonilla E, et al. N-acetylcysteine reduces disease activity by blocking mammalian target of rapamycin in T cells from systemic lupus erythematosus patients: a randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2012;64(9):2937–2946. doi:10.1002/art.34502
90. George PM, Wells AU, Jenkins RG. Pulmonary fibrosis and COVID-19: the potential role for antifibrotic therapy. Lancet Respir Med. 2020;8(8):807–815. doi:10.1016/S2213-2600(20)30225-3
91. Takayama K. In Vitro and Animal Models for SARS-CoV-2 research. Trends Pharmacol Sci. 2020;41(8):513–517. doi:10.1016/j.tips.2020.05.005
92. Rubin EJ, Longo DL, Baden LR. Interleukin-6 Receptor Inhibition in Covid-19 — cooling the Inflammatory Soup. N Eng J Med. 2021;384(16):1564–1565. doi:10.1056/NEJMe2103108
93. McElvaney OJ, Curley GF, Rose-John S, McElvaney NG. Interleukin-6: obstacles to targeting a complex cytokine in critical illness. Lancet Respir Med. 2021;9(6):643-654. doi:10.1016/S2213-2600(21)00103-X
94. Jones SA, Hunter CA. Is IL-6 a key cytokine target for therapy in COVID-19? Nat Rev Immunol. 2021;21(6):337–339. doi:10.1038/s41577-021-00553-8
95. Garbers C, Heink S, Korn T, Rose-John S. Interleukin-6: designing specific therapeutics for a complex cytokine. Nat Rev Drug Discov. 2018;17(6):395–412. doi:10.1038/nrd.2018.45
96. Jones SA, Jenkins BJ. Recent insights into targeting the IL-6 cytokine family in inflammatory diseases and cancer. Nat Rev Immunol. 2018;18(12):773–789. doi:10.1038/s41577-018-0066-7
97. Angriman F, Ferreyro BL, Burry L, et al. Interleukin-6 receptor blockade in patients with COVID-19: placing clinical trials into context. Lancet Respir Med. 2021;9(6):655-664. doi:10.1016/S2213-2600(21)00139-9
98. Bateman DN, Dear JW, Thanacoody HKR, et al. Reduction of adverse effects from intravenous acetylcysteine treatment for paracetamol poisoning: a randomised controlled trial. Lancet. 2014;383(9918):697–704. doi:10.1016/S0140-6736(13)62062-0
99. LiverTox: Clinical and research information on drug-induced liver injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012. Acetylcysteine. [Updated 2016 Nov 7]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548401. Accessed June 15, 2021.
100. Tarbox JA, Bansal A, Peiris AN. Angioedema. JAMA. 2018;319(19):2054. doi:10.1001/jama.2018.4860
101. Bailey B, McGuigan MA. Management of anaphylactoid reactions to intravenous N-acetylcysteine. Ann Emerg Med. 1998;31(6):710–715. doi:10.1016/s0196-0644(98)70229-x
102. Daoud A, Dalhoff KP, Christensen MB, Bøgevig S, Petersen TS. Two-bag intravenous N-acetylcysteine, antihistamine pretreatment and high plasma paracetamol levels are associated with a lower incidence of anaphylactoid reactions to N-acetylcysteine. Clin Toxicol. 2020;58(7):698–704. doi:10.1080/15563650.2019.1675886
103. Schmidt LE, Dalhoff K. Risk factors in the development of adverse reactions to N-acetylcysteine in patients with paracetamol poisoning. Br J Clin Pharmacol. 2001;51(1):87–91. doi:10.1046/j.1365-2125.2001.01305.x
104. Williams TC, Burgers WA. SARS-CoV-2 evolution and vaccines: Cause for concern? Lancet Respir Med. 2021;9(4):333–335. doi:10.1016/S2213-2600(21)00075-8
105. Baric RS. Emergence of a Highly Fit SARS-CoV-2 Variant. N Eng J Med. 2020;383(27):2684–2686. doi:10.1056/NEJMcibr2032888
106. Tatu AL, Nadasdy T, Nwabudike LC. Chitin-lipid interactions and the potential relationship between Demodex and SARS-CoV-2. Dermatol Ther. 2021;34(3):e14935. doi:10.1111/dth.14935
107. Baglivo M, Baronio M, Natalini G, et al. Natural small molecules as inhibitors of coronavirus lipid-dependent attachment to host cells: a possible strategy for reducing SARS-COV-2 infectivity? Acta Biomed. 2020;91(1):161–164. doi:10.23750/abm.v91i1.9402
108. Kubiak K, Sielawa H, Chen W, Dzika E. Endosymbiosis and its significance in dermatology. J Eur Acad Dermatol Venereol. 2018;32(3):347–354. doi:10.1111/jdv.14721
109. Kircik LH, Del Rosso JQ, Layton AM, Schauber J. Over 25 Years of Clinical Experience With Ivermectin: an Overview of Safety for an Increasing Number of Indications. J Drugs Dermatol. 2016;15(3):325–332.
110. Schaller M, Gonser L, Belge K, et al. Dual anti-inflammatory and anti-parasitic action of topical ivermectin 1% in papulopustular rosacea. J Eur Acad Dermatol Venereol. 2017;31(11):1907–1911. doi:10.1111/jdv.14437
111. Caly L, Druce JD, Catton MG, Jans DA, Wagstaff KM. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020;178:104787. doi:10.1016/j.antiviral.2020.104787
112. Chaccour C, Casellas A, Blanco-Di Matteo A, et al. The effect of early treatment with ivermectin on viral load, symptoms and humoral response in patients with non-severe COVID-19: a pilot, double-blind, placebo-controlled, randomized clinical trial. EClinicalMedicine. 2021;32:100720. doi:10.1016/j.eclinm.2020.100720
113. Sharmeen S, Skrtic M, Sukhai MA, et al. The antiparasitic agent ivermectin induces chloride-dependent membrane hyperpolarization and cell death in leukemia cells. Blood. 2010;116(18):3593–3603. doi:10.1182/blood-2010-01-262675
114. López-Medina E, López P, Hurtado IC, et al. Effect of Ivermectin on Time to Resolution of Symptoms Among Adults With Mild COVID-19: a Randomized Clinical Trial. JAMA. 2021;325(14):1426–1435. doi:10.1001/jama.2021.3071
115. Connors JM, Levy JH. COVID-19 and its implications for thrombosis and anticoagulation. Blood. 2020;135(23):2033–2040. doi:10.1182/blood.2020006000
116. Levi M, Thachil J, Iba T, Levy JH. Coagulation abnormalities and thrombosis in patients with COVID-19. Lancet Haematol. 2020;7(6):e438–e440. doi:10.1016/S2352-3026(20)30145-9
117. Wool GD, Miller JL. The Impact of COVID-19 Disease on Platelets and Coagulation. Pathobiology. 2021;88(1):15–27. doi:10.1159/000512007
118. Rostami M, Mansouritorghabeh H. D-dimer level in COVID-19 infection: a systematic review. Expert Rev Hematol. 2020;13(11):1265–1275. doi:10.1080/17474086.2020.1831383
119. Seitz R, Lerch L, Immel A, Egbring R. D-dimer tests detect both plasmin and neutrophil elastase derived split products. Ann Clin Biochem. 1995;32(Pt 2):193–195. doi:10.1177/000456329503200211
120. Ishii T, Doi K, Okamoto K, et al. Neutrophil elastase contributes to acute lung injury induced by bilateral nephrectomy. Am J Pathol. 2010;177(4):1665–1673. doi:10.2353/ajpath.2010.090793
121. Singh P, Schwartz RA. Disseminated intravascular coagulation: a devastating systemic disorder of special concern with COVID-19. Dermatol Ther. 2020;33(6):e14053. doi:10.1111/dth.14053
122. Sadowska AM, Manuel-y-Keenoy B, Vertongen T, et al. Effect of N-acetylcysteine on neutrophil activation markers in healthy volunteers: in vivo and in vitro study. Pharmacol Res. 2006;53(3):216–225. doi:10.1016/j.phrs.2005.11.003
123. Tirouvanziam R, Conrad CK, Bottiglieri T, Herzenberg LA, Moss RB, Herzenberg LA. High-dose oral N-acetylcysteine, a glutathione prodrug, modulates inflammation in cystic fibrosis. Proc Natl Acad Sci U S A. 2006;103(12):4628–4633. doi:10.1073/pnas.0511304103
124. Rubio ML, Martin-Mosquero MC, Ortega M, Peces-Barba G. Oral N-Acetylcysteine Attenuates Elastase-Induced Pulmonary Emphysema in Rats. CHEST. 2004;125(4):1500–1506. doi:10.1378/chest.125.4.1500
125. Mursaleen L, Noble B, Chan SHY, Somavarapu S. N-Acetylcysteine Nanocarriers Protect against Oxidative Stress in a Cellular Model of Parkinson’s Disease. Antioxidants. 2020;9(7):600. doi:10.3390/antiox9070600
126. Lancheros R, Guerrero CA, Godoy-Silva RD. Improvement of N-Acetylcysteine Loaded in PLGA Nanoparticles by Nanoprecipitation Method. J Nanotechnol. 2018;2018:3620373. doi:10.1155/2018/3620373
127. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2020;20(2):101–124. doi:10.1038/s41573-020-0090-8
128. Patra JK, Das G, Fraceto LF, et al. Nano based drug delivery systems: recent developments and future prospects. J Nanobiotechnology. 2018;16(1):71. doi:10.1186/s12951-018-0392-8
129. Sercombe L, Veerati T, Moheimani F, Wu SY, Sood AK, Hua S. Advances and Challenges of Liposome Assisted Drug Delivery. Front Pharmacol. 2015;6:286. doi:10.3389/fphar.2015.00286
130. Heurtault B, Saulnier P, Pech B, Proust JE, Benoit JP. Physico-chemical stability of colloidal lipid particles. Biomaterials. 2003;24(23):4283–4300. doi:10.1016/s0142-9612(03)00331-4
131. Aljihani SA, Alehaideb Z, Alarfaj RE, et al. Enhancing azithromycin antibacterial activity by encapsulation in liposomes/liposomal-N-acetylcysteine formulations against resistant clinical strains of Escherichia coli. Saudi J Biol Sci. 2020;27(11):3065–3071. doi:10.1016/j.sjbs.2020.09.012
132. Karimi Zarchi AA, Faramarzi MA, Gilani K, Ghazi-Khansari M, Ghamami G, Amani A. N-acetylcysteine-loaded PLGA nanoparticles outperform conventional N-acetylcysteine in acute lung injuries in vivo. Int J Polymeric Mater Polymeric Biomaterials. 2017;66(9):443–454. doi:10.1080/00914037.2016.1236339
133. Bailey MM, Berkland CJ. Nanoparticle formulations in pulmonary drug delivery. Med Res Rev. 2009;29(1):196–212. doi:10.1002/med.20140
134. Paranjpe M, Müller-Goymann CC. Nanoparticle-mediated pulmonary drug delivery: a review. Int J Mol Sci. 2014;15(4):5852–5873. doi:10.3390/ijms15045852
135. Danhier F, Ansorena E, Silva JM, Coco R, Le Breton A, Préat V. PLGA-based nanoparticles: an overview of biomedical applications. J Control Release. 2012;161(2):505–522. doi:10.1016/j.jconrel.2012.01.043
136. Maffezzoni E, Notargiacomo M, Agostini S, Gelardi M. Efficacy of a nasal spray containing N-acetylcysteine in hypertonic solution in the treatment of nonallergic chronic rhinitis with goblet cell metaplasia. J Biol Regul Homeost Agents. 2020;34(6):2345–2352. doi:10.23812/20-411-l
137. Macchi A, Terranova P, Castelnuovo P. Recurrent acute rhinosinusitis: a single blind clinical study of N-acetylcysteine vs ambroxol associated to corticosteroid therapy. Int J Immunopathol Pharmacol. 2012;25(1):207–217. doi:10.1177/039463201202500123
138. Cogo A, Chieffo A, Farinatti M, Ciaccia A. Efficacy of topical tuaminoheptane combined with N-acetyl-cysteine in reducing nasal resistance. A double-blind rhinomanometric study versus xylometazoline and placebo. Arzneimittelforschung. 1996;46(4):385–388.
139. Lane CJ, Redding DR, Gonzalez KA, Cardenas VJ, Boldogh I, Sur S. Topical N-acetyl cysteine (NAC) Reduces Late Phase Nasal Symptoms Following Ragweed Challenge. J Allergy Clin Immunol. 2009;123(2):S53. doi:10.1016/j.jaci.2008.12.170
140. Yilmaz B, Türkçü G, Şengül E, Gül A, Özkurt FE, Akdağ M. Efficacy of N-Acetylcysteine on Wound Healing of Nasal Mucosa. J Craniofac Surg. 2015;26(5):e422–6. doi:10.1097/scs.0000000000001880
141. Matsuyama T, Morita T, Horikiri Y, Yamahara H, Yoshino H. Enhancement of nasal absorption of large molecular weight compounds by combination of mucolytic agent and nonionic surfactant. J Control Release. 2006;110(2):347–352. doi:10.1016/j.jconrel.2005.09.047
142. Kim D, Kim YH, Kwon S. Enhanced nasal drug delivery efficiency by increasing mechanical loading using hypergravity. Sci Rep. 2018;8(1):168. doi:10.1038/s41598-017-18561-x
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.