Back to Journals » Infection and Drug Resistance » Volume 12
Multiresistant Fusarium Pathogens on Plants and Humans: Solutions in (from) the Antifungal Pipeline?
Authors Al-Hatmi AMS ![]() , de Hoog GS, Meis JF
, de Hoog GS, Meis JF ![]()
Received 9 September 2019
Accepted for publication 22 November 2019
Published 28 November 2019 Volume 2019:12 Pages 3727—3737
DOI https://doi.org/10.2147/IDR.S180912
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Abdullah MS Al-Hatmi,1–3 G Sybren de Hoog,2,3 Jacques F Meis3,4
1Ministry of Health, Directorate General of Health Services, Ibri, Oman; 2Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands; 3Centre of Expertise in Mycology Radboud University Medical Centre/Canisius Wilhelmina Hospital, Nijmegen, the Netherlands; 4Department of Medical Microbiology and Infectious Diseases, Canisius Wilhelmina Hospital, Nijmegen, the Netherlands
Correspondence: Abdullah MS Al-Hatmi
Directorate General of Health Services, PO Box 46, 516 Al-Akdhar, Ibri, Oman
Tel +968 25691905
Fax +968 25691915
Email [email protected]
Abstract: The fungal genus Fusarium contains numerous plant pathogens causing considerable economic losses. In addition, Fusarium species are emerging as opportunistic human pathogens causing both superficial and systemic infections. Appropriate treatment of Fusarium infections in a clinical setting of neutropenia is currently not available. ESCMID and ECMM joint guidelines, following the majority of published studies, suggest early therapy with amphotericin B and voriconazole, in conjunction with surgical debridement and reversal of immunosuppression. In this review, we elaborate on the trans-kingdom pathogenicity of Fusarium. Intrinsic resistance to several antifungal drugs and the evolution of antifungal resistance over the years are highlighted. Recent studies present novel compounds that are effective against some pathogenic fungi including Fusarium. We discuss the robust and dynamic antifungal pipeline, including results from clinical trials as well as preclinical data that might appear beneficial for patients with invasive fusariosis.
Keywords: Fusarium, trans-kingdom, novel compounds, antifungal pipeline, E1210, SCY-078, ASP2397, MGCD290, olorofim, AR-12, isavuconazole, efinaconazole, luliconazole
Trans-Kingdom Pathogenicity
Fusarium is a diverse genus of fungi containing several hundreds of species.1–3 Some plant pathogenic members of the genus are restricted to a single host species, whereas others have a broad host range. Economic losses in agriculture may be considerable, and the genus is listed as one of the most destructive plant pathogenic fungi.4,5 This genus is also frequently involved in vertebrate infection.6,7 Although the conditions in host tissues of plants and animals are very different, nearly all 24 taxa that have been described to occur in human infections as real cases8 have also been reported from plant diseases. Fusarium is capable of infecting plants as well as humans, a phenomenon known as trans-kingdom pathogenicity.9 This unusual ability has been demonstrated in numerous studies.10–18 Fusarium oxysporum f. sp. lycopersici, F. keratoplasticum, and F. falciforme are known for their pathogenicity to plants, but have also been reported from humans and other mammals.11–13 Similarly, F. pisi, F. temperatum, F. ramigenum, F. musae, Fusarium solani sensu stricto, and F. volatile have been recovered from living plants, while also their clinical relevance has been underlined.14,19–21 Cross-kingdom pathogenicity is in obvious conflict with plant host specificity. In F. oxysporum, small conditionally dispensable chromosomes carrying virulence factors, which are horizontally transmitted between germinating cells of different lineages in response to signals from a suitable habitat and which differentiate into infection hyphae,22,23 may explain the sudden outbreaks observed in agricultural settings. Three Fusarium oxysporum mitogen-activated protein kinases (MAPKs) have distinct and complementary roles in stress adaptation and cross-kingdom pathogenicity.24 Under selective pressure of host conditions, lineages show adaptation enhancing survival and replication. Segorbe et al25 underlined the role of MAPK genes that contribute to the regulation of development, stress response and virulence in plants and animals. Instantaneous use of windows of opportunity is instrumental for rapid expansion. Van Baarlen et al noted a molecular similarity between hypothetical virulence factors in plant and human pathogens,26 but among fungi in general such abilities are rarely combined8 and thus cannot be generalized; more often, opportunism is combined with extremotolerance.27 Cross-kingdom pathogenicity may thus be considered rather unique to Fusarium.
Fusarium oxysporum shows virulence in the Galleria mellonella infection model.28 In humans, the infection mechanism is largely unknown. In general, immunocompromised individuals are highly susceptible to develop Fusarium disseminated infections, especially during neutropenia.11,29,30 In otherwise healthy individuals, fusariosis generally remains a superficial infection;31 the fungi are quite commonly isolated from dermatological samples in the tropics.32 Fusarium keratitis, mostly initiated by traumatic inoculation of contaminated materials such as plant leaves,19 is a major public health concern with an estimated global burden of about 1–1.2 million cases annually.33
Evolving Taxonomy
The taxonomy of the genus Fusarium has been affected by changes in species concept. During the last centuries, the number of recognized species by traditional taxonomists varied enormously, from as few as nine species to several hundreds.34 In 1910, Apple and Wollenweber grouped all asexual fungi producing multicellular macroconidia with croissant shape from slimy sporodochia in Fusarium.35 Wollenweber and Reinking used the differences in morphology to organize the genus into 16 sections. These sections contained 65 species, 55 varieties, and 22 host-specific formae on the basis of the color of stroma, the presence and absence of sclerotia, and the length and number of septations in macroconidia.36 The characteristics used by those authors were the shape of conidia, microconidia, macroconidia, chlamydospores, basal foot cells and phialides. Also, the location of chlamydospores and other types of conidia was considered. Booth simplified this system to only 14 species.37 The taxonomy proposed by Gerlach and Nirenberg was similar to that of Wollenweber and Reinking, recognizing 21 species.38 Leslie and Summerell used morphological, biological and phylogenetic information for reclassification; they concluded that 70 species could be distinguished.34 At present, with the dawn of molecular sequencing, more than 300 species are recognized, grouped in 22 species complexes, all differing in morphology, host association, and particularly in molecular parameters.2 Fusarium was one of the first fungal groups where the term “species complex” was used for a series of closely related species. The term “species complex” has been defined,8 elaborating on early papers for use in clinical routine by Chen et al39 and Kwon-Chung et al,40 as a monophyletic group which are different at the molecular epidemiological level but are functionally indistinguishable. In other words, there are identifiable discontinuities in their features, but the differences are not meaningful for practice. Variations or discontinuities may lead to reproductive barriers and speciation. Whether the speciation process has advanced sufficiently remains a matter of debate with every single species cluster.
Geiser et al brought together a consortium of clinical and phytopathological experts and launched a plea for nomenclatural stability with preservation of the name Fusarium for all clinically relevant species complexes.41 The proposal was not effective, as Lombard et al moved the F. solani species complex to the genus Neocosmospora, and the F. dimerum species complex to a new genus Bisifusarium, on phylogenetic grounds.42 As these genera include species with substantial significance as plant and human pathogens, transfers have not widely been accepted. However, the same research group recently reported that 68 species are accepted in the genus Neocosmospora, 29 of them described as new, while 13 new combinations were made and 11 species remained as yet undescribed.43 For the sake of nomenclature stability in the clinical field, we adhere in this review to Fusarium as best known descriptor for fungi with morphological and ecological features in the sense of Wollenweber and Reinking,36 as agents of “fusariosis.”
Intrinsic Resistance to Antifungals
Most research on antifungals focuses on acquired resistance obtained by mutations in resistance genes, for example, the wealth of information available on Aspergillus fumigatus acquiring azole resistance triggered by exposure to agricultural antifungals in the environment.44–47 Among agricultural fungicides, difenoconazole had the lowest activity against Fusarium solani SC with MICs of >32 mg/mL.48 Herkert et al suggested a similar selective pressure on environmental Fusarium strains as that seen with Aspergillus.48 However, natural, intrinsic resistance, which is independent of previous antifungal exposure, has largely been neglected.49 Some fungi are intrinsically resistant to single drugs (e.g., C. krusei to fluconazole or C. lusitaniae to amphotericin B), while others are resistant to different classes of antifungals (e.g., C. auris, some strains resistant to all antifungals).50–52 Among the filamentous fungi, members of two adjacent orders are multiresistant, i.e., the Microascales (genera Scedosporium, Lomentospora, Scopulariopsis) and the Hypocreales (genera Acremonium, Fusarium, Trichoderma).53 Discovery of genes conveying intrinsic resistance may not only provide drug targets for the development of new antifungals, but may also repurpose antifungals with limited effectivity by blocking the intrinsic drug resistance genes of the pathogens.54
Nearly all Fusarium species are among those fungi that have inherent structural and functional characteristics to resist antifungals without prior exposure. The reason why this phenomenon occurs naturally in Fusarium and other members of the above orders is unknown. Resistance has been observed for amphotericin B, itraconazole, fluconazole and echinocandins, but variable MIC results are recorded with the newer triazoles (posaconazole, voriconazole, and isavuconazole).1 The molecular mechanisms of intrinsic resistance in Fusarium have not been described yet. However, a hypothetical molecular mechanism has been proposed by Fan et al who showed that CYP51 in Fusarium has three paralogues of sterol 14α-demethylase cytochrome P450 (CYP51A, -B, and -C), with CYP51C being restricted to Fusarium.55 The same authors stated that CYP51A deletion increased the sensitivity of Fusarium graminearum to azoles.55 Katiyar et al reported that mutations in the FKS1 gene,56 i.e., amino acid substitutions P647A and F639Y in FKS1, contribute to intrinsic echinocandin resistance in Fusarium solani. The major mechanism responsible for high-level azole resistance in clinical species of Candida is overexpression of plasma membrane efflux pumps,57 and this may also reduce azole susceptibility in Fusarium. Under the influence of azoles, various efflux mechanisms are triggered in F. graminearum, such as ATP-binding cassette (ABC) transporters to actively transport molecules across the cell membrane, reducing their impact on viability.58 Experiments with the agricultural antifungal tebuconazole indicated the presence of different resistance mechanisms in F. graminearum. One of the phenotypes conveyed resistance to azoles, whereas another was related to multidrug resistance.58
Heteroresistance is another example of variation in drug susceptibility within a population. It was already reported that in Candida albicans, Cryptococcus neoformans, Cryptococcus gattii, Aspergillus fumigatus and Aspergillus flavus, single cells can give rise to progeny with heterogeneous resistance phenotypes resistant to the azoles.59–61 This phenomenon may also be present in Fusarium.
Evolution of Antifungal Resistance
In recent decades, there has been much improper use of azoles, especially in agriculture. Resistance to azole fungicides in Aspergillus fumigatus as a human opportunist and Mycosphaerella fijiensis as a plant pathogen has been recognized during the last two decades, and the resistance mechanism is based on analogous genes (CYP51).62,63 In 1997, the first azole-resistant clinical isolate of Aspergillus fumigatus was reported,64 followed shortly thereafter by resistant A. fumigatus strains in the (agricultural) environment, exhibiting cross-resistance with fungicides.65
Due to the high fungicide pressure in the environment, resistance can also increase in Fusarium, although the main hypothesis is that resistance in Fusarium was already present prior to fungicide exposure. According to Lucas et al, in some organisms, the level of pre‐existing resistance is high and should, therefore, be considered as intrinsic resistance.66 In other organisms, processes such as increasing efflux pump activity or metabolism of toxins are not enough to confer intrinsic resistance to fungicides, but under high pressure of fungicides, resistance variation might occur.67,68 In addition to the problem of intrinsic variation of drug susceptibility among Fusarium species, we need to add the emerging issue of acquired resistance (if present), which refers to the ability of Fusarium species to evolutionarily develop mechanisms that lower their susceptibility toward certain antifungals. In this scenario, Fusarium, as a plant pathogen, as well as Aspergillus species, which are saprobic on plant debris, all occur in the environment,69 and therefore Fusarium is also exposed to the fungicides that are currently in use in agriculture. Consequently, it is useful to verify whether changes can be found in resistance profiles among clinical Fusarium isolates, in analogy to Aspergillus. We have checked this hypothesis by selecting Fusarium isolates from culture collections accessed before 1970 and after 1990 and tested their MICs against several antifungals including amphotericin B, itraconazole, posaconazole, voriconazole, isavuconazole, propiconazole, tebuconazole and difenoconazole using CLSI and EUCAST microdilution methods. We found that on the timescale and also comparing different methods, no statistically significant difference was revealed (p 0.122) between the MICs of strains from before 1970 and strains isolated after 1990. However, strains of Fusarium differed among each other in MIC values (Tables 1 and 2),1,70 showing that high MICs in Fusarium are strain specific. Current data show that some species/strains have higher MIC values than others. Our comparative antifungal susceptibility testing with clinical and environmental isolates from before 1970 and after 1990 showed that Fusarium indeed may have intrinsic resistance.
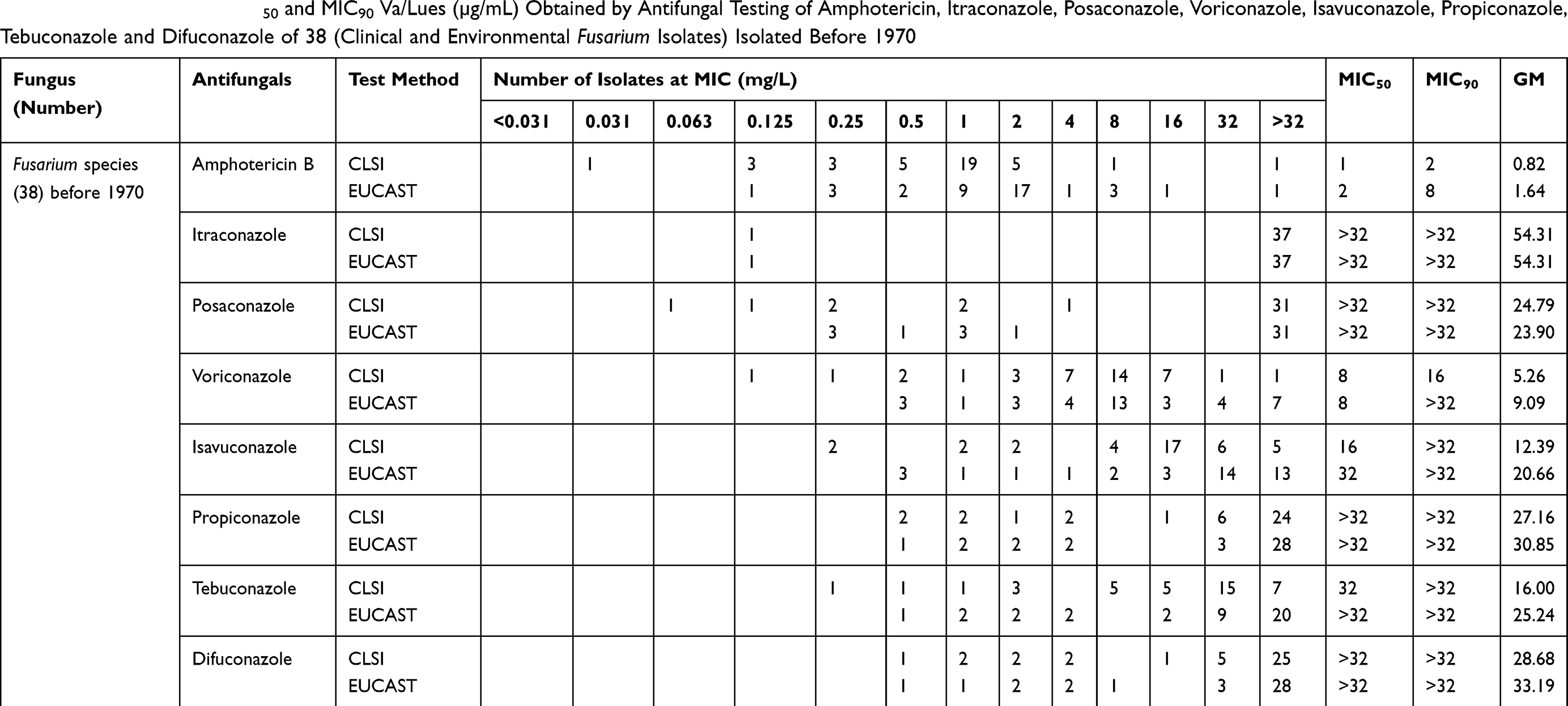 |
Table 1 GM, MIC Range, MIC50 and MIC90 Va/Lues (µg/mL) Obtained by Antifungal Testing of Amphotericin, Itraconazole, Posaconazole, Voriconazole, Isavuconazole, Propiconazole, Tebuconazole and Difuconazole of 38 (Clinical and Environmental Fusarium Isolates) Isolated Before 1970 |
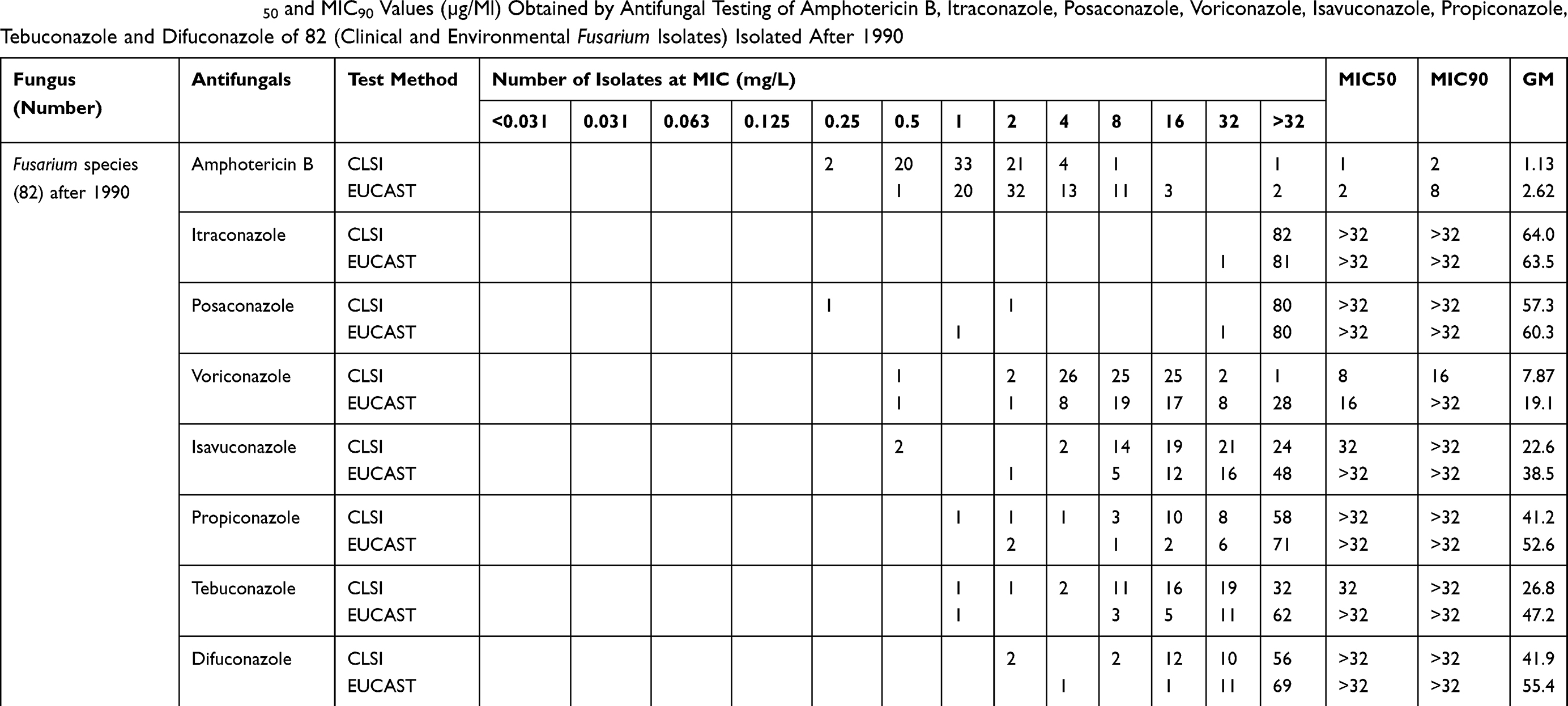 |
Table 2 GM, MIC Range, MIC50 and MIC90 Values (µg/Ml) Obtained by Antifungal Testing of Amphotericin B, Itraconazole, Posaconazole, Voriconazole, Isavuconazole, Propiconazole, Tebuconazole and Difuconazole of 82 (Clinical and Environmental Fusarium Isolates) Isolated After 1990 |
Current and New Drugs in the Pipeline
Here we discuss leading articles on antifungals that are available at present and those that are in the pipeline for use in patients with fusariosis (keratitis, onychomycosis and disseminated infections). The standard for treatment of keratitis due to Fusarium has not been determined, but is most commonly managed by topical application of natamycin or amphotericin B.71 Natamycin was found to be active against Fusarium species both in vitro and in vivo, and recently it has been recommended in combination with voriconazole as the mainstay of treatment for Fusarium keratitis.53 Early detection and installment of therapy are essential.72 Voriconazole has been used in regimens combining topical (10 mg/mL eye drops) and oral (400 mg/day) administration, with good results, particularly when a hypopyon is present.71 Recently, Oliveira et al and Kunt et al reported that chlorhexidine demonstrated in vitro antifungal activity against Fusarium strains isolated from keratitis lesions.73,74 Todokoro et al demonstrated that luliconazole exhibited strong in vitro antifungal activity against a broad range of filamentous fungi including Fusarium species.75
Combination therapy of natamycin with voriconazole, itraconazole or micafungin showed synergism,76 although Prajna et al reported in a clinical trial equal or inferior efficacy of 1% voriconazole compared to 5% natamycin alone in eye drops.77 Rees et al evaluated in vitro activities of natamycin and voriconazole in combination with four non-antifungal ophthalmic agents (5-fluorouracil, dorzolamide, EDTA and timolol).78 In eight Fusarium ocular isolates, resistance was noted to both natamycin and voriconazole. The data suggested that commonly used ophthalmic agents enhance the in vitro activity of antifungal drugs against drug-recalcitrant ocular fusariosis when used in combination.78 Posaconazole and ravuconazole are new azoles that have yet to be topically applied in ophthalmic settings.
Therapeutic outcomes of Fusarium onychomycoses, particularly of subungual cases, are variable. No standard treatment of onychomycosis due to Fusarium has as yet been identified. In general, itraconazole is applied either daily or intermittently.71 A second commonly used drug is terbinafine, sometimes combined with topical ciclopirox and amorolfine lacquer or with keratolytics such as urea.79,80 Tupaki-Sreepurna et al presented susceptibility profiles of 44 common nondermatophyte fungi including Fusarium against efinaconazole and showed excellent in vitro activity.81 Luliconazole and lanoconazole are new imidazole antifungal agents with broad-spectrum antifungal activity used clinically as topical drugs in the treatment of onychomycosis and dermatophytosis.82 Abastabar et al concluded that luliconazole, lanoconazole and efinaconazole exhibit potent in vitro activity against clinical and environmental Fusarium species, and these compounds might be an option for treating onychomycosis due to Fusarium.83 The same authors reported that the in vitro antifungal activity of efinaconazole, with a GM MIC of 0.85 µg/mL, was superior to that of amphotericin B, natamycin, other triazoles and echinocandins.83
Despite advances in therapy and early diagnosis, invasive fusariosis remains associated with high morbidity and with up to 70% mortality in case of dissemination.84 Research results confirm a high level of resistance, regardless of the species or strain of Fusarium involved. The high MIC levels are worrying and are imperative for the development of new drugs. The prognosis of patients with disseminated fusariosis is poor and is mainly associated with reversal of neutropenia, but novel antifungals and formulations may improve outcomes also.49 Isavuconazole has MICs against Fusarium species that are equivalent to or higher than other triazoles (1 to ≥16 μg/mL).69,85 In two clinical trials (SECURE and VITAL), seven patients with disseminated fusariosis were treated with isavuconazole as primary therapy, resulting in 44% 90-day survival.86,87
Fusarium antifungal resistance remains a significant problem for patients with compromised immunity and at high risk for invasive infection, and therefore there is an urgent need for novel therapeutic compounds and strategies. The following drugs are in the pipeline and have been studied and investigated in phase 1–2 preclinical trials (Table 3). ASP2397 is a new antifungal compound, producing its antifungal effects by disruption of the intracellular membrane. It has a modest in vitro antifungal activity (≤8 μg/mL) against Fusarium species.88 AR-12 (celecoxib derivative) has shown antifungal activity against different fungi such as Cryptococcus neoformans, Candida albicans and Scedosporium species and was active against Fusarium species at a MIC of 4 μg/mL.89,90 F901318 (olorofim, F2G) is a member of a novel class of antifungals, the orotomides, acting by interference of pyrimidine biosynthesis in the fungal cell. This compound displayed excellent activity against a broad range of pathogens.91 It has been used as an intravenous and oral agent for use in systemic mold infections.92 However, olorofim was tested against Fusarium species with variable results.93 Full inhibition was achieved with F. proliferatum (MIC 0.016 μg/mL), 50% inhibition with F. solani (MIC 1–2 μg/mL), while no inhibition was observed with F. dimerum.93 APX001 (E1210/1211, famnogepix) is an antifungal compound that is still under development.92 Reportedly, it has potent in vitro antifungal activity against molds that are difficult to treat, such as Fusarium, where MIC values of 0.015–0.25 μg/mL94 and 0.12 μg/mL95 were obtained. The arylamidine enfumafungin (MK-3118, SCY-078, ibrexafungerp) is a novel, orally bioavailable 1,3-β-d-glucan synthesis inhibitor. The compound was highly active against multidrug resistant Candida albicans and C. glabrata isolates,96 but showed no or poor activity against Fusarium species.97 However, the molecular variation SCY-078 T-2307 that inhibits fungal growth by interference with cellular metabolism98,99 has potent in vitro activity (0.125 μg/mL) against Fusarium solani.100 The histone deacetylase 2 inhibitor MGCD290 is effective in combination with both azoles and echinocandins in vitro and in animal models.101,102 The combination with voriconazole demonstrated synergy against six out of eight Fusarium isolates.101
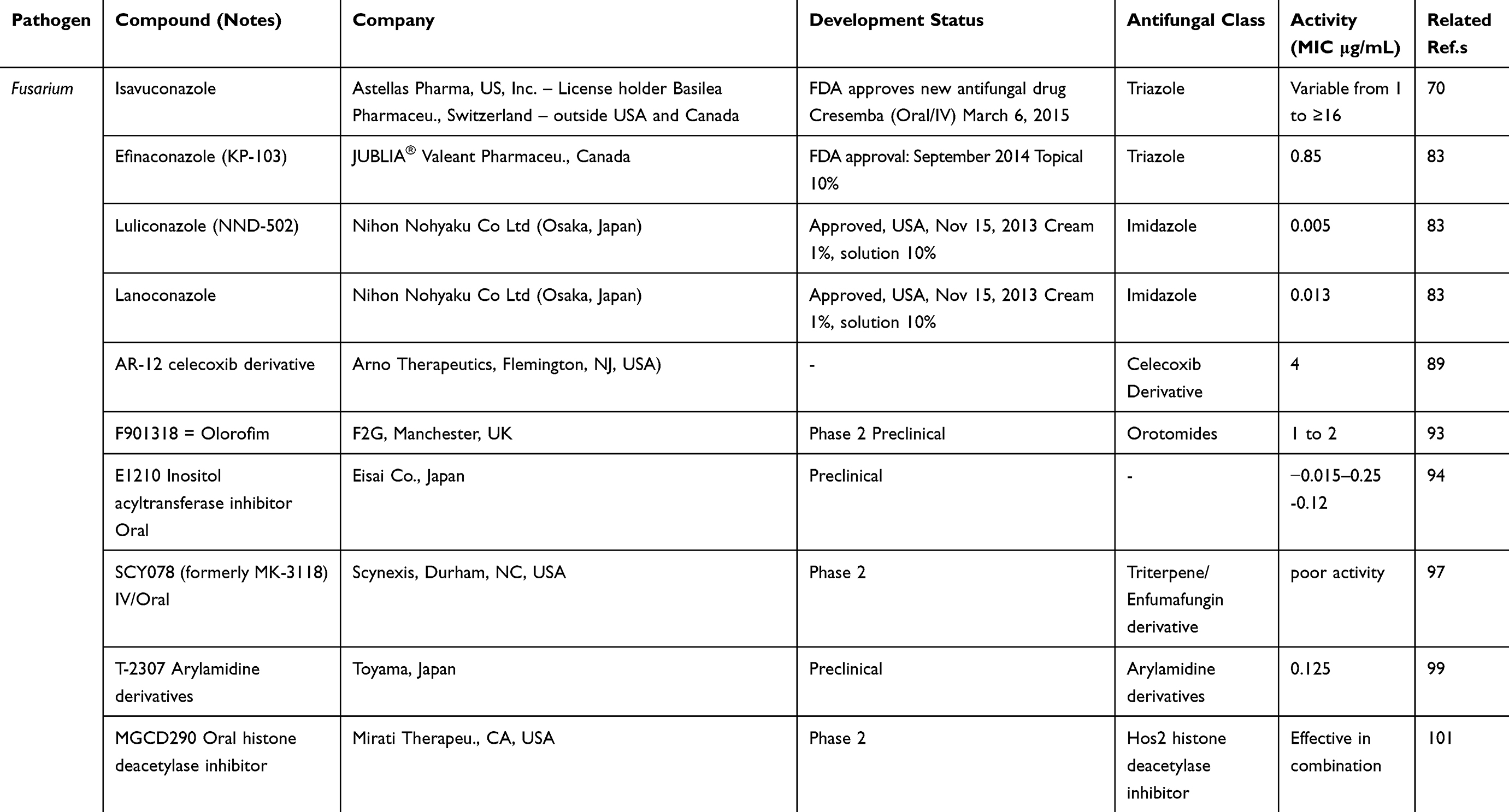 |
Table 3 Antifungal Product Type in the Pipeline, Pathogen, Phase of Clinical Trial and Expected Activity Against Fusarium |
In conclusion, Fusarium is one of the few fungi capable of infecting plants as well as humans, a phenomenon known as trans-kingdom pathogenicity. Intrinsic resistance and acquired resistance of Fusarium species are a threat to both human medicine and agriculture. Especially immunocompromised patients with longstanding neutropenia and disseminated fusariosis have a poor prognosis. Reversal of neutropenia is life-saving, but new classes of antifungals in the pipeline may improve the outcome of these severe opportunistic infections.
Disclosure
Jacques F Meis reports personal fees from Scynexis, personal fees from United Medical, personal fees from Gilead, personal fees from TEVA, grants from F2G, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Al-Hatmi AMS, Meis JF, de Hoog GS. Fusarium: molecular diversity and intrinsic drug resistance. PLoS Pathog. 2016;12(4):e1005464. doi:10.1371/journal.ppat.1005464
2. O’Donnell K, Rooney AP, Proctor RH, et al. Phylogenetic analyses of RPB1 and RPB2 support a middle cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet Biol. 2013;52:20–31. doi:10.1016/j.fgb.2012.12.004
3. O’Donnell K, Ward TJ, Robert VARG, Crous PW, Geiser DM, Kang S. DNA sequence-based identification of Fusarium: current status and future directions. Phytoparasitica. 2015;43:583–595. doi:10.1007/s12600-015-0484-z
4. Nelson PE, Dignani MC, Anaissie EJ. Taxonomy, biology, and clinical aspects of Fusarium species. Clin Microbiol Rev. 1994;7:479–504. doi:10.1128/CMR.7.4.479
5. Dean R, Van Kan JA, Pretorius ZA, et al. The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol. 2012;13(4):414–430. doi:10.1111/j.1364-3703.2011.00783.x
6. Sexton AC, Howlett BJ. Parallels in fungal pathogenesis on plant and animal hosts. Eukaryot Cell. 2006;5:1941–1949. doi:10.1128/EC.00277-06
7. Nucci M, Anaissie E. Fusarium infections in immunocompromised patients. Clin Microbiol Rev. 2007;20:695–704. doi:10.1128/CMR.00014-07
8. De Hoog GS, Guarro J, Gené J, et al. Atlas of Clinical Fungi.
9. Gauthier G, Keller N. Crossover fungal pathogens: the biology and pathogenesis of fungi capable of crossing kingdoms to infect plants and humans. Fungal Genet Biol. 2013;61:146–157. doi:10.1016/j.fgb.2013.08.016
10. Schroers HJ, Samuels GJ, Zhang N, Short DP, Juba J, Geiser DM. Epitypification of Fusisporium (Fusarium) solani and its assignment to a common phylogenetic species in the Fusarium solani species complex. Mycologia. 2016;108(4):806–819. doi:10.3852/15-255
11. Short DP, O’Donnell K, Thrane U, et al. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet Biol. 2013;53:59–70. doi:10.1016/j.fgb.2013.01.004
12. Ortoneda M, Guarro J, Madrid MP, et al. Fusarium oxysporum as a multi-host model for the genetic dissection of fungal virulence in plants and mammals. Infect Immun. 2004;72(3):1760–1766. doi:10.1128/IAI.72.3.1760-1766.2004
13. Nalim FA, Samuels GJ, Wijesundera RL, Geiser DM. New species from the Fusarium solani species complex derived from perithecia and soil in the old World tropics. Mycologia. 2011;103(6):1302–1330. doi:10.3852/10-307
14. Šišić A, Baćanović-Šišić J, Al-Hatmi AMS, et al. The ‘forma specialis’ issue in Fusarium: A case study in Fusarium solani f. sp. pisi. Sci Rep. 2018;8(1):1252. doi:10.1038/s41598-018-19779-z
15. Mehl HL, Epstein L. Fusarium solani species complex isolates conspecific with Fusarium solani f. sp. cucurbitae race 2 from naturally infected human and plant tissue and environmental sources are equally virulent on plants, grow at 37 degrees C and are interfertile. Environ Microbiol. 2007;9(9):2189–2199. doi:10.1111/j.1462-2920.2007.01333.x
16. Zhang N, O’Donnell K, Sutton DA, et al. Members of the Fusarium solani species complex that cause infections in both humans and plants are common in the environment. J Clin Microbiol. 2006;44(6):2186–2190. doi:10.1128/JCM.00120-06
17. Edwards ET. Studies on the Gibberella fujikuroi var. subglutinans the hitherto undescribed ascigerous stage of Fusarium moniliforme var. subglutinans and its pathogenicity on maize in New South Wales. Dep Agric New South Wales Sci Bull. 1935;49:1–68.
18. Schäfer K, Di Pietro A, Gow NA, MacCallum D. Murine model for Fusarium oxysporum invasive fusariosis reveals organ-specific structures for dissemination and long-term persistence. PLoS One. 2014;9(2):e89920. doi:10.1371/journal.pone.0089920
19. Al-Hatmi AM, Bonifaz A, de Hoog GS, et al. Keratitis by Fusarium temperatum, a novel opportunist. BMC Infect Dis. 2014;14:588. doi:10.1186/s12879-014-0588-y
20. Boral H, van Diepeningen A, Erdem E, et al. Mycotic keratitis caused by Fusarium solani sensu stricto (FSSC5): a case series. Mycopathologia. 2018;183:835–840. doi:10.1007/s11046-018-0280-7
21. Al-Hatmi AMS, Sandoval-Denis M, Nabet C, et al. Fusarium volatile, a new potential pathogen from a human respiratory sample. Fungal Syst Evol. 2019;4:71–181. doi:10.3114/fuse.2019.04.09
22. Ma LJ, van der Does HC, Borkovich KA, et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature. 2010;464:367–373. doi:10.1038/nature08850
23. Vlaardingerbroek I, Beerens B, Rose L, et al. Exchange of core chromosomes and horizontal transfer of lineage‐specific chromosomes in Fusarium oxysporum. Environm Microbiol. 2016;18:3702–3713. doi:10.1111/1462-2920.13281
24. Perez-Nadales E, Di Pietro A. The membrane mucin Msb2 regulates invasive growth and plant infection in Fusarium oxysporum. Plant Cell. 2011;23:1171–1185. doi:10.1105/tpc.110.075093
25. Segorbe D, Di Pietro A, Perez-Nadales E, Turra D. Three Fusarium oxysporum mitogen-activated protein kinases (MAPKs) have distinct and complementary roles in stress adaptation and cross-kingdom pathogenicity. Mol Plant Pathol. 2017;18(7):912–924. doi:10.1111/mpp.2017.18.issue-7
26. Van Baarlen P, Van Belkum A, Summerbell R, Crous PW, Thomma B. Molecular mechanisms of pathogenicity: how do pathogenic microorganisms develop cross-kingdom host jumps? FEMS Microbiol Rev. 2007;31:239–277. doi:10.1111/j.1574-6976.2007.00065.x
27. Gostincar C, Zajc J, Lenassi M, et al. Fungi between extremotolerance and opportunistic pathogenicity on humans. Fungal Divers. 2018;93:195–213. doi:10.1007/s13225-018-0414-8
28. Navarro-Velasco GY, Prados-Rosales RC, Ortíz-Urquiza A, Quesada-Moraga E, Di Pietro A. Galleria mellonella as model host for the trans-kingdom pathogen Fusarium oxysporum. Fungal Genet Biol. 2011;48(12):1124–1129. doi:10.1016/j.fgb.2011.08.004
29. Moretti ML, Busso-Lopes AF, Tararam CA, et al. Airborne transmission of invasive fusariosis in patients with hematologic malignancies. PLoS One. 2018;13:e0196426. doi:10.1371/journal.pone.0196426
30. Nucci M, Shoham S, Abdala E, et al. Outcomes of patients with invasive fusariosis who undergo further immunosuppressive treatments, is there a role for secondary prophylaxis? Mycoses. 2019;62(5):413–417. doi:10.1111/myc.2019.62.issue-5
31. Wickern GM. Fusarium allergic fungal sinusitis. J Allergy Clin Immunol. 1993;92:624–625. doi:10.1016/0091-6749(93)90087-V
32. van Diepeningen AD, Feng P, Ahmed S, et al. Spectrum of Fusarium infections in tropical dermatology evidenced by multilocus sequencing typing diagnostics. Mycoses. 2015;58:48–57. doi:10.1111/myc.12273
33. Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, White TC. Hidden killers: human fungal infections. Sci Transl Med. 2012;4(165):165rv13. doi:10.1126/scitranslmed.3004404
34. Leslie JF, Summerell BA. The Fusarium Laboratory Manual. Ames, IA: Blackwell Publishing; 2006.
35. Apple O, Wollenweber HW. Grundlagen einer Monographie der Gattung Fusarium (Link). In Arb K Biol Anst F Land U Forstw. 1910;8:1–207.
36. Wollenweber HW, Reinking OA. The Fusaria: Their Description Injurious Effects and Control. Vol. 8. Berlin: Paul Parey; 1935:1–135.
37. Booth C. The Genus Fusarium. Kew: Commonwealth Mycological Institute; 1971:237.
38. Gerlach W, Nirenberg H. The genus Fusarium-a pictorial atlas. Mitt Biol Bundes Anst Land- Forstw. 1982;209:1–406.
39. Chen M, Zeng J, De Hoog GS, et al. The “species complex” issue in clinically relevant fungi: a case study in Scedosporium apiospermum. Fungal Biol. 2016;120:137–146. doi:10.1016/j.funbio.2015.09.003
40. Kwon-Chung KJ, Bennett JE, Wickes BL, et al. The case for adopting the “species complex” nomenclature for the etiologic agents of cryptococcosis. mSphere. 2017;2:e00357–16. doi:10.1128/mSphere.00357-16
41. Geiser DM, Aoki T, Bacon CW, et al. One fungus, one name: defining the genus Fusarium in a scientifically robust way that preserves longstanding use. Phytopathology. 2013;103:400–408. doi:10.1094/PHYTO-07-12-0150-LE
42. Lombard L, van der Merwe NA, Groenewald JZ, Crous PW. Generic concepts in Nectriaceae. Stud Mycol. 2015;80:189–245. doi:10.1016/j.simyco.2014.12.002
43. Sandoval-Denis M, Lombard L, Crous PW. Back to the roots: a reappraisal of Neocosmospora. Persoonia. 2019;43:90–185.
44. Chowdhary A, Kathuria S, Xu J, Meis JF. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013;9(10):e1003633. doi:10.1371/journal.ppat.1003633
45. Chowdhary A, Meis JF. Emergence of azole resistant Aspergillus fumigatus and One Health: time to implement environmental stewardship. Environ Microbiol. 2018;20(4):1299–1301. doi:10.1111/emi.2018.20.issue-4
46. Alvarez-Moreno C, Lavergne RA, Hagen F, Morio F, Meis JF, Le Pape P. Fungicide-driven alterations in azole-resistant Aspergillus fumigatus are related to vegetable crops in Colombia, South America. Mycologia. 2019;111(2):217–224. doi:10.1080/00275514.2018.1557796
47. Meis JF, Chowdhary A, Rhodes JL, Fisher MC, Verweij PE. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos Trans R Soc Lond B Biol Sci. 2016;371(1709):20150460. doi:10.1098/rstb.2015.0460
48. Herkert PF, Al-Hatmi AMS, De Oliveira Salvador GL, et al. Molecular characterization and antifungal susceptibility of clinical Fusarium species from Brazil. Front Microbiol. 2019;10:737. doi:10.3389/fmicb.2019.00737
49. Wiederhold NP. Antifungal resistance: current trends and future strategies to combat. Infect Drug Resist. 2017;10:249–259. doi:10.2147/IDR.S124918
50. Walsh TJ, Groll A, Hiemenz J, Fleming R, Roilides E, Anaissie E. Infections due to emerging and uncommon medically important fungal pathogens. Clin Microbiol Infect. 2004;10(Suppl 1):48–66. doi:10.1111/j.1470-9465.2004.00839.x
51. Lackner M, Hagen F, Meis JF, et al. Susceptibility and diversity in the therapy-refractory genus Scedosporium. Antimicrob Agents Chemother. 2014;58(10):5877–5885. doi:10.1128/AAC.03211-14
52. Chowdhary A, Prakash A, Sharma C, et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009-17) in India: role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J Antimicrob Chemother. 2018;73(4):891–899. doi:10.1093/jac/dkx480
53. Al-Hatmi AMS, Bonifaz A, Ranque S, De Hoog GS, Verweij PE, Meis JF. Current antifungal treatment of fusariosis. Int J Antimicrob Agents. 2018;51(3):326–332. doi:10.1016/j.ijantimicag.2017.06.017
54. Scorzoni L, Silva ACAP, Marcos CM, et al. Antifungal therapy: new advances in the understanding and treatment of mycosis. Front Microbiol. 2017;8:1–23.
55. Fan J, Urban M, Parker JE, et al. Characterization of the sterol 14α-demethylases of Fusarium graminearum identifies a novel genus-specific CYP51 function. New Phytol. 2013;198:821–835. doi:10.1111/nph.12193
56. Katiyar SK, Edlind TD. Role for Fks1 in the intrinsic echinocandin resistance of Fusarium solani as evidenced by hybrid expression in Saccharomyces cerevisiae. Antimicrob Agents Chemother. 2009;53(5):1772–1778. doi:10.1128/AAC.00020-09
57. Cannon RD, Lamping E, Holmes AR, et al. Efflux-mediated antifungal drug resistance. Clin Microbiol Rev. 2009;22:291–321. doi:10.1128/CMR.00051-08
58. Becher R, Weihmann F, Deising HB, Wirsel SG. Development of a novel multiplex DNA microarray for Fusarium graminearum and analysis of azole fungicide responses. BMC Genomics. 2011;12:52. doi:10.1186/1471-2164-12-52
59. Calvet HM, Yeaman MR, Filler SG. Reversible fluconazole resistance in Candida albicans: a potential in vitro model. Antimicrob Agents Chemother. 1997;41:535–539. doi:10.1128/AAC.41.3.535
60. Sionov E, Chang YC, Garraffo HM, Kwon-Chung KJ. Heteroresistance to fluconazole in Cryptococcus neoformans is intrinsic and associated with virulence. Antimicrob Agents Chemother. 2009;53:2804–2815. doi:10.1128/AAC.00295-09
61. Tobin MB, Peery RB, Skatrud PL. Genes encoding multiple drug resistance-like proteins in Aspergillus fumigatus and Aspergillus flavus. Gene. 1997;200:11–23. doi:10.1016/S0378-1119(97)00281-3
62. Mellado E, Garcia-Effron G, Alcázar-Fuoli L, et al. A new Aspergillus fumigatus resistance mechanism conferring in vitro cross-resistance to azole antifungals involves a combination of cyp51A alterations. Antimicrob Agents Chemother. 2007;51:1897–1904. doi:10.1128/AAC.01092-06
63. Diaz-Trujillo C, da Cunha C, Chong P, Etienne Duveiller and Pawan K. Singh et al. Azole resistance is related to overexpression of the CYP51 gene in Mycosphaerella fijiensis. In: Book of Abstracts EPS PhD Autumn School ‘host-Microbe Interactomics’. the Netherlands: Wageningen; 2011:26–27.
64. Denning DW, Venkateswarlu K, Oakley KL, Anderson MJ, Manning NJ, Stevens DA. Itraconazole resistance in Aspergillus fumigatus. Antimicrob Agents Chemother. 1997;41:1364–1368. doi:10.1128/AAC.41.6.1364
65. Snelders E, Rijs AJ, Kema GH, et al. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl Environ Microbiol. 2009;75:4053–4057. doi:10.1128/AEM.00231-09
66. Lucas JA, Hawkins NJ, Fraaije BA. The evolution of fungicide resistance. Adv Appl Microbiol. 2015;90:29–92.
67. Kretschmer M, Leroch M, Mosbach A, et al. Fungicide-driven evolution and molecular basis of multidrug resistance in field populations of the grey mould fungus Botrytis cinerea. PLoS Pathog. 2009;5(12):e1000696. doi:10.1371/journal.ppat.1000696
68. Mouches C, Pasteur N, Berge JB, et al. Amplification of an esterase gene is responsible for insecticide resistance in a Californian Culex mosquito. Science. 1986;233:778–780. doi:10.1126/science.3755546
69. Verweij PE, Snelders E, Kema GHJ, Mellado E, Melchers WGJ. Azole resistance in Aspergillus fumigatus: a side-effect of environmental fungicide use? Lancet Infect Dis. 2009;9:789–795. doi:10.1016/S1473-3099(09)70265-8
70. Al-Hatmi AM, van Diepeningen AD, Curfs-Breuker I, de Hoog GS, Meis JF. Specific antifungal susceptibility profiles of opportunists in the Fusarium fujikuroi complex. J Antimicrob Chemother. 2015;70:1068–1071. doi:10.1093/jac/dku505
71. Thomas PA. Current perspectives on ophthalmic mycoses. Clin Microbiol Rev. 2003;16:730–797. doi:10.1128/CMR.16.4.730-797.2003
72. Qiu S, Zhao GQ, Lin J, et al. Natamycin in the treatment of fungal keratitis: a systematic review and meta-analysis. Int J Ophthalmol. 2015;8(3):597–602. doi:10.3980/j.issn.2222-3959.2015.03.29
73. Oliveira Dos Santos C, Kolwijck E, van der Lee HA, et al. In vitro activity of chlorhexidine compared with seven antifungal agents against 98 Fusarium isolates recovered from fungal keratitis patients. Antimicrob Agents Chemother. 2019;63(8):e02669–18. doi:10.1128/AAC.02669-18
74. Kunt Z, Yağmur M, Kandemir H, et al. In vitro efficacy of chlorhexidine and a riboflavin/UVA combination on fungal agents of keratitis. Curr Eye Res. 2019;14:1–5. doi:10.1080/02713683.2019.1652916
75. Todokoro D, Suzuki T, Tamura T, et al. Efficacy of luliconazole against broad-range filamentous fungi including Fusarium solani species complex causing fungal keratitis. Cornea. 2019;38(2):238–242. doi:10.1097/ICO.0000000000001812
76. Al-Hatmi AM, Meletiadis J, Curfs-Breuker I, Bonifaz A, Meis JF, De Hoog GS. In vitro combinations of natamycin with voriconazole, itraconazole and micafungin against clinical Fusarium strains causing keratitis. J Antimicrob Chemother. 2016;71:953–955. doi:10.1093/jac/dkv421
77. Prajna NV, Krishnan T, Mascarenhas J, et al. The mycotic ulcer treatment trial: a randomized trial comparing natamycin vs. voriconazole. JAMA Ophthalmol. 2013;131:422–429. doi:10.1001/jamaophthalmol.2013.1497
78. Rees CA, Bao B, Zegans ME, Cramera RA. Natamycin and voriconazole exhibit synergistic interactions with nonantifungal ophthalmic agents against Fusarium species ocular isolates. Antimicrob Agents Chemother. 2019;63:e02505–18. doi:10.1128/AAC.02505-18
79. Varon AG, Nouer SA, Barreiros G, et al. Superficial skin lesions positive for Fusarium are associated with subsequent development of invasive fusariosis. J Infect. 2014;68:85–89. doi:10.1016/j.jinf.2013.08.011
80. Galletti J, Negri M, Grassi FL, Kioshima-Cotica ÉS, Svidzinski TI. Fusarium spp. is able to grow and invade healthy human nails as a single source of nutrients. Eur J Clin Microbiol Infect Dis. 2015;34:1767–1772. doi:10.1007/s10096-015-2410-1
81. Tupaki-Sreepurna A, Jishnu BT, Thanneru V, et al. An assessment of in vitro antifungal activities of efinaconazole and itraconazole against common non-dermatophyte fungi causing onychomycosis. J Fungi. 2017;3(2):E20. doi:10.3390/jof3020020
82. Sahni K, Singh S, Dogra S. Newer topical treatments in skin and nail dermatophyte infections. Indian Dermatol Online J. 2018;9(3):149–158. doi:10.4103/idoj.IDOJ_281_17
83. Abastabar M, Al-Hatmi AMS, Vafaei Moghaddam M, et al. Potent activity of luliconazole, lanoconazole and eight comparators against molecularly characterized Fusarium species. Antimicrob Agents Chemother. 2018;62(5):e00009–18. doi:10.1128/AAC.00009-18
84. Ortoneda M, Capilla J, Pastor FJ, Pujol I, Guarro J. Efficacy of liposomal amphotericin B in treatment of systemic murine fusariosis. Antimicrob Agents Chemother. 2002;46:2273–2275. doi:10.1128/AAC.46.7.2273-2275.2002
85. Falci DR, Pasqualotto AC. Profile of isavuconazole and its potential in the treatment of severe invasive fungal infections. Infect Drug Resist. 2013;6:163–174. doi:10.2147/IDR.S51340
86. Cornely OA, Ostrosky-Zeichner L, Rahav G, et al. Outcomes in patients with invasive mold disease caused by Fusarium or Scedosporium spp. treated with isavuconazole: experience from the VITAL and SECURE trials.
87. Marty FM, Cornely OA, Mullane KM, et al. Isavuconazole for treatment of invasive fungal diseases caused by more than one fungal species. Mycoses. 2018;61(7):485–497. doi:10.1111/myc.2018.61.issue-7
88. Nakamura I, Yoshimura S, Masaki T, et al. ASP2397: a novel antifungal agent produced by Acremonium persicinum MF-347833. J Antibiot (Tokyo). 2017;70(1):45–51. doi:10.1038/ja.2016.107
89. Koselny K, Green J, DiDone L, et al. The celecoxib derivative AR-12 has broad spectrum antifungal activity in vitro and improves the activity of fluconazole in a murine model of cryptococcosis. Antimicrob Agents Chemother. 2016;60:7115–7127. doi:10.1128/AAC.01061-16
90. Koselny K, Green J, Favazzo L, et al. Antitumor/antifungal celecoxib derivative AR-12 is a non-nucleoside inhibitor of the ANL-family adenylating enzyme acetyl CoA synthetase. ACS Infect Dis. 2016;2(4):268–280. doi:10.1021/acsinfecdis.5b00134
91. Oliver JD, Sibley GE, Beckmann N, et al. F901318 represents a novel class of antifungal drug that inhibits dihydroorotate dehydrogenase. Proc Natl Acad Sci U S A. 2016;113(45):12809–12814. doi:10.1073/pnas.1608304113
92. Perfect JR. The antifungal pipeline: a reality check. Nat Rev Drug Discov. 2017;16(9):603–616. doi:10.1038/nrd.2017.46
93. Jørgensen KM, Astvad KMT, Hare RK, Arendrup MC. EUCAST determination of olorofim (F901318) susceptibility of mold species, method validation, and MICs. Antimicrob Agents Chemother. 2018;62:e00487–18. doi:10.1128/AAC.00487-18
94. Miyazaki M, Horii T, Hata K, et al. In vitro activity of E1210, a novel antifungal, against clinically important yeasts and molds. Antimicrob Agents Chemother. 2011;55:4652–4658. doi:10.1128/AAC.00291-11
95. Castanheira M, Duncanson FP, Diekema DJ, Guarro J, Jones RN, Pfaller MA. Activities of E1210 and comparator agents tested by CLSI and EUCAST broth microdilution methods against Fusarium and Scedosporium species identified using molecular methods. Antimicrob Agents Chemother. 2012;56(1):352–357. doi:10.1128/AAC.05414-11
96. Pfaller MA, Messer SA, Motyl MR, Jones RN, Castanheira M. Activity of MK-3118, a new oral glucan synthase inhibitor tested against Candida spp. by two international methods (CLSI and EUCAST). J Antimicrob Chemother. 2013;68:858–863. doi:10.1093/jac/dks466
97. Lamoth F, Alexander BD. Antifungal activities of SCY-078 (MK-3118) and standard antifungal agents against clinical non-Aspergillus mold isolates. Antimicrob Agents Chemother. 2015;59:4308–4311. doi:10.1128/AAC.00234-15
98. Mitsuyama J, Nomura N, Hashimoto K, et al. In vitro and in vivo antifungal activities of T-2307, a novel arylamidine. Antimicrob Agents Chemother. 2008;52(4):1318–1324. doi:10.1128/AAC.01159-07
99. Shibata T, Takahashi T, Yamada E, et al. T-2307 causes collapse of mitochondrial membrane potential in yeast. Antimicrob Agents Chemother. 2012;56:5892–5897. doi:10.1128/AAC.05954-11
100. Pianalto KM, Alspaugh JA. New horizons in antifungal therapy. J Fungi. 2016;2(4):E26. doi:10.3390/jof2040026
101. Pfaller MA, Messer SA, Georgopapadakou N, Martell LA, Besterman JM, Diekema DJ. Activity of MGCD290, a Hos2 histone deacetylase inhibitor, in combination with azole antifungals against opportunistic fungal pathogens. J Clin Microbiol. 2009;47:3797–3804. doi:10.1128/JCM.00618-09
102. Pfaller MA, Rhomberg PR, Messer SA, Castanheira M. In vitro activity of a Hos2 deacetylase inhibitor, MGCD290, in combination with echinocandins against echinocandin-resistant Candida species. Diagn Microbiol Infect Dis. 2015;81:259–263. doi:10.1016/j.diagmicrobio.2014.11.008
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.