")
Back to Journals » Cancer Management and Research » Volume 12
Modulation of MnSOD and FoxM1 Is Involved in Invasion and EMT Suppression by Isovitexin in Hepatocellular Carcinoma Cells
Authors Qiu Y, Cao X, Liu L, Cao X, Yuan Q, Li X, Cui Y, Xu C, Zou C, Ren K , Cao J
Received 8 January 2020
Accepted for publication 28 May 2020
Published 14 July 2020 Volume 2020:12 Pages 5759—5771
DOI https://doi.org/10.2147/CMAR.S245283
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
This paper has been retracted.
Yebei Qiu,1,2,* Xiaocheng Cao,1,2,* Lihua Liu,3,4 Xiaozheng Cao,3,4 Qing Yuan,5 Xiang Li,5 Yinghong Cui,1,5 Chang Xu,1,2 Chang Zou,4,6 Kaiqun Ren,1,2 Jianguo Cao1,2
1The Key Laboratory of Model Animals and Stem Cell Biology in Hunan Province, Medical College, Hunan Normal University, Changsha 410013, People’s Republic of China; 2The Key Laboratory of Study and Discover of Small Targeted Molecules of Hunan Province, Medical College, Hunan Normal University, Changsha 410013, People’s Republic of China; 3Pharmacy Department, The Second Clinical Medical School of Jinan University, Shenzhen People’s Hospital, Shenzhen 518020, People’s Republic of China; 4Shenzhen Public Service Platform on Tumor Precision Medicine and Molecular Diagnosis, Shenzhen People’s Hospital, Shenzhen 518020, People’s Republic of China; 5Department of Preclinical Medicine, Medical College, Hunan Normal University, Changsha, Hunan 410013, People’s Republic of China; 6Clinical Medical Research Center, The Second Clinical Medical School of Jinan University, Shenzhen People’s Hospital, Shenzhen 518020, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianguo Cao; Kaiqun Ren
Kaiqun Ren the Key Laboratory of Model Animals and Stem Cell Biology in Hunan Province, Medical College, Hunan Normal University, Changsha 410013, People’s Republic of China
Tel +86-0731-88912434
Email [email protected]; [email protected]
Background: Manganese superoxide dismutase (MnSOD) induces FoxM1 expression, subsequently contributing to migration in several cancer cells. Isovitexin (ISOV) was recently found to downregulate MnSOD and FoxM1, decreasing stemness in hepatocellular carcinoma (HCC) stem-like cells (HCSLCs). The current study aimed to determine whether inhibition of migration, invasion and EMT in HCSLCs by ISOV results from MnSOD/FoxM1 signaling blockade and subsequent Twist1, Slug, ZEB1 and MMP-2 downregulation.
Materials and Methods: We examined the migratory and invasive capabilities and EMT phenotype in HCC cells and their HCSLCs, respectively, by wound-healing assay, transwell invasion assay and Western blot after treatment with non-cytotoxic concentrations of ISOV, and explored the mechanism by which ISOV affects migration, invasion and EMT by MnSOD or FoxM1 knockdown and/or overexpression in HCSLCs or HCC cells.
Results: The results showed that ISOV not only downregulated MnSOD and FoxM1 but also suppressed the migratory and invasive capabilities and reversed the EMT phenotype in HCSLCs, which was reflected by elevated E-cadherin protein amounts, and reduced N-cadherin, Twist1, Slug, ZEB1 and MMP-2 protein levels. The suppressive effects of ISOV on the migratory and invasive capabilities and EMT phenotype could be potentiated by MnSOD or FoxM1 knockdown in HCSLCs, and attenuated by MnSOD or FoxM1 overexpression in HCC cells. Importantly, FoxM1 overexpression reversed MnSOD knockdown combined with ISOV suppression on the migratory and invasive capabilities and EMT phenotype in HCSLCs, while having little effects on MnSOD expression.
Conclusion: Collectively, the above findings demonstrated that ISOV suppresses migration, invasion and EMT in HCSLCs by blocking MnSOD/FoxM1 signaling subsequently inhibiting the expression of EMT-related transcription factors and MMP-2.
Keywords: hepatocellular carcinoma, cancer stem cell, isovitexin, epithelial-mesenchymal transition, MnSOD, FoxM1, Twist1, MMP-2
Introduction
Hepatocellular carcinoma (HCC) ranks fifth among malignancies in terms of incidence, and represents the third cause of malignant tumor-related death around the world; its low survival rate is due to a lack of efficient therapeutics.1,2 Although the cytologic pathogenesis of HCC is not completely elucidated, a small cell subset with stem cell characteristics, namely cancer stem cell-like cells (CSLCs) can initiate the tumor and promote cancer progression, recurrence and acquisition of resistance to chemotherapy.3,4 To date, it has been confirmed that epithelial-mesenchymal transition (EMT), an embryonic developmental process, confers stem-cell like features to cancer cells.5,6 Therefore, the development of agents targeting CSLCs to reverse EMT deserves further attention.
Forkhead box M1 (FoxM1) is a carcinogenic transcription factor that is abnormally upregulated in various cancers,including HCC.7,8 Suppression of FoxM1 has inhibitory effects on tumor progression and metastasis.9,10 Increased expression of FoxM1 was observed in HCC tissues, in association with poor prognosis of patients with HCC.11–14 FoxM1 silencing in mouse hepatocytes inhibits cell proliferation and reduces the formation of diethyl-nitrosamine-induced hepatoma.15 Our laboratory and others demonstrated that elevated FoxM1 by manganese superoxide dismutase (MnSOD) promotes migration and invasion.16,17 Whether and how FoxM1 upregulated by MnSOD induces the migratory and invasive capabilities and EMT phenotype in HCC stem-like cells (HCSLCs), thereby stimulating tumor progression, remains unknown.
It has been reported that MnSOD induces FoxM1 expression and promotes aggressiveness in lung cancer.17 Our recent study demonstrated that MnSOD is overexpressed in lung CSLCs from H460 cells, and confers carcinogenesis and lung CSLC properties through activation of the FoxM1 transcriptional factor.16 Because FoxM1 contributes to migration, invasion and EMT in various cancers,7,8,14 we initially aimed to evaluate whether FoxM1 upregulation by MnSOD overexpression leads to migration, invasion and EMT in HCSLCs.
Isovitexin (ISOV, apigenin-6-C-glucoside) has been shown to possess extensive biological activities.18 Natural flavone C-glycosides occur in different edible or medicinal plants.19,20 It is well known that ISOV and vitexin exert antitumor effects on HCC by targeting cell apoptosis and autophagy through regulation of apoptosis-related proteins such as Bax and Bcl-2 as well as the autophagy-associated protein LC3-II.21–24 We recently confirmed that ISOV inhibits stemness in HCSLCs.25 However, whether ISOV suppresses migration, invasion and EMT in HCSLCs remains unknown.
This work demonstrated enhanced migratory and invasive capabilities and induced EMT in HCSLCs compared with HCC cells. We firstly showed that ISOV suppressed migration and invasion, and reversed the EMT phenotype by downregulating MnSOD, FoxM1, Twist1, Slug, ZEB1 and MMP-2 in HCSLCs. These findings suggest MnSOD, and FoxM1 and its target proteins Twist1, Slug, ZEB1 and MMP-2 may promote migration, invasion and EMT, and ISOV might constitute a novel candidate for treating human HCC via suppression of migration and invasion as well as EMT inversion in HCSLCs.
Materials and Methods
Cell and Sphere Culture
MHCC97H and Sk-Hep-1 HCC cells as well as L-02 liver embryonic cells obtained from the Cell Bank of Chinese Academy of Sciences (Shanghai, China) were grown in DMEM (Invitrogen, Carlsbad, CA, USA) containing 10% FBS (Invitrogen) under a humid environment with 5% CO2 at 37°C.
To obtain HCSLCs,MHCC97H or Sk-Hep-1 cells were cultured in cancer stem cell medium (CSC-M) as described previously.25 The second-generation spheres were considered HCSLCs.25 For drug intervention, in primary sphere culture, cells were incubated with or without various concentrations of ISOV (Sigma-Aldrich St.; final concentrations of 5, 10 and 20 μM, respectively) in fresh CSC-M for 72h; then the second-generation spheres were cultured without ISOV.
Wound-Healing Assay
MHCC97Hcells (2×105) or Sk-Hep-1 cells (2×105), or respective HCSLCs (2×105) incubated with or without ISOV were cultured in DMEM (Invitrogen) with 10% FBS (Invitrogen) until 90% confluence. Then, a wound was created by scratching across the center of the well with a sterile 100 μL pipette tip. Next, the cells were washed and photographed in the same high-power field for analysis at 0 and 24h, respectively. MHCC97H cells, Sk-Hep-1 cells or respective HCSLCs treated with vehicle (0.1% DMSO) were used to standardize the number of migrated cells.
Invasion Assay
Cell invasion assay was performed in transwell systems with 8 μm pore membrane coating matrigel (Corning Incorporated, NY, USA). Briefly, MHCC97H cells (2×105) or Sk-Hep-1 cells (2×105), or respective HCSLC (2×105) incubated with or without ISOV in CSC-M without growth factors were plated in upper chambers (Corning Incorporated). CSC-M containing growth factors served as a chemoattractant in the bottom wells for 24h. Then, the invading cells were fixed with methanol, dyed with Wright’s stain, and mounted with Vectashield mounting medium (Vectorshield, Vector Laboratories). An optical microscope (Olympus DP72, Hamburg, Germany) was used for imaging (×200). Experiments were independently performed in triplicate.
Immunoblot
Immunoblot was carried out as described previously.16 Antibodies including anti-MnSOD, anti-FoxM1, anti-E-cadherin, anti-N-cadherin, anti-Twist, anti-MMP-2, anti-Slug, anti-ZEB1 (Cell Signaling Technology, Inc. USA) and anti-β-actin (Sigma, MA, USA) were used as primary antibodies. HRP-conjugated antibodies (Beyotime Institute; China) were used as secondary antibodies. Immunoreactive protein bands were detected using the enhanced chemiluminescence (ECL) reagent (Millipore; USA).
Adenoviruses and Infection
MHCC97H cells (2×105) or HCSLC cells (2×105) were infected with various adenoviruses harboring the indicated shRNA or cDNA at a multiplicity of infection (MOI) of 100, as described previously.16
Statistical Analysis
Statistical analyses were performed with the SPSS 20.0 software (IBM, Armonk, NY, USA) and SigmaPlot 12.5 (Systat Software Inc., San Jose, CA, USA). Data are mean±standard deviation (SD). Unpaired two-tailed Student’s t-test was used to compare various groups to controls. One-way ANOVA followed by post-hoc Tukey’s test was performed for pairwise comparison among groups p<0.05 was considered statistically significant.
Results
ISOV Inhibits Invasion and Reverses EMT Phenotype in HCSLCs from MHCC97H Cells
We initially performed immunoblot to analyze MnSOD and FoxM1 protein levels, and found that they were higher in HCSLCs compared with MHCC97H cells (Figure 1A). Meanwhile, cell migration and invasion rates were substantially elevated in HCSLCs compared with MHCC97H cells (Figure 1B and C). In addition, increased N-cadherin, Twist1, Slug, ZEB1 and MMP-2 amounts, and decreased E-cadherin levels, were found in HCSLCs compared with MHCC97H cells (Figures 1D and E; Supplementary Figure 1).Our results revealed that HCSLCs had higher migratory and invasive capabilities and enhanced EMT phenotype, which may be associated with increased MnSOD, FoxM1 and its target molecules Twist1, slug, ZEB1 and MMP-2 protein amounts.
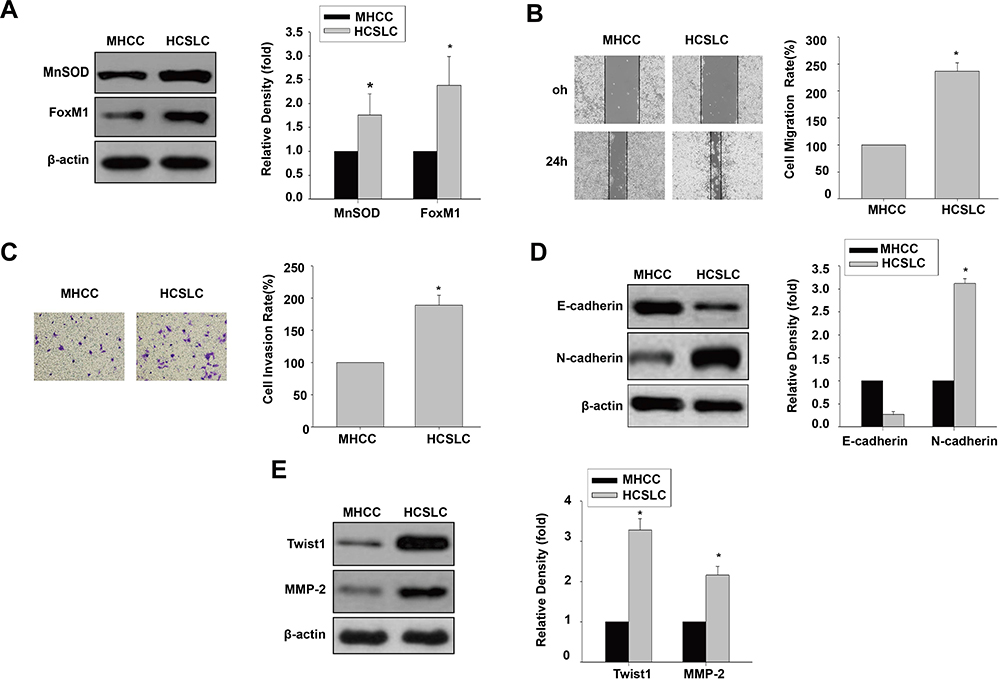 |
Figure 1 Comparison of invasion and EMT between HCSLCs and MHCC97H cells. MnSOD and FoxM1 protein levels (A), cell migratory and invasive capacities (B and C), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (D and E) in HCSLCs and MHCC97H Cells are shown (n=3; unpaired two-tailed Student’s t-test, *P<0.05 vs MHCC97H cells). |
ISOV and vitexin were shown to possess antitumor activities in hepatoma via cell apoptosis and autophagy.21–24 Therefore, we employed the CCK-8 kit to measure cell viability in liver embryonic L-02 cells, MHCC97H cells and the corresponding HCSLCs treated with ISOV. The results showed that ISOV remarkably suppressed MHCC97H cells and the corresponding HCSLCs compared with L02 cells (Figure 2A). IC50 was about 80 μM for MHCC97H cells and 40 μM for the corresponding HCSLCs (Figure 2A). We next examined the effects of ISOV on protein (MnSOD and FoxM1) levels, cell functions (migration and invasion) and EMT phenotype-related protein (E-cadherin, N-cadherin, Twist1, Slug, ZEB1 and MMP-2) levels in HCSLCs after treatment with non-cytotoxic concentrations (5.0, 10 and 20 μM) of ISOV. The results revealed that ISOV significantly downregulated MnSOD and FoxM1 in HCSLCs (Figure 2B). In addition, a substantial reduction of cell migration and invasion in HCSLCs was found after incubation with ISOV for 24h (Figure 2C and D). Furthermore, ISOV significantly altered the levels of the EMT-related proteins Twist1, Slug, ZEB1 and MMP-2, in a dose-dependent manner (Figures 2E and F; Supplementary Figure 2).Our data showed that ISOV could inhibit cell migration and invasion, and reverse EMT phenotype, which may be related to downregulation of MnSOD, FoxM1 and its downstream targets Twist1, Slug, ZEB1 and MMP-2.
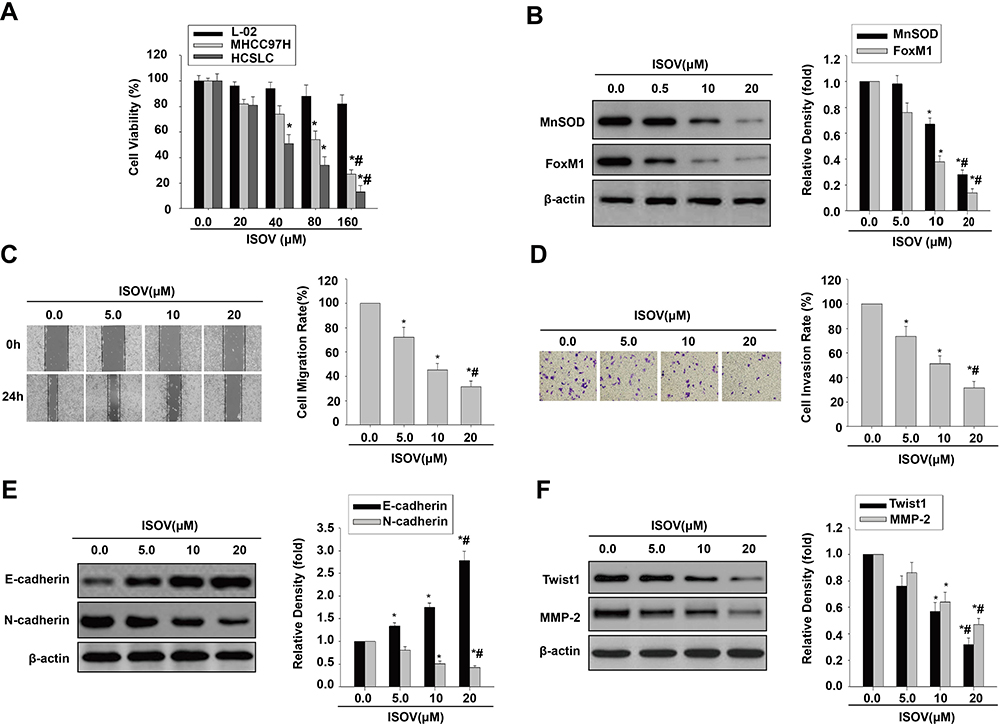 |
Figure 2 ISOV inhibits migration and invasion and reverses the EMT phenotype in HCSLCs. Effects of ISOV (20, 40, 80 and 160 μM) on cell viability (A) in L-02 cells, MHCC97H cells and HCSLCs (n=3; *P<0.05 vs vehicle (ISOV 0 μM); #P<0.05 vs 80 μM ISOV; one-way ANOVA). Effects of ISOV (5, 10, 20 μM) on MnSOD and FoxM1 protein levels (B) and cell migratory and invasive capacities (C and D), E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (E and F) in HCSLCs (n=3; *P<0.05 vs vehicle (ISOV 0 μM); #P<0.05 vs 5.0 μM ISOV; one-way ANOVA). |
Changes of MnSOD Expression Affect Cell Invasion and the EMT Phenotype in HCSLCs and MHCC97H Cells
We next demonstrated that MnSOD shRNA significantly downregulated MnSOD and FoxM1 in HCSLCs relative to vector control or non-transduced cells (Figure 3A). Substantial reductions of cell migration and invasion in HCSLCs after MnSOD silencing were found in comparison with vector control- or non-transduced cells (Figure 3B and C). In addition, decreased N-cadherin, Twist1, Slug, ZEB1 and MMP-2 levels as well as increased E-cadherin expression were observed in comparison with vector control or non-transduced MHCC97H cells (Figures 3D and E; Supplementary Figure 3A). Conversely, MnSOD overexpression significantly increased MnSOD and FoxM1 expression amounts in MHCC97H cells relative to vector control or non-transduced cells (Figure 3F). Substantially elevated cell migration and invasion in MHCC97H cells overexpressing MnSOD were found compared with vector control or non-transduced cells (Figure 3G and H). Meanwhile, increased N-cadherin, Twist1, Slug, ZEB1 and MMP-2 expression levels as well as decreased E-cadherin amounts were observed in MHCC97H cells overexpressing MnSOD relative to vector control or non-transduced cells (Figures 3I and J; Supplementary Figure 3B). Collectively, the above findings indicate that promotion of cell migration, invasion and EMT in HCSLCs may require elevated MnSOD expression.
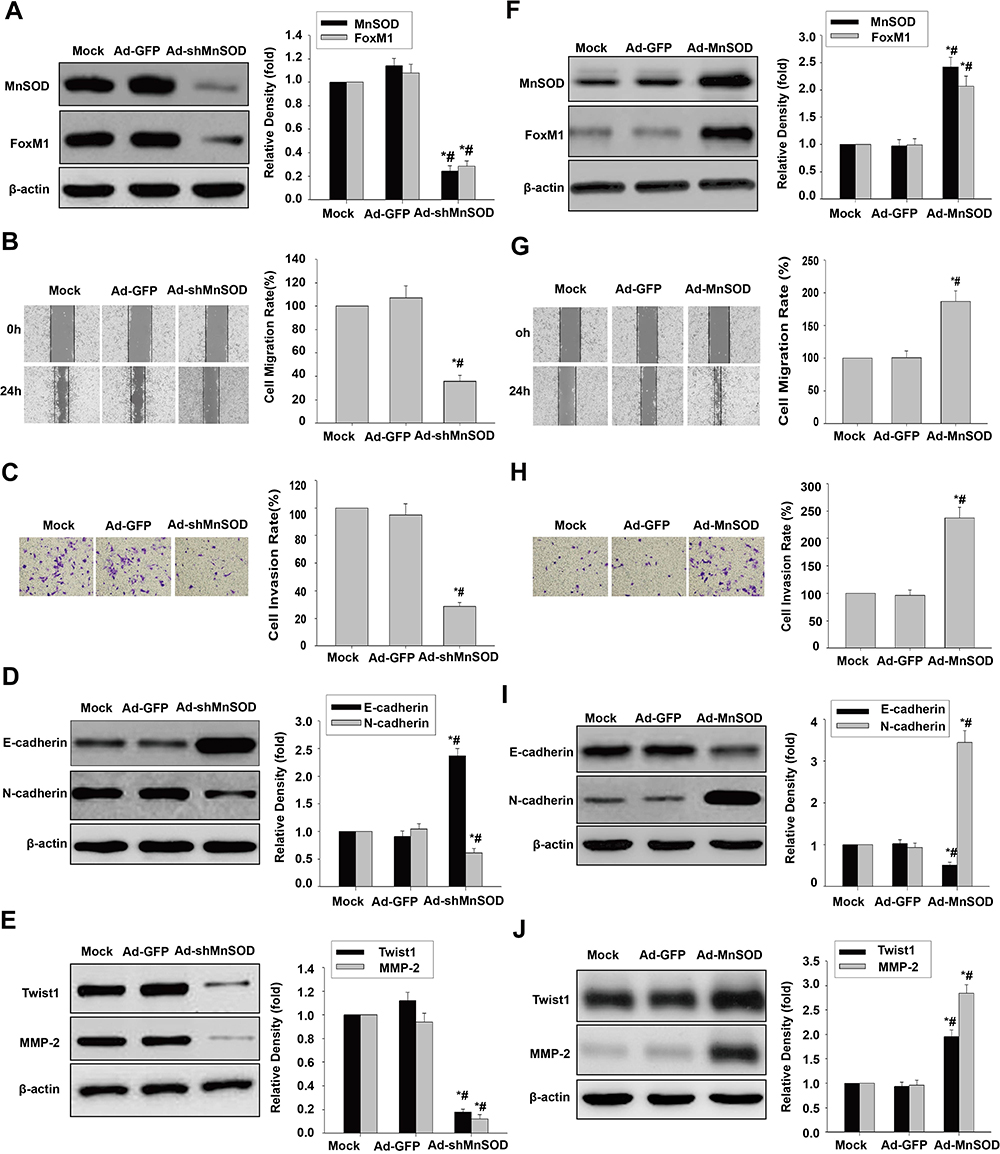 |
Figure 3 Effects of MnSOD silencing and overexpression on migration, invasion and EMT. Ad-GFP and Ad-shMnSOD were transduced into HCSLCs, respectively. MnSOD and FoxM1 protein levels (A), cell migratory and invasive capacities (B and C), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (D and E) in untreated-HCSLCs (Mock), harboring Ad-GFP or Ad-shMnSOD (n=3; *P<0.05 vs Mock; #P<0.05 vs Ad-GFP; one-way ANOVA) are shown. MHCC97H cells were transduced with Ad-GFP or Ad-MnSOD. MnSOD and FoxM1 protein levels (F), cell migratory and invasive capacities (G and H), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (I and J) in MHCC97H cells transfected with Ad-GFP or Ad-MnSOD (n=3; *P<0.05 vs Mock; #P<0.05 vs Ad-GFP; one-way ANOVA) are shown. |
Changes of MnSOD Expression Affect ISOV-Associated Inhibition of Migration and EMT in HCSLCs and MHCC97H Cells
For determining the relationship between MnSOD regulation and ISOV-associated repression of cell migration, invasion and EMT, MnSOD-knockdown HCSLCs and MHCC97H cells overexpressing MnSOD were incubated with or without ISOV. Silencing of MnSOD in combination with ISOV treatment more pronouncedly reduced MnSOD and FoxM1 levels compared with MnSOD silencing or ISOV alone (Figure 4A). Meanwhile, a starker reduction of cell migration and invasion was found after MnSOD knockdown combined with ISOV treatment in comparison with MnSOD knockdown or ISOV alone in HCSLCs (Figure 4B and C). In addition, more pronouncedly decreased N-cadherin, Twist1, Slug, ZEB1 and MMP-2 expression levels and increased E-cadherin expression were observed after MnSOD knockdown plus ISOV administration relative to MnSOD knockdown or ISOV treatment alone in HCSLCs (Figures 4D and E; Supplementary Figure 4A). Conversely, the suppressive effects of ISOV on MnSOD and FoxM1 expression as well as migration, invasion and EMT were nearly abrogated by MnSOD overexpression in MHCC97H cells (Figures 4F–J; Supplementary Figure 4B). Collectively, the above findings indicate that the suppressive effects of ISOV on cell migration, cell invasion and EMT in HCSLCs may require MnSOD downregulation.
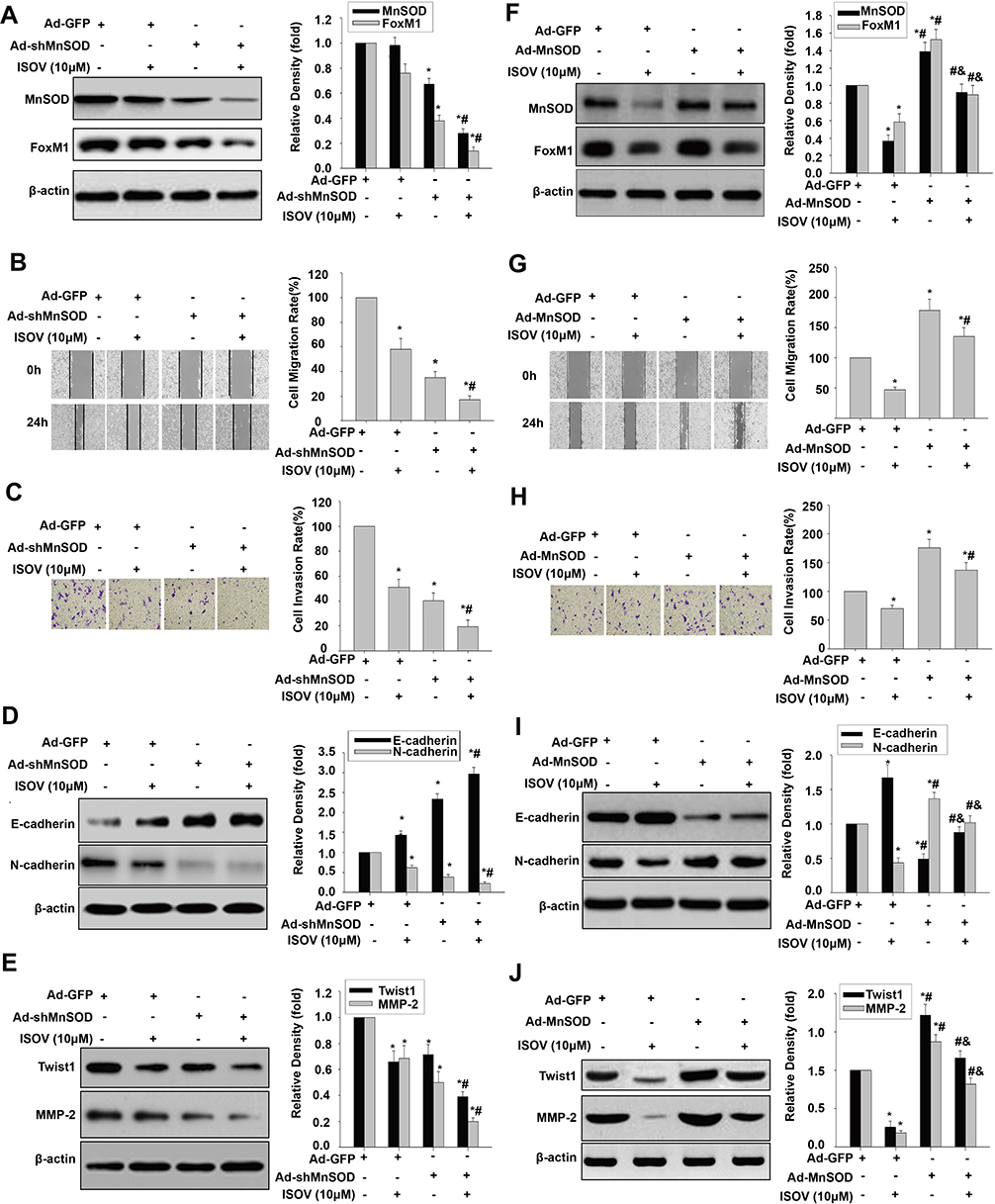 |
Figure 4 Effects of ISOV combined with MnSOD silencing and overexpression on migration, invasion and the EMT phenotype. MnSOD and FoxM1 protein levels (A), cell migratory and invasive capacities (B and C), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (D and E) in HCSLCs transfected with Ad-GFP or Ad-shMnSOD, and incubated with or without ISOV(10 μM) (n=3; *P<0.05 vs Ad-GFP; #P<0.05 vs Ad-GFP plus10.0 μM ISOV; one-way ANOVA) are shown. MHCC97H cells were transfected with Ad-GFP or Ad-MnSOD, and incubated with ISOV (10 μM). MnSOD and FoxM1 protein levels (F), cell migratory and invasive capacities (G and H), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (I and J) in MHCC97H cells transfected with Ad-GFP or Ad-MnSOD, incubated with or without ISOV(10 μM) (n=3; *P<0.05 vs Ad-GFP; #P<0.05 vs Ad-GFP plus 10.0 μM ISOV; &P<0.05 vs Ad-MnSOD alone; one-way ANOVA) are shown. |
Changes of FoxM1 Expression Affect Invasive Capability and EMT in HCSLCs and MHCC97H Cells
To analyze the link between FoxM1 and cell migration, invasion and EMT, FOXM1 shRNA and FoxM1 cDNA were transduced into HCSLCs and MHCC97H cells, respectively. FOXM1 knockdown remarkably decreased FoxM1 expression amounts, without affecting MnSOD expression (Figure 5A). Substantial decreases of cell migration and invasion were observed in HCSLC safter FoxM1 silencingrelative to vector- or non-transduced-control cells (Figure 5B and C). In addition, FoxM1 knockdown decreased N-cadherin, Twist1, Slug, ZEB1 and MMP-2 protein amounts, and upregulated E-cadherin in HCSLCs upon FoxM1 silencing compared with vector control or non-transduced HCSLCs (Figures 5D and E; Supplementary Figure 5A). Conversely, FoxM1 overexpression significantly elevated FoxM1 expression amounts, without affecting MnSOD expression (Figure 5F). Significantly increased cell migration and invasion in MHCC97H cells overexpressing FoxM1 were observed compared with vector control or non-transduced cells (Figure 5G and H). In addition, FoxM1 overexpression also elevated N-cadherin, Twist1, Slug, ZEB1 and MMP-2 expression levels, and downregulated E-cadherin compared with vector control or non-transduced MHCC97H cells (Figures 5I and J; Supplementary Figure 5B). Taken together, the above findings indicate that promotion of cell migration, invasion and EMT in HCSLCs require elevated FoxM1, and its downstream targets Twist1, Slug, ZEB1 and MMP-2 might be controlled by MnSOD.
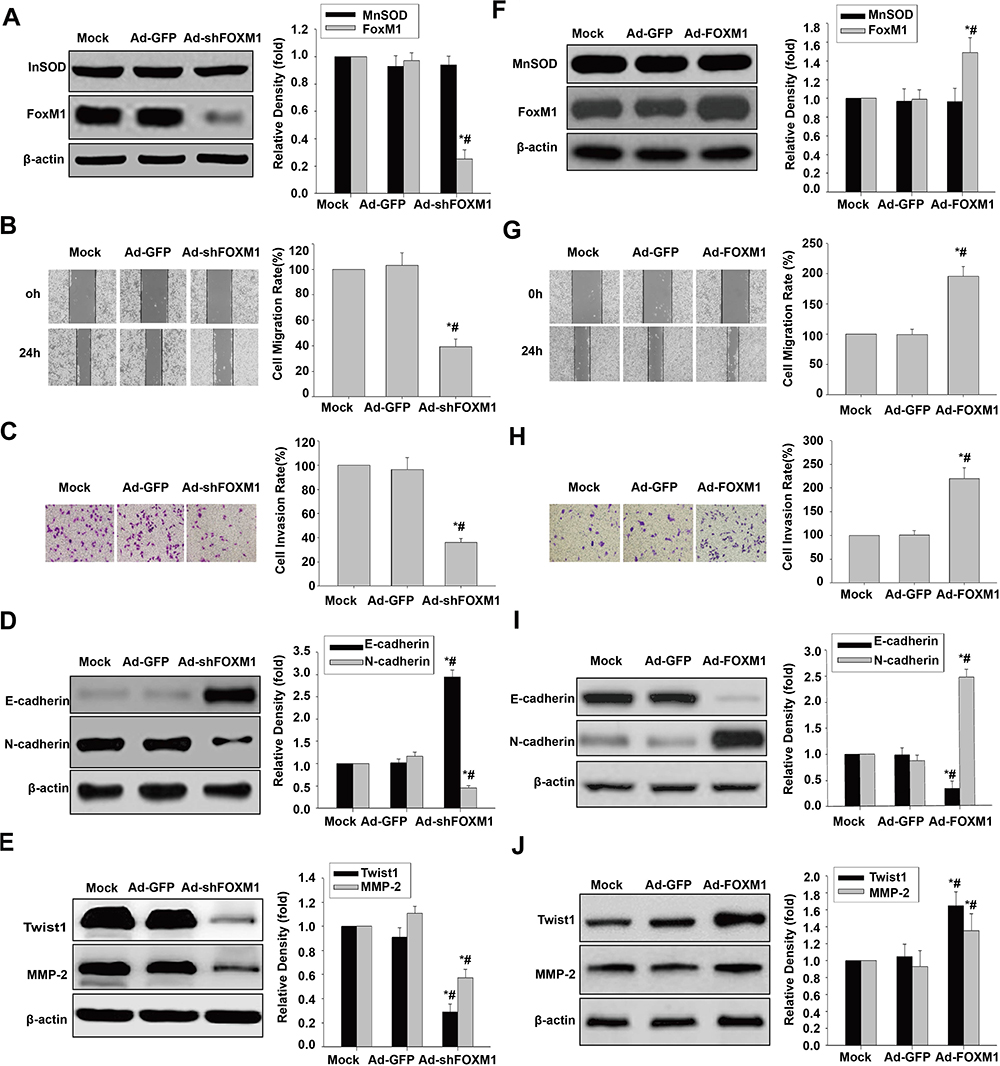 |
Figure 5 Effects of FoxM1 silencing and overexpression on migration, invasion and the EMT phenotype. MnSOD and FoxM1 protein levels (A), cell migratory and invasive capacities (B and C), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (D and E) in HCSLCs transfected with Ad-GFP or Ad-shFoxM1 (n=3; *P<0.05 vs Mock; #P<0.05 vs Ad-GFP;one-way ANOVA) are shown. MnSOD and FoxM1 protein levels (F), cell migratory and invasive capacities (G and H), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (I and J) in MHCC97H cells transfected with Ad-GFP or Ad-FoxM1 (n=3; *P<0.05 vs Mock; #P<0.05 vs Ad-GFP; one-way ANOVA) are shown. |
Changes of FoxM1 Expression Affect ISOV-Associated Inhibition of Invasion and EMT in HCSLCs or MHCC97H Cells
To determine whether ISOV inhibition of cell migration, invasion and EMT require FoxM1 repression, FoxM1 shRNA and FoxM1 cDNA were transduced into HCSLCs and MHCC97H cells, respectively. These cells were then treated with ISOV. We found that FoxM1 silencing significantly enhanced the suppressive effects of ISOV on FoxM1 expression, while ISOV-associated MnSOD downregulation was not altered after FoxM1 knockdown (Figure 6A). A more pronounced inhibition of cell migration and invasion was found after FoxM1 knockdown combined with ISOV relative to FoxM1 silencing or ISOV alone in HCSLCs (Figure 6B and C). In addition, FoxM1 knockdown also potentiated the downregulation of N-cadherin, Twist1,Slug, ZEB1 and MMP-2 as well as E-cadherin upregulation (Figures 6D and E; Supplementary Figure 6A) by ISOV. Conversely, repression of FoxM1 expression, cell migration, invasion and EMT phenotype induced by ISOV was nearly abrogated by overexpressing FoxM1 in MHCC97H cells (Figures 6F–6J; Supplementary Figure 6B). It should be noted that altered FoxM1 expression did not affect ISOV-associated suppression of MnSOD expression. Taken together, the above findings indicate that the suppressive effects of ISOV on cell migration, invasion and EMT in HCSLCs may require FoxM1 repression by MnSOD downregulation.
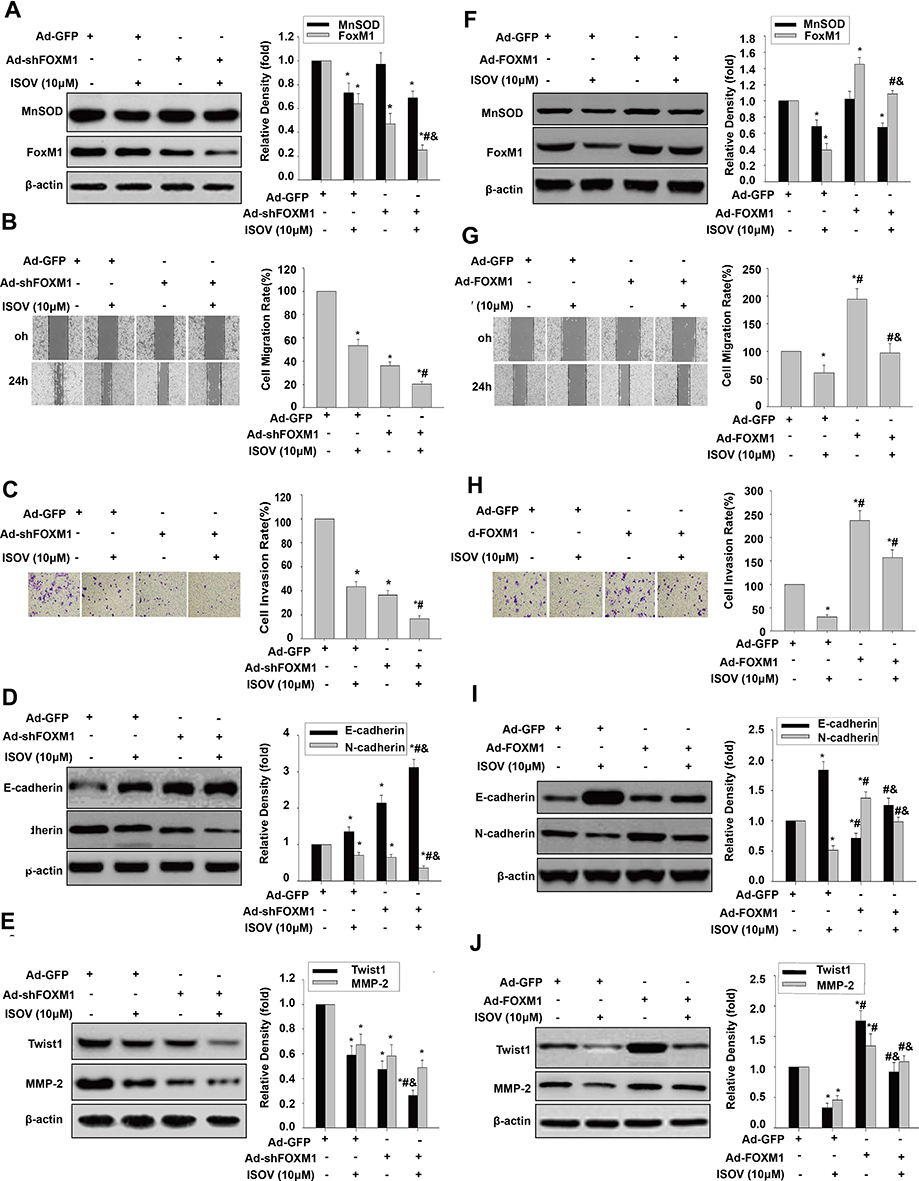 |
Figure 6 Effects of ISOV combined with FoxM1 silencing and overexpression on migration, invasion and the EMT phenotype. MnSOD and FoxM1 protein levels (A), cell migratory and invasive capacities (B and C), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (D and E) in HCSLCs transfected with Ad-GFP or Ad-shFoxM1, and incubated with or without ISOV (10 μM) (n=3; *P<0.05 vs Ad-GFP; #P<0.05 vs Ad-GFP plus 10.0 μM ISOV; &P<0.05 vs.Ad-shFoxM1 alone;one-way ANOVA) are shown. Ad-GFP and Ad-FoxM1 were transduced into MHCC97H cells incubated with or without ISOV (10 μM). MnSOD and FoxM1 protein levels (F), cell migratory and invasive capacities (G and H), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (I and J) in MHCC97H cells transfected with Ad-GFP or Ad-FoxM1, and incubated with or without ISOV (10 μM) (n=3; *P<0.05 vs Ad-GFP; #P<0.05 vs Ad-GFP plus 10.0 μM ISOV; &P<0.05 vs Ad- FoxM1 alone; one-way ANOVA). |
ISOV Inhibits Invasion and Reverses the EMT Phenotype in HCSLCs from SK-Hep-1 Cells
To determine whether ISOV represses cell migration, invasion and EMT by a mechanism similar to that observed in other HCC cell lines, spheres derived from the SK-Hep-1 cell line (SK-SC) were obtained. We then used the CCK-8 kit to measure cell viability in liver embryonic L-02 cells, SK-Hep-1 cells and SK-SC treated with ISOV. We found ISOV remarkably suppressed cell viability in SK-Hep-1 cells and SK-SC compared with L02 cells (Figure 7A). ISOV also significantly reduced the amounts of MnSOD and FoxM1 in SK-SC (Figure 7B). In addition, substantial reductions of cell migration and invasion in SK-SC were found after incubation with non-cytotoxic concentrations (10 and 20 μM) of ISOV for 24h (Figure 7C and D). Furthermore, ISOV significantly enhanced E-cadherin, attenuated N-cadherin, Twist1, Slug, ZEB1 and MMP-2 expressions in SK-SC (Figures 7E and F; Supplementary Figure 7).These results suggest that ISOV inhibited cell migration and invasion, and reversed the EMT phenotype by downregulating MnSOD, FoxM1, Twist1, Slug, ZEB1 and MMP-2, which was not limited to a specific type of cells.
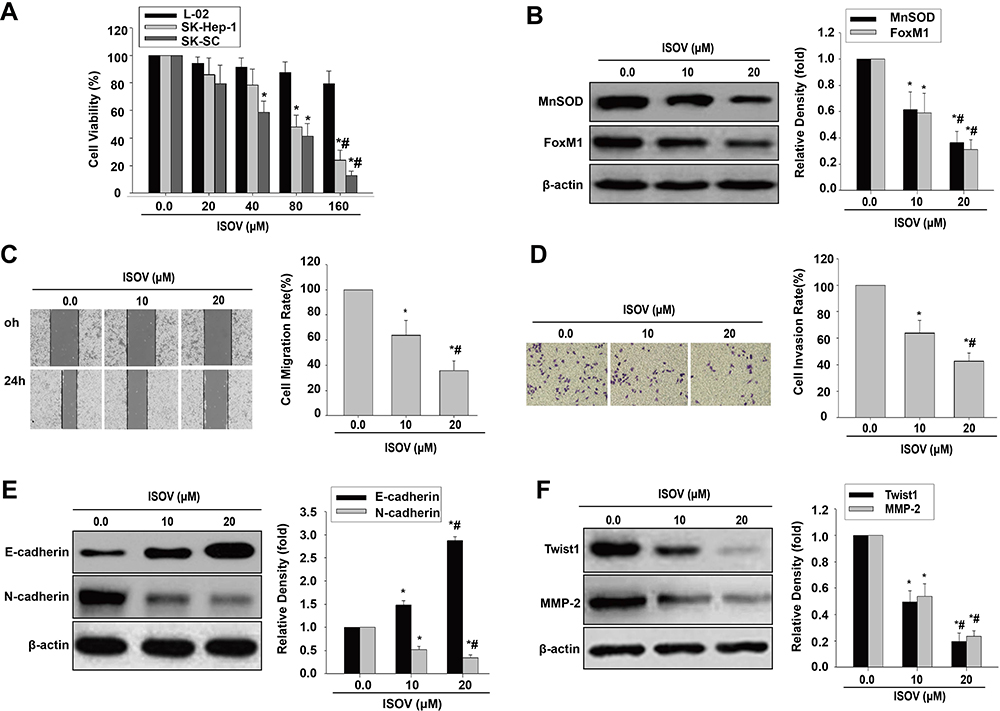 |
Figure 7 ISOV inhibits migratory and invasive capability and EMT in HCSLCs from SK-Hep-1 cells (SK-SC). Effects of ISOV (20, 40, 80 and 160 μM) on cell viability (A) in L-02 cells, SK-Hep-1 cells and SK-SC (n=3; *P<0.05 vs vehicle; #P<0.05 vs 80 μM ISOV; one-way ANOVA). Effects of ISOV (10, 20 μM) on MnSOD and FoxM1 protein levels (B), cell migratory and invasive capacities (C and D), and E-cadherin, N-cadherin, Twist1 and MMP-2 protein amounts (E and F) in SK-SC (n=3; *P<0.05 vs vehicle; #P<0.05 vs 10.0 μM ISOV; one-way ANOVA). |
Discussion
In the current study, we firstly clarified that elevated FoxM1 expression by MnSOD overexpression could promote migration, invasion and EMT in HCSLCs, indicating that the initiation, progression and metastasis of HCC is, at least partly, caused by MnSOD with the underlying mechanism involving FoxM1 upregulation. This is because EMT is a key step in the initiation, progression and metastasis of cancers.6,26–29 Importantly, we firstly demonstrated that the mechanisms underlying ISOV’s inhibitory effects on migratory and invasive capabilities as well as the EMT in HCSLCs occurred through downregulation of MnSOD, FoxM1, Twist1, Slug, ZEB1 and MMP-2. These findings suggest that HCSLCs present elevated migratory and invasive capabilities and induced EMT, which could significantly help in designing novel therapeutic agents targeting HCSLCs in HCC.30
MnSOD is considered an enzyme localized in the mitochondria; its overexpression impacts migration and invasion as well as the metabolic shift towards aerobic glycolysis in cancers.17,30 Our team and others have reported abnormal MnSOD is related to self-renewal in CSLCs.17,29,31–34 In the current study, we revealed that MnSOD may elicit migration and invasion as well as the EMT phenotype in HCSLCs.
It was shown that overexpression of MnSOD upregulates the cancerogenic transcriptional factor FoxM1 by releasing E2F1, thereby promoting the migration and invasion of lung cancer cells.17 Studies also showed that FoxM1 upregulated by MnSOD overexpressionis associated with stemness in CSLCs.17,29,31–34 Consistent with these results, the present study found that change of MnSOD expression markedly affected FoxM1 expression, whereas FoxM1 level change did not alter MnSOD expression. In addition, overexpression of FoxM1 reversed the inhibitory effects of MnSOD silencing on migratory and invasive abilities and EMT in MHCC97H cells. Accordingly, FoxM1 appears to be a downstream effector of MnSOD, with MnSOD/FoxM1 signaling promoting migration, invasion and EMT in HCSLCs.
EMT was initially identified as a process in embryogenesis. Subsequent studies confirmed that EMT mediates the maintenance of CSLC characteristics and enhances cell migratory and invasive capabilities in many cancers.35,36 The basic helix-loop-helix transcription factor Twist1 represses E-cadherin expression to endow the EMT phenotype in different cancers.27,37 Slug, also known as SNAIL2, is a transcription factor that regulates the EMT process in cancer progression.38 Slug can induce cancer cell metastasis and invasion by suppressing the epithelial phenotype and initiating EMT via binding to E-box DNA sequences found within the proximal promoter region of the E-cadherin gene.39,40 ZEB1 is considered a transcriptional repressor of epithelial genes involved in EMT.41 In the current study, we demonstrated that FoxM1 expression elevated by MnSOD overexpression raised Twist1, Slug and ZEB1 expression levels. Qian et al showed that Twist1 overexpression results in gastric cancer cell proliferation through FoxM1 upregulation.42 As FoxM1 upregulation may occur downstream of MnSOD as demonstrated above, the mutual activation of FoxM1 and Twist1, Slug and ZEB1 deserves further investigation.
FoxM1 has been reported to enhance MMP2-mediated invasion in human non-small cell lung cancer.43,44 MnSOD or FoxM1 overexpression elevated MMP2 levels, accompanied by increased invasion. The above responses were attenuated by MnSOD or FoxM1 shRNA, indicating MMP2 may be a downstream effector of the MnSOD/FoxM1 axis.
Our previous study showed that extracts harboring ISOV inhibit CSLC characteristics in lung cancer.19 ISOV and vitexin act as chemical isomers, exhibit strong cytotoxic activities against various cancer cells, including HCC cells, in vitro and in a xenograft tumor nude mouse model in vivo.21–25,45 However, no investigation on reversal of the EMT phenotype by ISOV has been reported so far. In the current study, we clearly provided evidence that ISOV represses migratory capability and reversed the EMT phenotype in HCSLCs by blocking MnSOD/FoxM1 signaling, subsequently downregulating Twist1, Slug, ZEB1 and MMP-2 in vitro. Nevertheless, the development of ISOV as a potential hepatoma chemopreventive and adjuvant chemotherapeutic agent warrants further investigation.
Conclusions
Collectively, our study demonstrated that repression of migration, invasion and EMT in HCSLCs by ISOV is due to MnSOD/FoxM1 signaling blockade, subsequently downregulating Twist1, Slug, ZEB1 and MMP-2. These findings suggest that ISOV may serve as a novel candidate for the treatment of HCC patients.
Abbreviations
ANOVA, analysis of variance; CSC-M, CSC-conditioned medium; CSLCs, cancer stem-like cells; EMT, epithelial-mesenchymal transition; FoxM1,Forkhead box M1; HCC, hepatocellular carcinoma; HCSLCs, hepatocellular carcinoma stem-like cells; ISOV, isovitexin; MnSOD, manganese superoxide dismutase; SD, standard deviation.
Data Sharing Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Ethics Approval and Informed Consent
The procedures were approved by the Ethics Committee of the Hunan Normal University.
Acknowledgments
The present study was supported by the National Natural Science Foundation of China (Nos. 81301894, 81302249, 81172375) and The Shenzhen Public Service Platform on Tumor Precision Medicine and Molecular Diagnosis, Shenzhen People’s Hospital.
Author Contributions
QYB and CXC performed and analyzed the experiments, wrote the paper. LLH, CXZ, YQ, LX, CYH, XC and ZC carried out the data collection, and data analysis. CJG and RKQ conceived and coordinated the study, designed, data analysis, revised the paper. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
3. Dawood S, Austin L, Cristofanilli M. Cancer stem cells: implications for cancer therapy. Oncology. 2014;28(12):1101–1107, 1110.
4. Pandit H, Li Y, Li X, Zhang W, Li S, Martin RCG. Enrichment of cancer stem cells via β-catenin contributing to the tumorigenesis of hepatocellular carcinoma. BMC Cancer. 2018;18(1):783. doi:10.1186/s12885-018-4683-0
5. Luo Y, Cui Y, Cao X, et al. 8-Bromo-7-methoxychrysin-blocked STAT3/Twist axis inhibits the stemness of cancer stem cell-like cell originated from SMMC-7721 cells. Acta Biochim Biophys Sin. 2017;49(5):458–464. doi:10.1093/abbs/gmx025
6. Li J, Zhou BP. Activation of β-catenin and Akt pathways by Twist are critical for the maintenance of EMT associated cancer stem cell-like characters. BMC Cancer. 2011;11:49. doi:10.1186/1471-2407-11-49
7. Yang N, Wang C, Wang Z, et al. FOXM1 recruits nuclear Aurora kinase A to participate in a positive feedback loop essential for the self-renewal of breast cancer stem cells. Oncogene. 2017;36(24):3428–3440. doi:10.1038/onc.2016.490
8. Kopanja D, Pandey A, Kiefer M, et al. Essential roles of FoxM1 in Ras-induced liver cancer progression and in cancer cells with stem cell features. J Hepatol. 2015;63(2):429–436. doi:10.1016/j.jhep.2015.03.023
9. Chen Y, Liu Y, Ni H, Ding C, Zhang X, Zhang Z. FoxM1 overexpression promotes cell proliferation and migration and inhibits apoptosis in hypopharyngeal squamous cell carcinoma resulting in poor clinical prognosis. Int J Oncol. 2017;51(4):1045–1054. doi:10.3892/ijo.2017.4094
10. Priller M, Poschl J, Abrao L, et al. Expression of FoxM1 is required for the proliferation of medulloblastoma cells and indicates worse survival of patients. Clin Cancer Res. 2011;17(21):6791–6801. doi:10.1158/1078-0432.CCR-11-1214
11. Shang R, Pu M, Li Y, Wang D. FOXM1 regulates glycolysis in hepatocellular carcinoma by transactivating glucose transporter 1 expression. Oncol Rep. 2017;37(4):2261–2269. doi:10.3892/or.2017.5472
12. Chen F, Bai G, Li Y, Feng Y, Wang L. A positive feedback loop of long noncoding RNA CCAT2 and FOXM1 promotes hepatocellular carcinoma growth. Am J Cancer Res. 2017;7(7):1423–1434.
13. Park YH, Kim SU, Kwon TH, et al. Peroxiredoxin II promotes hepatic tumorigenesis through cooperation with Ras/Forkhead box M1 signaling pathway. Oncogene. 2016;35(27):3503–3513. doi:10.1038/onc.2015.411
14. Meng FD, Wei JC, Qu K, et al. FoxM1 overexpression promotes epithelial-mesenchymal transition and metastasis of hepatocellular carcinoma. World J Gastroenterol. 2015;21(1):196–213. doi:10.3748/wjg.v21.i1.196
15. Wang X, Kiyokawa H, Dennewitz MB, Costa RH. The Forkhead Box m1b transcription factor is essential for hepatocyte DNA replication and mitosis during mouse liver regeneration. Proc Natl Acad Sci U S A. 2002;99(26):16881–16886. doi:10.1073/pnas.252570299
16. Fu Z, Cao X, Yang Y, Song Z, Zhang J, Wang Z. Upregulation of FoxM1 by MnSOD overexpression contributes to cancer stem-like cell characteristics in the lung cancer H460 cell line. Technol Cancer Res Treat. 2018;17:1533033818789635. doi:10.1177/1533033818789635
17. Chen PM, Wu TC, Shieh SH, et al. MnSOD promotes tumor invasion via upregulation of FoxM1-MMP2 axis and related with poor survival and relapse in lung adenocarcinomas. Mol Cancer Res. 2013;11(3):261–271. doi:10.1158/1541-7786.MCR-12-0527
18. Ganesan K, Xu B. Molecular targets of vitexin and isovitexin in cancer therapy: a critical review. Ann N Y Acad Sci. 2017;1401(1):102–113. doi:10.1111/nyas.13446
19. Cao X, Zou H, Cao J, et al. A candidate Chinese medicine preparation-Fructus Viticis Total Flavonoids inhibits stem-like characteristics of lung cancer stem-like cells. BMC Complement Altern Med. 2016;16:364. doi:10.1186/s12906-016-1341-4
20. Fu Y, Zu Y, Liu W, et al. Preparative separation of vitexin and isovitexin from pigeon pea extracts with macroporous resins. J Chromatogr A. 2007;1139(2):206–213. doi:10.1016/j.chroma.2006.11.015
21. Wang JG, Zheng XX, Zeng GY, Zhou YJ, Yuan H. Purified vitexin compound 1 induces apoptosis through activation of FOXO3a in hepatocellular carcinoma. Oncol Rep. 2014;31(1):488–496. doi:10.3892/or.2013.2855
22. Liu LH, Zhou YJ, Ding L, Zhang SZ, Sun J, Cao JG. Induction of apoptosis by VB1 in breast cancer cells: the role of reactive oxygen species and Bcl-2 family proteins. Int J Mol Med. 2014;33(2):423–430. doi:10.3892/ijmm.2013.1567
23. Wang J, Zheng X, Zeng G, Zhou Y, Yuan H. Purified vitexin compound 1 inhibits growth and angiogenesis through activation of FOXO3a by inactivation of Akt in hepatocellular carcinoma. Int J Mol Med. 2014;33(2):441–448. doi:10.3892/ijmm.2013.1587
24. Zhou Y, Liu YE, Cao J, et al. Vitexins, nature-derived lignan compounds, induce apoptosis and suppress tumor growth. Clin Cancer Res. 2009;15(16):5161–5169. doi:10.1158/1078-0432.CCR-09-0661
25. Cao X, Liu L, Yuan Q, et al. Isovitexin reduces carcinogenicity and stemness in hepatic carcinoma stem-like cells by modulating MnSOD and FoxM1. J Exp Clin Cancer Res. 2019;38(1):264. doi:10.1186/s13046-019-1244-6
26. Li C-W, Xia W, Huo L, et al. Epithelial-mesenchymal transition induced by TNF-α requires NF-κB-mediated transcriptional upregulation of Twist1. Cancer Res. 2012;72(5):1290–1300. doi:10.1158/0008-5472.CAN-11-3123
27. Dong W, Sun S, Cao X, et al. Exposure to TNF-α combined with TGF-β induces carcinogenesis in vitro via NF-κB/Twist axis. Oncol Rep. 2017;37(3):1873–1882. doi:10.3892/or.2017.5369
28. Li QQ, Xu JD, Wang WJ, et al. Twist1-mediated adriamycin-induced epithelial-mesenchymal transition relates to multidrug resistance and invasive potential in breast cancer cells. Clin Cancer Res. 2009;15(8):2657–2665. doi:10.1158/1078-0432.CCR-08-2372
29. Cui Y, Sun S, Ren K, et al. Reversal of liver cancer-associated stellate cell-induced stem-like characteristics in SMMC-7721 cells by 8-bromo-7-methoxychrysin via inhibiting STAT3 activation. Oncol Rep. 2016;35(5):2952–2962. doi:10.3892/or.2016.4637
30. Hart PC, Mao M, de Abreu AL, et al. MnSOD upregulation sustains the Warburg effect via mitochondrial ROS and AMPK-dependent signalling in cancer. Nat Commun. 2015;6:6053. doi:10.1038/ncomms7053
31. Kumar AP, Loo SY, Shin SW, et al. Manganese superoxide dismutase is a promising target for enhancing chemosensitivity of basal-like breast carcinoma. Antioxid Redox Signal. 2014;20(15):2326–2346. doi:10.1089/ars.2013.5295
32. Case AJ, Domann FE. Absence of manganese superoxide dismutase delays p53-induced tumor formation. Redox Biol. 2014;2:220–223. doi:10.1016/j.redox.2014.01.001
33. Lai Y, Yu X, Lin X, He S. Inhibition of mTOR sensitizes breast cancer stem cells to radiation-induced repression of self-renewal through the regulation of MnSOD and Akt. Int J Mol Med. 2016;37(2):369–377. doi:10.3892/ijmm.2015.2441
34. Iglesias-Bartolome R, Patel V, Cotrim A, et al. mTOR inhibition prevents epithelial stem cell senescence and protects from radiation-induced mucositis. Cell Stem Cell. 2012;11(3):401–414. doi:10.1016/j.stem.2012.06.007
35. Cao J, Wang X, Dai T, et al. Twist promotes tumor metastasis in basal-like breast cancer by transcriptionally upregulating ROR1. Theranostics. 2018;8(10):2739–2751. doi:10.7150/thno.21477
36. Shahin SA, Wang R, Simargi SI, et al. Hyaluronic acid conjugated nanoparticle delivery of siRNA against TWIST reduces tumor burden and enhances sensitivity to cisplatin in ovarian cancer. Nanomedicine. 2018;14(4):1381–1394. doi:10.1016/j.nano.2018.04.008
37. Lee TK, Poon RT, Yuen AP, et al. Twist overexpression correlates with hepatocellular carcinoma metastasis through induction of epithelial-mesenchymal transition. Clin Cancer Res. 2006;12(18):5369–5376. doi:10.1158/1078-0432.CCR-05-2722
38. Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cell movement and survival: implications in development and cancer. Development. 2005;132(14):3151–3161. doi:10.1242/dev.01907
39. Peinado H, Olmeda D, Cano A. Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nat Rev Cancer. 2007;7(6):415–428. doi:10.1038/nrc2131
40. An N, Zheng B. MiR-203a-3p inhibits pancreatic cancer cell proliferation, EMT, and apoptosis by regulating SLUG. Technol Cancer Res Treat. 2020;19:1533033819898729. doi:10.1177/1533033819898729
41. Kowalski-Chauvel A, Gouaze-Andersson V, Baricault L, et al. Alpha6-integrin regulates FGFR1 expression through the ZEB1/YAP1 transcription complex in glioblastoma stem cells resulting in enhanced proliferation and stemness. Cancers. 2019;11(3):406. doi:10.3390/cancers11030406
42. Qian J, Luo Y, Gu X, Zhan W, Wang X. Twist1 promotes gastric cancer cell proliferation through up-regulation of FoxM1. PLoS One. 2013;8(10):e77625. doi:10.1371/journal.pone.0077625
43. Zhao L, Liu L, Dong Z, Xiong J. miR-149 suppresses human non-small cell lung cancer growth and metastasis by inhibiting the FOXM1/cyclin D1/MMP2 axis. Oncol Rep. 2017;38(6):3522–3530. doi:10.3892/or.2017.6047
44. Hsieh NT, Huang CY, Li CC, Wang IC, Lee MF. MED28 and forkhead box M1 (FOXM1) mediate matrix metalloproteinase 2 (MMP2)-dependent cellular migration in human nonsmall cell lung cancer (NSCLC) cells. J Cell Physiol. 2019;234(7):11265–11275. doi:10.1002/jcp.27784
45. Liang X, Xu C, Cao X, Wang W. Isovitexin suppresses cancer stemness property and induces apoptosis of osteosarcoma cells by disruption of the DNMT1/miR-34a/Bcl-2 axis. Cancer Manag Res. 2019;11:8923–8936. doi:10.2147/CMAR.S222708
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.