")
Back to Journals » Infection and Drug Resistance » Volume 13
Molecular Epidemiological Survey of Prophages in MRSA Isolates in Taiwan
Authors Lin LC , Ge MC , Liu TP, Lu JJ
Received 13 November 2019
Accepted for publication 5 February 2020
Published 24 February 2020 Volume 2020:13 Pages 635—641
DOI https://doi.org/10.2147/IDR.S238495
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Lee-Chung Lin,1 Mao-Cheng Ge,1 Tsui-Ping Liu,1 Jang-Jih Lu1–3
1Department of Laboratory Medicine, Chang Gung Memorial Hospital, Taoyuan, Taiwan; 2Department of Medical Biotechnology and Laboratory Science, College of Medicine, Chang Gung University, Taoyuan, Taiwan; 3Department of Medicine, College of Medicine, Chang Gung University, Taoyuan, Taiwan
Correspondence: Jang-Jih Lu
Department of Laboratory Medicine, Chang Gung Memorial Hospital, Linkou, No. 5, Fu-Shin Street, Kweishan, Taoyuan 333, Taiwan
Email [email protected]
Introduction: The prevalence of methicillin-resistant Staphylococcus aureus (MRSA) type SCCmec IV or V is increasing in Taiwan. It has been suggested that the surface protein SasX is responsible for their transmission. However, the sasX gene was not detected in our SCCmec IV or V isolates. Since sasX was originally found in S. epidermidis and believed to be transferred to S. aureus by a prophage, studies were conducted to detect and type this prophage in our clinical isolates.
Materials and Methods: A total of 1192 MRSA isolates collected from 2006 to 2014 were examined. Multiplex PCRs were performed to determine SCCmec, sasX, and prophage types.
Results: The prevalence of SCCmec IV and V isolates was increased in recent years (from 2006 to 2014). The sasX gene was present in most SCCmec III isolates but was absent in SCCmec IV or V isolates. The Sa5 prophage was found only in SCCmec IV and SCCmec V (or Vt) isolates, and the Sa6 prophage was mainly present in SCCmec III isolates. MRSA isolates harboring prophage combinations Sa1, Sa2, and Sa3; Sa2 and Sa3; Sa2, Sa3, and Sa7; or Sa2 and Sa7 were mainly of SCCmec II, and those that harbored prophage combinations Sa3 and Sa6; Sa3, Sa6, and Sa7; or Sa3 and Sa7 were mostly of SCCmec III. The numbers of SCCmec II isolates containing prophages Sa2, Sa3, and Sa7 and those of SCCmec III isolates containing prophages Sa3 and Sa6 or Sa3, Sa6, and Sa7 were decreased from 2010 to 2014. The number of SCCmec IV isolates with prophage Sa3 or prophages Sa3 and Sa5 was decreased, but that of those with prophage Sa6 or prophages Sa2 and Sa3 was increased from 2010 to 2014.
Conclusion: The sasX gene was found to play no role in clonal selection of MRSA. The finding that different SCCmec types of MRSA harbored different types of prophages suggests that these prophages may affect the survival and clonal expansion of certain types of MRSA.
Keywords: MRSA, SCCmec, sasX, prophage
Background
Staphylococcus aureus is a common opportunistic pathogen colonized on mucosal or skin surface. Methicillin-resistant Staphylococcus aureus (MRSA) strains containing the staphylococcal cassette chromosome mec (SCCmec) have become the main cause of nosocomial infections.1 SCCmec is carried by a mobile genetic element, and its mecA gene encodes the penicillin-binding protein 2a (PBP2a) responsible for the methicillin resistance phenotype.2 More than six types of SCCmec have been discovered.3 SCCmec type II and type III MRSA strains are mainly found in hospitals and are commonly referred to as hospital-acquired MRSA (HA-MRSA).4 SCCmec type IV and type V strains are mostly distributed in communities and are called CA-MRSA.5 In addition to SCCmec typing, multilocus sequence typing (MLST) is also commonly done to classify MRSA isolates. Previous epidemiological studies have demonstrated that some MLST types are closely related to SCCmec types. For example, ST239 isolates are mostly SCCmec III, and ST59 isolates are mainly SCCmec IV or V.6
In the past decades, most MRSA isolates belonged to SCCmec II or III, but the prevalence of SCCmec IV or V isolates was increased in recent years.7,8 It has been reported that ST239 isolates containing the SasX surface protein are prevalent in China.9 SasX is one of the LPXTG-motif containing surface proteins in Staphylococcus aureus, and the sasX gene has been shown to be transmitted by the prophage ΦSPβ from Staphylococcus epidermidis.10 SasX is considered a virulence factor as it can enhance MRSA colonization.9
It has been shown that most S. aureus lysogenic phages belong to the Siphoviridae family.11 Based on the sequences of the integrase gene, seven major prophage types Sa1 to Sa7 have been determined.11,12
In order to understand the increase in the prevalence of SCCmec IV and V MRSA, we conducted an epidemiological study to investigate the roles of the sasX gene and prophages in the transmission of MRSA isolates.
Materials and Methods
Bacterial Strains
A total of 1192 MRSA isolates from the Chang Gung Memorial Hospital (CGMH) were used. These isolates were collected from 2006 to 2014 and stored at −80°C. All isolates were identified by MALDI-TOF and tested for oxacillin susceptibility as an indication of the presence or absence of SCCmec and further characterized by SCCmec typing. Three hundred and three randomly selected MRSA isolates from 2006 to 2010 were examined for the presence of the sasX gene. All isolates were grown on tryptic soy agar (TSA) plates at 37°C for 16 hrs for the studies.
DNA Preparation and PCR Analysis of sasX
MRSA cells of overnight cultures were pelleted and then resuspended in the lysis buffer (50 mM glucose, 10 mM EDTA, 25 mM Tris-HCL, pH 8.0) containing lysozyme (50 mg/mL), lysostaphin (5 mg/mL), and 10% SDS. Genomic DNA was isolated using the QIAGEN™ DNeasy extraction kit and stored at −20°C until used. The sasX gene was detected by PCR as described previously.13,14
SCCmec and Prophage Typing
SCCmec typing was achieved by multiplex PCR using two sets of primers to identify mecA and eight sets of primers to differentiate the five ccr genes as described previously.15 Prophage typing was also done by multiplex PCR as described.11 All primers used are listed in Table 1. Student t-test was performed to determine the significant difference between data from two different years.
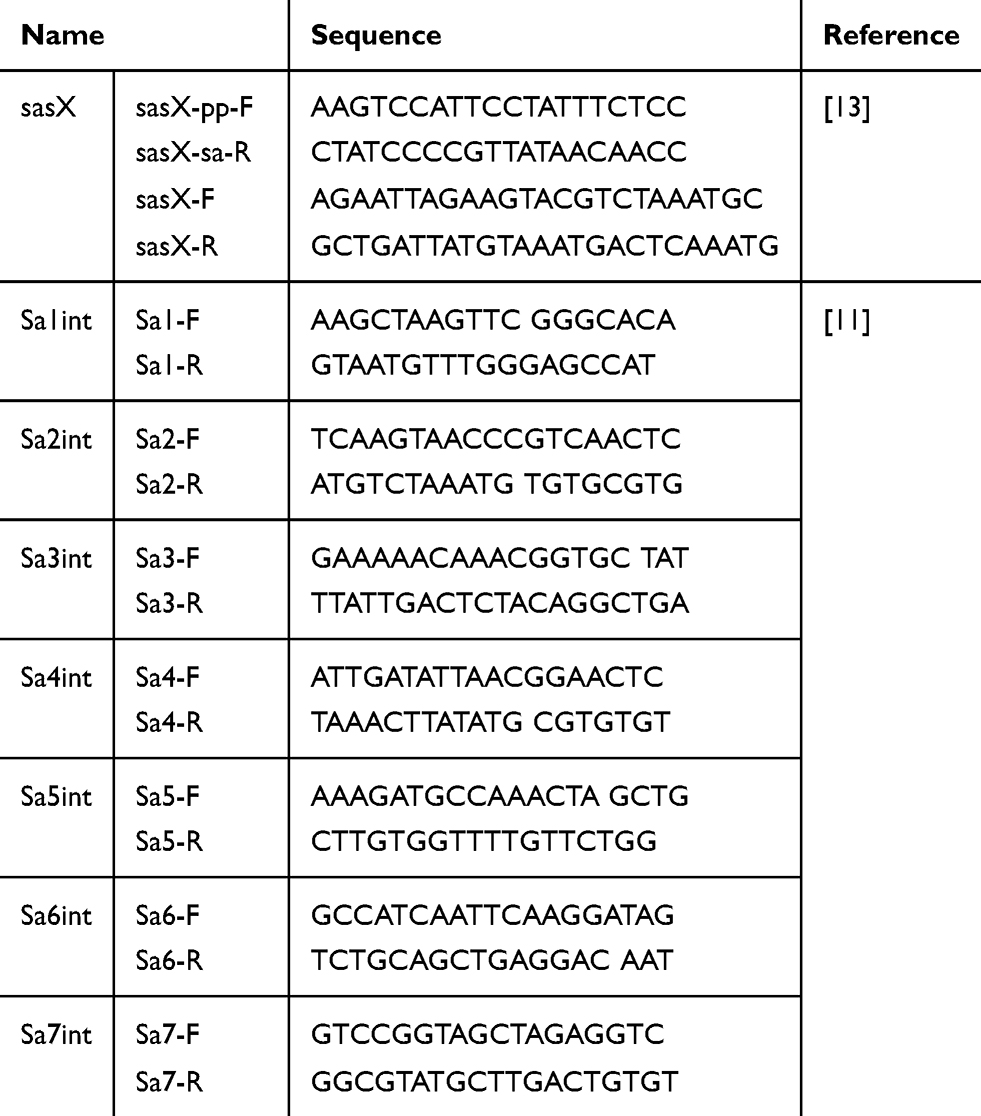 |
Table 1 Primers Used in This Study |
Results
Lack of sasX in SCCmec IV and V MRSA Isolates
To determine the prevalence of various types of MRSA in northern Taiwan, a total of 602 clinical isolates collected from 2006 to 2009 were SCCmec typed (Table 2). Results showed that the prevalence of SCCmec II MRSA was about the same from 2006 to 2009 (41% in 2006 and 42.1% in 2009), but that of SCCmec III MRSA was decreased from 44.6% in 2006 to 31.2% in 2009. However, the prevalence of SCCmec IV MRSA was increased from 8.6% in 2006 to 16.4% in 2009. The prevalence of SCCmec V MRSA was slightly increased, from 5% in 2006 to 7% in 2009. Since these 602 isolates were randomly selected from our collections from 2006 to 2009, there may be analytical bias. Therefore, a second study was performed on all isolates collected from 2010 (347 isolates) and 2014 (243 isolates). Results showed that the prevalence of SCCmec II MRSA was decreased from 14.4% in 2010 to 13.6% in 2014, and that of SCCmec III MRSA was decreased from 40.6% in 2010 to 33% in 2014. In contrast, the prevalence of SCCmec IV MRSA was increased from 22.5% in 2010 to 26.7% in 2014, and that of SCCmec V MRSA was increased from 15.3% to 22.2%. These results indicated that the prevalence of SCCmec II and III MRSA was decreasing and that of SCCmec IV and V MRSA was increasing from 2010 to 2014.
 |
Table 2 Number and Percentage of MRSA Isolates of Various SCCmec Types from 2006 to 2014 |
To determine whether the sasX gene plays any role in MRSA transmission, its presence in all MRSA isolates from 2006 to 2010 was determined (Table 3). Results showed that all SCCmec III and some SCCmec II isolates harbored sasX, but none of the SCCmec IV and SCCmec V isolates contained sasX.
 |
Table 3 The Prevalence of MRSA Isolates of Various SCCmec Types with the sasX Gene from 2006 to 2010 |
Types of Prophages in Various SCCmec Isolates
To understand the impact of prophages on the epidemiology of various types of MRSA, the prophages in all SCCmec isolates collected in 2010 (322 isolates) and 2014 (232 isolates) were typed by multiplex PCR of the integrase gene (Tables 4 and 5). Results showed that type Sa3 prophage was predominate and was present in isolates of all SCCmec groups. The Sa5 prophage was only found in SCCmec IV and SCCmec V (or Vt) isolates, and the Sa4 prophage was not present in any isolates (Table 4). Type Sa6 prophage was found in only 4–9.1% of SCCmec II and 1.9–5.6% of SCCmec V (or Vt) isolates but was present in 86.3–88.6% of SCCmec III isolates. Type Sa1 prophage was present in 1.4% of SCCmec III isolates, and type Sa7 was present in 1.5–2.6% of SCCmec IV and 5.7–9.3% SCCmec V (or Vt) isolates (Table 4).
 |
Table 4 Percentage of Various Types of SCCmec Isolates Harboring Prophages in 2010 and 2014 |
 |
Table 5 Prophage Types in Different SCCmec Isolates in 2010 and 2014 |
Some MRSA isolates were found to harbor multiple prophages (Table 5). The combination of prophages Sa1, Sa2, and Sa3; Sa2 and Sa3; Sa2, Sa3, and Sa7; or Sa2 and Sa7 was found only in SCCmec II isolates. Isolates that harbored prophages Sa3 and Sa6; Sa3, Sa6, and Sa7; or Sa3 and Sa7 were mainly SCCmec III. The number of SCCmec II isolates containing prophages Sa2, Sa3, and Sa7 was decreased from 15 in 2010 to 9 in 2014. The number of SCCmec III isolates with prophages Sa3 and Sa6 was decreased from 56 in 2010 to 22 in 2014, and that of those with prophages Sa3, Sa6, and Sa was decreased from 51 in 2010 to 25 in 2014. The number of SCCmec IV isolates with the Sa3 prophage was decreased from 25 in 2010 to 16 in 2014, and that of those with prophages Sa3 and Sa5 was decreased from 13 in 2010 to 7 in 2014. The number of SCCmec IV isolates with the Sa6 prophage was increased from 1 in 2010 to 9 in 2014, and that of those with prophages Sa2 and Sa3 was increased from 1 in 2010 to 11 in 2014. There were no significant changes in prophage types in SCCmec V isolates between 2010 and 2014.
Discussion
The change in the prevalence of various SCCmec types of MRSA has been reported.16–18 In this study, we found that both the prevalence (from 44.6% to 31.2%) and number (from 62 to 40) of SCCmec III isolates were decreased and those of SCCmec IV (prevalence from 8.6% to 16.4%, number from 13 to 21) and V (or Vt) (prevalence from 5% to 7%, number from 7 to 9) isolates were increased from 2006 to 2009. This trend is similar to that seen in other countries.19 Although the prevalence of SCCmec IV, V, or Vt isolates was increased from 2010 to 2014 (SCCmec IV, from 22.5% to 26.7%; SCCmec V or Vt, from 15.3% to 22.2%), there was a general decrease in the number of isolates of all SCCmec groups, except SCCmec V (or Vt) (53 in 2010 and 54 in 2014). The decrease in isolate number was most apparent for SCCmec III MRSA (from 141 in 2010 to 80 in 2014).
It has been speculated that the surface antigen gene sasX is responsible for the spreading of MRSA.13 However, we found that the sasX gene was not present in SCCmec IV and V isolates. This result suggests that the sasX gene plays no significant role in clonal selection or prevalence of certain types of MRSA in Taiwan.
Previous studies have shown that the sasX gene is transmitted from S. epidermidis to S. aureus by the ϕSPβ phage.10 This bacteriophage has been shown to carry genes encoding toxins and virulence factors.11,12 The integration of a bacteriophage into the genome of MRSA to become a prophage may affect its survival or virulence by affecting the expression of genes near the insertion sites.20,21 A significant finding was that 56 in 2010 and 22 in 2014 of SCCmec III isolates harbored prophages Sa3 and Sa6 and that 51 in 2010 and 25 in 2014 of SCCmec III isolates harbored prophages Sa3, Sa6, and Sa7. These results suggest an adverse effect of these prophages on the survival of SCCmec III isolates.
The mechanisms by which prophages affect the survival of MRSA isolates are unknown. It has been postulated that the bacterial immune system and phage integration affect MRSA clonal selection.11 The type I restriction-modification (R-M) system is a bacterial immune system, which may restrict the uptake of foreign DNA from other S. aureus strains.22,23 Since each MRSA clonal lineage has its own specific R-M system, it may allow intake of specific types of prophages. Our observation that prophage Sa5 existed only in SCCmec IV or V isolates and that Sa6 and Sa7 prophages were mainly present in SCCmec III isolates supports this hypothesis. It is possible that these prophages confer survival advantage to certain types of MRSA, e.g., SCCmec III, IV, and V.
The integration of phage genome into MRSA genome is another mechanism that may affect clonal selection.24–26 Prophage integration may mutate or affect the expression of host genes as evidenced by the observation that Sa3 and Sa6 prophage integration adversely affected the survival of S. aureus.20,21
The Sa3 prophage usually integrates into the hlb gene, which encodes the beta-toxin.20,21 The beta-toxin has been shown to play a role in colonization,27 biofilm formation,28,29 and virulence of MRSA.30 The inactivation of the hlb gene by Sa3 integration has been shown to weaken the biofilm formation ability.21,28,29 In conjunction with the delta-toxin, beta-toxin can enhance the pathogenicity of MRSA by making it resistant to degradation by phago‐endosomes.30 It has been demonstrated that the beta-toxin null strain SA113 failed in phagosomal escape, while complementation to recover beta-toxin production in SA113 enabled its escape from phagosomal degradation.30 The beta-toxin has also been shown to enhance the activities of sphingomyelinase and biofilm ligase, thus inhibiting IL-8 production by host cells.31 As IL-8 recruits neutrophils, the integration of Sa3 into the hlb gene would benefit the survival of MRSA.
The Sa6 prophage is usually inserted into the geh gene, which encodes a lipase. Inactivation of host lipase production would also benefit the survival of MRSA as lipase has been shown to degrade the antimicrobial fatty acid triglycerides produced by human skin.32 Mice immunized with human lipase have been shown to have an enhanced ability to defend S. aureus infection.33
Conclusions
Our data showed that the sasX surface antigen gene plays no role in clonal selection of a certain lineage of MRSA. The decrease in the number of SCCmec III isolates harboring both Sa3 and Sa6 prophages (from 56 in 2010 to 22 in 2014) and of those harboring Sa3, Sa6, and Sa7 prophages (from 51 in 2010 to 25 in 2014) suggests that the presence of these prophages make the isolates less advantageous in survival. The observation that prophage Sa5 existed only in SCCmec IV or V isolates and that the combination of Sa6 and Sa7 prophages was mainly found in SCCmec III isolates suggests that these prophages confer survival advantage to these types of MRSA. The mechanisms by which prophages affect the survival of MRSA isolates remain to be investigated. Results of this study would provide a guide for further epidemiological studies of the relationship between prophages and specific lineages of MRSA.
Ethics Approval and Consent to Participate
All MRSA isolates used in this study were obtained from Chang Gung Memorial Hospital bacterial bank. No ethics approval was required for the use of the isolates.
Data Sharing Statement
All data are included in this article.
Acknowledgments
We thank Dr. Chao-Hung Lee for assistance with the manuscript.
Funding
This work was supported by grants from the Chang Gung Memorial Hospital (CMRPG3F1721 and CMRPG3F1722) and the Ministry of Science and Technology, Taiwan (MOST 107-2320-B-182A-021-MY3 and MOST-107-2811-B-182A-002).
Disclosure
The authors declare no conflict of interest.
References
1. Jevons MP, Coe AW, Parker MT. Methicillin resistance in staphylococci. Lancet. 1963;1(7287):904–907. doi:10.1016/S0140-6736(63)91687-8
2. Bradley SF. Methicillin-resistant Staphylococcus aureus infection. Clin Geriatr Med. 1992;8(4):853–868. doi:10.1016/S0749-0690(18)30449-X
3. Katayama Y, Ito T, Hiramatsu K. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 2000;44(6):1549–1555. doi:10.1128/AAC.44.6.1549-1555.2000
4. Enright MC, Robinson DA, Randle G, Feil EJ, Grundmann H, Spratt BG. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc Natl Acad Sci U S A. 2002;99(11):7687–7692. doi:10.1073/pnas.122108599
5. Salgado CD, Farr BM, Calfee DP. Community-acquired methicillin-resistant Staphylococcus aureus: a meta-analysis of prevalence and risk factors. Clin Infect Dis. 2003;36(2):131–139. doi:10.1086/cid.2003.36.issue-2
6. Vandenesch F, Naimi T, Enright MC, et al. Community-acquired methicillin-resistant Staphylococcus aureus carrying Panton-Valentine leukocidin genes: worldwide emergence. Emerg Infect Dis. 2003;9(8):978–984. doi:10.3201/eid0908.030089
7. Deurenberg RH, Vink C, Kalenic S, Friedrich AW, Bruggeman CA, Stobberingh EE. The molecular evolution of methicillin-resistant Staphylococcus aureus. Clin Microbiol Infect. 2007;13(3):222–235. doi:10.1111/j.1469-0691.2006.01573.x
8. Huang YH, Tseng SP, Hu JM, Tsai JC, Hsueh PR, Teng LJ. Clonal spread of SCCmec type IV methicillin-resistant Staphylococcus aureus between community and hospital. Clin Microbiol Infect. 2007;13(7):717–724. doi:10.1111/j.1469-0691.2007.01718.x
9. Li M, Du X, Villaruz AE, et al. MRSA epidemic linked to a quickly spreading colonization and virulence determinant. Nat Med. 2012;18(5):816–819. doi:10.1038/nm.2692
10. Soderquist B, Andersson M, Nilsson M, et al. Staphylococcus epidermidis surface protein I (SesI): a marker of the invasive capacity of S. epidermidis? J Med Microbiol. 2009;58(Pt 10):1395–1397. doi:10.1099/jmm.0.008771-0
11. Goerke C, Pantucek R, Holtfreter S, et al. Diversity of prophages in dominant Staphylococcus aureus clonal lineages. J Bacteriol. 2009;191(11):3462–3468. doi:10.1128/JB.01804-08
12. Kahankova J, Pantucek R, Goerke C, Ruzickova V, Holochova P, Doskar J. Multilocus PCR typing strategy for differentiation of Staphylococcus aureus siphoviruses reflecting their modular genome structure. Environ Microbiol. 2010;12(9):2527–2538. doi:10.1111/j.1462-2920.2010.02226.x
13. Romling U, Balsalobre C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J Intern Med. 2012;272(6):541–561. doi:10.1111/joim.2012.272.issue-6
14. Kong H, Fang L, Jiang R, Tong J. Distribution of sasX, pvl, and qacA/B genes in epidemic methicillin-resistant Staphylococcus aureus strains isolated from East China. Infect Drug Resist. 2018;11:55–59. doi:10.2147/IDR.S153399
15. Turlej A, Hryniewicz W, Empel J. Staphylococcal cassette chromosome mec (Sccmec) classification and typing methods: an overview. Pol J Microbiol. 2011;60(2):95–103. doi:10.33073/pjm-
16. Chen CJ, Huang YC, Su LH, et al. Molecular epidemiology and antimicrobial resistance of methicillin-resistant Staphylococcus aureus bloodstream isolates in Taiwan, 2010. PLoS One. 2014;9(6):e101184. doi:10.1371/journal.pone.0101184
17. Chen CJ, Huang YC. New epidemiology of Staphylococcus aureus infection in Asia. Clin Microbiol Infect. 2014;20(7):605–623. doi:10.1111/1469-0691.12705
18. Chen CJ, Hsueh PR, Su LH, Chiu CH, Lin TY, Huang YC. Change in the molecular epidemiology of methicillin-resistant Staphylococcus aureus bloodstream infections in Taiwan. Diagn Microbiol Infect Dis. 2009;65(2):199–201. doi:10.1016/j.diagmicrobio.2009.05.020
19. David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev. 2010;23(3):616–687. doi:10.1128/CMR.00081-09
20. Lee CY, Iandolo JJ. Lysogenic conversion of staphylococcal lipase is caused by insertion of the bacteriophage L54a genome into the lipase structural gene. J Bacteriol. 1986;166(2):385–391. doi:10.1128/JB.166.2.385-391.1986
21. Carroll JD, Cafferkey MT, Coleman DC. Serotype F double- and triple-converting phage insertionally inactivate the Staphylococcus aureus beta-toxin determinant by a common molecular mechanism. FEMS Microbiol Lett. 1993;106(2):147–155. doi:10.1111/j.1574-6968.1993.tb05951.x
22. McCarthy AJ, Witney AA, Lindsay JA. Staphylococcus aureus temperate bacteriophage: carriage and horizontal gene transfer is lineage associated. Front Cell Infect Microbiol. 2012;2:6. doi:10.3389/fcimb.2012.00006
23. Waldron DE, Lindsay JA. Sau1: a novel lineage-specific type I restriction-modification system that blocks horizontal gene transfer into Staphylococcus aureus and between S. aureus isolates of different lineages. J Bacteriol. 2006;188(15):5578–5585. doi:10.1128/JB.00418-06
24. Brussow H, Canchaya C, Hardt WD. Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol Rev. 2004;68(3):560–602. doi:10.1128/MMBR.68.3.560-602.2004
25. Hargreaves KR, Kropinski AM, Clokie MR. Bacteriophage behavioral ecology: how phages alter their bacterial host’s habits. Bacteriophage. 2014;4:e29866. doi:10.4161/bact.29866
26. Davies EV, Winstanley C, Fothergill JL, James CE, Millard A. The role of temperate bacteriophages in bacterial infection. FEMS Microbiol Lett. 2016;363(5):fnw015. doi:10.1093/femsle/fnw015
27. Katayama Y, Baba T, Sekine M, Fukuda M, Hiramatsu K. Beta-hemolysin promotes skin colonization by Staphylococcus aureus. J Bacteriol. 2013;195(6):1194–1203. doi:10.1128/JB.01786-12
28. Huseby MJ, Kruse AC, Digre J, et al. Beta toxin catalyzes formation of nucleoprotein matrix in staphylococcal biofilms. Proc Natl Acad Sci U S A. 2010;107(32):14407–14412. doi:10.1073/pnas.0911032107
29. Sugimoto S, Iwamoto T, Takada K, et al. Staphylococcus epidermidis Esp degrades specific proteins associated with Staphylococcus aureus biofilm formation and host-pathogen interaction. J Bacteriol. 2013;195(8):1645–1655. doi:10.1128/JB.01672-12
30. Giese B, Glowinski F, Paprotka K, et al. Expression of delta-toxin by Staphylococcus aureus mediates escape from phago-endosomes of human epithelial and endothelial cells in the presence of beta-toxin. Cell Microbiol. 2011;13(2):316–329. doi:10.1111/j.1462-5822.2010.01538.x
31. Herrera A, Kulhankova K, Sonkar VK, et al. Staphylococcal beta-toxin modulates human aortic endothelial cell and platelet function through sphingomyelinase and biofilm ligase activities. MBio. 2017;8(2):e00273–e00217. doi:10.1128/mBio.00273-17
32. Cadieux B, Vijayakumaran V, Bernards MA, McGavin MJ, Heinrichs DE. Role of lipase from community-associated methicillin-resistant Staphylococcus aureus strain USA300 in hydrolyzing triglycerides into growth-inhibitory free fatty acids. J Bacteriol. 2014;196(23):4044–4056. doi:10.1128/JB.02044-14
33. Hu C, Xiong N, Zhang Y, Rayner S, Chen S. Functional characterization of lipase in the pathogenesis of Staphylococcus aureus. Biochem Biophys Res Commun. 2012;419(4):617–620. doi:10.1016/j.bbrc.2012.02.057
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.