Back to Journals » Veterinary Medicine: Research and Reports » Volume 6
Methicillin-resistant Staphylococcus aureus colonization and infection risks from companion animals: current perspectives
Authors Petinaki E, Spiliopoulou I
Received 28 June 2015
Accepted for publication 16 September 2015
Published 6 November 2015 Volume 2015:6 Pages 373—382
DOI https://doi.org/10.2147/VMRR.S91313
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Young Lyoo
Efthimia Petinaki,1 Iris Spiliopoulou2
1Department of Microbiology, School of Medicine, University of Thessalia, Larissa, 2Department of Microbiology, School of Medicine, University of Patras, Patras, Greece
Abstract: Methicillin-resistant Staphylococcus aureus (MRSA) remains one of the most virulent human pathogens and has also recently been recognized as such in the veterinary settings. Companion animals, including dogs, cats, horses, small exotic animals, wildlife animals, and livestock, may constitute a reservoir for MRSA transmission to humans and vice versa. The evolution, emergence, and risk factors for MRSA transmission among colonized or infected animals are reviewed in the present paper, and infection control practices are discussed.
Keywords: methicillin-resistant Staphylococcus aureus, companion animals, close contacts
Introduction
Methicillin-resistant Staphylococcus aureus infections in humans and animals
Methicillin-resistant S. aureus evolution
S. aureus is one of the most common pathogens among humans and animals. Since its description by Sir Alexander Ogston in 1882 and the German physician Friedrich J Rosenbach, it still remains a very successful bacterium.1 It shows a continuous evolution in terms of antibiotic resistance from the mid-1940s, when beta-lactamase producing strains were detected, till nowadays with the acquisition of vancomycin resistance determinants.1,2 S. aureus carries on the chromosome several genomic islands, including antibiotic resistance determinants and virulence genes.3 Genomic islands are mobile genetic elements and are classified according to the location of their attachment site and the evolution process of their integrase gene.3
Methicillin-resistant S. aureus (MRSA) was first isolated in 1960, after the introduction of semisynthetic penicillins in the clinical practice, and soon became a worldwide health problem, spreading in hospital settings.1,4 Methicillin resistance is due to the expression of a new penicillin-binding protein (PBP2a), a transpeptidase encoded mostly by mecA gene, located in a mobile genetic element, staphylococcal cassette chromosome (SCC), known as SCCmec.1,3 The generation of MRSA is a result of SCCmec integration into the oriC region of a methicillin-susceptible S. aureus (MSSA). Until now, eleven SCCmec types (I–XI) have been registered, but since whole-genome sequencing analysis continues new types are expected to be characterized.3 SCCmec types I–III were first identified in hospital-associated MRSA strains, carrying several additional antibiotic resistance determinants.3,4 Shorter in their length are SCCmec types IV and V, carrying only mec–gene complex expressing a more susceptible phenotype to antimicrobials and were first isolated from community-associated infections.3,4 Another mecA homolog is mecC that is located in SCCmec type XI and characterized in strains of animal and human origin.3
Methicillin-resistant S. aureus infections in humans
MRSA causes 80,000 invasive infections and 11,000 resultant deaths, as reported from the USA in 2011.5 Rates of asymptomatic nasal colonization in the general population range from 1.5% for MRSA to ≥30% for S. aureus at any given time.6,7 It constitutes the etiologic agent of a wide range of diseases causing skin and soft tissue infections (SSTIs), necrotizing pneumonia, toxin syndromes, systemic invasive infections, etc.4,8 This spectrum of diseases reflects the ability of strains to synthesize surface proteins that contribute to adhesion, toxins, and peptides mediating immune responses and enzymes contributing to their survival and virulence.9–12
Epidemiologic studies rely on molecular characterization of clonal lineages that were spread in the hospitals and the community. The most widely used methods are phage typing, followed by pulsed-field gel electrophoresis (PFGE), multilocus sequence typing (MLST), and spa typing.4 MLST classifies S. aureus isolates on the basis of allelic variation in seven housekeeping genes; therefore, clones consist of isolates with identical sequences at all seven loci and are assigned a unique sequence type (ST); clonal complexes comprise closely related STs differing by single nucleotide polymorphisms to a lesser than three loci.13 Nowadays, whole-genome sequence approaches provide information about clonal evolution, virulence, and antibiotic resistance, giving more concise information about this pathogen.4
Strains from nosocomial MRSA infections, characterized as health care-associated MRSA (HA-MRSA), are typically isolated after 72 hours of hospital admission and usually contain MRSA clones harboring SCCmec type I, II, or III (Table 1). Such strains are mainly associated with bacteremia, pneumonia, or urinary tract infections.4,8,14 Nosocomial MRSA is mostly reported as an adult pathogen, but it has also been associated with infections in pediatric and neonatal intensive care units.4,8,14 Some MRSA lineages such as those belonging to clonal complex CC30 and sequence type ST239 were among the most widely spread in the health care system, being responsible for HA-MRSA outbreaks.4 However, CC30 strains have reduced their virulence capacity, a fact that may lead to underestimation of their spread.4 ST239 strains harbor SCCmecIII element and have been classified into three major subclones (clades).4 It is remarkable that selected MRSA lineages are disseminated in every continent causing the majority of nosocomial infections, including bloodstream, catheter-related, surgical wounds infections, meningitis associated with shunt, deep seated abscesses, empyema, and bone or joint infections (Table 1).1,4,8,14–17 In the late 1990s, MRSA emerged as a community pathogen among patients without predisposing risk factors or any relation to health care settings (community-associated MRSA [CA-MRSA]).1,4 Isolates from CA-MRSA infections are typically recovered in <72 hours of hospital admission; they usually harbor SCCmec type IV or V and cause skin and SSTI (Table 1).1,4,9,12–17 These infections were identified as outbreaks in different countries; however, fatal cases were also soon reported.1,4 CA-MRSA clones were identified in USA, Europe, Latin America, Australia, and Asia. They were usually susceptible to non-beta-lactam antimicrobials, carried SCCmec types IV and V, phenol-soluble modulins, and Panton–Valentine leukocidin genes (PVL); recently, they have infiltrated the health care system.1,4,9,12 These strains are mainly associated with SSTIs; however, they may cause more invasive life-threatening infections, such as necrotizing pneumonia, musculoskeletal diseases, and septicemias, to a lesser extent (5%–10%).1,4,9,12,14 MRSA clones are transmitted within households and they can persist on surfaces.14
 | Table 1 Characteristics differentiating between health care and community-associated MRSA (HA-MRSA, CA-MRSA) infections in humans |
Furthermore, S. aureus causes food-borne diseases resulting from the consumption of preformed enterotoxins with contaminated food by toxin-producing strains.10 Sporadic cases are referred, but it seems that the incidence is much higher and underreported.
Methicillin-resistant S. aureus infections in animals
Apart from humans, MRSA has been recognized as an important pathogen of animals, including food chain ones, companion and exotic animals, as well as horses. Close proximity of animals with humans led to the spread of bacterial species, interchangeably from animals to humans and vice versa.16 Molecular typing revealed that MRSA strains with particular genetic characteristics (lineages or clones) are associated with specific hosts (animal species), whereas other lineages are widely disseminated (Table 2).16,18 Some paradigms include CC97, CC151, and CC130 strains, which were very rarely isolated from humans, causing bovine mastitis; ST398 clone was first isolated from pigs and later from other animal species and humans, whereas ST1 clone of human origin was later recovered from cattle and pigs in Europe (Table 2).16,18 In most cases, MRSA clones isolated from companion animals differ from those of food chain animals and are of human origin. However, this issue has not been definitely resolved since a study from Belgium demonstrated that ST398 strains are spread among pigs, humans, and pets living in the same farm.19 MRSA infections among companion animals including horses mainly concern SSTIs and surgical wounds.16,19,20 In cattle, they cause mastitis, whereas exudative epidermitis is reported among pigs.20 A large-scale study in Germany revealed that CC22 and CC5 MRSA causing wound infections in humans predominated among infected dogs and cats, whereas CC398 strains were isolated from horses, suggesting that this lineage was adapted to a new host (Table 2).21,22
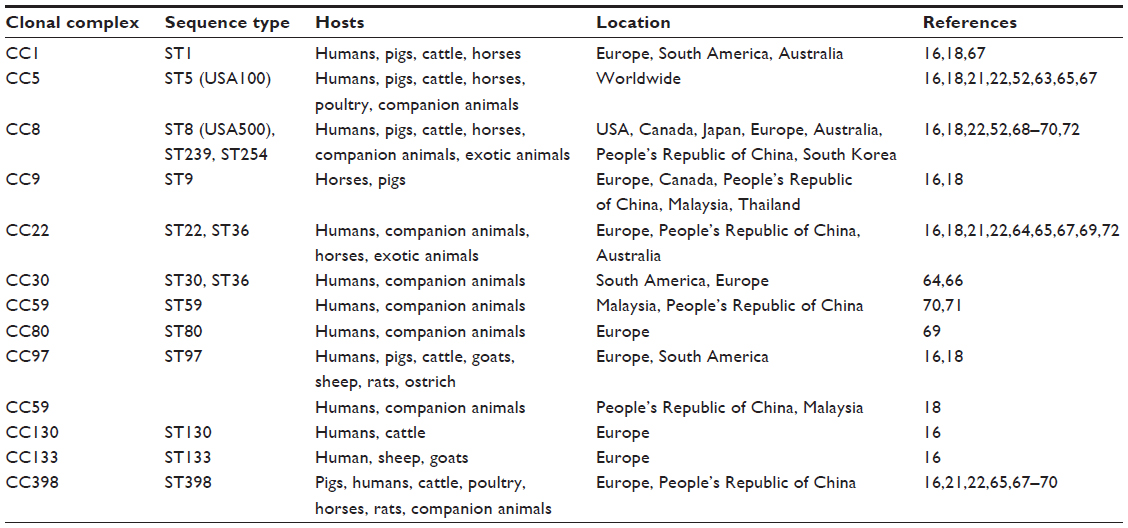 | Table 2 Methicillin-resistant S. aureus (MRSA) most common clones identified in humans and animals according to location |
Emergence of MRSA in veterinary settings
The predominant staphylococcal species colonizing or infecting animals vary according to animal species. Although S. aureus is the main species in humans, other coagulase-positive staphylococci, such as Staphylococcus pseudintermedius, predominate in domestic animals. However, an increased incidence of human infections due to MRSA probably led to the spread of MRSA in veterinary species.23 Faires et al evaluated the rate of MRSA transmission from infected animals to humans and vice versa. Upon MRSA-infected animal identification, at least one MRSA-colonized household person was identified in >25% (6/22, 27.3%) of the cases. In contrast, only one out of eight (12.5%) MRSA-infected humans had an MRSA-colonized pet.24 The increasing number of published sporadic MRSA infections, MRSA-infectious diseases, and MRSA outbreaks in veterinary medicine justifies their recognition as a “New Emerging Pathogen” today.25
The first MRSA isolation was described in 1972, from a bovine mastitis case, and ever since, several reports from animal-linked populations were documented.26–29 MRSA strains are isolated from a number of vertebrate hosts, including cats, dogs, horses, cattle, chickens, rabbits, and pigs.30 In cattle, MRSA is a major mastitis pathogen that is spread through cow-to-cow, usually during milking.31 It is interesting that even though MRSA is commonly isolated from milk, bacteria have been infrequently recovered from cases of clinical and subclinical bovine mastitis.32,33 Although MRSA can be isolated from cattle, infection is relatively rare.
In 1997, Hartmann et al reported one MRSA strain isolated from a postoperative wound infection of a horse,29 whereas in 1999, Seguin et al reported an equine outbreak taken place in a veterinary hospital.34 In horses, the types of MRSA involved in colonization or infection vary depending on the region studied. Canadian MRSA-5, a rare strain to humans, is the most frequent one isolated in the country from horses and horse personnel. In Europe, other MRSA strains have been identified including ST398, a type of porcine origin (Table 2).35,36 Recently, according to whole-genome analytical and epidemiologic studies, S. aureus ST398 strains are clustered into two distinct phylogenetic clades, a livestock and a basal human clade.37,38 Main characteristics of the former clade are the loss of bacteriophage ΦSa3 and the acquisition of a Tn916-like transposon carrying the tetM gene; human S. aureus ST398 clade harbor ΦSa3 and the human-specific immune evasion cluster genes that carry the scn and chp, encoding a staphylococcal complement inhibitor and a chemotaxis inhibitor protein, respectively.37,38
The first report of MRSA isolation in swine populations was described in France.39 Swine-associated MRSA colonization and infection have now been detected across Europe, in North America, and Asia, with a rate of colonization among pigs up to 49%.40–43 The emergence of MRSA ST398 in swine and the association of human colonization with such strains among veal calf surgeons suggest that cattle may serve as a reservoir for MRSA colonization or infection.44 This is strengthened by the fact that ST398 strains colonize humans who work with swine or veal calves in Europe. In addition, farm rats are colonized by ST398 MRSA, suggesting that farm rodent populations may play a role in dissemination and persistence of such strains during swine operations.45 Furthermore, dust samples taken from swine operations harbor MRSA, suggesting that a transmission route from farm animals to dust followed by colonization of rats and other farm rodent populations may play a role as environmental reservoirs for human colonization and infection.43
All avian species appear to be susceptible to staphylococcal infections and are common wherever poultry are raised. S. aureus is the usual causative agent, but other staphylococci (Staphylococcus hyicus and Staphylococcus cohnii) may also be found. The rate of methicillin resistance varies among regions. Although USA meat is usually disinfected, recent studies have demonstrated that 25% of chicken and turkey retail samples were positive for S. aureus, whereas 3.9% and 1.7%, respectively, were positive for MRSA in Michigan.46 The prevalence of S. aureus in turkey and chicken in Iowa was 19.4% and 17.8%, respectively, without any MRSA isolation.47 Most of poultry MRSA belong to ST5, a clone of human origin.48 This clone was also detected in turkeys, but with a lower frequency. Other turkey MRSA isolates belong to CC9 and CC398, which are both livestock-associated lineages.49
Dogs and cats may become colonized or infected with S. aureus, including MRSA. Usually, they are colonized by other staphylococcal species, several of which express methicillin resistance. Various studies clearly demonstrate that MRSA strains found in companion animals are frequently identical to human epidemic clones.16–18,28,50–52 Associated infections usually include pyoderma or postoperative wounds infections, and infrequently otitis, urinary tract infections, and arthropathies.53–55
Detection of S. aureus and specifically MRSA in wildlife is scarce, although healthy carriage or infections have been reported in red squirrels (exudative dermatitis),56 black bear (endocarditis),57 zebra (cutaneous granuloma),58 raccoon,59 and dolphins (pyogenic meningoencephalitis).60
Prevalence of MRSA in companion animals; reservoirs
Identification of MRSA in companion and livestock animals has expanded the interest in understanding the genetic evolution and adaptation of such strains in different hosts. With molecular analyses, it is proven that the bacterium always evolves, encountering environmental conditions, response to selective pressure from host-specific environment, competition with microorganisms of the host’s normal flora, and antimicrobial usage, which varies among host species.18 Knowledge of MRSA host adaptation and animal reservoirs is of importance for the implementation of control practices not only in hospital and veterinary settings but also in the household. The host adaptation ability has been proven for MRSA CC133 and CC398 that include the human lineage with immune evasion cluster genes such as chp, sak, and scn carried on ΦSa3 bacteriophage, and the animal lineage carrying vwb gene.18,37,38 Human lineages are isolated from people without any contact with farm animals, whereas farmers and veterinarians may be colonized by lineages of animal origin (Table 2).18 Such an event has not been as yet characterized among MRSA clones from companion animals. With today’s expansion of companion animal market, besides dogs, cats, horses, as well as small exotic animals that live in households should be included.61 In USA, it is estimated that 62% of the population lives with a pet.61 The majority host dogs, followed by cats, birds, reptiles, or other small animals.61 Moreover, zoo parks and aquariums are included in the same group, since several animal species are in close contact with humans.
MRSA studies in animals have proved that nares, mouth, and perineum constitute the major sites of colonization,62 whereas the belly area that is often in close contact with humans is usually colonized with staphylococci.63 Misic et al have investigated the microbiota flora from sites that usually are colonized by S. aureus among companion animals and their household contacts.62 S. aureus and MRSA carriage in cats were not associated with changes in the rest microbial community, whereas in dogs, carriage was associated with changes of the microbiota flora.62 Moreover, pets share common microbiota flora with their households and not with other pets outside their house.62 This finding was verified by the fact that people who did not have a pet carried microbiota more similar to each other as compared to people who did have pets in their household.62
Molecular typing shows that cats and dogs are colonized or infected by HA-MRSA from humans in close contact, such as CC5, CC8, CC22, CC59, and CC239 (Table 2). In horses, MRSA lineages of animal origin are characterized: CC1, CC5, CC8, CC22, CC72, and CC398, whereas among exotic and wildlife animals, a variety of clones have been detected (Table 2).16,18,21,22,52,63–72
In a study performed in the Greater London area among healthy and veterinary-treated dogs, cats, and horses, it was shown that healthy animals carried MRSA in a percentage of 0.66%, 0.46%, and 0%, respectively, whereas the prevalence was higher among treated animals (3.23%, 2.16%, and 1.97%).64 The overall MRSA carriage was low (1.5%), and this result should be taken into consideration by veterinary personnel and household contacts. Clonal analysis showed the presence of CC22 and CC30 clones in cats and dogs and CC22, CC8/ST239, and ST398 in horses.64 Pets carried HA-MRSA clones, whereas ST398 characterization of animal origin suggests that more epidemiologic studies are needed.64
Kottler et al screened healthy pets for MRSA colonization, whereas their households were assigned into three groups: veterinary personnel, health care workers, and those without any contact with the health care system.73 No differences were found in MRSA carriage among the human groups (MRSA in total 5.63%), whereas MRSA prevalence among pets was 3.41%. Only four person–pet pairs had identical PFGE banding patterns. However, it was not apparent whether the corresponding strains originated from the pet or the owner.73
Molecular analysis of MRSA recovered from humans and animals in Portugal showed that CC5 and CC22 clones isolated from pets were the same as the human HA-MRSA and CA-MRSA lineages carrying also toxin genes, PVL included.65 Of importance is that one strain belonged to ST105-II (CC5), the same ST as a vancomycin-resistant S. aureus that had earlier been described in the country. This result poses a threat of whether pets could be a reservoir for virulent MRSA, as well as vancomycin-resistant S. aureus spread.65
Estimation of MRSA infections in companion animals was performed over a 5-year period in France. A total of 23 strains were identified (1.8%), which in their majority carried enterotoxin genes and to a lesser extend tst and leukotoxins’ genes.74 Sixteen strains were assigned to the Lyon clone, SCCmec-IV, the most prevalent human HA-MRSA one.74 It is of interest the identification of one USA300 strain in the aforementioned collection.74
Higher MRSA rates were detected in a large study performed in Germany during 2010–2012 among dogs, cats, and horses with wound infections. S. aureus was identified in 5.8%, 12.2%, and 22.8% of samples, respectively.21,22 MRSA accounted for 62.7% of canine, 46.4% of feline, and 41.3% of equine isolates, corresponding to an epidemiologic infection prevalence of 3.6%, 5.7%, and 9.4%, respectively.21,22 MRSA genotyping showed that the infection source for dogs and cats is humans, since the identified clones (CC22 and CC5) cause wound infections predominately in the country.21,22 On the contrary, clone CC398 seems to replace former CC8 among horses.21,22
In a comprehensive study from Austrian companion animals (dogs, cats, rabbits) and horses with MRSA infections, ST398-SCCmec type IVa was isolated from all species, whereas three strains carried enterotoxin genes.67 Similar results were published from the Netherlands, where outbreaks in horses with postsurgical wound infections were investigated in a veterinary hospital and animal clinics.68 The predominant MRSA clones that spread within horses and the environment were the ST398 followed by ST8.68
Morris et al, in USA, studying household contacts and their pets (dogs and cats) previously diagnosed with SSTIs, found that 11.6% of the pets were MRSA-positive.17 The decreased yield over time suggests that MRSA carriage was rather transient in pets.17 Clonal analysis showed the predominance of USA100 (HA-MRSA) and the presence of USA300 and USA400 (CA-MRSA) PFGE types. This result may be explained by the fact that USA100 is the major human nasal colonizer in USA.17 Moreover, molecular analysis could not prove a direct person to pet transmission.17 Predominance of USA100 clone (ST5-SCCmecII) was also identified in six healthy companion animals in Athens, Georgia, by Davis et al.63 Similar results were found in a multicenter study with participation of five institutions in USA, where, households and pets (dogs, cats, and reptiles) of patients with MRSA SSTIs were sampled.75 MRSA prevalence in pets was 8%, a population that may serve as a reservoir for human reinfection.75 Moreover, authors showed that mouth is the most sensitive anatomical site for MRSA colonization screening studies.75
Nasal staphylococcal colonization was performed in a veterinary clinic in Rio de Janeiro among 130 companion animals. Only one cat was found MRSA positive. The strain was classified as a PVL-negative ST30-SCCmecIV carrying enterotoxins and phenol-soluble modulins α3 genes, already characterized as the human clone causing SSTIs.66
Presence of S. aureus and MRSA was investigated also among wild animals in a pilot study conducted in Central Iowa, including 37 species. Three animals carried MRSA (2.6%) and seven MSSA (6.1%), whereas three S. aureus were PVL positive.76 Thus, it was shown that MRSA is spread in wild animals, even though more studies are needed to estimate its prevalence in this population.
Among pets and especially dogs, S. pseudintermedius predominates as a colonizer of coagulase-positive staphylococci species, as reported also from Lithuania. Among pets, 5.3% were positive for methicillin-resistant staphylococci, but none was MRSA.77 It seems that MRSA carriage is transient in pets. More long-term studies are needed among healthy and infected pets, veterinary personnel and environment, as well as household contacts in order to clarify whether pets, other household animals, humans, or the environment is the reservoir of MRSA. Studies performed in different countries have shown that among companion animals in close contact with humans, cats, and dogs carry MRSA lineages of human origin, whereas horses and pets living in farms carry MRSA of animal origin. More studies are needed to estimate MRSA epidemiology in more exotic animal species that may come in contact with humans.
Transmission risk of MRSA to humans and vice versa
Pets, including cats, dogs, horses, and exotic species, may carry MSSA and MRSA.23 Significant risk factors for MRSA infection in dogs and cats are the number of antimicrobial courses, the number of days admitted to veterinary clinics, and surgical implants.78 Although S. aureus is not the primary staphylococcal species causing SSTIs in companion animals, current literature demonstrates that domestic pets can carry or may be infected by the same MRSA epidemic strains that cause SSTIs in humans.79,80 Therefore, pets can harbor MRSA and it is conceivable that they may be a source of cross-transmission to human family members. Until now, little is known about pet owners and the role of contact with infected pets. Loeffler et al80 demonstrated for the first time an occupational risk in small animal general practitioners by MRSA carriage. In the same study, high risk groups for MRSA carriage were veterinary staff and owners of MRSA infected pets, despite having no direct hospital links. In another study, Loeffler et al failed to prove direct transmission of MRSA in a rescue kennel from a dog with wound infection to other animals.81 Even though ten carriage isolates were recovered from healthy dogs that were assigned in the same clone as the infecting one (CC22), no direct transmission was proved, but rather environmental exposure.81
Although the role of pet animals in the transmission of MRSA in the community is not well defined, pet owners and humans who work in farms and veterinary hospitals seem to be at greater risk of MRSA colonization or infection, emphasizing a possible transmission of MRSA between animals and humans.52,62,63,65,69,82 Given that the incidence of MRSA carriage in companion animals has increased substantially,69 intimate contact between pets and their owners creates favorable conditions for MRSA transmission. On the other hand, several case reports note that infections of humans persist until the pet (as well as any other colonized or infected cohabitant) is treated with antimicrobials to which bacteria are susceptible.50,51,83,84 This is reinforced by the finding that pets may be colonized with the same strains causing infection to human households.52 Risk factors for MRSA infection in animals were investigated in Germany, where the number of veterinary personnel, previous antimicrobial therapy, and surgical wound infections constituted statistically significant factors, a result comparable to similar factors assigned for human MRSA infections.21,22 There are increasing reports suggesting that pet animals may play a role in household MRSA transmission; it is not clear whether pets are colonized when owners are infected or whether the pet is exposed to MRSA from the colonized owner or a contaminated environment. Thus, it is obvious that companion animals represent important targets for intervention to curb further transmission of MRSA in the household and the community.85 It is notable that, in addition to contact with veterinary clinics, surgery and antimicrobial use, contact with children and licking behaviors constitute another risk factor for pet colonization.86 Interaction between children and pets within households includes direct face-to-face contact through licking or biting behavior or indirect contact through shared objects.87
In a multicenter study from People’s Republic of China involving six participating veterinary hospitals, a large collection of nasal swabs from healthy and infected pets, as well as, hospitals’ personnel was processed.70 A total of 22 MRSA strains were identified (prevalence in pets 1.1%), the majority being classified into ST59-SCCmecIV clone related to the human one, whereas one strain belonged to ST398-SCCmecV previously detected also in humans. Moreover, one MRSA strain from a veterinarian and two from dogs hospitalized in the same hospital were genetically identical (ST239-SCCmecIII), suggesting in-hospital transmission.70
Risk factors for MRSA carriage were investigated among veterinarians and veterinary technicians in a small animal clinic in Japan. Veterinarians (MRSA prevalence 22.9%) and sex (male, MRSA prevalence 29.2%) were risk factors for MRSA colonization.88 Even though contact with MRSA-infected pets was reported, no association of this factor with MRSA carriage in the personnel was statistically proved.88 Potential transmission of MRSA among veterinary staff, environment, and dogs was detected by molecular analysis of recovered strains in Malaysia, a country with high MRSA prevalence.71
Even though pets are colonized with human MRSA lineages in most cases, in a study from Belgium including six farms with MRSA ST398-positive pigs, cluster analysis and similarity % showed that MRSA ST398 of animal origin was disseminated among humans, environment, pets, and rodents, suggesting animal-to-animal transmission with environment playing also a role in this route.19
Pets other than dogs and cats may also be important factors in transmission. MRSA has been identified in parrots and other birds, hamsters and small ruminants, iguanas, and turtle. Even though these animals have less frequent contact with humans as compared to cats and dogs and transmit MRSA to a lesser extent, a parrot, a rabbit, a guinea pig, a bat, and a turtle were found to be infected by the human lineage ST22-SCCmec IV in a small animal clinic in Berlin.72
Strain relatedness between staphylococcal isolates from people and animals within households tends to be similar to those between human household members. Fifty percent to 67% of households bear indistinguishable MRSA strains. Although discovery of related strains in both humans and animals suggests that transmission has occurred, it does not show the direction of movement (from people to animals, vice versa, or from a common source). Moreover, some staphylococcal lineages may be better adapted to multispecies colonization than others.
Conclusion and future perspectives
MRSA is a prevalent problem in human and veterinary medicine. Veterinarians should be aware of the potential spread of MRSA in various animal species. Even though limited risk factors have been reported to be implicated in MRSA transmission between animals and humans, detailed analysis of outbreaks when occurred and the resulted control have proved that infection control practices such as the “search and destroy” policy applied in hospital settings are of the same importance in veterinary practice. The search and destroy policy consists of screening for MRSA carriage in patients, households including pets and environment, decolonization treatment, and follow-up.89 Therefore, proper antimicrobial usage, education, hand hygiene, and general screening for MRSA in veterinary personnel should be applied, particularly by introducing wound infection surveillance, decolonization, and follow-up surveillance studies. Moreover, it seems that besides companion animals and veterinarians the environment is also of great importance. Taking into account the fact that MRSA can survive on surfaces from 4 days to >51 days depending on the material tested, infection control practices should also involve environmental measures when outbreaks occur.14,89,90 Although the factors that promote person to pet cross-transmission are not as yet clearly elucidated, pet owners, especially immunocompromised patients, should also be educated regarding common sense practices to help mitigate the risk; such recommendations include both “social distancing” and hygiene practices. Other suggestions may be prevention of pets from licking the infected person or sleeping on the person’s bed, covering wounds and abrasions, avoidance of contact with exudates or excretions, daily washing of the pet’s food and water dishes, frequent laundering of pet bedding, and proper disposal of pet waste. Finally, household pets do not need to be removed if a family member acquires an MRSA infection. However, general precautions, such as good hand hygiene to limit interspecies transmission are encouraged. Additional social measures to limit contact during periods of active shedding (ie, clinical illnesses) may also be necessary.
The conclusion of this review is that certain MRSA genotypes are able to infect a specific spectrum of animals. A worldwide finding is that dogs and cats, as well as small exotic animals living in close contact with humans may be colonized or infected by human HA-MRSA or CA-MRSA clones circulating in a particular country. On the other hand, MRSA lineages of animal origin have been recovered from horses. This result is also observed in close contact farm pets, rodents, and humans. Therefore, hygiene practices are also recommended for farm animals and their close contacts.
Strategies to break the cycle of MRSA infection between animals and humans in close contact must be applied. Since all potential risk factors for MRSA transmission from animals to humans and vice versa have not been completely elucidated, broader studies in different settings and molecular analysis of recovered strains are needed in order to give an insight into MRSA evolution in animal and human environment.
Acknowledgments
We thank Professor ED Anastassiou for the critical review and corrections and Dr N Giormezis for his contribution in the format of the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
Stryjewski ME, Corey GR. Methicillin-resistant Staphylococcus aureus: an evolving pathogen. Clin Infect Dis. 2014;58(Suppl 1):S10–S19. | |
Chang S, Sievert DM, Hageman JC, et al; Vancomycin-Resistant Staphylococcus aureus Investigative Team. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N Engl J Med. 2003;348(14):1342–1347. | |
Hiramatsu K, Ito T, Tsubakishita S, et al. Genomic basis for methicillin resistance in Staphylococcus aureus. Infect Chemother. 2013; 45(2):117–136. | |
Uhlemann AC, Otto M, Lowy FD, DeLeo FR. Evolution of community- and healthcare-associated methicillin-resistant Staphylococcus aureus. Infect Genet Evol. 2014;21:563–574. | |
Dantes R, Mu Y, Belflower R, et al; Emerging Infections Program-Active Bacterial Core Surveillance MRSA Surveillance Investigators. National burden of invasive methicillin-resistant Staphylococcus aureus infections, United States, 2011. JAMA Intern Med. 2013;173(21):1970–1978. | |
Mainous AG 3rd, Hueston WJ, Everett CJ, Diaz VA. Nasal carriage of Staphylococcus aureus and methicillin-resistant S. aureus in the United States, 2001–2002. Ann Fam Med. 2006;4(2):132–137. | |
Gorwitz RJ, Kruszon-Moran D, McAllister SK, et al. Changes in the prevalence of nasal colonization with Staphylococcus aureus in the United States, 2001–2004. J Infect Dis. 2008;197(9):1226–1234. | |
Iwamoto M, Mu Y, Lynfield R, et al. Trends in invasive methicillin-resistant Staphylococcus aureus infections. Pediatrics. 2013;132(4):e817–e824. | |
Otto M. Phenol-soluble modulins. Int J Med Microbiol. 2014; 304(2):164–169. | |
Kadariya J, Smith TC, Thapaliya D. Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health. Biomed Res Int. 2014;2014:827965. | |
Aman MJ, Adhikari RP. Staphylococcal bicomponent pore-forming toxins: targets for prophylaxis and immunotherapy. Toxins. 2014; 6(3):950–972. | |
Shallcross LJ, Fragaszy E, Johnson AM, Hayward AC. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: a systematic review and meta-analysis. Lancet Infect Dis. 2013;13(1):43–54. | |
Multi Locus Sequence Typing [database on the Internet]. Available from http://saureus.mlst.net. Accessed 15th May, 2015. | |
Knox J, Uhlemann AC, Lowy FD. Staphylococcus aureus infections: transmission within households and the community. Trends Microbiol. 2015;23(7):437–444. | |
Song JH, Hsueh PR, Chung DR, et al; ANSORP Study Group. Spread of methicillin-resistant Staphylococcus aureus between the community and the hospitals in Asian countries: an ANSORP study. J Antimicrob Chemother. 2011;66(5):1061–1069. | |
Pantosti A. Methicillin-resistant Staphylococcus aureus associated with animals and its relevance to human health. Front Microbiol. 2012;3:127. | |
Morris DO, Lautenbach E, Zaoutis T, Leckerman K, Edelstein PH, Rankin SC. Potential for pet animals to harbour methicillin-resistant Staphylococcus aureus when residing with human MRSA patients. Zoonoses Public Health. 2012;59(4):286–293. | |
McCarthy AJ, Lindsay JA, Loeffler A. Are all meticillin-resistant Staphylococcus aureus (MRSA) equal in all hosts? Epidemiological and genetic comparison between animal and human MRSA. Vet Dermatol. 2012;23(4):267–275, e53–e54. | |
Pletinckx LJ, Verhegghe M, Crombé F, et al. Evidence of possible methicillin-resistant Staphylococcus aureus ST398 spread between pigs and other animals and people residing on the same farm. Prev Vet Med. 2013;109(3–4):293–303. | |
Cuny C, Friedrich A, Kozytska S, et al. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) in different animal species. Int J Med Microbiol. 2010;300(2–3):109–117. | |
Vincze S, Brandenburg AG, Espelage W, et al. Risk factors for MRSA infection in companion animals: results from a case-control study within Germany. Int J Med Microbiol. 2014;304(7):787–793. | |
Vincze S, Stamm I, Kopp PA, et al. Alarming proportions of methicillin-resistant Staphylococcus aureus (MRSA) in wound samples from companion animals, Germany 2010–2012. PLoS One. 2014;9(1):e85656. | |
Cohn LA, Middleton JR. A veterinary perspective on methicillin-resistant Staphylococci. J Vet Emerg Crit Care. 2010;20(1):31–45. | |
Faires MC, Tater KC, Weese JS. An investigation of methicillin-resistant Staphylococcus aureus colonization in people and pets in the same household with an infected person or infected pet. J Am Vet Med Assoc. 2009;235(5):540–543. | |
Walther B, Friedrich AW, Brunnberg L, Wieler LH, Lubke-Becker A. Methicillin-resistant Staphylococcus aureus (MRSA) in veterinary medicine: a “new emerging pathogen”? Berl Munch Terarztl Wochenschr. 2006;119(5–6):222–232. | |
Devriese LA, Van Damme LR, Fameree L. Methicillin (cloxacillin)-resistant Staphylococcus aureus strains isolated from bovine mastitis cases. Zentralbl Veterinarmed B. 1972;19(7):598–605. | |
Scott GM, Thomson R, Malone-Lee J, Ridgway GL. Cross-infection between animals and man: possible feline transmission of Staphylococcus aureus infection in humans? J Hosp Infect. 1988;12(1):29–34. | |
Cefai C, Ashurst S, Owens C. Human carriage of methicillin-resistant Staphylococcus aureus linked with pet dog. Lancet. 1994; 344(8921):539–540. | |
Hartmann FA, Trostle SS, Klohnen AA. Isolation of methicillin-resistant Staphylococcus aureus from a postoperative wound infection in a horse. J Am Vet Med Assoc. 1997;211(5):590–592. | |
Leonard FC, Markey BK. Meticillin-resistant Staphylococcus aureus in animals: a review. Vet J. 2008;175(1):27–36. | |
Smith TH, Fox LK, Middleton JR. Outbreak of mastitis caused by one strain of Staphylococcus aureus in a closed dairy herd. J Am Vet Med Assoc. 1998;212(4):553–556. | |
Monecke S, Kuhnert P, Hotzel H, Slickers P, Ehricht R. Microarray based study on virulence-associated genes and resistance determinants of Staphylococcus aureus isolates from cattle. Vet Microbiol. 2007; 125(1–2):128–140. | |
Wang Y, Wu CM, Lu LM, Ren GW, Cao XY, Shen JZ. Macrolide-lincosamide-resistant phenotypes and genotypes of Staphylococcus aureus isolated from bovine clinical mastitis. Vet Microbiol. 2008; 130(1–2):118–125. | |
Seguin JC, Walker RD, Caron JP, et al. Methicillin-resistant Staphylococcus aureus outbreak in a veterinary teaching hospital: potential human-to-animal transmission. J Clin Microbiol. 1999;37(5):1459–1463. | |
de Neeling AJ, van den Broek MJ, Spalburg EC, et al. High prevalence of methicillin resistant Staphylococcus aureus in pigs. Vet Microbiol. 2007;122(3–4):366–372. | |
Cuny C, Strommenger B, Witte W, Stanek C. Clusters of infections in horses with MRSA ST1, ST254, and ST398 in a veterinary hospital. Microb Drug Resist. 2008;14(4):307–310. | |
Sarrou S, Liakopoulos A, Chasioti M, et al. Dissemination of methicillin-susceptible CC398 Staphylococcus aureus strains in a rural Greek area. PLoS One. 2015;10(4):e0122761. | |
Abdelbary MM, Wittenberg A, Cuny C, et al. Phylogenetic analysis of Staphylococcus aureus CC398 reveals a sub-lineage epidemiologically associated with infections in horses. PLoS One. 2014;9(2):e88083. | |
Armand-Lefevre L, Ruimy R, Andremont A. Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg Infect Dis. 2005;11(5):711–714. | |
Khanna T, Friendship R, Dewey C, Weese JS. Methicillin resistant Staphylococcus aureus colonization in pigs and pig farmers. Vet Microbiol. 2008;128(3–4):298–303. | |
Smith TC, Male MJ, Harper AL, et al. Methicillin-resistant Staphylococcus aureus (MRSA) strain ST398 is present in midwestern US swine and swine workers. PLoS One. 2009;4(1):e4258. | |
Pomba C, Hasman H, Cavaco LM, da Fonseca JD, Aarestrup FM. First description of meticillin-resistant Staphylococcus aureus (MRSA) CC30 and CC398 from swine in Portugal. Int J Antimicrob Agents. 2009;34(2):193–194. | |
Wagenaar JA. Lifestock associated MRSA: epidemiology in animal production, transmission to humans and characterization of the strain. Tijdschr Diergeneeskd. 2009;134(24):1032–1035. | |
Tenhagen BA, Vossenkuhl B, Käsbohrer A, et al. Methicillin-resistant Staphylococcus aureus in cattle food chains – prevalence, diversity, and antimicrobial resistance in Germany. J Anim Sci. 2014; 92(6):2741–2751. | |
van de Giessen AW, van Santen-Verheuvel MG, Hengeveld PD, Bosch T, Broens EM, Reusken CB. Occurrence of methicillin-resistant Staphylococcus aureus in rats living on pig farms. Prev Vet Med. 2009; 91(2–4):270–273. | |
Bhargava K, Wang X, Donabedian S, Zervos M, de Rocha L, Zhang Y. Methicillin-resistant Staphylococcus aureus in retail meat, Detroit, Michigan, USA. Emerg Infect Dis. 2011;17(6):1135–1137. | |
Hanson BM, Dressler AE, Harper AL, et al. Prevalence of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus (MRSA) on retail meat in Iowa. J Infect Public Health. 2011;4(4):169–174. | |
Smyth DS, Feil EJ, Meaney WJ, et al. Molecular genetic typing reveals further insights into the diversity of animal-associated Staphylococcus aureus. J Med Microbiol. 2009;58(pt 10):1343–1353. | |
Monecke S, Ruppelt A, Wendlandt S, et al. Genotyping of Staphylococcus aureus isolates from diseased poultry. Vet Microbiol. 2013;162(2–4):806–812. | |
Manian FA. Asymptomatic nasal carriage of mupirocin-resistant, methicillin-resistant Staphylococcus aureus (MRSA) in a pet dog associated with MRSA infection in household contacts. Clin Infect Dis. 2003;36(2):e26–e28. | |
van Duijkeren E, Wolfhagen MJ, Box AT, Heck ME, Wannet WJ, Fluit AC. Human-to-dog transmission of methicillin-resistant Staphylococcus aureus. Emerg Infect Dis. 2004;10(12):2235–2237. | |
Ferreira JP, Anderson KL, Correa MT, et al. Transmission of MRSA between companion animals and infected human patients presenting to outpatient medical care facilities. PLoS One. 2011; 6(11):e26978. | |
Kania SA, Williamson NL, Frank LA, Wilkes RP, Jones RD, Bemis DA. Methicillin resistance of Staphylococci isolated from the skin of dogs with pyoderma. Am J Vet Res. 2004;65(9):1265–1268. | |
Jones RD, Kania SA, Rohrbach BW, Frank LA, Bemis DA. Prevalence of oxacillin- and multidrug-resistant Staphylococci in clinical samples from dogs: 1,772 samples (2001–2005). J Am Vet Med Assoc. 2007;230(2):221–227. | |
Kruse H, Hofshagen M, Thoresen SI, Bredal WP, Vollset I, Soli NE. The antimicrobial susceptibility of Staphylococcus species isolated from canine dermatitis. Vet Res Commun. 1996;20(3):205–214. | |
Simpson VR, Davison NJ, Kearns AM, et al. Association of a lukM-positive clone of Staphylococcus aureus with fatal exudative dermatitis in red squirrels (Sciurus vulgaris). Vet Microbiol. 2013; 162(2–4):987–991. | |
McBurney S, Veitch AM, Daoust PY. Bacterial valvular endocarditis in a black bear from Labrador. J Wildl Dis. 2000;36(4):788–791. | |
Pandey GS, Nomura Y, Kobayashi K, Fujise H, Yamada T. Cutaneous staphylococcal granuloma in a free living zebra (Equus burchelli) in Zambia. J Vet Med Sci. 1998;60(1):137–138. | |
Hamir AN. Systemic Staphylococcus aureus infection in a free-ranging raccoon (Procyon lotor). J Wildl Dis. 2010;46(2):665–668. | |
Colgrove GS, Migaki G. Cerebral abscess associated with stranding in a dolphin. J Wildl Dis. 1976;12(2):271–274. | |
Messenger AM, Barnes AN, Gray GC. Reverse zoonotic disease transmission (zooanthroponosis): a systematic review of seldom-documented human biological threats to animals. PLoS One. 2014;9(2):e89055. | |
Misic AM, Davis MF, Tyldsley AS, et al. The shared microbiota of humans and companion animals as evaluated from Staphylococcus carriage sites. Microbiome. 2015;3:2. | |
Davis JA, Jackson CR, Fedorka-Cray PJ, et al. Carriage of methicillin-resistant Staphylococci by healthy companion animals in the US. Lett Appl Microbiol. 2014;59(1):1–8. | |
Loeffler A, Pfeiffer DU, Lindsay JA, Soares Magalhaes RJ, Lloyd DH. Prevalence of and risk factors for MRSA carriage in companion animals: a survey of dogs, cats and horses. Epidemiol Infect. 2011;139(7):1019–1028. | |
Couto N, Belas A, Kadlec K, Schwarz S, Pomba C. Clonal diversity, virulence patterns and antimicrobial and biocide susceptibility among human, animal and environmental MRSA in Portugal. J Antimicrob Chemother. 2015;70(9):2483–2487. | |
Quitoco IM, Ramundo MS, Silva-Carvalho MC, et al. First report in South America of companion animal colonization by the USA1100 clone of community-acquired meticillin-resistant Staphylococcus aureus (ST30) and by the European clone of methicillin-resistant Staphylococcus pseudintermedius (ST71). BMC Res Notes. 2013;6:336. | |
Loncaric I, Kunzel F, Licka T, Simhofer H, Spergser J, Rosengarten R. Identification and characterization of methicillin-resistant Staphylococcus aureus (MRSA) from Austrian companion animals and horses. Vet Microbiol. 2014;168(2–4):381–387. | |
van Duijkeren E, Moleman M, Sloet van Oldruitenborgh-Oosterbaan MM, et al. Methicillin-resistant Staphylococcus aureus in horses and horse personnel: an investigation of several outbreaks. Vet Microbiol. 2010;141(1–2):96–102. | |
Loeffler A, Lloyd DH. Companion animals: a reservoir for methicillin-resistant Staphylococcus aureus in the community? Epidemiol Infect. 2010;138(5):595–605. | |
Zhang W, Hao Z, Wang Y, et al. Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and veterinary staff in China. Vet J. 2011;190(2):e125–e129. | |
Aklilu E, Zakaria Z, Hassan L, Hui Cheng C. Molecular relatedness of methicillin-resistant S. aureus isolates from staff, environment and pets at University Veterinary Hospital in Malaysia. PLoS One. 2012; 7(8):e43329. | |
Walther B, Wieler LH, Friedrich AW, et al. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from small and exotic animals at a university hospital during routine microbiological examinations. Vet Microbiol. 2008;127(1–2):171–178. | |
Kottler S, Middleton JR, Perry J, Weese JS, Cohn LA. Prevalence of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus carriage in three populations. J Vet Intern Med. 2010;24(1):132–139. | |
Haenni M, Saras E, Châtre P, et al. A USA300 variant and other human-related methicillin-resistant Staphylococcus aureus strains infecting cats and dogs in France. J Antimicrob Chemother. 2012;67(2):326–329. | |
Iverson SA, Brazil AM, Ferguson JM, et al. Anatomical patterns of colonization of pets with staphylococcal species in homes of people with methicillin-resistant Staphylococcus aureus (MRSA) skin or soft tissue infection (SSTI). Vet Microbiol. 2015;176(1–2):202–208. | |
Wardyn SE, Kauffman LK, Smith TC. Methicillin-resistant Staphylococcus aureus in central Iowa wildlife. J Wildl Dis. 2012; 48(4):1069–1073. | |
Ruzauskas M, Couto N, Kerziene S, et al. Prevalence, species distribution and antimicrobial resistance patterns of methicillin-resistant Staphylococci in Lithuanian pet animals. Acta Vet Scand. 2015;57:27. | |
Soares Magalhães RJ, Loeffler A, Lindsay J, et al. Risk factors for methicillin-resistant Staphylococcus aureus (MRSA) infection in dogs and cats: a case-control study. Vet Res. 2010;41(5):55. | |
Weese JS, Dick H, Willey BM, et al. Suspected transmission of methicillin-resistant Staphylococcus aureus between domestic pets and humans in veterinary clinics and in the household. Vet Microbiol. 2006;115(1–3):148–155. | |
Loeffler A, Pfeiffer DU, Lloyd DH, Smith H, Soares-Magalhaes R, Lindsay JA. Meticillin-resistant Staphylococcus aureus carriage in UK veterinary staff and owners of infected pets: new risk groups. J Hosp Infect. 2010;74(3):282–288. | |
Loeffler A, Pfeiffer DU, Lindsay JA, Soares-Magalhaes R, Lloyd DH. Lack of transmission of methicillin-resistant Staphylococcus aureus (MRSA) between apparently healthy dogs in a rescue kennel. Vet Microbiol. 2010;141(1–2):178–181. | |
Stein RA. Methicillin-resistant Staphylococcus aureus – the new zoonosis. Int J Infect Dis. 2009;13(3):299–301. | |
Sing A, Tuschak C, Hormansdorfer S. Methicillin-resistant Staphylococcus aureus in a family and its pet cat. N Engl J Med. 2008; 358(11):1200–1201. | |
van Duijkeren E, Wolfhagen MJ, Heck ME, Wannet WJ. Transmission of a Panton-Valentine leucocidin-positive, methicillin-resistant Staphylococcus aureus strain between humans and a dog. J Clin Microbiol. 2005;43(12):6209–6211. | |
Bender JB, Waters KC, Nerby J, Olsen KE, Jawahir S. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from pets living in households with MRSA-infected children. Clin Infect Dis. 2012; 54(3):449–450. | |
Lefebvre SL, Reid-Smith RJ, Waltner-Toews D, Weese JS. Incidence of acquisition of methicillin-resistant Staphylococcus aureus, Clostridium difficile, and other health-care-associated pathogens by dogs that participate in animal-assisted interventions. J Am Vet Med Assoc. 2009;234(11):1404–1417. | |
Oehler RL, Velez AP, Mizrachi M, Lamarche J, Gompf S. Bite-related and septic syndromes caused by cats and dogs. Lancet Infect Dis. 2009;9(7):439–447. | |
Ishihara K, Saito M, Shimokubo N, Muramatsu Y, Maetani S, Tamura Y. Epidemiological analysis of methicillin-resistant Staphylococcus aureus carriage among veterinary staff of companion animals in Japan. J Vet Med Sci. 2014;76(12):1627–1629. | |
Böcher S, Skov RL, Knudsen MA, et al. The search and destroy strategy prevents spread and long-term carriage of methicillin-resistant Staphylococcus aureus: results from the follow-up screening of a large ST22 (E-MRSA 15) outbreak in Denmark. Clin Microbiol Infect. 2010; 16(9):1427–1434. | |
Neely AN, Maley MP. Survival of enterococci and Staphylococci on hospital fabrics and plastic. J Clin Microbiol. 2000;38(2):724–726. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.