")
Back to Journals » Infection and Drug Resistance » Volume 15
Minocycline Susceptibility and tetB Gene in Carbapenem-Resistant Acinetobacter baumannii in Taiwan
Authors Yang JL, Yang CJ, Chuang YC , Sheng WH, Chen YC , Chang SC
Received 6 January 2022
Accepted for publication 29 April 2022
Published 2 May 2022 Volume 2022:15 Pages 2401—2408
DOI https://doi.org/10.2147/IDR.S357344
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Jia-Ling Yang,1 Chia-Jui Yang,2,3 Yu-Chung Chuang,1 Wang-Huei Sheng,1 Yee-Chun Chen,1 Shan-Chwen Chang1
1Department of Internal Medicine, National Taiwan University Hospital, Taipei, Taiwan, Republic of China; 2Department of Internal Medicine, Far Eastern Memorial Hospital, New Taipei City, Taiwan; 3School of Medicine, National Yang Ming Chiao Tung University, Taipei, Taiwan
Correspondence: Yu-Chung Chuang, Division of Infectious Diseases, Department of Internal Medicine, National Taiwan University Hospital, No. 7 Chung-Shan South Road, Zhongzheng Dist, Taipei, 100225, Taiwan, Republic of China, Tel +886 2 2312 3456 ext 65054, Fax +886 2 2397 1412, Email [email protected]
Purpose: In this study, we evaluated the minocycline susceptibility rate in carbapenem-resistant Acinetobacter baumannii (CRAB) clinical strains, and the association between tetB carriage and minocycline susceptibility in CRAB.
Patients and Methods: A total of 100 genetically unrelated CRAB clinical strains from bloodstream infection were randomly collected from a medical center in Taiwan. An argument for a new minocycline susceptibility breakpoint of 1 mg/L was suggested based on pharmacokinetic (PK) and pharmacodynamic (PD) studies. Strains with minocycline minimum inhibitory concentrations (MICs) of > 1 mg/L were classified as PK-PD non-susceptible. TetB carriage was detected by polymerase chain reaction (PCR).
Results: Fifty-five (55%) CRAB strains were susceptible to minocycline according to the Clinical and Laboratory Standards Institute (CLSI) criteria, among which 98.2% (54/55) were PK-PD non-susceptible. The minocycline MIC50/90 was 4/16 mg/L. Ninety-seven (97%) strains carried tetB. All of the tetB-positive strains and 66.7% (2/3) of the tetB-negative strains were PK-PD non-susceptible. By statistical analysis, tetB carriage was significantly correlated with PK-PD non-susceptibility (P = 0.03) and a higher minocycline MIC (P = 0.02). The sensitivity and specificity of the tetB PCR for predicting PK-PD non-susceptibility were 98% and 100%, respectively.
Conclusion: At our institute, most CRAB strains were PK-PD non-susceptible and most carried tetB gene. Recognizing the minocycline MIC and tetB status may be essential when using minocycline to treat CRAB-related infections.
Keywords: CRAB, minocycline PK-PD non-susceptibility, multidrug resistance, efflux
Introduction
Carbapenem-resistant Acinetobacter baumannii (CRAB) represents an escalating global public health threat.1 A substantial proportion of the CRAB isolates are also extensively drug-resistant (XDR).2–4 The therapeutic options available for CRAB infections are limited, and the most effective and safe regimens remain unknown.5 Minocycline, a semisynthetic tetracycline derivative, is proven to exert in vitro antibacterial activity against A. baumannii.6 Clinical experiences suggest that minocycline is an attractive therapeutic option for drug-resistant A. baumannii infections.7,8 Among the XDR A. baumannii complex, minocycline is the second-most active agent after colistin, with a susceptibility rate of 67.3–72.7%.4,9 However, debates regarding the minocycline susceptibility breakpoint for A. baumannii remain. The Clinical and Laboratory Standards Institute (CLSI) minocycline breakpoint for A. baumannii is 4 mg/L. The European Committee on Antimicrobial Susceptibility Testing (EUCAST) has not defined a minocycline breakpoint for A. baumannii. Many experts have suggested the need for clinical validation or re-evaluation of the CLSI breakpoint.10,11 Although favorable clinical experiences are reported in using minocycline at dose of 400 mg daily to treat A. baumannii infections, the cases are relatively few and most regimens employ minocycline combined a second or third agent rather than monotherapy.12 An in vitro pharmacodynamic (PD) study using a pharmacokinetic (PK) model demonstrates the emergence of resistance in A. baumannii strains with minocycline MICs of 3 or 4 mg/L under exposure to 400 mg/day minocycline.6 Based on the PK and PD studies,6,13 Tsakris et al have estimated a new susceptibility breakpoint of 1 mg/L for A. baumannii when daily dose of 400 mg minocycline is administered.11
Minocycline resistance in A. baumannii is mainly mediated by the narrow-spectrum efflux pump, TetB, which belongs to the major facilitator superfamily.14 A. baumannii strains lacking the tetB gene reportedly have lower minocycline minimum inhibitory concentrations (MICs) than do those carrying tetB.15,16 The prevalence of tetB in A. baumannii has increased over recent years, and the minocycline MICs for tetB-carried A. baumannii have also increased.15–17 This phenomenon may be achieved by acquiring pre-existing resistance determinants via the mobile genetic elements, followed by amplification in response to selection, as is commonly observed in multi-drug resistant A. baumannii.18 For example, insertion sequence is found to play a role in spreading tetB.19 Furthermore, tetB is found in AbaR-type genomic antibiotic resistance islands or transposons, by which that import tetB and other antibiotic resistance genes into the globally disseminated A. baumannii clones.20–22 Another important factor may involve the clonal spread of tetB-carried A. baumannii.19,23 The AdeABC efflux pump, belonging to the resistance-nodulation-cell division efflux system, is associated with the development of tigecycline resistance. Minocycline is reportedly a weaker substrate of AdeABC pumps than tigecycline.24 Thus, tigecycline-resistant A. baumannii isolates may remain susceptible to minocycline.14,24 TetB testing is considered a valuable tool for gene-based susceptibility evaluation.16 A global study shows that the presence of tetB is highly associated with MICs above the CLSI susceptibility breakpoint of 4 mg/L, and the lack of tetB gene could be a marker of CLSI-defined minocycline susceptibility in A. baumannii.16
In Taiwan, the minocycline susceptibility rate is 85% among all species of A. baumannii complex isolates, including A. baumannii, A. nosocomialis and A. pittii.25 The minocycline susceptibility rate of CRAB, the bacterium of critical-priority, is unknown. Furthermore, the extent of tetB carriage is also unknown. In this study, we evaluated the minocycline susceptibility rate and the prevalence rate of tetB carriage in CRAB. We also analyzed the possible association involving tetB carriage and the minocycline susceptibility.
Materials and Methods
Study Samples and Species Identification
This study was conducted at the National Taiwan University Hospital (NTUH), Taipei, a 2600-bed medical center located in Taipei City, Taiwan. Clinical A. baumannii isolates from patients with bloodstream infections (BSIs) were collected between 2014 and 2017 as previously described.26 Only the first episode was included if the patient had experienced multiple episodes of A. baumannii BSIs, and only one isolate was included for each episode. Between 2014 and 2016, the A. baumannii complex isolates primarily identified by conventional biochemical methods were collected for species identification according to the 16S–23S ribosomal RNA gene intergenic spacer region, as previously described.27 Isolates of A. nosocomialis and A. pittii were excluded.28 In 2017, the A. baumannii isolates were identified directly by the hospital microbiological laboratories using matrix-assisted laser desorption ionization-time of flight mass spectrometry Bruker Biotyper system (MicroFlex LT; Bruker Daltonik GmbH, Bremen, Germany), which was showed to be a rapid, simple, and reliable method to discriminate A. baumannii from other A. pittii and A. nosocomialis.29 Among the A. baumannii isolates collected, those preliminarily identified as carbapenem-resistant by the hospital clinical laboratory were collected for further microbiological studies. Imipenem or meropenem resistance was confirmed by antimicrobial susceptibility testing as described below. This study was approved by the Research Ethics Committee of NTUH (NTUH 201008047R). The isolates were collected as part of routine clinical practice, and the requirement for informed consent was waived by the ethics committee.
Pulsed-Field Gel Electrophoresis
Randomly selected CRAB isolates were subjected to pulsed-field gel electrophoresis (PFGE) to determine the clonal relatedness.30 Strains with relatedness ≥80% were defined as PFGE-related strains. A total of 100 PFGE-unrelated CRAB strains were collected for further minocycline antimicrobial susceptibility testing and tetB gene detection.
Antimicrobial Susceptibility Testing
The MICs of amikacin, ampicillin/sulbactam, ceftazidime, cefepime, ciprofloxacin, levofloxacin, imipenem and meropenem for A. baumannii strains were determined using Sensititre GNX3F (Trek Diagnostics, West Sussex, UK). The MICs of minocycline, tigecycline, and colistin were determined using broth microdilution. The MIC results were interpreted according to the CLSI recommendations and/or the EUCAST criteria for all antimicrobial agents.31,32 The CLSI minocycline breakpoints for A. baumannii were as follows: MIC ≤4 mg/L for susceptibility, 8 mg/L for intermediate resistance, and ≥16 mg/L for resistance. EUCAST did not issue minocycline breakpoints for A. baumannii. Strains with minocycline MICs of >1 mg/L were classified as PK-PD non-susceptible.11 For tigecycline, the non-species-related PK-PD breakpoint defined by EUCAST as ≤0.5 mg/L for susceptibility was used.
Detection of tetB Gene
tetB carriage was detected using polymerase chain reaction (PCR). The primers used for amplification of the tetB gene were as previously described.16 tetB_F (5′-TAC GTG AAT TTA TTG CTT CGG-3′) and tetB_R (5′-ATA CAG CAT CCA AAG CGC AC-3′). PCR was performed using TaKaRa TaqTM (TaKaRa Beijing, China) under the following conditions: 95°C for 5 min; followed by 35 cycles of 95°C for 20s, 55°C for 20s and 72°C for 1 min, with a final extension step of 72°C for 5 min. The PCR products were analyzed by a 1% agarose gel electrophoresis.
Statistical Analysis
A two-sample Student’s t-test of proportions was used to compare the susceptibility rates to the different antimicrobial agents. The rank-sum test was used to compare the MICs between the two groups. Fisher’s exact test was used to compare categorical variables between the two groups. Two-sided P values <0.05 were considered to be statistically significant. The sensitivity, specificity, positive predictive value (PPV) and negative predictive value (NPV) of the tetB testing in predicting minocycline susceptibility were determined. Data were analyzed using Stata software (version 14; StataCorp, College Station, TX, USA).
Results
Antimicrobial Susceptibility Testing
A total of 100 genetically unrelated CRAB clinical strains were subjected to antimicrobial susceptibility testing for minocycline, colistin, tigecycline, and other antimicrobial agents (Table 1). The colistin MIC50/90 was 0.25/1 mg/L. The tigecycline MIC50/90 was 2/4 mg/L, and none of the CRAB strains was susceptible to tigecycline with MIC of ≤0.5 mg/L. The minocycline MIC50/90 was 4/16 mg/L. The minocycline susceptibility rate of the 100 CRAB strains was 55%, which as interpreted according to the CLSI criteria. Among the 55 CLSI-defined minocycline-susceptible CRAB strains, 54 strains (98.2%) were PK-PD non-susceptible (MICs >1 mg/L). The susceptibility rate to sulbactam, ceftazidime, cefepime and piperacillin–tazobactam for all 100 strains from 0% to 4%, and susceptibility rate to amikacin was 13%. Ninety-nine strains (99%) were PK-PD non-susceptible (MICs >1 mg/L). None of the strains exhibited a minocycline MIC of ≤0.5 mg/L, and only one strain (1%) had a minocycline MIC of 1 mg/L.
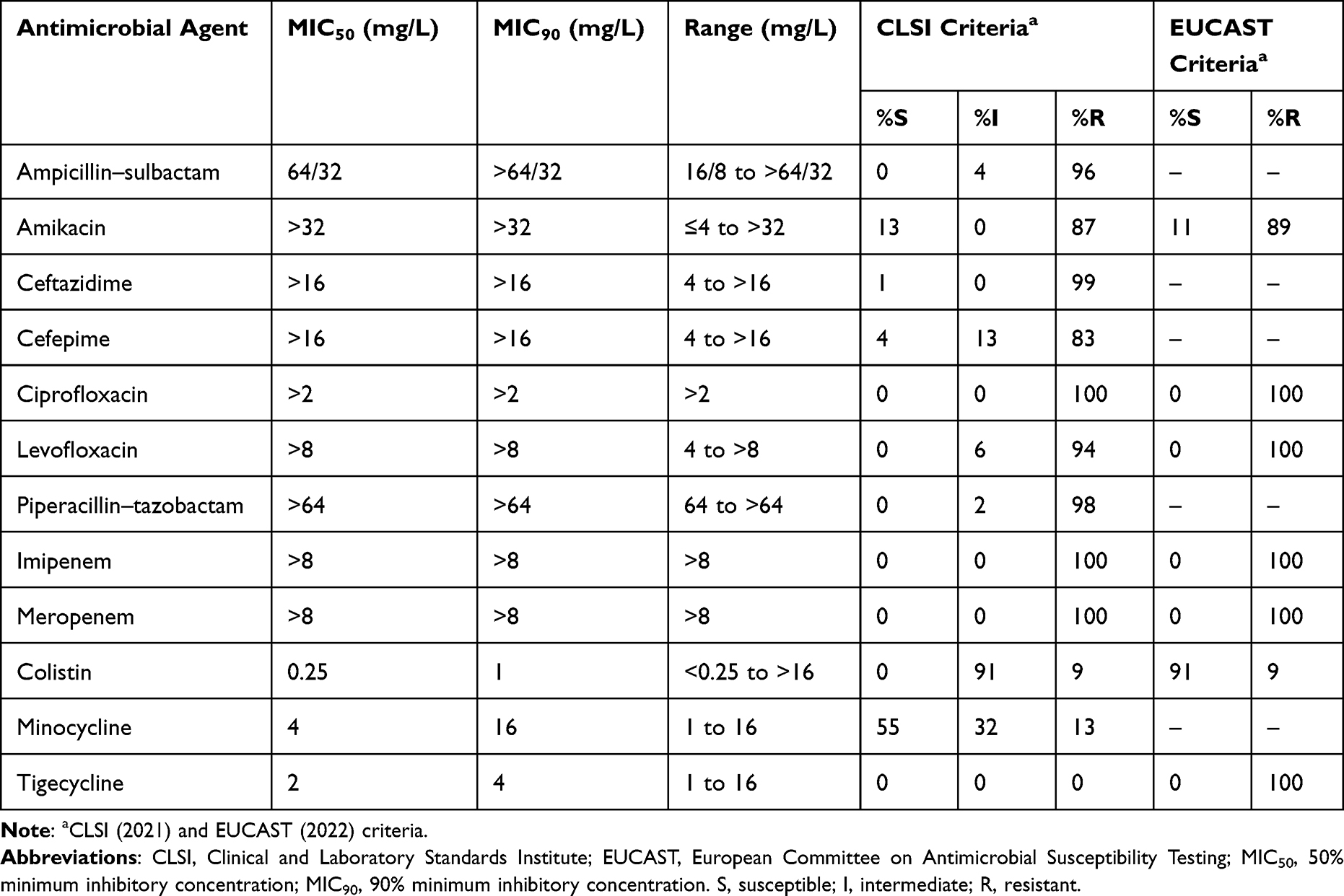 |
Table 1 In vitro Antimicrobial Activity of Antimicrobial Agents Against 100 Genetically Unrelated Carbapenem-Resistant Acinetobacter baumannii Strains |
TetB Carriage and Correlation with Susceptibility Testing
There was a high positive rate of tetB carriage in CRAB. Of the 100 tested strains, 97 (97%) were positive for tetB using PCR. The median minocycline MICs were 4 mg/L (range, 2–16 mg/L) for tetB-positive strains and 2 mg/L (range, 1–4 mg/L) for tetB-negative strains. By statistical analysis, tetB carriage was significantly correlated with a higher minocycline MIC (P = 0.02). The distribution of minocycline MICs of tetB-positive and -negative strains presented a modest degree of separation (Figure 1). All the three tetB-negative strains and 53.6% (52/97) of the 97 tetB-positive strains were susceptible to minocycline according to the CLSI criteria. tetB carriage was not significantly correlated with CLSI-defined minocycline non-susceptibility (P = 0.25). All of the tetB-positive strains and 66.7% (2/3) of the tetB-negative strains were PK-PD non-susceptible. tetB carriage was significantly correlated with minocycline PK-PD non-susceptibility (MIC >1 mg/L) (P = 0.03).
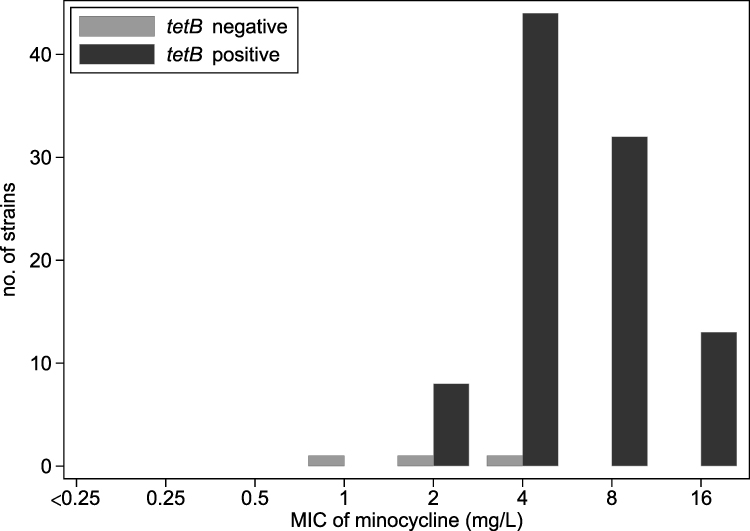 |
Figure 1 Distribution of minocycline minimum inhibitory concentrations (MICs) of the 100 carbapenem-resistant Acinetobacter baumannii strains. |
All of the 100 CRAB strains were resistant to tigecycline, with MICs of >0.5 mg/L. For tetB-positive strains, the median tigecycline MIC was 2 mg/L (range, 1–16 mg/L), and the MIC50/90 was 2/4 mg/L. For tetB-negative strains, the median tigecycline MIC was 2 mg/L (range, 2–16 mg/L). tetB carriage was not correlated with a higher tigecycline MIC (P = 0.23). The sensitivity and specificity of the tetB PCR for predicting CLSI-defined minocycline non-susceptibility (MIC >4 mg/L) were 100% and 5.5%, respectively, and the PPV and NPV were 46.4% and 100%, respectively. The sensitivity and specificity of the tetB PCR to predict minocycline PK-PD non-susceptibility (MIC >1 mg/L) were 98% and 100%, respectively, and the PPV and NPV were 100% and 33.3%, respectively. The area under the receiver operating characteristic (ROC) curve of using tetB PCR to predict CLSI-defined minocycline non-susceptibility (MIC >4 mg/L) and minocycline PK-PD non-susceptibility (MIC >1 mg/L) were 0.5273 and 0.9899, respectively.
Discussion
Since safe and effective therapeutic options for CRAB infections are severely limited, characterization of the clinical isolates by phenotypic and molecular methods is important to provide information on the epidemiological characteristics of such pathogens.33 In the present study, we tested the susceptibility of 100 genetically unrelated CRAB clinical strains to minocycline. Among the tested antimicrobial agents, minocycline was the most active agent against CRAB, according to the CLSI criteria. Fifty-five CRAB strains (55%) were susceptible to minocycline. In addition, among the 100 CRAB strains analyzed here, 99% were PK-PD non-susceptible, with minocycline MICs of >1 mg/L. Most of the tested strains (97%) carried tetB gene.
When the CLSI susceptibility breakpoint was applied, the susceptibility rate to minocycline was 55%. However, there remains ongoing debates regarding minocycline susceptibility breakpoints. Tsakris et al estimated the minocycline susceptibility breakpoint for A. baumannii using a Monte Carlo simulation based on PD data. The breakpoints were expected to be 0.5 mg/L and 1 mg/L when minocycline were administered at doses of 200 mg/day and 400 mg/day, respectively.11 In the present study, the percentage (1%) of CRAB strains with relatively low minocycline MIC (≤1 mg/L) was much lower than those reported in other regions, ie, there were 15.9% US and 18.4% UK CRAB isolates with minocycline MICs of ≤0.5 mg/L, and 32.7% CRAB isolates with minocycline MICs of ≤1 mg/L in both regions.15,34
The clinical efficacy and the optimal dose of minocycline for treating A. baumannii infections remain unclear. In infections caused by A. baumannii with minocycline MICs of 3 or 4 mg/L, clinical failure occurred in a substantial proportion of critically ill patients receiving minocycline at the dose of 200 mg/day.35 Minocycline at the dose of 400 mg/day combined with a second or third active drug was suggested based on a few favorable clinical experiences.12 An in vitro PD study,6 which revealed the emergence of resistance in A. baumannii strains with minocycline MICs of 3 or 4 mg/L under exposure to 400 mg/day minocycline,6 also favored combination therapy rather than minocycline monotherapy. A recent in vitro PD study suggested that a minocycline dose of 700 mg/day was the pharmacodynamically optimized dose against CRAB strains with minocycline MICs of 2 mg/L, when minocycline was used in combination with polymyxin B and sulbactam.36 The high doses of minocycline, such as 700 mg or 800 mg/day, had been reported in the literature for non-infectious diseases. A Phase I study revealed that minocycline administered at a dose of 300 mg twice daily was the maximum-tolerated dose in healthy adult subjects.37 Among the CRAB strains in the present study, there were only 1% with a relatively low minocycline MIC of ≤1 mg/L and 9% with minocycline MICs of 2 mg/L. Considering the high percentage of CRAB strains with minocycline MIC of >1 mg/L and the unresolved safety issues involving high-dose minocycline, the role of minocycline in treating CRAB-related infections in clinical settings needs be defined, and additional clinical studies are warranted.
In addition to the increasing prevalence of tetB-carried A. baumannii over recent years, the minocycline MICs for tetB-carried A. baumannii also increased.15–17 A substantial proportion of A. baumannii carried tetB in the USA and worldwide. In one report, 77.6% of the 107 US CRAB strains collected between 2009 and 2015 carried tetB. The minocycline MICs were relatively low (≤1 mg/L) for approximately 15% of the tetB-positive strains.15 Among clinical A. baumannii isolates collected worldwide between 1998 and 2015, 64.0% (165/258) carried tetB. There were almost all (99.4%, 164/165) of the tetB-positive isolates with minocycline MICs of ≥4 mg/L, and only 0.6% of the tetB-positive isolates with relatively a low minocycline MIC (≤1 mg/L).16 In the present study, there were 91.8% (89/97) of the 97 tetB-positive strains with minocycline MICs of ≥4 mg/L, and only 8.2% (8/97) with minocycline MICs of 2 mg/L. The minocycline MICs were >1 mg/L for all of the 97 tetB-positive CRAB strains. The distribution of minocycline MICs in our tetB-carried CRAB strains was similar to that observed in global tetB-carried A. baumannii isolates.16 In that study, the presence of tetB is highly associated with MICs above the CLSI-defined susceptibility breakpoint, as well as MICs above the PK-PD susceptibility breakpoint of 1 mg/L.16 However, the role of tetB testing in discriminating minocycline PK-PD susceptibility in CRAB should be interpreted with caution, as the number of tetB-negative CRAB strains in our study was insufficient to determine such an association. Moreover, the application of tetB testing for predicting minocycline MICs might vary in different populations of A. baumannii, especially the minocycline MICs are relatively low for a substantial proportion of tetB-carried A. baumannii.15,38 One study exploring on the tetB-carried A. baumannii isolates with minocycline MICs of ≤1 mg/L found that there were minocycline-resistant subpopulations, which increased after incubation with minocycline. However, such subpopulations were not detected among isolates not carrying tetB.38 That study implies the induction of the TetB efflux pump in tetB-carried isolates under exposure to minocycline. Detection of tetB status might still have a role in discriminating minocycline non-susceptibility or the likelihood of developing resistance during exposure to minocycline when minocycline is chosen as a therapeutic regimen for CRAB infections. In addition, the high prevalence of tetB-carried CRAB with minocycline MICs of >1 mg/L in our hospital also warrants further investigations.
This study had several strengths. First, we focused on CRAB, which ranked highest for research of any effective drug, for example, minocycline. Second, all CRAB strains were identified as genetically unrelated by PFGE. Therefore, our results regarding minocycline susceptibility for CRAB may be robustly representative. Third, we clarified whether the CRAB strains carried tetB to detect a potential relatively higher minocycline MIC. However, this study also had several limitations. First, we did not examine the tetB gene expression levels in our CRAB strains. Although tetB carriage is statistically correlated with minocycline MIC above the PK-PD susceptibility breakpoint of 1 mg/L and a higher minocycline MIC, antimicrobial resistance might result from differences in tetB gene expression.39 The association between tetB gene expression levels and minocycline MIC in our CRAB strains warrants further investigation. Whether our CRAB strains express tetB constitutively,39 and whether the mechanisms regulating tetB expression, such as Tet repressor protein encoded by tetR, differ in A. baumannii from other genus might also warrant exploration.40 Furthermore, we did not assess whether other efflux pumps might have contributed to the relatively higher minocycline MICs detected in our CRAB strains. However, the TetB efflux pump has been reported to be the primary mechanism of minocycline resistance.41 Finally, this study reported the minocycline susceptibility rate of CRAB clinical strains in a single medical center. Nevertheless, we had highlighted the importance of obtaining the information regarding local minocycline MICs and tetB gene prevalence.
Conclusion
The findings of this study suggest that although minocycline was the most active agent against CRAB clinical strains according to the CLSI criteria at our institute, most of our CRAB strains might be classified as PK-PD non-susceptible with minocycline MICs of >1 mg/L. Most of the CRAB strains carried tetB gene. Recognizing the minocycline MIC and the tetB status may be essential when using minocycline to treat CRAB-related infections.
Acknowledgments
The authors thank the NTUH Third Core Facility for technical assistance and facility support.
Author Contributions
All authors made a significant contribution to the work reported, including in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all of these areas; took part in drafting, or critically reviewing the article; gave final approval of the version to be submitted; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was partly supported by the Ministry of Science and Technology, Taiwan [MOST-109-2628-B-00-041, MOST-109-2314-B-002-235-MY3], the National Taiwan University Hospital [NTUH. 110-S4901], and the Far Eastern Memorial Hospital. National Taiwan University Hospital Joint Research Program [108-FTN14, 109-FTN09, 110-FTN10].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
2. Strateva T, Sirakov I, Stoeva T, et al. Carbapenem-resistant Acinetobacter baumannii: current status of the problem in four Bulgarian university hospitals (2014–2016). J Glob Antimicrob Resist. 2019;16:266–273. doi:10.1016/j.jgar.2018.10.027
3. Kuo SC, Chang SC, Wang HY, et al. Emergence of extensively drug-resistant Acinetobacter baumannii complex over 10 years: nationwide data from the Taiwan Surveillance of Antimicrobial Resistance (TSAR) program. BMC Infect Dis. 2012;12(1):200. doi:10.1186/1471-2334-12-200
4. Flamm RK, Shortridge D, Castanheira M, Sader HS, Pfaller MA. In vitro activity of minocycline against U.S. Isolates of Acinetobacter baumannii-Acinetobacter calcoaceticus species complex, Stenotrophomonas maltophilia, and Burkholderia cepacia complex: results from the SENTRY Antimicrobial Surveillance Program, 2014 to 2018. Antimicrob Agents Chemother. 2019;63(11):e01154–e011519. doi:10.1128/AAC.01154-19
5. Doi Y. Treatment options for carbapenem-resistant gram-negative bacterial infections. Clin Infect Dis. 2019;69(Suppl 7):S565–S575. doi:10.1093/cid/ciz830
6. Alfouzan WA, Noel AR, Bowker KE, Attwood MLG, Tomaselli SG, MacGowan AP. Pharmacodynamics of minocycline against Acinetobacter baumannii studied in a pharmacokinetic model of infection. Int J Antimicrob Agents. 2017;50(6):715–717. doi:10.1016/j.ijantimicag.2017.06.026
7. Ritchie DJ, Garavaglia-Wilson A. A review of intravenous minocycline for treatment of multidrug-resistant Acinetobacter infections. Clin Infect Dis. 2014;59(Suppl 6):S374–S380. doi:10.1093/cid/ciu613
8. Fragkou PC, Poulakou G, Blizou A, et al. The role of minocycline in the treatment of nosocomial infections caused by multidrug, extensively drug and pandrug resistant Acinetobacter baumannii: a systematic review of clinical evidence. Microorganisms. 2019;7(6):159. doi:10.3390/microorganisms7060159
9. Gales AC, Seifert H, Gur D, Castanheira M, Jones RN, Sader HS. Antimicrobial susceptibility of Acinetobacter calcoaceticus-Acinetobacter baumannii complex and Stenotrophomonas maltophilia clinical isolates: results from the SENTRY Antimicrobial Surveillance Program (1997–2016). Open Forum Infect Dis. 2019;6(Suppl1):S34–S46. doi:10.1093/ofid/ofy293
10. Wong D, Nielsen TB, Bonomo RA, Pantapalangkoor P, Luna B, Spellberg B. Clinical and pathophysiological overview of Acinetobacter infections: a century of challenges. Clin Microbiol Rev. 2017;30(1):409–447. doi:10.1128/CMR.00058-16
11. Tsakris A, Koumaki V, Dokoumetzidis A. Minocycline susceptibility breakpoints for Acinetobacter baumannii: do we need to re-evaluate them? J Antimicrob Chemother. 2019;74(2):295–297. doi:10.1093/jac/dky448
12. Pogue JM, Neelakanta A, Mynatt RP, Sharma S, Lephart P, Kaye KS. Carbapenem-resistance in gram-negative bacilli and intravenous minocycline: an antimicrobial stewardship approach at the Detroit Medical Center. Clin Infect Dis. 2014;59(Suppl 6):S388–S393. doi:10.1093/cid/ciu594
13. Sklenar I, Spring P, Dettli L. One-dose and multiple-dose kinetics of minocycline in patients with renal disease. Agents Actions. 1977;7(3):369–377. doi:10.1007/BF01969570
14. Coyne S, Courvalin P, Perichon B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob Agents Chemother. 2011;55(3):947–953. doi:10.1128/AAC.01388-10
15. Wang P, McElheny CL, Mettus RT, Shanks RMQ, Doi Y. Contribution of the TetB efflux pump to minocycline susceptibility among carbapenem-resistant Acinetobacter baumannii strains. Antimicrob Agents Chemother. 2017;61(10):e01176–e011117. doi:10.1128/AAC.01176-17
16. Lomovskaya O, Sun D, Rubio-Aparicio D, et al. Absence of TetB identifies minocycline-susceptible isolates of Acinetobacter baumannii. Int J Antimicrob Agents. 2018;52(3):404–406. doi:10.1016/j.ijantimicag.2018.04.006
17. Huys G, Cnockaert M, Vaneechoutte M, et al. Distribution of tetracycline resistance genes in genotypically related and unrelated multiresistant Acinetobacter baumannii strains from different European hospitals. Res Microbiol. 2005;156(3):348–355. doi:10.1016/j.resmic.2004.10.008
18. Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol Rev. 2018;31(4):e00088–e000817. doi:10.1128/CMR.00088-17
19. Vilacoba E, Almuzara M, Gulone L, et al. Emergence and spread of plasmid-borne tet(B): ISCR2in minocycline-resistant Acinetobacter baumannii isolates. Antimicrob Agents Chemother. 2013;57(1):651–654. doi:10.1128/AAC.01751-12
20. Seputiene V, Povilonis J, Suziedeliene E. Novel variants of AbaR resistance islands with a common backbone in Acinetobacter baumannii isolates of European clone II. Antimicrob Agents Chemother. 2012;56(4):1969–1973. doi:10.1128/AAC.05678-11
21. Nigro SJ, Hall RM. Antibiotic resistance islands in A320 (RUH134), the reference strain for Acinetobacter baumannii global clone 2. J Antimicrob Chemother. 2012;67(2):335–338. doi:10.1093/jac/dkr447
22. Nigro SJ, Hall RM. Tn6167, an antibiotic resistance island in an Australian carbapenem-resistant Acinetobacter baumannii GC2, ST92 isolate. J Antimicrob Chemother. 2012;67(6):1342–1346. doi:10.1093/jac/dks037
23. Zhang X, Li F, Awan F, Jiang H, Zeng Z, Molecular LW. Epidemiology and clone transmission of carbapenem-resistant Acinetobacter baumannii in ICU rooms. Front Cell Infect Microbiol. 2021;11:633817. doi:10.3389/fcimb.2021.633817
24. Lomovskaya O, Sun D, King P, Dudley MN. Tigecycline (TIG) but not minocycline (MINO) selects for clinically relevant efflux-mediated resistance (R) in Acinetobacter spp.
25. Yang YS, Lee Y, Tseng KC, et al. In vivo and In vitro efficacy of minocycline-based combination therapy for minocycline-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2016;60(7):4047–4054. doi:10.1128/AAC.02994-15
26. Chuang YC, Cheng A, Sun HY, et al. Microbiological and clinical characteristics of Acinetobacter baumannii bacteremia: implications of sequence type for prognosis. J Infect. 2019;78(2):106–112. doi:10.1016/j.jinf.2018.10.001
27. Chang HC, Wei YF, Dijkshoorn L, Vaneechoutte M, Tang CT, Chang TC. Species-level identification of isolates of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex by sequence analysis of the 16S–23S rRNA gene spacer region. J Clin Microbiol. 2005;43(4):1632–1639. doi:10.1128/JCM.43.4.1632-1639.2005
28. Chuang YC, Sheng WH, Li SY, et al. Influence of genospecies of Acinetobacter baumannii complex on clinical outcomes of patients with Acinetobacter bacteremia. Clin Infect Dis. 2011;52(3):352–360. doi:10.1093/cid/ciq154
29. Hsueh P-R, Kuo L-C, Chang T-C, et al. Evaluation of the bruker biotyper matrix-assisted laser desorption ionization–time of flight mass spectrometry system for identification of blood isolates of Acinetobacter species. J Clin Microbiol. 2014;52(8):3095–3100. doi:10.1128/JCM.01233-14
30. Seifert H, Dolzani L, Bressan R, et al. Standardization and interlaboratory reproducibility assessment of pulsed-field gel electrophoresis-generated fingerprints of Acinetobacter baumannii. J Clin Microbiol. 2005;43(9):4328–4335. doi:10.1128/JCM.43.9.4328-4335.2005
31. EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters. Version 12.0; 2022. Available from: https://www.eucast.org.
32. Performance CLSI. Standards for Antimicrobial Susceptibility Testing. CLSI supplement M100.
33. Matthew GD, Stefania Z, Ádám LN, Ibrahim B, Márió G. Insights on carbapenem-resistant Acinetobacter baumannii: phenotypic characterization of relevant isolates. Acta Biologica Szegediensis. 2021;65(1):85–92. doi:10.14232/abs.2021.1.85-92
34. Livermore DM, Mushtaq S, Warner M, Woodford N. In vitro activity of eravacycline against carbapenem-resistant Enterobacteriaceae and Acinetobacter baumannii. Antimicrob Agents Chemother. 2016;60(6):3840–3844. doi:10.1128/AAC.00436-16
35. Goff DA, Bauer KA, Mangino JE. Bad bugs need old drugs: a stewardship program’s evaluation of minocycline for multidrug-resistant Acinetobacter baumannii infections. Clin Infect Dis. 2014;59(Suppl 6):S381–S387. doi:10.1093/cid/ciu593
36. Beganovic M, Daffinee KE, Luther MK, LaPlante KL. Minocycline alone and in combination with polymyxin B, meropenem, and sulbactam against carbapenem-susceptible and -resistant Acinetobacter baumannii in an in vitro pharmacodynamic model. Antimicrob Agents Chemother. 2021;65(3):e01680–e016820. doi:10.1128/AAC.01680-20
37. Cornely OA, Arenz D, Barraud O, et al. Phase I study to evaluate the safety and pharmacokinetics of single and multiple ascending doses of intravenous minocycline in healthy adult subjects. Open Forum Infect Dis. 2018;5(Supp 1):S425–S426. doi:10.1093/ofid/ofy210.1218
38. Rodriguez CH, Nastro M, Vay C, Famiglietti A. In vitro activity of minocycline against multidrug-resistant Acinetobacter baumannii isolates: evaluation of clinical breakpoints and review of literature. Microb Drug Resist. 2021;27(11):1560–1563. doi:10.1089/mdr.2020.0398
39. Coyne S, Guigon G, Courvalin P, Perichon B. Screening and quantification of the expression of antibiotic resistance genes in Acinetobacter baumannii with a microarray. Antimicrob Agents Chemother. 2010;54(1):333–340. doi:10.1128/AAC.01037-09
40. Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev. 2001;65(2):232–260. doi:10.1128/MMBR.65.2.232-260.2001
41. Asadi A, Abdi M, Kouhsari E, et al. Minocycline, focus on mechanisms of resistance, antibacterial activity, and clinical effectiveness: back to the future. J Glob Antimicrob Resist. 2020;22:161–174. doi:10.1016/j.jgar.2020.01.022
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.