Back to Journals » Journal of Pain Research » Volume 15
Metabolomics and Microbiomics: New Potential Strategies in Chronic Pain Syndrome
Authors Piras C, Pintus BM, Noto A ![]() , Maurizio E
, Maurizio E ![]() , Fanos V, Mario M, Mussap M
, Fanos V, Mario M, Mussap M ![]() , Atzori L, Sardo S
, Atzori L, Sardo S ![]() , Gabriele F
, Gabriele F ![]()
Received 23 December 2021
Accepted for publication 2 March 2022
Published 11 March 2022 Volume 2022:15 Pages 723—731
DOI https://doi.org/10.2147/JPR.S354516
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Qi Fang
Cristina Piras,1 Bruno Maria Pintus,2 Antonio Noto,1 Maurizio Evangelista,3 Vassilios Fanos,4 Mario Musu,2 Michele Mussap,4 Luigi Atzori,1 Salvatore Sardo,2 Gabriele Finco2
1Department of Biomedical Sciences, University of Cagliari, Monserrato, 09042, Italy; 2Department of Medical Sciences and Public Health, University of Cagliari, Monserrato, 09042, Italy; 3Department of Anesthesiology and Pain Medicine, Cattolica University, Rome, 00168, Italy; 4Department of Surgical Science, University of Cagliari, Monserrato, 09042, Italy
Correspondence: Antonio Noto, Email [email protected]
Abstract: Chronic pain affects almost 20% of the European adult population and it significantly reduces patients’ quality of life. Chronic pain is considered a multidimensional experience determined by the interaction of several genetic and environmental factors. The effect of specific genetic contributions is often unclear, and the interpretation of the results from studies focused on genetic influences on pain has been complicated by the existence of multiple pain phenotypes. A step forward from genetics could be given by the application of metabolomics and microbiomics tools. Metabolomics is a powerful approach for hypothesis generation in biology, and it aims to analyze low molecular weight compounds, either metabolic intermediates or metabolic end-products, resulting from human or microbial metabolism. Microbiomics is a fast-growing field in which all the microbes are examined together, and as a result, its perturbation may indicate the development of chronic diseases. By applying these methodologies for the study of chronic pain, several differences have been identified. The alteration of the choline-PAF pathway is an intriguing finding recognized by several groups. In our opinion, metabolomics and microbiomics techniques will allow significant progress into the medical field. Patients may benefit from the possibility of being stratified and classified based on their metabolic and microbial profile, which, in the next future, may lead to personalized therapy.
Keywords: pain, chronic pain, metabolomics, microbiomics, biomarkers
Definition and Classification of Pain
Pain has always been part of the human experience despite not being recognized by the scientific world apart from the recent past. During the evolution of superior organisms, pain had a fundamental role as a physiological message to undertake a reaction for maintaining physical integrity; however, when it loses this protective function, it leads to maladaptive responses that ultimately damage the host, a pathological condition named pain syndrome.1 Its definition dates back to 1979 when the International Association for the Study of Pain (IASP) defined pain as “unpleasant sensory and emotional experience associated with a real or a potential tissue damage” that leads to a diminished quality of life by producing a negative emotional state.2,3 A slight revision was carried out in 2018 by the IASP that formed a task force to evaluate this classification, which became “An unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage”.4
By definition, pain is categorized into two main types: nociceptive and neuropathic. Nociceptive pain occurs when the somatic or visceral tissue is threatened or damaged by noxious stimuli through the activation of specific pain receptors called nociceptors. Neuropathic pain is caused by structural damage and dysfunction affecting the somatosensory neural pathways in the peripheral or central nervous system (CNS).5 Pain is further classified according to its duration: acute pain has a clear temporal and qualitative relationship with the noxious stimulus, it is perceived immediately after an injury, while chronic pain is continuous or recurrent and persists beyond the average expected period of healing, and for classification purposes, this time window has been set at three months.6 Chronic pain is considered a multidimensional experience determined by the interaction of several factors such as somatosensory, social-emotional, and motivational, whose pathophysiological mechanisms are not fully understood. The Global Burden of Disease reviews have recently named it one of the most prominent causes of disability worldwide.7 The lack of knowledge has limited the development of effective therapeutic options, constraining patients to use drug therapies characterized by significant adverse effects and, in some cases, poor therapeutic relief.
Furthermore, chronic pain is increasing proportionally with the age of subjects ranging from 14.3% in 18–25-year-olds to 62% in the over 75 age group.8 The number of subjects with chronic pain in the USA is calculated to be close to 100 million, with an annual expenditure near to $635 million.9 These numbers are likely to increase further due to aging people and better treatment of long-term conditions such as stroke and cancer.
Nowadays, it is essential to emphasize that the management of this condition is affected by at least two significant gaps of knowledge: 1) an objective measure of the intensity of pain, 2) a univocal identification of the molecular basis and pathways of chronic pain. The former is still performed by applying standard tools to interview the patient about the intensity, quality, and duration of the sensation. Everyone reacts uniquely to a given painful stimulus, based on experience and what is called his/her own “pain threshold”, and each person can assess, according to its parameter, how severe pain is affecting him or herself. Measurement tools in pain medicine range from unidimensional intensity-oriented tools to multidimensional questionnaire, such that they provide useful hints for recognizing the different pain phenotypes and assess therapy.
The most used pain scale is called Visual Analogue Scale (VAS). VAS is a one-dimensional approach that consists of a 10 cm horizontal or vertical line with the two extreme values labelled as “No pain” and “worst possible pain”. In this case, the patient is asked to place a mark on the line where the level corresponds to the perceived intensity of pain. The distance in centimeters from the lower end of the line to the patient’s sign is used as a numerical index of pain intensity.10 A more sophisticated evaluation method is the McGill Pain Questionnaire (MPQ) which represents a multidimensional feeling that includes different sensations of pain such as quantitative, qualitative, and unpleasantness of pain.11
Undoubtedly, the subjectivity of pain experience precludes the possibility of a true measure. For this reason, it is crucial to introduce and develop new methods for identifying molecular and biochemical alterations induced by pain. In this regard, genomics approaches led to the discovery of heritability features, which mainly were characteristic of rare human pain disorders.12
Chronic Pain: From Genes to Metabolites
Over 350 genes have been reported to be relevant to pain and analgesia in the last two decades.13 The most common genetic variations, which can affect several aspects of this experience, are caused by changes in a single nucleotide sequence (SNP).14,15
One of the first pain conditions described by a genetic defect was the rare mutation of the voltage-gated sodium channels (VGSC), causing “Erythromelalgia”.16 SCN9A is the gene encoding the human voltage-gated sodium channel type 9 subunit alpha, and the protein translated from the gene is called Nav1.7. Following this discovery, several mutations of Nav1.7 resulting in both exaggeration or absence of pain have been described and linked to human monogenic pain syndromes. Among them, the SNPs Arg1150Trp and rs6746030 have been associated with an enhanced excitation of dorsal root ganglia.
Similarly, variations have been found in three intron SNPs rs4820242, rs2284015, and rs2284017 of the CACNG2 gene, which encodes a type 1 transmembrane α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor (AMPA) regulatory peptide that increased susceptibility to chronic pain after nerve injury in mastectomy patients.17 Other SNPs were also associated with a significant reduction of chronic pain. For example, the minor allele of the SNP Ala1073Val; rs6795970 in SCN10A was significantly involved in visceral pain perception causing a shift in channel activation, thus reducing repetitive firing of dorsal root ganglion neurons and attenuating mechanical pain sensitivity.18 The identification of these mutations was, in principle, crucial for addressing personalized pharmacotherapy. Regrettably, the biology of pain is more complex and heterogeneous, so these findings should be considered just a small piece of the big puzzle, to which post-transcriptional, post-translational, and epigenetic modifications should be added.19 As an example, recent work by Agarwal et al showed that post-translational modification of SUMO conjugation of proteins (Covalent modification by small ubiquitin-related modifier polypeptides named SUMOylation) is also involved in chronic pain, in a particular condition named diabetic peripheral neuropathy (DPN). DPN affects approximately 21% of patients with diabetes, and authors identified a channel TRPV1, novel molecular targets of SUMOylation that critically regulated its activity. In particular, the function resulted impaired by diabetes-induced de-SUMOylation and metabolic imbalance eliciting neuropathy and sensory loss. Authors suggested that modulation of the SUMOylation status of specific molecular targets may yield new strategies for treatment and reversal of chronic pain.20 A way to bridge the gap between post-translational, epigenetic modifications and biomarkers identification is metabolomics, which provides a comprehensive snapshot of physiological processes offering insights into the regulatory networks that influence cellular processes.
For this reason, the number of studies that aimed to identify an altered metabolic profile characterizing chronic pain patients has increased significantly in the last few years.
A typical metabolic profile is achieved by measuring a panel of metabolites. Usually, several hundred, produced in response to several stimuli. The development of specialized analytical platforms such as Nuclear Magnetic Resonance (1H-NMR) spectroscopy, gas chromatography (GC), and liquid chromatography (LC) coupled with mass spectrometry (MS) are useful to provide a comprehensive metabolic fingerprint. Data analysis is also crucial to determine the number of relationships between grouping and feature variables and determine and facilitate discrimination among groups. As demonstrated by a constant increase of MEDLINE PubMed records related to the topic of metabolomics of pain in both animal and human subjects, this approach is gaining wider interest in the academic community and the most relevant findings are discussed in this review.
An intriguing study about the application of metabolomics as a tool for biomarker discovery in pain syndrome was published by Patti et al, who used the tibial nerve transection (TNT) on Sprague-Dawley rat as a model of neuropathic pain.21 TNT rat induces allodynia, a condition in which generally innocuous stimuli elicit a pain response that persists for at least nine weeks post-surgery. Tissues from dorsal horn, dorsal root ganglia (DRG), tibial nerve, and plasma samples were collected from these rats, in which the lipid content was analyzed by mass spectrometry technique. The metabolic content from TNT rats was compared with that of a sham surgery group resulting in an alteration of hundreds of molecules, 94% of which came from ipsilateral dorsal horn tissue samples. Among the important metabolites, TNT animals were characterized by upregulation of ceramide (d18:1:16:0), phosphatidylcholines, platelet activation factor (PAF), N, N-dimethyl sphingosine (DMS), and downregulation of several diacylglycerols.
The increase of PAF can be considered of extreme interest because of its role in the pathophysiology of pain. PAF is a potent phospholipid activator and mediator of inflammation, mediated through a G-protein-linked receptor named PAFr. PAFr is expressed on the surface of various cells and tissues, and the PAF/PAFr system seems to play an essential role in modulating pain signaling.21
In line with Patti G., a study published by Motoyama et al has indirectly identified PAF as a crucial pain mediator.22 The authors studied the efficacy of PAF antagonists, namely CV-309, BN 50739, and WEB 2086, administered to mice suffering from neuropathic pain caused by peripheral nerve injury. Interestingly, by administering PAF antagonists and intravenous and oral administration, the authors demonstrated a potent and long-lasting anti-allodynic effect, providing new insights into the PAF/PAFr interaction. Further support to this hypothesis was provided by our group that studied the lipid profile of patients affected by fibromyalgia, a disorder characterized by chronic pain and fatigue. By analyzing the plasma metabolomic profile, two molecules resulted significantly different between the patients and the healthy controls: 1-tetradecanoyl-sn-glycerol-3-phosphocholine [PC(14:0/0:0)] and 1-hexadecanoyl-sn-glycero-3-phosphocholine[PC(16:0/0:0)].23 Interestingly, these metabolites had a chemical structure very similar to the physiological PAF, holding therefore high probability to interact with PAFr and a role in pain perception. Another piece supporting this theory has been highlighted by Morita et al, who performed an intrathecal administration of PAF in mice producing neuropathic pain characterized by severe tactile allodynia.24,25 In addition, in our last experiment conducted on plasma samples collected from fibromyalgia patients, we identified the molecule choline, which was also identified in our first experiment conducted in patients affected by nociceptive or neuropathic pain.26,27 Considering all these results together, it seems that the choline-PAF pathway is highly involved in the activation and propagation of pain, and in our opinion, deserves a more in-depth study. A simplified schematic representation of this hypothesis is indicated in Figure 1.
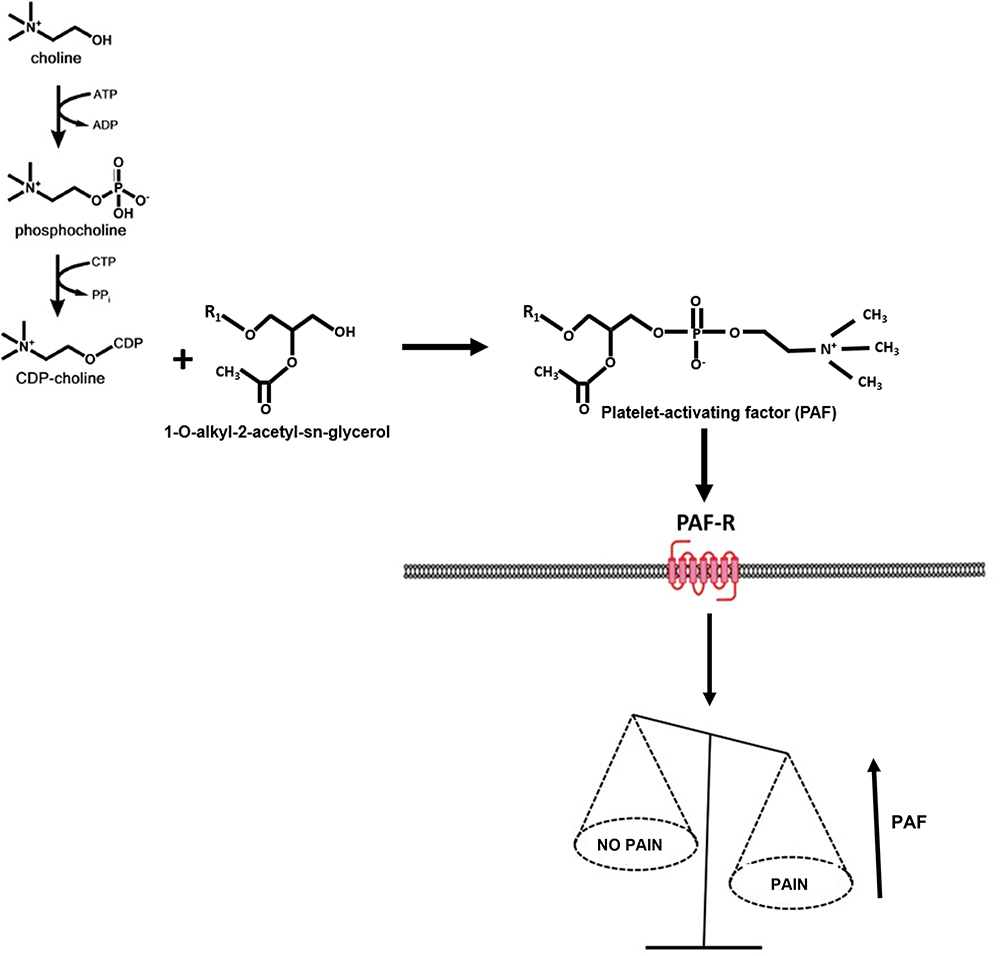 |
Figure 1 A simplified schematic representation of the hypothesized Platelet-activating factor (PAF) pathway in chronic pain syndrome. |
Other researchers focused their attention on the polar metabolites that also appeared to play a role in chronic pain. Among them, Fais et al studied 22 subjects with fibromyalgia compared to a controls group.28 These patients displayed high levels of inosine, hypoxanthine, xanthine, and a low level of adenosine. In agreement with Fais et al was the study performed by Sawynok et al, who showed that fibromyalgia patients had an excessive conversion of adenosine to inosine.29 A shared explanation was the increase of the enzyme adenosine deaminase, which led to an impairment of the purine pathway in chronic pain patients.
A Cutting-Edge Topic for Chronic Pain Research: The Gut Microbiota
A new area of research focused on the analysis of the microbiome (genome) or/and the microbiota (microorganisms) is emerging in the medical field, being driven by an interest on the way bacteria and host interact in health and diseases. To date, only a tiny percentage of the bacteria that comprise the human microbiome have been isolated, identified, and studied. However, recent technological advances have made it feasible to be entirely analyzed.
The application of cutting-edge technologies, such as metagenomics, and metabolomics, have provided significant insight into the complexity of relationships between the microbiome and the host. Identifying microbial homeostasis and/or its alteration, named eubiosis/dysbiosis, is considered the new Rosetta Stone of medicine since it may help to unlock the obscure signs of several diseases. In particular, the role of bacteria seems to be essential for the exacerbation of many neurological diseases, including Alzheimer’s disease, Parkinson’s disease, autism spectrum disorder, traumatic brain injury, and chronic pain syndromes.30–34
Although, the study of microbiota has been relegated for many years to an inconspicuous role and an uncertain meaning, nowadays, it is one of the most promising fields of medicine. There are about 100 trillion microbes that make up the human microbiota, a number that is ten times higher than that of eukaryotic cells of the whole body, with an estimated weight of about 1.5–2 kg.35,36
Considering pain syndromes, some studies have examined the fecal content of microorganisms, while others were focused on the microbial molecules released by microorganisms in the circulatory system and eventually eliminated by the kidney. Malatji et al performed urine metabolomics analysis in 18 fibromyalgia patients compared to a control group. Hippuric, succinic, and lactic acids resulted significantly increased in those patients suggesting a metabolic shift towards anaplerosis and alteration of the microbiota.37,38
Fibromyalgia was investigated by Clos-Garcia et al, who studied the serum metabolome and the fecal microbiome of 105 fibromyalgia patients and 54 healthy controls.39 The serum analysis revealed elevated levels of glutamate, while the microbiome composition resulted in a reduction in bacteria from Bifidobacterium and Lactobacillus genera, which transform glutamate into GABA. According to the authors, this could explain the elevation of glutamate serum levels in fibromyalgia.
Likewise, Minerbi et al studied the alterations of the microbial content of fibromyalgia patients compared to a control group by using 16S rRNA gene amplification, whole-genome sequencing, and targeted metabolomics.40 By using different variance abundance analyses, significant differences were identified in 19 bacterial taxa. Faecalibacterium prausnitzii and Bacteroides uniformis were found in lower relative abundance in fibromyalgia patients, whereas higher relative abundance was observed for Intestinimonas butyriciproducens, Flavonifractor plautii, Butyricoccus desmolans, Eisenbergiella tayi, and Eisenbergiella massiliensis. In particular, authors focused their attention on the role of Faecalibacterium prausnitzii, which had a lower abundance than healthy controls. The authors hypothesized that since they were less represented, the production of the metabolite butyrate, a well-known anti-inflammatory agent, was also reduced, leading to a pro-inflammatory and pro-nociceptive effect. According to the authors, these results demonstrated a dysbiotic profile in these patients that might be related to the chronic pain syndrome.
It is essential to highlight how several authors reported the role of some small microbial metabolites, including acetate, butyrate, and propionate (SCFAs), showing immunomodulatory and anti-inflammatory properties in the intestine with beneficial effects in diseases such as chronic pain, neurodegenerative diseases, and depression.41–44 These metabolites are produced by the microbiota and, due to their potential anti-inflammatory effects, appear to play an important role in the pathogenesis of chronic pain. It has been reported that SCFAs may affect the central nervous system by modulating energy metabolism, neuroinflammation, and the blood-brain barrier. Different receptors for SCFA have been discovered in different types of cells, including neuronal ones.45
Tryptophan is also considered an important molecule in pain syndrome being a precursor of the neuromodulator serotonin.46 It is an essential amino acid yielded through dietary supplementation, its deficiency has been considered the major mechanism underlying the comorbidity of chronic pain.47,48 Its effect is modulated by the gut microflora, in turn, producing tryptophan derivatives that seem to have a role in modulating inflammation, neurotransmission, and immune response and therefore, promoting the progression of pain. A possible explanation regards the ability of gut bacteria to metabolize tryptophan into indole and its derivatives, which are ligands for AhR receptor playing an important role in maintaining intestinal homeostasis by the modulation of barrier integrity and inflammatory response.49 A recent study has shown that oral supplementation of tryptophan or AhR ligand-producing Lactobacillus spp. (eubiosis) can ameliorate colitis symptoms, suggesting that tryptophan metabolites have an important role in mucosal immune homeostasis and chronic pain.
An intriguing topic is the modulation of microbiota, which may help restore or maintain a favorable balance in the ecosystem, improving human health. In particular, probiotics that are selected live strains microorganisms that confer a health benefit on the host might influence the systemic health. For example, a recent study has demonstrated that using a specific probiotic strain was able to ameliorate visceral pain. This study was carried out by Savino et al, who performed a randomized, blinded, placebo-controlled clinical trial to study intestinal colic of infants.50 The enrolled children were divided into two groups. The intervention group was treated with the probiotic Lactobacillus Reuteri DSM 17938. Urine samples were collected from each subject before starting the supplementation and at the end of the study period. The metabolomics analysis indicated that the Lactobacillus reuteri group had a different pattern of metabolites for the class of nutrients (sugars, amino acids, carboxylic acids) that resulted in more abundance after the study period with a significant reduction of pain. In light of these promising results, and although the mechanisms underlying the effects of the microbiota modulation on visceral pain perception remain to be elucidated, authors suggested expanding the cohort and including other kinds of pain syndrome.
A possible explanation regards the bidirectional communication between the gut and the brain named the gut-brain axis (GBA) which it is thought to regulate the brain homeostasis including emotional functions. In particular, microbial metabolites serve as messengers capable of crossing the brain membrane and acting as neuromodulators, which regulate the functioning of brain circuits. Dysregulation of the gut microbiome-metabolome and the ensuing disruption of the GBA are thought to contribute to the pathogenesis of chronic pain.51
A summary of principal metabolites, their corresponding bacterial sources, and their potential role are reported in Table 1.
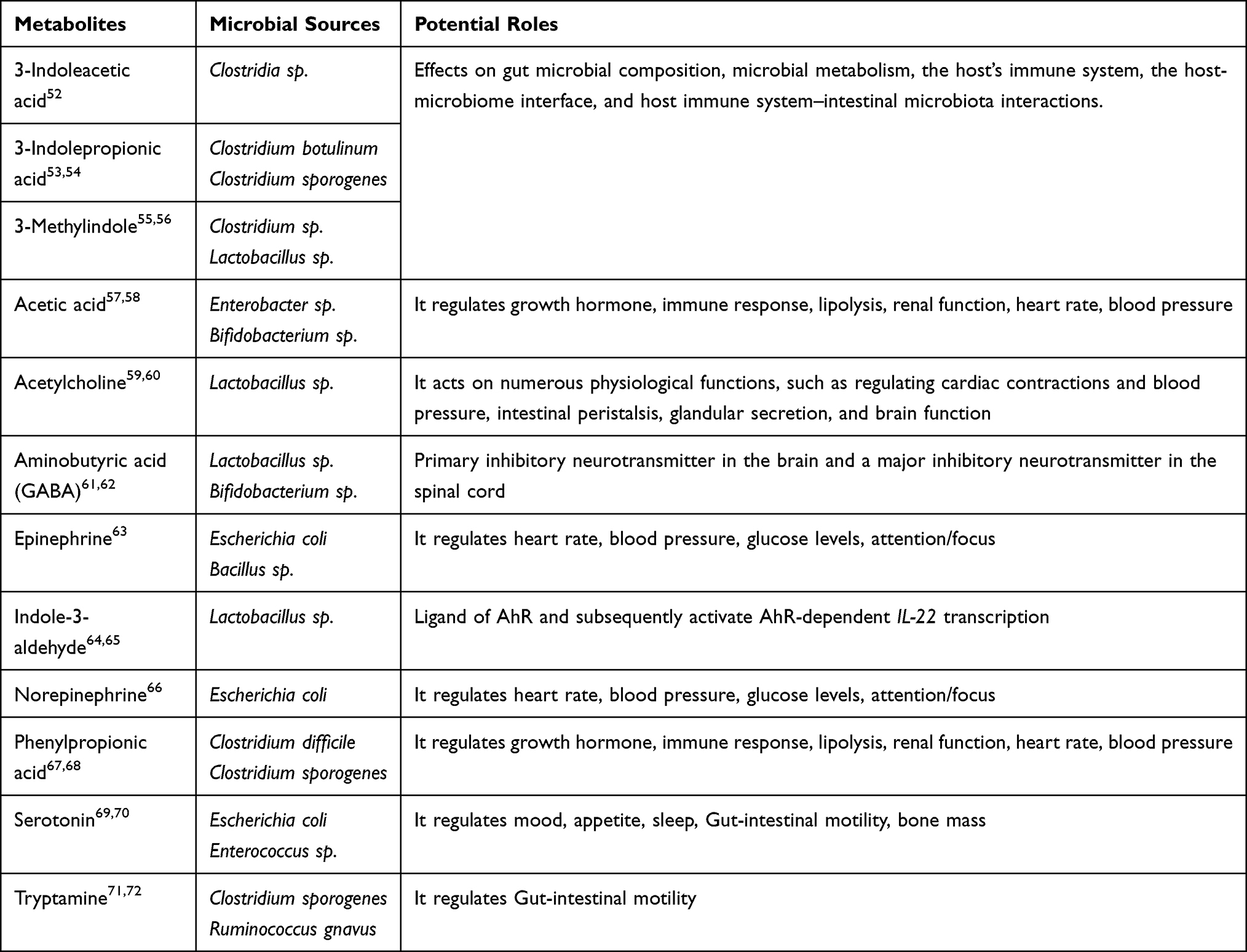 |
Table 1 Metabolites, Their Corresponding Bacterial Sources and Potential Roles |
Conclusions, Limitations and Challenge
Chronic pain management remains challenging; so far, omics technologies such as metabolomics and microbiomics offer a new way to classify chronic pain syndromes. By understanding the pathways involved in the development of chronic pain, new therapeutic targets may be discovered, and patients can be stratified based on their metabolic profile. Of course, it must be mentioned that the bench-to-bedside route of the patient in omics studies of chronic pain syndromes is still far away. The idea of performing metabolomics and microbiomics analysis in each patient is also unaffordable since these techniques are laborious and expensive, and unsuitable for screening vast populations. Of course, once a series of biomarkers are discovered and validated, these patterns of metabolites may become analytical kits routinely used for detection and quantification.
In addition, limitations of metabolomics and microbiomics should be mentioned.
Metabolomics studies are sensitive to various environmental stimuli, including diet, stress, drugs, lifestyle, and disease, therefore a particular caution during experimental design should be made.
A technical limitation of metabolomics is that a comprehensive view of all the metabolites present in a sample is still not possible due to the considerable differences in terms of concentrations, ranging between the picomolar to the micromolar.
While, concerning microbiomics, pioneering papers have shown correlations between diseases with shifts in the microbiota at the phylum level (ie, in the Firmicutes/Bacteroidetes ratio). Today, a limitation step regards the comprehension of the metabolic capacity of the intestinal microbiota, which has not been fully understood.
Of course, a definitive test for a causative effect of dysbiosis could be provided by prospective studies where microbiota composition is regularly established for all participants before the development of a certain disease.
Disclosure
The authors report no potential conflicts of interest for this work.
References
1. Todd EM, Kucharski A. Pain: historical perspectives. In: Warfield CA, Bajwa ZH, editors. Principles and Practice of Pain Medicine. McGraw-Hill; 2004:1–10.
2. Mingote S, Chuhma N, Kalmbach A, et al. Dopamine neuron dependent behaviors mediated by glutamate cotransmission. eLife. 2017;6:e27566. doi:10.7554/eLife.27566
3. Berger A, Dukes EM, Oster G. Clinical characteristics and economic costs of patients with painful neuropathic disorders. J Pain. 2004;5(3):143–149. doi:10.1016/j.jpain.2003.12.004
4. Srinivasa NR, Carr DB, Cohen M, et al. The revised international association for the study of pain definition of pain: concepts, challenges, and compromises. Pain. 2020;161(9):1976–1982. doi:10.1097/j.pain.0000000000001939
5. Scholz J, Finnerup NB, Attal N, et al. The IASP classification of chronic pain for ICD-11: chronic neuropathic pain. Pain. 2019;160(1):53–59. doi:10.1097/j.pain.0000000000001365
6. Treede RD, Rief W, Barke A, et al. Chronic pain as a symptom or a disease: the IASP classification of chronic pain for the international classification of diseases (ICD-11). Pain. 2019;160(1):19–27. doi:10.1097/j.pain.0000000000001384
7. Vos T, Flaxman AD, Naghavi M, et al. Years lived with disability (ylds) for 1160 sequelae of 289 diseases and injuries 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380(9859):2163–2196. doi:10.1016/S0140-6736(12)61729-2
8. Fayaz A, Croft P, Langford RM, Donaldson LJ, Jones GT. Prevalence of chronic pain in the UK: a systematic review and meta-analysis of population studies. BMJ Open. 2016;6(6):e010364. doi:10.1136/bmjopen-2015-010364
9. Institute of Medicine (US) Committee on Advancing Pain Research, Care, and Education. Relieving Pain in America: A Blueprint for Transforming, Prevention, Care, Education and Research. Washington (DC): National Academies Press (US); 2011. PMID: 22553896 Bookshelf ID: NBK91497. doi:10.17226/13172
10. Delgado DA, Lambert BS, Boutris N, et al. Validation of digital visual analog scale pain scoring with a traditional paper-based visual analog scale in adults. J Am Acad Orthop Surg Glob Res Rev. 2018;2(3):e088. doi:10.5435/JAAOSGlobal-D-17-00088
11. Melzack R. The McGill pain questionnaire: major properties and scoring methods. Pain. 1975;1(3):277–299. doi:10.1016/0304-3959(75)90044-5
12. Tang S, Jing H, Song F, et al. MicroRNAs in the spinal microglia serve critical roles in neuropathic pain. Mol Neurobiol. 2021;58:132–142. doi:10.1007/s12035-020-02102-1
13. Lacroix-Fralish ML, Ledoux JB, Mogil JS. The pain genes database: an interactive web browser of pain-related transgenic knockout studies. Pain. 2007;131(1–2):
14. Kuo C, Diatchenko L, Zaykin D. Discovering multilocus association with complex phenotypes. In: Belfer I, Diatchenko L, editors. Pain Genetics- Basic to Translational Science. Oxford: Wiley-Blackwell; 2013:99–114.
15. Sung YS, Ambron RT. Pathways that elicit long-term changes in gene expression in nociceptive neurons following nerve injury: contributions to neuropathic pain. Neurol Res. 2004;26(2):195–203. doi:10.1179/016164104225013761
16. Yang Y, Wang Y, Li S, et al. Mutations in SCN9A, encoding a sodium channel alpha subunit, in patients with primary erythermalgia. J Med Genet. 2004;41(3):171–174. doi:10.1136/jmg.2003.012153
17. Nissenbaum J, Devor M, Seltzer Z, et al. Susceptibility to chronic pain following nerve injury is genetically affected by CACNG2. Genome Res. 2010;20(9):1180–1190. doi:10.1101/gr.104976.110
18. Gonzalez-Lopez E, Kawasawa Y, Walter V, et al. Homozygosity for the SCN10A polymorphism rs6795970 is associated with hypoalgesic inflammatory bowel disease phenotype. Front Med. 2018;5:324. doi:10.3389/fmed.2018.00324
19. Denk F, McMahon SB. Chronic pain: emerging evidence for the involvement of epigenetics. Neuron. 2012;73(3):435–444. doi:10.1016/j.neuron.2012.01.012
20. Agarwal N, Taberner FJ, Rojas DR, et al. SUMOylation of enzymes and ion channels in sensory neurons protects against metabolic dysfunction, neuropathy, and sensory loss in diabetes. Neuron. 2020;107(6):1141–1159. doi:10.1016/j.neuron.2020.06.037
21. Patti GJ, Yanes O, Shriver LP, et al. Metabolomics implicates altered sphingolipids in chronic pain of neuropathic origin. Nat Chem Biol. 2012;8(3):232–234. doi:10.1038/nchembio.767
22. Motoyama N, Morita K, Kitayama T, et al. Pain-releasing action of platelet-activating factor (PAF) antagonists in neuropathic pain animal models and the mechanisms of action. Eur J Pain. 2013;17(8):1156–1167. doi:10.1002/j.1532-2149.2013.00289.x
23. Caboni P, Liori P, Kumar A, et al. Metabolomics analysis and modeling suggest a Lysophosphocholines-PAF Receptor interaction in Fibromyalgia. PLoS One. 2014;9(9):e107626. doi:10.1371/journal.pone.0107626
24. Morita K, Morioka N, Abdin J, Kitayama S, Nakata Y, Dohi T. Development of tactile allodynia and thermal hyperalgesia by intrathecally administered platelet-activating factor in mice. Pain. 2004;111(3):351–359. doi:10.1016/j.pain.2004.07.016
25. Morita K, Kitayama T, Morioka N, Dohi T. Glycinergic mediation of tactile allodynia induced by platelet-activating factor (PAF) through glutamateNO-cyclic GMP signalling in spinal cord in mice. Pain. 2008;138(3):525–536. doi:10.1016/j.pain.2008.01.030
26. Piras C, Conte S, Pibiri M, et al. Metabolomics and psychological features in fibromyalgia and electromagnetic sensitivity. Sci Rep. 2020;10(1):20418. doi:10.1038/s41598-020-76876-8
27. Finco G, Locci E, Mura P, et al. Can urine metabolomics be helpful in differentiating neuropathic and nociceptive pain? A proof-of-concept study. PLoS One. 2016;11(3):e0150476. doi:10.1371/journal.pone.0150476
28. Fais A, Cacace E, Corda M, et al. Purine metabolites in fibromyalgia syndrome. Clin Biochem. 2013;46:37–39. doi:10.1016/j.clinbiochem.2012.09.009
29. Sawynok J. Adenosine receptor targets for pain. Neuroscience. 2016;338:1–18. doi:10.1016/j.neuroscience.2015.10.031
30. Ochoa-Reparaz J, Mielcarz DW, Ditrio LE, et al. Role of gut commensal microflora in the development of experimental autoimmune encephalomyelitis. J Immunol. 2009;183(10):6041–6050. doi:10.4049/jimmunol.0900747
31. Noto A, Fanos V, Barberini L, et al. The urinary metabolomics profile of an Italian autistic children population and their unaffected siblings autism. J Matern Fetal Neonatal Med. 2014;2:46–52. doi:10.3109/14767058.2014.954784
32. Rumah KR, Linden J, Fischetti VA, Vartanian T. Isolation of Clostridium perfringens type B in an individual at first clinical presentation of multiple sclerosis provides clues for environmental triggers of the disease. PLoS One. 2013;8(10):e76359. doi:10.1371/journal.pone.0076359
33. Friedland RP. Mechanisms of molecular mimicry involving the microbiota in neurodegeneration. J Alzheimers Dis. 2015;45:349–362. doi:10.3233/JAD-142841
34. Tremlett H, Fadrosh DW, Faruqi AA, et al.; US Network of Pediatric MS Centers. Associations between the gut microbiota and host immune markers in pediatric multiple sclerosis and controls. BMC Neurol. 2016;16(1):182. doi:10.1186/s12883-016-0703-3
35. Marchesi JR, Adams DH, Fava F, et al. The gut microbiota and host health: a new clinical frontier. Gut. 2016;65(2):330–339. doi:10.1136/gutjnl-2015-309990
36. Zhu X, Han Y, Du J, Liu R, Jin K, Yi W. Microbiota-gut-brain axis and the central nervous system. Oncotarget. 2017;8(32):53829–53838. doi:10.18632/oncotarget.17754
37. Malatji BG, Meyer H, Mason S, et al. A diagnostic biomarker profile for fibromyalgia syndrome based on an NMR metabolomics study of selected patients and controls. BMC Neurol. 2017;17:88. doi:10.1186/s12883-017-0863-9
38. Malatji BG, Mason S, Mienie LJ, et al. The GC–MS metabolomics signature in patients with fibromyalgia syndrome directs to dysbiosis as an aspect contributing factor of FMS pathophysiology. Metabolomics. 2019;15:54. doi:10.1007/s11306-019-1513-6
39. Clos-Garcia M, Andres-Marin N, Fernandez-Eulate G, et al. Gut microbiome and serum metabolome analyses identify molecular biomarkers and altered glutamate metabolism in fibromyalgia. EBioMedicine. 2019;46:499–511. doi:10.1016/j.ebiom.2019.07.031
40. Minerbi A, Gonzalez E, Brereton NJB, et al. Altered microbiome composition in individuals with fibromyalgia. Pain. 2019;160(11):2589–2602. doi:10.1097/j.pain.0000000000001640
41. Deng FL, Pan JX, Zheng P, et al. Metabonomics reveals peripheral and central short-chain fatty acid and amino acid dysfunction in a naturally occurring depressive model of macaques. Neuropsychiatr Dis Treat. 2019;15:1077–1088. doi:10.2147/NDT.S186071
42. Unger MM, Spiegel J, Dillmann KU, et al. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Parkinsonism Relat Disord. 2016;32:66–72. doi:10.1016/j.parkreldis.2016.08.019
43. Freidin MB, Wells HRR, Potter T, Livshits G, Menni C, Williams FMK. Metabolomic markers of fatigue: association between circulating metabolome and fatigue in women with chronic widespread pain. Biochim Biophys Acta Mol Basis Dis. 2018;1864:601–606. doi:10.1016/j.bbadis.2017.11.025
44. Li S, Hua D, Wang Q, et al. The role of bacteria and its derived metabolites in chronic pain and depression: recent findings and research progress. Int J Neuropsychopharmacol. 2020;23(1):26–41. doi:10.1093/ijnp/pyz061
45. Bolognini D, Tobin AB, Milligan G, Moss CE. The pharmacology and function of receptors for short-chain fatty acids. Mol Pharmacol. 2016;89:388–398. doi:10.1124/mol.115.102301
46. Parker G, Brotchie H. Gender differences in depression. Int Rev Psychiatry. 2010;22(5):429–436. doi:10.3109/09540261.2010.492391
47. Thor KB, Kirby M, Viktrup L. Serotonin and noradrenaline involvement in urinary incontinence, depression and pain: scientific basis for overlapping clinical efficacy from a single drug, duloxetine. Int J Clin Pract. 2007;61:1349–1355. doi:10.1111/j.1742-1241.2007.01433.x
48. Benson C, Mifflin K, Kerr B, Jesudasan SJ, Dursun S, Baker G. Biogenic amines and the amino acids GABA and glutamate: relationships with pain and depression. Mod Trends Pharmacopsychiatry. 2015;30:67–79.
49. Lamas B, Natividad JM, Sokol H. Aryl hydrocarbon receptor and intestinal immunity. Mucosal Immunol. 2018;11:1024–1038. doi:10.1038/s41385-018-0019-2
50. Savino F, Fanos V, Noto A, Biggio D, Fattuoni C, Barberini L. Urinary metabolome of infants with colic treated with Lactobacillus reuteri DSM 17938: a pilot randomized trial. Minerva Pediatr. 2021. doi:10.23736/S0026-4946.20.06128-9
51. Dinan TG, Cryan JF. The microbiome-gut-brain axis in health and disease. Gastroenterol Clin North Am. 2017;46(1):77–89. doi:10.1016/j.gtc.2016.09.007
52. Dodd D, Spitzer MH, Van Treuren W, et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature. 2017;551:648–652. doi:10.1038/nature24661
53. Jellet JJ, Forrest TP, Macdonald IA, Marrie TJ, Holdeman LV. Production of indole-3-propanoic acid and 3-(p-hydroxyphenyl)propanoic acid by Clostridium sporogenes: a convenient thin-layer chromatography detection system. Can J Microbiol. 1980;26(4):448–453. doi:10.1139/m80-074
54. Qi C, Wang P, Fu T, et al. A comprehensive review for gut microbes: technologies, interventions, metabolites and diseases. Brief Funct Genomics. 2021;20(1):42–60. doi:10.1093/bfgp/elaa029
55. Ma Q, Meng N, Li Y, Wang J. Occurrence, impacts, and microbial transformation of 3-methylindole (skatole): a critical review. J Hazard Mater. 2021;416:126181. doi:10.1016/j.jhazmat.2021.126181
56. Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9:3294. doi:10.1038/s41467-018-05470-4
57. Evenepoel P, Meijers BKI, Bammens BRM, Verbeke K. Uremic toxins originating from colonic microbial metabolism. Kidney Int. 2009;114:S12–S19. doi:10.1038/ki.2009.402
58. Umena T, Shime N, Asahara T, Nomoto K, Itoi T. A pilot study of Bifidobacterium breve in neonates undergoing surgery for congenital heart disease. J Intensive Care. 2014;2:36. doi:10.1186/2052-0492-2-36
59. John SK, Chandrapragasam V, Dey P. Impact of gut microbiome lactobacillus spp. in brain function and its medicament towards Alzheimer’s disease pathogenesis. J Pure Appl Microbiol. 2021;15(3):1029–1043. doi:10.22207/JPAM.15.3.02
60. Guzman MG, Toral M, Romero M, et al. Antihypertensive effects of probiotics Lactobacillus strains in spontaneously hypertensive rats. Mol Nutr Food Res. 2015;59(11):2326–2336. doi:10.1002/mnfr.201500290
61. Kigerl KA, Mostacada K, Popovich PG. Gut microbiota are disease-modifying factors after traumatic spinal cord injury. Neurotherapeutics. 2018;15:60–67. doi:10.1007/s13311-017-0583-2
62. Moloney RD, Johnson AC, O’Mahony SM, et al. Stress and the Microbiota–Gut–Brain Axis in Visceral Pain: relevance to Irritable Bowel Syndrome. CNS Neurosci Ther. 2016;22:102–117. doi:10.1111/cns.12490
63. Wishart DS. Metabolomics for investigating physiological and pathophysiological processes. Physiol Rev. 2019;99(4):1819–1875. doi:10.1152/physrev.00035.2018
64. Zhu S, Jiang Y, Xu K, et al. The progress of gut microbiome research related to brain disorders. J Neuroinflammation. 2020;17(1):25. doi:10.1186/s12974-020-1705-z
65. Zelante T, Puccetti M, Giovagnoli S, Romani L. Regulation of host physiology and immunity by microbial indole-3-aldehyde. Curr Opin Immunol. 2021;70:27–32. doi:10.1016/j.coi.2020.12.004
66. De Weerth C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neurosci Biobehav Rev. 2017;83:458–471. doi:10.1016/j.neubiorev.2017.09.016
67. Liu Y, Hou Y, Wang G, Zheng X, Hao H. Gut microbial metabolites of aromatic amino acids as signals in host–microbe interplay. Trends Endocrinol Metab. 2020;31(11):818–834. doi:10.1016/j.tem.2020.02.012
68. Wikoff WR, Anfora AT, Liu J, et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc Natl Acad Sci USA. 2009;106(10):3698–3703. doi:10.1073/pnas.0812874106
69. De Vadder F, Grasset E, Mannerås Holm L, et al. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc Natl Acad Sci USA. 2018;115(25):6458–6463. doi:10.1073/pnas.1720017115
70. Yano JM, Yu K, Donaldson GP, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015;161(2):264–276. doi:10.1016/j.cell.2015.02.047
71. Bhattarai Y, Jie S, Linden DR, et al. Bacterially derived tryptamine increases mucus release by activating a host receptor in a mouse model of inflammatory bowel disease. iScience. 2020;23(12):101798. doi:10.1016/j.isci.2020.101798
72. Henke MT, Brown EM, Cassily CD, et al. Capsular polysaccharide correlates with immune response to the human gut microbe Ruminococcus gnavus. Proc Natl Acad Sci USA. 2021;118(20):e2007595118. doi:10.1073/pnas.2007595118
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.