Back to Journals » OncoTargets and Therapy » Volume 13
miR-100 Inhibits Cell Growth and Proliferation by Targeting HOXA1 in Nasopharyngeal Carcinoma
Authors He W, Huang Y, Jiang C, Zhu Y, Wang L ![]() , Zhang W, Huang W, Zhou T
, Zhang W, Huang W, Zhou T ![]() , Tang S
, Tang S
Received 26 August 2019
Accepted for publication 21 November 2019
Published 20 January 2020 Volume 2020:13 Pages 593—602
DOI https://doi.org/10.2147/OTT.S228783
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Tohru Yamada
Weifeng He, 1,* Yun Huang, 2,* Cheng chuan Jiang, 1 Yuan Zhu, 3 Ling Wang, 4 Weiwei Zhang, 1 Weiguo Huang, 2 Ting Zhou, 1, 5 Sanyuan Tang 1
1Department of Oncology, Brain Hospital of Hunan Province, Changsha 410007, Hunan Province, People’s Republic of China; 2Cancer Research Institute, Hengyang Medical College of University of South China, Hengyang 421001, Hunan Province, People’s Republic of China; 3People’s Hospital of Changshou Chongqing, Chongqing 401220, People’s Republic of China; 4Yi Chang Central People’s Hospital, Yichang 443000, Hubei Province, People’s Republic of China; 5Department of Clinical Pharmacy, College of Pharmacy, Hunan University of Traditional Chinese Medicine, Changsha 410007, Hunan Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Sanyuan Tang
Department of Oncology, Brain Hospital of Hunan Province, Changsha, Hunan Province, People’s Republic of China
Tel +86 15273156823
Email [email protected]
Ting Zhou
Department of Oncology, Brain Hospital of Hunan Province, Changsha, Hunan Province, People’s Republic of China
Tel/Fax +86 18374819626
Email [email protected]
Background: Increasing evidence indicates that the dysregulation of miRNAs plays a vital role in tumorigenesis and progression of nasopharyngeal carcinoma (NPC). Thus, it is necessary to further investigate the function and mechanism of miRNAs in NPC.
Methods: miR-100 expression was analyzed using publicly available databases and then tested using quantitative RT-PCR in NPC tissues and cell lines. MTT and colony formation assays and xenograft tumor model were used to test the NPC cell growth and proliferation abilities while modulating miR-100 expression. The target of miR-100 was predicted with TargetScan and validated with luciferase reporter assay, quantitative RT-PCR, and Western blot.
Results: The expression of miR-100 was significantly reduced in NPC tissues and cell lines. Overexpression of miR-100 obviously suppressed NPC cell growth and proliferation, whereas silencing miR-100 promoted NPC cell growth and proliferation in vitro. HOXA1 (homeobox A1) was validated as a direct target of miR-100, and restoring HOXA1 expression could reverse the inhibitive effect of miR-100 on NPC cell growth and proliferation. The mRNA and protein expression of HOXA1 was increased in NPC cell lines. Furthermore, ectopic expression of miR-100 inhibited xenograft tumor growth in vivo.
Conclusion: Taken together, our findings suggest that miR-100 could suppress NPC growth and proliferation through targeting HOXA1, providing a novel target for the miRNA-mediated therapy for patients with NPC in the future.
Keywords: miR-100, nasopharyngeal carcinoma, proliferation, invasion, HOXA1
Introduction
Nasopharyngeal carcinoma (NPC) is a malignant epithelial tumor with the highest incidence in China, which accounts for 47.7% of the new cases worldwide.1 As its unique anatomical location, radiotherapy is the primary treatment for NPC patients. During the past years, with the use of intensity-modulated radiation therapy and its combination with chemotherapy, the 5-year survival rate of NPC patients has been significantly improved. However, about 30% of NPC patients eventually die of relapse or metastasis.2 Therefore, it is very urgent to carry out relevant studies to clarify the underlying mechanism of NPC tumorigenesis and development and provide new molecular targets for future treatment.
MiRNAs are small non-coding RNAs, and it can enhance mRNA degradation or inhibit its translation through base pairing with the 3′-untranslated region (3′-UTR) of mRNA.3 Many studies demonstrate that miRNA expression changes significantly in various tumors, making it as biomarkers for tumor diagnosis, subclassify, and prognosis prediction.4–6 More importantly, the changed miRNAs play very important roles in the process of tumorigenesis, development, and progression.7–9 It has been also found that miRNA expression is dysregulated in NPC,10–12 and the dysregulated miRNAs can promote NPC cell proliferation, invasion, and angiogenesis, including miR-125b, miR-506, miR-142-3p, miR-23a, and so on.13–16 Thus, previous studies indicate that miRNA plays vital roles in NPC carcinogenesis and development. It also should be noted that further exploration of the function and mechanism of miRNA involved in NPC development and progression would provide novel therapeutic targets.
Based on analyzing several publicly available microarray data, we found that miR-100 was obviously decreased in NPC. It has been reported that miR-100 is dysregulated in many types of cancers, and it functions as either a tumor suppressor or promoter in different tumor types.17,18 In NPC, miR-100 was found to be decreased and it can suppress NPC cell migration and invasion through targeting IGF1R.19 However, no study has elucidated the function and mechanisms of miR-100 in NPC growth and proliferation. Therefore, in our present study, we performed functional studies and found that overexpression of miR-100 inhibited NPC cell growth and proliferation in vitro, while silencing miR-100 had an opposite function. Mechanism studies indicated that HOXA1 was a direct target of miR-100, and the restoration of HOXA1 expression reversed the inhibitive effect of miR-100 overexpression on NPC cell growth and proliferation. Our findings suggest that miR-100 functions as a tumor suppressor in NPC, which might serve as a treatment target for NPC patients.
Methods
Cell Culture
The NPC cell lines, including CNE1, CNE2, HNE1, HONE1, SUNE1, and C666-1, were maintained in RPMI-1640 medium (Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Gibco); and the human immortalized nasopharyngeal epithelial cell line NP69 was maintained in KSFM medium (Gibco) supplemented with bovine pituitary extract (BD Biosciences, San Diego, CA, USA) in a humidified atmosphere at 37°C with 5% CO2. All NPC cell lines and NP69 were generously provided by Professor Zhiqiang Xiao (Central South University), and the use of these cell lines was approved by the Institutional Ethical Review Board of the Brain Hospital of Human Province.
RNA Isolation and Quantitative RT-PCR
Total RNA was extracted using the TRIzol Reagent (Invitrogen, Carlsbad, CA, USA), and then the cDNA was synthesized using a reverse transcriptase (Promega, Madison, WI, USA). Quantitative PCR was conducted using the SYBR Green reagents (Invitrogen). The primers for miR-100 were purchased from the RiboBio (Guangzhou), and for HOXA1 was synthesized: GGCTCGCCTCAATACATTCAC (forward) and CGTACTCTCCAACTTTCCCTG (reverse). All of the experiments were conducted in triplicate and normalized to the expression of U6 or GAPDH. The relative expression was calculated with the 2− ΔΔCT equation.
Transient Transfection and Stably Transfected Cell Line Construction
The miR-100 mimics or inhibitor and its controls were purchased from the GenePharma (Suzhou). The HOXA1 plasmid and its empty vector were purchased from the FulenGen (Guangzhou). Cells were transfected using the Lipofectamine 2000 reagent (Invitrogen) and harvested for assays after 48 hrs. The sequence of pri-miR-100 was amplified and cloned into the pSin-EF2-puromycin lentiviral plasmid (Addgene, Cambridge, MA, USA). The lenti-miR-Ctrl or lenti-miR-100 was used to transfect SUNE1 cells and then selected using puromycin.
MTT and Colony Formation Assay
Cells were seeded into 96-well plates with a density of 1000 NPC cells per well, and then incubated for 0–4 days. On each day, the cells were stained with MTT dye (Sigma, St. Louis, MO, USA), and the absorbance was tested at 490 nm using a spectrophotometer. In addition, cells were seeded into 6-well plates with a density of 400 cells and cultured for 10 days. Colonies were fixed and stained, and then counted with an inverted microscope.
Luciferase Reporter Assay
The wild type (WT) and mutant (MT) 3′-UTR sequences of HOXA1 were amplified and inserted into the psiCHECK™ vector (Promega). Then, the reporter plasmids, together with p-TK Renilla plasmid and miR-100 mimics or control were used to transfect NPC cells using Lipofectamine 2000 (Invitrogen). Cells were harvested 48 h later, and the luciferase signals were tested using the Dual-Luciferase Reporter Assay System (Promega).
Western Blot
Total protein was extracted from NPC cells with RIPA buffer, separated with 9% gels, and then transfected to PVDF membranes (Millipore, Billerica, MA, USA). The membranes were firstly incubated with anti-HOXA1 antibody (Abcam, Cambridge, MA, USA), and then with secondary antibody (Sigma). The α-tubulin was used as a loading control, and the bands were obtained with enhanced chemiluminescence.
Xenograft Tumor Growth
Female BALB/c nude mice aged 3~4 weeks were purchased from the Charles River Laboratories (Beijing). SUNE1 cells stably expressing miR-100 or miR-Ctrl were injected into the dorsal flank of mice. The tumor size was measured every 3 days for the calculation of the tumor volume. On day 30, the mice were sacrificed, and the tumors were weighted. All animal study protocols were performed following the guidelines of the Experimental Animal Care and Use Ethics Committee of the Brain Hospital of Hunan Province.
Statistical Analyses
All experiments were conducted at least three times, and all results were shown as mean ± SD. SPSS 22.0 (SPSS Inc., Chicago, IL, USA) was used for data analysis by comparing with Student’s t-test, and p<0.05 was considered as a significant difference.
Results
miR-100 Is Reduced in NPC Clinical Samples and Cell Lines
We firstly analyzed the biggest publicly available NPC microarray data (GSE32960) and found that miR-100 was significantly reduced in NPC tissues (n=312) compared with normal nasopharynx tissues (n=18) (Figure 1A, p<0.001). Then, miR-100 was verified to be reduced in NPC tissues based on the other two publicly available microarray data with the accession number GSE36682 (Figure 1B, p<0.001) and GSE43039 (Figure 1C, p=0.049). In addition, we detected the miR-100 expression in the immortalized nasopharyngeal epithelial cell line NP69 and six NPC cell lines using quantitative RT-PCR, and found that miR-100 was obviously decreased in all NPC cell lines (Figure 1D, all p<0.05). These results indicate that miR-100 is reduced in NPC tissue samples and cell lines.
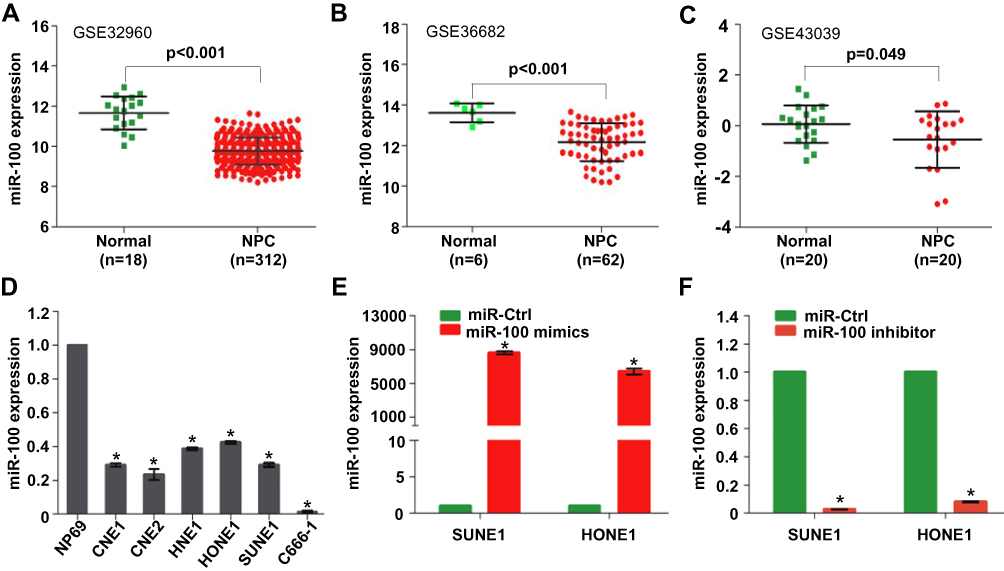 |
Figure 1 Mir-100 is reduced in NPC clinical samples and cell lines. (A–C) Relative miR-100 expression in NPC tissues and normal nasopharynx tissues in three publicly available microarray data. (D) Relative expression of miR-100 in the immortalized nasopharyngeal epithelial cell line NP69 and six NPC cell lines. (E, F) Relative expression of miR-100 in SUNE1 and HONE1 cells transfected with miR-100 mimics, miR-100 inhibitor or miR-Ctrl. U6 was used as the endogenous control. Data are presented as the mean ± SD, and the p values were calculated using the Student’s t-test; * p<0.05. |
Overexpression of miR-100 Inhibits NPC Cell Growth and Proliferation
We transiently transfected SUNE1 and HONE1 cells with miR-100 mimics or its control (miR-Ctrl), and confirmed the transfection efficiency with quantitative RT-PCR (Figure 1E, all p<0.05). Then, we did functional experiments including MTT and colony formation assays. MTT assay showed that NPC cells transfected with miR-100 mimics grew more slowly than cells transfected with miR-Ctrl (Figure 2A and B, p<0.05). Colony formation assay found that NPC cells transfected with miR-100 mimics formed fewer and smaller colonies than cells transfected with miR-Ctrl (Figure 2C and D, p<0.05). These results indicate that overexpression of miR-100 inhibits NPC cell growth and proliferation.
 |
Figure 2 Overexpression of miR-100 inhibits NPC cell growth and proliferation. (A, B) Cell growth viabilities of SUNE1 and HONE1 cells transfected with miR-100 mimics or miR-Ctrl determined by the MTT assay. (C–D) Representative images and quantification of colonies of SUNE1 and HONE1 cells transfected with miR-100 mimics or miR-Ctrl determined by the colony formation assay. Data are presented as the mean ± SD, and the p values were calculated using the Student’s t-test; * p<0.05. |
Silencing miR-100 Promotes NPC Cell Growth and Proliferation
We also transiently transfected SUNE1 and HONE1 cells with miR-100 inhibitor or its control (miR-Ctrl) and verified the transfection efficiency with quantitative RT-PCR (Figure 1F, p<0.05). Then, we also conducted the same functional experiments as mentioned above. We found that the reduction of miR-100 expression distinctly promoted SUNE1 and HONE1 cell growth and increased their colony formation rates (Figure 3A–D, p<0.05). These results indicate that silencing miR-100 promotes NPC cell growth and proliferation.
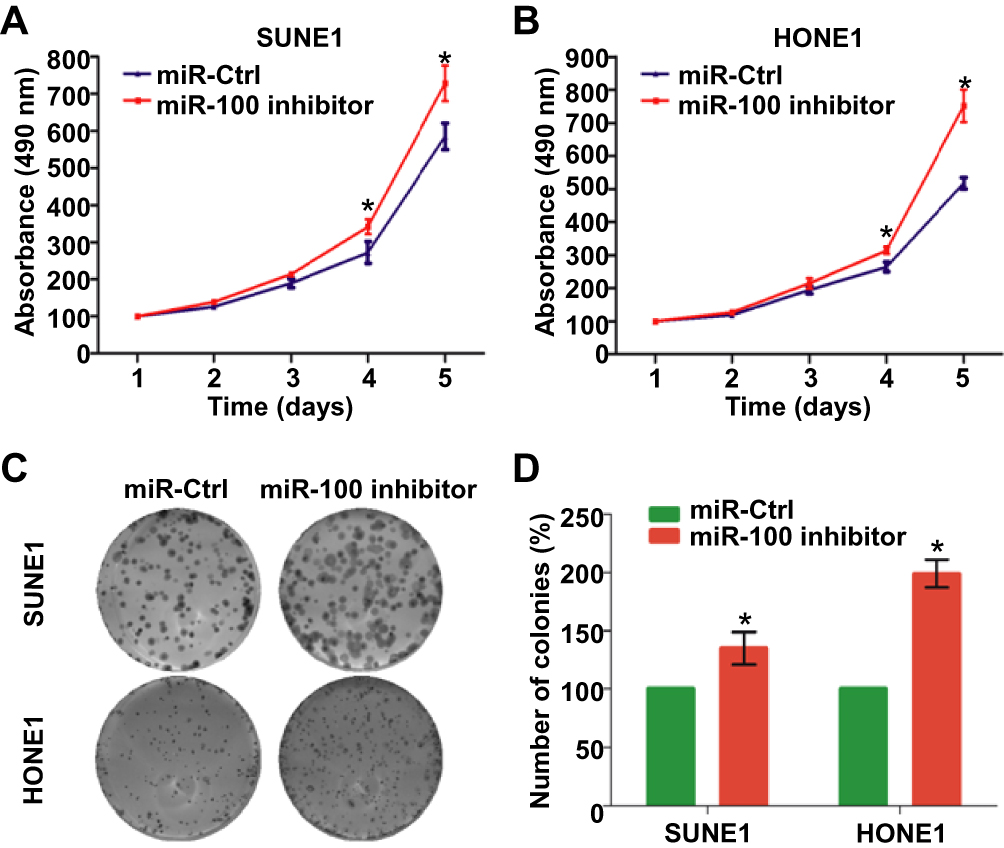 |
Figure 3 Silencing of miR-100 promotes NPC cell growth and proliferation. (A, B) Cell growth viabilities of SUNE1 and HONE1 cells transfected with miR-100 inhibitor or miR-Ctrl determined by the MTT assay. (C, D) Representative images and quantification of colonies of SUNE1 and HONE1 cells transfected with miR-100 inhibitor or miR-Ctrl determined by the colony formation assay. Data are presented as the mean ± SD, and the p values were calculated using the Student’s t-test; * p<0.05. |
HOXA1 Is a Direct Target of miR-100 and Its Expression Is Increased in NPC
We predicted the potential target of miR-100 using the online database TargetScan and selected HOXA1 for further validation because of its crucial effect on cell growth. Figure 4A shows the putative binding site between miR-100 and HOXA1. Luciferase report assay found that overexpression of miR-100 could inhibit the luciferase activities of the HOXA1 3′-UTR WT reporter plasmid, but not the HOXA1 3′-UTR MT reporter plasmid (Figure 4B, p<0.05). Then, quantitative PCR and Western blot showed that overexpression of miR-100 could suppress the mRNA and protein expression of HOXA1 (Figure 4C and D, p<0.05). Furthermore, HOXA1 mRNA and protein were upregulated in NPC cell lines (Figure 4E and F). These results indicate that HOXA1 is a direct target of miR-100 in NPC.
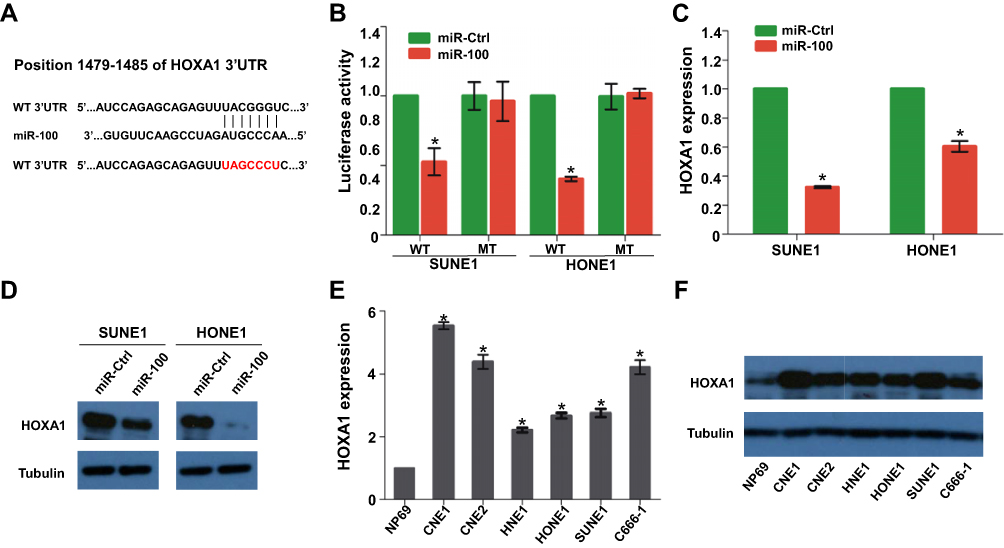 |
Figure 4 HOXA1 is a direct target of miR-100 and its expression is decreased in NPC (A) Wild-type (WT) and mutant (MT) 3′-UTR of HOXA1 with putative bind site of miR-100. (B) Relative luciferase activities of SUNE1 and HONE1 cells determined by the luciferase reporter assay. (C, D) The mRNA and protein expression of HOXA1 in SUNE1 and HONE1 cells transfected with miR-100 mimics or miR-Ctrl. (E, F) The mRNA and protein expression of HOXA1 in NPC cell lines determined by quantitative RT-PCR and Western blot. The data are presented as the mean ± SD; p values were calculated using Student’s t-test; * p<0.05. |
Restoration of HOXA1 Reverses the Inhibitive Effects of miR-100 in NPC
We then investigate whether HOXA1 was essential for the regulatory effect of miR-100 on NPC cell growth and proliferation. We transiently co-transfected SUNE1 and HONE1 cells with miR-100 mimics and HOXA1 plasmid or its empty vector, and then did MTT and colony formation assays. The functional results showed that the ectopic expression of HOXA1 reversed the inhibitive effects of miR-100 on NPC cell growth and proliferation (Figure 5A–D, p<0.05). These results indicate that HOXA1 is a functional target of miR-100 in NPC.
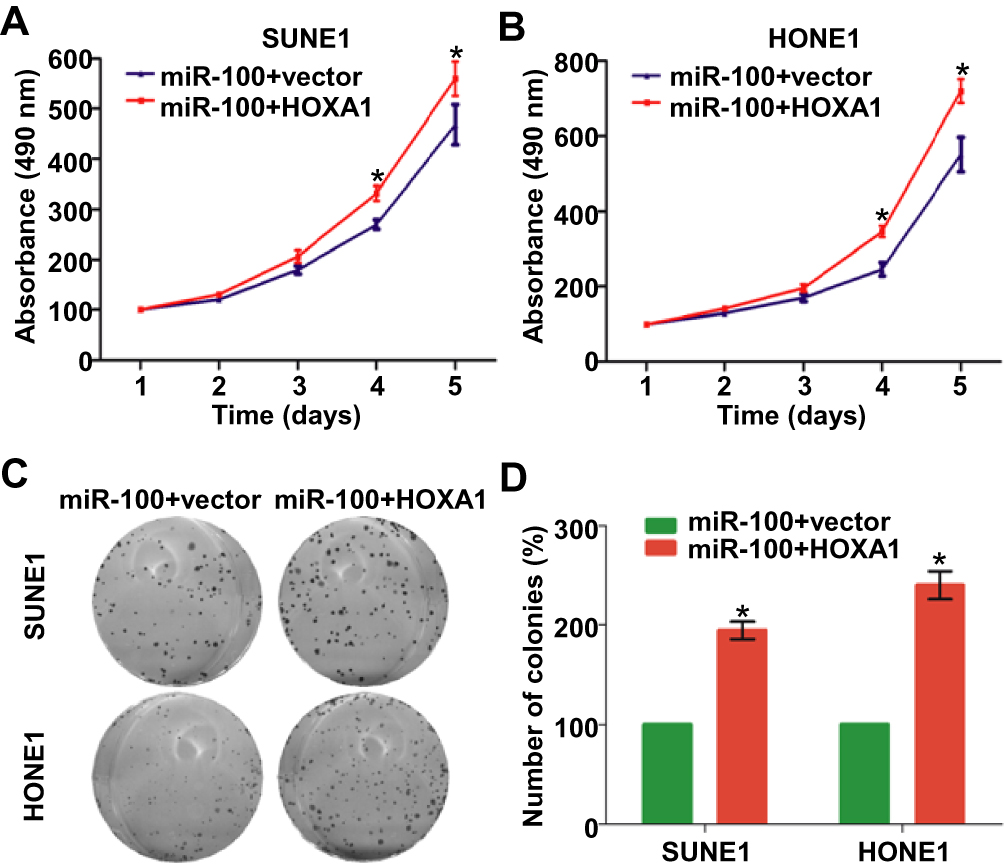 |
Figure 5 Restoration of HOXA1 reverses the inhibitive effect of miR-100 in NPC (A, B) Cell growth viabilities of SUNE1 and HONE1 cells transfected with miR-100 mimics and HOXA1 plasmids or empty vector determined by the MTT assay. (C, D) Representative images and quantification of colonies of SUNE1 and HONE1 cells transfected with miR-100 mimics and HOXA1 plasmids or empty vector determined by colony formation assay. Data are presented as the mean ± SD, and the p values were calculated using the Student’s t-test; * p<0.05. |
Overexpression of miR-100 Inhibits NPC Xenograft Tumor Growth
We constructed xenograft tumor growth model through injecting SUNE1 cells that stably expressing miR-100 or miR-Ctrl into the dorsal flank of nude mice, and all of the mice formed xenograft tumors (Figure 6A). The tumors formed in the miR-100 overexpression group grew more slowly and smaller than the miR-Ctrl group (Figure 6B, p<0.05). In addition, the tumor size and weight were significantly different between the two groups (Figure 6C and D, p<0.05). The expression of miR-100 was increased in xenograft tumor tissues of miR-100 overexpression group than the miR-Ctrl group (Figure 6E, p<0.05). These results indicate that overexpression of miR-100 inhibits NPC xenograft tumor growth.
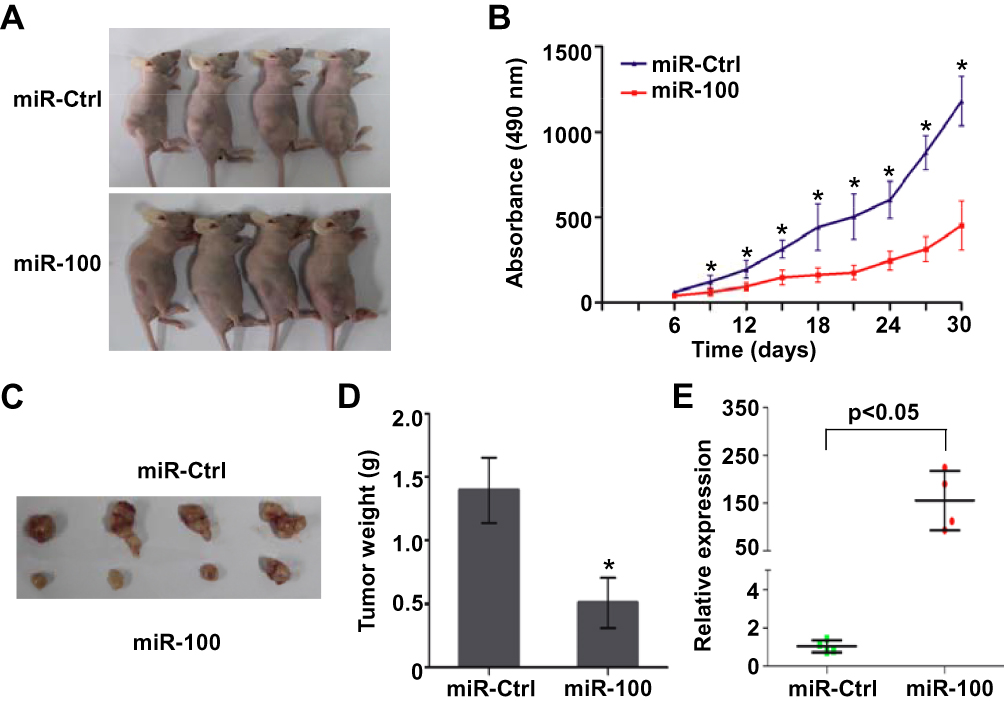 |
Figure 6 Overexpression of miR-100 inhibits NPC xenograft tumor growth (A) Xenograft tumor growth models in nude mice were constructed by injecting with SUNE1 cells stably expressing miR-100 or miR-Ctrl, and the xenograft tumors were formed. (B) The growth curves of the tumor volumes. (C) Representative images of the excised tumors. (D) The weights of the excised tumors. (E) Relative expression of miR-100 in xenograft tumors of the miR-100 overexpression group and miR-Ctrl group. Data are presented as the mean ± SD, and the p values were calculated using the Student’s t-test; * p<0.05. |
Discussion
Recurrence and metastasis are two major causes of treatment failure and death for patients with NPC, and it is urgent to better understanding the molecular mechanisms related to NPC tumorigenesis and progression, which would guide a more personalized therapy for NPC. In our present study, we reported that miR-100 was decreased in NPC and functioned as a tumor suppressor by inhibiting NPC cell growth and proliferation.
Recently, miRNAs have been reported to be dysregulated in NPC based on genome-wide profiling.10,12 The dysregulated miRNAs play an important role in NPC tumorigenesis and progression by promoting NPC cell proliferation, invasion, and angiogenesis.13–16 It has been found that miR-125b is upregulated in NPC, and it can promote NPC cell proliferation and inhibit cell apoptosis by targeting A20 and activating the NF-kB signaling pathway.13 miR-506 can inhibit NPC tumor growth and metastasis through inactivating the Wnt/β-catenin signaling pathway by targeting LHX2.14 EZH2-DNMT1-mediated epigenetic silencing of miR-142-3p promotes NPC cell invasion and metastasis by targeting ZEB2.15 miR-23a can promote NPC cell growth, migration, and angiogenesis by targeting TSGA10.16 Here, in our present study, we found that miR-100 was obviously decreased in NPC tissue samples by analyzing three publicly available microarray data. Functional experiments verified that miR-100 could suppress NPC cell growth and proliferation in vitro and inhibit xenograft tumor growth in vivo. It is worth noting that a recent study reports that miR-100 can inhibit NPC cell migration and invasion by targeting IGF1R.19 These findings indicate that miR-100 plays important roles in NPC development and progression.
miR-100, as a member of the miR-99 family, has been reported to be dysregulated in many types of cancers, and it can function as either a tumor suppressor or promoter, which depends on the tumor types and microenvironment.17,18 Downregulation of miR-100 has been found in esophageal squamous cell carcinoma, non-small cell lung cancer, breast cancer, and so on.20–22 On the other hand, upregulation of miR-100 is observed in small cell lung cancer, renal cell carcinoma, and pancreatic adenocarcinoma, and so on.23–25 It has been also found that miR-100 play vital roles in various biological processes, such as cell proliferation, apoptosis, cell cycle, migration, differentiation, and angiogenesis.17,18 For example, miR-100 inhibits breast cancer proliferation and survival by targeting IGF2.22 miR-100 inhibits gastric cancer tumor growth and metastasis and growth by targeting ZBTB7A.26 miR-100 promotes cell differentiation and survival in acute myeloid leukemia by targeting RBSP3.27 miR-100 promotes hepatocellular carcinoma cell metastasis by enhancing ICMT-Rac1 signaling.28 In this study, we found that miR-100 was decreased in NPC and functioned as a tumor-suppressive miRNA, enriching the understanding of the function and mechanism of miR-100 in tumors.
As we have known, miRNAs exert their function by base-pairing with the 3′-UTR of their target genes.3 Many target genes of miR-100 have been identified and verified, including mTOR, IGFR, PLK1, AKT1, RAP1B, FGFR3, and so on.29–34 As each miRNA can regulate multiple different target genes, our study predicted HOXA1 as a potential direct target of miR-100 with the public available database TargetScan. We then verified HOXA1 as the target of miR-100 in NPC using luciferase report assay, quantitative RT-PCR and Western blot. Similarly, HOXA1 was also found to be a direct target of miR-100 in lung cancer and breast cancer.23,35 It should be noted that HOXA1 can be regulated by several other miRNAs, such as miR-30b, miR-99a, and miR-577, and so on.36–38 More importantly, HOXA1 has been reported as an oncogene in various types of cancers.39–41 In our study, we found that HOXA1 was increased in NPC and it was regulated by miR-100, thereby enriching the understanding of miR-100/HOXA1 signaling pathway involvement in NPC tumorigenesis and progression.
In conclusion, our study revealed that miR-100 expression was reduced, and functioned as a tumor suppressor in NPC. Ectopic expression of miR-100 could suppress NPC cell growth and proliferation in vitro, and inhibit xenograft tumor growth in vivo by targeting HOXA1. Our findings make a better understanding of the mechanisms involved in miR-100-mediated NPC tumorigenesis and progression, further providing an opportunity for the exploration of novel miRNA-based targeted treatments for NPC patients.
Acknowledgments
This work was supported by grants from Health Commission of Hunan Province (20190339 and B20180849); Natural Science Foundation of Hunan Province (2018JJ6120 and 2019JJ40159); and The Foundation of Brain Hospital of Hunan Province (2018). The funders had no role in the study design, data collection, analysis, decision to publish or the preparation of the manuscript.
Author Contributions
TZ and ST designed the study. WH, YH, CJ, YZ, WL, and WZ carried out all of the experiments, prepared figures and drafted the manuscript. All authors participated in the data analysis and the interpretation of all of the results, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
2. Sun X, Su S, Chen C, et al. Long-term outcomes of intensity-modulated radiotherapy for 868 patients with nasopharyngeal carcinoma: an analysis of survival and treatment toxicities. Radiother Oncol. 2014;110:398–403. doi:10.1016/j.radonc.2013.10.020
3. Vishnoi A, Rani S. MiRNA biogenesis and regulation of diseases: an overview. Methods Mol Biol. 2017;1509:1–10.
4. Kong YW, Ferland-McCollough D, Jackson TJ, Bushell M. microRNAs in cancer management. Lancet Oncol. 2012;13(6):e249–e258. doi:10.1016/S1470-2045(12)70073-6
5. Chen B, Xia Z, Deng YN, et al. Emerging microRNA biomarkers for colorectal cancer diagnosis and prognosis. Open Biol. 2019;9(1):180212. doi:10.1098/rsob.180212
6. Adhami M, Haghdoost AA, Sadeghi B, Malekpour Afshar R. Candidate miRNAs in human breast cancer biomarkers: a systematic review. Breast Cancer. 2018;25(2):198–205. doi:10.1007/s12282-017-0814-8
7. Ebert MS, Sharp PA. Roles of microRNAs in conferring robustness to biological processes. Cell. 2012;149(3):515–524. doi:10.1016/j.cell.2012.04.005
8. Rupaimoole R, Calin GA, Lopez-Berestein G, Sood AK. miRNA deregulation in cancer cells and the tumor microenvironment. Cancer Discov. 2016;6(3):235–246. doi:10.1158/2159-8290.CD-15-0893
9. Lin S, Gregory RI. MicroRNA biogenesis pathways in cancer. Nat Rev Cancer. 2015;15(6):321–333. doi:10.1038/nrc3932
10. Chen HC, Chen GH, Chen YH, et al. MicroRNA deregulation and pathway alterations in nasopharyngeal carcinoma. Br J Cancer. 2009;100(6):1002–1011.
11. Li T, Chen JX, Fu XP, et al. microRNA expression profiling of nasopharyngeal carcinoma. Oncol Rep. 2011;25(5):1353–1363. doi:10.3892/or.2011.1204
12. Liu N, Chen NY, Cui RX, et al. Prognostic value of a microRNA signature in nasopharyngeal carcinoma: a microRNA expression analysis. Lancet Oncol. 2012;13(6):633–641. doi:10.1016/S1470-2045(12)70102-X
13. Zheng Z, Qu JQ, Yi HM, et al. MiR-125b regulates proliferation and apoptosis of nasopharyngeal carcinoma by targeting A20/NF-kB signaling pathway. Cell Death Dis. 2017;8(6):e2855. doi:10.1038/cddis.2017.211
14. Liang TS, Zheng YJ, Wang J, Zhao JY, Yang DK, Liu ZS. MicroRNA-506 inhibits tumor growth and metastasis in nasopharyngeal carcinoma through the inactivation of the Wnt/β-catenin signaling pathway by down-regulating LHX2. J Exp Clin Cancer Res. 2019;38(1):97. doi:10.1186/s13046-019-1023-4
15. Li YQ, He Q, Wen X, et al. EZH2-DNMT1-mediated epigenetic silencing of miR-142-3p promotes metastasis through targeting ZEB2 in nasopharyngeal carcinoma. Cell Death Differ. 2019;26(6):1089–1106. doi:10.1038/s41418-018-0208-2
16. Bao L, You B, Shi S, et al. Metastasis-associated miR-23a from nasopharyngeal carcinoma-derived exosomes mediates angiogenesis by repressing a novel target gene TSGA10. Oncogene. 2018;37(21):2873–2889. doi:10.1038/s41388-018-0183-6
17. Qin C, Huang RY, Wang ZX. Potential role of miR-100 in cancer diagnosis, prognosis, and therapy. Tumour Biol. 2015;36(3):1403–1409. doi:10.1007/s13277-015-3267-8
18. Li C, Gao Y, Zhang K, et al. Multiple roles of miroRNA-100 in human cancer and its therapeutic potential. Cell Physiol Biochem. 2015;37(6):2143–2159. doi:10.1159/000438572
19. Sun X, Liu X, Wang Y, Yang S, Chen Y, Yuan T. miR-100 inhibits the migration and invasion of nasopharyngeal carcinoma by targeting IGF1R. Oncol Lett. 2018;15(6):8333–8338. doi:10.3892/ol.2018.8420
20. Zhou SM, Zhang F, Chen XB, et al. miR-100 suppresses the proliferation and tumor growth of esophageal squamous cancer cells via targeting CXCR7. Oncol Rep. 2016;35(6):3453–3459. doi:10.3892/or.2016.4701
21. Liu J, Lu KH, Liu ZL, Sun M, De W, Wang ZX. MicroRNA-100 is a potential molecular marker of non-small cell lung cancer and functions as a tumor suppressor by targeting polo-like kinase 1. BMC Cancer. 2012;12:519. doi:10.1186/1471-2407-12-519
22. Gebeshuber CA, Martinez J. miR-100 suppresses IGF2 and inhibits breast tumorigenesis by interfering with proliferation and survival signaling. Oncogene. 2013;32(27):3306–3310. doi:10.1038/onc.2012.372
23. Xiao F, Bai Y, Chen Z, et al. Downregulation of HOXA1 gene affects small cell lung cancer cell survival and chemoresistance under the regulation of miR-100. Eur J Cancer. 2014;50:1541–1554. doi:10.1016/j.ejca.2014.01.024
24. Wang G, Chen L, Meng J, Chen M, Zhuang L, Zhang L. Overexpression of microRNA-100 predicts an unfavorable prognosis in renal cell carcinoma. Int Urol Nephrol. 2013;45:373–379. doi:10.1007/s11255-012-0374-y
25. Huang JS, Egger ME, Grizzle WE, McNally LR. MicroRNA-100 regulates IGF1-receptor expression in metastatic pancreatic cancer cells. Biotech Histochem. 2013;88:397–402. doi:10.3109/10520295.2012.762460
26. Shi DB, Wang YW, Xing AY, et al. C/EBPα-induced miR-100 expression suppresses tumor metastasis and growth by targeting ZBTB7A in gastric cancer. Cancer Lett. 2015;369(2):376–385. doi:10.1016/j.canlet.2015.08.029
27. Zheng YS, Zhang H, Zhang XJ, et al. MiR-100 regulates cell differentiation and survival by targeting RBSP3, a phosphatase-like tumor suppressor in acute myeloid leukemia. Oncogene. 2012;31(1):80–92. doi:10.1038/onc.2011.208
28. Zhou HC, Fang JH, Luo X, et al. Downregulation of microRNA-100 enhances the ICMT-Rac1 signaling and promotes metastasis of hepatocellular carcinoma cells. Oncotarget. 2014;5(23):12177–12188. doi:10.18632/oncotarget.2601
29. Xu C, Zeng Q, Xu W, et al. MiRNA-100inhibits human bladder urothelial carcinogenesis by directly targeting mTOR. Mol Cancer Ther. 2013;12:207–219. doi:10.1158/1535-7163.MCT-12-0273
30. Tovar V, Alsinet C, Villanueva A, et al. IGF activation in a molecular subclass of hepatocellular carcinoma and pre-clinical efficacy of IGF-1R blockage. J Hepatol. 2010;52:550–559. doi:10.1016/j.jhep.2010.01.015
31. Li BH, Zhou JS, Ye F, et al. ReducedmiR-100 expression in cervical cancer and precursors and its carcinogenic effect through targeting PLK1 protein. Eur J Cancer. 2011;47:2166–2174. doi:10.1016/j.ejca.2011.04.037
32. Jin Y, Tymen SD, Chen D, et al. MicroRNA-99 family targets AKT-mTOR signaling pathway in dermal wound healing. PLoS ONE. 2013;8(5):e64434. doi:10.1371/journal.pone.0064434
33. Peng H, Luo J, Hao H, et al. MicroRNA-100 regulates SW620 colorectal cancer cell proliferation and invasion by targeting RAP1B. Oncol Rep. 2014;31(5):2055–2062. doi:10.3892/or.2014.3075
34. Luan Y, Zhang S, Zuo L, Zhou L. Overexpression of miR-100 inhibits cell proliferation, migration, and chemosensitivity in human glioblastoma through FGFR3. Onco Targets Ther. 2015;8:3340–3391. doi:10.2147/OTT.S85677
35. Chen D, Sun Y, Yuan Y, et al. miR-100 induces epithelial-mesenchymal transition but suppresses tumorigenesis, migration and invasion. PLoS Genet. 2014;10:e1004177. doi:10.1371/journal.pgen.1004177
36. Li Q, Zhang X, Li N, Liu Q, Chen D. miR-30b inhibits cancer cell growth, migration, and invasion by targeting homeobox A1 in esophageal cancer. Biochem Biophys Res Commun. 2017;485(2):506–512. doi:10.1016/j.bbrc.2017.02.016
37. Wang JG, Tang WP, Liao MC, Liu YP, Ai XH. MiR-99a suppresses cell invasion and metastasis in nasopharyngeal carcinoma through targeting HOXA1. Onco Targets Ther. 2017;10:753–761. doi:10.2147/OTT.S126781
38. Han S, Liu Z, Wang Y, et al. MicroRNA-577 inhibits the migration and invasion of hepatocellular carcinoma cells by targeting homeobox A1. Oncol Rep. 2018;39(6):2987–2995. doi:10.3892/or.2018.6388
39. Wang H, Liu G, Shen D, et al. HOXA1 enhances the cell proliferation, invasion and metastasis of prostate cancer cells. Oncol Rep. 2015;34(3):1203–1210. doi:10.3892/or.2015.4085
40. Yuan C, Zhu X, Han Y, et al. Elevated HOXA1 expression correlates with accelerated tumor cell proliferation and poor prognosis in gastric cancer partly via cyclin D1. J Exp Clin Cancer Res. 2016;35:15. doi:10.1186/s13046-016-0294-2
41. Taminiau A, Draime A, Tys J, et al. HOXA1 binds RBCK1/HOIL-1 and TRAF1 and modulates the TNF/NF-kB pathway in a transcription-independent manner. Nucleic Acids Res. 2016;44(15):4349–7331. doi:10.1093/nar/gkw606
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.