Back to Journals » OncoTargets and Therapy » Volume 13
MiR-519d and miR-328-3p Combinatorially Suppress Breast Cancer Progression
Authors Ma H, Liu T, Xu Y, Wang X, Wang J, Liu X ![]()
Received 15 September 2020
Accepted for publication 8 December 2020
Published 18 December 2020 Volume 2020:13 Pages 12987—12997
DOI https://doi.org/10.2147/OTT.S281962
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr XuYu Yang
Haiming Ma,1 Tao Liu,2 Yanhua Xu,3 Xinying Wang,4 Jin Wang,1 Xiaokang Liu1
1Department of Oncology, Guangrao County People’s Hospital, Dongying City, Shandong Province 257300, People’s Republic of China; 2Department of General Surgery, Guangrao County People’s Hospital, Dongying City, Shandong Province 257300, People’s Republic of China; 3Department of Anesthesiology, Guangrao County People’s Hospital, Dongying City, Shandong Province 257300, People’s Republic of China; 4Department of Pathology, Guangrao County People’s Hospital, Dongying City, Shandong Province 257300, People’s Republic of China
Correspondence: Xiaokang Liu
Department of Oncology, Guangrao County People’s Hospital, No. 180 Huayuan Road, Guangrao County, Dongying City, Shandong Province 257300, People’s Republic of China
Tel +86-18562040137
Email [email protected]
Background: MiR-519d and miR-328-3p had tumor-regulatory properties in different cancers, but their combinatorial effects and potential common target in breast cancer had not been fully reported. This research targeted to study the underlying mechanism of how miR-519d and miR-328-3p cooperatively suppressed breast cancer.
Methods: MiR-519d and miR-328-3p expressions in breast cancer tissues and cells were assessed and Ki-67 expression was also checked. DLR assay was executed to verify whether Ki-67 was a common target of miR-519d and miR-328-3p. Western blot, flow cytometry, colony formation, wound healing and transwell assays were applied to examine the inhibitory roles of these two miRNAs on the malignant behaviors of breast cancer cells and the potential molecular mechanism.
Results: Impeded miR-519d and miR-328-3p expressions and enhanced Ki-67 expression were detected in breast cancer tissues and cells. Ki-67 was confirmed as a target of these two miRNAs. MiR-519d and miR-328-3p hampered cell proliferation and blocked cell cycle via binding to Ki-67 and they also suppressed migration and invasion. The combinatorial effects of two miRNAs were much stronger than a single miRNA.
Conclusion: Our findings proved that miR-519d and miR-328-3p played combinatorial anti-cancer roles in breast cancer by directly targeting a common target Ki-67. Our study suggested that these two miRNAs might own the potential to become novel therapeutic biomarkers involved in the diagnosis and therapy of breast cancer.
Keywords: breast cancer, miR-519d, miR-328-3p, Ki-67
Introduction
Breast cancer is the most dangerous cancerous killer of women.1 Its morbidity and mortality are rising sharply in this decade.2 In addition, with the development of modern diagnostic methods, more and more studies started focusing on the non-invasive diagnostic molecular biomarkers such as microRNAs which contributed to the metastasis of breast cancer.3 However, the survival rate of patients carrying metastatic breast cancer has dramatically increased in recent years due to a variety of intrinsic and extrinsic factors.2 Besides that, the proliferation ability and highly activated cell cycle are the crucial factors contributing to metastasis of different cancers.4 Therefore, to find the effective diagnostic and treatment method of breast cancer, there is an urgent requirement for a clearer understanding of the mechanisms associated with metastatic breast cancer at the molecular level.
MicroRNA is a type of small, non-coding single-stranded RNA molecules, predominantly bind to the 3′-UTRs of complementary sequences of mRNAs for translational repression or degradation.5 Focusing on miR-519d and miR-328-3p, on the one hand, they are found to be ectopically expressed in different cancers. For example, miR-519d6 and miR-328-3p expressions are suppressed in breast cancer.7 On the other hand, these two miRNAs affect some cellular behaviors involved in cancer progression such as cell proliferation, migration, invasion and regulation of cell cycle via targeting different molecules. For instance, miR-519d targets STAT3 and inhibits migration, invasion, proliferation, and survival of breast cancer cells.8 MiR-328-3p targets GRM4 and suppresses metastasis of breast cancer.9 Therefore, more targets of miR-519d and miR-328-3p needed to be identified and whether they could target a common target in the mediation of breast cancer need to be studied.
Antigen Ki-67 exists in G1, S, G2 and mitosis phases of the cell cycle which is also related to cell proliferation.10 Moreover, it is a predictive biomarker involved in breast cancer diagnosis.11 Different studies have reported the clinical and prognostic roles of Ki-67 in different types of cancers such as gastric cancer12 and breast cancer.13 Besides, Ki-67 expression can be mediated by specific miRNA–mRNA interactions.14 A study has indicated that miR-519d targets Ki-67 to suppress the growth of liver cancer cells.15 Therefore, not only is the dysregulation of Ki-67 associated with cancer prognosis, but also it is integrated with some potential mechanisms involved in proliferation and cell cycle of cancer progression.
Currently, few pieces of evidences can build up the connection between miR-519d, miR-328-3p and Ki-67 in breast cancer and clarify the underlying targeting relationship and action mechanism between them. Therefore, this study aimed to find the potential mechanism of miR-519d, miR-328-3p and Ki-67 in the inhibition of breast cancer.
Materials and Methods
Cancer Tissue Samples
The breast cancer tissues and corresponding tumor-adjacent normal tissues were resected from 10 patients at our department and the samples were histopathologically verified. The patients had not received any therapy before sample collection. The fresh tissues were quickly frozen in liquid nitrogen until use. The informed written consents were signed by all subjects. The protocol of this research was approved by the Ethical Committee of Guangrao County People’s Hospital.
Immunohistochemistry Assay
The specimens were fixed using 10% formalin (Solarbio, China) and embedded in the paraffin wax (Solarbio, China) followed by slicing. After deparaffinization using xylene (Solarbio, China) and rehydration by ethanol (Solarbio, China). The primary antibody against Ki-67 (Beyotime, China) was applied and incubated at 4°C for 16 h. The DAB Histostaining kit (Solarbio, China) was selected to localize Ki-67 in clinical tissues.
Cell Culture
The breast cancer cell lines (MDA-MB-231, MCF-7 and SK-BR-3) and normal breast epithelial cell line MCF-10A were obtained from the Cell Bank of the Chinese Academy of Sciences and cultivated according to the instructions. In detail, MDA-MB-231 was inoculated in Leibovitz’s L-15 (Procell, China) containing 10% FBS (Procell, China) with 1% penicillin and streptomycin (P/S) (Procell, China). MCF-7 was cultivated in MEM (Procell, China) with 0.01mg/mL insulin (Procell, China), 10% FBS and 1% P/S. SK-BR-3 was maintained in McCoy’s 5A (Procell, China) with 10% FBS and 1% P/S whereas MCF-10A was maintained in DMEM/F12 (Procell, China) with 10% FBS, 0.01mg/mL insulin with 10% FBS and 1% P/S. MCF-7, SK-BR-3 and MCF-10A cell lines were kept in a humidified incubator (Herocell 180, Radobio, China) supplemented with 5% CO2 at 37°C while MDA-MB-231 was cultured in 100% air at 37°C.
Transfection
The cultivated cells were seeded into 6-well plates of 2×105 cells/well and incubated for 12 h. At 70–80% confluence, the cells were divided into 5 groups (MCF-7: blank, mimic control, miR-519d mimic, inhibitor control and miR-519d inhibitor; MDA-MB-231: blank, mimic control, miR-328-3p mimic, inhibitor control and miR-328-3p inhibitor) and the HiPerFect Transfection Reagent (Qiagen, USA) was selected to complete the transfection process. Briefly, 150 ng miRNA oligonucleotides and 4 μL reagent were added into 100 μL RPMI-1640 and mixed well. The mixture was then transferred into the cell culture medium. 48 h after transfection, cells were collected for the subsequent detections.
The 70 bp Ki-67 cDNA of the 3ʹUTR region with the whole coding sequence was constructed into the pcDNA3.1 vector (ThermoFisher, USA). The breast cancer cell lines were transfected with pcDNA3.1 vector containing Ki-67 or empty vector via Lipofectamine 3000 as purchased from the supplier (ThermoFisher, USA). 48 h after incubation, the cells were harvested and sent to subsequent experiments.
qRT-PCR
The RNAeasy Animal RNA Isolation kit (Beyotime, China) was used to extract total RNA. cDNA was converted from mature miRNAs through the application of Hifair cDNA Synthesis SuperMix kit (Yeasen, China). To examine the expression of miR-519d, miR-328-3p and Ki-67, qRT-PCR was executed on QuantStudio 7 Flex (ThermoFisher, USA) using the BeyoFast SYBR Green qPCR Mix kit (Beyotime, China). The primers are summarized in Table 1. Quantification of RNA levels was normalized with U6 or GAPDH using the 2−ΔΔCt cycle threshold method.
 |
Table 1 Primers Used for qRT-PCR |
Western Blot
The total proteins from the cultured cells were extracted with the Lysis Buffer (Beyotime, China). After that, a bicinchoninic acid protein assay kit (Beyotime, China) was applied to evaluate the protein concentration. After separation by 10% SDS-PAGE, the samples were transferred onto the PVDF membrane (Merck Millipore, Germany). 5% non-fat milk was applied to block the unbinding sites of the membrane for 2 h and the primary rabbit antibodies against Ki-67 (1:1000), PHH3 (1:100), cyclin D1 (1:500), pRb (1:1000), ppRb (1:500), E2F1 (1:500), α-tubulin (1:500) and GAPDH (1:1000) (Beyotime, China) were added and incubated for 12 h at 4°C. The blots were rinsed and then the horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody (1:2000; Beyotime, China) was applied and incubated for 1 h. Next, after rinsing, the bands on the membrane were stained by the chemiluminescence kit (Beyotime, China) and visualized. The intensity was quantified using ImageJ 1.53b software (NIH, USA).
BrdU Incorporative Assay
The cells were treated with 100 μM BrdU for 0.5 h and fixed with 4% formaldehyde for 10 min, followed by denaturation neutralization for 20 min. After that, 5% bovine serum albumin was added for blocking for 0.5 h and the cells were stained using rabbit anti-BrdU (1:1000) antibodies (Beyotime, China) for 1 h. Next, the secondary anti-rabbit antibody conjugated to Alexa Fluor 555 (Beyotime, China) was applied and the cells were kept for an extra 1 h. Finally, the cells were mounted with Vectashield mounting reagent (Vector, USA) containing DAPI that stained the nucleus and observed under the microscope (Olympus CKX53, Japan).
DLR Assay
The wild type and mutated sequences of 3ʹ-UTR fragment of Ki-67 containing the binding sites of miR-519d and miR-328-3p were synthesized from cDNA using ProFlex PCR (ThermoFisher, USA) and inserted into pmiRGLO vectors (Promega, USA) to create the wild-type (pmiRGLO-Ki-67-wt) and mutant (pmiRGLO-Ki-67-mt) plasmids. The cells were co-transfected when the confluence reached 80% with 10 pmol miR-519d mimic or miR-328-3p or mimic control and 100 ng of wt/mt 3ʹ-UTR of Ki-67 pmirGLO constructs using Lipofectamine (Invitrogen, USA). 48 h after transfection, the luciferase activity which represented the influence of miR-519d or miR-328-3p on Ki-67 was measured using the Dual-Luciferase Reporter Gene Assay Kit (Yeasen, China) and GloMax 20/20 Luminometer (Promega, USA). The luminescence activity of the firefly luciferase substrate was normalized to the Renilla luciferase substrate.
Flow Cytometry
Cells were harvested 48 h after transfection and rinsed three times with PBS followed by fixation with 75% ethanol at 4°C for 12 h. After that, the cells were resuspended in PBS with RNase A (Beyotime, China) and incubated with propidium iodide (Beyotime, China) for 1 h. The cell cycle analysis was conducted by Cytoflex S (Beckman Coulter, USA).
Colony Formation Assay
48h after transfection, the cells were inoculated in 6-well plates of 1000 cells/well and kept at 37°C for 14 days. The medium was discarded and refreshed every 3 days. On day 14, after rinsed three times, the cells were fixed with 4% polymerized formaldehyde (Beyotime, China) for 0.5 h and stained with 2.5% crystal violet (Beyotime, China) for another 0.5 h. The 6-well plates were rinsed three times by PBS and set to be air-dried. The colonies from each well were counted via microscopy (Olympus CKX53, Japan).
Wound Healing Migration Assay
The transfected cells were transferred to the 6-well plates. After confluence reaching 80%, the cell layer was scratched using a 10 μL pipette tip to make the wound. The supernatant was removed and the corresponding fresh medium was added and the cells were continually incubated. The images were captured at 0 h and 48 h by microscopy (CKX53, Olympus, Japan). The relative migration rate was calculated by (original wound area/remaining wound area) *100%.
Transwell Invasion Assay
The cells were seeded into the top of the 24-well chamber with 8 μm pore size precoated with 80 μL Matrigel (BD, China). The lower chamber containing 800 µL corresponding medium mentioned in the cell lines and culture section above was filled with 10% FBS. After cultured for 24 h, the cotton swab was applied to erase the cells on the upper surface of the chamber, and the cells invaded into the lower surface were fixed by 4% polyformaldehyde fix solution (Beyotime, China). Then, 0.5% crystal violet (Beyotime, China) was added to stain the cells and the numbers were counted by microscopy (Olympus CKX53, Japan).
Statistical Analysis
The experiments were independently conducted in triplicate and repeated three times. Data were expressed as mean ± SD. Statistical analyses were conducted using SPSS 26.0 (SPSS Inc, USA) and Graphpad Prism 8.4.2 (Graphpad LLC, USA). Student’s t-test and one-way ANOVA test followed by post hoc analysis were conducted to determine the statistical differences between groups under different conditions. P<0.05 was considered a statistically significant difference. All graphs were plotted using Graphpad Prism 8.0.2 (GraphPad, USA) and Adobe Photoshop CS6 13.0.1 (Adobe Systems Incorporated, USA).
Results
The Low Expressions of miR-519d, miR-328-3p and High Expression of Ki-67 in Breast Cancer
At first, qRT-PCR was executed to evaluate the expressions of miR-519d and miR-328-3p in clinical tissues. The results revealed that both of their expressions were lower in cancerous tissues (Figure 1A). Besides, similar results of qRT-PCR were obtained in the cultivated cells. The expressions of miR-519d and miR-328-3p were lower in three breast cancer cell lines compared with the normal cell line (Figure 1B). After that, the expression of Ki-67 in tissues and cultivated cells was subsequently determined. The results indicated that Ki-67 was highly expressed in breast cancer tissues (Figure 1C–E) and three cell lines (Figure 1F and G) at both mRNA and protein levels compared with the normal tissues and the normal cell line. These results summarized that the expressions of miR-519d and miR-328-3p were decreased whereas Ki-67 expression was strengthened in breast cancer.
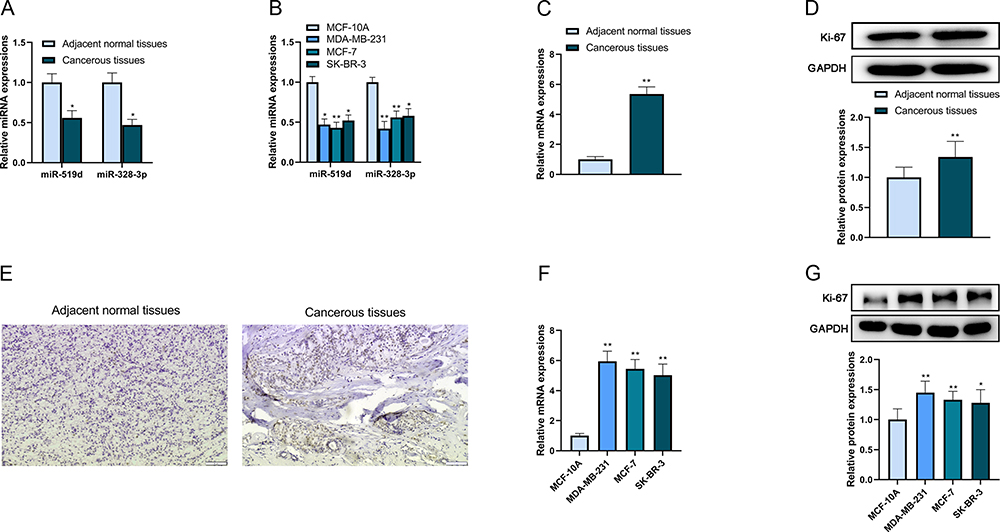 |
Figure 1 The low expressions of miR-519d, miR-328-3p and high expression of Ki-67 in breast cancer. (A) The expression levels of miR-519d and miR-328-3p in clinical tissues were analyzed by qRT-PCR. (B) The expression levels of miR-519d and miR-328-3p in cultivated cell lines were analyzed by qRT-PCR. (C) The mRNA expression level of Ki-67 in clinical tissues was analyzed by qRT-PCR. (D) The protein expression level of Ki-67 in clinical tissue (representative) was analyzed by Western blot. (E) The protein expression of Ki-67 in clinical tissue (representative) was analyzed by immunohistochemistry. Scale bar: 50 μm. (F) The mRNA expression level of Ki-67 in cultivated cell lines was analyzed by qRT-PCR. (G) The protein expression level of Ki-67 in cultivated cell lines was analyzed by Western blot. *P < 0.05 and **P < 0.01. |
miR-519d and miR-328-3p Inhibited the Cancerous Behaviors of Breast Cancer Cells
After the transfection of MCF-7 (blank, mimic control and miR-519d mimic) and MDA-MB-231 (blank, mimic control and miR-328-3p mimic) cell lines, subsequent experiments were conducted to check the roles of miR-519d and miR-328-3p on the cellular behaviors of breast cancer cells. The transfection efficiency was checked by qRT-PCR. The MCF-7 cells transfected with miR-519d mimic expressed higher levels of miR-519d compared with the blank and mimic control group. Similar results were found in MDA-MB-231 cells after miR-328-3p transfection (Figure 2A). Then, it was also observed by flow cytometry that the proportion of cells in the G1 phase was upregulated while the number of cells in the S phase was downregulated after miR-519d or miR-328-3p mimic transfection (Figure 2B). The overexpression of miR-519d or miR-328-3p suppressed cell proliferation which was revealed by the results of BrdU incorporative assay (Figure 2C) and colony formation assay (Figure 2D). After that, a downregulation of PHH3 expression level was shown by Western blot, because PHH3 only existed in the mitosis of cells, which could better represent the proliferation ability (Figure 2E). And miR-519d and miR-328-3p could also cause a decrease in the abilities of cell migration (Figure 2F) and invasion (Figure 2G). These results showed that miR-519d and miR-328-3p triggered the arrest of the cell cycle and inhibited cancerous behaviors in MCF-7 and MDA-MB-231 cell lines.
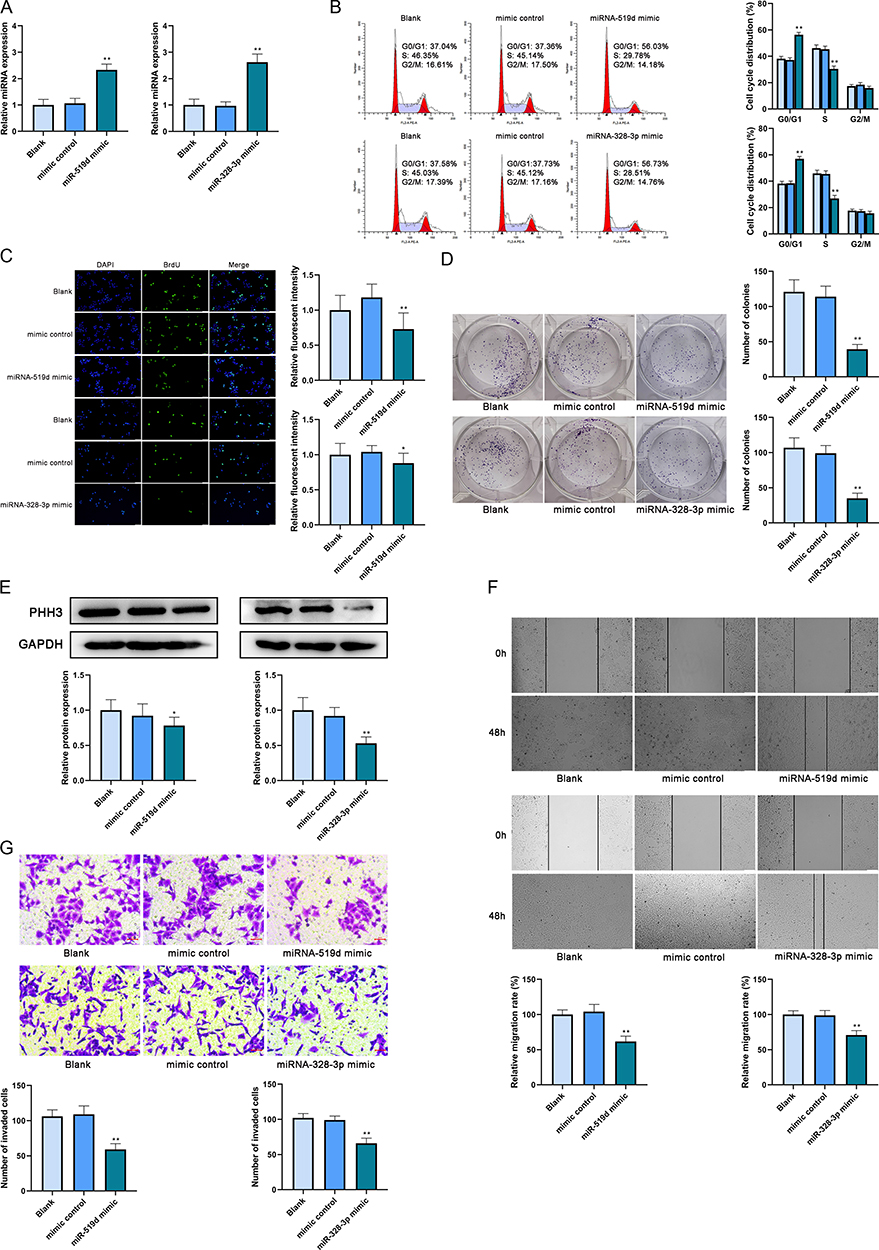 |
Figure 2 MiR-519d and miR-328-3p inhibited the cancerous behaviors of breast cancer cells. (A) The transfection efficiency was analyzed by qRT-PCR. (B) The cell cycle after transfection was analyzed by flow cytometry. (C) The proliferation of cells after transfection was analyzed by BrdU incorporative assay. Scale bar: 50 μm. (D) The proliferation of cells after transfection was analyzed by colony formation assay. (E) The protein expression level of PHH3 was analyzed by Western blot. (F) The migration of cells after transfection was analyzed by wound-healing assay. Scale bar: 100 μm. (G) The invasion of cells after transfection was analyzed by transwell assay. Scale bar: 5 μm. *P < 0.05 and **P < 0.01. |
Ki-67 Was a Target of miR-519d and miR-328-3p
Ki-67 was predicted as a target of miR-519d and miR-328-3p by ENCORI (Figure 3A). After that, the MCF-7 cell line was divided into 4 groups and co-transfected, respectively, by miR-519d mimic + pmirGLO-Ki-67-wt/pmirGLO-Ki-67-mt or miR-control + pmirGLO-Ki-67-wt/pmirGLO-Ki-67-mt. The MDA-MB-231 cell line was also divided into 4 groups and co-transfected, respectively, by miR-328-3p mimic + pmirGLO-Ki-67-wt/pmirGLO-Ki-67-mt or miR-control + pmirGLO-Ki-67-wt/pmirGLO-Ki-67-mt. Then the results of DLR assay showed that the relative luciferase activity in the MCF-7 cells transfected with miR-519d mimic + pmirGLO-Ki-67-wt reduced significantly compared with the other groups and similar outcomes were detected in MDA-MB-231 cells which reported that the relative luciferase activity in the miR-328-3p mimic + pmirGLO-Ki-67-wt reduced significantly (Figure 3B). These results confirmed that Ki-67 was a target of miR-519d and miR-328-3p.
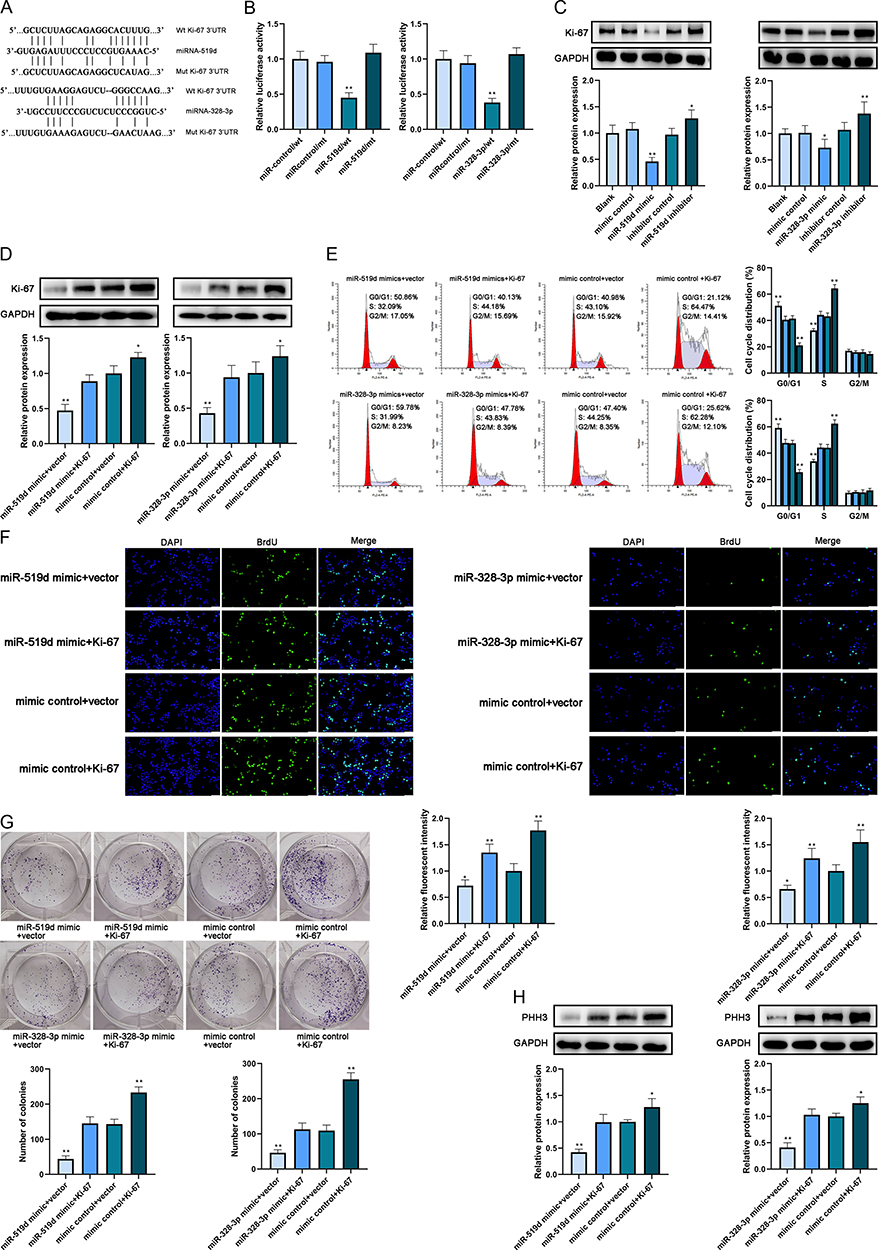 |
Figure 3 MiR-519d and miR-328-3p targeted Ki-67 individually to mediate the proliferation of breast cancer cells. (A) The binding site of miR-519d and miR-328-3p on Ki-67 was predicted by ENCORI. (B) The binding relationship was verified by dual-luciferase reporter assay. (C) The protein expression level of Ki-67 in cultivated cell lines after transfection was analyzed by Western blot. (D) The protein expression level of Ki-67 in cultivated cell lines after transfection was analyzed by Western blot. (E) The cell cycle after transfection was analyzed by flow cytometry. (F) The proliferation of cells after transfection was analyzed by BrdU incorporative assay. Scale bar: 50 μm. (G) The proliferation of cells after transfection was analyzed by colony formation assay. (H) The protein expression level of PHH3 was analyzed by Western blot. *P < 0.05 and **P < 0.01. |
After that, Western blot was conducted after transfection of MCF-7 (blank, mimic control, miR-519d mimic, inhibitor control and miR-519d inhibitor) and MDA-MB-231 cell lines (blank, mimic control, miR-328-3p mimic, inhibitor control and miR-328-3p inhibitor) and the outcomes pointed out that miR-519d and miR-328-3p diminished the expression of Ki-67 expression in two cell lines, respectively (Figure 3C). The use of miR-519d inhibitor and miR-328-3p inhibitor could increase the Ki-67 expression. These results above proved that Ki-67 was a target of miR-519d and miR-328-3p.
MiR-519d and miR-328-3p Targeted Ki-67 Individually to Mediate the Proliferation of Breast Cancer Cells
To verify whether miR-519d and miR-328-3p affected the cell cycle and proliferation of MCF-7 and MDA-MB-231 cell lines, respectively, by targeting Ki-67, the cells were divided into four groups (MCF7: miR-519d mimic+empty vector, miR-519d mimic+Ki-67, mimic control+empty vector, mimic control+Ki-67; MDA-MB-231: miR-328-3p mimic+empty vector, miR-328-3p mimic+Ki-67, mimic control+empty vector, mimic control+Ki-67) and transfected, respectively. Western blot was executed after transfection to check the protein expression of Ki-67 and the results indicated that mimic control+Ki-67 group in both cell lines expressed the highest level of Ki-67 while the miR-519d mimic+empty vector and miR-328-3p mimic+empty vector expressed the lowest level of Ki-67 in these two co-transfected cell lines, respectively (Figure 3D). These results proved that the overexpression of miR-519d and miR-328-3p could inhibit the endogenous and ectopic expressions of Ki67.
Then, to check the roles of miR-519d and miR-328-3p on cell cycle and proliferation via targeting Ki-67 in these four groups of two cell lines above, flow cytometry, BrdU incorporative assay and colony formation assays were conducted. It was reported by the results of flow cytometry that the percentage of cells in the G1 phase accrued in the group transfected with miRNA+empty vector and decreased in the group transfected with mimic control+Ki-67. And the proportion of cells in the S phase showed opposite results in these four groups of both cell lines (Figure 3E). Furthermore, the results of BrdU incorporative assay (Figure 3F), colony formation assay (Figure 3G) and PHH3 expression detected by Western blot (Figure 3H) in these two cell lines also revealed that the group co-transfected with miR-519d+empty vector and miR-328-3p showed lower DNA synthetic ability and proliferative potential whereas the group co-transfected with mimic control+Ki-67 showed stronger DNA synthesis and proliferation compared with the other groups. To summarize, overexpression of miR-519d impeded the malignant behaviors of the MCF-7 cell line and transfection of miR-328-3p also exerted the anti-tumor effects in the MDA-MB-231 cell line.
Co-Transfection and the Combinatorial Effects of miR-519d and miR-328-3p
Ki-67 had been identified to be inhibited by miR-519d as well as miR-328-3p in MCF-7 and MDA-MB-231 cells, respectively, and individually. Therefore, it is postulated that whether a synergistic regulation of Ki-67 by these two miRNAs in breast cancer cells existed. To verify the combined tumor-suppressive roles of both miR-519d and miR-328-3p in one breast cancer cell line, MDA-MB-231 cells were transfected by miR-519d or miR-328-3p individually or co-transfected by miR-519d and miR-328-3p (blank, mimic control, miRNA-519d mimic, miRNA-328-3p mimic, miRNA-519d mimic + miRNA-328-3p mimic) and Western blot was executed to examine the expression of Ki-67 protein in cells. The results indicated that the Ki-67 protein expression was the lowest in the group co-transfected by miRNA-519d mimic + miRNA-328-3p mimic compared with the other groups (Figure 4A). After that, a series of assays were performed to examine the cellular effect of the co-transfection of two miRNAs in one breast cancer cell line. The outcomes of flow cytometry reported that the proportion of cells in the G1 phase increased in the miRNA-519d mimic + miRNA-328-3p mimic group compared with miRNA-519d mimic group and miRNA-328-3p mimic group (Figure 4B). In addition to this, cell cycle regulatory protein cyclin D1 could accelerate cancer cell cycle G1-S transition and retinoblastoma protein (pRb) is a tumor suppressor protein and its hyperphosphorylated (inactivated) form ppRb could trigger the release of transcription factor E2F1 to enhance G1-S transition.16 Overexpression of these two miRNAs also significantly decreased the protein expression levels of cyclin D1, ppRb and E2F1 (Figure 4C) which indicated that the co-transfection of these two miRNAs dominantly arrested the cell cycle at G1-S transition and the impact of co-transfection of two miRNAs was much stronger than the transfection of a single miRNA. Cell cycle was associated with proliferation, and the lowest proliferative ability was also found in the same group after BrdU incorporative assay (Figure 4D), colony formation assay (Figure 4E) and PHH3 expression detected by WB (Figure 4F). Then the results of wound healing (Figure 4G) and transwell assay (Figure 4H) also proved that the group co-transfected with two miRNAs showed stronger inhibitory effects on migration and invasion of breast cancer cells compared with the blank group and the groups transfected by the single miRNA. To conclude, in the MDA-MB-231 cell line, overexpression of miR-328-3p exerted stronger inhibitory effects on the Ki-67 expression, cell proliferation, G1-S transition, migration and invasion than miR-519d. The transfection of two miRNAs suppressed the malignant behaviors of breast cancer cells more effectively through targeting Ki-67 than the transfection of a single miRNA.
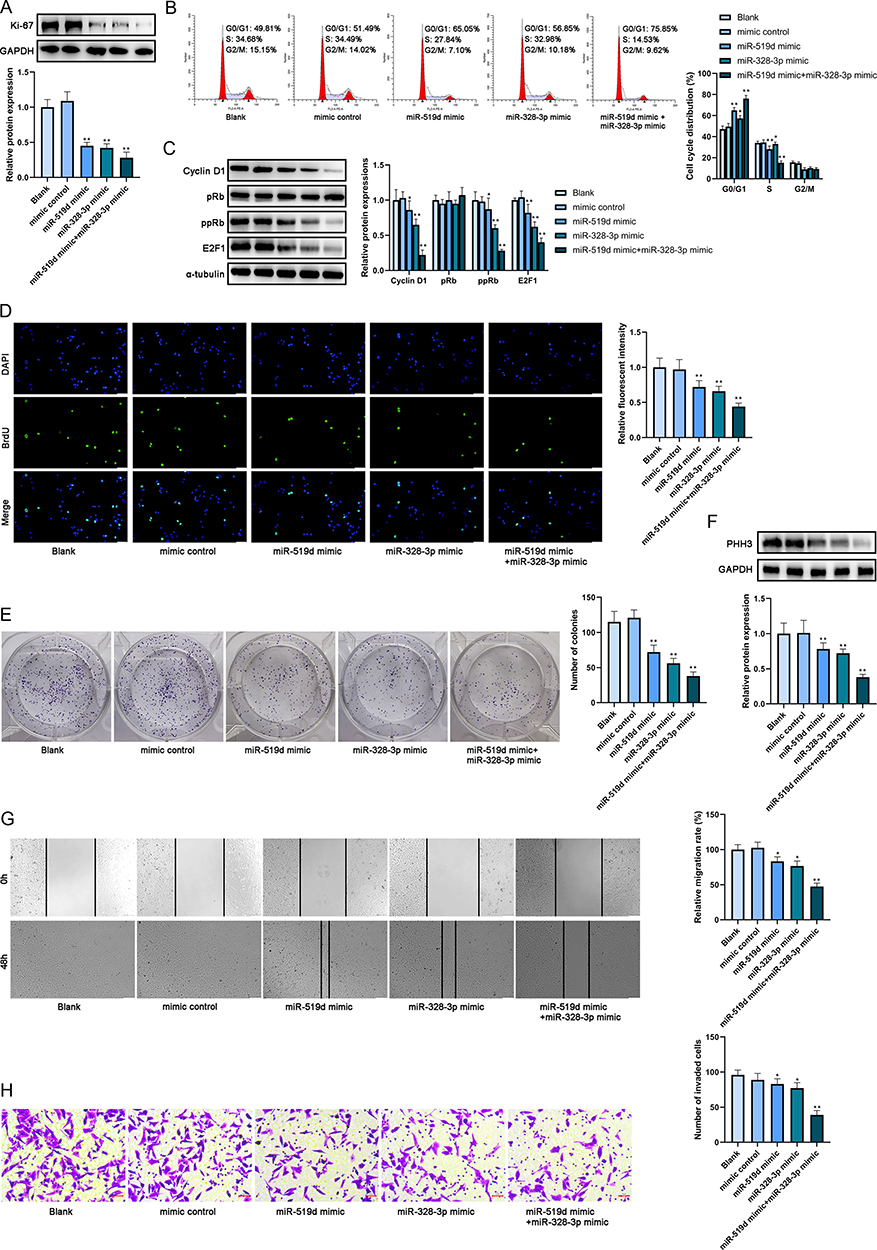 |
Figure 4 Co-transfection and the combinatorial effects of miR-519d and miR-328-3p. (A) The protein expression level of Ki-67 in cultivated cell lines after transfection was analyzed by Western blot. (B) The cell cycle after transfection was analyzed by flow cytometry. (C) The protein expression levels of cyclin D1, pRb, ppRb and E2F1 in cultivated cell lines after transfection were analyzed by Western blot. (D) The proliferation of cells after transfection was analyzed by BrdU incorporative assay. Scale bar: 100 μm. (E) The proliferation of cells after transfection was analyzed by colony formation assay. (F) The protein expression level of PHH3 was analyzed by Western blot. (G) The migration of cells after transfection was analyzed by wound-healing assay. Scale bar: 100 μm. (H) The invasion of cells after transfection was analyzed by transwell assay. Scale bar: 5 μm. *P < 0.05 and **P < 0.01. |
Discussion
In this research, the roles of miRNA-519d and miRNA-328-3p in breast cancer cells were studied. In clinical breast cancer tissues and cultured cells, low expressions of these two miRNAs were detected by qRT-PCR. Both of them showed their function to suppress cancerous behaviors of breast cancer cells. Meanwhile, the enhanced expression of Ki-67 was found in breast cancer tissues and cell lines in comparison to normal tissues and the normal breast epithelial cell line and Ki-67 was verified as a target of miRNA-519d and miRNA-328-3p. Last but not least, miRNA-519d and miRNA-328-3p can combinatorially regulate the proliferation and cell cycle of breast cancer cells to show anti-cancer effects through directly targeting Ki-67.
Up to date, although breast cancer was one of the most studied cancer, there was still abundant hidden molecular treasure required to be excavated. Furthermore, the diagnostic approaches of breast cancer relied on the patients’ serum level of biological molecules that were not developed satisfactorily and the lack of biomarkers and molecular drugs were crucial issues of the diagnosis and treatment of breast cancer. Ki-67 was one of the representative biomarkers with the proto-oncogenic and proliferative roles and overexpression of Ki-67 were detected in different cancers such as lung cancer17 and breast cancer.18 The immunohistochemistry method was broadly applied in the diagnosis of breast cancer and Ki-67 positive cells were often associated with the clinical stages of breast cancer.11 Although the overexpression of Ki-67 was correlated with cancer metastasis and survival rate and it also contributed to the proliferation of breast cancer cells and lymph node metastasis,19 the actual roles of Ki-67 were still poorly understood and there have been only a few published studies that correlate the expression of Ki-67 with miRNA targeting and regulation.14
MiRNAs were small non-coding RNAs that endogenously mediated the expression and function of target mRNAs and they had become a hot research spot in this decade. The aberrant expressions of miRNAs were discovered to be linked to breast cancer such as miR-19a20 and miR-190.21 Different sophisticated databases could be used to predict the target of different miRNAs in combination with a variety of biological techniques to confirm these predictions and verify their effective roles in different cancers. On the one hand, focused on miR-519d, it had already shown its function as a tumor suppressor gene through the regulation of different targets in breast cancer. For instance, it suppressed the cancerous behaviors of breast cancer cells via targeting MMP3.6 It also inhibited the metastasis and progression of breast cancer through binding to LIMK1.22 Moreover, Hou’s research had already indicated that miR-519d could bind to Ki-67 and involved in liver cancer suppression which was consistent with our results conducted in breast cancer cells.15 On the other hand, focused on miR-328-3p, it owned tumor-suppressive function in breast cancer9 and osteosarcoma23 via targeting different molecules. In this research, we found that in breast cancer cells, miR-519d and miR-328-3p directly targeted Ki-67 to reduce its expression to suppress proliferation and block cell cycle progression and the anti-cancer effects of miR-519d were weaker than miR-328-3p which filled the research blank. Interestingly, because Ki-67 is a proliferative biomarker, the underlying reason for how these two miRNAs controlled cell migration and invasion needed to be further studied.
In conclusion, we presented in this research for the first time that miR-519d and miR-328-3p were novel regulators of Ki-67 which exerted anti-cancer effects combinatorially in breast cancer. Our data provided a tentative method for the diagnosis and therapy of breast cancer and indicated a rational direction for further research.
Abbreviations
miRNA, microRNA; 3′-UTRs, 3′-untranslated regions; mRNA, messenger RNA; OSCC, oral squamous cell carcinoma; STAT3, signal transducer and activator of transcription 3; GRM4, glutamate metabotropic receptor 4; DLR, dual-luciferase reporter; PHH3, phosphohistone H3; MMP3, matrix metalloproteinase 3; LIMK1, LIM domain kinase 1.
Data Sharing Statement
The analyzed data sets generated during the present study are available from the corresponding author on reasonable request.
Ethics Approval
The methodologies of this research were approved by the Ethics Committee of Guangrao County People’s Hospital and obeyed the principles of the Declaration of Helsinki.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This work was supported by China Medical Hand-in-hand Project Committee and Beijing Medical Association Award Foundation (No. YXJL2015-138).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Ji F, Yang CQ, Li XL, et al. Risk of breast cancer-related death in women with a prior cancer. Aging. 2020;12(7):5894–5906. doi:10.18632/aging.102984
2. Youn HJ, Han WA. Review of the epidemiology of breast cancer in Asia: focus on risk factors. Asian Pac J Cancer Prev. 2020;21(4):867–880. doi:10.31557/APJCP.2020.21.4.867
3. Zografos E, Zagouri F, Kalapanida D, et al. Prognostic role of microRNAs in breast cancer: a systematic review. Oncotarget. 2019;10(67):7156–7178.
4. Urooj T, Wasim B, Mushtaq S, Shah SNN, Shah M. Cancer cell-derived secretory factors in breast cancer-associated lung metastasis: their mechanism and future prospects. Curr Cancer Drug Targets. 2020;20(3):168–186. doi:10.2174/1568009620666191220151856
5. Alitheen NBM, Osman MA. The regulatory role of MicroRNAs in breast cancer. Int J Mol Sci. 2019.
6. Chu C, Liu X, Bai X, et al. MiR-519d suppresses breast cancer tumorigenesis and metastasis via targeting MMP3. Int J Biol Sci. 2018;14(2):228–236. doi:10.7150/ijbs.22849
7. Liu T, Ye P, Ye Y, Lu S, Han B. Circular RNA hsa_circRNA_002178 silencing retards breast cancer progression via microRNA-328-3p-mediated inhibition of COL1A1. J Cell Mol Med. 2020;24(3):2189–2201. doi:10.1111/jcmm.14875
8. Deng X, Zhao Y, Wang B. miR-519d-mediated downregulation of STAT3 suppresses breast cancer progression. Oncol Rep. 2015;34(4):2188–2194. doi:10.3892/or.2015.4160
9. Xiao B, Chen D, Zhou Q, et al. Glutamate metabotropic receptor 4 (GRM4) inhibits cell proliferation, migration and invasion in breast cancer and is regulated by miR-328-3p and miR-370-3p. BMC Cancer. 2019;19(1):891. doi:10.1186/s12885-019-6068-4
10. Chen WJ, He DS, Tang RX, Ren FH, Chen G. Ki-67 is a valuable prognostic factor in gliomas: evidence from a systematic review and meta-analysis. Asian Pac J Cancer Prev. 2015;16(2):411–420. doi:10.7314/APJCP.2015.16.2.411
11. Kurbel S, Dmitrović B, Marjanović K, Vrbanec D, Juretić A. Distribution of Ki-67 values within HER2 & ER/PgR expression variants of ductal breast cancers as a potential link between IHC features and breast cancer biology. BMC Cancer. 2017;17(1):231. doi:10.1186/s12885-017-3212-x
12. Xiong DD, Zeng CM, Jiang L, Luo DZ, Chen G. Ki-67/MKI67 as a predictive biomarker for clinical outcome in gastric cancer patients: an updated meta-analysis and systematic review involving 53 studies and 7078 patients. J Cancer. 2019;10(22):5339–5354. doi:10.7150/jca.30074
13. Zhu X, Chen L, Huang B, et al. The prognostic and predictive potential of Ki-67 in triple-negative breast cancer. Sci Rep. 2020;10(1):225.
14. Liu Y, Tang K, Yan W, et al. Identifying Ki-67 specific miRNA-mRNA interactions in malignant astrocytomas. Neurosci Lett. 2013;546:36–41. doi:10.1016/j.neulet.2013.04.030
15. Hou YY, Cao WW, Li L, et al. MicroRNA-519d targets MKi67 and suppresses cell growth in the hepatocellular carcinoma cell line QGY-7703. Cancer Lett. 2011;307(2):182–190. doi:10.1016/j.canlet.2011.04.002
16. Ahlin C, Lundgren C, Embretsén-Varro E, Jirström K, Blomqvist C, Fjällskog ML. High expression of cyclin D1 is associated to high proliferation rate and increased risk of mortality in women with ER-positive but not in ER-negative breast cancers. Breast Cancer Res Treat. 2017;164(3):667–678. doi:10.1007/s10549-017-4294-5
17. Wei DM, Chen WJ, Meng RM, et al. Augmented expression of Ki-67 is correlated with clinicopathological characteristics and prognosis for lung cancer patients: an up-dated systematic review and meta-analysis with 108 studies and 14,732 patients. Respir Res. 2018;19(1):150. doi:10.1186/s12931-018-0843-7
18. Ishibashi N, Nishimaki H, Maebayashi T, et al. Changes in the Ki-67 labeling index between primary breast cancer and metachronous metastatic axillary lymph node: a retrospective observational study. Thorac Cancer. 2019;10(1):96–102. doi:10.1111/1759-7714.12907
19. Aziz S, Wik E, Knutsvik G, et al. Evaluation of tumor cell proliferation by Ki-67 expression and mitotic count in lymph node metastases from breast cancer. PLoS One. 2016;11(3):e0150979. doi:10.1371/journal.pone.0150979
20. Anfossi S, Giordano A, Gao H, et al. High serum miR-19a levels are associated with inflammatory breast cancer and are predictive of favorable clinical outcome in patients with metastatic HER2+ inflammatory breast cancer. PLoS One. 2014;9(1):e83113. doi:10.1371/journal.pone.0083113
21. Yu Y, Luo W, Yang ZJ, et al. miR-190 suppresses breast cancer metastasis by regulation of TGF-β-induced epithelial-mesenchymal transition. Mol Cancer. 2018;17(1):70. doi:10.1186/s12943-018-0818-9
22. Li D, Song H, Wu T, et al. MiR-519d-3p suppresses breast cancer cell growth and motility via targeting LIM domain kinase 1. Mol Cell Biochem. 2018;444(1–2):169–178. doi:10.1007/s11010-017-3241-4
23. Shi J, An G, Guan Y, et al. miR-328-3p mediates the anti-tumor effect in osteosarcoma via directly targeting MMP-16. Cancer Cell Int. 2019;19(1):104. doi:10.1186/s12935-019-0829-7
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.