Back to Journals » Cancer Management and Research » Volume 10
miR-300 rs12894467 polymorphism may be associated with susceptibility to primary lung cancer in the Chinese Han population
Authors Liu Z ![]() , Lin Y, Kang S, Xu Q, Xiong W, Cai L
, Lin Y, Kang S, Xu Q, Xiong W, Cai L ![]() , He F
, He F
Received 27 April 2018
Accepted for publication 30 June 2018
Published 17 September 2018 Volume 2018:10 Pages 3579—3588
DOI https://doi.org/10.2147/CMAR.S172514
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Zhiqiang Liu,1,2 Yong Lin,3 Shuling Kang,4 Qiuping Xu,2 Weimin Xiong,2 Lin Cai,2 Fei He2,5
1The United Innovation of Mengchao Hepatobiliary Technology Key Laboratory of Fujian Province, Mengchao Hepatobiliary Hospital of Fujian Medical University, Fuzhou, Fujian, China; 2Department of Epidemiology and Health Statistics, School of Public Health, Fujian Medical University, Fuzhou, Fujian, China; 3Clinical Laboratory, Zhongshan Hospital, Medical College of Xiamen University, Xiamen, Fujian, China; 4Fuzhou Center for Disease Control and Prevention, Fuzhou, Fujian, China; 5Key Laboratory of Ministry of Education for Gastrointestinal Cancer, Fujian Medical University, Fuzhou, Fujian, China
Objective: The etiology of lung cancer has been attributed to both environmental and genetic factors. In this study, we investigated the association between five miRNA gene single-nucleotide polymorphisms (SNPs) and the risk of lung cancer, and explored the interaction between genetic and environmental factors in the Han people of China, the ethnic group that represents >90% of the population of the country.
Methods: This case–control study included 1,067 cases and 1,085 controls. Epidemiological data were collected by in-person interviews using a standard questionnaire. Matrix-assisted laser desorption/ionization time of flight mass spectrometry was applied to genotype the selected miRNA gene SNPs. Unconditional logistic regression and stratified analysis were used to analyze the associations between these SNPs and lung cancer, and to calculate the adjusted odds ratios (ORs) and 95% confidence intervals (CIs). Crossover analysis, logistic regression, and the Excel table made by Andersson were used to analyze the combined and interaction effects of gene–environment.
Results: The rs12894467 CC/CT genotype was associated with a significantly increased risk for lung cancer in women (adjusted OR =1.46, 95% CI=1.01–2.10). Smokers carrying the CC/CT genotype were associated with a significantly decreased risk of lung cancer, the adjusted OR was 0.75 (95% CI: 0.57–0.98). In the dominant model, rs12894467 and gender were associated with a positive multiplicative interaction; rs12894467 and smoking were associated with a negative multiplicative interaction.
Conclusion: The rs12894467 polymorphism was potentially associated with primary lung cancer in the Han Chinese population and had an interactive relationship with environmental factors.
Keywords: microRNAs, single-nucleotide polymorphism, lung cancer, case–control studies, susceptibility
Introduction
Of the malignant tumors, lung cancers are currently the greatest threat to human health. In males, the worldwide age-standardized morbidity and mortality rates for lung cancer are 34.2/100,000 and 30.0/100,000; in females, the corresponding rates are 13.6/100,000 and 11.1/100,000, respectively (GLOBOCAN 2012, International Agency for Research on Cancer).1 Among newly diagnosed cancer cases worldwide, lung cancer accounted for 13.0% of the total, and 19.4% of the fatal cancer cases were attributable to primary tumors in the lung, which ranked first in both malignant neoplasms.1 The data for China also showed that there were 733,000 new lung cancer cases and 591,000 new lung cancer deaths, which ranked lung tumors first in cancer incidence and mortality.2
MicroRNAs (miRNAs) are highly conserved, endogenous non-protein-encoded small molecules consisting of single-stranded noncoding RNA with lengths of 21–24 nucleotides that can be fully or partially complementary to the target sequence of the 3’-untranslated region (3’-UTR) of the target mRNAs. miRNAs can cause degradation or translation inhibition of the target mRNAs at the post-transcriptional level. They can negatively regulate gene expression and play an important role in gene regulation.3 About 30% of the coding protein genes in humans are regulated by miRNAs.4 These miRNAs can act as oncogenes and tumor suppressor genes,5 which affect tumor occurrence, development, and prognosis.6,7 The importance of miRNAs in lung cancer has been further defined in recent years.8–10 It was reported that miRNA alterations might play a crucial role in the inflammation and epithelial-to-mesenchymal transitions that are associated with the pathogenesis, diagnosis, treatment, and prognosis of lung cancer.11
The development of lung cancer is a complex process. Under the same environmental exposures, individuals with varying genetic backgrounds have different susceptibilities to lung cancer. Single-nucleotide polymorphisms (SNPs) are third-generation molecular genetic markers, and a large number of studies have shown that SNPs not only can change the type of amino acids in peptide products, causing individual phenotypic differences, but also can affect the genetic susceptibility, treatment, and prognosis of the disease.12–15 miRNA gene SNPs (including SNPs in pri-miRNA, pre-miRNA, and mature miRNA) can affect the maturation process and expression level of miRNAs, resulting in disorders or deletions in miRNA gene regulation networks,16 and have been implicated in cancer initiation and development.5
In recent years, a few studies have discussed the relationship between miRNA gene SNPs (especially rs2910164 and rs11614913) and lung cancer susceptibility.17–19 However, some of the conclusions from these studies have been controversial.20,21 The genesis of lung cancer can be associated with both environmental and genetic factors, with tobacco smoking being recognized as the most important risk factor for neoplasms of the lung. In recent years, it has been reported that smoking can lead to development of lung cancer after regulatory dysfunction by miRNA22,23 and that the toxins in cigarette smoke can cause genetic or epigenetic damage to miRNAs, including changes in SNPs.24 We propose that smoking may be an effect modification factor for the association between miRNA gene SNPs and lung cancer. Thus, our study elucidated the association between miRNA gene SNPs and lung cancer in the Han Chinese population, and further analyzed the combined and interactive effects of genetic and environmental factors, understanding of which may lead to better prevention and control of lung cancer.
Materials and methods
Study subjects
We recruited 1,067 patients with primary lung cancer from three hospitals (The First Affiliated Hospital of Fujian Medical University, Fujian Medical University Union Hospital, and Fuzhou General Hospital of Nanjing Military Command), and 1,085 gender- and age-frequency matched (±2 years) healthy controls were selected from the communities between January 2006 and December 2012. All cases and controls were residents of Fujian Province. This study was approved by the Institutional Review Board of Fujian Medical University (Fuzhou, China) and all participants signed informed consent forms.
Data and sample collections
All epidemiological data were obtained by in-person interviews using a standardized questionnaire, which collected information on general demographic characteristics, body mass index (BMI), smoking, exposure to environmental tobacco smoke (ETS), alcohol consumption, tea use, history of lung diseases, occupational exposure to hazardous substances, family history of cancer, living near to sources of pollution or industrial toxic emissions, type of residence, ventilation status, type of cooking fuel, exposure to cooking fumes, use of exhaust fans or a range hood in the kitchen, cooking oil temperature, and whether or not their home had been decorated in the past 10 years. All research variables were clearly defined.
A 5 mL non-fasting blood sample was collected from each case with a vacuum tube. A saliva sample or 5 mL non-fasting blood sample was collected from each control using the Oragene DNA Self-collection kit (DNA Genotek, OraSure Technologies, Inc., Ottawa, Ontario, Canada) or a vacuum tube.
Selection of SNPs
Based on the constantly updated website of the miRNA Public Registry: miRBase (http://www.mirbase.org/), and by using the dbSNP database (http://www.ncbi.nlm.nih.gov/snp) and the Variation Viewer (http://www.ncbi.nlm.nih.gov/variation/view/), SNPs with a minor allele frequency (MAF) >5% in the Han population were selected. The results showed that in this population, pre-miR-146a rs2910164 and miR-196a rs11614913 were in the first three. By searching for any miRNA gene SNPs that were related to tumor susceptibility or prognosis, the other three SNPs (miR-26a1 rs7372209, miR-27a rs895819, and miR-300 rs12894467) were identified (Table S1).
Genotyping
Genomic DNA was extracted from the blood samples by protease K digestion and phenol-chloroform extraction and purification according to standard procedures. The genomic DNA was stored at –20°C until being subjected to SNP genotyping with the Sequenom platform according to the manufacturer’s iPLEX Application Guide (Sequenom Inc., San Diego, CA, USA). The samples were scanned through a matrix-assisted laser desorption ionization-time-of-flight mass spectrometry system and genotyped with a MassArray Type 3.4 (Sequenom Inc.). Approximately 10% of the samples (randomly selected) were rerun for quality control purposes. Genotyping call rates were >90% (case group: 96.0% for rs2910164, 94.3% for rs11614913, 94.7% for rs7372209, 94.3% for rs895819, and 96.0% for rs12894467; and control group: 97.5% for rs2910164, 96.9% for rs11614913, 97.9% for rs7372209, 94.6% for rs895819, and 95.9% for rs12894467) and the concordance rate reached 99.5%.
Statistical methods
Differences in distributions of selected demographic factors between cases and controls were compared using chi-squared tests. Unconditional logistic regression models were used to estimate the odds ratios (ORs) and 95% confidence intervals (CIs) of the associations between SNPs and lung cancer in dominant, recessive, and additive genetic models. A multinomial unconditional logistic regression model was used to evaluate the association between SNPs and different pathological types of lung cancer. Potential confounders were selected based on prior knowledge of lung cancer. The relative excess risk of interaction (RERI), attributable proportion of interaction (API), synergy index (S), and their 95% CI were calculated by the Excel table made by Andersson25 to evaluate the additive interaction between miRNA gene SNPs and environmental factors, RERI = OR11− OR01− OR10+1, API = RERI/OR11 and S = (OR11−1)/[(OR01−1) + (OR10−1)]. If there is no additive interaction, the 95% CI of the RERI and API contains 0 and the 95% CI of S contains 1. The combined effects and multiplication interaction were analyzed by crossover analysis and logistic regression model. Hardy–Weinberg equilibrium was conducted using a goodness-of-fit chi-squared test for each SNP among the controls. All statistical analyses were conducted using Stata 14.0 software (Stata Corp, College Station, TX, USA). A P value <0.05 was considered to be statistically significant.
Results
Baseline demographic characteristics
In the current study, 1,067 cases and 1,085 controls were included. The mean age of the cases was 58.63±10.73 years; that of the control group was 59.00±10.83 years (t=–0.800, P=0.424). There was no significant difference in age, gender, marital status, passive smoking, and tea drinking, whereas there were significant differences between their histories of occupational exposure to hazardous substances, family history of cancer, education, BMI, smoking, drinking alcohol, and history of lung diseases. Of the 1,067 cases of lung cancer, there were 509 (47.7%) with lung adenocarcinomas, 297 (27.8%) squamous cell carcinoma, 86 (8.1%) small cell carcinoma, 65 (6.1%) alveolar cell carcinoma, 33 (3.1%) adenosquamous carcinoma, 25 (2.3%) large cell carcinoma, and 52 (4.9%) other types (Table 1).
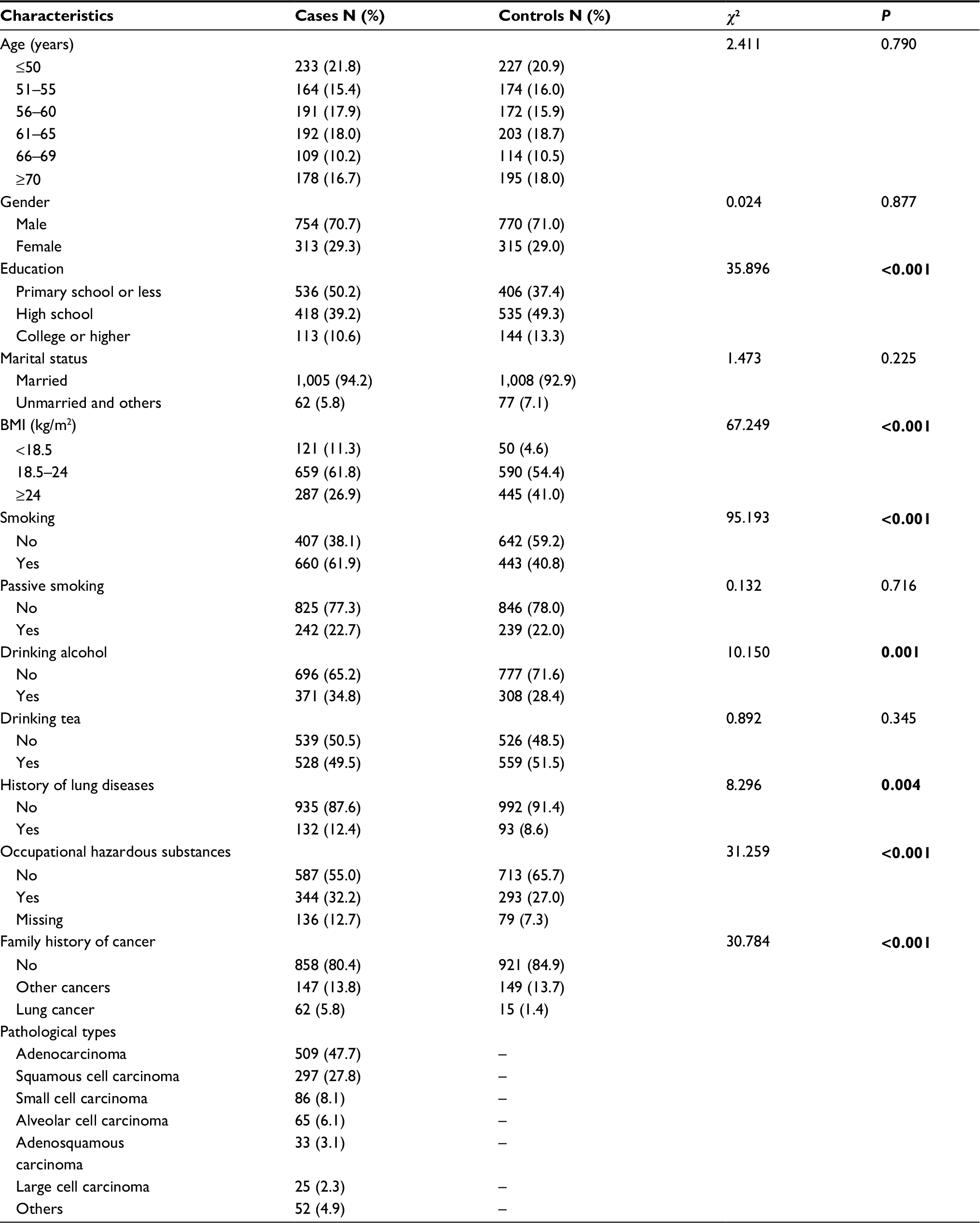 | Table 1 Subject characteristics by case and control groups Note: The values in bold represent P<0.05. Abbreviation: BMI, body mass index. |
Association between single miRNA gene SNP and lung cancer
The distribution of each SNP in the control group was in accordance with the Hardy–Weinberg equilibrium (P>0.05). We investigated the relationship between three genetic models (dominant, recessive, and additive) of miRNA gene SNPs and primary lung cancers of different pathological types. However, rs2910164, rs11614913, rs7372209, rs895819, and rs12894467 were not associated with different pathological types of primary lung cancer (P>0.05) (Table 2).
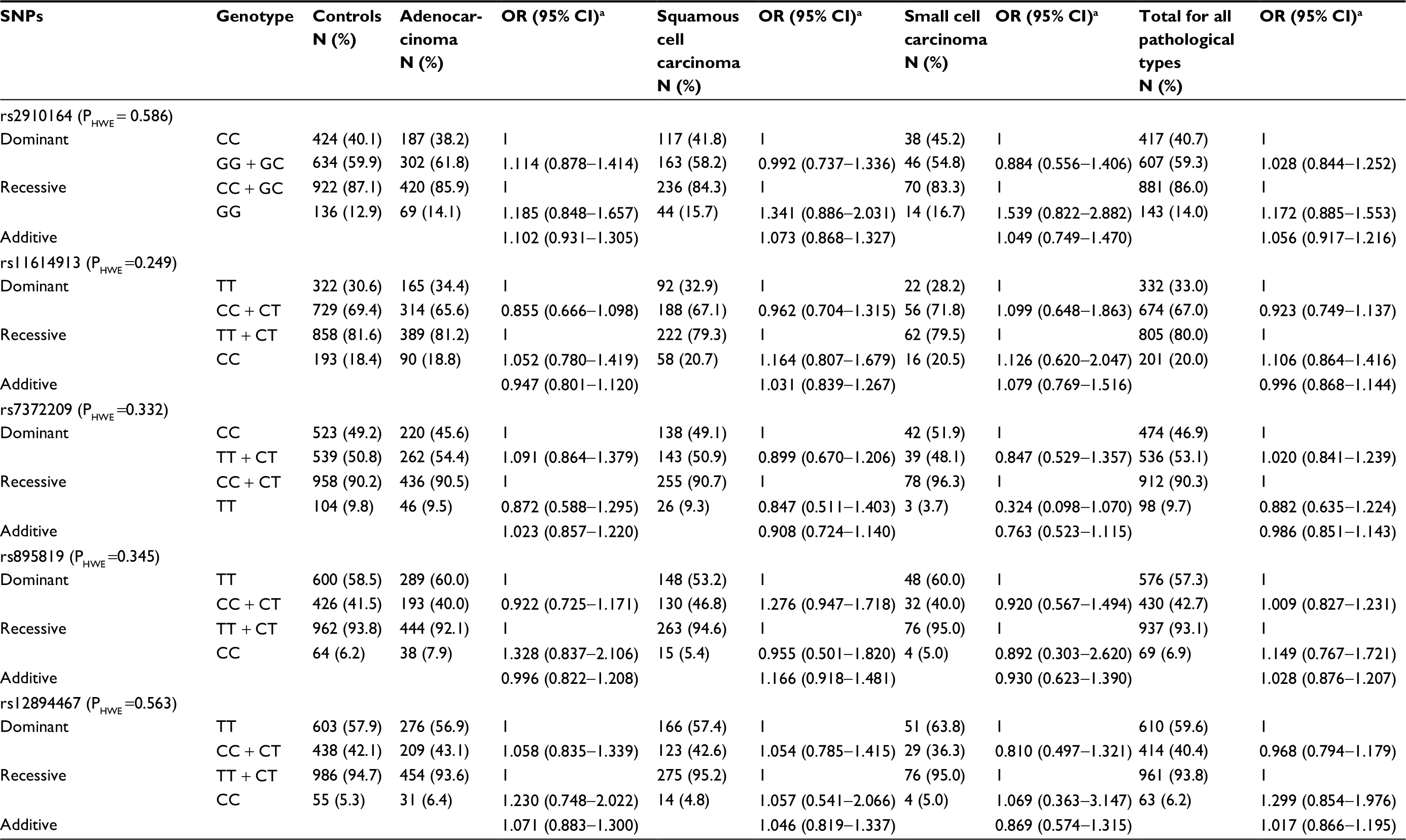 | Table 2 The relationship between miRNA gene SNPs and different pathological types of primary lung cancer Note: aAdjustment of age, gender, education, marital status, BMI, drinking alcohol, drinking tea, history of lung diseases, occupational exposure to hazardous substances, family history of cancer, smoking, ETS, living nearby the polluting enterprises, residential type, ventilation status, type of cooking fuel, exposure to cooking fumes, using exhaust fans or range hood, cooking oil temperature, and house decorated or not in the past 10 years. Abbreviations: BMI, body mass index; CI, confidence interval; ETS, environmental tobacco smoke; HWE, Hardy–Weinberg equilibrium; OR, odds ratio. |
After stratified analysis by gender and smoking status, and adjusting for possible confounders, we determined that in the dominant genetic model, the rs12894467 CC/CT genotype was associated with a significantly increased risk for lung cancer in women (with an adjusted OR =1.46, 95% CI=1.01–2.10); smokers carrying the CC/CT genotype were associated with a significantly decreased risk of lung cancer, the adjusted OR was 0.75 (95% CI: 0.57–0.98). The association of rs12894467 and lung cancer showed significant heterogeneity in different gender (P=0.009) and smoking status (P=0.004). There were no statistically significant associations between the rs2910164, rs11614913, rs7372209 and rs895819, and lung cancer (Figure 1).
 | Figure 1 Stratified analysis of the association between miRNA gene SNPs and the risk of primary lung cancer in the Han population. Notes: A and B are stratified by gender in dominant and recessive models respectively. C and D are stratified by smoking status in dominant and recessive models respectively. aAdjustment of age, gender, education, marital status, BMI, drinking alcohol, drinking tea, history of lung diseases, occupational exposure to hazardous substances, family history of cancer, smoking, ETS, living nearby the polluting enterprises, residential type, ventilation status, type of cooking fuel, exposure to cooking fumes, using exhaust fans or range hood, cooking oil temperature, and house decorated or not in the past 10 years. Abbreviations: BMI, body mass index; CI, confidence interval; ETS, environmental tobacco smoke; OR, odds ratio; SNPs, single-nucleotide polymorphisms. |
Combined and interaction effects of rs12894467 and gender or smoking
After adjustment for possible confounding factors, analysis showed that in the dominant model, rs12894467 and gender showed a positive multiplicative interaction, and rs12894467 and smoking had a negative multiplicative interaction. In the rs12894467 dominant model, women with CC/CT genotypes and smokers carrying the TT genotype had a higher risk of lung cancer. Additive interactive effects were not detected (Table 3).
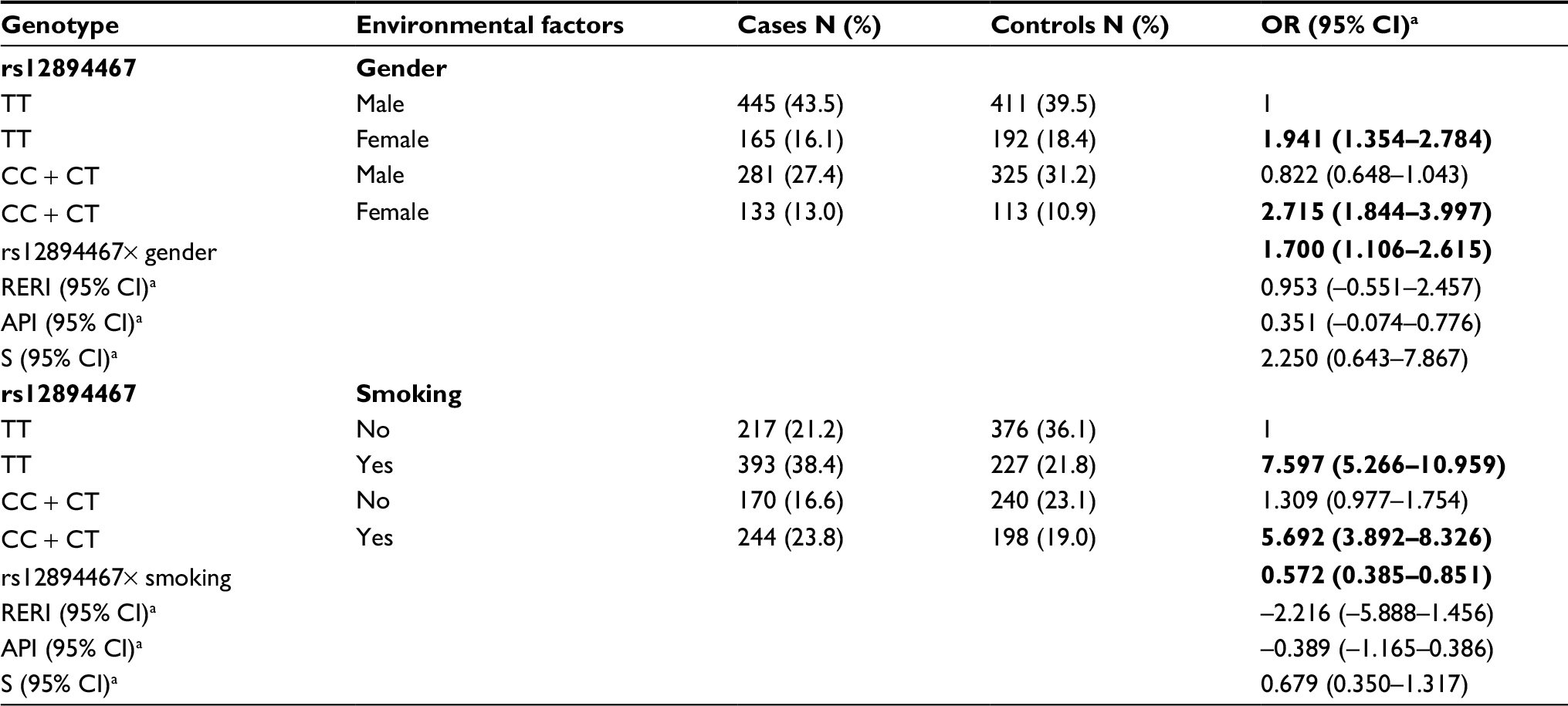 | Table 3 The combined and interaction effects of rs12894467 and gender or smoking in dominant model Notes: aAdjustment of age, gender, education, marital status, BMI, drinking alcohol, drinking tea, history of lung diseases, occupational exposure to hazardous substances, family history of cancer, smoking, ETS, living nearby the polluting enterprises, residential type, ventilation status, type of cooking fuel, exposure to cooking fumes, using exhaust fans or range hood, cooking oil temperature, and house decorated or not in the past 10 years. The values in bold represent P<0.05. Abbreviations: API, attributable proportion of interaction; CI, confidence interval; ETS, environment tobacco smoke; OR, odds ratio; RERI, relative excess risk of interaction; S, synergy index. |
Discussion
At present, the process for biosynthesis of miRNA has been basically determined. First, in the nucleus, the gene sequence encoding the miRNA is transcribed as a long primary miRNA (pri-miRNA), and then the pri-miRNA is cleaved to a length of about 60–70 nucleotides, a miRNA precursor with a stem-loop structure (pre-miRNA).26–28 The pre-miRNA is then transported from the nucleus to the cytoplasm by a transporter protein and cleaved into the siRNA-like incompletely matched double-stranded RNA consisting of 21–24 nucleotides.29–31 Finally, miRNAs* and miRNAs are generated by RNA helicase. Mature miRNAs are involved in RNA-induced silencing complex (RISC), and miRNAs* are degraded. miRNAs mainly mediate post-transcriptional gene regulation through two mechanisms: degradation and translation inhibition of target mRNAs.
miRNA gene SNPs can affect the miRNA maturation process, similar to the process in common genes, and miRNA gene transcription is also regulated by the promoter.32 The mechanism of miRNA gene SNPs is similar to the general gene SNPs, both the SNPs in coding and regulatory area can affect the transcription and synthesis of miRNA. They thus can affect the miRNA and target mRNA interaction, resulting in miRNA posttranscriptional gene regulation network abnormalities, and thus closely related to tumor susceptibility.
Jeon et al33 found that the CG + GG genotype of pre-miR-146a rs2910164 in the Korean population reduced the risk of lung cancer (relative to the CC genotype), with an adjusted OR of 0.80 (95% CI: 0.66–0.96), this relationship was only found in non-smokers. A meta-analysis34 also showed that the rs2910164 was associated with the susceptibility of lung cancer in the East Asian population (G vs C: OR =0.92, 95% CI: 0.85–0.99; CG + GG vs CC: OR =0.86, 95% CI: 0.76–0.99). In the Chinese population,17 miR-146a rs2910164 was associated with risk of non-small-cell lung cancer, and the frequency distribution of the CC genotype and C allele was greater in non-small-cell lung cancer patients than in the control group (P=0.03). However, our study showed that there was no significant correlation between rs2910164 and all or different pathological types of lung cancer in the Han population in China, even after stratified analysis by gender and smoking status. One meta-analysis20 did not report an association of rs2910164 with susceptibility to lung cancer, consistent with the results of our study. At present, the research about rs2910164 and susceptibility to lung cancer is still limited, and the results of the meta-analyses were unstable. In addition, studies of different races, research designs, and sample sizes may lead to different results, so further research will be required to clarify these issues.
Tian et al6 conducted a case–control study to investigate the association of SNPs in four pre-miRNAs with the susceptibility of lung cancer. It was found that SNP rs11614913 in miR-196a was associated with the susceptibility of lung cancer, and the risk of lung cancer in people with genotype CC was 25% higher than those with TT/CT (OR =1.25, 95% CI: 1.01–1.54). Meta-analysis19 also showed that rs11614913 allele C could increase the risk of lung cancer in Asian populations (C vs T: OR =1.12, 95% CI: 1.03–1.22; and CC vs TT: OR =1.26, 95% CI: 1.07–1.49), and several other meta-analyses18,35,36 demonstrated similar results. Our study found that the association between rs11614913 CC genotype and lung cancer was not statistically significant, and the 95% CI includes 1, but the estimated OR value is greater than 1, which indicates that this result might reflect an insufficient sample size, and a failure to identify the association of this potential micro-susceptibility biomarker with lung cancer. In addition, the variability of the results might be attributed to the different populations and research designs, different data analysis methods, and so on. Therefore, the association of this SNP with lung cancer will require a larger sample size and more relevant studies for further validation.
Studies have shown that pri-miR-26a1 rs7372209 is associated with premalignant oral lesions.37 Our results showed that the relationship between rs7372209 and lung cancer susceptibility was not statistically significant, which is similar to the results of studies of cervical cancer38 and esophageal cancer.39,40 Similarly, Yin et al41 found no association between this SNP and the risk of lung cancer in non-smoking female populations in China. Moreover, Yoon et al42 did not find an association between rs7372209 and the prognosis of non-small-cell lung cancer. The pre-miR-27a rs895819 CC mutation has been reported to increase the risk of gastric cancer43 and cervical cancer.38 Only one case–control study with 560 lung cancer patients and 568 healthy controls in China reported that the rs895819 CC mutation could increase the risk of non-small-cell lung cancer.44 Our study was based on a larger sample size but found no association between rs895819 and the risk of lung cancer. Further research using multi-center studies and larger sample sizes may be needed to fully validate the association between rs895819 and the risk of lung cancer.
miRNA-300 is closely related to a variety of tumors.45,46 Shen et al46 reported that miR-300 expression was upregulated in gastric cancer, and could promote gastric cancer cell proliferation and invasion by targeting P53. Xue et al47 showed that upregulation of miR-300 promoted proliferation and invasion of osteosarcoma by targeting BRD7. miRNA-300 was also commonly upregulated in glioma tissues, it could enhance the self-renewal of glioma stem-like cells and reduce differentiation toward both astrocyte and neural fates.48 Yu et al49 reported that miR-300 in human epithelial cancer inhibited the epithelial-to-mesenchymal transition and metastases by targeting Twist, and that in addition miR-300 could inhibit experimental lung metastasis in vivo. The rs12894467 is a SNP in the pre-miR-300 ring region that has been associated with risk of pediatric acute lymphoblastic leukemia,50 but not with risk of acquired immune deficiency syndrome (AIDS)-related non-Hodgkin’s lymphoma.51 So far, no published studies have addressed possible associations of rs12894467 with lung cancers. After stratification by gender and smoking status, our study found a significant association between rs12894467 and primary lung cancer. Further interactive analysis demonstrated that smoking and gender were both effect modification factors for the association between rs12894467 and lung cancer, and this may highlight new strategies for lung cancer risk stratification and prevention.
Because of the unavoidable selection and memory biases in case–control studies, and because the same miRNAs may play different regulatory roles in different races and tissues, the variable study designs and sample sizes may also affect the relationships between miRNA gene SNPs and lung cancer. Therefore, the results of our study still need more rigorous design of larger multicenter studies to further verify and further explore the mechanisms responsible for the results of our study.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (81402738), the Fujian Program for Outstanding Young Researchers in University awarded by the Education Department of Fujian (2017B019), the National Key Research and Development Program of China (2017YFC0907100), and the Fujian Provincial Natural Science Foundation Project (2016J01355). We would also like to express our appreciation to the patients who participated in our study.
Disclosure
The authors report no conflicts of interest in this work.
References
Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–E386. | ||
Chen W, Zheng R, Zhang S, Zeng H, Zou X, He J. Report of Cancer Incidence and Mortality in China, 2013. China Cancer. 2017;26(1):1–7. | ||
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. | ||
Ambros V. The functions of animal microRNAs. Nature. 2004; 431(7006):350–355. | ||
Slaby O, Bienertova-Vasku J, Svoboda M, Vyzula R. Genetic polymorphisms and microRNAs: new direction in molecular epidemiology of solid cancer. J Cell Mol Med. 2012;16(1):8–21. | ||
Tian T, Shu Y, Chen J, et al. A functional genetic variant in microRNA-196a2 is associated with increased susceptibility of lung cancer in Chinese. Cancer Epidemiol Biomarkers Prev. 2009;18(4):1183–1187. | ||
Hu Z, Chen J, Tian T, et al. Genetic variants of miRNA sequences and non-small cell lung cancer survival. J Clin Invest. 2008;118(7):2600–2608. | ||
Zhao L, Ni X, Zhao L, et al. MiroRNA-188 Acts as tumor suppressor in non-small-cell lung cancer by targeting MAP3K3. Mol Pharm. 2018;15(4):1682–1689. | ||
Du X, Qi F, Lu S, Li Y, Han W. Nicotine upregulates FGFR3 and RB1 expression and promotes non-small cell lung cancer cell proliferation and epithelial-to-mesenchymal transition via downregulation of miR-99b and miR-192. Biomed Pharmacother. 2018;101:656–662. | ||
Mao G, Liu Y, Fang X, et al. Tumor-derived microRNA-494 promotes angiogenesis in non-small cell lung cancer. Angiogenesis. 2015;18(3):373–382. | ||
Castro D, Moreira M, Gouveia AM, Pozza DH, De Mello RA. MicroRNAs in lung cancer. Oncotarget. 2017;8(46):81679–81685. | ||
Roses AD. Pharmacogenetics place in modern medical science and practice. Life Sci. 2002;70(13):1471–1480. | ||
Przeworski M, Hudson RR, Di Rienzo A. Adjusting the focus on human variation. Trends Genet. 2000;16(7):296–302. | ||
Kim WH, Min KT, Jeon YJ, et al. Association study of microRNA polymorphisms with hepatocellular carcinoma in Korean population. Gene. 2012;504(1):92–97. | ||
Giacalone A, Montalto G, Giannitrapani L, et al. Association between single nucleotide polymorphisms in the cyclooxygenase-2, tumor necrosis factor-a, and vascular endothelial growth factor-A genes, and susceptibility to hepatocellular carcinoma. OMICS. 2011;15(3):193–196. | ||
Campayo M, Navarro A, Viñolas N, et al. A dual role for KRT81: a miR-SNP associated with recurrence in non-small-cell lung cancer and a novel marker of squamous cell lung carcinoma. PLoS One. 2011;6(7):e22509. | ||
Jia Y, Zang A, Shang Y, et al. MicroRNA-146a rs2910164 polymorphism is associated with susceptibility to non-small cell lung cancer in the Chinese population. Med Oncol. 2014;31(10):194. | ||
He B, Pan Y, Cho WC, et al. The association between four genetic variants in microRNAs (rs11614913, rs2910164, rs3746444, rs2292832) and cancer risk: evidence from published studies. PLoS One. 2012;7(11): e49032. | ||
Yuan Z, Zeng X, Yang D, Wang W, Liu Z. Effects of common polymorphism rs11614913 in Hsa-miR-196a2 on lung cancer risk. PLoS One. 2013;8(4):e61047. | ||
Xu L, Tang W. The associations of nucleotide polymorphisms in mir-196a2, mir-146a, mir-149 with lung cancer risk. Cancer Biomark. 2015;15(1):57–63. | ||
Hao X, Xia L, Qu R, Yang X, Jiang M, Zhou B. Association between miR-146a rs2910164 polymorphism and specific cancer susceptibility: an updated meta-analysis. Fam Cancer. 2018;17(3):459–468. | ||
Van Pottelberge GR, Mestdagh P, Bracke KR, et al. MicroRNA expression in induced sputum of smokers and patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2011;183(7):898–906. | ||
Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6(11):857–866. | ||
Russ R, Slack FJ. Cigarette-smoke-induced dysregulation of microRNA expression and its role in lung carcinogenesis. Pulm Med. 2012;2012:791234–791239. | ||
Andersson T, Alfredsson L, Källberg H, Zdravkovic S, Ahlbom A. Calculating measures of biological interaction. Eur J Epidemiol. 2005;20(7):575–579. | ||
Lee Y, Jeon K, Lee JT, Kim S, Kim VN. MicroRNA maturation: stepwise processing and subcellular localization. Embo J. 2002;21(17):4663–4670. | ||
Lee Y, Ahn C, Han J, et al. The nuclear RNase III Drosha initiates microRNA processing. Nature. 2003;425(6956):415–419. | ||
Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the Microprocessor complex. Nature. 2004;432(7014):231–235. | ||
Bagga S, Bracht J, Hunter S, et al. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell. 2005;122(4):553–563. | ||
Hutvágner G, Mclachlan J, Pasquinelli AE, Bálint E, Tuschl T, Zamore PD. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science. 2001;293(5531):834–838. | ||
Ketting RF, Fischer SE, Bernstein E, Sijen T, Hannon GJ, Plasterk RH. Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Genes Dev. 2001;15(20):2654–2659. | ||
Johnson SM, Lin SY, Slack FJ. The time of appearance of the C. elegans let-7 microRNA is transcriptionally controlled utilizing a temporal regulatory element in its promoter. Dev Biol. 2003;259(2):364–379. | ||
Jeon HS, Lee YH, Lee SY, et al. A common polymorphism in pre-microRNA-146a is associated with lung cancer risk in a Korean population. Gene. 2014;534(1):66–71. | ||
Fan X, Wu Z. Effects of four single nucleotide polymorphisms in microRNA-coding genes on lung cancer risk. Tumour Biol. 2014;35(11):10815–10824. | ||
Chu H, Wang M, Shi D, et al. Hsa-miR-196a2 Rs11614913 polymorphism contributes to cancer susceptibility: evidence from 15 case-control studies. PLoS One. 2011;6(3):e18108. | ||
Wang G, Wang W, Gao W, Lv J, Fang J. Two functional polymorphisms in microRNAs and lung cancer risk: a meta-analysis. Tumour Biol. 2014;35(3):2693–2699. | ||
Clague J, Lippman SM, Yang H, et al. Genetic variation in MicroRNA genes and risk of oral premalignant lesions. Mol Carcinog. 2010;49(2):183–189. | ||
Xiong XD, Luo XP, Cheng J, Liu X, Li EM, Zeng LQ. A genetic variant in pre-miR-27a is associated with a reduced cervical cancer risk in southern Chinese women. Gynecol Oncol. 2014;132(2):450–454. | ||
Zhang J, Huang X, Xiao J, et al. Pri-miR-124 rs531564 and pri-miR-34b/c rs4938723 polymorphisms are associated with decreased risk of esophageal squamous cell carcinoma in Chinese populations. PLoS One. 2014;9(6):e100055. | ||
Wei J, Zheng L, Liu S, et al. miR-196a2 rs11614913 T > C polymorphism and risk of esophageal cancer in a Chinese population. Hum Immunol. 2013;74(9):1199–1205. | ||
Yin Z, Li H, Cui Z, et al. Polymorphisms in pre-miRNA genes and cooking oil fume exposure as well as their interaction on the risk of lung cancer in a Chinese nonsmoking female population. Onco Targets Ther. 2016;9:395–401. | ||
Yoon KA, Yoon H, Park S, et al. The prognostic impact of microRNA sequence polymorphisms on the recurrence of patients with completely resected non-small cell lung cancer. J Thorac Cardiovasc Surg. 2012;144(4):794–807. | ||
Sun Q, Gu H, Zeng Y, et al. Hsa-mir-27a genetic variant contributes to gastric cancer susceptibility through affecting miR-27a and target gene expression. Cancer Sci. 2010;101(10):2241–2247. | ||
Ma JY, Yan HJ, Yang ZH, Gu W. Rs895819 within miR-27a might be involved in development of non small cell lung cancer in the Chinese Han population. Asian Pac J Cancer Prev. 2015;16(5):1939–1944. | ||
Wang L. The Upregulation of miR-300 in Colorectal Cancer Affects the Malignancy of the Tumor Cells. Chongqing, China: Third Military Medical University; 2015. | ||
Shen Z, Li C, Zhang K, et al. The up-regulation of miR-300 in gastric cancer and its effects on cells malignancy. Int J Clin Exp Med. 2015;8(5):6773–6783. | ||
Xue Z, Zhao J, Niu L, An G, Guo Y, Ni L. Up-regulation of miR-300 promotes proliferation and invasion of osteosarcoma by targeting BRD7. PLoS One. 2015;10(5):e0127682. | ||
Zhang D, Yang G, Chen X, et al. mir-300 promotes self-renewal and inhibits the differentiation of glioma stem-like cells. J Mol Neurosci. 2014;53(4):637–644. | ||
Yu J, Xie F, Bao X, Chen W, Xu Q. miR-300 inhibits epithelial to mesenchymal transition and metastasis by targeting Twist in human epithelial cancer. Mol Cancer. 2014;13:121. | ||
López-López E, Gutiérrez-Camino Á, Piñán MÁ, et al. Pharmacogenetics of microRNAs and microRNAs biogenesis machinery in pediatric acute lymphoblastic leukemia. PLoS One. 2014;9(3):e91261. | ||
Peckham-Gregory EC, Thapa DR, Martinson J, et al. MicroRNA-related polymorphisms and non-Hodgkin lymphoma susceptibility in the Multicenter AIDS Cohort Study. Cancer Epidemiol. 2016;45:47–57. |
Supplementary material
 | Table S1 The information about selected miRNA gene SNPs Abbreviations: MAF, minor allele frequency; SNP, single-nucleotide polymorphism. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.