Back to Journals » Journal of Inflammation Research » Volume 18
Microbiome–Gut–Brain Axis: Implications for the Links Between Inflammatory Bowel Disease and Neuropsychiatric Disorders
Authors Tian CM, Yang MF, Kong C, Luo D, Yue NN, Zhao HL, Zhang Y ![]() , Lu JP, Liang YJ
, Lu JP, Liang YJ ![]() , Song Y, Wei DR, Yao J
, Song Y, Wei DR, Yao J ![]() , Wang LS
, Wang LS ![]() , Li DF
, Li DF ![]()
Received 29 December 2024
Accepted for publication 1 July 2025
Published 23 September 2025 Volume 2025:18 Pages 13183—13212
DOI https://doi.org/10.2147/JIR.S514838
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam Bachstetter
Cheng-Mei Tian,1,2,* Mei-Feng Yang,3,* Chen Kong,1,* Duo Luo,4,* Ning-Ning Yue,1 Hai-Lan Zhao,1 Yuan Zhang,5 Jian-Ping Lu,6 Yu-Jie Liang,6,7 Yang Song,1 Dao-Ru Wei,8 Jun Yao,1 Li-Sheng Wang,1 De-Feng Li1
1Department of Gastroenterology, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, Guangdong, 518020, People’s Republic of China; 2Department of Emergency, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; the First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, Guangdong, 518020, People’s Republic of China; 3Department of Hematology, Yantian District People’s Hospital, Shenzhen, Guangdong, 518020, People’s Republic of China; 4Department of Geriatrics, Guangzhou First People’s Hospital, Guangzhou Medical University, Guangzhou, 510180, People’s Republic of China; 5Department of Medical Administration, Huizhou Institute of Occupational Diseases Control and Prevention, Huizhou, Guangdong, 516000, People’s Republic of China; 6Department of Child and Adolescent Psychiatry, Shenzhen Institute of Mental Health, Shenzhen Kangning Hospital, Shenzhen, 518020, People’s Republic of China; 7Jining Medical University, Jining, 272000, People’s Republic of China; 8Department of Rehabilitation, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, Guangdong, 518020, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dao-Ru Wei, Department of Rehabilitation, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), No. 1017, Dongmen North Road, Luohu District, Shenzhen, 518020, People’s Republic of China, Tel +86 755 25533018, Email [email protected] De-Feng Li, Department of Gastroenterology, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; the First Affiliated Hospital, Southern University of Science and Technology), No. 1017, Dongmen North Road, Luohu District, Shenzhen, 518020, People’s Republic of China, Tel +86 755 25533018, Email [email protected]
Abstract: The brain and gastrointestinal tract are closely interconnected as important sensory organs processing signals from both environmental and internal cues. Recent studies have shown that dysregulation of the gut microbiota in inflammatory bowel disease activates the gut immune system. The cross-talk mechanism along the gut-brain axis is implicated in the development of neuropsychiatric disorders such as autism, depression, anxiety, Alzheimer’s disease, and Parkinson’s disease. Here, we discuss the molecular mechanisms involved in signaling across the gut-brain axis, including the immune and neuroendocrine system, intestinal permeability, microbial composition, and bacterial extracellular vesicles. We focus on the link between specific inflammatory bowel disease, microbial genera and psychiatric and neurological disorders, and propose that the results of preclinical and clinical studies open up the possibility of targeting the gut microbiota to treat neuropsychiatric disorders that are altered by gut interactions.
Plain Language Summary: There is a strong bidirectional communication between the brain and the digestive system. Dysregulation of the gut microbiota (GM) in inflammatory bowel disease (IBD) activates the gut immune system and contributes to brain disorders. Targeting the gut-brain axis has the potential to treat both neurological disorders such as depression, autism, Alzheimer’s disease, and Parkinson’s syndrome, and to ameliorate intestinal disorders such as IBD.
Keywords: Gut–brain axis, GBA, microbiome–gut–brain axis, MGBA, inflammatory bowel disease, IBD, neuropsychiatric disorders, probiotics, psychobiotics
Graphical Abstract:

Introduction
The gut has certain self-regulatory functions. However, the gut is regulated by multiple systems such as neural, immune, and endocrine systems.1 In addition, the gut is influenced by a complex intestinal environment constructed by the gut microbiota (GM), chemicals and physical conditions, in which GM serves the most important extrinsic factor.1 Studies have shown that disorders of the GM are involved in the pathogenesis and development of inflammatory bowel disease (IBD), and interventions in disorders of the GM have become an important tool in the treatment of IBD.2
The brain is a regulatory organ, the high-level center of the nervous system. Through the nervous system, the brain regulates the functional activity of almost all organs of the body.3 The brain is also a functional organ capable of generating consciousness through mental-psychological and behavioral activities.3 At the same time, changes in gut function and alterations in the GM transmit information to the brain through multiple pathways, affect the regulatory and higher functions of the brain.4 It has been found that IBD is often accompanied by neuropsychiatric symptoms, increasing the prevalence of each other, and there is a reciprocal facilitation effect in treatment.2 There is also a strong link between disturbances in the GM and brain dysfunction. There is growing evidence of a gut-brain associated with aspects of their own or their children’s neurodevelopment, neuropsychiatric disorders and the incidence of neurodegenerative diseases (Figure 1).4
 |
Figure 1 Active role of gut microbiota in brain development. The gut microbiome plays a crucial role in various processes of brain development, such as the establishment of the blood-brain barrier (BBB), neurogenesis, myelination, microglia maturation, the development and maintenance of the microglial cells, the development of the HPA axis, and the stress response. |
This review discusses the influence of the GM in the occurrence and development of IBD, and focuses on the involvement of GM in the occurrence and development of neuropsychiatric disorders, such as depression, anxiety disorders, autism spectrum disorder (ASD), schizophrenia (SCZ), Alzheimer’s disease (AD), and Parkinson’s disease (PD). In addition to discuss the comorbidity of IBD and mental health disorders, this review explores the potential therapeutic value of the GM for IBD and neuropsychiatric disorders by summarizing the effects of the GM on the gut and brain. We searched for disease-specific diagnostic markers based on structural and omics changes in GM. Using GM as a therapeutic target, we study the effects of diet or medication on GM, and explore the therapeutic effects of GM interventions such as gut microecological regulators and GM transplantation on IBD or neuropsychiatric disorders.
Microbiome–Gut–Brain Axis (MGBA)
The gut and brain are important organs in the human body, with their own distinct anatomical locations, independent structures, and specific physiological functions.5 However, they are regulated by several systems of the human body, influenced by the surrounding environment, and had a complete regulatory system.5 The developing brain requires the availability of dietary components from the gut for the maturation and normal function of nerve cells. There is now clear evidence that gut microbes directly contribute to brain development and play an active role in neurodevelopmental processes (Figure 2) including, neurogenesis, microglia maturation and myelination. These processes are critical in shaping animal behavior and cognition.
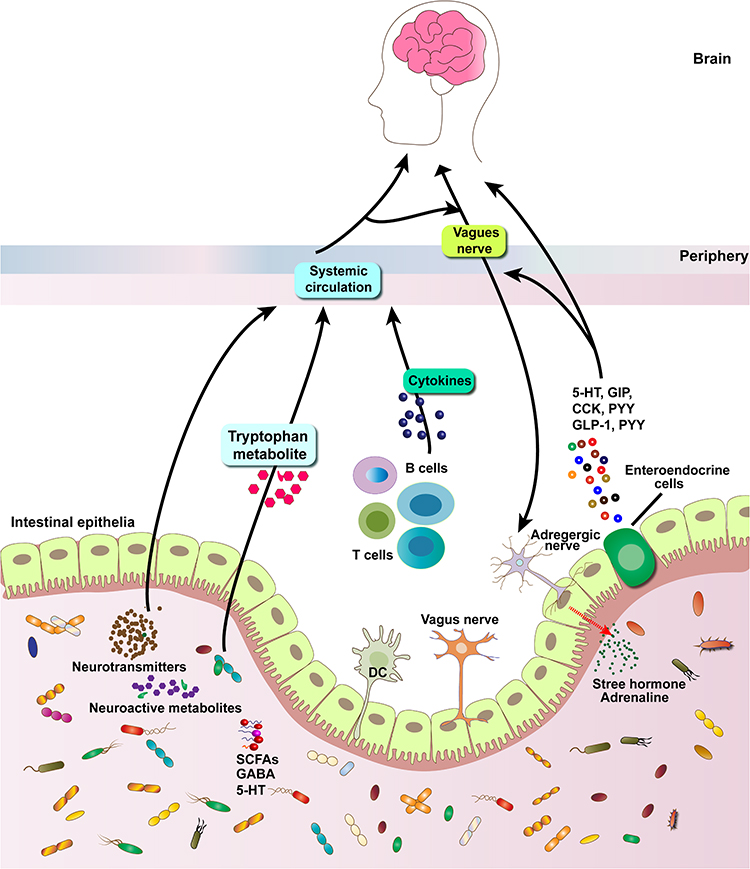 |
Figure 2 Schematic diagram outlining known bi-directional communication pathways between the brain-gut-microbiome, including immunomodulatory responses, neuronal innervation, gut endocrine and microbial metabolite signaling. Short-chain fatty acids, γ-aminobutyric acid, 5-hydroxytryptamine, norepinephrine, and dopamine, such as SCFAs produced by many commensal bacteria, have this neuro-immunomodulatory role. Intestinal mucosal cells (green) contain more than 90% of the body’s serotonin (5-HT). Abbreviations: CCK, cholecystokinin; GLP-1, glucagon-like peptide-1; PYY, peptide YY; 5-HT, serotonin. |
The digestive tract, whose main task is to process ingested food and defend against invading microorganisms, as well as plays an important habitat for microorganisms, forming the “second gene pool” of the host and having a great impact on host health or disease.2,6 The gut is home to a large number of regulatory cells of the host, such as nerve cells, immune cells and endocrine cells, being “second brain” of the host, with strong defensive and regulatory capabilities.6 The functional activity of the brain is not only regulated by the nervous system, but also associated with the immune and endocrine systems.7 Moreover, the brain is also influenced by the microorganisms colonized or invaded the host.4 The general connection between gut and brain, termed the gut-brain axis (GBA), establishes neural and circulatory networks that enable mutual interaction, regulation and control.8
GM is classified as resident or transient microorganisms. Resident microorganisms are able to adhere and colonize the intestinal tract, and persist and multiply in the gut.9 Passage microorganisms cannot adhere and colonize, and are easily carried away by feces.7 The GM exchanges material, energy and information in the intestinal microenvironment.10 Microorganisms accompany food into the gut, and they carry a large number of antigenic components, and produce a large number of metabolites.11 Antigenic molecules are recognized by intestinal immune cells, and activated intestinal immune responses.12 Some metabolites are able to cross the intestinal barrier, and reach the target cells through the intestinal neural or circulatory network, affecting the function of the host target organ.13 GM play an important role in GBA by influencing the physiological functions of the gut and brain through multiple pathways, as well as being regulated by the gut and nervous system.14 GM join the GBA as a foreign “biological organ” to form the MGBA (Figure 2).14,15 Studies have shown that psychiatric disorders, such as depression, anxiety disorders, ASD, SCZ, and neurodegenerative pathologies including AD and PD, are associated with gut or GM4,16–18(Table 1).
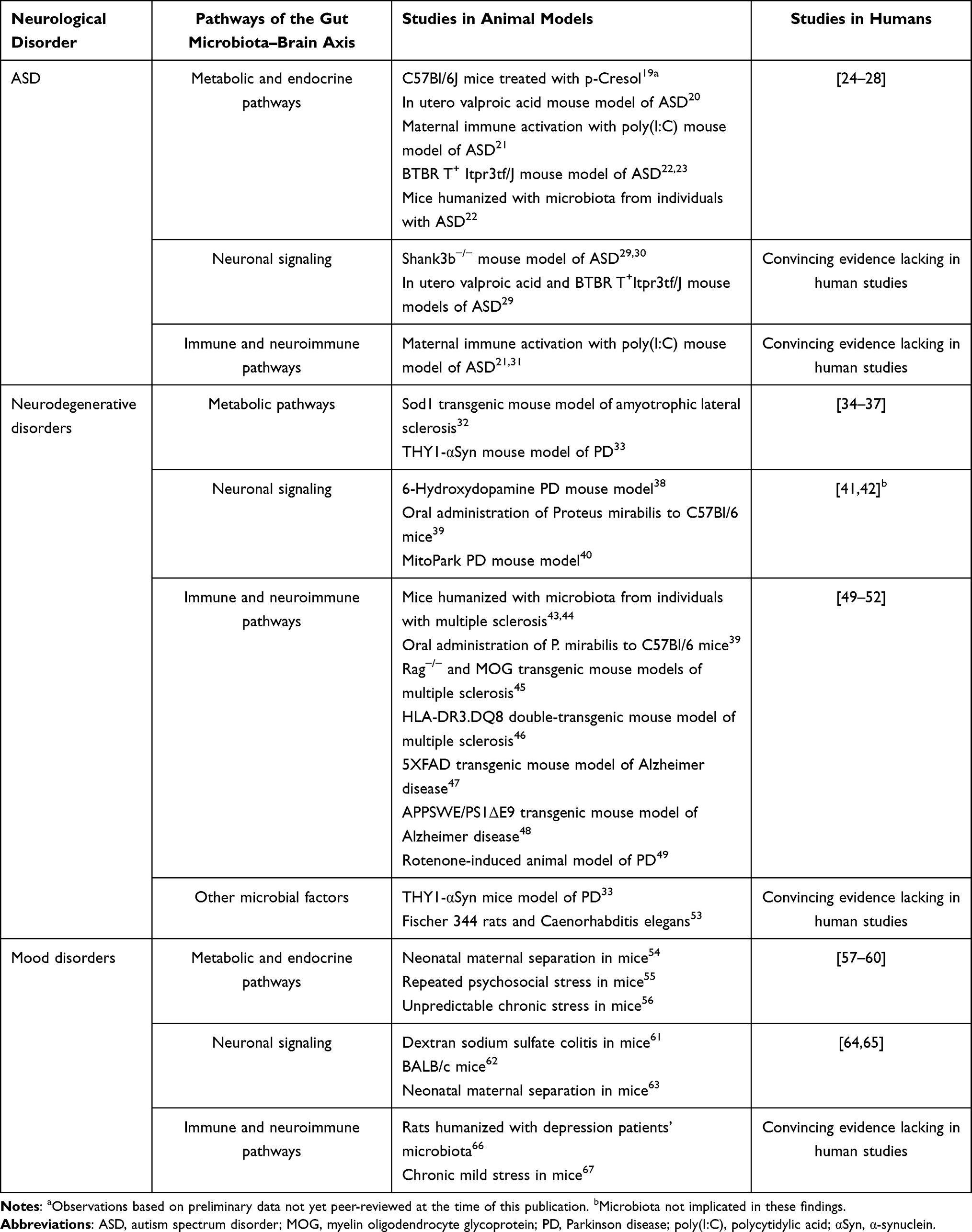 |
Table 1 Microbiota-Gut–Brain Axis Pathways and Neurological Disorders |
MGBA Communication Pathway
The mechanism of MGBA action, involving interactions among GM, the gut, and the brain, remains incompletely elucidated.68,69 The MGBA is the interconnection among cell-cell, cell-microorganism, as well as the transport and effects of GM-derived bioactive molecules in the host body.70 Physical barriers such as the intestinal epithelium and blood-brain barrier restrict direct microbial-host contact, necessitating chemical signaling via GM-derived molecules.71 GM derived signaling molecule crossing the intestinal barrier acts directly on intestinal target cells, including peripheral neurons, intestinal immune cells, and intestinal endocrine cells, converting them into neural signals, immune signals, and endocrine signals and becoming pathways to regulate host target organs.14,72 Accordingly, MGBA communication pathways are broadly classified into peripheral neural, immune, endocrine, and integrated communication pathways.73,74 The GM affects the development and function of the brain through multiple pathways such as peripheral nerve, immune, and endocrine72,75 (Table 2).
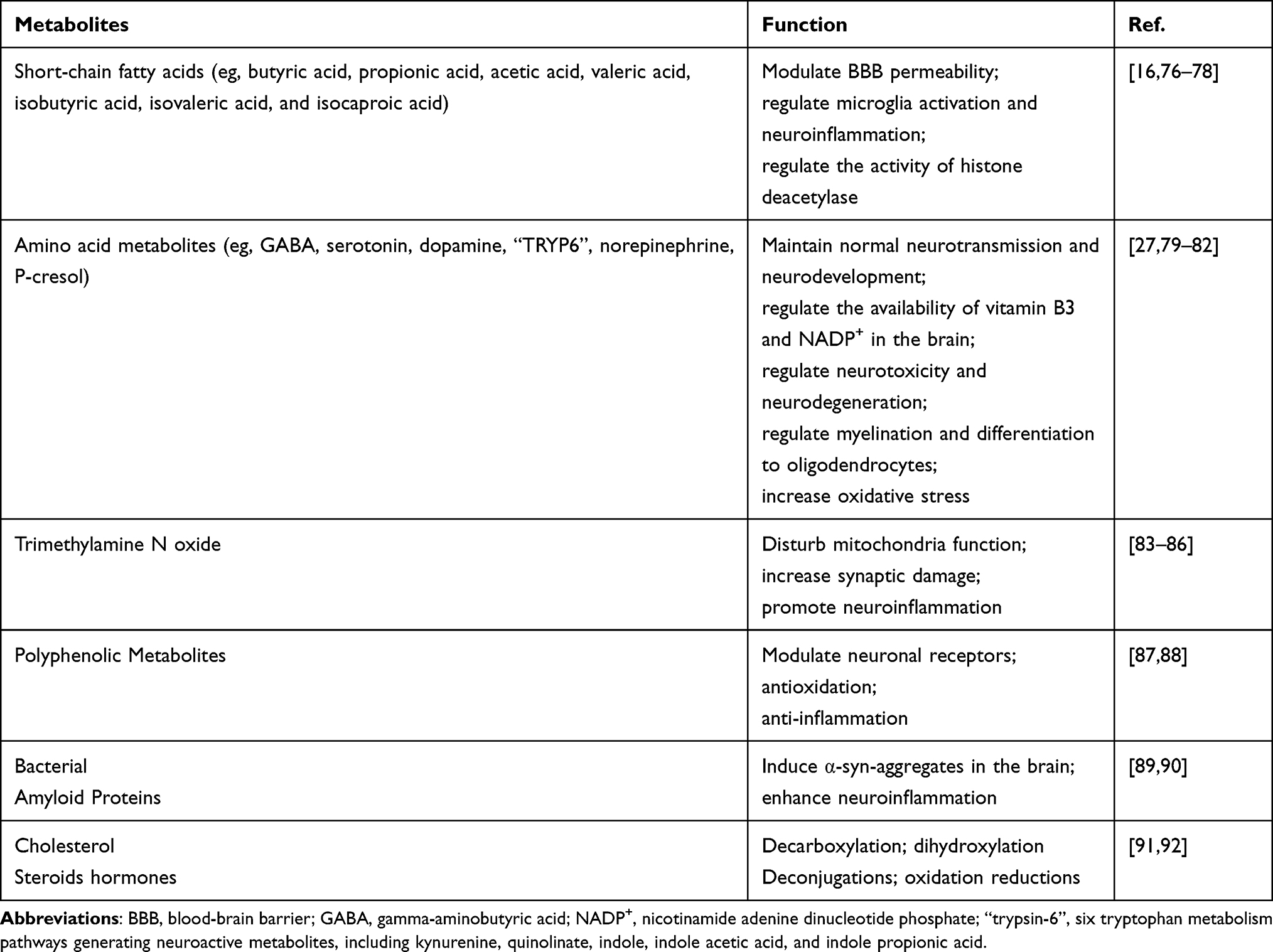 |
Table 2 Roles of Gut Microbial Metabolites on Brain Development |
Peripheral Nerve Communication Pathways
The gut’s peripheral neural network comprises autonomic components (sympathetic and parasympathetic/vagal pathways) and the intrinsic enteric nervous system (ENS).6,93 The centers of the ENS are located around the digestive tract (Figure 3). These low-level nerve centers regulate most of the physiological activities of the gut with a high degree of autonomy.93 Their regulation is automated and mechanized, and occurred by “stress” and “emergency” responses.94 The peripheral nerves of the gut cannot directly sense or regulate GM. However, the metabolites of GM contain neurotransmitters or neuromodulators, which act as important neural signaling molecules, and have effects on both the peripheral gut nerves and the brain6 (Table 3).
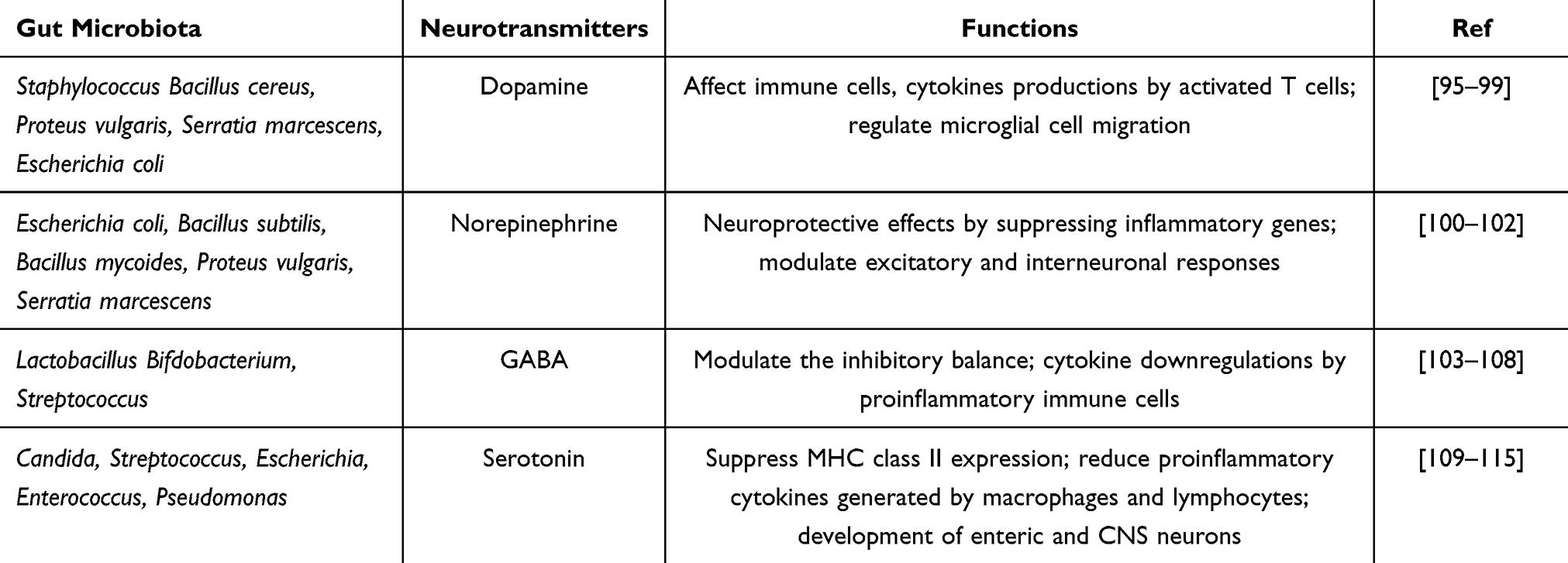 |
Table 3 | Gut Microbiota-Derived Neurotransmitters and Their Potential Functions in Brain Development |
 |
Figure 3 The distribution of the enteric neuronal network. The submucosal plexus, on the other hand, is located in the submucosal layer, where extrinsic sympathetic and parasympathetic fibers enter the entire layer of the intestinal wall and form a complex neural network with intrinsic enteric nerves and EGCs. The enteric neural network regulates enterocytes through neurotransmitters, neurotrophic factors and receptors. |
GM metabolize tyrosine, tryptophan, and glutamate to produce a variety of important neurotransmitters, including dopamine, norepinephrine, 5-hydroxytryptamine (5-HT), and histamine.116,117 Of these, acetylcholine and norepinephrine are the most important autonomic neurotransmitters.118 Acetylcholine acts on nicotinic receptors in the cerebral cortex after being released from nerve endings.116 The metabolism of tyrosine by GM produces precursors of 5-HT, which affects mental activity in the brain.119,120 In addition, GM metabolites that enter the circulation can affect the blood brain barrier (BBB). Decreased levels of short-chain fatty acid (SCFA) may disrupt the integrity of the BBB.121,122 Dietary supplementation with SCFA or SCFA-producing probiotic strains reverse the damage to the BBB, and maintain its integrity.121–124
The dense neural network of the gut is unable to directly contact the GM, but is able to sense some of the GM metabolite changes.125 The vagus nerve (VN), which distributes large number of intestinal regulatory peptides and intestinal metabolite receptors, is the main pathway by which the GM influences the nervous system.126,127 The VN is capable of transmitting information about gastrointestinal tract function and GM metabolites to the central autonomic network of the nervous system and brain through the intestinal visceral sensory afferent nerves.125,128 After integrating the afferent information via the central nervous system (CNS), it affects the function of relevant target organs via efferent nerves.128,129 In addition, intestinal VN can inhibit peri-intestinal inflammation and decrease intestinal permeability, and reduce the damage of GM and metabolites to the intestinal epithelial barrier.124,130 Under stress, VN is inhibited and has deleterious effects on the intestinal epithelial barrier and GM, generating an excessive immune response to GM.130 SCFA produced by GM metabolism acts as an informative molecule recognizing by VN receptors and having an activating effect on the VN.126 Toll-like receptor 4 (TLR4) expressed on VN afferent fibers is also capable of recognizing GM-derived LPS, which influence brain function by the ENS or VN modulates intestinal permeability.49,129
Immune Communication Pathways
GM carry large number of antigenic components that can be recognized by the receptors of intestinal immune cells.131,132 Under physiological conditions, it can induce normal immune response and promote the establishment of intestinal immune function.8 Under pathological condition, it can induce excessive immunity of intestinal mucosa and become an important pathogenesis of IBD.8,117,133 Cytokines produced by activated immune cells can pass through the BBB and enter the brain, thus affecting the development and physiological activities of the host brain.134 Activated intestinal immune cells can also enter the brain through the circulating network, affecting the immune function of microglia and astrocytes.117 In addition, intestinal colonized GM can induce host immune tolerance and work with host immune cells to defend against exogenous pathogens through colonization resistance.8,132,134
Endocrine Communication Pathways
The intestine is the largest endocrine organ in the human body, and enteroendocrine cells (EEC) constitute 1% of intestinal epithelial cells.135,136 The endocrine cells of the intestinal mucosa gather abundant receptors on their surface, which can receive signaling molecules from various sources such as GM metabolites and cytokines, and regulate the hormone secretion of endocrine cells in a feedback manner.131,136 Through the change of hormone level, it affects the function of target organs.131 In addition, some GM metabolites can act on extraintestinal endocrine cells through blood circulation to regulate the secretion of specific hormones.133 It has been found that GM metabolites can affect the secretion of 5-HT, substance P, cholecystokinin (CCK), gamma-aminobutyric acid (GABA) and other neuroactive mediators by enteroendocrine cells.116,117,119,120,137 Another study has found that colonic L cells are able to receive indole metabolites produced by the metabolism of intestinal flora, affecting the secretion of glucagon-like peptide-1 (GLP-1), then stimulating colonic VN activity.138
Integrated Communication Pathways
The regulatory systems of organs are complex and intertwined, and the communication pathway of MGBA may involve multiple regulatory systems such as neural, immune, and endocrine systems, forming a comprehensive communication pathway.69,73,139,140 The hypothalamic–pituitary–adrenal (HPA) axis involves the nervous and endocrine systems, serving as a key integrative pathway within the MGBA.131,141 Firstly, many hormones secreted by the HPA axis can directly regulate intestinal physiological functions and influence the composition of GM. Corticotropin releasing hormone (CRH) and glucocorticoids can also increase intestinal epithelial permeability and cause intestinal epithelial barrier dysfunction.140,142 The dysfunction of the intestinal epithelial barrier can change the composition of GM and affect the balance of intestinal microecology.131,143 Secondly, multiple metabolites of GM can activate the HPA axis, which in turn affects intestinal and brain function. SCFAs produced by gut bacteria can reduce the expression of genes encoding proteins involved in the HPA axis and can also activate the HPA axis by releasing lipopolysaccharides and peptidoglycan.144 Escherichia coli (E. coli) can produce Clp-β proteins that mimic α-melanocortin (α-MSH) to stimulate the release of Melanocortin precursor, which promotes the synthesis of Adrenocorticotropic hormone (ATCH).145–147 The metabolites of GM can influence the secretion of neuroactive mediators by enteroendocrine cells, including 5-HT, substance P, growth inhibitor, CCK, GABA, adenosine triphosphate (ATP), leptin, and orexin.141,142 These mediators can influence peri-intestinal nerve and brain function, exhibiting a cascading effect of GM on endocrine cells and neurons.141,144
Study on the Correlation Between GM and IBD
MGBA plays an important role in the pathogenesis and disease activity of IBD. GM disorders are the most important exogenous cause of IBD, and IBD patients are almost always accompanied by GM changes2,132,148–150 (Figure 4).
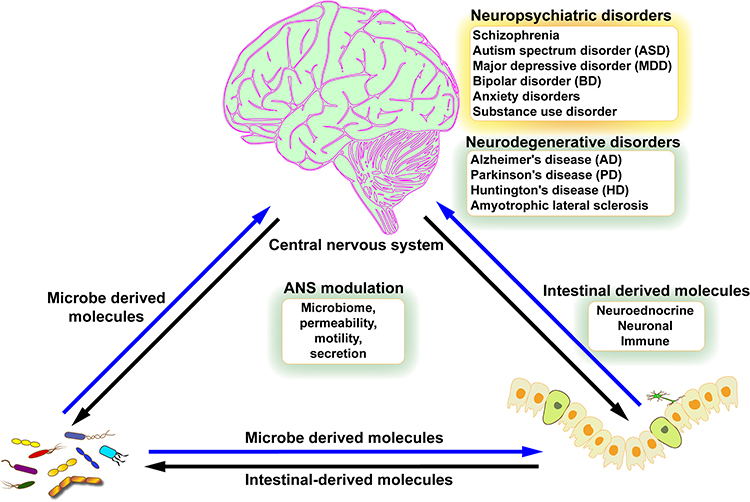 |
Figure 4 Systems biology model of brain-gut-gut microbiota interactions. The gut microbiota communicates with the gut connectome via microbial metabolites, and changes in gut function can affect the gut microbiota. In the presence of disturbances such as psychosocial stress, alterations in the gain of these two-way interactions can change the stability and behavior of the system, manifesting as brain-gut disturbances. |
Studies on the Effect of GM Disorders on IBD
GM disorders affect intestinal peristalsis, disrupt the intestinal barrier, and lead to recurrent intestinal infections and abnormal intestinal immunity, which plays an important role in the pathogenesis and disease activity of IBD.151 GM carries antigenic components that may cause abnormal intestinal immunity.151 GM metabolites can also be involved in the pathogenesis of IBD through neurological, immunological, endocrine, and other regulatory pathways causing intestinal dysfunction and disruption of the intestinal barrier.152 It has been found that butyrate is a SCFA produced by intestinal microbial metabolism, which provides energy to promote the production of mucin and adenosine monophosphate of colonic cells, and shape the mucosal immune system by regulating the differentiation of relevant T cells.152,153 Thibault et al found that the number of some butyrate-producing bacteria and the amount of butyrate were reduced in Ulcerative colitis (UC) patients, reflecting the effect of GM metabolites on IBD.153,154 Therefore, regulating GM disorders has become an important target for IBD treatment.155,156 Probiotic supplementation and fecal microbiota transplantation (FMT) are commonly used in the treatment of clinical IBD, and their safety and efficacy have been recognized.156–158 Indirectly, this reflects the important role of GM disorders in IBD pathogenesis and disease activity.2
Study of GM Changes in Patients with IBD
It has been found that IBD patients suffer from intestinal microecological imbalance accompanied by a decrease in the diversity and abundance of the bacterial flora.151 For instance, there was a decrease in the number of intestinal commensal bacteria, such as Bacteroides and Firmicutes, and an increase in the number of harmful bacteria, such as Aspergillus phylum and Actinomycetes phylum.152 Common causative organisms of intestinal infections present in patients with IBD include Clostridium difficile (C. difficile), Campylobacter spp., adherent-invasive E. coli, and Salmonella spp.153 There are also studies on the intestinal flora during the active phase of IBD that found a significant increase in the number of bacteria such as Enterobacteriaceae, Enterococci, yeasts, Bacteroides, and Peptococcus, and a significant decrease in the number of bacteria such as Lactobacillus, Fusobacterium, and Bifidobacterium in the colon, as compared to the remission phase.153,154 In addition, the intestinal fungal community of IBD patients had an altered compositional structure with increased diversity and abundance as well as elevated intestinal mucosal colonization density compared to the healthy population.155,156 The intestinal viral community of IBD patients had an increased diversity of viruses as well as an increased chance of intestinal viral infections compared to the healthy population.2 Whether changes in the intestinal fungal and viral communities are associated with the pathogenesis of IBD remains to be studied in depth.156–158
Study on the Correlation Between GM and Neuropsychiatric Disorders
Depression, anxiety, ASD, SCZ, PD, AD and other neuropsychiatric disorders are affected by MGBA.7,68 GM can interfere with the development of the brain, and affect the physiological functions of the brain.4 There is a distant distance between GM and the brain, and the metabolites of GM and the brain also have intestinal barriers, blood-brain barriers, needing to use the neural and circulatory network in order to reach the brain.141,142 The main mutual communication between GM and the brain pathway is the peri-intestinal nerves, which combine with the immune system or endocrine system to establish an integrated communication pathway between GM and the brain.159,160 For example, in the HPA axis, GM-derived signaling molecules influence brain function through endocrine organs and central nervous system transit151 (Table 4).
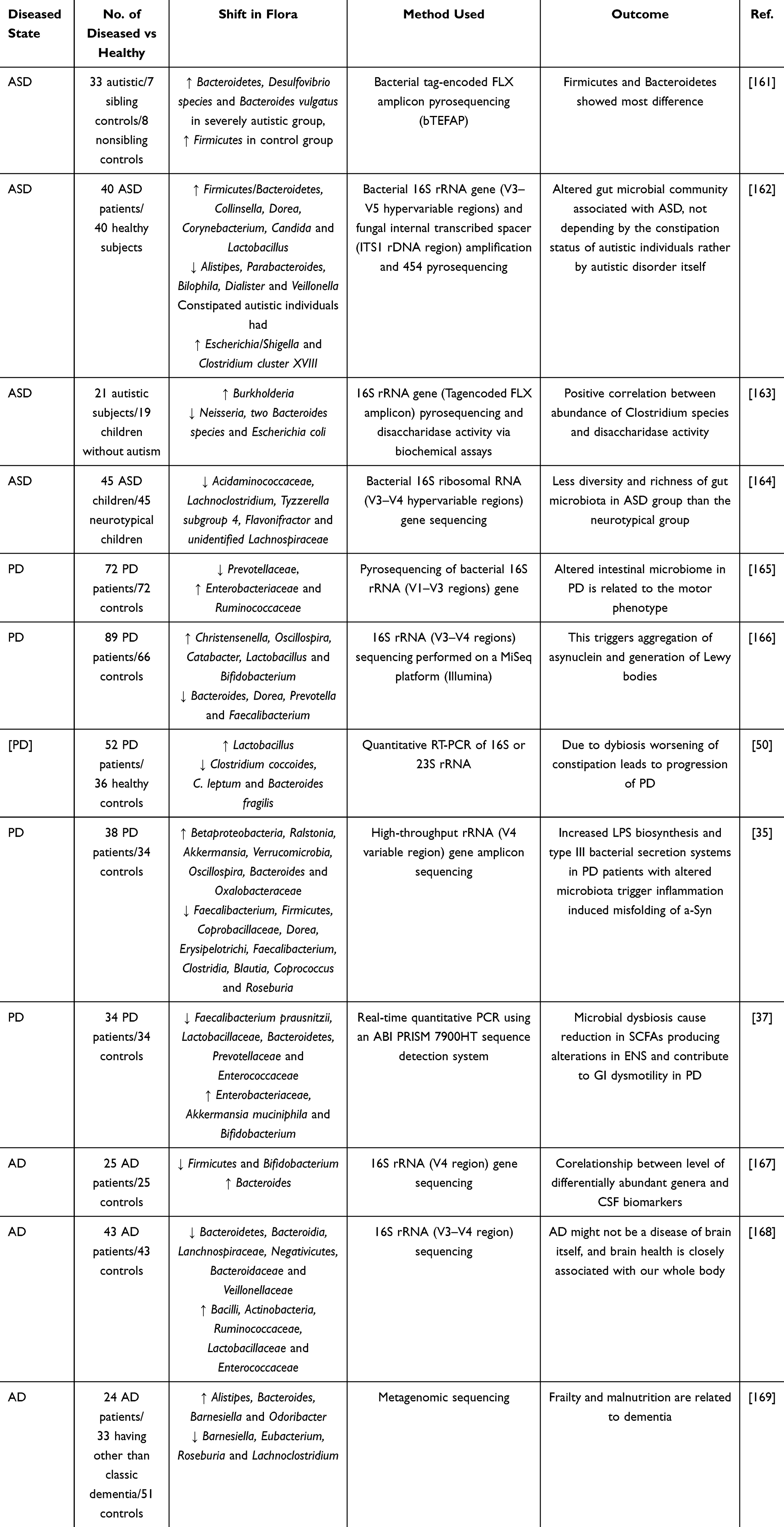 |
Table 4 Study on the Correlation Between Gut Microbiota (GM) and Neuropsychiatric Disorders |
GM and Depression or Anxiety
Emotion is a higher function of the brain with relatively independent functional areas.170 It is generally accepted that the emotional region of the cerebral cortex is in the prefrontal lobe, and subcortical emotional centers include the hypothalamus, limbic system, and brainstem network structures.170,171 Depression or anxiety both manifest as emotional disorders and they often occur together.172,173 The main features of depression are depressed mood and loss of interest, while anxiety disorders are characterized by mood anxiety and panic attacks.173 They can be accompanied by emotional experiences such as guilt, despair, worry, and irritability.173,174 The pathogenesis of depression and anxiety is unclear and influenced by individual health and illness, as well as by external factors, such as social and environmental factors.174 GM also plays an important role in the development of host affective disorders.145
It has been found that germ-free mice attenuate stress responses in the HPA axis and less depressive-like behaviors than normal mice.140,141 Implantation of fecal bacteria from depressed patients increased the incidence of depressive symptoms in mice compared to implantation of normal human fecal bacteria.57 Aizawa et al found that the intestinal probiotic levels of Bifidobacterium and Lactobacilli were significantly lower in patients with major depression than in healthy individuals.175,176 Patients with major depression had an increased diversity of fecal flora, with an increase in Aspergillus and Actinobacteria and a decrease in the abundance of the thick-walled phylum, such as Bacteroides.176 Patients had a higher abundance of pro-inflammatory bacteria in the gut, such as Enterobacter, whereas SCFA-producing bacteria were reduced in abundance, such as Fecal bacilli and fecal cocci.176–178 These metabolite changes may lead to intestinal immune dysfunction.179
GM and GM metabolites affect the emotional function areas of the brain through neural, immune, and endocrine pathways, resulting in affective disorders such as depression or anxiety.174,180 Human mood is regulated by important neurotransmitters such as dopamine and 5-HT, while GM has an important influence on the biosynthesis of dopamine and 5-HT.119,120 GM disorders affect the synthesis of dopamine and 5-HT, which has an impact on the regulation of mood and makes it easy to develop symptoms of depression or anxiety.116,119 It was found that protein or messenger ribonucleic acid (mRNA) expression of brain-derived neurotrophic factor was reduced in a mouse model of intestinal inflammation, and anxiety-like behavior was increased in mice.120,181 Their anxiety behaviors were normalized after probiotic treatment, which suggests that the state of intestinal inflammation is associated with anxiety-like behavior, and can be improved by regulating GM disorders.181 It has been shown that GM dysbiosis is an important trigger of intestinal inflammation, and the resulting increase in pro-inflammatory cytokines promotes the risk of depression or anxiety symptoms in humans.182–184 Another experiment found that pretreatment with probiotics reduced c-fos proto-oncogene protein (C-FOS) expression and increased brain-derived neurotrophic factor (BDNF) expression in different regions of the brain in rats under stress, as a result, the abnormal proliferation and morphological changes of neurons in the hippocampus could be avoided.182,184
GM and ASD
ASD is a group of neurodevelopmental disorders. The main manifestations are different degrees of behavioral stereotypes, language developmental disorders and social disorders.185,186 The etiology of ASD is not known, and mainly related to genetic variations, neurodevelopmental disorders, immune dysfunction and other factors.187 Studies have shown that GM may affect the onset and course of ASD, and intervening in GM disorders can improve the clinical symptoms of ASD, making it a potential target for ASD treatment.188,189
It was found that reduced social motivation and interest in novelty can be improved in germ free (GF) mice after colonization with gut bacteria.190 Suggesting a role for GM in regulating social interactions and interest.188 Additionally, the GM composition and metabolites were significantly altered in children with ASD, mainly by a decrease in the number of Bacteroides and a decrease in the ratio of Bacteroides to thick-walled bacteria, and an increase in the number of Lactobacillus, Clostridium, Desulfovibrio and Enterobacter.188,191–193 Bifidobacterium are among the earliest probiotics to colonize the neonatal gut, and their reduced abundance may affect neurodevelopment and contribute to the pathogenesis of ASD.194 Plasma metabolites (including nicotinamide ribose, Inosine 5’-Monophosphate (IMP), iminodiacetate, methyl succinate, galacturonate, valerylglycine, sarcosine, and leucylglycine) levels were significantly reduced in a study that accurately assessed plasma metabolites in children with ASD.194 The study found that C. difficile, through its metabolite propionic acid, can trigger ASD in rats.191,195 Treatment of Clostridium intestinalis infections with antibiotics, such as vancomycin, significantly improved psychiatric symptoms in children with ASD.194,196 Treatment of refractory C. difficile infections with fecal transplants has been shown to improve psychiatric symptoms in children with ASD.197,198 A study in a mouse model of ASD showed that treatment with C. difficile improved small intestinal osmolarity, altered the composition of the GM, and alleviated ASD-related symptoms.195,199 Another study showed that mice transplanted with fecal flora from ASD patients showed changes in blood and fecal GM metabolites, and exhibited ASD-like behavioral deficits.190 The ASD-like behavioral defects in some mice can be ameliorated by supplementation with probiotics or metabolites.190,200
GM and SCZ
The main characteristic of SCZ is that the individual’s way of perceiving reality is severely impaired and he or she experiences persistent psychological and behavioral difficulties.201,202 Clinical manifestations include persistent delusions and hallucinations, disorganized thinking and behavior, which may be accompanied by anxiety, depression, social withdrawal, aggressive violence, and suicidality.203,204 The etiology and pathogenesis of SCZ have not been elucidated. It is currently believed that SCZ is related to the individual’s genetics, brain development and structure, psychological cognition, and behavioral stress, as well as social, environmental, and other external factors.203,204 Many studies suggest that GM may influence the onset and course of SCZ.201,205
The most frequently reported changes in GM composition in SCZ patients were Clostridium, Enterococcus, Prevotella, and especially increased abundance of Lactobacillus spp.4,206,207 The intestinal flora of SCZ patients contained a wide range of parthenogenetic anaerobes, such as Lactobacillus fermentum, Enterococcus faecalis, and alkalophilic bacillus, which are rare in the intestinal tracts of healthy individuals.208,209 Through macro-genomic sequencing and 16s ribosomal RNA (rRNA) sequencing, it has been found that the GM composition of SCZ patients shows higher α-diversity at the genus level and higher β-diversity at the species level compared to controls.200,210 It has also found that glutamate synthase (GOGAT) is more active in the intestines of SCZ patients than that in healthy controls, which correlated with GM-associated intestinal IgA levels.208,211,212
Studies have found that GF mice are less likely to develop SCZ symptoms than normal mice.208 SCZ-related behaviors can be induced in GF mice by transplantation of SCZ patient feces, accompanied by altered levels of glutamate, glutamine, and GABA in the hippocampus, but could not be identified to be associated with a specific intestinal bacterial strain.213 Some studies have found that microbial infections during fetal life may increase the risk of developing SCZ in children, but have not delved into which specific pathogenic microbial infections.214,215 Animal studies have found that rodents with an altered gut core microbiome after treatment with sub-chronic phencyclidine (sub-PCP) can produce SCZ-like symptoms.216
GM and AD
AD is a neurodegenerative disease that occurs in the brain and manifests itself as a combined impairment of cognitive content and skills, accompanied with a decline in behavioral abilities.217,218 The etiology of AD is complex and unknown, with more than 30 possible influencing factors and hypotheses.219 In terms of pathogenesis, the more accepted hypotheses include β-amyloid (Ap) deposition, tau protein hyperphosphorylation, and abnormal glucose metabolism in the brain caused by insulin resistance.220 Many studies have shown that GM play an important role in various hypotheses of AD pathogenesis.221–223
Animal studies have found that GF mice have a lower incidence of AD compared to conventionally raised mice from the same background.222 Amyloid plaques and neuroinflammatory manifestations were not seen in the brain tissue of GF mice, which showed GM played an important role in the pathogenesis of AD.224,225 Clinical studies have found differences in the GM composition of AD patients compared to the healthy population. The richness and diversity of GM was reduced in AD patients compared to the healthy population, with a decrease in the Firmicutes, Bifidobacterium, and an increase in the Bacteroides.226–228 One controlled study found a significant increase in the abundance of Escherichia and Shigella, which was involved in the inflammatory response, in fecal samples from AD patients.168 GM alterations were found to be closely associated with pathological deposition of Ap and hyperphosphorylation of tau proteins.229,230 GM metabolites, such as short-chain fatty acids, can interfere with Ap deposition and help prevent AD.76,123
GM and PD
PD is also a neurodegenerative disease that occurs in the brain, manifested by impairment of the brain’s motor regulatory function and movement disorders, such as resting tremor and bradykinesia, which may be accompanied by non-motor symptoms including sensory disturbances, psychosomatic disorders, and gastrointestinal dysfunction.231,232 The main pathological feature of PD is the deposition of α-synuclein (α-syn) and aggregation of Lewy bodies in dopaminergic neurons in the dense nigral region of the midbrain, contributing to neuronal degeneration and death as well as significant reduction of dopamine content in the striatum.232,233 The exact etiology of the pathological changes associating with PD is unclear, and genetic factors, environmental factors, ageing, and oxidative stress may be involved.234 Many studies have shown that GM influences the pathologic process of PD through metabolites.70,235
Several studies have found significant alterations in GM and metabolites in PD mouse models and PD patients compared to healthy controls.235,236 This was mainly characterized by increased abundance of lactobacillus and decreased abundance of Prevotella and Puccinia spp.235,237 Studies of GM composition in PD patients have revealed a decrease in SCFA-producing intestinal strains such as Prevotellaceae, E. faecalis, and Gastric cocci, a decrease in intestinal strains with anti-inflammatory effects such as Coprococcus, and E. faecalis, and an increase in bacteria involved in lipid metabolism and in increasing intestinal permeability.236 It has also been observed that changes in GM metabolites such as SCFA and lipopolysaccharide (LPS) may influence the onset or development of PD.34,238 Some studies have found that the deposition of α-synuclein in the ENS cause by abnormal intestinal barrier function and GM disorders may be correlated with the pathogenesis of PD.215,239 It has been pointed out that bacterial neurotoxins, such as LPS, enter the bloodstream through the disrupted intestinal epithelial barrier, and produce inflammatory cytokines by immune cells through nuclear factor kappa-B (NF-κB) and TLR4, leading to systemic inflammatory responses.49,240 Moreover, bacterial LPS and inflammatory cytokines (including TNF-α, IL-1β, and IL-6) can induce BBB destruction, causing α-syn accumulation in dopaminergic neurons in the substantia nigra.241,242
Correlation Studies of IBD and Neuropsychiatric Disorders
IBD is often accompanied by neuropsychiatric disorders, and MGBA may be a link by which they establish a connection.243,244 This connection is mainly manifested in comorbidities and it is difficult to clarify that there is a causal relationship between them.132,149,245 The gut is distant from the brain and various physiologic barriers exist.246 However, pathological changes such as inflammatory response, barrier disruption, and intestinal microecological disorders occurring in IBD can send signals to the brain through the peripheral nerves, the immune system, or the endocrine system, affecting the brain’s regulatory or higher neural functions.246,247 In turn, the brain affects the structure and function of the gut by regulating the nervous system, immune system or endocrine system, which is an important pathogenesis of IBD.245,248
IBD with Anxiety or Depression
Studies have shown that there is a significant increase in the prevalence of depression and anxiety in people with IBD compared to the general population, especially during the active phase of IBD.150,249 The correlation between IBD and anxiety or depression is significantly higher compared to other chronic diseases such as hypertension and diabetes.250 There is a high prevalence of symptoms of anxiety and depression in patients with IBD, with up to a third of patients affected by anxiety symptoms and a quarter affected by depression symptoms.251 Prevalence was also increased in patients with active disease: half of these patients met criteria for anxiety symptoms and a third met criteria for depression symptoms.250,252
GM is a target of MGBA and has become an important target for IBD treatment.239,253 Studies have found that treatment targeting GM can improve IBD symptoms.249,250 And with the improvement of IBD symptoms, patients’ depression or anxiety symptoms also improved significantly. Some studies have reported that medication for IBD also has a palliative effect on anxiety or depression.250,254 It is not clear whether this palliative effect is associated with remission of IBD or if it is simply a medication effect.249 Anxiolytic or antidepressant therapy for people with IBD has also been shown to be helpful in the remission of IBD.249,255 Addressing psycho-spiritual disorders in patients with IBD can improve their compliance and satisfaction with treatment and enhance the outcome of IBD.249,255 Other studies have shown that antidepressant or anxiety medications can relieve IBD symptoms or reduce the risk of IBD recurrence and improve quality of life, but evidence that can be characterized is lacking.255,256
IBD with ASD
A meta-analysis showed that the prevalence of gastrointestinal dysfunction was significantly higher in children with ASD than in healthy children.194 Gastrointestinal symptoms in children with ASD may also be related to the severity of ASD.257,258 Some studies have shown that about 40% of ASD patients have experienced gastrointestinal disorders, and both psychiatric and gastrointestinal symptoms in ASD patients have been associated with GM disorders.259 One study found that children with ASD are often diagnosed with co-occurring disorders including IBD, suggesting a strong association between ASD and IBD.260,261 Another study found that interventions targeting GM improved IBD symptoms and was also able to alleviate psychiatric symptoms of ASD.262 And targeting ASD alone was observed to alleviate gastrointestinal symptoms as well as alter GM composition in patients with IBD.194,261
IBD with AD or PD
For patients with IBD, screening for AD or PD has found that IBD modestly increases the risk of developing AD or PD.149,263 Dietary factors and gastrointestinal dysfunction are correlated with the onset and progression of AD or PD.149 Patients with AD or PD are also frequently associated with gastrointestinal dysfunction, which may be an influential factor in the pathogenesis of IBD.149,264 Both IBD and AD or PD are associated with MGBA, a common pathway for their correlative linkage.149 Studies have shown that GM plays a key role not only in the pathogenesis of IBD, but also in the pathogenesis of AD or PD.265,266 Studies have shown that treatments targeting intestinal infections, microecological imbalances, and intestinal dysbiosis may also affect neurological symptoms of AD or PD.267 Early anti-inflammatory therapies for IBD may reduce the risk of developing AD or PD.72 Intervening in GM disorders in patients with IBD may be an early preventive measure to reduce the risk of developing AD/PD by altering intestinal permeability, modulating inflammatory responses, and reducing the aggregation of α-synuclein in the ENS.72,268
The Role of the GM in IBD and Neuropsychiatric Disorders
The clinical translation of MGBA focuses on GM applications in the diagnosis and treatment of IBD and neuropsychiatric disorders.11 In terms of diagnosis, current studies have identified abnormalities in GM composition and its metabolites of IBD and neuropsychiatric disorders.68,267 However, these abnormalities lack specificity and sensitivity, and are difficult to assist in the diagnosis of IBD and neuropsychiatric disorders.18,68 Therapeutically, intervening in GM disorders has achieved better results in IBD treatment, whereas therapeutic research in neuropsychiatric disorders is still in the beginning stage.11,155 Using GM as a therapeutic target, it will intervene in the GM through gut microecological modifiers, GM transplantation, dietary modification and drug selection, and explore the therapeutic effects of these interventions on IBD or neuropsychiatric disorders, respectively.11,72,157
Intestinal Microecological Regulators
Gut microecology is a dynamic community of GM and the environment in which they live. Foreign microorganisms are constantly introduced with food, and intestinal microorganisms are constantly excreted with feces.269–271 It is also influenced by the host’s genetics, immunity, age, diet, medication, and gastrointestinal dynamics in a state of dynamic change.272 The maintenance of a long-term and stable coexistence between the GM and the host is called intestinal microecological equilibrium.273 Dysbiosis refers to the disruption or transformation of this coexistence.272 Gut microecological regulators can correct the intestinal microecological imbalance to a certain extent and increase beneficial intestinal microorganisms.158,274,275 Gut microecological regulators include probiotics, prebiotics and synbiotics.272
Probiotics
The vast majority of gut microorganisms are bacteria.276 Intestinal flora can be categorized into beneficial, neutral and harmful bacteria according to their relationship with the host.276,277 Probiotics are selected from beneficial bacteria or strains of bacteria, cultured in vitro to make food or drugs, and after taking them, they can go through the test of gastric acid and bile to reach the intestinal colonization.278,279 At present, the probiotics used in human body mainly include Lactobacillus, Bifidobacterium, Enterococcus, Streptococcus, Bacillus, Clostridium, and Saccharomyces.277,280
Intestinal colonized probiotics form a microbial film on the surface of the intestinal mucosa have colonization resistance (CR) effect, such as inhibiting the growth and activity of pathogenic bacteria, enhancing mucosal immunity, and maintaining the intestinal mucosal barrier.278 Intestinal probiotics can exert anti-inflammatory effects and reduce gastrointestinal symptoms in IBD subjects through CR effects.281 Rembaeken et al randomized 83 patients with active UC after remission induced by standard treatment to mesalazine or E. coli Nissle, and found that there was no statistically significant difference between the two groups in the rate of UC recurrence.281,282 Lichtenstein et al found that VSL#3 enriched with eight probiotic species, including Saccharomyces boulardii, were useful in maintaining clinical remission in patients with Crohn’s disease (CD).283,284
Certain strains of enteric bacteria have also been shown to be effective in improving behavioral symptoms of neuropsychiatric disorders such as anxiety and depression, ASD, AD, and PD, and are referred to as “psychotropic probiotics”.285,286 “Psychotropic probiotics” are part of the probiotic family, which can obtain psychotropic benefits, such as anti-anxiety and anti-depression by influencing the host’s mood, cognition and other mental activities.286 Several preclinical studies and clinical reports on Bifidobacterium spp. and Lactobacillus spp. have found that supplementation with these two “psychotropic probiotics” improves mood, reduces anxiety, and enhances cognitive function, which can significantly improve CNS function and alleviate psychiatric symptoms.287,288 Lactobacillus spp. and Bifidobacterium spp. as probiotics can alleviate PD-related symptoms.289 Bacillus spp. as probiotics have the ability to convert L-tyrosine to L-DOPA, thus improving dopamine production and secretion, and relieving PD-related symptoms.290 In addition, probiotics are able to reduce the levels of pro-inflammatory factors in the body, such as IL-6, IL-1t3, TNF-a, and microglia activation markers, suggesting that they have the ability to reduce the overall level of inflammation.291 Targeting the GM with probiotics becomes a possible treatment for a variety of neuropsychiatric disorders.286
Prebiotics
Prebiotics are substances that are not digested by the gastrointestinal tract, but selectively become “food” for one or a few beneficial intestinal bacteria, stimulating the growth and activity of the beneficial bacteria.292,293 Prebiotics only selectively stimulate the growth of beneficial bacteria and do not promote the growth of harmful bacteria, and do not increase the digestive burden of the gastrointestinal tract and its nutritional needs.294 The most basic prebiotics are carbohydrates, which can be derived from food or formed when food is refined and processed.292 Currently, commonly used prebiotics include oligofructose, oligogalactose, oligosaccharides, and isomaltooligosaccharides.292 The combined preparation of probiotics and prebiotics is called symbiotic.295,296 It is a new generation of microecological regulator, which plays the physiological functions of probiotics and prebiotics at the same time, so that probiotics and prebiotics can coordinate their effects, fight against diseases together and maintain the microecological balance of the organism.297,298
A meta-analysis showed that prebiotics can significantly reduce the intestinal mucosal injury score and improve the index of intestinal inflammatory response in a rat model of IBD.296 DrabliIlska et al found that inulin, which is rich in oligofructose, can improve the metabolism of amino acids in the plasma and urine of children with CD, and maintain intestinal permeability, thus mitigate the gastrointestinal symptoms of children with CD.299 It has been found that prebiotics containing galactose and oligofructose exerts antidepressant and anxiolytic effects, which can reverse depression or anxiety symptoms triggered by chronic stress.300 Prebiotics with N-acetylcysteine administered for 8 weeks can reduce irritability and repetitive behaviors in infants with ASD compared to controls.301 Another study has also shown that lactulose-containing prebiotics can ameliorate cognitive deficits through autophagic and anti-inflammatory pathways in AD mouse models.302
FMT
FMT involves transferring a healthy human fecal sample carrying intestinal flora and metabolites into a patient’s intestine to re-establish normal functioning intestinal flora in the treatment of intestinal and extra-intestinal diseases.303 Fecal samples can be infused directly into the patient’s colon via colonoscopy, enema, or made into lyophilized material for oral feeding.304,305 FMT ensures the integrity and bioactivity of the intestinal flora, and the quantity and quality of bacteria reaching the intestine.306 FMT makes it easier to establish the structure of the healthy intestinal flora or model the intestinal flora of a specific disease.305 Clinically, FMT has been shown to be effective in the treatment of recurrent and refractory C. difficile infections (CDI).304,305,307,308 FMT application in IBD treatment has also resulted in significant clinical remission.305 There is also a large body of research exploring the FMT use in the treatment of other intestinal and non-intestinal related diseases. Study has shown that FMT can improve the diversity of the GM and alleviate gastrointestinal symptoms.306 Patients with a variety of neuropsychiatric disorders, including anxiety and depression, ASD, and PD, also benefit from FMT treatment, with no serious adverse events occurring during treatment or during follow-up.309
One study conducted in vitro culture of GM transplantation in ASD mice, which resulted in a significant reduction of anxiety-like behavior in mice.310 FMT treatment in AD mice by enema can change the composition of the GM in mice, and thus effectively alleviated the cognitive dysfunction of AD, which indirectly showed that changes in the composition of the GM relating to cognitive dysfunction.311 In another study, FMT was performed by colonoscopy on ASD patients, and the GM of ASD patients was transformed to a healthy state, and the ASD-related symptoms of patients were significantly improved.195 However, more relevant studies are still needed to demonstrate the effect of FMT on ASD patients.312
Dietary Modification
Diet is the main source of nutrients for the GM and a pathway for invasion by foreign microorganisms.313 Different dietary structures carry different bacterial species and nutritional elements that have a direct impact on GM. Studies have found that gut microecology can be altered by dietary modifications.313,314 Westernized diets and ketogenic diets lack carbohydrates available to gut microbes, leading to a decrease in gut microecological diversity.315–317 Vegetarian diets and Mediterranean diets with adequate dietary fiber intake are effective in improving the diversity and stability of gut microecology.318
The aim of regulating intestinal microecological imbalance and combating MGBA-associated diseases is achieved through the development of a rational and individualized dietary regimen that alters intestinal microbial composition and metabolites.314,315,319 It has been found that a vegetarian diet or the addition of SCFA to the diet will increase the level of Unified Parkinson’s Disease Rating Scale (UDPRS III) and reduce the effective daily dose of levodopa equivalents, a potentially effective non-pharmacological treatment for PD.320 It has been shown that breastfeeding for 6 months reduces the chances of ASD manifestation, while formula feeding is associated with increased C. difficile abundance in the gut.321
Drug Use
Chemical drugs can also affect GM parasitized in the human body, causing intestinal microecological imbalance and associated diseases.322 Conversely, GM also affects host metabolism and absorption of chemical drugs.323,324 The interactions between chemical drugs and gut microbes need to be taken into account when using chemical drugs, such as antibiotics, immunomodulators, anticancer and antipsychotic drugs.325 In a large-scale, in vitro screening study of more than 1000 drugs against 40 representatives of gut commensals, it was found that 24% of the drugs tested exhibited growth inhibition against at least one bacterial strain.326
Antibiotics can directly affect the gut microflora, potentially eliminating pathogenic bacteria as well as depleting probiotics.323 For intestinal infections, the pros and cons need to be weighed and assessed individually to rationalize the choice of antibiotics in the clinic.322 Both pathogenic and probiotic bacteria should be eliminated to avoid intestinal microecological imbalance. Rifaximin is an orally non-absorbable antibiotic.327 Prantera et al showed that rifaximin induced remission of moderately active CD with low side effects.328 Rifaximin achieves its therapeutic effect in IBD mainly through local anti-infective and repair of intestinal epithelial barrier damage in the gastrointestinal tract, but large-scale clinical trials are still needed to confirm this.329,330
A significant negative correlation was found between some antipsychotic drugs and microbial diversity, especially antidepressants and antipsychotics had the highest correlation.331,332 Selective 5-hydroxytryptamine reuptake inhibitors (SSRIs), such as fluoxetine, sertraline, and citalopram, have shown some antimicrobial activity against strains of Bacillus, Clostridium, Enterococcus, Pseudomonas, and Staphylococcus.326,333 It has been found that the tricyclic antidepressant amitriptyline has antimicrobial activity against intestinal pathogenic bacteria, such as Bacillus spp., Staphylococcus spp. and Vibrio cholerae.334 Promethazine exerts growth inhibition against Yersinia pestis and E. coli, which can cause intestinal infections.335
Conclusions and Perspectives
IBD and neuropsychiatric disorders exhibit strong associations with the MGBA, both characterized by distinct GM alterations.72 GM dysbiosis is recognized as a key contributor to IBD pathogenesis and is implicated in the neurobiological pathways underlying psychiatric disorders.11 Current interventions targeting gut dysbiosis-including probiotics, FMT, and dietary modulation—demonstrate moderate efficacy in IBD (clinical remission rates 40–60%) and emerging potential in neuropsychiatric disorders (eg, 30% reduction in depression scores).336 A clear causal relationship between IBD and neuropsychiatric disorders is difficult to establish based on current research. However, IBD and neuropsychiatric disorders have a significantly higher chance of co-occurring with each other, increasing each other’s incidence and prevalence to varying degrees.268 They have also demonstrated a reciprocal and mutually reinforcing effect in treatment, mainly in terms of interventions on intestinal microecology.268 In terms of clinical translation, it is still difficult to assist in the diagnosis of IBD and neuropsychiatric diseases due to the technical limitations of gut microbiological testing and the lack of gut microecological alterations with high specificity and sensitivity. In terms of therapeutic transformation, intestinal microecological regulators are abundant and diverse, including probiotics, prebiotics, synbiotics, and postbiotics, which are able to regulate GM composition and metabolites more accurately. The technologies of donor screening, bacterial fluid preparation, and transplantation pathway for FMT have matured, and are gradually expanding the scope of clinical application and standardizing the criteria for clinical application. Many studies have shown that GM has emerged as a therapeutic target for IBD and a potential therapeutic target for neuropsychiatric disorders.
GM in the host intestine is large in number and diverse, and affected by multiple factors such as host diet, medication, and health, disease. There are still great challenges in GM detection techniques and research methods. Macroscopic correlation studies have failed to adequately take into account the complex changes and interfering factors of GM, and homogenization and standardization are difficult to achieve. Evidence for the existence of correlation between GM and neuropsychiatric disorders is still insufficient. Microcosmic correlation studies are still scarce, and more studies at the molecular level are needed to explore the extent and mechanisms of correlation. Moreover, while therapeutic of the GM holds transformative potential, critical challenges impede clinical translation. Standardization hurdles, prebiotics lack dose-response consensus and FMT protocols vary dangerously in donor screening, processing, and delivery routes. Patient heterogeneity introduces further complexity—baseline microbiome architecture of FMT responsiveness, while host genetics reduce probiotic engraftment. Long-term efficacy remains precarious due to ecological succession dynamics. In the future, it is necessary to continuously improve the detection technology of GM, conduct extensive experimental comparative studies, and continuously explore the pathways and effect mechanisms of MGBA. It will be more reliable to verify the correlation between GM and specific diseases and promote the clinical translation of MGBA in IBD and neuropsychiatric diseases.
Abbreviations
5-HT, 5-hydroxytryptamine; AD, Alzheimer’s disease; Ap, β-amyloid; ASD, autism spectrum disorder; ATP, adenosine triphosphate; BBB, Blood Brain Barrier; BDNF, Brain-derived neurotrophic factor; CCK, cholecystokinin; CD, Crohn’s disease; C. difficile, Clostridium difficile; CDI, Clostridium difficile infections; C-FOS, c-fos proto-oncogene protein; CNS, central nervous system; CR, colonization resistance; CRH, corticotropin releasing hormone; EEC, enteroendocrine cells; ENS, enteric nervous system; E. coli, Escherichia coli; FMT, fecal microbiota transplantation; GABA, gamma-aminobutyric acid; GBA, Gut–brain axis; GF, germ free; GLP-1, glucagon-like peptide-1; GM, gut microbiota; GOGAT, Glutamate synthase; HPA, hypothalamic–pituitary–adrenal axis; IMP, Inosine 5’-Monophosphate; IBD, Inflammatory bowel disease; IL-1β, interleukin-1β; IL-1t3 interleukin-1t3; IL-6, interleukin-6; L-DOPA, 3,4-Dihydroxyphenyl L-Alanine; LPS, lipopolysaccharide; mRNA, messenger ribonucleic acid; MGBA, microbiome–gut–brain; NF-κB, nuclearfactorkappa-B; PD, Parkinson’s disease; rRNA, ribosomal RNA; SCFA, short-chain fatty acid; SCZ, schizophrenia; SSRIs, Selective 5-hydroxytryptamine reuptake inhibitors; subPCP, subchronic phencyclidine; TLR4, Toll-like receptor 4; TNF-α, tumor necrosis factor-α; UC, Ulcerative colitis; UDPRS III, Unified Parkinson’s Disease Rating Scale; VN, vagus nerve; α-MSH, mimic alpha melanopsin; α-syn, α-synuclein; Ap, β-amyloid.
Acknowledgments
This work was supported by Science and Technology Innovation Committee of Shenzhen (JCYJ20210324113802006, JCYJ2022053015180024 and JCYJ20210324113613035). Science and Technology Innovation Committee of Shenzhen (KCXFZ2021102016454300), and the Shenzhen Key Medical Discipline Construction Fund (No. SZXK042), Sanming Project of Medicine in Shenzhen (No. SZSM202311025). Guangzhou Health Science and Technology Project (20241A011008).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas, took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published, have agreed on the journal to which the article has been submitted, and agree to be accountable for all aspects of the work.
Disclosure
No conflicts of interest, financial or otherwise, are declared by the authors.
References
1. Ramos GP, Papadakis KA. Mechanisms of disease: inflammatory bowel diseases. Mayo Clin Proc. 2019;94(1):155–165. doi:10.1016/j.mayocp.2018.09.013
2. Nishida A, Inoue R, Inatomi O, Bamba S, Naito Y, Andoh A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin J Gastroenterol. 2018;11(1):1–10. doi:10.1007/s12328-017-0813-5
3. Muhammad F, Fan B, Wang R, et al. The molecular gut-brain axis in early brain development. Int J Mol Sci. 2022;23(23):15389. doi:10.3390/ijms232315389
4. Warner BB. The contribution of the gut microbiome to neurodevelopment and neuropsychiatric disorders. Pediatr Res. 2019;85(2):216–224. doi:10.1038/s41390-018-0191-9
5. Chandra S, Alam MT, Dey J, et al. Healthy gut, healthy brain: the gut microbiome in neurodegenerative disorders. Curr Top Med Chem. 2020;20(13):1142–1153. doi:10.2174/1568026620666200413091101
6. Furness JB. The enteric nervous system and neurogastroenterology. Nat Rev Gastroenterol Hepatol. 2012;9(5):286–294. doi:10.1038/nrgastro.2012.32
7. Sorboni SG, Moghaddam HS, Jafarzadeh-Esfehani R, Soleimanpour S. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin Microbiol Rev. 2022;35(1):e0033820. doi:10.1128/CMR.00338-20
8. Marano G, Mazza M, Lisci FM, et al. The microbiota-gut-brain axis: psychoneuroimmunological insights. Nutrients. 2023;15(6):1496. doi:10.3390/nu15061496
9. Kuziel GA, Rakoff-Nahoum S. The gut microbiome. Curr Biol. 2022;32(6):R257–r264. doi:10.1016/j.cub.2022.02.023
10. Cryan JF, O’Riordan KJ, Cowan CSM, et al. The Microbiota-Gut-Brain Axis, Physiol Rev. 2019;99(4):1877–2013.
11. Long-Smith C, O’Riordan KJ, Clarke G, Stanton C, Dinan TG, Cryan JF. Microbiota-gut-brain axis: new therapeutic opportunities. Annu Rev Pharmacol Toxicol. 2020;60:477–502. doi:10.1146/annurev-pharmtox-010919-023628
12. Agirman G, Yu KB, Hsiao EY. Signaling inflammation across the gut-brain axis. Science. 2021;374(6571):1087–1092. doi:10.1126/science.abi6087
13. Margolis KG, Cryan JF, Mayer EA. The microbiota-gut-brain axis: from motility to mood. Gastroenterology. 2021;160(5):1486–1501. doi:10.1053/j.gastro.2020.10.066
14. Chen P, Zhang L, Feng Y, et al. Brain-gut axis and psychiatric disorders: a perspective from bibliometric and visual analysis. Front Immunol. 2022;13:1047007. doi:10.3389/fimmu.2022.1047007
15. Mayer EA, Nance K, Chen S. The Gut-Brain Axis. Annu Rev Med. 2022;73:439–453. doi:10.1146/annurev-med-042320-014032
16. Sampson TR, Mazmanian SK. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe. 2015;17(5):565–576. doi:10.1016/j.chom.2015.04.011
17. Morais LH, Schreiber HLT, Mazmanian SK. The gut microbiota-brain axis in behaviour and brain disorders. Nat Rev Microbiol. 2021;19(4):241–255. doi:10.1038/s41579-020-00460-0
18. Naveed M, Zhou QG, Xu C, et al. Gut-brain axis: a matter of concern in neuropsychiatric disorders…! Prog Neuropsychopharmacol Biol Psychiatry. 2021;104:110051. doi:10.1016/j.pnpbp.2020.110051
19. Bermudez-Martin P, Becker JAJ, Caramello N, et al. The microbial metabolite p-Cresol induces autistic-like behaviors in mice by remodeling the gut microbiota. Microbiome. 2021;9(1):157. doi:10.1186/s40168-021-01103-z
20. de Theije CG, Wopereis H, Ramadan M, et al. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav Immun. 2014;37:197–206. doi:10.1016/j.bbi.2013.12.005
21. Hsiao EY, McBride SW, Hsien S, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155(7):1451–1463. doi:10.1016/j.cell.2013.11.024
22. Sharon G, Cruz NJ, Kang DW, et al. Human Gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell. 2019;177(6):1600–1618.e17. doi:10.1016/j.cell.2019.05.004
23. Golubeva AV, Joyce SA, Moloney G, et al. Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. EBioMedicine. 2017;24:166–178. doi:10.1016/j.ebiom.2017.09.020
24. Needham BD, Adame MD, Serena G, et al. Plasma and fecal metabolite profiles in autism spectrum disorder. Biol Psychiatry. 2021;89(5):451–462. doi:10.1016/j.biopsych.2020.09.025
25. Emond P, Mavel S, Aïdoud N, et al. GC-MS-based urine metabolic profiling of autism spectrum disorders. Anal Bioanal Chem. 2013;405(15):5291–5300. doi:10.1007/s00216-013-6934-x
26. Ming X, Stein TP, Barnes V, Rhodes N, Guo L. Metabolic perturbance in autism spectrum disorders: a metabolomics study. J Proteome Res. 2012;11(12):5856–5862. doi:10.1021/pr300910n
27. Kang DW, Ilhan ZE, Isern NG, et al. Differences in fecal microbial metabolites and microbiota of children with autism spectrum disorders. Anaerobe. 2018;49:121–131. doi:10.1016/j.anaerobe.2017.12.007
28. Li DF, Wang JY, Yang MF, et al. Factors associated with diagnostic accuracy, technical success and adverse events of endoscopic ultrasound-guided fine-needle biopsy: a systematic review and meta-analysis. J Gastroenterol Hepatol. 2020;35(8):1264–1276. doi:10.1111/jgh.14999
29. Sgritta M, Dooling SW, Buffington SA, et al. Mechanisms underlying microbial-mediated changes in social behavior in mouse models of autism spectrum disorder. Neuron. 2019;101(2):246–259.e6. doi:10.1016/j.neuron.2018.11.018
30. Tabouy L, Getselter D, Ziv O, et al. Dysbiosis of microbiome and probiotic treatment in a genetic model of autism spectrum disorders. Brain Behav Immun. 2018;73:310–319. doi:10.1016/j.bbi.2018.05.015
31. Kim S, Kim H, Yim YS, et al. Maternal gut bacteria promote neurodevelopmental abnormalities in mouse offspring. Nature. 2017;549(7673):528–532. doi:10.1038/nature23910
32. Blacher E, Bashiardes S, Shapiro H, et al. Potential roles of gut microbiome and metabolites in modulating ALS in mice. Nature. 2019;572(7770):474–480. doi:10.1038/s41586-019-1443-5
33. Sampson TR, Debelius JW, Thron T, et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of parkinson’s disease. Cell. 2016;167(6):1469–1480.e12. doi:10.1016/j.cell.2016.11.018
34. Cirstea MS, Yu AC, Golz E, et al. Microbiota composition and metabolism are associated with gut function in parkinson’s disease. Mov Disord. 2020;35(7):1208–1217. doi:10.1002/mds.28052
35. Keshavarzian A, Green SJ, Engen PA, et al. Colonic bacterial composition in Parkinson’s disease. Mov Disord. 2015;30(10):1351–1360. doi:10.1002/mds.26307
36. Bedarf JR, Hildebrand F, Coelho LP, et al. Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naïve Parkinson’s disease patients. Genome Med. 2017;9(1):39. doi:10.1186/s13073-017-0428-y
37. Unger MM, Spiegel J, Dillmann KU, et al. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Parkinsonism Relat Disord. 2016;32:66–72. doi:10.1016/j.parkreldis.2016.08.019
38. Castelli V, d’Angelo M, Lombardi F, et al. Effects of the probiotic formulation SLAB51 in in vitro and in vivo Parkinson’s disease models. Aging. 2020;12(5):4641–4659. doi:10.18632/aging.102927
39. Choi JG, Kim N, Ju IG, et al. Oral administration of proteus mirabilis damages dopaminergic neurons and motor functions in mice. Sci Rep. 2018;8(1):1275. doi:10.1038/s41598-018-19646-x
40. Hsieh TH, Kuo CW, Hsieh KH, et al. Probiotics alleviate the progressive deterioration of motor functions in a mouse model of parkinson’s disease. Brain Sci. 2020;10(4):206. doi:10.3390/brainsci10040206
41. Svensson E, Horváth-Puhó E, Thomsen RW, et al. Vagotomy and subsequent risk of Parkinson’s disease. Ann Neurol. 2015;78(4):522–529. doi:10.1002/ana.24448
42. Liu B, Fang F, Pedersen NL, et al. Vagotomy and Parkinson disease: a Swedish register-based matched-cohort study. Neurology. 2017;88(21):1996–2002. doi:10.1212/WNL.0000000000003961
43. Cekanaviciute E, Yoo BB, Runia TF, et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc Natl Acad Sci U S A. 2017;114(40):10713–10718. doi:10.1073/pnas.1711235114
44. Berer K, Gerdes LA, Cekanaviciute E, et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc Natl Acad Sci U S A. 2017;114(40):10719–10724. doi:10.1073/pnas.1711233114
45. Lee YK, Menezes JS, Umesaki Y, Mazmanian SK. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. 2011;108(Suppl 1):4615–4622. doi:10.1073/pnas.1000082107
46. Mangalam A, Shahi SK, Luckey D, et al. Human gut-derived commensal bacteria suppress CNS inflammatory and demyelinating disease. Cell Rep. 2017;20(6):1269–1277. doi:10.1016/j.celrep.2017.07.031
47. Mezö C, Dokalis N, Mossad O, et al. Different effects of constitutive and induced microbiota modulation on microglia in a mouse model of alzheimer’s disease. Acta Neuropathol Commun. 2020;8(1):119. doi:10.1186/s40478-020-00988-5
48. Minter MR, Zhang C, Leone V, et al. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer’s disease. Sci Rep. 2016;6:30028. doi:10.1038/srep30028
49. Perez-Pardo P, Dodiya HB, Engen PA, et al. Role of TLR4 in the gut-brain axis in Parkinson’s disease: a translational study from men to mice. Gut. 2019;68(5):829–843. doi:10.1136/gutjnl-2018-316844
50. Hasegawa S, Goto S, Tsuji H, et al. Intestinal dysbiosis and lowered serum lipopolysaccharide-binding protein in parkinson’s disease. PLoS One. 2015;10(11):e0142164. doi:10.1371/journal.pone.0142164
51. Wang X, Sun G, Feng T, et al. Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer’s disease progression. Cell Res. 2019;29(10):787–803. doi:10.1038/s41422-019-0216-x
52. Peter I, Dubinsky M, Bressman S, et al. Anti-tumor necrosis factor therapy and incidence of parkinson disease among patients with inflammatory bowel disease. JAMA Neurol. 2018;75(8):939–946. doi:10.1001/jamaneurol.2018.0605
53. Chen SG, Stribinskis V, Rane MJ, et al. Exposure to the functional bacterial amyloid protein curli enhances alpha-synuclein aggregation in aged fischer 344 rats and caenorhabditis elegans. Sci Rep. 2016;6:34477. doi:10.1038/srep34477
54. Gareau MG, Jury J, MacQueen G, Sherman PM, Perdue MH. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut. 2007;56(11):1522–1528. doi:10.1136/gut.2006.117176
55. van de Wouw M, Boehme M, Lyte JM, et al. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain-gut axis alterations. J Physiol. 2018;596(20):4923–4944. doi:10.1113/JP276431
56. Marin IA, Goertz JE, Ren T, et al. Microbiota alteration is associated with the development of stress-induced despair behavior. Sci Rep. 2017;7:43859. doi:10.1038/srep43859
57. Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21(6):786–796. doi:10.1038/mp.2016.44
58. Valles-Colomer M, Falony G, Darzi Y, et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol. 2019;4(4):623–632. doi:10.1038/s41564-018-0337-x
59. Allen AP, Hutch W, Borre YE, et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry. 2016;6(11):e939. doi:10.1038/tp.2016.191
60. Dalile B, Vervliet B, Bergonzelli G, Verbeke K, Van Oudenhove L. Colon-delivered short-chain fatty acids attenuate the cortisol response to psychosocial stress in healthy men: a randomized, placebo-controlled trial. Neuropsychopharmacology. 2020;45(13):2257–2266. doi:10.1038/s41386-020-0732-x
61. Bercik P, Park AJ, Sinclair D, et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol Motil. 2011;23(12):1132–1139. doi:10.1111/j.1365-2982.2011.01796.x
62. O’Leary OF, Felice D, Galimberti S, et al. GABAB(1) receptor subunit isoforms differentially regulate stress resilience. Proc Natl Acad Sci U S A. 2014;111(42):15232–15237. doi:10.1073/pnas.1404090111
63. Desbonnet L, Garrett L, Clarke G, Kiely B, Cryan JF, Dinan TG. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience. 2010;170(4):1179–1188. doi:10.1016/j.neuroscience.2010.08.005
64. Strandwitz P, Kim KH, Terekhova D, et al. GABA-modulating bacteria of the human gut microbiota. Nat Microbiol. 2019;4(3):396–403. doi:10.1038/s41564-018-0307-3
65. Pinto-Sanchez MI, Hall GB, Ghajar K, et al. Probiotic bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: a pilot study in patients with irritable bowel syndrome. Gastroenterology. 2017;153(2):448–459.e8. doi:10.1053/j.gastro.2017.05.003
66. Kelly JR, Borre Y, C OB, et al. Transferring the blues: depression-associated gut microbiota induces neurobehavioural changes in the rat. J Psychiatr Res. 2016;82:109–118. doi:10.1016/j.jpsychires.2016.07.019
67. Li N, Wang Q, Wang Y, et al. Oral probiotics ameliorate the behavioral deficits induced by chronic mild stress in mice via the gut microbiota-inflammation axis. Front Behav Neurosci. 2018;12:266. doi:10.3389/fnbeh.2018.00266
68. Kim YK, Shin C. The microbiota-gut-brain axis in neuropsychiatric disorders: pathophysiological mechanisms and novel treatments. Curr Neuropharmacol. 2018;16(5):559–573. doi:10.2174/1570159X15666170915141036
69. Rogers GB, Keating DJ, Young RL, Wong ML, Licinio J, Wesselingh S. From gut dysbiosis to altered brain function and mental illness: mechanisms and pathways. Mol Psychiatry. 2016;21(6):738–748. doi:10.1038/mp.2016.50
70. Tan AH, Lim SY, Lang AE. The microbiome-gut-brain axis in Parkinson disease - from basic research to the clinic. Nat Rev Neurol. 2022;18(8):476–495. doi:10.1038/s41582-022-00681-2
71. Chakrabarti A, Geurts L, Hoyles L, et al. The microbiota-gut-brain axis: pathways to better brain health. Perspectives on what we know, what we need to investigate and how to put knowledge into practice. Cell Mol Life Sci. 2022;79(2):80. doi:10.1007/s00018-021-04060-w
72. Toledo ARL, Monroy GR, Salazar FE, et al. Gut-Brain axis as a pathological and therapeutic target for neurodegenerative disorders. Int J Mol Sci. 2022;23(3):1184. doi:10.3390/ijms23031184
73. Lee A, Lee JY, Jung SW, et al. Brain-gut-microbiota axis. Korean J Gastroenterol. 2023;81(4):145–153. doi:10.4166/kjg.2023.028
74. Suganya K, Koo BS. Gut-brain axis: role of gut microbiota on neurological disorders and how probiotics/prebiotics beneficially modulate microbial and immune pathways to improve brain functions. Int J Mol Sci. 2020;21(20):7551. doi:10.3390/ijms21207551
75. Person H, Keefer L. Psychological comorbidity in gastrointestinal diseases: update on the brain-gut-microbiome axis. Prog Neuropsychopharmacol Biol Psychiatry. 2021;107:110209. doi:10.1016/j.pnpbp.2020.110209
76. Wenzel TJ, Gates EJ, Ranger AL, Klegeris A. Short-chain fatty acids (SCFAs) alone or in combination regulate select immune functions of microglia-like cells. Mol Cell Neurosci. 2020;105:103493. doi:10.1016/j.mcn.2020.103493
77. Braniste V, Al-Asmakh M, Kowal C, et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014;6(263):263ra158. doi:10.1126/scitranslmed.3009759
78. Stilling RM, van de Wouw M, Clarke G, Stanton C, Dinan TG, Cryan JF. The neuropharmacology of butyrate: the bread and butter of the microbiota-gut-brain axis? Neurochem Int. 2016;99:110–132. doi:10.1016/j.neuint.2016.06.011
79. Gill SR, Pop M, Deboy RT, et al. Metagenomic analysis of the human distal gut microbiome. Science. 2006;312(5778):1355–1359. doi:10.1126/science.1124234
80. Parthasarathy A, Cross PJ, Dobson RCJ, Adams LE, Savka MA, Hudson AO. A three-ring circus: metabolism of the three proteogenic aromatic amino acids and their role in the health of plants and animals. Front Mol Biosci. 2018;5:29. doi:10.3389/fmolb.2018.00029
81. Song S, Liu J, Zhang F, Hong JS. Norepinephrine depleting toxin DSP-4 and LPS alter gut microbiota and induce neurotoxicity in α-synuclein mutant mice. Sci Rep. 2020;10(1):15054. doi:10.1038/s41598-020-72202-4
82. Shandilya S, Kumar S, Kumar Jha N, Kumar Kesari K, Ruokolainen J. Interplay of gut microbiota and oxidative stress: perspective on neurodegeneration and neuroprotection. J Adv Res. 2022;38:223–244. doi:10.1016/j.jare.2021.09.005
83. Li D, Ke Y, Zhan R, et al. Trimethylamine-N-oxide promotes brain aging and cognitive impairment in mice. Aging Cell. 2018;17(4):e12768. doi:10.1111/acel.12768
84. Govindarajulu M, Pinky PD, Steinke I, et al. Gut metabolite TMAO induces synaptic plasticity deficits by promoting endoplasmic reticulum stress. Front Mol Neurosci. 2020;13:138. doi:10.3389/fnmol.2020.00138
85. Brunt VE, LaRocca TJ, Bazzoni AE, et al. The gut microbiome-derived metabolite trimethylamine N-oxide modulates neuroinflammation and cognitive function with aging. Geroscience. 2021;43(1):377–394. doi:10.1007/s11357-020-00257-2
86. Filosa S, Di Meo F, Crispi S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen Res. 2018;13(12):2055–2059. doi:10.4103/1673-5374.241429
87. Feng X, Li Y, Brobbey Oppong M, Qiu F. Insights into the intestinal bacterial metabolism of flavonoids and the bioactivities of their microbe-derived ring cleavage metabolites. Drug Metab Rev. 2018;50(3):343–356. doi:10.1080/03602532.2018.1485691
88. Gasperotti M, Passamonti S, Tramer F, et al. Fate of microbial metabolites of dietary polyphenols in rats: is the brain their target destination? ACS Chem Neurosci. 2015;6(8):1341–1352. doi:10.1021/acschemneuro.5b00051
89. Friedland RP, Chapman MR. The role of microbial amyloid in neurodegeneration. PLoS Pathog. 2017;13(12):e1006654. doi:10.1371/journal.ppat.1006654
90. Sampson TR, Challis C, Jain N, et al. A gut bacterial amyloid promotes α-synuclein aggregation and motor impairment in mice. Elife. 2020;9. doi:10.7554/eLife.53111
91. Needham BD, Kaddurah-Daouk R, Mazmanian SK. Gut microbial molecules in behavioural and neurodegenerative conditions. Nat Rev Neurosci. 2020;21(12):717–731. doi:10.1038/s41583-020-00381-0
92. Diviccaro S, FitzGerald JA, Cioffi L, et al. Gut steroids and microbiota: effect of gonadectomy and sex. Biomolecules. 2022;12(6):767. doi:10.3390/biom12060767
93. Furness JB. Types of neurons in the enteric nervous system. J Auton Nerv Syst. 2000;81(1–3):87–96. doi:10.1016/S0165-1838(00)00127-2
94. Gershon MD, Margolis KG. The gut, its microbiome, and the brain: connections and communications. J Clin Invest. 2021;131(18). doi:10.1172/JCI143768
95. Luqman A, Nega M, Nguyen MT, Ebner P, Götz F. SadA-expressing Staphylococci in the Human gut show increased cell adherence and internalization. Cell Rep. 2018;22(2):535–545. doi:10.1016/j.celrep.2017.12.058
96. Roshchina VV. New trends and perspectives in the evolution of neurotransmitters in microbial, plant, and animal cells. Adv Exp Med Biol. 2016;874:25–77.
97. Sarkar C, Basu B, Chakroborty D, Dasgupta PS, Basu S. The immunoregulatory role of dopamine: an update. Brain Behav Immun. 2010;24(4):525–528. doi:10.1016/j.bbi.2009.10.015
98. Färber K, Pannasch U, Kettenmann H. Dopamine and noradrenaline control distinct functions in rodent microglial cells. Mol Cell Neurosci. 2005;29(1):128–138. doi:10.1016/j.mcn.2005.01.003
99. Borodovitsyna O, Flamini M, Chandler D. Noradrenergic Modulation of cognition in health and disease. Neural Plast. 2017;2017:6031478. doi:10.1155/2017/6031478
100. Tsavkelova EA, Botvinko IV, Kudrin VS, Oleskin AV. Detection of neurotransmitter amines in microorganisms with the use of high-performance liquid chromatography. Dokl Biochem. 2000;372(1–6):115–117.
101. Zafra F, Lindholm D, Castrén E, Hartikka J, Thoenen H. Regulation of brain-derived neurotrophic factor and nerve growth factor mRNA in primary cultures of hippocampal neurons and astrocytes. J Neurosci. 1992;12(12):4793–4799. doi:10.1523/JNEUROSCI.12-12-04793.1992
102. O’Donnell J, Zeppenfeld D, McConnell E, Pena S, Nedergaard M. Norepinephrine: a neuromodulator that boosts the function of multiple cell types to optimize CNS performance. Neurochem Res. 2012;37(11):2496–2512. doi:10.1007/s11064-012-0818-x
103. Barrett E, Ross RP, O’Toole PW, Fitzgerald GF, Stanton C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J Appl Microbiol. 2012;113(2):411–417. doi:10.1111/j.1365-2672.2012.05344.x
104. Takanaga H, Ohtsuki S, Hosoya K, Terasaki T. GAT2/BGT-1 as a system responsible for the transport of gamma-aminobutyric acid at the mouse blood-brain barrier. J Cereb Blood Flow Metab. 2001;21(10):1232–1239. doi:10.1097/00004647-200110000-00012
105. Al-Sarraf H. Transport of 14C-gamma-aminobutyric acid into brain, cerebrospinal fluid and choroid plexus in neonatal and adult rats. Brain Res Dev Brain Res. 2002;139(2):121–129. doi:10.1016/S0165-3806(02)00537-0
106. Shyamaladevi N, Jayakumar AR, Sujatha R, Paul V, Subramanian EH. Evidence that nitric oxide production increases gamma-amino butyric acid permeability of blood-brain barrier. Brain Res Bull. 2002;57(2):231–236. doi:10.1016/S0361-9230(01)00755-9
107. Bjurstöm H, Wang J, Ericsson I, et al. GABA, a natural immunomodulator of T lymphocytes. J Neuroimmunol. 2008;205(1–2):44–50. doi:10.1016/j.jneuroim.2008.08.017
108. Wu C, Sun D. GABA receptors in brain development, function, and injury. Metab Brain Dis. 2015;30(2):367–379. doi:10.1007/s11011-014-9560-1
109. Evrensel A, Ceylan ME. The gut-brain axis: the missing link in depression. Clin Psychopharmacol Neurosci. 2015;13(3):239–244. doi:10.9758/cpn.2015.13.3.239
110. Abbott NJ. Inflammatory mediators and modulation of blood-brain barrier permeability. Cell Mol Neurobiol. 2000;20(2):131–147. doi:10.1023/A:1007074420772
111. Sternberg EM, Wedner HJ, Leung MK, Parker CW. Effect of serotonin (5-HT) and other monoamines on murine macrophages: modulation of interferon-gamma induced phagocytosis. J Immunol. 1987;138(12):4360–4365. doi:10.4049/jimmunol.138.12.4360
112. Kubera M, Maes M, Kenis G, Kim YK, Lasoń W. Effects of serotonin and serotonergic agonists and antagonists on the production of tumor necrosis factor alpha and interleukin-6. Psychiatry Res. 2005;134(3):251–258. doi:10.1016/j.psychres.2004.01.014
113. Liu MT, Kuan YH, Wang J, Hen R, Gershon MD. 5-HT4 receptor-mediated neuroprotection and neurogenesis in the enteric nervous system of adult mice. J Neurosci. 2009;29(31):9683–9699. doi:10.1523/JNEUROSCI.1145-09.2009
114. McVey Neufeld KA, Perez-Burgos A, Mao YK, Bienenstock J, Kunze WA. The gut microbiome restores intrinsic and extrinsic nerve function in germ-free mice accompanied by changes in calbindin. Neurogastroenterol Motil. 2015;27(5):627–636. doi:10.1111/nmo.12534
115. Jiang SH, Li J, Dong FY, et al. Increased serotonin signaling contributes to the warburg effect in pancreatic tumor cells under metabolic stress and promotes growth of pancreatic tumors in mice. Gastroenterology. 2017;153(1):277–291.e19. doi:10.1053/j.gastro.2017.03.008
116. Roth W, Zadeh K, Vekariya R, Ge Y, Mohamadzadeh M. Tryptophan metabolism and gut-brain homeostasis. Int J Mol Sci. 2021;22(6):2973. doi:10.3390/ijms22062973
117. Sudo N. Biogenic amines: signals between commensal microbiota and gut physiology. Front Endocrinol. 2019;10:504. doi:10.3389/fendo.2019.00504
118. Chen Y, Xu J, Chen Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients. 2021;13(6):1.
119. Hata T, Asano Y, Yoshihara K, et al. Regulation of gut luminal serotonin by commensal microbiota in mice. PLoS One. 2017;12(7):e0180745. doi:10.1371/journal.pone.0180745
120. McLean PG, Borman RA, Lee K. 5-HT in the enteric nervous system: gut function and neuropharmacology. Trends Neurosci. 2007;30(1):9–13. doi:10.1016/j.tins.2006.11.002
121. Guo C, Huo YJ, Li Y, Han Y, Zhou D. Gut-brain axis: focus on gut metabolites short-chain fatty acids. World J Clin Cases. 2022;10(6):1754–1763. doi:10.12998/wjcc.v10.i6.1754
122. Liu S, Gao J, Liu K, Zhang HL. Microbiota-gut-brain axis and Alzheimer’s disease: implications of the blood-brain barrier as an intervention target. Mech Ageing Dev. 2021;199:111560. doi:10.1016/j.mad.2021.111560
123. Silva YP, Bernardi A, Frozza RL. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol. 2020;11:25. doi:10.3389/fendo.2020.00025
124. O’Riordan KJ, Collins MK, Moloney GM, et al. Short chain fatty acids: microbial metabolites for gut-brain axis signalling. Mol Cell Endocrinol. 2022;546:111572. doi:10.1016/j.mce.2022.111572
125. Siopi E, Galerne M, Rivagorda M, et al. Gut microbiota changes require vagus nerve integrity to promote depressive-like behaviors in mice. Mol Psychiatry. 2023;28(7):3002–3012. doi:10.1038/s41380-023-02071-6
126. Tan C, Yan Q, Ma Y, Fang J, Yang Y. Recognizing the role of the vagus nerve in depression from microbiota-gut brain axis. Front Neurol. 2022;13:1015175. doi:10.3389/fneur.2022.1015175
127. Forsythe P, Bienenstock J, Kunze WA. Vagal pathways for microbiome-brain-gut axis communication. Adv Exp Med Biol. 2014;817:115–133.
128. Cawthon CR, de La Serre CB. Gut bacteria interaction with vagal afferents. Brain Res. 2018;1693(Pt B):134–139. doi:10.1016/j.brainres.2018.01.012
129. Bonaz B, Bazin T, Pellissier S. The vagus nerve at the interface of the microbiota-gut-brain axis. Front Neurosci. 2018;12:49. doi:10.3389/fnins.2018.00049
130. Goswami C, Iwasaki Y, Yada T. Short-chain fatty acids suppress food intake by activating vagal afferent neurons. J Nutr Biochem. 2018;57:130–135. doi:10.1016/j.jnutbio.2018.03.009
131. Ge T, Yao X, Zhao H, et al. Gut microbiota and neuropsychiatric disorders: implications for neuroendocrine-immune regulation. Pharmacol Res. 2021;173:105909. doi:10.1016/j.phrs.2021.105909
132. Masanetz RK, Winkler J, Winner B, Günther C, Süß P. The gut-immune-brain axis: an important route for neuropsychiatric morbidity in inflammatory bowel disease. Int J Mol Sci. 2022;23(19):11111. doi:10.3390/ijms231911111
133. Villageliú DN, Rasmussen S, Lyte M. A microbial endocrinology-based simulated small intestinal medium for the evaluation of neurochemical production by gut microbiota. FEMS Microbiol Ecol. 2018;94(7). doi:10.1093/femsec/fiy096
134. Marietta E, Horwath I, Taneja V. Microbiome, immunomodulation, and the neuronal system. Neurotherapeutics. 2018;15(1):23–30. doi:10.1007/s13311-017-0601-4
135. Engelstoft MS, Egerod KL, Lund ML, Schwartz TW. Enteroendocrine cell types revisited. Curr Opin Pharmacol. 2013;13(6):912–921. doi:10.1016/j.coph.2013.09.018
136. Adriaenssens AE, Reimann F, Gribble FM. Distribution and stimulus secretion coupling of enteroendocrine cells along the intestinal tract. Compr Physiol. 2018;8(4):1603–1638. doi:10.1002/j.2040-4603.2018.tb00048.x
137. Gheorghe CE, Martin JA, Manriquez FV, Dinan TG, Cryan JF, Clarke G. Focus on the essentials: tryptophan metabolism and the microbiome-gut-brain axis. Curr Opin Pharmacol. 2019;48:137–145. doi:10.1016/j.coph.2019.08.004
138. Yamane S, Inagaki N. Regulation of glucagon-like peptide-1 sensitivity by gut microbiota dysbiosis. J Diabetes Investig. 2018;9(2):262–264. doi:10.1111/jdi.12762
139. Cheng LK, O’Grady G, Du P, Egbuji JU, Windsor JA, Pullan AJ. Gastrointestinal system. Wiley Interdiscip Rev Syst Biol Med. 2010;2(1):65–79. doi:10.1002/wsbm.19
140. Vodička M, Ergang P, Hrnčíř T, et al. Microbiota affects the expression of genes involved in HPA axis regulation and local metabolism of glucocorticoids in chronic psychosocial stress. Brain Behav Immun. 2018;73:615–624. doi:10.1016/j.bbi.2018.07.007
141. Frankiensztajn LM, Elliott E, Koren O. The microbiota and the hypothalamus-pituitary-adrenocortical (HPA) axis, implications for anxiety and stress disorders. Curr Opin Neurobiol. 2020;62:76–82. doi:10.1016/j.conb.2019.12.003
142. de Weerth C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neurosci Biobehav Rev. 2017;83:458–471. doi:10.1016/j.neubiorev.2017.09.016
143. Misiak B, Łoniewski I, Marlicz W, et al. The HPA axis dysregulation in severe mental illness: can we shift the blame to gut microbiota? Prog Neuropsychopharmacol Biol Psychiatry. 2020;102:109951. doi:10.1016/j.pnpbp.2020.109951
144. Kasarello K, Cudnoch-Jedrzejewska A, Czarzasta K. Communication of gut microbiota and brain via immune and neuroendocrine signaling. Front Microbiol. 2023;14:1118529. doi:10.3389/fmicb.2023.1118529
145. Lima-Ojeda JM, Rupprecht R, Baghai TC. Gut microbiota and depression: pathophysiology of depression: hypothalamic-pituitary-adrenal axis and microbiota-gut-brain axis. Nervenarzt. 2020;91(12):1108–1114. doi:10.1007/s00115-020-01029-1
146. Tiwari K, Singh M, Kumar P, Mukhopadhyay K. Binding of cationic analogues of α-MSH to lipopolysaccharide and disruption of the cytoplasmic membranes caused bactericidal action against Escherichia coli. Sci Rep. 2022;12(1):1987. doi:10.1038/s41598-022-05684-z
147. Dominique M, Breton J, Guérin C, et al. Effects of macronutrients on the in vitro production of ClpB, a bacterial mimetic protein of α-MSH and its possible role in satiety signaling. Nutrients. 2019;11(9):2115. doi:10.3390/nu11092115
148. Guan Q. A comprehensive review and update on the pathogenesis of inflammatory bowel disease. J Immunol Res. 2019;2019:7247238. doi:10.1155/2019/7247238
149. Kim JS, Chen MH, Wang HE, Lu CL, Wang YP, Zhang B. Inflammatory bowel disease and neurodegenerative diseases. Gut Liver. 2023;17(4):495–504. doi:10.5009/gnl220523
150. Abautret-Daly Á, Dempsey E, Parra-Blanco A, Medina C, Harkin A. Gut-brain actions underlying comorbid anxiety and depression associated with inflammatory bowel disease. Acta Neuropsychiatr. 2018;30(5):275–296. doi:10.1017/neu.2017.3
151. Banfi D, Moro E, Bosi A, et al. Impact of microbial metabolites on microbiota-gut-brain axis in inflammatory bowel disease. Int J Mol Sci. 2021;22(4):1.
152. Li H, Sun J, Du J, et al. Clostridium butyricum exerts a neuroprotective effect in a mouse model of traumatic brain injury via the gut-brain axis. Neurogastroenterol Motil. 2018;30(5):e13260. doi:10.1111/nmo.13260
153. Sun J, Xu J, Yang B, et al. Effect of clostridium butyricum against microglia-mediated neuroinflammation in alzheimer’s disease via regulating gut microbiota and metabolites butyrate. Mol Nutr Food Res. 2020;64(2):e1900636. doi:10.1002/mnfr.201900636
154. Bourassa MW, Alim I, Bultman SJ, Ratan RR. Butyrate, neuroepigenetics and the gut microbiome: can a high fiber diet improve brain health? Neurosci Lett. 2016;625:56–63. doi:10.1016/j.neulet.2016.02.009
155. Aggeletopoulou I, Konstantakis C, Assimakopoulos SF, Triantos C. The role of the gut microbiota in the treatment of inflammatory bowel diseases. Microb Pathog. 2019;137:103774. doi:10.1016/j.micpath.2019.103774
156. Emge JR, Huynh K, Miller EN, et al. Modulation of the microbiota-gut-brain axis by probiotics in a murine model of inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol. 2016;310(11):G989–98. doi:10.1152/ajpgi.00086.2016
157. Nishida A, Nishino K, Sakai K, Owaki Y, Noda Y, Imaeda H. Can control of gut microbiota be a future therapeutic option for inflammatory bowel disease? World J Gastroenterol. 2021;27(23):3317–3326. doi:10.3748/wjg.v27.i23.3317
158. Airola C, Severino A, Porcari S, et al. Future modulation of gut microbiota: from eubiotics to FMT, engineered bacteria, and phage therapy. Antibiotics. 2023;12(5):868. doi:10.3390/antibiotics12050868
159. Reid G, Abrahamsson T, Bailey M, et al. How do probiotics and prebiotics function at distant sites? Benef Microbes. 2017;8(4):521–533. doi:10.3920/BM2016.0222
160. Liu G, Yu Q, Tan B, et al. Gut dysbiosis impairs hippocampal plasticity and behaviors by remodeling serum metabolome. Gut Microbes. 2022;14(1):2104089. doi:10.1080/19490976.2022.2104089
161. Finegold SM, Dowd SE, Gontcharova V, et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe. 2010;16(4):444–453. doi:10.1016/j.anaerobe.2010.06.008
162. Strati F, Cavalieri D, Albanese D, et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome. 2017;5(1):24. doi:10.1186/s40168-017-0242-1
163. Kushak RI, Winter HS, Buie TM, Cox SB, Phillips CD, Ward NL. Analysis of the duodenal microbiome in autistic individuals: association with carbohydrate digestion. J Pediatr Gastroenterol Nutr. 2017;64(5):e110–e116. doi:10.1097/MPG.0000000000001458
164. Ma B, Liang J, Dai M, et al. Altered gut microbiota in chinese children with autism spectrum disorders. Front Cell Infect Microbiol. 2019;9:40. doi:10.3389/fcimb.2019.00040
165. Scheperjans F, Aho V, Pereira PA, et al. Gut microbiota are related to parkinson’s disease and clinical phenotype. Mov Disord. 2015;30(3):350–358. doi:10.1002/mds.26069
166. Petrov VA, Saltykova IV, Zhukova IA, et al. Analysis of gut microbiota in patients with parkinson’s disease. Bull Exp Biol Med. 2017;162(6):734–737. doi:10.1007/s10517-017-3700-7
167. Vogt NM, Kerby RL, Dill-McFarland KA, et al. Gut microbiome alterations in Alzheimer’s disease. Sci Rep. 2017;7(1):13537. doi:10.1038/s41598-017-13601-y
168. Zhuang ZQ, Shen LL, Li WW, et al. Gut microbiota is altered in patients with alzheimer’s disease. J Alzheimers Dis. 2018;63(4):1337–1346. doi:10.3233/JAD-180176
169. Haran JP, Bhattarai SK, Foley SE, et al. Alzheimer’s disease microbiome is associated with dysregulation of the anti-inflammatory P-glycoprotein pathway. mBio. 2019;10(3). doi:10.1128/mBio.00632-19
170. Šimić G, Tkalčić M, Vukić V, et al. Understanding emotions: origins and roles of the amygdala. Biomolecules. 2021;11(6):823. doi:10.3390/biom11060823
171. Critchley HD, Garfinkel SN. Interoception and emotion. Curr Opin Psychol. 2017;17:7–14. doi:10.1016/j.copsyc.2017.04.020
172. Ménard C, Hodes GE, Russo SJ. Pathogenesis of depression: insights from human and rodent studies. Neuroscience. 2016;321:138–162. doi:10.1016/j.neuroscience.2015.05.053
173. Miller K, Massie MJ. Depression and anxiety. Cancer J. 2006;12(5):388–397. doi:10.1097/00130404-200609000-00008
174. Qian X, Li Q, Zhu H, et al. Bifidobacteria with indole-3-lactic acid-producing capacity exhibit psychobiotic potential via reducing neuroinflammation. Cell Rep Med. 2024;5:101798. doi:10.1016/j.xcrm.2024.101798
175. Fuke N, Takagi T, Higashimura Y, et al. Lactobacillus brevis KB290 with Vitamin A ameliorates murine intestinal inflammation associated with the increase of CD11c+ macrophage/CD103- dendritic cell ratio. Inflamm Bowel Dis. 2018;24(2):317–331. doi:10.1093/ibd/izx003
176. Aizawa E, Tsuji H, Asahara T, et al. Possible association of bifidobacterium and lactobacillus in the gut microbiota of patients with major depressive disorder. J Affect Disord. 2016;202:254–257. doi:10.1016/j.jad.2016.05.038
177. Kim IB, Park SC, Kim YK. Microbiota-gut-brain axis in major depression: a new therapeutic approach. Adv Exp Med Biol. 2023;1411:209–224.
178. Wu M, Tian T, Mao Q, et al. Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl Psychiatry. 2020;10(1):350. doi:10.1038/s41398-020-01038-3
179. Kronsten VT, Tranah TH, Pariante C, Shawcross DL. Gut-derived systemic inflammation as a driver of depression in chronic liver disease. J Hepatol. 2022;76(3):665–680. doi:10.1016/j.jhep.2021.11.008
180. Hu X, Li Y, Wu J, et al. Changes of gut microbiota reflect the severity of major depressive disorder: a cross sectional study. Transl Psychiatry. 2023;13(1):137. doi:10.1038/s41398-023-02436-z
181. O’Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res. 2015;277:32–48. doi:10.1016/j.bbr.2014.07.027
182. Bastiaanssen TFS, Cowan CSM, Claesson MJ, Dinan TG, Cryan JF. Making sense of … the microbiome in psychiatry. Int J Neuropsychopharmacol. 2019;22(1):37–52. doi:10.1093/ijnp/pyy067
183. Chiba S, Numakawa T, Ninomiya M, Richards MC, Wakabayashi C, Kunugi H. Chronic restraint stress causes anxiety- and depression-like behaviors, downregulates glucocorticoid receptor expression, and attenuates glutamate release induced by brain-derived neurotrophic factor in the prefrontal cortex. Prog Neuropsychopharmacol Biol Psychiatry. 2012;39(1):112–119. doi:10.1016/j.pnpbp.2012.05.018
184. Haghighat N, Rajabi S, Mohammadshahi M. Effect of synbiotic and probiotic supplementation on serum brain-derived neurotrophic factor level, depression and anxiety symptoms in hemodialysis patients: a randomized, double-blinded, clinical trial. Nutr Neurosci. 2021;24(6):490–499. doi:10.1080/1028415X.2019.1646975
185. Reynoso C, Rangel MJ, Melgar V. Autism spectrum disorder: etiological, diagnostic and therapeutic aspects. Rev Med Inst Mex Seguro Soc. 2017;55(2):214–222.
186. Joon P, Kumar A, Parle M. What is autism? Pharmacol Rep. 2021;73(5):1255–1264. doi:10.1007/s43440-021-00244-0
187. Sharma SR, Gonda X, Tarazi FI. Autism spectrum disorder: classification, diagnosis and therapy. Pharmacol Ther. 2018;190:91–104. doi:10.1016/j.pharmthera.2018.05.007
188. Ahrens AP, Hyotylainen T, Petrone JR, et al. Infant microbes and metabolites point to childhood neurodevelopmental disorders. Cell. 2024;187:1853–1873e15. doi:10.1016/j.cell.2024.02.035
189. Su Q, Wong OWH, Lu W, et al. Multikingdom and functional gut microbiota markers for autism spectrum disorder. Nat Microbiol. 2024;9:2344–2355. doi:10.1038/s41564-024-01739-1
190. Xiao L, Yan J, Yang T, et al. Fecal microbiome transplantation from children with autism spectrum disorder modulates tryptophan and serotonergic synapse metabolism and induces altered behaviors in germ-free mice. mSystems. 2021;6(2). doi:10.1128/msystems.01343-20
191. Kang DW, Adams JB, Vargason T, Santiago M, Hahn J, Krajmalnik-Brown R. Distinct fecal and plasma metabolites in children with autism spectrum disorders and their modulation after microbiota transfer therapy. mSphere. 2020;5(5). doi:10.1128/mSphere.00314-20
192. Needham BD, Tang W, Wu WL. Searching for the gut microbial contributing factors to social behavior in rodent models of autism spectrum disorder. Dev Neurobiol. 2018;78(5):474–499. doi:10.1002/dneu.22581
193. Davoli-Ferreira M, Thomson CA, McCoy KD. Microbiota and microglia interactions in ASD. Front Immunol. 2021;12:676255. doi:10.3389/fimmu.2021.676255
194. Fattorusso A, Di Genova L, Dell’Isola GB, Mencaroni E, Esposito S. Autism spectrum disorders and the gut microbiota. Nutrients. 2019;11(3):521. doi:10.3390/nu11030521
195. Kang DW, Adams JB, Gregory AC, et al. Microbiota transfer therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: an open-label study. Microbiome. 2017;5(1):10. doi:10.1186/s40168-016-0225-7
196. Finegold SM, Downes J, Summanen PH. Microbiology of regressive autism. Anaerobe. 2012;18(2):260–262. doi:10.1016/j.anaerobe.2011.12.018
197. Frye RE, Slattery J, MacFabe DF, et al. Approaches to studying and manipulating the enteric microbiome to improve autism symptoms. Microb Ecol Health Dis. 2015;26:26878. doi:10.3402/mehd.v26.26878
198. Brown CC, Manis MM, Bohm NM, Curry SR. Oral vancomycin for secondary prophylaxis of clostridium difficile infection. Ann Pharmacother. 2019;53(4):396–401. doi:10.1177/1060028018815170
199. Kang DW, Adams JB, Coleman DM, et al. Long-term benefit of microbiota transfer therapy on autism symptoms and gut microbiota. Sci Rep. 2019;9(1):5821. doi:10.1038/s41598-019-42183-0
200. Zhernakova A, Kurilshikov A, Bonder MJ, et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science. 2016;352(6285):565–569. doi:10.1126/science.aad3369
201. Vafadari B. Stress and the role of the gut-brain axis in the pathogenesis of schizophrenia: a literature review. Int J Mol Sci. 2021;22(18):9747. doi:10.3390/ijms22189747
202. Zamanpoor M. Schizophrenia in a genomic era: a review from the pathogenesis, genetic and environmental etiology to diagnosis and treatment insights. Psychiatr Genet. 2020;30(1):1–9. doi:10.1097/YPG.0000000000000245
203. Wahbeh MH, Avramopoulos D. Gene-environment interactions in schizophrenia: a literature review. Genes. 2021;12(12):1850. doi:10.3390/genes12121850
204. Schmitt A, Falkai P, Papiol S. Neurodevelopmental disturbances in schizophrenia: evidence from genetic and environmental factors. J Neural Transm. 2023;130(3):195–205. doi:10.1007/s00702-022-02567-5
205. Wu H, Jiawei X, Wen Z, et al. Microbiome-gut-brain profiles in schizophrenia and their potential link to cognitive performance: findings from a case-control study. Schizophr Bull. 2025. doi:10.1093/schbul/sbaf028
206. Munawar N, Ahsan K, Muhammad K, et al. Hidden role of gut microbiome dysbiosis in schizophrenia: antipsychotics or psychobiotics as therapeutics? Int J Mol Sci. 2021;22(14):7671. doi:10.3390/ijms22147671
207. Li S, Song J, Ke P, et al. The gut microbiome is associated with brain structure and function in schizophrenia. Sci Rep. 2021;11(1):9743. doi:10.1038/s41598-021-89166-8
208. Samochowiec J, Misiak B. Gut microbiota and microbiome in schizophrenia. Curr Opin Psychiatry. 2021;34(5):503–507. doi:10.1097/YCO.0000000000000733
209. Ghorbani M, Rajandas H, Parimannan S, et al. Understanding the role of gut microbiota in the pathogenesis of schizophrenia. Psychiatr Genet. 2021;31(2):39–49. doi:10.1097/YPG.0000000000000270
210. Zhu F, Ju Y, Wang W, et al. Metagenome-wide association of gut microbiome features for schizophrenia. Nat Commun. 2020;11(1):1612. doi:10.1038/s41467-020-15457-9
211. Burbaeva G, Boksha IS, Starodubtseva LI, et al. Glutamate dysmetabolism in patients with schizophrenia. Vestn Ross Akad Med Nauk. 2007;3:19–24.
212. Xu R, Wu B, Liang J, et al. Altered gut microbiota and mucosal immunity in patients with schizophrenia. Brain Behav Immun. 2020;85:120–127. doi:10.1016/j.bbi.2019.06.039
213. Zheng P, Zeng B, Liu M, et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci Adv. 2019;5(2):eaau8317. doi:10.1126/sciadv.aau8317
214. Opler MG, Susser ES. Fetal environment and schizophrenia. Environ Health Perspect. 2005;113(9):1239–1242. doi:10.1289/ehp.7572
215. Kneeland RE, Fatemi SH. Viral infection, inflammation and schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2013;42:35–48. doi:10.1016/j.pnpbp.2012.02.001
216. Pyndt Jørgensen B, Krych L, Pedersen TB, et al. Investigating the long-term effect of subchronic phencyclidine-treatment on novel object recognition and the association between the gut microbiota and behavior in the animal model of schizophrenia. Physiol Behav. 2015;141:32–39. doi:10.1016/j.physbeh.2014.12.042
217. Khan S, Barve KH, Kumar MS. Recent advancements in pathogenesis, diagnostics and treatment of alzheimer’s disease. Curr Neuropharmacol. 2020;18(11):1106–1125. doi:10.2174/1570159X18666200528142429
218. Jia L, Ke Y, Zhao S, et al. Metagenomic analysis characterizes stage-specific gut microbiota in alzheimer’s disease. Mol Psychiatry. 2025;30(9):3951–3962. doi:10.1038/s41380-025-02973-7
219. Rostagno AA. Pathogenesis of alzheimer’s disease. Int J Mol Sci. 2022;24(1):107. doi:10.3390/ijms24010107
220. Zhang H, Zheng Y. β amyloid hypothesis in alzheimer’s disease:pathogenesis, prevention, and management. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2019;41(5):702–708. doi:10.3881/j.issn.1000-503X.10875
221. Jiang C, Li G, Huang P, Liu Z, Zhao B. The gut microbiota and alzheimer’s disease. J Alzheimers Dis. 2017;58(1):1–15. doi:10.3233/JAD-161141
222. Xi J, Ding D, Zhu H, et al. Disturbed microbial ecology in Alzheimer’s disease: evidence from the gut microbiota and fecal metabolome. BMC Microbiol. 2021;21(1):226. doi:10.1186/s12866-021-02286-z
223. Benichou Haziot C, Birak KS. Therapeutic potential of microbiota modulation in alzheimer’s disease: a review of preclinical studies. J Alzheimers Dis Rep. 2023;7(1):415–431. doi:10.3233/ADR-220097
224. Leblhuber F, Ehrlich D, Steiner K, et al. The immunopathogenesis of alzheimer’s disease is related to the composition of gut microbiota. Nutrients. 2021;13(2):361. doi:10.3390/nu13020361
225. Zhuang Z, Yang R, Wang W, Qi L, Huang T. Associations between gut microbiota and Alzheimer’s disease, major depressive disorder, and schizophrenia. J Neuroinflammation. 2020;17(1):288. doi:10.1186/s12974-020-01961-8
226. Liu S, Gao J, Zhu M, Liu K, Zhang HL. Gut microbiota and dysbiosis in alzheimer’s disease: implications for pathogenesis and treatment. Mol Neurobiol. 2020;57(12):5026–5043. doi:10.1007/s12035-020-02073-3
227. Wang Y, Li L, Zhao X, et al. Intestinal microflora changes in patients with mild alzheimer’s disease in a Chinese cohort. J Alzheimers Dis. 2022;88(2):563–575. doi:10.3233/JAD-220076
228. Hung CC, Chang CC, Huang CW, Nouchi R, Cheng CH. Gut microbiota in patients with alzheimer’s disease spectrum: a systematic review and meta-analysis. Aging. 2022;14(1):477–496. doi:10.18632/aging.203826
229. Braak H, Braak E. Neuropathological stageing of alzheimer-related changes. Acta Neuropathol. 1991;82(4):239–259. doi:10.1007/BF00308809
230. Hampel H, Blennow K, Shaw LM, Hoessler YC, Zetterberg H, Trojanowski JQ. Total and phosphorylated tau protein as biological markers of Alzheimer’s disease. Exp Gerontol. 2010;45(1):30–40. doi:10.1016/j.exger.2009.10.010
231. Vázquez-Vélez GE, Zoghbi HY. Parkinson’s disease genetics and pathophysiology. Annu Rev Neurosci. 2021;44:87–108. doi:10.1146/annurev-neuro-100720-034518
232. Ye H, Robak LA, Yu M, Cykowski M, Shulman JM. Genetics and pathogenesis of parkinson’s syndrome. Annu Rev Pathol. 2023;18:95–121. doi:10.1146/annurev-pathmechdis-031521-034145
233. Schapira AH, Jenner P. Etiology and pathogenesis of parkinson’s disease. Mov Disord. 2011;26(6):1049–1055. doi:10.1002/mds.23732
234. Alexoudi A, Alexoudi I, Gatzonis S. Parkinson’s disease pathogenesis, evolution and alternative pathways: a review. Rev Neurol. 2018;174(10):699–704. doi:10.1016/j.neurol.2017.12.003
235. DeWeerdt S. Parkinson’s gut-microbiota links raise treatment possibilities. Nature. 2025. doi:10.1038/d41586-025-01253-2
236. Sun MF, Shen YQ. Dysbiosis of gut microbiota and microbial metabolites in parkinson’s disease. Ageing Res Rev. 2018;45:53–61. doi:10.1016/j.arr.2018.04.004
237. Nishiwaki H, Ito M, Ishida T, et al. Meta-analysis of gut dysbiosis in parkinson’s disease. Mov Disord. 2020;35(9):1626–1635. doi:10.1002/mds.28119
238. Aho VTE, Houser MC, Pereira PAB, et al. Relationships of gut microbiota, short-chain fatty acids, inflammation, and the gut barrier in Parkinson’s disease. Mol Neurodegener. 2021;16(1):6. doi:10.1186/s13024-021-00427-6
239. Haneishi Y, Furuya Y, Hasegawa M, Picarelli A, Rossi M, Miyamoto J. Inflammatory bowel diseases and gut microbiota. Int J Mol Sci. 2023;24(4):3817. doi:10.3390/ijms24043817
240. Zhao Z, Ning J, Bao XQ, et al. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome. 2021;9(1):226. doi:10.1186/s40168-021-01107-9
241. Sui YT, Bullock KM, Erickson MA, Zhang J, Banks WA. Alpha synuclein is transported into and out of the brain by the blood-brain barrier. Peptides. 2014;62:197–202. doi:10.1016/j.peptides.2014.09.018
242. Block ML, Zecca L, Hong JS. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat Rev Neurosci. 2007;8(1):57–69. doi:10.1038/nrn2038
243. Bernstein CN, Hitchon CA, Walld R, et al. Increased Burden of psychiatric disorders in inflammatory bowel disease. Inflamm Bowel Dis. 2019;25(2):360–368. doi:10.1093/ibd/izy235
244. Gluch P, Swiatek A, Dudek P, Fabisiak A, Talar-Wojnarowska R. Neurological manifestations and psychiatric disorders in the course of inflammatory bowel diseases. J Gastrointestin Liver Dis. 2022;31(1):107–118. doi:10.15403/jgld-3965
245. Bernstein CN. the brain-gut axis and stress in inflammatory bowel disease. Gastroenterol Clin North Am. 2017;46(4):839–846. doi:10.1016/j.gtc.2017.08.006
246. Eldridge F, Raine T. Understanding and addressing the psychological burden of IBD. J Crohns Colitis. 2022;16(2):177–178. doi:10.1093/ecco-jcc/jjab130
247. Marrie RA, Walld R, Bolton JM, et al. Increased incidence of psychiatric disorders in immune-mediated inflammatory disease. J Psychosom Res. 2017;101:17–23. doi:10.1016/j.jpsychores.2017.07.015
248. Eugenicos MP, Ferreira NB. Psychological factors associated with inflammatory bowel disease. Br Med Bull. 2021;138(1):16–28. doi:10.1093/bmb/ldab010
249. Bisgaard TH, Allin KH, Keefer L, Ananthakrishnan AN, Jess T. Depression and anxiety in inflammatory bowel disease: epidemiology, mechanisms and treatment. Nat Rev Gastroenterol Hepatol. 2022;19(11):717–726. doi:10.1038/s41575-022-00634-6
250. Marrie RA, Graff LA, Fisk JD, Patten SB, Bernstein CN. The relationship between symptoms of depression and anxiety and disease activity in IBD over time. Inflamm Bowel Dis. 2021;27(8):1285–1293. doi:10.1093/ibd/izaa349
251. Hu S, Chen Y, Chen Y, Wang C. Depression and anxiety disorders in patients with inflammatory bowel disease. Front Psychiatry. 2021;12:714057. doi:10.3389/fpsyt.2021.714057
252. Neuendorf R, Harding A, Stello N, Hanes D, Wahbeh H. Depression and anxiety in patients with inflammatory bowel disease: a systematic review. J Psychosom Res. 2016;87:70–80. doi:10.1016/j.jpsychores.2016.06.001
253. Timmer A, Preiss JC, Motschall E, Rücker G, Jantschek G, Moser G. Psychological interventions for treatment of inflammatory bowel disease. Cochrane Database Syst Rev. 2011;2:Cd006913.
254. Tiles-Sar N, Neuser J, de Sordi D, et al. Psychological interventions for inflammatory bowel disease: a systematic review and component network meta-analysis protocol. BMJ Open. 2022;12(6):e056982. doi:10.1136/bmjopen-2021-056982
255. Fiest KM, Bernstein CN, Walker JR, et al. Systematic review of interventions for depression and anxiety in persons with inflammatory bowel disease. BMC Res Notes. 2016;9(1):404. doi:10.1186/s13104-016-2204-2
256. Gracie DJ, Irvine AJ, Sood R, Mikocka-Walus A, Hamlin PJ, Ford AC. Effect of psychological therapy on disease activity, psychological comorbidity, and quality of life in inflammatory bowel disease: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2017;2(3):189–199. doi:10.1016/S2468-1253(16)30206-0
257. Kim JY, Choi MJ, Ha S, et al. Association between autism spectrum disorder and inflammatory bowel disease: a systematic review and meta-analysis. Autism Res. 2022;15(2):340–352. doi:10.1002/aur.2656
258. Doshi-Velez F, Avillach P, Palmer N, et al. Prevalence of inflammatory bowel disease among patients with autism spectrum disorders. Inflamm Bowel Dis. 2015;21(10):2281–2288. doi:10.1097/MIB.0000000000000502
259. Lee M, Krishnamurthy J, Susi A, et al. Association of Autism spectrum disorders and inflammatory bowel disease. J Autism Dev Disord. 2018;48(5):1523–1529. doi:10.1007/s10803-017-3409-5
260. McPartland J, Volkmar FR. Autism and related disorders. Handb Clin Neurol. 2012;106:407–418.
261. Horvath K, Perman JA. Autism and gastrointestinal symptoms. Curr Gastroenterol Rep. 2002;4(3):251–258. doi:10.1007/s11894-002-0071-6
262. Fulceri F, Morelli M, Santocchi E, et al. Gastrointestinal symptoms and behavioral problems in preschoolers with autism spectrum disorder. Dig Liver Dis. 2016;48(3):248–254. doi:10.1016/j.dld.2015.11.026
263. Szandruk-Bender M, Wiatrak B, Szeląg A. The risk of developing alzheimer’s disease and parkinson’s disease in patients with inflammatory bowel disease: a meta-analysis. J Clin Med. 2022;11(13):3704. doi:10.3390/jcm11133704
264. Kim GH, Lee YC, Kim TJ, et al. Risk of neurodegenerative diseases in patients with inflammatory bowel disease: a nationwide population-based cohort study. J Crohns Colitis. 2022;16(3):436–443. doi:10.1093/ecco-jcc/jjab162
265. Sugihara K, Kamada N. Diet-microbiota interactions in inflammatory bowel disease. Nutrients. 2021;13(5):1533. doi:10.3390/nu13051533
266. Temko JE, Bouhlal S, Farokhnia M, Lee MR, Cryan JF, Leggio L. The microbiota, the gut and the brain in eating and alcohol use disorders: a ‘ménage à trois’? Alcohol Alcohol. 2017;52(4):403–413. doi:10.1093/alcalc/agx024
267. Elmaleh DR, Downey MA, Kundakovic L, Wilkinson JE, Neeman Z, Segal E. New approaches to profile the microbiome for treatment of neurodegenerative disease. J Alzheimers Dis. 2021;82(4):1373–1401. doi:10.3233/JAD-210198
268. Zeng R, Wang J, Zheng C, et al. Lack of causal associations of inflammatory bowel disease with parkinson’s disease and other neurodegenerative disorders. Mov Disord. 2023;38(6):1082–1088. doi:10.1002/mds.29386
269. Fu Q, Song T, Ma X, Cui J. Research progress on the relationship between intestinal microecology and intestinal bowel disease. Animal Model Exp Med. 2022;5(4):297–310. doi:10.1002/ame2.12262
270. Floch MH. Intestinal microecology in health and wellness. J Clin Gastroenterol. 2011;45:S108–10. doi:10.1097/MCG.0b013e3182309276
271. Gomaa EZ. Human gut microbiota/microbiome in health and diseases: a review. Antonie van Leeuwenhoek. 2020;113(12):2019–2040. doi:10.1007/s10482-020-01474-7
272. Dixit K, Chaudhari D, Dhotre D, Shouche Y, Saroj S. Restoration of dysbiotic human gut microbiome for homeostasis. Life Sci. 2021;278:119622. doi:10.1016/j.lfs.2021.119622
273. Kataoka K. The intestinal microbiota and its role in human health and disease. J Med Invest. 2016;63(1–2):27–37. doi:10.2152/jmi.63.27
274. Li HY, Zhou DD, Gan RY, et al. Effects and mechanisms of probiotics, prebiotics, synbiotics, and postbiotics on metabolic diseases targeting gut microbiota. A Narrative Review, Nutrients. 2021;13(9):1.
275. Liu X, Cao S, Zhang X. Modulation of gut microbiota-brain axis by probiotics, prebiotics, and diet. J Agric Food Chem. 2015;63(36):7885–7895. doi:10.1021/acs.jafc.5b02404
276. Adak A, Khan MR. An insight into gut microbiota and its functionalities. Cell Mol Life Sci. 2019;76(3):473–493. doi:10.1007/s00018-018-2943-4
277. Pesce M, Seguella L, Del Re A, et al. Next-generation probiotics for inflammatory bowel disease. Int J Mol Sci. 2022;23(10):5466. doi:10.3390/ijms23105466
278. So D, Quigley EMM, Whelan K. Probiotics in irritable bowel syndrome and inflammatory bowel disease: review of mechanisms and effectiveness. Curr Opin Gastroenterol. 2023;39(2):103–109. doi:10.1097/MOG.0000000000000902
279. De Greef E, Vandenplas Y, Hauser B, Devreker T, Veereman G. The use of probiotics in IBD and IBS. Minerva Pediatr. 2014;66(5):491–500.
280. Abraham BP, Quigley EMM. Probiotics in inflammatory bowel disease. Gastroenterol Clin North Am. 2017;46(4):769–782. doi:10.1016/j.gtc.2017.08.003
281. Adam B, Liebregts T, Holtmann G. Maintaining remission of ulcerative colitis with the probiotic Escherichia Coli Nissle 1917 is as effective as with standard mesalazine. Z Gastroenterol. 2006;44(3):267–269. doi:10.1055/s-2005-858972
282. Rembacken BJ, Snelling AM, Hawkey PM, Chalmers DM, Axon AT. Non-pathogenic Escherichia coli versus mesalazine for the treatment of ulcerative colitis: a randomised trial. Lancet. 1999;354(9179):635–639. doi:10.1016/S0140-6736(98)06343-0
283. Orel R, Kamhi Trop T. Intestinal microbiota, probiotics and prebiotics in inflammatory bowel disease. World J Gastroenterol. 2014;20(33):11505–11524. doi:10.3748/wjg.v20.i33.11505
284. Ganji-Arjenaki M, Rafieian-Kopaei M. Probiotics are a good choice in remission of inflammatory bowel diseases: a meta analysis and systematic review. J Cell Physiol. 2018;233(3):2091–2103. doi:10.1002/jcp.25911
285. Magalhães-Guedes KT. Psychobiotic Therapy: method to reinforce the immune system. Clin Psychopharmacol Neurosci. 2022;20(1):17–25. doi:10.9758/cpn.2022.20.1.17
286. Cheng LH, Liu YW, Wu CC, Wang S, Tsai YC. Psychobiotics in mental health, neurodegenerative and neurodevelopmental disorders. J Food Drug Anal. 2019;27(3):632–648. doi:10.1016/j.jfda.2019.01.002
287. De Oliveira FL, Salgaço MK, de Oliveira MT, et al. Exploring the Potential of lactobacillus helveticus R0052 and bifidobacterium longum r0175 as promising psychobiotics using SHIME. Nutrients. 2023;15(6):1521. doi:10.3390/nu15061521
288. Lewis ED, Antony JM, Crowley DC, et al. Efficacy of lactobacillus paracasei HA-196 and bifidobacterium longum R0175 in alleviating symptoms of irritable Bowel Syndrome (IBS): a randomized, placebo-controlled study. Nutrients. 2020;12(4):1159. doi:10.3390/nu12041159
289. Ilie OD, Paduraru E, Robea MA, et al. The possible role of bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 on locomotor activity and oxidative stress in a rotenone-induced zebrafish model of parkinson’s disease. Oxid Med Cell Longev. 2021;2021:9629102. doi:10.1155/2021/9629102
290. Surwase SN, Jadhav JP. Bioconversion of L-tyrosine to L-DOPA by a novel bacterium Bacillus sp. JPJ, Amino Acids. 2011;41(2):495–506. doi:10.1007/s00726-010-0768-z
291. Feng YK, Wu QL, Peng YW, et al. Oral P. gingivalis impairs gut permeability and mediates immune responses associated with neurodegeneration in LRRK2 R1441G mice. J Neuroinflammation. 2020;17(1):347. doi:10.1186/s12974-020-02027-5
292. Roberfroid M, Gibson GR, Hoyles L, et al. Prebiotic effects: metabolic and health benefits. Br J Nutr. 2010;104(2):S1–63. doi:10.1017/S0007114510003363
293. Neri-Numa IA, Pastore GM. Novel insights into prebiotic properties on human health: a review. Food Res Int. 2020;131:108973. doi:10.1016/j.foodres.2019.108973
294. Holscher HD. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes. 2017;8(2):172–184. doi:10.1080/19490976.2017.1290756
295. Sanders ME, Merenstein DJ, Reid G, Gibson GR, Rastall RA. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat Rev Gastroenterol Hepatol. 2019;16(10):605–616. doi:10.1038/s41575-019-0173-3
296. Quigley EMM. Prebiotics and probiotics in digestive health. Clin Gastroenterol Hepatol. 2019;17(2):333–344. doi:10.1016/j.cgh.2018.09.028
297. Chunchai T, Thunapong W, Yasom S, et al. Decreased microglial activation through gut-brain axis by prebiotics, probiotics, or synbiotics effectively restored cognitive function in obese-insulin resistant rats. J Neuroinflammation. 2018;15(1):11. doi:10.1186/s12974-018-1055-2
298. Roselli M, Finamore A. Use of synbiotics for ulcerative colitis treatment. Curr Clin Pharmacol. 2020;15(3):174–182. doi:10.2174/1574884715666191226120322
299. Drabińska N, Krupa-Kozak U, Jarocka-Cyrta E. Intestinal permeability in children with celiac disease after the administration of oligofructose-enriched inulin into a gluten-free diet-results of a randomized, placebo-controlled, pilot trial. Nutrients. 2020;12(6):1736. doi:10.3390/nu12061736
300. Burokas A, Arboleya S, Moloney RD, et al. Targeting the microbiota-gut-brain axis: prebiotics have anxiolytic and antidepressant-like effects and reverse the impact of chronic stress in mice. Biol Psychiatry. 2017;82(7):472–487. doi:10.1016/j.biopsych.2016.12.031
301. Ghanizadeh A, Moghimi-Sarani E. A randomized double blind placebo controlled clinical trial of N-acetylcysteine added to risperidone for treating autistic disorders. BMC Psychiatry. 2013;13:196. doi:10.1186/1471-244X-13-196
302. Lee YS, Lai DM, Huang HJ, et al. Prebiotic lactulose ameliorates the cognitive deficit in alzheimer’s disease mouse model through macroautophagy and chaperone-mediated autophagy pathways. J Agric Food Chem. 2021;69(8):2422–2437. doi:10.1021/acs.jafc.0c07327
303. Antushevich H. Fecal microbiota transplantation in disease therapy. Clin Chim Acta. 2020;503:90–98. doi:10.1016/j.cca.2019.12.010
304. Ianiro G, Segal JP, Mullish BH, et al. Fecal microbiota transplantation in gastrointestinal and extraintestinal disorders. Future Microbiol. 2020;15:1173–1183. doi:10.2217/fmb-2020-0061
305. Ooijevaar RE, Terveer EM, Verspaget HW, Kuijper EJ, Keller JJ. Clinical application and potential of fecal microbiota transplantation. Annu Rev Med. 2019;70:335–351. doi:10.1146/annurev-med-111717-122956
306. Mahmoudi H, Hossainpour H. Application and development of fecal microbiota transplantation in the treatment of gastrointestinal and metabolic diseases: a review. Saudi J Gastroenterol. 2023;29(1):3–11. doi:10.4103/sjg.sjg_131_22
307. Ianiro G, Bibbò S, Porcari S, et al. Fecal microbiota transplantation for recurrent C. difficile infection in patients with inflammatory bowel disease: experience of a large-volume European FMT center. Gut Microbes. 2021;13(1):1994834. doi:10.1080/19490976.2021.1994834
308. Haber SL, Raney CRK, Larson TL, Lau JP. Fecal microbiota transplantation for recurrent Clostridioides difficile infection. Am J Health Syst Pharm. 2019;76(13):935–942. doi:10.1093/ajhp/zxz078
309. Yang D, Zhao D, Ali Shah SZ, et al. The Role of the gut microbiota in the pathogenesis of parkinson’s disease. Front Neurol. 2019;10:1155. doi:10.3389/fneur.2019.01155
310. The Alzheimer’s Association. 2020 Alzheimer’s disease facts and figures. Alzheimers Dement. 2020. doi:10.1002/alz.12068
311. Sun J, Xu J, Ling Y, et al. Fecal microbiota transplantation alleviated Alzheimer’s disease-like pathogenesis in APP/PS1 transgenic mice. Transl Psychiatry. 2019;9(1):189. doi:10.1038/s41398-019-0525-3
312. Żebrowska P, Łaczmańska I, Łaczmański Ł. Future directions in reducing gastrointestinal disorders in children with ASD using fecal microbiota transplantation. Front Cell Infect Microbiol. 2021;11:630052. doi:10.3389/fcimb.2021.630052
313. Beam A, Clinger E, Hao L. Effect of diet and dietary components on the composition of the gut microbiota. Nutrients. 2021;13(8):2795. doi:10.3390/nu13082795
314. Berding K, Vlckova K, Marx W, et al. Diet and the microbiota-gut-brain axis: sowing the seeds of good mental health. Adv Nutr. 2021;12(4):1239–1285. doi:10.1093/advances/nmaa181
315. Wolter M, Grant ET, Boudaud M, et al. Leveraging diet to engineer the gut microbiome. Nat Rev Gastroenterol Hepatol. 2021;18(12):885–902. doi:10.1038/s41575-021-00512-7
316. Malesza IJ, Malesza M, Walkowiak J, et al. High-fat, western-style diet, systemic inflammation, and gut microbiota: a narrative review. Cells. 2021;10(11):3164. doi:10.3390/cells10113164
317. Paoli A, Mancin L, Bianco A, Thomas E, Mota JF, Piccini F. Ketogenic diet and microbiota: friends or enemies? Genes. 2019;10(7):534. doi:10.3390/genes10070534
318. Merra G, Noce A, Marrone G, et al. Influence of mediterranean diet on human gut microbiota. Nutrients. 2020;13(1):7. doi:10.3390/nu13010007
319. Gubert C, Kong G, Renoir T, Hannan AJ. Exercise, diet and stress as modulators of gut microbiota: implications for neurodegenerative diseases. Neurobiol Dis. 2020;134:104621. doi:10.1016/j.nbd.2019.104621
320. Chen SJ, Chen CC, Liao HY, et al. Association of fecal and plasma levels of short-chain fatty acids with gut microbiota and clinical severity in patients with parkinson disease. Neurology. 2022;98(8):e848–e858. doi:10.1212/WNL.0000000000013225
321. Schultz ST, Klonoff-Cohen HS, Wingard DL, et al. Breastfeeding, infant formula supplementation, and autistic disorder: the results of a parent survey. Int Breastfeed J. 2006;1:16. doi:10.1186/1746-4358-1-16
322. Weersma RK, Zhernakova A, Fu J. Interaction between drugs and the gut microbiome. Gut. 2020;69(8):1510–1519. doi:10.1136/gutjnl-2019-320204
323. Kim DH. Gut microbiota-mediated drug-antibiotic interactions. Drug Metab Dispos. 2015;43(10):1581–1589. doi:10.1124/dmd.115.063867
324. Le Bastard Q, Al-Ghalith GA, Grégoire M, et al. Systematic review: human gut dysbiosis induced by non-antibiotic prescription medications. Aliment Pharmacol Ther. 2018;47(3):332–345. doi:10.1111/apt.14451
325. Walsh J, Griffin BT, Clarke G, Hyland NP. Drug-gut microbiota interactions: implications for neuropharmacology. Br J Pharmacol. 2018;175(24):4415–4429. doi:10.1111/bph.14366
326. Maier L, Pruteanu M, Kuhn M, et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature. 2018;555(7698):623–628. doi:10.1038/nature25979
327. Pimentel M. Review of rifaximin as treatment for SIBO and IBS. Expert Opin Investig Drugs. 2009;18(3):349–358. doi:10.1517/13543780902780175
328. Prantera C, Lochs H, Grimaldi M, Danese S, Scribano ML, Gionchetti P. Rifaximin-extended intestinal release induces remission in patients with moderately active Crohn’s disease. Gastroenterology. 2012;142(3):473–481.e4. doi:10.1053/j.gastro.2011.11.032
329. Sartor RB. Review article: the potential mechanisms of action of rifaximin in the management of inflammatory bowel diseases. Aliment Pharmacol Ther. 2016;43(Suppl 1):27–36. doi:10.1111/apt.13436
330. Lopetuso LR, Napoli M, Rizzatti G, Gasbarrini A. The intriguing role of Rifaximin in gut barrier chronic inflammation and in the treatment of Crohn’s disease. Expert Opin Investig Drugs. 2018;27(6):543–551. doi:10.1080/13543784.2018.1483333
331. Seeman MV. What is the significance of the impact of antipsychotics on the gut microbiome? Expert Opin Drug Metab Toxicol. 2023;19(3):125–127. doi:10.1080/17425255.2023.2200161
332. Chen A, Park TY, Li KJ, DeLisi LE. Antipsychotics and the microbiota. Curr Opin Psychiatry. 2020;33(3):225–230. doi:10.1097/YCO.0000000000000594
333. Manchia M, Fontana A, Panebianco C, et al. Involvement of gut microbiota in schizophrenia and treatment resistance to antipsychotics. Biomedicines. 2021;9(8):875. doi:10.3390/biomedicines9080875
334. Luppens SB, Kara D, Bandounas L, et al. Effect of veillonella parvula on the antimicrobial resistance and gene expression of Streptococcus mutans grown in a dual-species biofilm. Oral Microbiol Immunol. 2008;23(3):183–189. doi:10.1111/j.1399-302X.2007.00409.x
335. Müller RT, Pos KM. The assembly and disassembly of the AcrAB-TolC three-component multidrug efflux pump. Biol Chem. 2015;396(9–10):1083–1089. doi:10.1515/hsz-2015-0150
336. Baldi S, Mundula T, Nannini G, Amedei A. Microbiota shaping - the effects of probiotics, prebiotics, and fecal microbiota transplant on cognitive functions: a systematic review. World J Gastroenterol. 2021;27(39):6715–6732. doi:10.3748/wjg.v27.i39.6715
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.