Back to Journals » Infection and Drug Resistance » Volume 12
Molecular characterization, serotypes and phenotypic and genotypic evaluation of antibiotic resistance of the Klebsiella pneumoniae strains isolated from different types of hospital-acquired infections
Authors Ranjbar R ![]() , Kelishadrokhi AF, Chehelgerdi M
, Kelishadrokhi AF, Chehelgerdi M ![]()
Received 27 December 2018
Accepted for publication 19 February 2019
Published 20 March 2019 Volume 2019:12 Pages 603—611
DOI https://doi.org/10.2147/IDR.S199639
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Reza Ranjbar,1 Ali Fatahian Kelishadrokhi,2 Mohammad Chehelgerdi2
1Molecular Biology Research Center, Systems Biology and Poisonings Institute, Baqiyatallah University of Medical Sciences, Tehran, Iran; 2Biotechnology Research Center, Shahr-e Kord Branch, Islamic Azad University, Shahr-e Kord, Iran
Purpose: Virulent and resistant Klebsiella pneumoniae strains are considered as one of the most significant causes of hospital-acquired infections. The present investigation was done to study the distribution of virulence factors, capsule serotypes and phenotypic and genotypic evaluation of antibiotic resistance of the K. pneumoniae strains isolated from hospital-acquired infections.
Patients Materials and methods: Two hundred and sixty different types of hospital-acquired infections were collected and cultured. Antibiotic resistance pattern of K. pneumoniae isolates and their molecular characterization were studied using disk diffusion and PCR, respectively.
Results: One hundred and fifty out of 260 (44.22%) hospital-acquired infections harbored K. pneumoniae. Urine samples (63.75%) had the highest prevalence of K. pneumoniae, while wound (33.33%) had the lowest. K. pneumoniae strains harbored the highest prevalence of resistance against ampicillin (100%), cefuroxime (100%), amoxicillin/clavulanic acid (95.65%) and ceftazidime (95.52%). FimH-1 (93.04%), traT (92.17%), mrkD (84.34%), and entB (80.86%) were the most commonly detected virulence genes. AcrAB (96.52%) and tolC (85.21%) were the most commonly detected antibiotic resistance genes. Prevalence of ompK35 and ompK36 virulence genes were 75.65% and 79.13%, respectively. Prevalence of K1 and K2-positive serotypes were 27.82% and 6.96%, respectively.
Conclusions: High prevalence of resistance against several types of antibiotics and simultaneous presence of some virulence factors and multi-drug resistance genes pose an important public health issue.
Keywords: Klebsiella pneumoniae, antibiotic resistance pattern, antibiotic resistance genes, capsule serotypes, virulence genes, hospital infections
Introduction
Hospital-acquired infections including urinary tract infections (UTIs), respiratory, skin and soft tissue, blood, gastrointestinal and finally reproductive infections are considered as the most important causes of hospitalization. Hospital-acquired infections are estimated for around 10–60% of hospitalizations and more than 800,000 deaths annually in the USA.1,2 The presence of multi-drug resistant pathogenic bacteria has increased the impact of hospital-acquired infections.1,2
Klebsiella pneumoniae is a Gram-negative, non-motile, encapsulated, lactose-fermenting, facultative anaerobic, and rod shape bacterium belonging to the Enterobacteriaceae family.3,4 K. pneumoniae is also an important pathogen in nosocomial infections such as pneumonia, bacteremia, UTIs, pyogenic liver abscesses, wounds and burns infections around the world.3 Furthermore, K. pneumoniae infections are difficult to treat because of the presence of certain virulence factors and also the emergence of antibiotic resistance.3–6
Pathogenicity of K. pneumoniae is due to the presence of certain virulence genes which encode specific virulence factors that allow the bacterium to attack the immune system and cause many kinds of diseases.3,5,6 A set of virulence factors contributing to virulence and pathogenicity in K. pneumoniae strains, including the capsular serotype (particularly capsule serotypes K1 and K2), lipopolysaccharide, iron-scavenging systems, and adhesions , have been introduced in the K. pneumoniae strains isolated from hospital-acquired infections. Iron acquisition systems are essential for the growth of pathogenic bacteria. Furthermore, the iron chelator siderophore allows bacteria to take up protein-bound iron from the host cells.6,7 Virulence-associated genes include those encoding regulators of mucoid phenotype A (rmpA), type 1 and type 3 adhesins (fimH-1, mrkD), aerobactin (iron siderophore) synthase (iucC), bacteriocin biosynthesis [enterobactin (entB), and yersiniabactin (irP-1)], and serum resistance-associated outer membrane lipoprotein (traT) have a predominant role in pathogenicity of K. pneumoniae strains isolated from hospital-acquired infections.6,8 The above-mentioned virulence factors have an essential role in adhesion and invasion of K. pneumoniae strains to host tissues.
K. pneumoniae strains isolated from hospital-acquired infections have emerging antibiotic resistance. Epidemiological investigations revealed that K. pneumoniae strains isolated from hospital-acquired infections harbored a high prevalence of resistance against commonly used groups of antibiotics, especially penicillins, aminoglycosides, tetracyclines, macrolides, lincosamides, folate inhibitors, fluoroquinolones, and phenicols.9,10 Gram-negative bacteria like K. pneumoniae have developed several mechanisms of resistance against commonly used antimicrobials. The presence of certain antibiotic resistance genes is one of the most important causes of the occurrence of antibiotic resistance. Genes coding for the multi-drug efflux pump system AcrAB-TolC and MdtK and porin coding genes (OmpK35 and OmpK36) have high clinical importance in the occurrence of antibiotic resistance in K. pneumoniae strains.9,10 These genes predominantly cause the occurrence of resistance against several kinds of antibiotic groups. Thus, it is important to detect antibiotic resistance genes in the K. pneumoniae strains isolated from hospital-acquired infections.
Data on the prevalence, antibiotic resistance properties and distribution of virulence genes of K. pneumoniae strains are scarce in Iran. Therefore, the current research was done to assess the prevalence rate, distribution of virulence genes and phenotypic and genotypic evolution of antibiotic resistance of K. pneumoniae strains isolated from different types of hospital-acquired infections.
Materials and methods
Samples
From April to October 2017, a total of 260 hospital-acquired infections including urine (n=80), wound (n=45), blood (n=64) and sputum (n=71) samples were randomly collected from hospitalized patients. All specimens were collected from hospitalized patients of Shohada Lenjan, Imam Khomeini Falavarjan, Sadooghi and Al-Zahra hospitals of Isfahan province, Iran. Midstream urine was collected in a sterile condition to decrease potential bacterial, cellular and artifactual contamination. Samples were immediately transferred to the laboratory in coolers with ice-packs.
Bacterial isolation
All samples were immediately streaked onto MacConkey Agar (MCA, Merck, Germany) and incubated at 37°C for 24 h. Bacterial colonies were streaked again on Brilliant Green Agar (BGA, Merck) and xylose lysine deoxycholate agar (XLD, Merck) and incubated at 37°C for 24 h. Colonies were streaked again on HiCrome UTI agar (HiMedia Laboratories, Mumbai, India). All isolates were subjected to standard confirmatory tests, which included oxidase and catalase, growth on SIM (sulfide, indole, motility), Simon citrate, MR-VP (methyl red – Voges Proskauer), lysine iron agar, Kligler agar, phenylalanine agar, urea agar, malonate, blood agar, and MacConkey agar. K. pneumoniae isolates were stored in tryptic soy broth (TSB, Merck) containing 20% glycerol at −70°C for further characterization.
K. pneumoniae strains were sub-cultured overnight in Luria-Bertani broth (Merck) and genomic DNA was extracted from typical colonies of K. pneumoniae using DNA extraction kit (Fermentas, Leon-Rot, Germany) according to the manufacturer’s instruction. All of the positive colonies were confirmed another time using PCR-based amplification of the 16SrRNA gene. PCR was performed according to the method described previously.11
Antibiotic susceptibility pattern
Antibiotic susceptibility pattern of K. pneumoniae isolates was performed according to the Kirby–Bauer disk diffusion method.12 Mueller–Hinton agar (Merck) medium was used for this purpose. Principles of the Clinical and Laboratory Standards Institute (CLSI) were used for this purpose.12 Antibiotic resistance pattern of the K. pneumoniae strains was assessed against 15 commonly used antibiotic agents including amoxicillin/clavulanic acid (20/10 μg), ampicillin (10 μg), chloramphenicol (30 μg), gentamicin (10 μg), piperacillin/tazobactam (110/10 μg), ceftazidime (30 μg), cefuroxime (30 μg), aztreonam (30 μg), imipenem (10 μg), tobramycin (10 μg), amikacin (30 μg), tetracycline (30 μg), ciprofloxacin (5 μg), nalidixic acid (30 μg), and co-trimoxazole (30 μg) (Thermo Fisher Scientific Oxoid Ltd, Basingstoke, UK). All of the inoculated plates were aerobically incubated at 37°C for 18–24 h in an aerobic atmosphere. Results were interpreted based on the instruction provided by the CLSI (2014).12 K. pneumoniae ATCC 4352 was used as a quality control organism.
PCR amplification of antibiotic resistance genes and virulence markers
The frequency of rmpA, fimH-1, mrkD, traT, entB, Irp-1, and IucC virulence genes, OmpK35, OmpK36, mdtK, tolC and AcrAB antibiotic resistance genes and K1 and K2 capsule serotypes were assessed using PCR.13 A set of primers introduced in a previous study were used for this purpose.13 A programmable DNA thermo-cycler (Eppendorf Mastercycler 5330, Eppendorf-Nethel-Hinz GmbH, Hamburg, Germany) was used in all PCR reactions. PCR grade water and K. pneumoniae standard strains (ATCC 18883, ATCC 43816, ATCC 4352 and ATCC 10031) were used as negative and positive controls, respectively. Ten microliters of PCR product were exposed to electrophoresis in a 2% agarose gel in 1× TBE buffer at 80 V for 30 min, stained with SYBR Green. The UVI doc gel documentation systems (Grade GB004, Jencons PLC, London, UK) was applied for analysis of images.
Statistical analysis
Statistics were subjected to Microsoft Office Excel (version 15; Microsoft Corp., Redmond, WA, USA). Statistical analysis was performed by means of the SPSS 21.0 statistical software (IBM Corporation, Armonk, NY, USA). The chi-square test and Fisher’s exact two-tailed test were applied to measure any significant relationship. P-value <0.05 was considered as the statistically significant level.
Ethics statement
The study was approved by the Ethical Council of Research of the Molecular Biology Research Center, Systems Biology and Poisonings Institute, Baqiyatallah University of Medical Sciences, Tehran, Iran. (Consent Ref Number BMSU-2017). Verification of this research project and the licenses related to the sampling process were approved by the Prof Reza Ranjbar (Approval Ref Number 2017/45). All clinical samples were taken from volunteer hospitalized patients. Written informed consent was also obtained from all of the study patients or their parents.
Results
Table 1 characterizes the prevalence of K. pneumoniae strains in different types of hospital-acquired infections. One hundred and fifteen of 260 (44.22%) hospital-acquired infections harbored K. pneumoniae. All positive strains were also approved using the 16SrRNA-based PCR amplification. Urine samples (63.75%) had the highest prevalence of K. pneumoniae, while wound (33.33%) had the lowest. Statistically significant difference was seen between types of samples and prevalence of K. pneumoniae (P<0.05).
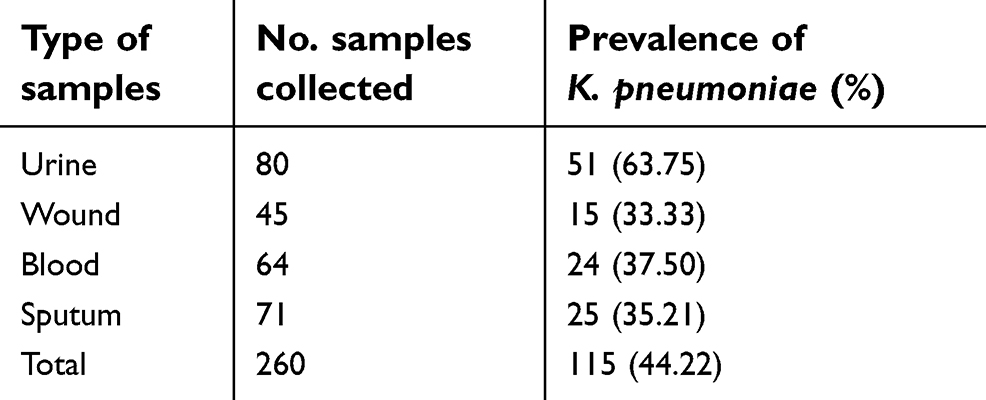 | Table 1 Prevalence of K. pneumoniae strains isolated from different types of hospital-acquired infections |
Table 2 characterizes the antibiotic resistance pattern of K. pneumoniae strains isolated from different types of hospital-acquired infections. K. pneumoniae strains harbored the highest prevalence of resistance against ampicillin (100%), cefuroxime (100%), amoxicillin/clavulanic acid (95.65%), ceftazidime (95.52%), piperacillin/tazobactam (84.34%), tobramycin (80.80%), ciprofloxacin (80%), nalidixic acid (80%), co-trimoxazole (77.39%) and aztreonam (74.78%) antibiotic agents. K. pneumoniae strains harbored the lowest prevalence of resistance against chloramphenicol (43.47%), tetracycline (53.91%) and amikacin (56.52%). Statistically significant difference was seen between types of samples and the prevalence of antibiotic resistance (P<0.05).
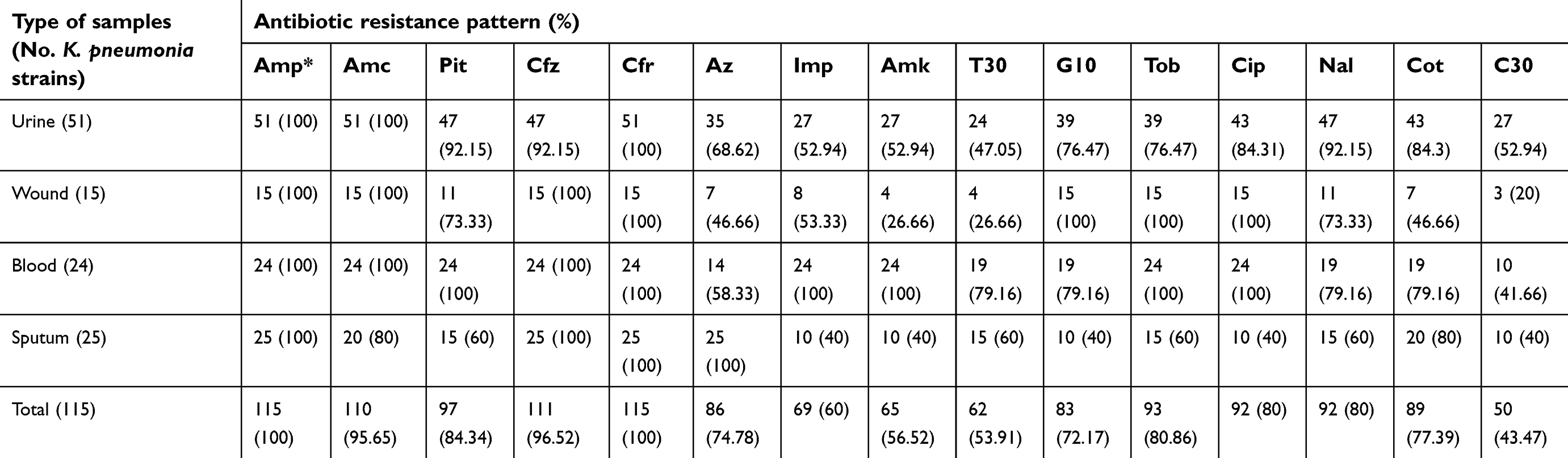 | Table 2 Antibiotic resistance pattern of K. pneumoniae strains isolated from different types of hospital-acquired infections |
Table 3 characterizes the distribution of virulence genes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections. FimH-1 (93.04%), traT (92.17%), mrkD (84.34%), and entB (80.86%) were the most commonly detected virulence genes amongst the K. pneumoniae strains. Prevalence of rmpA (13.91%) and jrp1 (31.30%) was lower than other detected virulence genes. Statistically significant difference was seen between types of samples and the prevalence of virulence genes (P<0.05).
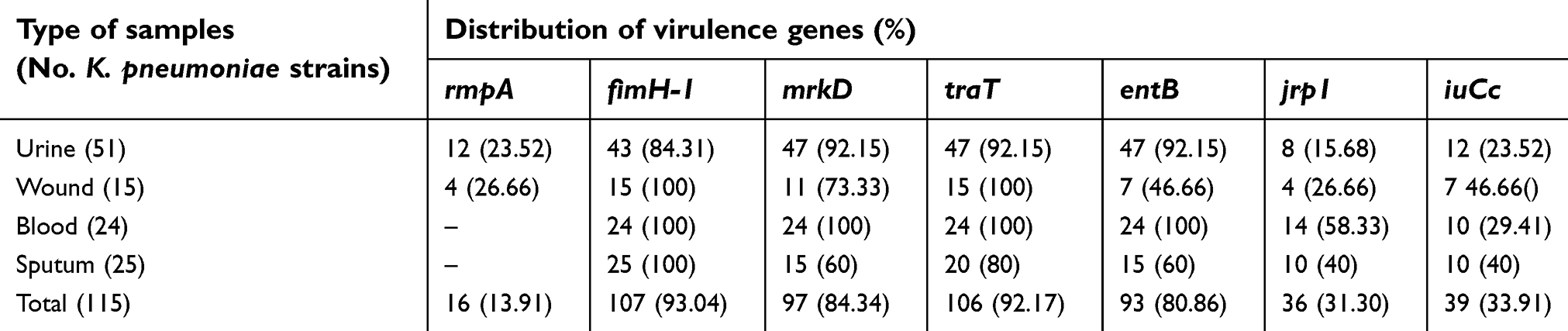 | Table 3 Distribution of virulence genes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections |
Table 4 characterizes the distribution of antibiotic resistance genes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections. AcrAB (96.52%) and tolC (85.21%) were the most commonly detected antibiotic resistance genes of the K. pneumoniae strains isolated from different types of hospital-acquired infections. MdtK (29.54%) had the lowest prevalence amongst all detected antibiotic resistance genes. Prevalence of ompK35 and ompK36 virulence genes were 75.65% and 79.13%, respectively. Statistically significant difference was seen between types of samples and the prevalence of antibiotic resistance genes (P<0.05).
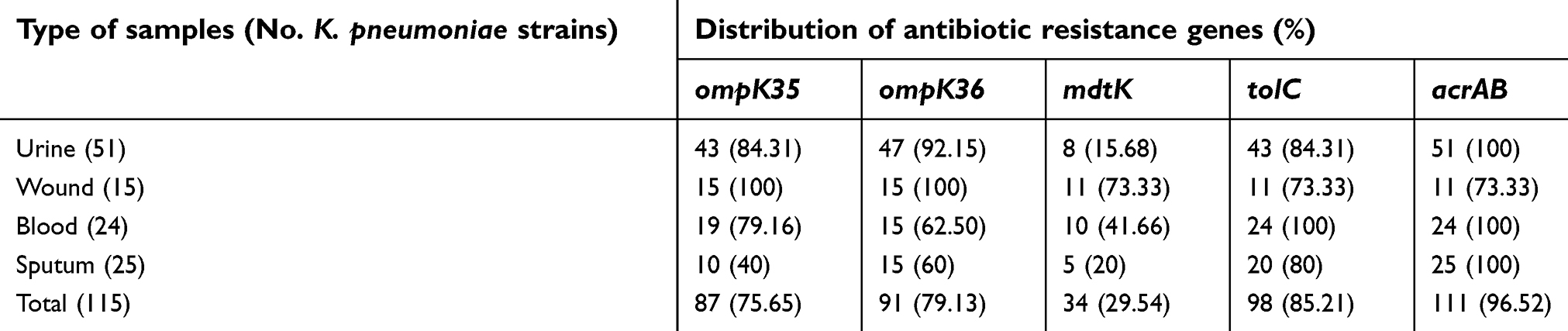 | Table 4 Distribution of antibiotic resistance genes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections |
Table 5 characterizes the distribution of capsule serotypes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections. Prevalence of K1 and K2-positive serotypes of the K. pneumoniae strains isolated from different types of hospital-acquired infections were 27.82% and 6.96%, respectively. Serotypes of 65.21% of the studied K. pneumoniae strains were not related to K1 and K2 serotypes.
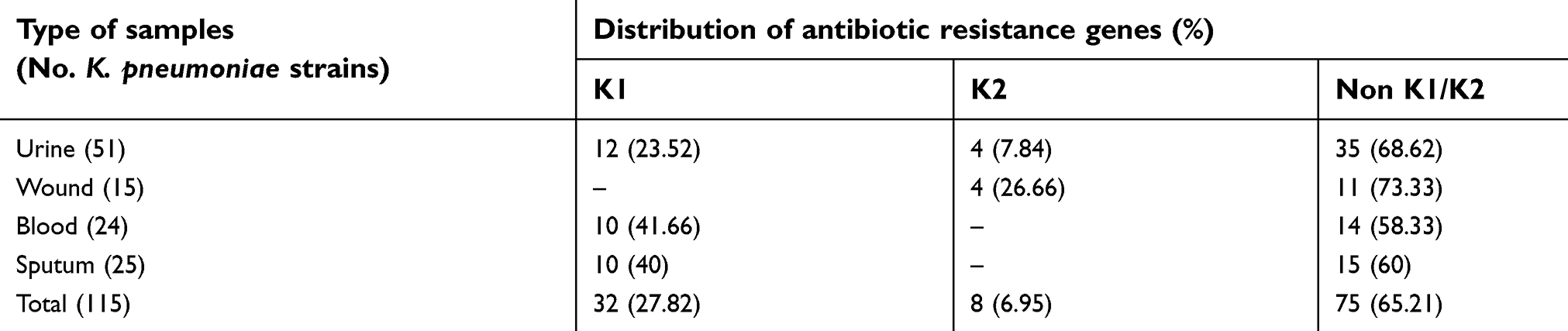 | Table 5 Distribution of capsule serotypes amongst the K. pneumoniae strains isolated from different types of hospital-acquired infections |
Discussion
K. pneumoniae is an important cause of multidrug-resistant infections worldwide.13,14 Epidemiological studies have highlighted the emergence of multidrug-resistant and virulent K. pneumoniae strains isolated from hospital-acquired infections.13,14 Antibiotic resistant and virulent bacteria caused more severe diseases for longer periods of time.
The present research was done to assess the prevalence, distribution of virulence genes and genotypic and phenotypic analyses of antibiotic resistance amongst the K. pneumoniae strains isolated from hospital-acquired infections. Prevalence of K. pneumoniae in different types of human hospital infections was 44.22%. Furthermore, the prevalence of K. pneumoniae strains in urine, wound, blood, and sputum samples were 63.75%, 33.33%, 37.50%, and 35.21%, respectively. Lev et al15 reported that the total prevalence of K. pneumoniae in hospital-acquired infections in hospitals of Moscow was 16.66%. They characterized that prevalence of K. pneumoniae in respiratory, urine, wound, cerebrospinal fluid, blood, and rectal swab specimens were 57%, 30%, 5%, 4%, 3%, and 1%, respectively. Akter et al16 reported that prevalence of K. pneumoniae strains in hospital-acquired infections of Bangladesh hospitals was 19.72%. They characterized that the most prevalent infection caused by K. pneumoniae isolates was UTIs (27–36%), followed by swab (8–10%) or pus (3–6%). Similar results were found earlier by Riaz et al (Pakistan),17 Lina et al (Bangladesh),18 Sarathbabu et al (India),19 and Heidary et al (Iran).20 Moreover, higher prevalence of K. pneumoniae strains in UTIs has been reported from North Africa,21 Nepal,22 USA,23 and Taiwan.24
We found that K. pneumoniae strains harbored the highest prevalence of resistance against ampicillin, cefuroxime, amoxicillin/clavulanic acid, ceftazidime, piperacillin/tazobactam, tobramycin, ciprofloxacin, nalidixic acid, co-trimoxazole, and aztreonam antibiotic agents. High prevalence of antibiotic-resistant K. pneumoniae strains was also reported from the cases of hospital-acquired infections in Canada,25 Nepal,25 Australia,26 China27, and Pakistan.28 Higher fatality rate was also reported for infections caused by antibiotic-resistant K. pneumoniae.29 Prevalence of antibiotic-resistant K. pneumoniae strains in Asian and Latin American countries had a range between 40% and 55%.30,31 In a study conducted in Saudi Arabia,32 K. pneumoniae strains isolated from hospital-acquired infections exhibited high prevalence of resistance against ceftriaxone (58.93%), meropenem (5.60%), gentamicin (11.10%), piperacillin (11.10%), amikacin (6.70%), and ciprofloxacin (6.70%) antibiotic agents, which was similar to our findings. Mekki et al33 reported the higher prevalence of resistance of K. pneumoniae strains against cotrimoxazole (100%), gentamicin (100%), nalidixic acid (100%), cefuroxime (100%), ciprofloxacin (97.37%), nitrofurantoin (97.37%), and amikacin (39.47%). Seibert et al34 reported that K. pneumoniae strains isolated from hospital infections harbored the high prevalence of resistance against amikacin (8.50%), gentamicin (42.60%), ceftriaxone (66%), cefepime (55.60%), imipenem (80%), and meropenem (83%).
K. pneumoniae strains isolated from the clinical samples of the present study harbored high prevalence of fimH-1, traT, mrkD, entB, rmpA, and jrp1 virulence genes. This phenomenon characterizes the high pathogenicity of K. pneumoniae strains. These virulence genes were also predominant in the K. pneumoniae strains isolated from hospital-acquired infections in China,35 UK,36 France,37 and Brazil.38 Type 1 fimbriae is the most frequent adhesive factor in K. pneumoniae strains. Presence of this gene can lead to UTIs. Type 1 fimbrial adhesion can also arbitrate the binding of K. pneumoniae strains to tissue cells of the urinary and respiratory tracts. MrkD protein is a vital factor in binding bacterial pathogens to the collagen molecules of the mammalian cells.13 The majority of K. pneumoniae clinical isolates usually express type 1 fimbrial adhesins.13 In the current investigation, the two genes coding for these adhesive structures (fimH-1 and MrkD) were detected in all types of infections. The plasmidic traT gene encodes an outer membrane protein involved in bacterial conjugation and blocks the complement-mediated cascade, and act as an invasion.13 W noticed the traT gene in 92.17% of K. pneumoniae isolates. The incidence of traT gene in our isolates was relatively high as it was frequently associated with the K1 capsule serotype.13 The majorityf of Enterobacteriaceae strains contain genes encoding iron uptake systems, including aerobactin or enterochelin. These siderophores have double roles as they can also prevent T cell proliferation in addition to their role in enhancing iron uptake.39–41 The iron siderophores aerobactin synthase gene (IucC), enterobactin biosynthesis gene (entB), and yersinibactin biosynthesis gene (Irp-1) were detected in 33.91%, 31.30%, and 80.86% of the K. pneumoniae isolates, respectively. Highly pathogenic Yersinia strains have high-pathogenicity island (HPI) that comprise the gene Irp-1. This HPI is also predominant in Klebsiella and other enterobacteria, including K. pneumoniae, K. oxytoca, E. coli, Citrobacterspecies, and Enterobacter species.39–41
The capsular serotypes K1 and K2 are associated with the main virulent strains of K. pneumoniae.42 Feizabadi et al have shown that K1 and K2 serotypes were detected in 11.2% and 14.6% of K. pneumoniae strains isolated from hospital-acquired infections, respectively.43 K1 and K2 serotypes were detected in 27.82% and 6.96% of the K. pneumoniae isolates of our study, respectively. Higher distribution of the K1 capsular serotype was also reported from K. pneumoniae strains isolated from hospital-acquired infections in Japan44 and Australia.45
The final section of our research was conducted on the distribution of some of the most important antibiotic resistance genes of the K. pneumoniae strains. AcrAB, tolC, MdtK, ompK35, and ompK36 were predominant amongst the K. pneumoniae strains isolated from hospital-acquired infections. Antibiotic efflux pumps act as one of the most important mechanisms of antimicrobial resistance in K. pneumoniae.46–48 The increased efflux of the antimicrobial agent leads to the reduction of its intracellular concentration, which can increase bacterial survival. The AcrAB efflux pump was more frequent than mdtK. The presence of the efflux pump system (AcrAB-TolC) was significantly associated with the antibiotic resistance pattern.46–48 Furthermore, most of the K. pneumoniae strains were missing either the AcrAB efflux pump or the tolC outer membrane protein or both. It has been reported that loss of ompK35 and ompK36 led to an increase in carbapenem, ciprofloxacin, and chloramphenicol resistance.46–48 Similarly, in our study, resistant K. pneumoniae strains harbored high distribution of ompK35 and ompK36 resistance genes. Both OmpK35 and OmpK36 also play a role in K. pneumoniae virulence and infection.46–48 Deletion of OmpK36 and OmpK35 can lead to the reduction in virulence of highly virulent strains and can increase their susceptibility to neutrophil phagocytosis. In our investigation, both Ompk35 and OmpK36 genes were simultaneously detected in the majority of K. pneumoniae isolates and especially those of urine and wound samples. Their distributions were variable though in sputum and blood samples. A direct correlation between efflux pumps and virulence of pathogenic bacteria was reported by Padilla et al (2010).49 Several genes essential for intracellular invasion and survival were downregulated in mutant strains lacking acrAB-tolC efflux pumps.46–49
Conclusion
We identified a high prevalence of K. pneumoniae strains with a considerable distribution of putative virulence factors, antibiotic resistance genes, and capsular serotypes, and a high prevalence of antibiotic resistance of the K. pneumoniae strains isolated from different hospital-acquired infections. K. pneumoniae strains exhibit high prevalence of resistance against ampicillin, cefuroxime, amoxicillin/clavulanic acid, ceftazidime, piperacillin/tazobactam, tobramycin, ciprofloxacin, nalidixic acid, co-trimoxazole, and aztreonam antibiotic agents, high distribution of fimH-1 (93.04%), traT (92.17%), mrkD (84.34%), and entB (80.86%) virulence genes, considerable prevalence of acrAB, tolC, mdtK, ompK35 and ompK36 antibiotic resistance genes, and finally significant incidence of K1 capsular serotype. High prevalence of resistance against several types of antibiotic agents and the simultaneous presence of some virulence factors and resistance genes pose an important public health issue regarding the worse condition of hospital infections. However, further studies are required to find other epidemiological aspects of the K. pneumoniae strains and a correlation between virulence genes and antibiotic resistance properties of the K. pneumoniae strains.
Abbreviations
Amp, ampicillin; AMX, amoxicillin; rmpA, regulators of mucoid phenotype A; IucC, aerobactin (iron siderophore) synthase; entB, enterobactin; irP-1, yersiniabactin; traT, serum resistance-associated outer membrane lipoprotein; CLR, clarithromycin; CLSI, Clinical Laboratory Standards Institute.
Data sharing statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We would like to thank the Clinical Research Development Center of Baqiyatallah Hospital for their kind cooperation. The authors also gratefully acknowledge all the staff of Biotechnology Research Center, Shahr-e Kord Branch, Islamic Azad University, Shahr-e Kord in southwest Iran for their sincere support.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Reed D, Kemmerly SA. Infection control and prevention: a review of hospital-acquired infections and the economic implications. Ochsner J. Epub ahead of print 2009. doi:10.1043/1524-5012-9.1.27
2. Lobdell KW, Stamou S, Sanchez JA. Hospital-acquired infections. Surg Clin North Am. 2012;92(1):65–77. doi:10.1016/j.suc.2011.11.003
3. Lin WH, Wang MC, Tseng CC. et al. Clinical and microbiological characteristics of Klebsiella pneumoniae isolates causing community-acquired urinary tract infections. Infection. Epub ahead of print 2010. doi:10.1007/s15010-010-0049-5
4. Ranjbar R, Memariani H, Sorouri R. et al. Distribution of virulence genes and genotyping of CTX-M-15-producing Klebsiella pneumoniae isolated from patients with community-acquired urinary tract infection (CA-UTI). Microb Pathog. Epub ahead of print 2016. doi:10.1016/j.micpath.2016.10.002
5. Cristina ML, Sartini M, Ottria G, et al. Epidemiology and biomolecular characterization of carbapenem-resistant Klebsiella pneumoniae in an Italian hospital. J Prev Med Hyg. 2016;57(3):E149.
6. Highsmith AK, Jarvis WR. Klebsiella pneumoniae: selected virulence factors that contribute to pathogenicity. Infect Control. Epub ahead of print 1985. doi:10.1017/S0195941700062640
7. Lin CT, Wu CC, Chen YS, et al. Fur regulation of the capsular polysaccharide biosynthesis and iron-acquisition systems in Klebsiella pneumoniae CG43. Microbiology. 2011;157(2):419–429. doi:10.1099/mic.0.044065-0
8. Lawlor MS, O’Connor C, Miller VL. Yersiniabactin is a virulence factor for Klebsiella pneumoniae during pulmonary infection. Infect Immun. Epub ahead of print 2007. doi:10.1128/IAI.00372-06
9. Shi W, Li K, Ji Y. et al. Carbapenem and cefoxitin resistance of Klebsiella pneumoniae strains associated with porin ompK36 loss and DHA-1 β-lactamase production. Brazilian J Microbiol. Epub ahead of print 2013. doi:10.1590/S1517-83822013000200015
10. Sun J, Deng Z, Yan A. Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochem Biophys Res Commun. Epub ahead of print 2014. doi:10.1016/j.bbrc.2014.05.090
11. Liu Y, Liu C, Zheng W. et al. PCR detection of Klebsiella pneumoniae in infant formula based on 16S–23S internal transcribed spacer. Int J Food Microbiol. Epub ahead of print 2008. doi:10.1016/j.ijfoodmicro.2008.03.005
12. Wayne P. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement M100-S25, Clinical and Laboratory Standards Institute. 2015:240.
13. Wasfi R, Elkhatib WF, Ashour HM. Molecular typing and virulence analysis of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from Egyptian hospitals. Sci Rep. Epub ahead of print 2016. doi:10.1038/srep38929
14. Beceiro A, Tomás M, Bou G. Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin Microbiol Rev. Epub ahead of print 2013. doi:10.1128/CMR.00059-12
15. Lev AI, Astashkin EI, Kislichkina AA, et al. Comparative analysis of Klebsiella pneumoniae strains isolated in 2012–2016 that differ by antibiotic resistance genes and virulence genes profiles. Pathog Glob Health. 2018;112(3):142–151. doi:10.1080/20477724.2018.1460949
16. Akter J, Masudul Azad Chowdhury AM, Forkan MA. Study on prevalence and antibiotic resistance pattern of Klebsiella isolated from clinical samples in South East Region of Bangladesh. Am J Drug Discov Dev. Epub ahead of print 2014. doi:10.3923/ajdd.2014.73.79
17. Riaz S, Faisal M, Hasnain S. Prevalence and comparison of beta-lactamase producing Escherichia coli and Klebsiella spp. from clinical and environmental sources in Lahore, Pakistan. African J Microbiol Res. Epub ahead of print 2012. doi:10.5897/AJMR11.1457
18. Lina TT, Rahman SR, Gomes DJ. Multiple-antibiotic resistance mediated by plasmids and integrons in uropathogenic Escherichia coli and Klebsiella pneumoniae. Bangladesh J Microbiol. 2007;24(1):19–23. doi:10.3329/bjm.v24i1.1231
19. Sarathbabu DR, Ramani DTV, Rao DKB, et al. Antibiotic susceptibility pattern of Klebsiella pneumoniae isolated from sputum, urine and pus samples. J Pharm Biol Sci. 2016;11;5(29):1470-1475.
20. Heidary M, Nasiri MJ, Dabiri H. Prevalence of drug-resistant Klebsiella pneumoniae in Iran : a review article. Iran J Public Heal. 2018;47(3):317–326.
21. El Bouamri MC, Arsalane L, El Kamouni Y. et al. Antimicrobial susceptibility of urinary Klebsiella pneumoniae and the emergence of carbapenem-resistant strains: a retrospective study from a university hospital in Morocco, North Africa. African J Urol. Epub ahead of print 2015. doi:10.1016/j.afju.2014.10.004
22. Chaudhary P, Bhandari D, Thapa K, et al. Prevalence of extended spectrum beta-lactamase producing Klebsiella Pneumoniae isolated from urinary tract infected patients. J Nepal Health Res Counc. 2004; 1;22(2):87.
23. Paczosa MK, Mecsas J. Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol Mol Biol Rev. Epub ahead of print 2016. doi:10.1128/MMBR.00078-15
24. Lin WP, Wang JT, Chang SC. et al. The antimicrobial susceptibility of Klebsiella pneumoniae from community settings in Taiwan, a trend analysis. Sci Rep. Epub ahead of print 2016. doi:10.1038/srep36280
25. Chander A, Shrestha CD. Prevalence of extended spectrum beta lactamase producing Escherichia coli and Klebsiella pneumoniae urinary isolates in a tertiary care hospital in Kathmandu, Nepal. BMC Res Notes. Epub ahead of print 2013. doi:10.1186/1756-0500-6-487
26. Osthoff M, McGuinness SL, Wagen AZ. et al. Urinary tract infections due to extended-spectrum beta-lactamase-producing Gram-negative bacteria: identification of risk factors and outcome predictors in an Australian tertiary referral hospital. Int J Infect Dis. Epub ahead of print 2015. doi:10.1016/j.ijid.2015.03.006
27. Zhang J, Zhou K, Zheng B. et al. High prevalence of ESBL-producing Klebsiella pneumoniae causing community-onset infections in China. Front Microbiol. Epub ahead of print 2016. doi:10.3389/fmicb.2016.01830
28. Hasan Ejaz H, ul-Haq K, Zafa A, et al. Urinary tract infections caused by extended spectrum β-lactamase (ESBL) producing Escherichia coli and Klebsiella pneumoniae. African J Biotechnol. 2011;10(73):16661–16666. doi:10.5897/AJB11.2449
29. Mehrgan H, Rahbar M. Prevalence of extended-spectrum β-lactamase-producing Escherichia coli in a tertiary care hospital in Tehran, Iran. Int J Antimicrob Agents. 2008;31(2):147–151. doi:10.1016/j.ijantimi-cag.2007.09.008
30. Babypadmini S, Appalaraju B. Extended spectrum -lactamases in urinary isolates of Escherichia coli and Klebsiella pneumoniae - prevalence and susceptibility pattern in a tertiary care hospital. Indian J Med Microbiol. 2004;22(3):172–174.
31. Aminzadeh Z, Sadat Kashi M, Sha’bani M. Bacteriuria by extended-spectrum Beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae: isolates in a governmental hospital in South of Tehran, Iran. Iran J Kidney Dis. 2008;2(4):197–200.
32. Al-Zahrani A, Akhtar N. Susceptibility patterns of extended spectrum ß-Lactamase (ESBL)-producing Escherichia coli and Klebsiella pneumoniae isolated in a teaching hospital. Pakistan J Med Res. 2004. Available from: http://www.pmrc.org.pk/PJMR44_2/susceptibilitypatterns of extended spectrum%DF-lactamase(esbl)-producingescherichiacoliandklebsiellapneumoniaeisolatedinateachinghospital.pdf.
33. Mekki AH, Hassan AN, Elsayed DEM. Extended spectrum beta lactamases among multi drug resistant Escherichia coli and Klebsiella species causing urinary tract infections in Khartoum. J Bacteriol Res. 2010;2:18–21.
34. Seibert G, Hörner R, Meneghetti BH, et al. Nosocomial infections by Klebsiella pneumoniae carbapenemase producing enterobacteria in a teaching hospital. Einstein (Sao Paulo). 2014;12(3):282–286. doi:10.1590/s1679-45082014ao3131
35. Jian-li W, Yuan-yuan S, Shou-yu G. et al. Serotype and virulence genes of Klebsiella pneumoniae isolated from mink and its pathogenesis in mice and mink. Sci Rep. Epub ahead of print 2017. doi:10.1038/s41598-017-17681-8
36. Turton JF, Payne Z, Coward A. et al. Virulence genes in isolates of Klebsiella pneumoniae from the UK during 2016, including among carbapenemase gene-positive hypervirulent K1-st23 and ‘non-hypervirulent’ types ST147, ST15 and ST383. J Med Microbiol. Epub ahead of print 2018. doi:10.1099/jmm.0.000653
37. Compain F, Babosan A, Brisse S, et al. Multiplex PCR for detection of seven virulence factors and K1/K2 capsular serotypes of Klebsiella pneumoniae. J Clin Microbiol. 2014;52(12):4377–4380. doi:10.1128/JCM.00749-14
38. Souza Lopes AC, Rodrigues JF, Cabral AB. et al. Occurrence and analysis of irp2 virulence gene in isolates of Klebsiella pneumoniae and Enterobacter spp. from microbiota and hospital and community-acquired infections. Microb Pathog. Epub ahead of print 2016. doi:10.1016/j.micpath.2016.04.018
39. Schubert S, Cuenca S, Fischer D. et al. High-pathogenicity island of Yersinia pestis in enterobacteriaceae isolated from blood cultures and urine samples: prevalence and functional expression. J Infect Dis. Epub ahead of print 2000. doi:10.1086/315831
40. Autenrieth I, Hantke K, Heesemann J. Immunosuppression of the host and delivery of iron to the pathogen: a possible dual role of siderophores in the pathogenesis of microbial infections? Med Microbiol Immunol. Epub ahead of print 1991. doi:10.1007/BF00206117
41. Carniel E. The Yersinia high-pathogenicity island: an iron-uptake island. Microbes Infect. Epub ahead of print 2001. doi:10.1016/S1286-4579(01)01412-5
42. Turton JF, Baklan H, Siu LK. et al. Evaluation of a multiplex PCR for detection of serotypes K1, K2 and K5 in Klebsiella sp. and comparison of isolates within these serotypes. FEMS Microbiol Lett. Epub ahead of print 2008. doi:10.1111/j.1574-6968.2008.01208.x
43. Feizabadi MM, Raji N, Delfani S. Identification of Klebsiella pneumoniae K1 and K2 capsular types by PCR and Quellung test. Jundishapur J Microbiol. Epub ahead of print 2013. doi:10.5812/jjm.7585
44. Ikeda M, Mizoguchi M, Oshida Y, et al. Clinical and microbiological characteristics and occurrence of Klebsiella pneumoniae infection in Japan. Int J Gen Med. 2018;11:293.
45. Jenney AW, Clements A, Farn JL. et al. Seroepidemiology of Klebsiella pneumoniae in an Australian Tertiary Hospital and its implications for vaccine development. J Clin Microbiol. Epub ahead of print 2006. doi:10.1128/JCM.44.1.102-107.2006
46. Piddock LJV. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev. Epub ahead of print 2006. doi:10.1128/CMR.19.2.382-402.2006
47. Chen JH, Siu LK, Fung CP. et al. Contribution of outer membrane protein K36 to antimicrobial resistance and virulence in Klebsiella pneumoniae. J Antimicrob Chemother. Epub ahead of print 2010. doi:10.1093/jac/dkq056
48. Padilla E, Llobet E, Doménech-Sánchez A. et al. Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob Agents Chemother. Epub ahead of print 2010. doi:10.1128/AAC.00715-09
49. Tsai YK, Fung CP, Lin JC. et al. Klebsiella pneumoniae Outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob Agents Chemother. Epub ahead of print 2011. doi:10.1128/AAC.01275-10
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.