")
Back to Journals » OncoTargets and Therapy » Volume 12
MicroRNA-618 Directly Targets Metadherin mRNA To Suppress The Malignant Phenotype Of Osteosarcoma Cells By Reducing PTEN-AKT Pathway Output
Authors Li B, Zhao J, Zhao Q, Wu D, Zhang C, Zhao K, Song Y, Gao C
Received 13 June 2019
Accepted for publication 7 October 2019
Published 15 November 2019 Volume 2019:12 Pages 9795—9807
DOI https://doi.org/10.2147/OTT.S219440
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
This paper has been retracted.
Bohan Li, 1 Jie Zhao, 2 Qian Zhao, 3 Dongjin Wu, 2 Cheng Zhang, 2 Kun Zhao, 2 Yang Song, 2 Chunzheng Gao 2
1Health Management Center, The Second Hospital of Shandong University, Shandong 250033, People’s Republic of China; 2Department of Spine Surgery, The Second Hospital of Shandong University, Shandong 250033, People’s Republic of China; 3Jinan Central Hospital, The Affiliated Hospital of Shandong University, Shandong 250013, People’s Republic of China
Correspondence: Dongjin Wu
Department of Spine Surgery, The Second Hospital of Shandong University, Shandong 250033, People’s Republic of China
Email [email protected]
Purpose: Dysregulation of microRNA-618 (miR-618) has been observed in multiple types of human cancer. However, whether miR-618 is implicated in osteosarcoma (OS) initiation and progression is still unclear. Hence, we measured the expression of miR-618 in OS tissues and cell lines. In addition, the roles of miR-618 and the mechanisms underlying its activities in OS cells were examined.
Methods: The expression status of miR-618 in OS was analyzed by reverse-transcription quantitative PCR. The regulatory roles of miR-618 overexpression in OS were explored by the Cell Counting Kit-8 assay, flow-cytometric analysis, Transwell cell migration and invasion assays, and a tumor xenograft experiment.
Results: The results revealed that the expression of miR-618 was notably lower in OS tissues and cell lines, and that the low miR-618 expression significantly correlated with the clinical stage and distant metastasis among patients with OS. Exogenous miR-618 expression significantly suppressed OS cell proliferation, migration, and invasion and induced apoptosis in vitro as well as slowed tumor growth in vivo. Mechanism investigation indicated that metadherin (MTDH) is a direct target gene of miR-618 in OS cells. A knockdown of MTDH mimicked the tumor-suppressive effects of miR-618 upregulation on OS cells. Notably, resumption of MTDH expression attenuated the miR-618–mediated reduction in OS cell growth and metastasis in vitro. In addition, miR-618 overexpression reduced the PTEN–AKT pathway output in OS cells both in vitro and in vivo through downregulation of MTDH.
Conclusion: To the best of our knowledge, this is the first study to show that miR-618 exerts crucial tumor-suppressive actions in OS pathogenesis by directly targeting MTDH mRNA and reducing PTEN–AKT pathway output. These results will help to elucidate the functions of miR-618 in OS and suggest that this miRNA may be investigated as a therapeutic target in this disease.
Keywords: microRNA-618, osteosarcoma, metadherin, proliferation, invasion
Introduction
Osteosarcoma (OS), deriving from primitive bone-forming mesenchymal cells, is the most prevalent malignant bone tumor.1 OS most commonly occurs in children and adolescents and accounts for ~5% of childhood cancer cases and 8.9% of cancer-associated deaths among children.2 Owing to remarkable advances in the therapeutic techniques, such as wide tumor excision, chemotherapy, radiotherapy, and immunotherapy, the clinical efficacy of OS management has improved greatly.3 However, the prognosis of patients with OS remains unsatisfactory, with a 5-year overall-survival rate of only 30%.4 Most patients with OS eventually present with metastasis and/or recurrence.5 The details of the OS pathogenesis remain largely unknown; this situation may hamper the search for promising therapeutic targets. Therefore, it is important to fully elucidate the mechanisms of OS carcinogenesis and progression because they may point to novel and effective therapeutic modalities that will improve the prognosis of patients with OS.
MicroRNAs (miRNAs) are a large group of endogenous single-stranded short noncoding RNA molecules with the length of ~18–24 nucleotides.6 MiRNAs modulate gene expression at both the transcriptional and post-transcriptional levels via binding to the 3′‐untranslated region (3′‐UTR) of target mRNAs, thereby promoting degradation of messenger RNAs (mRNAs) or suppressing their translation into protein.7 The alteration in miRNA expression has been frequently reported in almost all human cancer types.8–10 Numerous miRNAs have been confirmed to be aberrantly expressed in OS and to function as oncogenic RNAs or tumor suppressors.11 An increasing number of studies have documented the involvement of miRNAs in the control over multiple steps of OS onset and progression, including cell proliferation, apoptosis, cell cycle, and metastasis.12–14 Hence, miRNAs have become promising biomarkers for the diagnosis and targets for the treatment of OS.
Dysregulation of miR-618 has been observed in prostate cancer15 and thyroid carcinoma.16,17 Nonetheless, whether miR-618 is implicated in the OS initiation and progression is still unclear. In this study, we first measured the expression of miR-618 in OS tissues and cell lines. Then, the correlation between miR-618 expression and clinical parameters of patients with OS was evaluated. A series of functional experiments was conducted to assess and validate the influence of miR-618 on the initiation and progression of OS. Moreover, the mechanisms underlying the activities of miR-618 in OS cells were examined.
Materials And Methods
Clinical Tissue Specimens
A total of 41 patients with OS who underwent surgical resection in The Second Hospital of Shandong University were recruited. None of these patients had received anticancer therapies, such as chemotherapy, radiotherapy, or immunotherapy. All the tissue specimens were frozen in liquid nitrogen immediately after surgical resection and then stored at −80°C. The Ethics Committee of the Second Hospital of Shandong University approved the study protocol, and all the patients provided written informed consent.
Cell Culture And Transient Transfection
Four human OS cell lines (SAOS-2, MG-63, U2OS, and HOS) and the normal human osteoblast hFOB1.19 cell line were purchased from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). All these cell lines were incubated at 5% CO2 and 37°C in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% of fetal bovine serum (FBS; both from Gibco, Invitrogen, Carlsbad, CA, USA) and 1% of a penicillin/streptomycin solution (Sigma‐Aldrich, St. Louis, MO).
The synthetic miR-618 mimics and miRNA mimic negative control (miR-NC) were purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, Guangdong, China). The MTDH-overexpressing plasmid was generated by inserting the MTDH cDNA lacking its 3′-UTR into the pCMV vector. This plasmid was chemically synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The small interfering RNA (siRNA) against MTDH (si-MTDH) was acquired from Qiagen GmbH (Hilden, Germany) and used to knock down endogenous MTDH expression. Negative control siRNA (si-NC) served as a control for si-MTDH. RNA oligonucleotides and the plasmid were transfected into cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
RNA Extraction And Reverse-Transcription Quantitative PCR (RT-qPCR)
The TRIzol Reagent (Invitrogen; Thermo Fisher Scientific) was employed for total-RNA isolation from the tissue specimens and cells. The concentration of total RNA was determined on a Nanodrop 2000 (Thermo Fisher Scientific). Total RNA was reversely transcribed into cDNA using the miScript Reverse Transcription Kit (Qiagen GmbH). Thereafter, qPCR was performed to measure miR-618 expression with the miScript SYBR Green PCR Kit (Qiagen GmbH). To determine MTDH mRNA expression, reverse transcription was carried out using the PrimeScript RT Reagent Kit (Takara Bio, Dalian, China). Next, qPCR was carried out by means of the SYBR Premix Ex Taq™ Kit (Takara Bio, Dalian, China) and an Applied Biosystems 7500 Real-time PCR System (Thermo Fisher Scientific). Small nuclear RNA U6 served as the internal reference for miR‐618, and GAPDH was the internal control for MTDH. Relative gene expression was calculated by the 2−ΔΔCq method.18
A Cell Counting Kit-8 (CCK-8) Assay
Transfected cells were seeded in 96-well plates at a density of 3 × 103 cells/well. Five replicate wells were set up for each group. After cultivation for 0, 24, 48, or 72 h, the CCK-8 assay was carried out by the addition of 10 μL of the CCK-8 reagent (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) into every well. The cells were incubated at 37°C and 5% CO2 for additional 2 h, and the absorbance value of each well was measured on a spectrophotometric plate reader (Infinite® 200 PRO; Tecan Group, Ltd., Mannedorf, Switzerland).
Flow-Cytometric Analysis
Cell apoptosis was assessed using the Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (BioLegend, San Diego, CA, USA). Briefly, transfected cells were treated with EDTA-free 0.25% Trypsin (Gibco, Invitrogen) and rinsed twice with ice-cold phosphate-buffered saline. The cells were then transferred into a flow tube and resuspended in 100 µL of binding buffer, followed by incubation at room temperature in the dark for 20 min with 5 μL of Annexin V-FITC and 5 μL of a propidium iodide solution. The percentages of apoptotic cells were determined on a flow cytometer (FACScan; BD Biosciences, San Jose, CA, USA).
Transwell Migration And Invasion Assays
A Transwell chamber (Costar™; Corning, Inc., Corning, NY, USA) coated with Matrigel (BD Biosciences) was applied to evaluate the cellular invasion capacity. In particular, the transfected cells were collected after 48 h of incubation and then resuspended in the FBS-free culture medium. A total of 200 μL of the suspension containing 5 × 104 cells was seeded in the upper chambers. The lower chambers were filled with 500 μL of DMEM containing 20% of FBS, which served as a chemoattractant. Following 24 h incubation at 37°C and 5% CO2, the cells that moved through the membrane were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet. Finally, the noninvading cells were removed, and the invading cells were photographed under an inverted microscope (Olympus IX83; Olympus Corporation, Tokyo, Japan). Five random visual fields of each chamber were selected for quantification. The migratory ability of cells was assessed with the experimental procedures similar to the invasion assay, except that the Transwell chambers were not precoated with Matrigel.
A Tumor Xenograft Experiment
All the animal experimental procedures were approved by the Ethics Committee of The Second Hospital of Shandong University, and were carried out in accordance with the Animal Protection Law of the People’s Republic of China-2009. Cells transfected with the miR-618 mimics or miR-NC were subcutaneously injected into the upper flank of 4- to 5-week-old nude mice (Shanghai Laboratory Animal Center; Shanghai, China). The width and length of tumor xenografts was measured every week with calipers. All the mice were euthanized 4 weeks after the inoculation. The tumor xenografts were excised and weighed, and their volume was calculated via the following formula: tumor volume (mm3) = (length × width2)/2.
Bioinformatics Analysis And A Luciferase Reporter Assay
TargetScan 7.1 (http://www.targetscan.org/) and miRanda (http://www.microrna.org) were employed to predict the potential targets of miR-618. MTDH was found to be a candidate target gene of miR-618.
The 3′-UTR fragment of the human MTDH gene containing the predicted wild-type (wt) or mutant (mut) miR-618–binding site was amplified by Shanghai GenePharma Co., Ltd. The 3′-UTR fragments were then inserted into the pMIR-REPORT vector (Promega, Madison, WI, USA) to construct the luciferase reporter plasmids: pMIR-MTDH-3ʹ-UTR-wt and pMIR-MTDH-3ʹ-UTR-mut. The luciferase reporter assay was conducted as follows: cells were seeded in 24-well plates, then cotransfected with either the miR-618 mimics or miR-NC and either pMIR-MTDH-3ʹ-UTR-wt or pMIR-MTDH-3ʹ-UTR-mut using Lipofectamine 2000. The transfected cells were harvested at 48 h post-transfection, and the luciferase activity was determined by means of a Dual-Luciferase Reporter Assay System (Promega). The firefly luciferase activity was normalized to that of Renilla luciferase.
Protein Extraction And Western Blot Analysis
Tissues or cells were lysed using the Active Protein Extraction Kit (KGP1050; Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) containing protease inhibitors (Millipore, Billerica, MA). The concentration of the total protein extracted from tissues or cells was measured with the Enhanced BCA Protein Assay Kit (Beyotime Institute of Biotechnology, Shanghai, China). Equal amounts of total protein were loaded for SDS-PAGE on 10% polyacrylamide gels and then transferred to polyvinylidene difluoride membranes (Millipore). After blocking with 5% skimmed milk for 2 h, the membranes were incubated overnight at 4°C with primary antibodies against MTDH (cat. No. sc-517220; Santa Cruz Biotechnology, Dallas, TX, USA), PTEN (cat. No. ab77161; Abcam, Cambridge, MA, USA), AKT (cat. No. sc-81434; Santa Cruz Biotechnology), phospho- (p-)AKT (cat. No. sc-514032; Santa Cruz Biotechnology), or GAPDH (cat. No. ab125247; Abcam). Next, the membranes were washed with Tris-buffered saline supplemented with 0.05% of Tween 20 (TBST) three times and incubated with a horseradish peroxidase–conjugated goat anti-mouse IgG antibody (cat. No. ab6789; Abcam) as a secondary antibody at room temperature for 2 h. Immunoreactivity was visualized with Enhanced Chemiluminescence Reagents (ECL; Pierce; Thermo Fisher Scientific).
Statistical Analysis
All the results were expressed as mean ± standard deviation. Student’s t test was performed to evaluate the differences between two groups. Comparisons among multiple groups were conducted by one-way analysis of variance followed by Bonferroni’s post hoc test. The association between miR-618 and clinical characteristics of the patients with OS was assessed by the χ2 test. Spearman correlation analysis was carried out to determine the correlation between miR-618 and MTDH mRNA levels among the OS tissue samples. All statistical analyses were performed using Statistical Package for the Social Sciences (SPSS) version 16.0 (SPSS, Inc., Chicago, IL, USA), and differences were defined as statistically significant if the P value was less than 0.05.
Results
Expression Of miR-618 Is Low In OS And Is Associated With Poor Clinical Outcomes
First of all, RT-qPCR was carried out to measure miR-618 expression in the 41 pairs of OS tissue samples and adjacent normal tissue (ANT) samples. MiR-618 was found to be significantly downregulated in OS tissue samples when compared with ANTs (Figure 1A, P < 0.05). In addition, the expression level of miR-618 was determined in four OS cell lines: SAOS-2, MG-63, U2OS, and HOS. Normal human osteoblast cell line hFOB1.19 served as a control. The results revealed that miR-618 expression was lower in all the four tested OS cell lines than in hFOB1.19 cells (Figure 1B, P < 0.05). The 41 patients with OS were separated into low and high miR-618 expression groups according to the median value as a cutoff. Underexpression of miR-618 correlated with the clinical stage (P = 0.005) and distant metastasis (P = 0.011) but not with age (P = 0.505), gender (P = 0.326), or tumor size (P = 0.277; Table 1). These results suggested that miR-618 expression may be closely associated with the initiation and progression of OS.
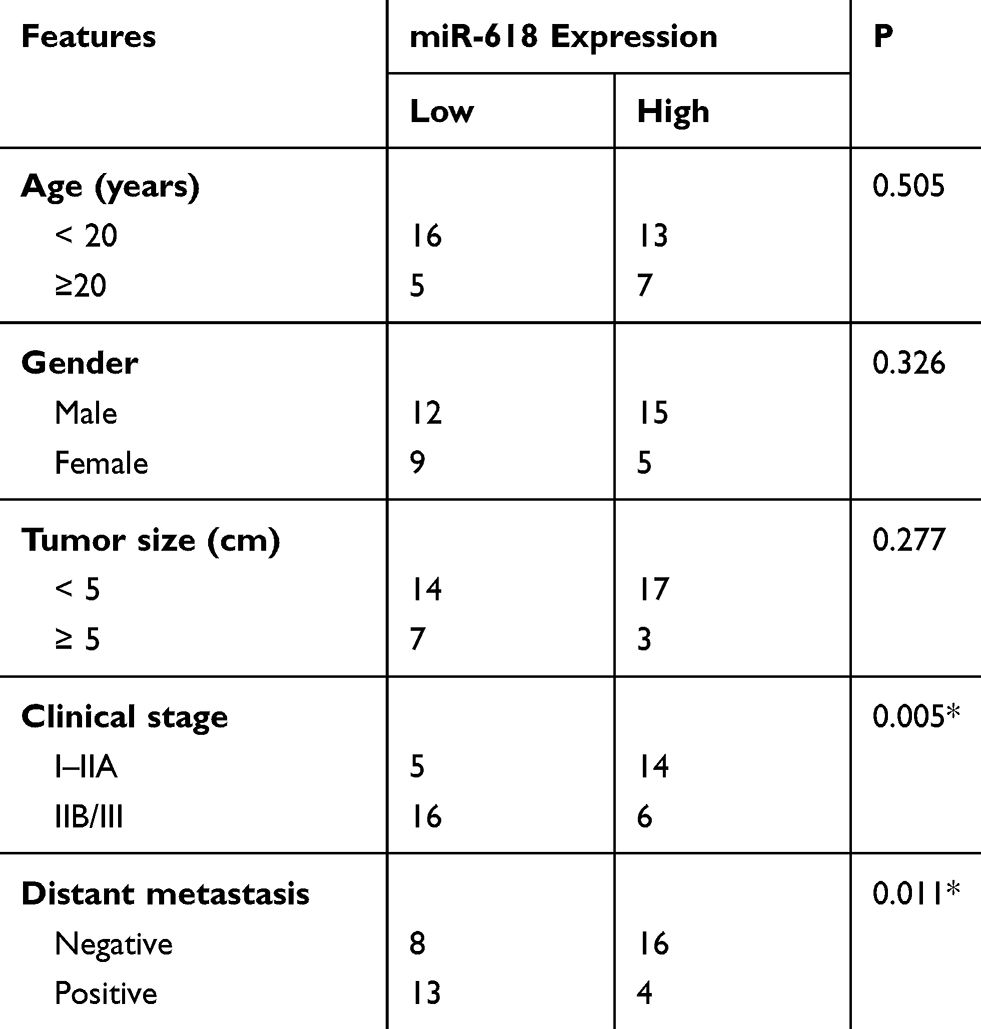 |
Table 1 The Association Between miR-618 Expression And Clinicopathological Features In Patients With OS |
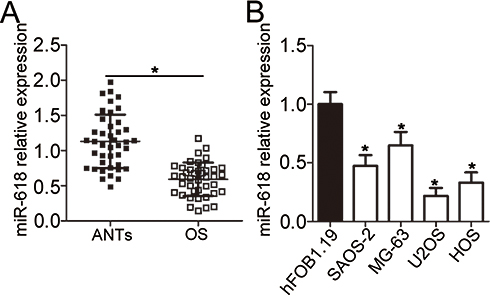 |
Figure 1 MiR-618 is underexpressed in OS tissues and cell lines. (A) The expression level of miR-618 was assessed in 41 pairs of OS tissue samples and ANTs by RT-qPCR. *P < 0.05 vs group “ANTs.” (B) RT-qPCR was carried out to measure miR-618 expression in the human hFOB1.19 normal osteoblast cell line and four OS cell lines (SAOS-2, MG-63, U2OS, and HOS). *P < 0.05 vs the hFOB1.19 group. |
MiR-618 Suppresses OS Cell Proliferation, Migration, And Invasion And Induces Apoptosis
To investigate the functions of miR-618 in OS progression, we transfected the miR-618 mimics into U2OS and HOS cells, and then transfection efficiency was evaluated by RT-qPCR (Figure 2A, P < 0.05). The results of the CCK-8 assay revealed that U2OS and HOS cells with miR-618 upregulation had weaker proliferative (Figure 2B, P < 0.05) capacity than did the cells transfected with miR-NC. Additionally, the influence of miR-618 overexpression on the apoptosis of OS cells was analyzed by flow cytometry. Transfection with the miR-618 mimics notably elevated the percentages of apoptotic U2OS and HOS cells (Figure 2C, P < 0.05). Furthermore, recovery of miR-618 expression obviously attenuated migratory (Figure 2D, P < 0.05) and invasive (Figure 2E, P < 0.05) abilities of U2OS and HOS cells. Taken together, these data indicated that miR-618 inhibited the growth and metastasis of OS cells in vitro.
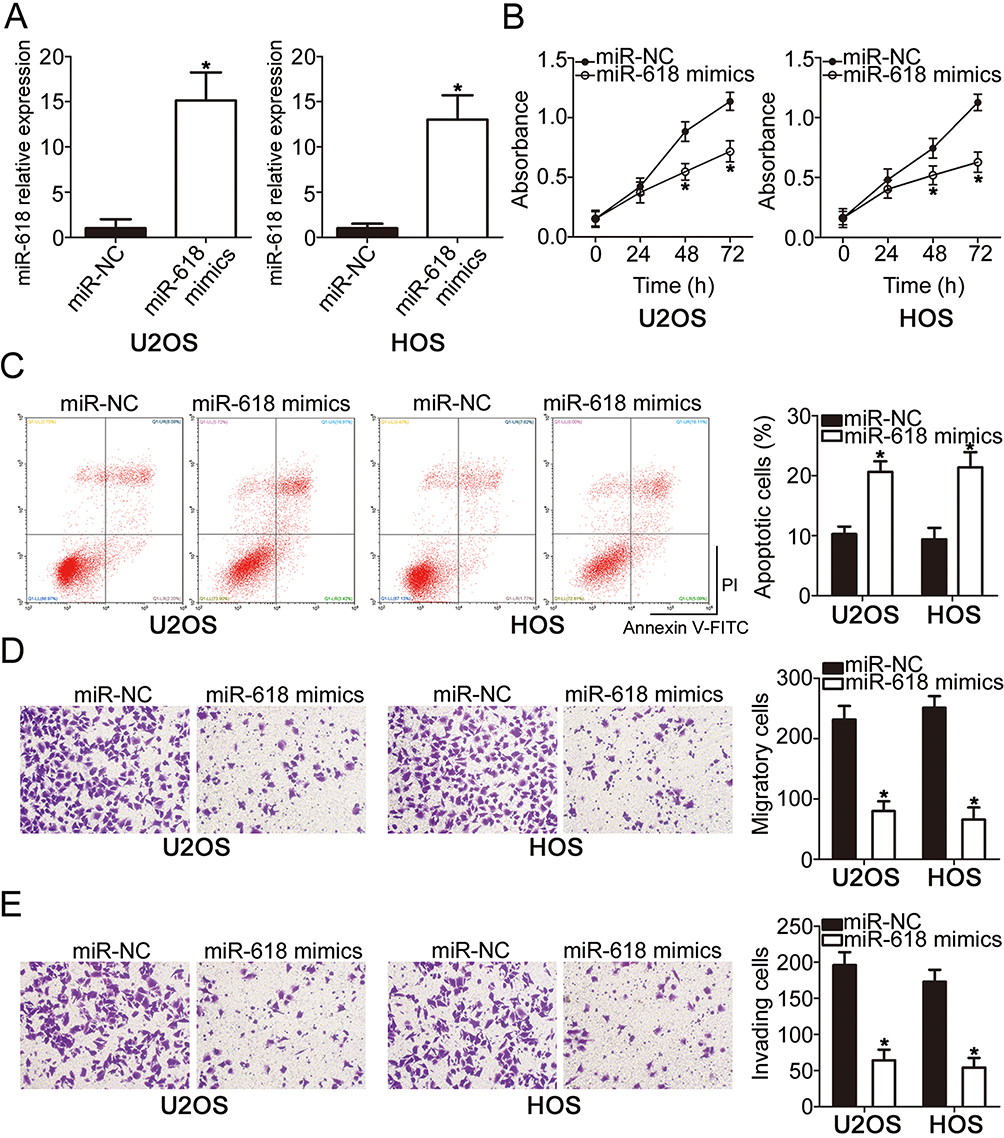 |
Figure 2 Resumption of miR-618 expression restrains the growth and metastasis of U2OS and HOS cells. (A) The expression level of miR-618 was measured in U2OS and HOS cells following transfection with the miR-618 mimics or miR-NC. *P < 0.05 vs group “miR-NC.” (B, C) The CCK-8 assay and flow-cytometric analysis were performed to examine the proliferation and apoptosis of miR-618 mimic–transfected or miR-NC–transfected U2OS and HOS cells. *P < 0.05 vs the miR-NC group. (D, E) The impact of miR-618 overexpression on U2OS and HOS cell migration and invasion was assessed in Transwell cell migration and invasion assays. *P < 0.05 vs the miR-NC group. |
MTDH Is A Direct Target Gene Of miR-618 In OS Cells
To gain insight into the mechanisms by which miR-618 suppresses the malignant phenotype of OS cells, we performed bioinformatics analysis to predict the putative targets of miR-618 and found that the 3′-UTR of MTDH contains a highly conserved binding site for miR-618 (Figure 3A). The luciferase reporter assay was carried out to test whether the 3′-UTR of MTDH could be directly targeted by miR-618 in OS cells. The results indicated that resumption of miR-618 expression significantly suppressed the luciferase activity generated by plasmid pMIR-MTDH-3ʹ-UTR-wt in U2OS and HOS cells (P < 0.05), but the suppressive effect was not observed in the cells harboring pMIR-MTDH-3ʹ-UTR-mut (Figure 3B). We next increased miR-618 expression in U2OS and HOS cells to test whether the expression of MTDH changed in response. Introduction of miR-618 evidently reduced MTDH expression in U2OS and HOS cells at mRNA (Figure 3C, P < 0.05) and protein levels (Figure 3D, P < 0.05). Furthermore, we detected the expression of MTDH protein in four OS cell lines and hFOB1.19. The analysis indicated that MTDH protein was overexpressed in four OS cell lines relative to that in hFOB1.19 (Figure 3E, P < 0.05). Moreover, we quantified MTDH expression in the 41 OS tissue samples and matching ANTs, revealing that the expression of MTDH mRNA was higher in OS tissue samples than in ANTs (Figure 3F, P < 0.05). The analysis of correlation between the expression levels of miR-618 and MTDH was conducted, and an inverse correlation between miR-618 and MTDH among the OS tissue samples was validated (Figure 3G; R2 = 0.3195, P = 0.0001). Taken together, these results meant that MTDH is a direct target gene of miR-618 in OS cells.
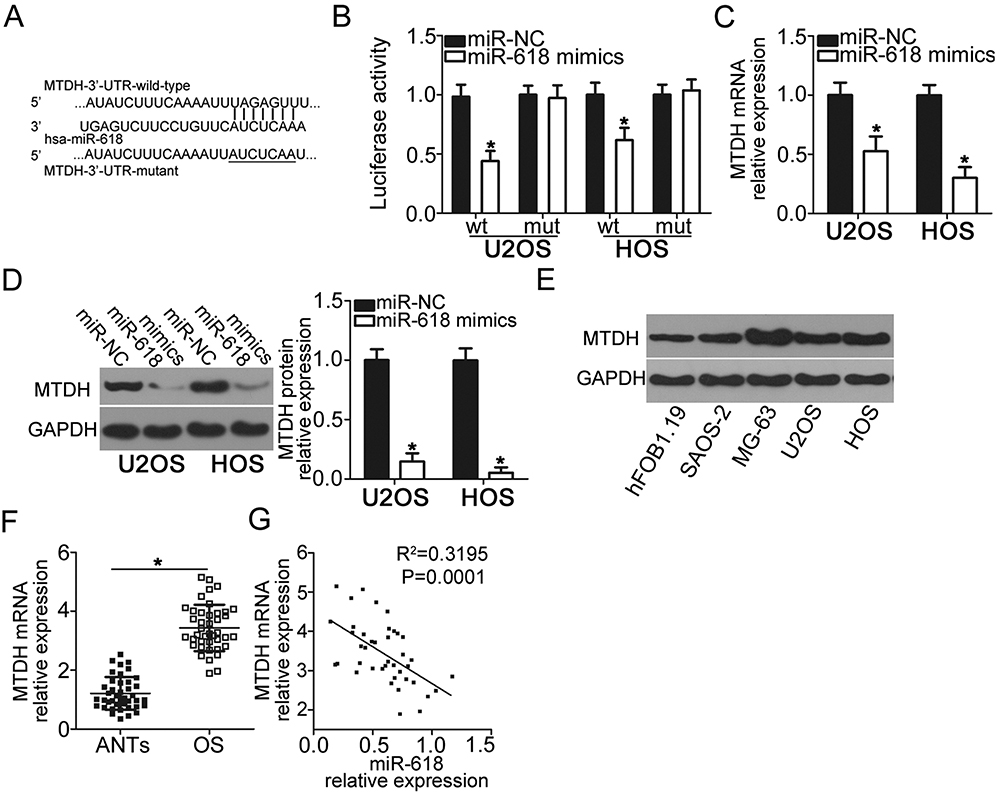 |
Figure 3 MTDH is a direct target gene of miR-618 in OS cells. (A) The 3′-UTR of the MTDH mRNA contains a potential miR-618–binding site. The mutant 3′-UTR region of the MTDH mRNA is also shown. (B) Plasmid pMIR-MTDH-3ʹ-UTR-wt or pMIR-MTDH-3ʹ-UTR-mut along with the miR-618 mimics or miR-NC was transfected into U2OS and HOS cells. After 48 h culture, the luciferase reporter assay was conducted to determine the luciferase activity. *P < 0.05 vs group miR-NC. (C, D) U2OS and HOS cells were transfected with the miR-618 mimics or miR-NC. The mRNA and protein levels of MTDH were measured by RT-qPCR and Western blot analysis, respectively. *P < 0.05 vs the miR-NC group. (E) Western blot analysis was conducted to detect MTDH protein expression in the human hFOB1.19 normal osteoblast cell line and four OS cell lines. (F) The mRNA expression of MTDH was detected in 41 pairs of OS tissue samples and ANT samples using RT-qPCR. *P < 0.05 vs ANTs. (G) Spearman correlation analysis was applied to assess the correlation between expression levels of miR-618 and MTDH mRNA in OS tissues. R2 = 0.3195, P = 0.0001. |
The MTDH Knockdown Simulates The Tumor-Suppressive Effects Of miR-618 In OS Cells
To explore the functions of MTDH in OS, si-MTDH was utilized to knock down endogenous MTDH expression in U2OS and HOS cells. Following si-MTDH transfection, Western blot analysis confirmed that the protein level of MTDH was efficiently knocked down in U2OS and HOS cells (Figure 4A, P < 0.05). Functional assays revealed that the downregulation of MTDH significantly slowed the proliferation (Figure 4B, P < 0.05), increased apoptosis (Figure 4C, P < 0.05), and attenuated migration (Figure 4D, P < 0.05) and invasiveness (Figure 4E, P < 0.05) of U2OS and HOS cells. Consequently, the MTDH knockdown exerted the effects similar to those of miR-618 upregulation in OS cells, thus confirming MTDH as a functional target of miR-618 in OS cells.
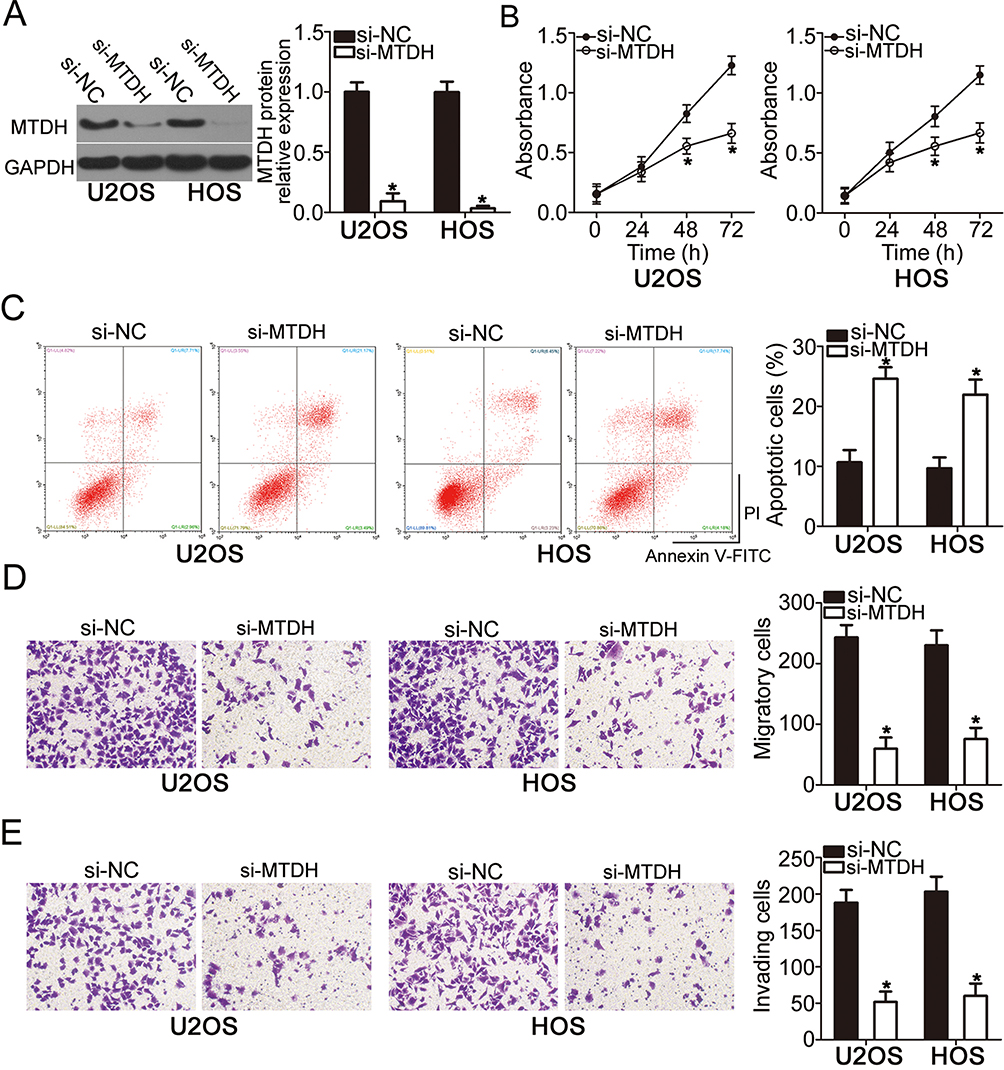 |
Figure 4 The knockdown of MTDH suppresses the proliferation, migration, and invasiveness but induces the apoptosis of U2OS and HOS cells. U2OS and HOS cells were transfected with either si-MTDH or si-NC and studied in the following assays. (A) At 72 h after transfection, the protein level of MTDH was determined by Western blot analysis. *P < 0.05 vs group si-NC. (B–E) The proliferation, apoptosis, migration, and invasion were assessed by the CCK-8 assay, flow-cytometric analysis, and Transwell migration and invasion assays, respectively. *P < 0.05 vs the si-NC group. |
MTDH Restoration Attenuates The Actions Of miR-618 Overexpression On OS Cells
To further clarify whether the decrease in MTDH expression by miR-618 upregulation was responsible for the suppression of OS aggressiveness, we restored MTDH expression in miR-618–overexpressing U2OS and HOS cells via cotransfection with MTDH-overexpressing plasmid pCMV-MTDH (Figure 5A, P < 0.05). Functional assays revealed that the effects of miR-618 upregulation on U2OS and HOS cell proliferation (Figure 5B, P < 0.05), apoptosis (Figure 5C, P < 0.05), migration (Figure 5D, P < 0.05), and invasion (Figure 5E, P < 0.05) were partially reversed by the recovery of MTDH expression. Thus, miR-618 performed tumor-suppressive functions in OS cells by downregulating MTDH.
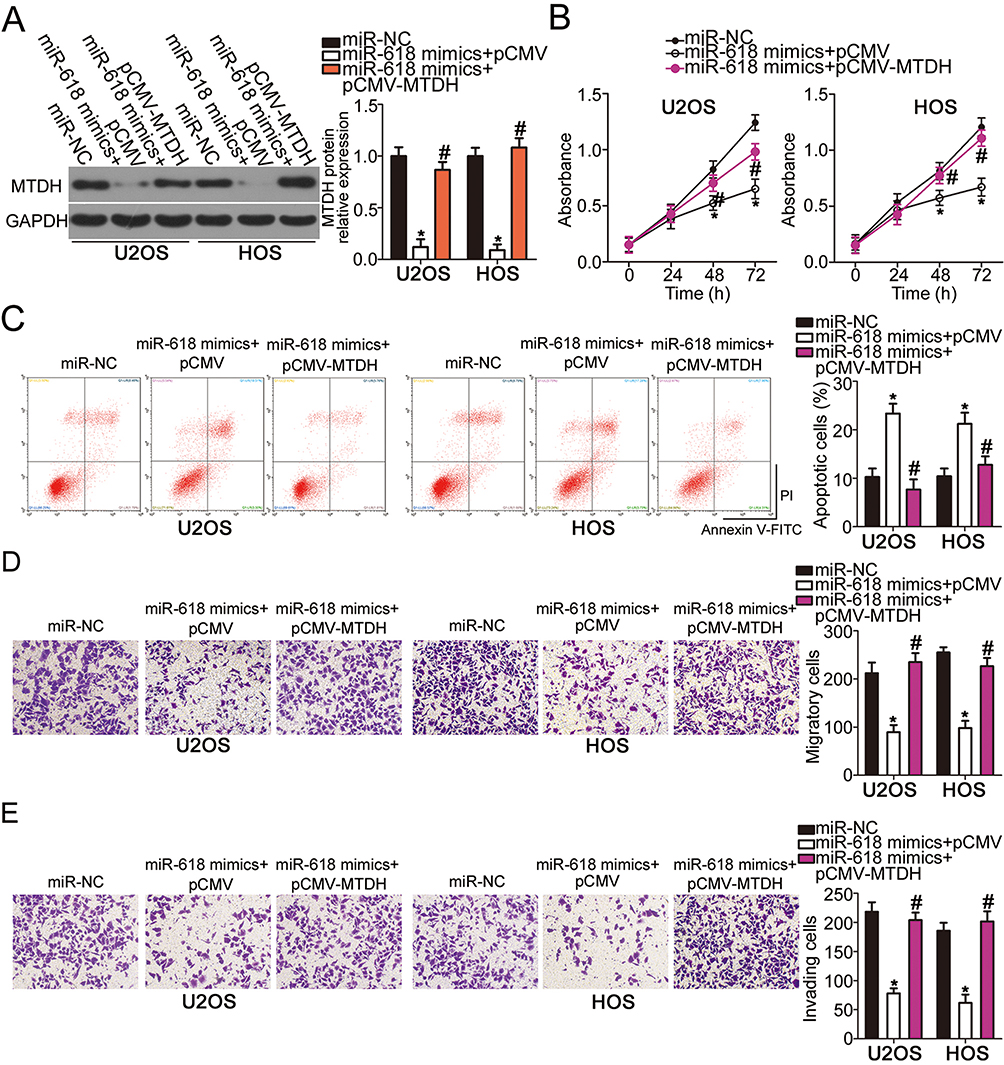 |
Figure 5 MTDH downregulation is required for the miR-618–driven inhibition of U2OS and HOS cell growth and metastasis in vitro. MiR-618–overexpressing U2OS and HOS cells were transfected with either plasmid pCMV-MTDH or the empty pCMV vector. (A) Transfected cells were collected after 72 h of incubation and subjected to Western blot analysis for the determination of MTDH protein expression. *P < 0.05 vs group miR-NC. #P < 0.05 vs the “miR-618 mimics+pCMV” group. (B–E) The proliferation, apoptosis, migration, and invasiveness of U2OS and HOS cells treated as described above were investigated by the CCK-8 assay, flow-cytometric analysis, and Transwell migration and invasion assays, respectively. *P < 0.05 vs group miR-NC. #P < 0.05 vs the miR-618 mimics+pCMV group. |
MiR-618 Decreases PTEN–AKT Signaling Output By Targeting MTDH mRNA In OS Cells
The PTEN–AKT pathway has been reported to be regulated by MTDH.19–21 Having identified MTDH as a direct target gene of miR-618, we next attempted to test whether miR-618 affects the PTEN–AKT pathway via MTDH downregulation. Hence, protein levels of PTEN, p-AKT, and AKT in U2OS and HOS cells after cotransfection with the miR-618 mimics and either plasmid pCMV-MTDH or the empty pCMV vector were evaluated by Western blotting. The upregulation of miR-618 significantly increased PTEN and decreased p-AKT amounts in U2OS and HOS cells, while total AKT expression was unaffected (Figure 6). Notably, restoration of MTDH expression partially reversed the changes in PTEN and p-AKT protein levels caused by miR-618 overexpression (Figure 6A). Similarly, we investigated whether MTDH knockdown was able to mimic the influence of miR-618 overexpression on PTEN–AKT pathway in OS cells. As expected, interference of MTDH expression increased PTEN and reduced p-AKT expression in U2OS and HOS cells (Figure 6B). These results meant that miR-618 diminished the PTEN–AKT signaling output in OS cells by directly targeting MTDH mRNA and downregulating MTDH.
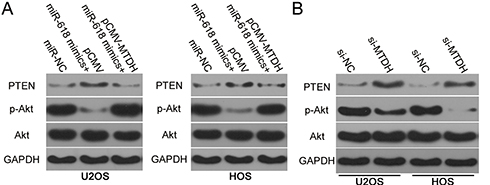 |
Figure 6 MiR-618 inhibits activation of the PTEN–Akt pathway in OS cells by targeting MTDH. (A) pCMV-MTDH or pCMV was transfected into U2OS and HOS cells in the presence of the miR-618 mimics. After that, protein levels of PTEN, p-AKT, and AKT were assayed by Western blotting. (B) Western blotting was utilized to determine PTEN, p-AKT, and AKT expression in U2OS and HOS cells after si-NC or si-MTDH injection. |
MiR-618 Slows The OS Tumor Growth In Vivo
Next, a tumor xenograft experiment was conducted to examine the impact of miR-618 on OS cell tumorigenicity in vivo. HOS cells transfected with the miR-618 mimics were inoculated into nude mice, and miR-NC–transfected cells served as a control. The volume and weight of tumor xenografts derived from the miR-618 mimic–transfected HOS cells were notably lower (Figure 7A and B, P < 0.05; Figure 7C, P < 0.05) than those in the miR-NC group. Next, RT-qPCR was carried out to quantitate miR-618 expression in the tumor xenografts. Higher miR-618 expression was observed in the tumor xenograft of the miR-618 mimic group as compared with the miR-NC group (Figure 7D, P < 0.05). Furthermore, the protein levels of MTDH, PTEN, p-AKT, and AKT in the tumor xenografts were determined via Western blot analysis. The results revealed that MTDH and p-AKT protein amounts evidently decreased whereas the PTEN protein amount significantly increased in the miR-618 mimic–treated nude mouse group (Figure 7E). Taken together, these results implied that miR-618 inhibited the tumor growth of OS cells in vivo, and the growth inhibition was achieved through inhibition of the MTDH–PTEN–AKT pathway output.
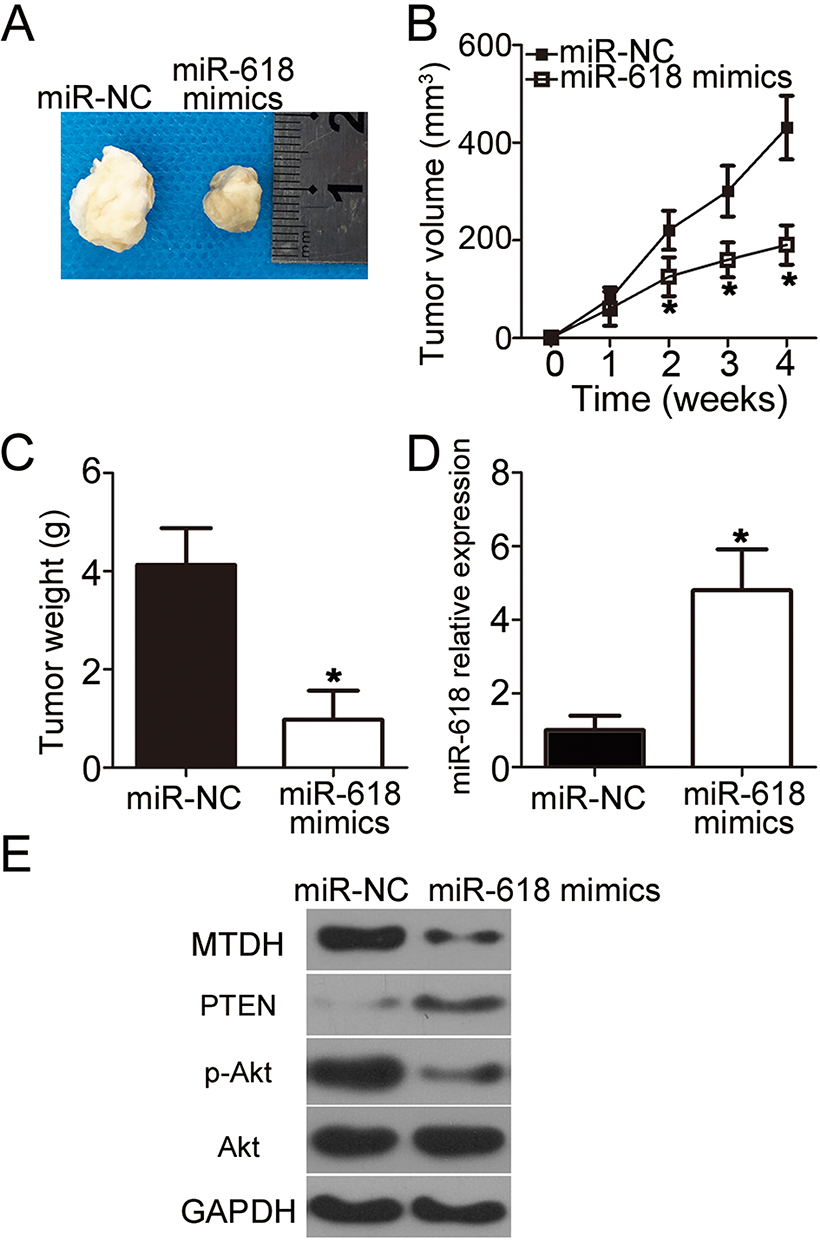 |
Figure 7 MiR-618 upregulation impairs OS tumor growth in vivo. (A) Representative images of tumor xenografts derived from HOS cells transfected with the miR-618 mimics or miR-NC. (B) The volume of tumor xenografts in the miR-618 mimic group was smaller than that in the miR-NC group. *P < 0.05 compared with group miR-NC. (C) Tumor xenografts in the miR‐618 mimic group and miR-NC group were excised and weighed at 4 weeks after implantation. *P < 0.05 vs miR-NC. (D) RT-qPCR was performed to analyze miR-618 expression in the tumor xenografts. *P < 0.05 vs group miR-NC. (E) Protein amounts of MTDH, PTEN, p-AKT, and AKT in tumor xenografts were quantified by Western blotting. |
Discussion
An increasing number of studies has shown that the accumulation of genetic and epigenetic alterations may be closely related to OS initiation and progression.22–24 A variety of miRNAs are dysregulated in OS and contribute to the tumorigenic processes.25–27 Hence, further research into the miRNAs that play important roles in the aggressive behaviors of OS is necessary to identify candidate targets for the treatment of patients with OS. MiR-618 is underexpressed in prostate cancer15 and thyroid carcinoma.16,17 Patients with prostate cancer harboring a low miR-618 level have worse outcomes than do the patients with high miR-618 levels.15 However, the expression level of miR-618 in OS has remained unclear. In this study, for the first time, we demonstrated that miR-618 is downregulated in both OS tissues and cell lines. Low miR-618 expression correlated with the clinical stage and distant metastasis among our patients with OS. These results suggest that miR-618 might be an effective biomarker for the prognosis of OS.
MiR-618 performs tumor-suppressive functions in carcinogenesis and cancer progression. For instance, miR-618 upregulation suppresses the metastasis and promotes mesenchymal–epithelial transition of prostate cancer cells by directly targeting Forkhead box p2.15 Resumption of miR-618 expression restricts thyroid cancer cell growth and metastasis and induces G2–M arrest via the blockade of X-linked inhibitor of apoptosis protein and via deactivation of the PI3K–AKT signaling pathway.16,17 Nevertheless, little is known about the specific roles of miR-618 in OS. Herein, functional experiments revealed that overexpression of miR-618 inhibited OS cell proliferation, migration, and invasion and promoted apoptosis in vitro. Besides, exogenous miR-618 expression retarded OS growth in vivo. Previous study revealed that autophagy induction is a mechanism for cell detah. However, we did not tested whether autophagy induction was related with the proliferation inhibition caused by miR-618 overexpression. It was a limitation of our study, and we will resolve it in our following investigations. This study provides clues to the profound involvement of miR-618 in OS and suggests that miR-618 might be a potential target for treating patients with OS.
MiRNAs play their important part in the tumorigenesis and tumor progression by directly regulating the expression of their target genes.28 Accordingly, we next attempted to identify the direct target gene that is involved in the anticancer actions of miR-618 in OS cells. Bioinformatics analysis was performed first, to predict the putative target of miR-618. The 3′-UTR of MTDH was found to contain a highly conserved binding site for miR-618. Then, the luciferase activity assay was performed to verify the targeting of miR-618 to the 3′-UTR of MTDH mRNA. Furthermore, miR-618 overexpression successfully decreased endogenous MTDH expression at both mRNA and protein levels in OS cells. MTDH turned out to be upregulated in OS tissue samples, and the upregulation of MTDH inversely correlated with miR-618 expression. Suppression of MTDH expression simulated the tumor-suppressive action of miR-618 overexpression in OS cells. Subsequent rescue experiments confirmed that restoration of MTDH expression partially reversed the miR-618–mediated tumor-suppressive effects on OS cells. These observations provided sufficient evidence to designate MTDH as a direct target gene of miR-618 in OS cells.
MTDH, also known as astrocyte-elevated gene 1, is located in chromosomal region 8q22.29 It is reported to be upregulated in various cancers and is associated with cancer progression.30–32 Increased MTDH expression significantly correlates with gender, clinical stages, classification, metastasis, differentiation, and poor survival of patients with OS.33 High MTDH expression also strongly correlates with the poorer prognosis of patients with OS.33,34 MTDH performs oncogenic functions in the malignant progression of OS by regulating cell proliferation, apoptosis, migration, invasion, metastasis, epithelial–mesenchymal transition, and chemoresistance.33–36 Notably, MTDH has been reported to be directly targeted and regulated by various miRNAs in different human cancers. For example, miR-136,37 miR-342-3p,38 miR-448,39 and miR-50640 directly target MTDH mRNA to inhibit the malignant progression of OS. Hence, targeting MTDH by miRNAs is a promising modality for the prevention and treatment of OS.
Conclusion
Our results for the first time revealed that miR‐618 functions as a tumor suppressor during OS progression by directly targeting MTDH and reducing PTEN–AKT pathway output. Hence, this study provides functional evidence fully supporting the hypothesis that miR-618 is a promising target for the management of OS.
Abbreviations
3′-UTR, 3′-untranslated region; ANT, adjacent normal tissue; CCK-8, Cell Counting Kit-8; DMEM, Dulbecco’s Modified Eagle’s Medium; FBS, fetal bovine serum; FITC, fluorescein Isothiocyanate; miRNA, miR, microRNA; mut, mutant; NC, negative control; OS, osteosarcoma; p-AKT, phospho-AKT; RT-qPCR, reverse-transcription quantitative PCR; TBST, Tris-buffered saline with 0.05% of Tween 20; wt, wild-type.
Availability Of Data And Materials
The datasets used and/or analyzed during this study are available from the corresponding author on reasonable request.
Author Contributions
Chunzheng Gao designed this study and performed all statistical analyses. Bohan Li, Jie Zhao, and Qian Zhao carried out the RT-qPCR, Western blotting, and luciferase reporter assays. The CCK-8 and Transwell invasion assays were conducted by Dongjin Wu, Cheng Zhang, and Kun Zhao. Yang Song performed the tumor xenograft assay. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Ethics Approval And Consent To Participate
The Ethics Committee of the Second Hospital of Shandong University approved the study protocol, and all the patients provided written informed consent. All the animal experimental procedures were approved by the Ethics Committee of the Second Hospital of Shandong University and were carried out in accordance with the Animal Protection Law of the People’s Republic of China-2009.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Ottaviani G, Jaffe N. The epidemiology of osteosarcoma. Cancer Treat Res. 2009;152:3–13.
2. Siegel HJ, Pressey JG. Current concepts on the surgical and medical management of osteosarcoma. Expert Rev Anticancer Ther. 2008;8(8):1257–1269. doi:10.1586/14737140.8.8.1257
3. Bielack SS, Hecker-Nolting S, Blattmann C, Kager L. Advances in the management of osteosarcoma. F1000Research. 2016;5:2767. doi:10.12688/f1000research
4. Anderson ME. Update on survival in osteosarcoma. Orthop Clin North Am. 2016;47(1):283–292. doi:10.1016/j.ocl.2015.08.022
5. Marina N, Gebhardt M, Teot L, Gorlick R. Biology and therapeutic advances for pediatric osteosarcoma. Oncologist. 2004;9(4):422–441. doi:10.1634/theoncologist.9-4-422
6. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
7. Makarova JA, Shkurnikov MU, Wicklein D, et al. Intracellular and extracellular microRNA: an update on localization and biological role. Prog Histochem Cytochem. 2016;51(3–4):33–49. doi:10.1016/j.proghi.2016.06.001
8. Link A, Kupcinskas J. MicroRNAs as non-invasive diagnostic biomarkers for gastric cancer: current insights and future perspectives. World J Gastroenterol. 2018;24(30):3313–3329. doi:10.3748/wjg.v24.i30.3313
9. Iqbal MA, Arora S, Prakasam G, Calin GA, Syed MA. MicroRNA in lung cancer: role, mechanisms, pathways and therapeutic relevance. Mol Aspects Med. 2018. doi:10.1016/j.mam.2018.07.003
10. To KK, Tong CW, Wu M, Cho WC. MicroRNAs in the prognosis and therapy of colorectal cancer: from bench to bedside. World J Gastroenterol. 2018;24(27):2949–2973. doi:10.3748/wjg.v24.i27.2949
11. Ram Kumar RM, Boro A, Fuchs B. Involvement and clinical aspects of microRNA in osteosarcoma. Int J Mol Sci. 2016;17(6):877. doi:10.3390/ijms17060877
12. Wang X, Peng L, Gong X, Zhang X, Sun R, Du J. miR-423-5p inhibits osteosarcoma proliferation and invasion through directly targeting STMN1. Cell Physiol Biochem. 2018;50(6):2249–2259. doi:10.1159/000495085
13. Wang DZ, Jing SF, Hao SB, Huang XY, Miao QT, Gao JF. MiR-218 promotes apoptosis of U2OS osteosarcoma cells through targeting BIRC5. Eur Rev Med Pharmacol Sci. 2018;22(20):6650–6657. doi:10.26355/eurrev_201810_16140
14. Lin ZW, Zhang W, Jiang SD, Wei WB, Li XF. Inhibition of microRNA-940 suppresses the migration and invasion of human osteosarcoma cells through the secreted frizzled-related protein 1-mediated Wnt/beta-catenin signaling pathway. J Cell Biochem. 2018.
15. Song XL, Tang Y, Lei XH, Zhao SC, Wu ZQ. miR-618 inhibits prostate cancer migration and invasion by targeting FOXP2. J Cancer. 2017;8(13):2501–2510. doi:10.7150/jca.17407
16. Yi L, Yuan Y. MicroRNA-618 modulates cell growth via targeting PI3K/Akt pathway in human thyroid carcinomas. Indian J Cancer. 2015;52(Suppl 3):E186–E189. doi:10.4103/0019-509X.186577
17. Cheng Q, Zhang X, Xu X, Lu X. MiR-618 inhibits anaplastic thyroid cancer by repressing XIAP in one ATC cell line. Ann D’endocrinologie. 2014;75(4):187–193. doi:10.1016/j.ando.2014.01.002
18. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
19. Zhang L, Li C, Cao L, et al. microRNA-877 inhibits malignant progression of colorectal cancer by directly targeting MTDH and regulating the PTEN/Akt pathway. Cancer Manag Res. 2019;11:2769–2781. doi:10.2147/CMAR.S194073
20. Li L, Zhang H. MicroRNA-379 inhibits cell proliferation and invasion in glioma via targeting metadherin and regulating PTEN/AKT pathway. Mol Med Rep. 2018;17(3):4049–4056. doi:10.3892/mmr.2017.8361
21. Li J, Li C, Li H, et al. MicroRNA30a5p suppresses tumor cell proliferation of human renal cancer via the MTDH/PTEN/AKT pathway. Int J Mol Med. 2018;41(2):1021–1029. doi:10.3892/ijmm.2017.3269
22. Varshney J, Scott MC, Largaespada DA, Subramanian S. Understanding the osteosarcoma pathobiology: a comparative oncology approach. Vet Sci. 2016;17(6):3. doi:10.3390/vetsci3010003
23. Li B, Ye Z. Epigenetic alterations in osteosarcoma: promising targets. Mol Biol Rep. 2014;41(5):3303–3315. doi:10.1007/s11033-014-3193-7
24. Tang N, Song WX, Luo J, Haydon RC, He TC. Osteosarcoma development and stem cell differentiation. Clin Orthop Relat Res. 2008;466(9):2114–2130. doi:10.1007/s11999-008-0335-z
25. Zhang XD, Wang YN, Feng XY, Yang JY, Ge YY, Kong WQ. Biological function of microRNA-30c/SOX9 in pediatric osteosarcoma cell growth and metastasis. Eur Rev Med Pharmacol Sci. 2018;22(1):70–78. doi:10.26355/eurrev_201801_14102
26. Zhang H, He QY, Wang GC, et al. miR-422a inhibits osteosarcoma proliferation by targeting BCL2L2 and KRAS. Biosci Rep. 2018;38(2).
27. Sun X, Dai G, Yu L, Hu Q, Chen J, Guo W. miR-143-3p inhibits the proliferation, migration and invasion in osteosarcoma by targeting FOSL2. Sci Rep. 2018;8(1):606. doi:10.1038/s41598-017-18739-3
28. Harries LW. Long non-coding RNAs and human disease. Biochem Soc Trans. 2012;40(4):902–906. doi:10.1042/BST20120020
29. Anttila V, Stefansson H, Kallela M, et al. Genome-wide association study of migraine implicates a common susceptibility variant on 8q22.1. Nat Genet. 2010;42(10):869–873.
30. Ma Z, Chen Y, Dong S, et al. AEG-1 mRNA expression in non-small cell lung cancer is associated with increased tumor angiogenesis. Pathol Res Pract. 2017;213(10):1257–1263. doi:10.1016/j.prp.2017.09.003
31. Huang LL, Wang Z, Cao CJ, et al. AEG-1 associates with metastasis in papillary thyroid cancer through upregulation of MMP2/9. Int J Oncol. 2017;51(3):812–822. doi:10.3892/ijo.2017.4074
32. Wu S, Yang L, Wu D, et al. AEG-1 induces gastric cancer metastasis by upregulation of eIF4E expression. J Cell Mol Med. 2017;21(12):3481–3493. doi:10.1111/jcmm.2017.21.issue-12
33. Wang F, Ke ZF, Sun SJ, et al. Oncogenic roles of astrocyte elevated gene-1 (AEG-1) in osteosarcoma progression and prognosis. Cancer Biol Ther. 2011;12(6):539–548. doi:10.4161/cbt.12.6.16301
34. Tang J, Shen L, Yang Q, Zhang C. Overexpression of metadherin mediates metastasis of osteosarcoma by regulating epithelial-mesenchymal transition. Cell Prolif. 2014;47(5):427–434. doi:10.1111/cpr.2014.47.issue-5
35. Liu B, Wu Y, Peng D. Astrocyte elevated gene-1 regulates osteosarcoma cell invasion and chemoresistance via endothelin-1/endothelin A receptor signaling. Oncol Lett. 2013;5(2):505–510. doi:10.3892/ol.2012.1056
36. Wang F, Ke ZF, Wang R, Wang YF, Huang LL, Wang LT. Astrocyte elevated gene-1 (AEG-1) promotes osteosarcoma cell invasion through the JNK/c-Jun/MMP-2 pathway. Biochem Biophys Res Commun. 2014;452(4):933–939. doi:10.1016/j.bbrc.2014.09.009
37. Guo T, Pan G. MicroRNA-136 functions as a tumor suppressor in osteosarcoma via regulating metadherin. Cancer Biomarkers. 2018;22(1):79–87. doi:10.3233/CBM-170970
38. Zhang S, Liu L, Lv Z, Li Q, Gong W, Wu H. MicroRNA-342-3p inhibits the proliferation, migration, and invasion of osteosarcoma cells by targeting astrocyte-elevated gene-1 (AEG-1). Oncol Res. 2017;25(9):1505–1515. doi:10.3727/096504017X14886485417426
39. Jiang W, Wang S, Sun Y, Jiang Y, Yu T, Wang J. Overexpression of microRNA-448 inhibits osteosarcoma cell proliferation and invasion through targeting of astrocyte elevated gene-1. Mol Med Rep. 2017;16(4):5713–5721. doi:10.3892/mmr.2017.7249
40. Yao J, Qin L, Miao S, Wang X, Wu X. Overexpression of miR-506 suppresses proliferation and promotes apoptosis of osteosarcoma cells by targeting astrocyte elevated gene-1. Oncol Lett. 2016;12(3):1840–1848. doi:10.3892/ol.2016.4827
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.