Back to Journals » Cancer Management and Research » Volume 12
miR-148a Affects Polarization of THP-1-Derived Macrophages and Reduces Recruitment of Tumor-Associated Macrophages via Targeting SIRPα
Authors Ma D, Zhang Y, Chen G, Yan J ![]()
Received 12 November 2019
Accepted for publication 5 August 2020
Published 4 September 2020 Volume 2020:12 Pages 8067—8077
DOI https://doi.org/10.2147/CMAR.S238317
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Beicheng Sun
Donghe Ma,1 Yan Zhang,2 Guanghua Chen,3 Jia Yan1
1Digestive Department, University-Town Hospital of Chongqing Medical University, Chongqing 401331, People’s Republic of China; 2Neurology Department, People’s Hospital of Hechuan Chongqing, Chongqing 401520, People’s Republic of China; 3Dermatology Department, Children’s Hospital of Chongqing Medical University, Chongqing 400014, People’s Republic of China
Correspondence: Jia Yan Tel +86-17782351875
Fax +86-23-65715900
Email [email protected]
Purpose: The objective of this study was to investigate the effect of miR-148a on the polarization and recruitment of tumor-associated macrophages (TAMs).
Methods: In human monocyte THP-1 cells, M1 or M2 differentiation was induced by phorbol 12-myristate 13-acetate (PMA) with specific induction supplements and identified using flow cytometry and ELISA. To alter cellular miR-148a expression level, THP-1 cells were transfected with miR-148a mimics or inhibitors. A dual-luciferase assay was used to determine whether miR-148a could directly regulate the expression of signal regulatory protein α (SIRPα). Expression of miR-148a and SIRPα was detected with RT-PCR or Western blot. A co-culture system of THP-1 cells and colorectal cancer SW480 cells was used for TAM induction. The recruitment of macrophage to SW480 cells was measured using chemotaxis assay. In SW480 cells, apoptosis induced by macrophages was detected using flow cytometry and a xenograft assay. Macrophage infiltration was detected by immunofluorescence assay in tumor tissues.
Results: miR-148a over-expression increased M1-related CD86 expression in THP-1 cells and promoted differentiation to M1-like macrophages. Inhibition of miR-148a increased M2-related CD206 expression and promoted differentiation to M2-like macrophages. In the co-culture system, THP-1 cells were induced to the M2-like state by SW480 cells. The level of miR-148a negatively correlated with the levels of M2-related cytokines. Additionally, miR-148a expression level was negatively associated with macrophage recruitment by colorectal cancer cells. Furthermore, in miR-148a over-expression, the number of macrophages recruited by SW480 cells was reduced. Meanwhile, cancer cell apoptosis induced by macrophages was enhanced. Thus, miR-148a expression was beneficial for the transformation of macrophages, from an immune-suppressive status to an immune-promoting status. These anti-cancer effects of miR-148a were related to the down-regulation of SIRPα in macrophages, directly targeted by miR-148a. A xenograft assay showed that the co-inoculation of macrophages over-expressing miR-148a reduced subcutaneous tumorigenesis and M2 macrophage infiltration.
Conclusion: miR-148a promoted the differentiation of M0 macrophages into anti-tumor classical activation type M1, and reduced TAM recruitment by targeting SIRPα to inhibit colorectal cancer cell viability.
Keywords: colorectal cancer, tumor-associated macrophage, microRNA-148a, polarization, signal regulatory protein alpha
Plain Language Summary
- Background: Polarization of tumor-associated macrophages (TAMs) presents important effect on the growth of tumor cells, angiogenesis, and local infiltration. Recent studies have revealed that the polarization of TAMs can be regulated by miRNAs.
- Question: Does miR-148a, a down-regulated miRNA, affect the regulation of TAMs in colorectal cancer?
- What did we do: 1) Induce the polarization of human monocyte THP-1 cells in vitro and examine the relationship between miR-148a expression and polarization status. 2) Investigate whether miR-148a expression affects macrophage recruitment using in vitro chemotaxis assay. 3) Finally, assess whether miR-148 enhances the anti-tumor effect of macrophages using a xenografts assay.
- What we found: 1) miR-148a promoted polarization of anti-tumor M1 type macrophages while inhibiting the polarization of pro-tumor M2 type macrophages. 2) In the co-culture system, THP-1 cells could be induced into the M2-like state by colorectal cancer cells. 3) miR-148a reduced macrophage recruitment by directly targeting SIRPα, and increased apoptosis of cancer cells in the co-culture system. 3) Over-expression of miR-148a in macrophages reduced subcutaneous tumorigenesis and infiltration of M2 macrophages in vivo.
- What do these results mean: miR-148a can play a multi-level anti-tumor role by participating in macrophage polarization and recruitment. Therefore, miR-148a may possess potential therapeutic value in colorectal cancer.
Introduction
According to the newly released GLOBOCAN statistics 2018, colorectal cancer is the third most commonly diagnosed cancer (6.1% of the total cases) and the second leading cause of cancer death (9.2% of the total cancer deaths).1 Distant metastasis and recurrence of colorectal cancer are the main factors associated with poor prognosis, which is closely influenced by microenvironment composed of tumors and stroma.2 In recent years, the importance of the immune microenvironment in cancer metastasis has gained considerable attention. Tumor-associated macrophages (TAMs), the most abundant inflammatory infiltrating cells in tumor stromal cells, are an important component of the tumor microenvironment. TAMs can be regulated by tumor cells and differentiate into anti-tumor classical activation type (M1 type) or pro-tumor alternative activation type (M2 type).3 Moreover, an increased number of M2 TAMs has been highly correlated with tumor metastasis, angiogenesis, and poor prognosis.4 Regarding the molecular mechanism, M2 TAMs can produce several cytokines necessary for tumor cell growth and angiogenesis, including transforming growth factor β (TGF-β), vascular endothelial growth factor (VEGF), epidermal growth factor (EGF).5 M2 TAMs synthesize and secrete proteolytic enzymes such as matrix metalloproteinase (MMP) and cathepsin, which provide necessary conditions for invasion, infiltration, and migration of tumor cells.6,7 Furthermore, M2 TAMs can inhibit humoral and cellular immunity against cancer cells via several pathways, maintaining tumor cells in a state of immune tolerance and to escape elimination by the body.8
Notably, TAM regulation is an important anti-tumor strategy. Reprogramming of TAMs toward M1-like macrophages can suppress tumor progression, which can be achieved through chemotherapy, radiotherapy, and immunotherapy.9 Currently, studies have shown that polarization of TAMs can be regulated by microRNAs (miRNAs). miR-19a-3p inhibits the progression of breast cancer by down-regulating M2-like macrophage polarization by targeting Fra-1 expression.10 miR-301a-3p was enriched in pancreatic cancer cell-derived exosomes, promoting malignant behaviors by increasing polarization toward M2 macrophages.11 In patients with advanced colorectal cancer, miR-148a was an independent prognostic factor. miR-148a was down-regulated in colorectal cancer tissues and was associated with shorter overall survival.12 The effect of miR-148a on TAMs remained unclear. Therefore, the objective of the current study was to investigate the effect of miR-148a on the polarization and recruitment of TAMs in colorectal cancer cells.
Materials and Methods
Cell Lines and Cell Cultures
Human peripheral blood monocyte THP-1 cells (ATCC, Manassas, USA) were used for TAM induction. Colorectal adenocarcinoma SW480 cells (ATCC) were used for in vitro experiments. Cells were maintained with RPMI 1640 (HyClone, Logan, USA) supplemented with 10% fetal bovine serum (HyClone) in 5% CO2 atmosphere at 37ºC. M1 and M2 polarization was induced as previously reported with marginal modifications.13 To induce M0 macrophage, THP-1 cells were activated with 100 nM phorbol 12-myristate 13-acetate (PMA, Sigma, St Louis, USA) in serum-free RPMI medium for 48 hours. After cell adherence, floating cells were carefully removed. M1 polarization was induced by incubating M0 cells with 100 pg/mL of lipopolysaccharides (LPS, R&D Systems, Minneapolis, USA) and 20 ng/mL of interferon-γ (IFN-γ, R&D). M2 polarization was induced by incubation with 20 ng/mL of interleukin-4 (IL-4, R&D).
Macrophage Marker Identification
After induction of polarization for 2 days, cells were harvested and resuspended in FACS tubes (106 cells in 100 uL PBS). Simultaneously, culture supernatants were collected and tested for specific cytokines (IL-1β and IL-12 as M1 marker, and IL-10 as M2 marker) using ELISA kits (eBioscience, San Diego, USA). Subsequently, macrophages were labeled with 5 μL of specific primary antibodies against relative surface markers (CD68-PE as pan macrophage marker,14 CD86-APC as an M1 marker,15 and CD206-FITC as an M2 marker,16 all obtained from eBioscience) on ice for 30 minutes. The stained cells were washed twice with cold PBS, resuspended in 400 μL of PBS, and detected for surface markers using flow cytometry (BD LSRII system, BD Biosciences, Franklin Lakes, USA). THP-1 cells in M0 state were processed in the same manner as the control.
Transfection and Luciferase Activity Assay
THP-1 cells were seeded in 6-well plates and induced into M0 cells. On reaching 80% confluency, 100 pmol of precursor for miR-148a mimics (5ʹ-UCAGUGCACUACAGAACUUUGU-3ʹ),17 miR-148a inhibitor (5ʹ-AGUCACGUGAUGUCUUGAAACA),17 or scramble control (5ʹ-CAGUACUUUUGUGUAGUACAA-3ʹ) (all synthesized by RiboBio Co. Ltd., Guangzhou, China) was transfected into M0 cells using Lipofectamine RNAiMAX reagent (Invitrogen, Carlsbad, USA). After 48 hours of transfection, cells were harvested and used for further molecular assays. To verify the transcriptional regulation of signal regulatory protein α (SIRPα) by miR-148a, a dual-luciferase reporter assay was performed using the Dual-Luciferase Reporter Assay System (Promega, Madison, USA). A double-stranded DNA oligo nucleotide encompassing the 3ʹ-UTR of SIRPα was cloned into the pmirGLO vector (Promega) to form the SIRP-WT plasmid. Mutagenesis of the predicted miR-148a seed sequence within the 3ʹ-UTR of SIRPα was performed by GeneArt Site-Directed mutagenesis system (Invitrogen). Mutated sequences were inserted into the pmirGLO vector to form the SIRP-MU plasmid. SW480 cells were co-transfected with the SIRP-WT plasmid or the SIRP-MU plasmid, a pRL-SV40 vector containing Renilla luciferase (Promega), and miR-148a mimics or scramble. After 48 hours of incubation, luciferase activities were detected following the manufacturer’s protocol.
RNA Extraction and Quantitative RT-PCR
Total RNA was extracted from tissue samples using TRIzol reagent (Invitrogen) in accordance with instructions. RNA concentration was calculated by detecting OD260/280. RNA was then reverse transcribed to cDNA using First-Stand cDNA Synthesis SuperMix kit (TransGen Biotech, Beijing, China). The expression of miR-148a and SIRPα was quantified with qPCR in 20 μL reaction volume using Top Green qPCR SuperMix (TransGen). The PCR reactions were carried out on 7500 Real-Time PCR System (Applied Biosystems Inc., Stockholm, Sweden). The PCR condition was set as: pre-denaturation at 95°C for 5 min; denaturation at 95°C for 30 s, annealing at 58°C for 1 min, extension at 72°C for 30 s for a total of 40 cycles. The primers used are as follows: 5ʹ-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACAAAG-3 as miR-148a RT primer; 5ʹ-TCAGTGCACTACAGAACTTTGT-3ʹ and 5ʹ-GCTGTCAACGATACGCTACGT-3ʹ for miR-148a amplification; 5ʹ-AAAATATGGAACGCTTCACGAATTTG-3ʹ as U6 RT primer; 5ʹ-CTCGCTTCGGCAGCACA-3ʹ and 5ʹ-AACGCTTCACGAATTTGCGT-3ʹ for U6 amplification; 5ʹ-CCCCACATTGCCACATACCT-3ʹ and 5ʹ-GATTTTCTCCTCCCCACCCT-3ʹ for SIRPα amplification; 5ʹ-AACGGATTTGGTCGTTTGGG-3ʹ and 5ʹ-CCTGGAAGATGGTGATGGGAT-3ʹ for GAPDH amplification. U6 and GAPDH were used as an internal control for miR-148a and SIRPα, respectively. The relative expression of miR-148a and SIRPα was normalized using the 2−ΔΔCT method.
Protein Extraction and Western Blot
Cells were harvested and lysed using RIPA lysis buffer (Beyotime Biotechnology, Shanghai, China). The supernatants containing soluble proteins were collected by centrifugation. The protein concentrations were measured using bicinchoninic acid protein assay. Equal amounts of proteins (50 μg) were resolved in 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride membrane (Millipore, Temecula, USA), and blocked with 5% skim milk. The membranes were probed with primary antibodies specific for the targeted protein overnight at 4°C. The primary antibody used at 1:1000 dilutions was: rabbit monoclonal anti-SIRPα (Abcam Inc., Cambridge, UK), and rabbit polyclonal anti-GAPDH (Abcam). GAPDH was used as an internal control. The membranes were then probed with goat anti-rabbit IgG (horseradish peroxidase-conjugated, Abcam) at room temperature for 1 h. The bands were visualized using enhanced chemiluminescence kit (Beyotime). The relative protein expression levels were analyzed with ImageJ software.
Co-Culture System and Transwell Chemotaxis Assay
A co-culture system of THP-1 cells and SW480 cells was established using 24-well transwell plates (Millipore). SW480 cells in the logarithmic growth phase were harvested and suspended in serum-free medium to reach a density of 1×105 cells/mL. One mL of the SW480 cell suspension was seeded in the lower chambers. Untransfected or transfected THP-1 cells (all M0 status) were suspended in serum-free medium and added to the upper chambers (1×106 cells in 200 μL medium). In order to study the effect of tumor cells on the polarization of macrophages, inserts with 3 μm polycarbonate membrane were used. In this case, M0 cells were co-cultured with the SW480 cells without direct contact. Macrophages without co-culture were set as the control. After 48 h, culture medium in the upper chamber was collected for detection of cytokine markers. In order to study the macrophage recruitment by tumor cells, inserts with 8 μm polycarbonate membrane were used. In this case, macrophages can migrate through the membrane. After 24 hours of co-culturing tumor cells and macrophages, the upper chambers were collected. The non-migrated macrophage cells were carefully removed, and the macrophage cells that migrated to the lower surface were fixed with formalin and stained with 0.1% crystal violet. Six random visual fields in each well were visualized under a microscope for cell counting. The SW480 cells in the lower chambers were harvested and stained with annexin V-FITC and propidium iodide (Beyotime Biotechnology, Shanghai, China). The apoptosis rate (105 cells/sample) was detected with flow cytometer.
Xenograft Assay
Female BALB/c nude mice (8-week old, purchased from SLAC Laboratory Animal Company, Shanghai, China) were used to perform the xenograft assay. The study protocol was approved by the Institutional Ethics Committee of Experimental Animal of Chongqing Medical University (Approval number XXXX). The animal experiments were performed following the National Institutes of Health guide for the care and use of laboratory animals and the AVMA euthanasia guidelines 2013. All mice were maintained in well-ventilated animal facility with sufficient food and water. Nine mice were randomly divided into three groups (3 mice per group). Untransfected THP-1 cells (M0 status) as well as THP-1 cells (M0 status) transfected with miR-148a mimics were used in the xenograft assay. Each mouse was injected with a mixture of 5×106 cells (M0 macrophages and SW480 cells mixed at 1:10 in 100 μL PBS) in the dorsal flanks of the nude mice. Inoculation with SW480 cells was used as a control. After 3 weeks of inoculation, the mice were sacrificed under anesthesia to assess tumor formation. Tumor volume and weight were measured as previously reported.18 Tumor tissues were then fixed in 10% neutral-buffered formalin and embedded in paraffin for histopathological assessment. Paraffin-embedded tumor tissues were cut into 5-μm thick sections. Infiltration of M2-like macrophages was detected by immunofluorescence as previously described.19 Briefly, the tissue sections were blocked with 10% goat serum and incubated with mouse anti-CD68 and rabbit anti-CD206, and then visualized using Alexa Fluor Plus 555 goat anti-mouse IgG and Alexa Fluor 488 goat anti-rabbit IgG (all antibodies obtained from eBioscience). Then, DAPI nuclear staining buffer was added and incubated at room temperature for 5 min. The sections were mounted with fluorescence quenching buffer and observed under a Zeiss confocal microscope (LSM 780, Carl Zeiss, Germany) at 100× magnification.
Statistical Analysis
All the data were processed with GraphPad Prism 6 (GraphPad Software Inc., San Diego, USA). Continuous variables are expressed as mean ± standard deviation (SD). Two groups were compared by Student’s t-test. Multiple comparisons of tumor volume were performed by Mann–Whitney U-test; other multiple comparisons were performed by one-way ANOVA followed by Tukey’s post-test. P values <0.05 were considered statistical significant.
Results
miR-148a is Differentially Expressed During Macrophage Polarization
After induction using PMA, THP-1 cells changed from suspension to adherent growth, with the morphology gradually changing from round to irregular shape. After 48 hours, THP-1 cells formed numerous protrusions and pseudopods and became M0 cells. Sequential M1 or M2 polarization was induced in the presence of LPS+IFN-γ or IL-4, respectively. The expression of recognized macrophage markers, CD68, CD86, and CD206, was analyzed by immunofluorescence staining (Figure 1A). The pan macrophage marker, CD68, was expressed in both M1-like and M2-like subtypes. The ratio of CD86+ cells significantly increased in M1-like macrophages; the ratio of CD206+ cells significantly increased in M2-like macrophages. Compared with M0 cells, expression of anti-inflammatory factors, IL-1β and IL-12, was up-regulated by LPS+IFN-γ induction; simultaneously, expression of proinflammatory factor, IL-10, was up-regulated by IL-4 induction (Figure 1B). These results indicated that the monocyte-to-macrophage induction system was established. Next, we examined the expression of miR-148a in different macrophage subtypes. As shown in Figure 1C, miR-148a was differentially expressed. Compared with THP-1 cells, miR-148a was up-regulated in M1-like macrophages and down-regulated in M2-like macrophages. These results suggested that miR-148a might play a role in macrophage polarization.
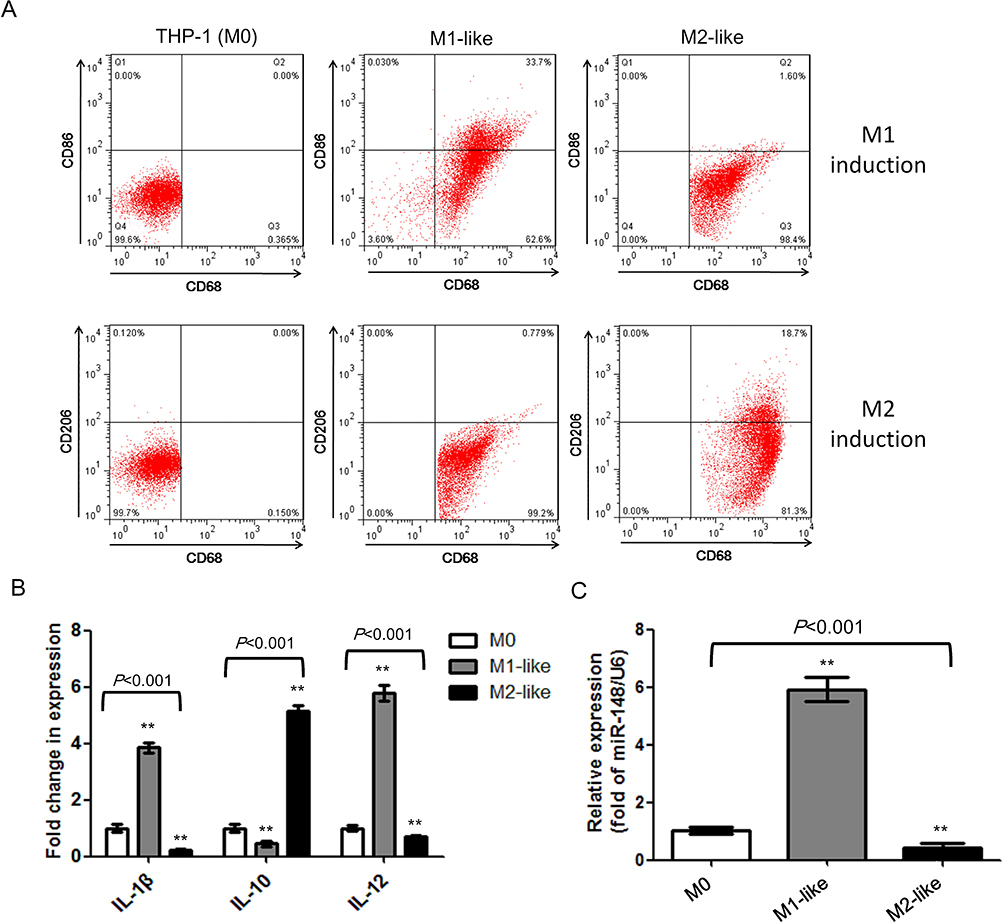 |
Figure 1 miR-148a is differentially expressed during macrophage polarization. (A) Detection of macrophage markers, CD68, CD86 and CD206, on cell surface by flow cytometry. (B) Detection of expression level of macrophage-related cytokines. (C) Detection of the expression level of miR-148a in different macrophages subtypes. **P<0.01 vs THP-1. |
miR-148a Induces Re-Polarization from M2-Like Macrophages to M1-Like Macrophages
Next, we investigated the effect of miR-148a on macrophage re-polarization. To provoke or block miR-148a expression, M0 cells were transfected with mimics or inhibitor of miR-148a, respectively (Figure 2A). Then, polarization was induced in M0 cells, and the expression of specific cytokines and surface antigens were evaluated. Transfecting miR-148a mimics significantly increased the expression of M1 macrophage-related IL-1β and IL-12, while transfecting miR-148a inhibitor increased the expression of M2 macrophage-related IL-10 (Figure 2B). miR-148a over-expression significantly increased the percentage of CD86+ cells, while inhibiting miR-148a increased the percentage of CD206+ cells (Figure 2C). CD68 expression was not affected by the different transfectants. The results suggest that M1-like macrophages were re-polarized to M2-like phenotypes when down-regulating miR-148a and up-regulating miR-148a was beneficial for the re-polarization of the M2-like phenotype to the M1-like phenotype.
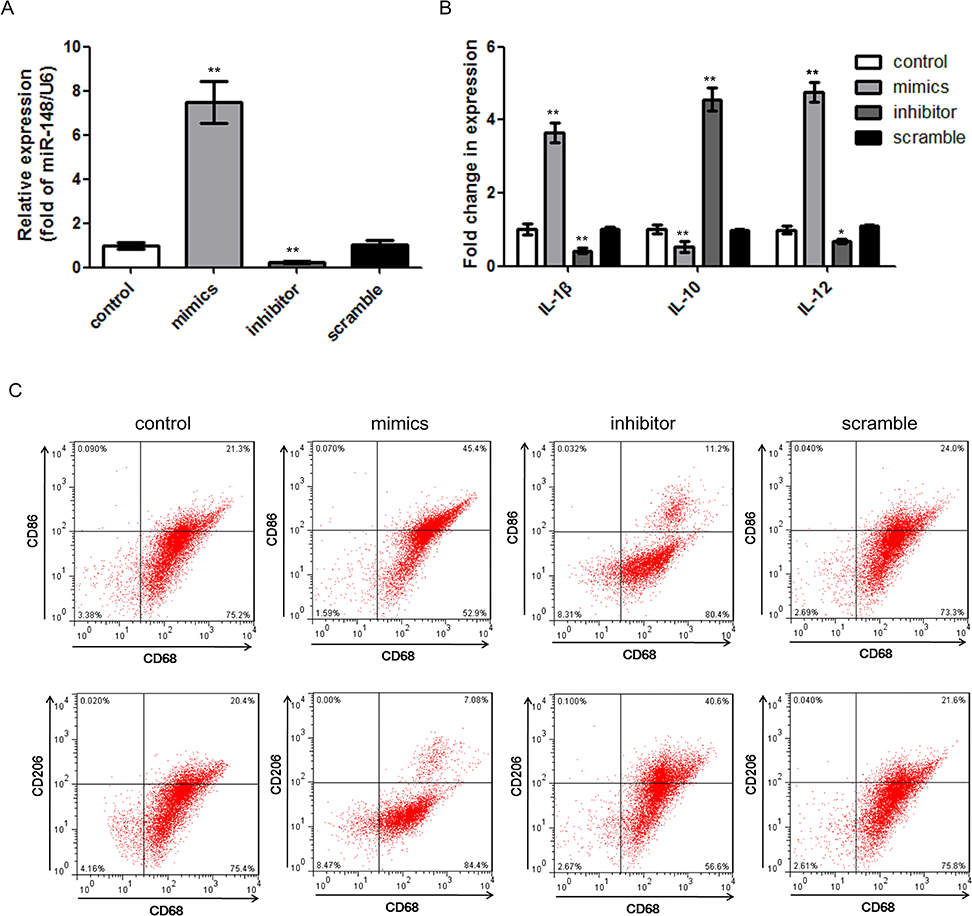 |
Figure 2 miR-148a effects on re-polarization ofM1-like or M2-like macrophages. (A) Expression of miR-148a in different transfectants of M0 cells. (B) Expression level of macrophage-related cytokines in different transfectants. (C) Re-polarization of M2-like macrophages into M1-like macrophages by increasing miR-148s and M1-like macrophages into M2-like macrophages by inhibiting miR-148a, respectively. **P<0.01 vs M0 control; *P<0.05 vs M0 control. |
miR-148a Reduced Recruitment of Macrophages by Targeting SIRPα
A previous study has shown that there exist binding sites between the 3ʹ-UTR region of SIRPα and miR-148a.20 SIRPα is a receptor on the surface of macrophages, which can be recognized and bound by CD47 on the surface of tumor cells. Typically, CD47 is highly expressed on the surface of tumor cells. Through combination with SIRPα, tumor cells can avoid phagocytosis by macrophages, thus achieving immune escape.21 In other words, SIRPα is a negative regulator of macrophage phagocytosis. We hypothesized that miR-148a might affect the biological activity of macrophages by regulating the SIRPα expression level. We first detected the SIRPα expression levels in different macrophage subtypes (Figure 3A). Compared with M0 cells, SIRPα was down-regulated in M1-like cells and up-regulated in M2-like cells; the difference was significant (P=0.002). We performed a dual-luciferase reporter assay and confirmed that miR-148a could directly target SIRPα (Figure 3B and C).
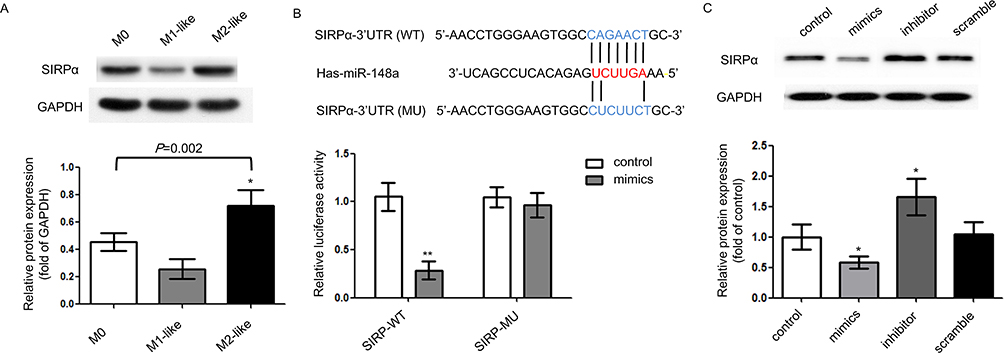 |
Figure 3 miR-148a directly targets SIRPα. (A) SIRPα expression in different macrophage subtypes. (B) Direct regulation between miR-148a and SIRPα by dual-luciferase reporter assay. (C) Relative protein expression of SIRPα in different transfectants. **P<0.01 vs control; *P<0.05 vs control. |
Then, we investigated whether miR-148a expression affected the recruitment of macrophages using a co-culture system. The effect of co-culture on the macrophage polarization is shown in Figure 4A. Compared with solely cultured THP-1 (M0 state), THP-1 cells co-cultured with SW480 cells resembled the characteristics of M2-like macrophages, ie, low expression of IL-1β and IL-12, and high expression of IL-10. In other words, SW480-conditioned THP-1 cells differentiated into TAMs. Over-expression of miR-148a in TAMs promoted the expression of cytokines similar to the profile of M1-like macrophages. A chemotaxis assay (Figure 4B) showed that low expression of miR-148a could enhance the transmembrane behavior of macrophages, ie, more macrophages could be recruited by cancer cells. Simultaneously, the induction of SW480 cell apoptosis was decreased (Figure 4C). These results suggest that over-expression of miR-148a, which was associated with low expression of SIRPα, could reduce transmembrane signaling in macrophages, inducing further apoptosis in cancer cells.
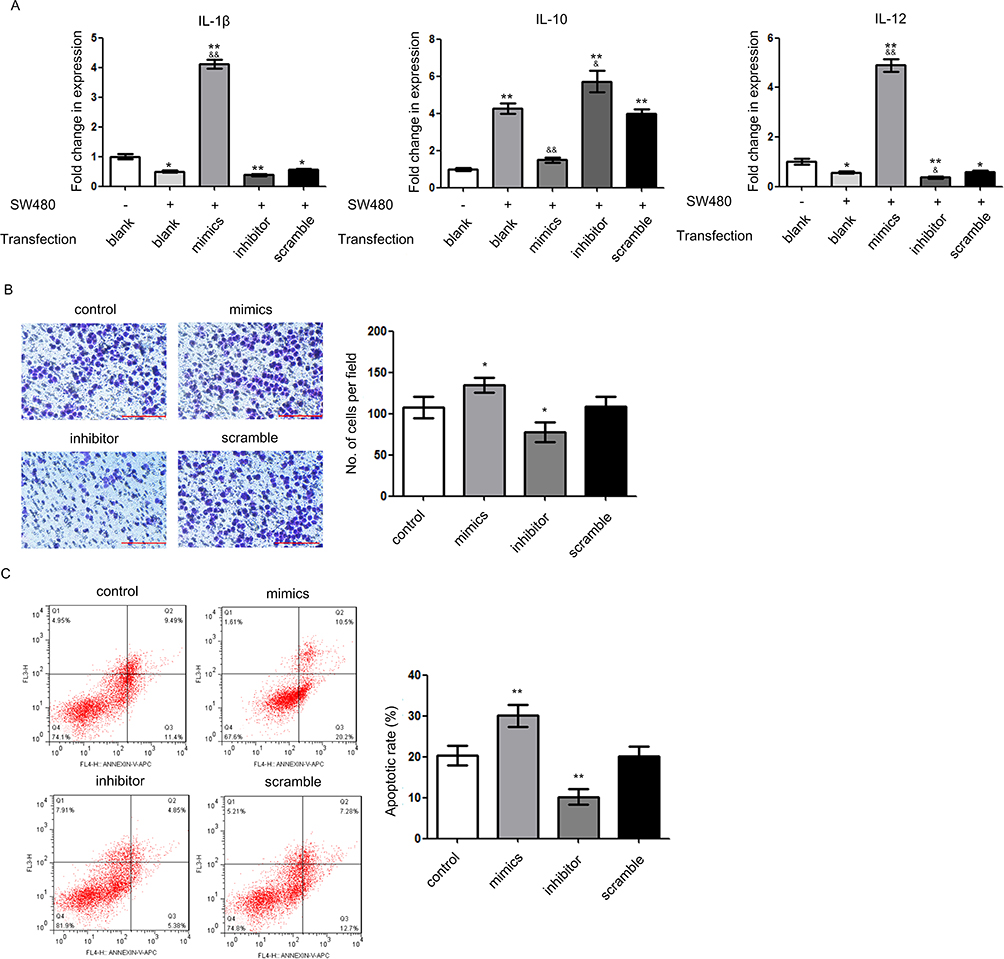 |
Figure 4 miR-148a reduces recruitment of tumor-associated macrophages. (A) Expression level of macrophage-related cytokines in different co-culture systems. *P<0.05 vs blank control without SW480 co-culture; **P<0.01 vs blank control without SW480 co-culture; &P<0.05 vs blank control with SW480 co-culture; &&P<0.01 vs blank control with SW480 co-culture. (B) Migration ability of different transfectants by chemotaxis assay. Scale bar=200 μm. *P<0.05 vs control. (C) Induction of tumor cell apoptosis by different transfectants. **P<0.01 vs control. |
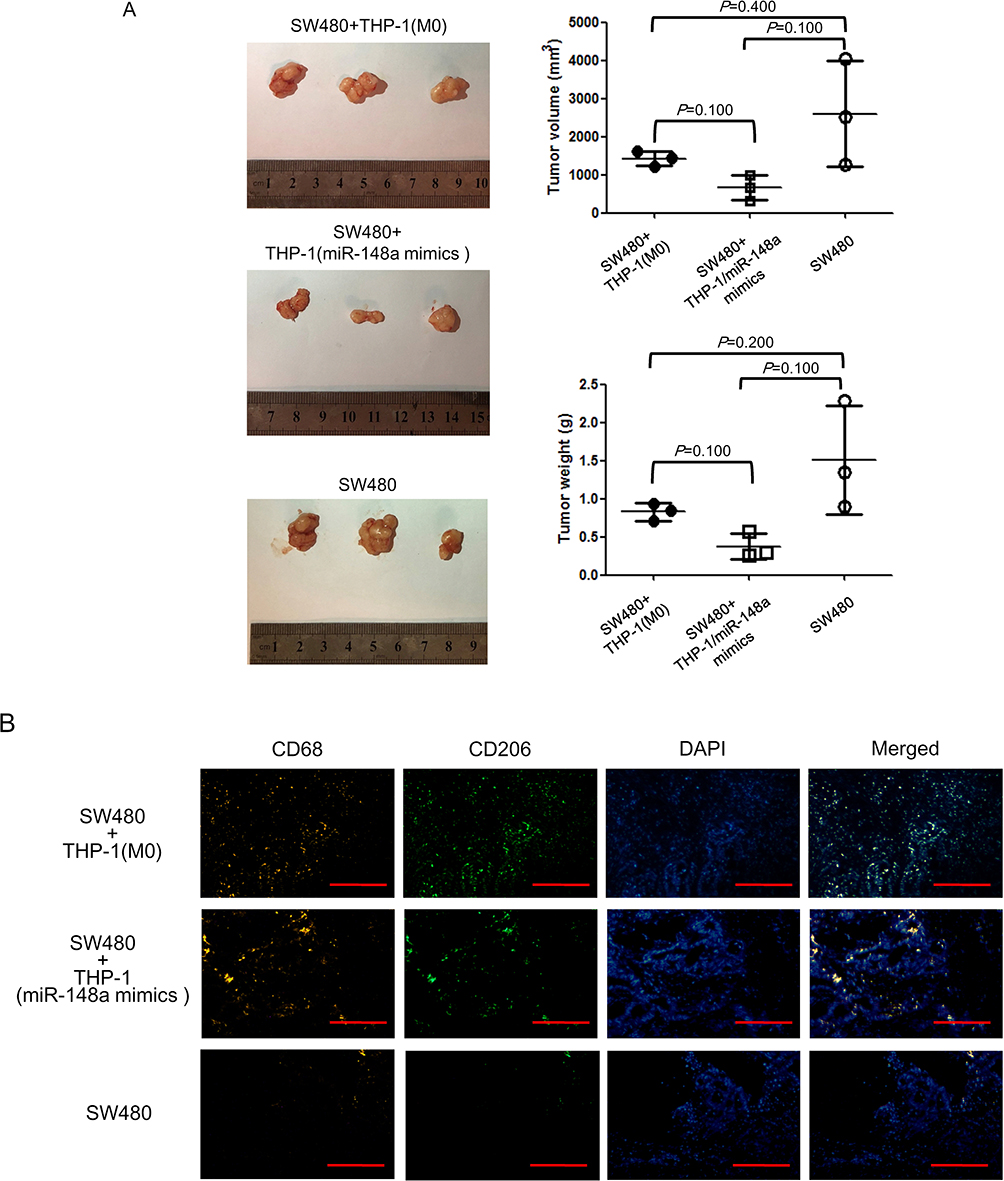 |
Figure 5 miR-148a enhances anti-tumor effect of macrophages in vivo. (A) Xenograft assay in nude mice. (B) Infiltration of macrophages in subcutaneous tumor by immunofluorescence assay. Scale bar=200 μm. |
miR-148a Enhanced the Anti-Tumor Effect of Macrophages in vivo
In the above results, we demonstrated that low expression of miR-148a was beneficial for the polarization of M2-like macrophages, as well as macrophage recruitment. In order to further determine the effect of miR-148a on the in vivo function of macrophages, we induced untransfected THP-1 cells and THP-1 cells over-expressing miR-148a into M0 status, mixed with SW480 cells, and inoculated them into nude mice. As shown in Figure 5A the subcutaneous tumors of mice inoculated with SW480 cells were relatively large. Following inoculation with the mixture of SW480 cells and macrophages, the subcutaneous tumor size was reduced. Furthermore, compared with the co-inoculation of THP-1 cells, the co-inoculation of THP-1 cells transfected with miR-148a mimics was more effective in inhibiting subcutaneous tumor formation. However, the difference among the three groups was not significant, which could be attributed to the small sample size of animals. In subcutaneous tumors, the expression of TAM-related CD markers was detected by immunofluorescence (Figure 5A). The pan-macrophage marker, CD68, was generally expressed in co-inoculated macrophages, while the M2 macrophage marker, CD206, was reduced in co-inoculated THP-1 cells transfected with miR-148a mimics. The results further suggest that increasing miR-148a expression in macrophages helps inhibit the production of M2-like macrophages, thus enhancing the anti-tumor effect of macrophages in vivo.
Discussion
Several clinical and preclinical studies have shown that the recruitment and infiltration of TAMs are crucial for the occurrence and prognosis of cancer.22,23 A large cohort study of colorectal cancer patients has revealed that decreased lymphatic vessels and reduced immune cytotoxicity were “two hallmarks of the metastatic process”, suggesting that the immune status was a more important factor contributing to cancer spread than the genomic instability of cancer cells.24 The current study revealed a potential mechanism of TAM regulation. Over-expression of miR-148a could suppress colorectal cancer by inhibiting polarization of M2-like macrophages, reducing the recruitment of macrophages via SIRPα targeting, and further strengthening the killing effect of macrophages both in vitro and in vivo.
The anti-tumor effect of miR-148a has been extensively investigated in multiple cancers. Most of the studies have discussed the effect of miR-148a on the biological characteristics of tumor cells. In colorectal cancer, miR-148a inhibits cell proliferation, reduces migration, and promotes apoptosis by targeting Bcl-2 and ErbB3.25,26 miR-148a suppresses the expression of stem cell markers and enhances chemoresistance of SW480 cells by regulating the WNT10b and β-catenin signaling pathways.27 Our study further investigated the effect of miR-148a on TAMs. Our results suggest that miR-148a can promote M1 macrophage polarization and regulate the secretion of inflammatory cytokine markers. Similar results have been reported in a recent study by Huang et al, where miR-148a promoted M1 polarization of murine bone marrow-derived macrophages and enhanced the killing ability of macrophages upon Notch activation; over-expression of miR-148a increased production of reactive oxygen species by directly targeting phosphatase and tensin homolog (PTEN).28
In addition to affecting macrophage polarization, we also observed that miR-148a could affect macrophage recruitment by targeting SIRPα. The CD47-SIRPα interaction is crucial for the immune escape of tumor cells. Tumor cells recognize and bind to SIRPα on macrophages through CD47 surface antigen, to transmit “don’t eat me” signal to macrophages.29 Blocking the CD47-SIRPα signaling pathway could be developed as an attractive candidate strategy for cancer therapy. In several preclinical studies, the application of anti-SIRPα antibodies has enhanced the phagocytosis of tumor cells, and this anti-tumor action was significantly associated with selective depletion of macrophages.30–34 Currently, little is known regarding the miRNA regulation of SIRPα expression. Another study has identified SIRPA, which encodes SIPRα, as a common target gene of miR-17, miR-20a, and miR-106a.35 In macrophages, the up-regulation of these three miRNAs led to SIRPα reduction and macrophage activation,35 which is consistent with our results. These studies indicate that miRNAs can be an alternative regulatory factor in SIRPα blocking. Another possible regulatory mechanism is that tumor cells regulate macrophage polarization through exosome derived miRNAs.36–38 Through co-culture of macrophages and SW480 cells, we observed that macrophages differentiated into M2-like state. This co-culture system has been used in various researches as an in vitro model of TAM induction.13,39,40 However, this experiment only suggests that some substances secreted by tumor cells affect macrophage polarization, not necessarily the exosome derived miRNAs.
Collectively, the findings of this study suggest that miR-148a can play a multiple-level anti-tumor role by participating in macrophage polarization and recruitment. However, data regarding the regulatory effect of miR-148a on TAMs remain limited. Further studies are crucial to determine whether miR-148a regulates macrophage proliferation and elucidate underlying mechanisms. Simultaneously, considering that TAMs are only a part of the tumor microenvironment, the regulatory effect of miR-148a on other microenvironment components, such as cancer-associated fibroblasts and stromal endothelial cells, deserves further in-depth investigation.
Disclosure
The authors report no conflicts of interests in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
3. Squadrito ML, De Palma M. Macrophage regulation of tumor angiogenesis: implications for cancer therapy. Mol Aspects Med. 2011;32(2):123–145. doi:10.1016/j.mam.2011.04.005
4. Qian B, Pollard JW. Macrophage diversity enhances tumor progression and metastasis. Cell. 2010;141(1):39–51. doi:10.1016/j.cell.2010.03.014
5. Riabov V, Gudima A, Wang N, Mickley A, Orekhov A, Kzhyshkowska J. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front Physiol. 2014;5:75. doi:10.3389/fphys.2014.00075
6. Afik R, Zigmond E, Vugman M, et al. Tumor macrophages are pivotal constructors of tumor collagenous matrix. J Exp Med. 2016;213(11):2315–2331. doi:10.1084/jem.20151193
7. Vinnakota K, Zhang Y, Selvanesan BC, et al. M2‐like macrophages induce colon cancer cell invasion via matrix metalloproteinases. J Cell Physiol. 2017;232(12):3468–3480. doi:10.1002/jcp.25808
8. Mantovani A, Schioppa T, Porta C, Allavena P, Sica A. Role of tumor-associated macrophages in tumor progression and invasion. Cancer Metastasis Rev. 2006;25(3):315–322. doi:10.1007/s10555-006-9001-7
9. Genard G, Lucas S, Michiels C. Reprogramming of tumor-associated macrophages with anticancer therapies: radiotherapy versus chemo- and immunotherapies. Front Immunol. 2017;8:828. doi:10.3389/fimmu.2017.00828
10. Yang J, Zhang Z, Chen C, et al. MicroRNA-19a-3p inhibits breast cancer progression and metastasis by inducing macrophage polarization through downregulated expression of Fra-1 proto-oncogene. Oncogene. 2014;33(23):3014–3023. doi:10.1038/onc.2013.258
11. Wang X, Luo G, Zhang K, et al. Hypoxic tumor-derived exosomal miR-301a mediates M2 macrophage polarization via PTEN/PI3Kγ to promote pancreatic cancer metastasis. Cancer Res. 2018;78(16):4586–4598. doi:10.1158/0008-5472.CAN-17-3841
12. Takahashi M, Cuatrecasas M, Balaguer F, et al. The clinical significance of MiR-148a as a predictive biomarker in patients with advanced colorectal cancer. PLoS One. 2012;7(10):e46684. doi:10.1371/journal.pone.0046684
13. Genin M, Clement F, Fattaccioli A, Raes M, Michiels C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer. 2015;15:577. doi:10.1186/s12885-015-1546-9
14. Iqbal AJ, Mcneill E, Kapellos TS, et al. Human CD68 promoter GFP transgenic mice allow analysis of monocyte to macrophage differentiation in vivo. Blood. 2014;124(15):e33–444. doi:10.1182/blood-2014-04-568691
15. Ying W, Cheruku PS, Bazer FW, Safe SH, Zhou B. Investigation of macrophage polarization using bone marrow derived macrophages. J Vis Exp. 2013;(76):50323.
16. Igarashi Y, Nawaz A, Kado T, et al. Partial depletion of CD206-positive M2-like macrophages induces proliferation of beige progenitors and enhances browning after cold stimulation. Sci Rep. 2018;8(1):14567. doi:10.1038/s41598-018-32803-6
17. Deng Y, Wang J, Huang M, Xu G, Wei W, Qin H. Inhibition of miR‐148a‐3p resists hepatocellular carcinoma progress of hepatitis C virus infection through suppressing c‐Jun and MAPK pathway. J Cell Mol Med. 2019;23(2):1415–1426. doi:10.1111/jcmm.14045
18. Naito S, von Eschenbach AC, Giavazzi R, Fidler IJ. Growth and metastasis of tumor cells isolated from a human renal cell carcinoma implanted into different organs of nude mice. Cancer Res. 1986;46(8):4109–4115.
19. Sun X, Gao D, Gao L, et al. Molecular imaging of tumor-infiltrating macrophages in a preclinical mouse model of breast cancer. Theranostics. 2015;5(6):597–608. doi:10.7150/thno.11546
20. Xu M, Qin Y, Qu J, et al. Evaluation of five candidate genes from GWAS for association with oligozoospermia in a Han Chinese population. PLoS One. 2013;8(11):e80374. doi:10.1371/journal.pone.0080374
21. Barclay AN, Van den Berg TK. The interaction between signal regulatory protein alpha (SIRPα) and CD47: structure, function, and therapeutic target. Annu Rev Immunol. 2014;32:25–50. doi:10.1146/annurev-immunol-032713-120142
22. Fridman WH, Pages F, Sautes-Fridman C, Galon J. The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer. 2012;12(4):298–306. doi:10.1038/nrc3245
23. Tauriello DVF, Calon A, Lonardo E, Batlle E. Determinants of metastatic competency in colorectal cancer. Mol Oncol. 2017;11(1):97–119. doi:10.1002/1878-0261.12018
24. Mlecnik B, Bindea G, Kirilovsky A, et al. The tumor microenvironment and immunoscore are critical determinants of dissemination to distant metastasis. Sci Transl Med. 2016;8(327):327ra26. doi:10.1126/scitranslmed.aad6352
25. Zhang H, Li Y, Huang Q, et al. MiR-148a promotes apoptosis by targeting Bcl-2 in colorectal cancer. Cell Death Differ. 2011;18(11):1702–1710. doi:10.1038/cdd.2011.28
26. Zhao W, Zheng J, Wei G, Yang K, Wang G, Sun X. miR‑148a inhibits cell proliferation and migration through targeting ErbB3 in colorectal cancer. Oncol Lett. 2019;18(3):2530–2536. doi:10.3892/ol.2019.10581
27. Shi L, Xi J, Xu X, Peng B, Zhang B. MiR-148a suppressed cell invasion and migration via targeting WNT10b and modulating β-catenin signaling in cisplatin-resistant colorectal cancer cells. Biomed Pharmacother. 2019;109:902–909. doi:10.1016/j.biopha.2018.10.080
28. Huang F, Zhao J, Wang L, et al. miR-148a-3p mediates Notch signaling to promote the differentiation and M1 activation of macrophages. Front Immunol. 2017;8:1327. doi:10.3389/fimmu.2017.01327
29. Murata Y, Kotani T, Ohnishi H, Matozaki T. The CD47-SIRPα signalling system: its physiological roles and therapeutic application. J Biochem. 2014;155(6):335–344. doi:10.1093/jb/mvu017
30. Yanagita T, Murata Y, Tanaka D, et al. Anti-SIRPα antibodies as a potential new tool for cancer immunotherapy. JCI Insight. 2017;2(1):e89140. doi:10.1172/jci.insight.89140
31. Murata Y, Tanaka D, Hazama D, et al. Anti‐human SIRPα antibody is a new tool for cancer immunotherapy. Cancer Sci. 2018;109(5):1300–1308. doi:10.1111/cas.13548
32. Zhao XW, van Beek EM, Schornagel K, et al. CD47-signal regulatory protein-α (SIRPα) interactions form a barrier for antibody-mediated tumor cell destruction. Proc Natl Acad Sci U S A. 2011;108(45):18342–18347. doi:10.1073/pnas.1106550108
33. Bener G, Felix A, Sanchez de Diego C, Pascual Fabregat I, Ciudad CJ, Noe V. Silencing of CD47 and SIRPα by Polypurine reverse Hoogsteen hairpins to promote MCF-7 breast cancer cells death by PMA-differentiated THP-1 cells. BMC Immunol. 2016;17(1):32. doi:10.1186/s12865-016-0170-z
34. Ring NG, Herndler-Brandstetter D, Weiskopf K, et al. Anti-SIRPα antibody immunotherapy enhances neutrophil and macrophage antitumor activity. Proc Natil Acad Sci U S A. 2017;114(49):E10578–E10585. doi:10.1073/pnas.1710877114
35. Zhu D, Pan C, Li L, et al. MicroRNA-17/20a/106a modulate macrophage inflammatory responses through targeting signal-regulatory protein α. J Allergy Clin Immunol. 2013;132(2):426–436. doi:10.1016/j.jaci.2013.02.005
36. Wang D, Wang X, Si M, et al. Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages. Cancer Lett. 2020;474:36–52. doi:10.1016/j.canlet.2020.01.005
37. Ham S, Lima LG, Chai EPZ, et al. Breast cancer-derived exosomes alter macrophage polarization via gp130/STAT3 signaling. Front Immunol. 2018;9:871. doi:10.3389/fimmu.2018.00871
38. Chen X, Ying X, Wang X, Wu X, Zhu Q, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer deliver microRNA-940 to induce macrophage M2 polarization. Oncol Rep. 2017;38(1):522–528. doi:10.3892/or.2017.5697
39. Wang Z, Xu L, Hu Y, et al. miRNA let-7b modulates macrophage polarization and enhances tumor-associated macrophages to promote angiogenesis and mobility in prostate cancer. Sci Rep. 2016;6(1):25602. doi:10.1038/srep25602
40. Gao Y, Fang P, Li WJ, et al. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol Immunol. 2020;117:20–28. doi:10.1016/j.molimm.2019.10.026
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.