")
Back to Journals » Journal of Inflammation Research » Volume 16
Metabolomic Profiling of Lungs from Mice Reveals the Variability of Metabolites in Pneumocystis Infection and the Metabolic Abnormalities in BAFF-R-Deficient Mice
Authors Rong HM, Kang HYJ, Tong ZH
Received 24 October 2022
Accepted for publication 14 March 2023
Published 27 March 2023 Volume 2023:16 Pages 1357—1373
DOI https://doi.org/10.2147/JIR.S394608
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Monika Sharma
Heng-Mo Rong,* Han-Yu-Jie Kang,* Zhao-Hui Tong
Department of Respiratory and Critical Care Medicine, Beijing Institute of Respiratory Medicine, Beijing Chao-Yang Hospital, Capital Medical University, Beijing, 100020, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhao-Hui Tong, Department of Respiratory and Critical Care Medicine, Beijing Institute of Respiratory Medicine, Beijing Chao-yang Hospital, Capital Medical University, NO. 8, Gong Ti South Road, Chao yang District, Beijing, 100020, People’s Republic of China, Tel +86 13910930309, Email [email protected]
Purpose: The incidence of Pneumocystis pneumonia (PCP) in patients without human immunodeficiency virus (HIV) has been increasing. In this study, we aimed to investigate the metabolic changes in Pneumocystis infection and the metabolic abnormalities in B-cell-activating factor receptor (BAFF-R)-deficient mice with Pneumocystis infection.
Methods: The important function of B cells during Pneumocystis infection is increasingly recognized. In this study, a Pneumocystis-infected mouse model was constructed in BAFF-R–/– mice and wild-type (WT) mice. Lungs of uninfected WT C57BL/6, WT Pneumocystis-infected, and BAFF-R–/– Pneumocystis-infected mice were used for metabolomic analyses to compare the metabolomic profiles among the groups, with the aim of exploring the metabolic influence of Pneumocystis infection and the influence of mature B-cell deficiency during infection.
Results: The results indicated that many metabolites, mainly lipids and lipid-like molecules, were dysregulated in Pneumocystis-infected WT mice compared with uninfected WT C57BL/6 mice. The data also demonstrated significant changes in tryptophan metabolism, and the expression levels of key enzymes of tryptophan metabolism, such as indoleamine 2,3-dioxygenase 1 (IDO1), were significantly upregulated. In addition, B-cell development and function might be associated with lipid metabolism. We found a lower level of alitretinoin and the abnormalities of fatty acid metabolism in BAFF-R–/– Pneumocystis-infected mice. The mRNA levels of enzymes associated with fatty acid metabolism in the lung were upregulated in BAFF-R–/– Pneumocystis-infected mice and positively correlated with the level of IL17A, thus suggesting that the abnormalities of fatty acid metabolism may be associated with greater inflammatory cell infiltration in the lung tissue of BAFF-R–/– Pneumocystis-infected mice compared with the WT Pneumocystis-infected mice.
Conclusion: Our data revealed the variability of metabolites in Pneumocystis-infected mice, suggesting that the metabolism plays a vital role in the immune response to Pneumocystis infection.
Keywords: Pneumocystis pneumonia, metabolomics, B lymphocytes, inflammation
Introduction
Pneumocystis pneumonia (PCP) is a life-threatening complication in immunosuppressed subjects without human immunodeficiency virus (HIV).1 Given the increase in organ transplantation and the widespread use of immunosuppressive drugs, the incidence of PCP in the non-HIV immunocompromised population has been rising.2 Intensive care unit (ICU) admission, mechanical ventilation rates, and mortality are higher in HIV-negative patients than in HIV-positive patients after PCP infection.3,4 Thus, it is necessary to better understand the immune mechanism of HIV-negative PCP and explore new therapeutic targets to improve the current treatment.
CD4+ T cells play a crucial role in the immune response to Pneumocystis by recruiting effector cells to the lungs.5 Previous studies have shown that the loss of CD4+ T cells makes the murine susceptible to Pneumocystis infection.6,7 Many studies have demonstrated that diverse CD4+ T-cell subsets, such as Th1 cells and Th17 cells, are induced after Pneumocystis infection.8–10 In addition, there is growing evidence that B cells are also crucial in the immune response against Pneumocystis infection.11,12 B cells are responsible for antibody production and antigen presentation.12 According to studies in murine models, anti-CD20 antibody therapy makes mice susceptible to Pneumocystis pneumonia.13 In addition to the generation of Pneumocystis-specific antibodies, B cells are also crucial for priming CD4+ T cells for the clearance of Pneumocystis.14 Overall, previous studies have indicated that B cells are both effector cells and responsible for priming CD4+ T cells.11
As for B cells, our previous studies have suggested that corticosteroid-treated hosts with PCP have apparent suppression in B cells immunity, which emphasizes the importance of B cells in the anti-Pneumocystis defense.15 In addition, we used B-cell-activating factor receptor-deficient mice (BAFF-R–/– mice) to explore the B-cell-mediated function in Pneumocystis infection. B-cell-activating factor (BAFF) —a member of the tumor necrosis factor (TNF) family—is fundamental for the survival of B cells.16 BAFF receptor (BAFF-R) is the receptor for BAFF and is the key for BAFF-mediated mature B- cell survival,17 which means that BAFF-R deficiency results in B-cell maturation disorders. Our previous research evaluated the role of BAFF-R in host defense against Pneumocystis and showed that the percentages of mature B cells, naïve B cells, and functional B cells expressing IgM and IgD antibodies in the lung and blood declined in BAFF-R–/– mice compared with wild-type (WT) mice infected with Pneumocystis at 2 weeks. BAFF-R deficiency also resulted in impaired production of Pneumocystis-specific antibodies, delayed clearance of Pneumocystis, and more significant inflammatory cell infiltration of the lungs in BAFF-R–/– mice, although no significant respiratory distress was seen in Pneumocystis-infected WT mice and BAFF-R-deficient mice, and there was no significant difference in body weight between the groups;18 however, the mechanism still needs to be further explored.
Recent studies have shown that metabolism plays a critical role in regulating the function of immune cells.19,20 In infectious diseases, metabolism is also important in the body’s immune defense to prevent pathogens. For example, a previous study showed that increased glycolysis could enhance the effector function of CD8+ T cells in HIV infection.21 In SARS-CoV-2 infection, it has been found that enhanced B-cell metabolism after illness leads to the downregulation of CD19 expression in B cells, resulting in immunodeficiency in recovered patients.22 Untargeted metabolomics is a commonly used method. This method can detect all of the measurable metabolites in a sample to help understand the current body state.23 Several recent studies have shown significant changes in serum metabolic composition in COVID-19 patients compared with healthy individuals, and these changes are associated with disease severity.24–26 However, the metabolic changes in PCP remain unclear.
In this study, we performed metabolomic analysis to explore the effects of Pneumocystis infection on metabolic profiles so as to develop new potential therapeutic targets. In addition, to further explore the role of mature B cells in Pneumocystis infection, we performed a metabolomic analysis of mouse lung tissues to compare the metabolomic profiles between WT Pneumocystis-infected mice and BAFF-R–/– Pneumocystis-infected mice so as to reveal the effects of mature B-cell deficiency on metabolism.
Materials and Methods
Mice
WT C57BL/6 mice and severe combined immunodeficient (SCID) mice were purchased from Vital River Laboratory Animal Co., Ltd. (Beijing, China). BAFF-R–/– mice on a C57BL/6 background were obtained from The Jackson Laboratory (Bar Harbor, ME, USA). All of the mice used in this study were 6-8-week-old females. They were bred in specific pathogen-free conditions. The study was approved by the Capital Medical University Animal Care and Use Committee and followed the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals and the Guidelines for Chinese Regulation for the Use and Care of Laboratory Animals.
Pneumocystis Lung Infection
Pneumocystis murina was obtained from the American Type Culture Collection and maintained in SCID mice as previously described.15,18 In brief, the lungs from SCID mice infected with Pneumocystis murina were homogenized and were pelleted at 1,000 g for 10 min at 4°C. Phosphate-buffered saline (PBS) was used to resuspend the pellet, and then the lung homogenates were stained by Diff Quick (Baxter, McGraw Park, IL). We microscopically determined the number of Pneumocystis cysts, which was adjusted to a concentration of 1×107 cysts/mL, and these were frozen to establish subsequent Pneumocystis infection models. The murine models of Pneumocystis infection were prepared by intratracheally inoculating with 1×106 Pneumocystis cysts diluted in 100 μL PBS. Control mice were intratracheally injected with lung homogenates from uninfected SCID mice.
Sample Preparation and Extraction for Untargeted Metabolomics
According to our prior study,18 the most severe lung inflammation and the highest Pneumocystis load were observed at 2 weeks after infection. Thus, mouse lung tissues were collected at 2 weeks after Pneumocystis infection and stored at −80° C until use. The lung tissues were thawed on ice. The lung tissue (50 ± 2 mg) was homogenized with 500 µL of 70% methanol at 30 Hz for 3 min. After homogenization, the mixture was shaken for 5 min and incubated on ice for 15 min. Then, the mixture was centrifuged at 12,000 rpm for 10 min at 4° C, and 400 µL of the supernatant was collected in another tube. A total of 500 μL ethyl acetate/methanol was added into the precipitate of the centrifuge tube, and the mixture was oscillated for 5 min and incubated on ice for 15 min. Next, 400 µL of the supernatant was collected after centrifugation at 12,000 rpm for 10 min at 4°C. The two supernatants were merged and concentrated. Concentration was achieved using a freeze concentrator (7310038, LABCONCO, Thermo) at 4°C and 3000 r/min overnight. Then 100 µL of 70% methanol was added, and an ultrasound was performed for 3 min. After centrifugation at 12,000 rpm for 3 min at 4°C, 60 µL of the supernatant was collected for LC-MS analysis.
HPLC Conditions (T3)
All of the sample extracts were analyzed by the LC-MS system. The analytical conditions were as follows: UPLC: column, Waters ACQUITY UPLC HSS T3 C18 (1.8 µm, 2.1 mm × 100 mm); column temperature, 40°C; flow rate, 0.4 mL/min; injection volume, 2uL; solvent system, water (0.1% formic acid): acetonitrile (0.1% formic acid); gradient program, 95:5 V/V at 0 min, 10:90 V/V at 11.0 min, 10:90 V/V at 12.0 min, 95:5 V/V at 12.1 min, and 95:5 V/V at 14.0 min.
Identification of the Metabolic Information
The raw data obtained by LC-MS analysis were first converted into mzML format by ProteoWizard software. Peak extraction, alignment, and retention time correction were performed by the XCMS program. The SVR method was used to correct the peak area and filter the peaks with a loss rate > 50% in each group of samples. Then, the metabolic identification information was acquired by searching the laboratory’s self-built database and integrating the public database and metDNA.
Real-Time PCR
RNAs were purified from the homogenized lung tissue by TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) and reverse-transcribed using the PrimeScript TM II 1st Strand cDNA Synthesis Kit (Takara Bio, Dalian, China). cDNA (1 μg) was used as template for RT-PCR, which was performed using SYBR PreMix Ex Taq TM II ROX plus (Takara) and the ABI Prism 7500 Sequence Detection System (Applied Biosystems, Foster City, CA). Gene expression was calculated using the 2−ΔΔCT method after normalization to β-actin (Actb). The Primers’ sequences are listed in Supplemental Table 1.
Statistical Analysis
Statistical analysis was performed by the R program. The statistical significance was set at p < 0.05. Univariate statistical analysis included Student’s t-test and variance multiple analysis. Multivariate statistical analysis included principal component analysis (PCA) and orthogonal partial least-squares discriminant analysis (OPLS-DA). PCA was performed by the base package in R software (version 3.5.0). OPLS-DA of metabolomic data was performed by MetaboAnalystR (version 1.0.1) in R software (version 3.5.0). The significantly different metabolites between the groups were filtered and confirmed by combining the results of the variable importance in projection (VIP) values (VIP > 1.0), t-test (P < 0.05), and fold change (FC) values (FC ≥ 2.0 or FC ≤ 0.5). The significantly different metabolites were used for pathway analysis with the Kyoto Encyclopedia of Genes and Genomes biological pathway database, and metabolic pathways with P < 0.05 were defined as being significantly enriched.
Results
Metabolic Profiling of Lung Samples
For untargeted metabolomic analysis, lung samples were obtained from seven uninfected WT mice in the control group, seven WT Pneumocystis-infected mice, and seven BAFF-R–/– Pneumocystis-infected mice. PCA was used to investigate the degree of variation between the groups in both ion modes, and a grouping trend between the groups could be observed (Figure 1). Then, OPLS-DA was performed to further explore the differences between the groups in electrospray ionization positive (ESI+) and negative (ESI-) modes. As shown in Figure 2, a clearer separation between the groups was observed.
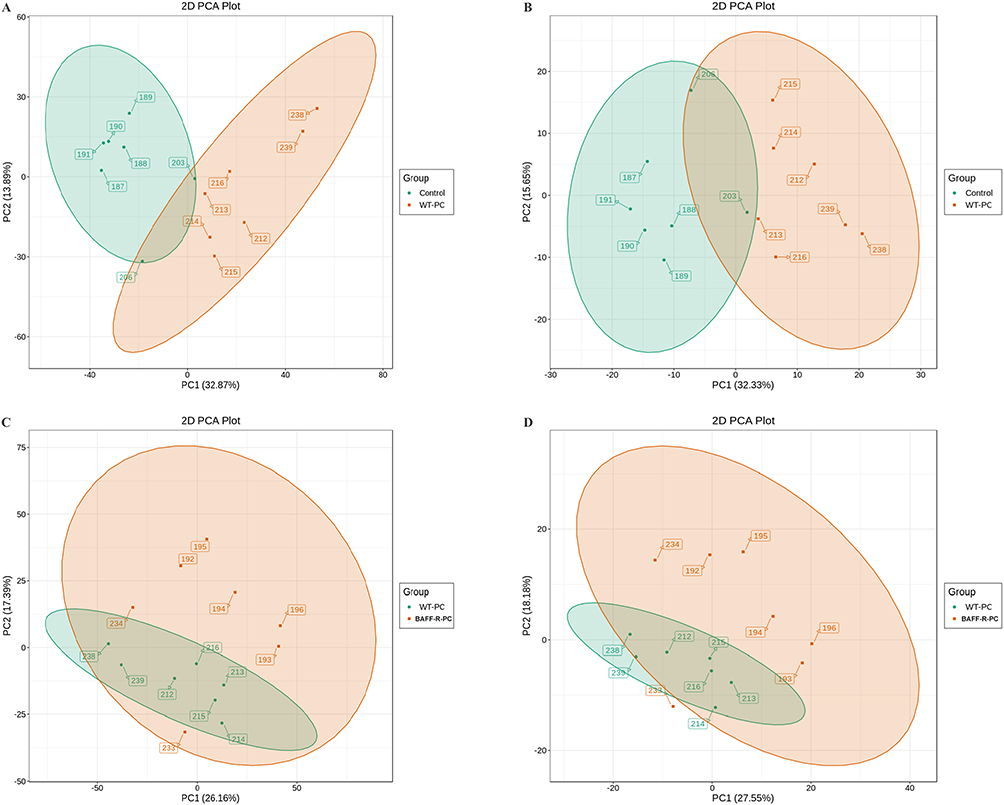 |
Figure 1 Scatter plot of scores based on the PCA model. (A) Positive ion (ESI+) scan between the control group and Pneumocystis-infected wild-type (WT) mice; (B) Negative ion (ESI−) scan between the control group and Pneumocystis-infected WT mice; (C) ESI+ scan between the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice; (D) ESI− scan between the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice. |
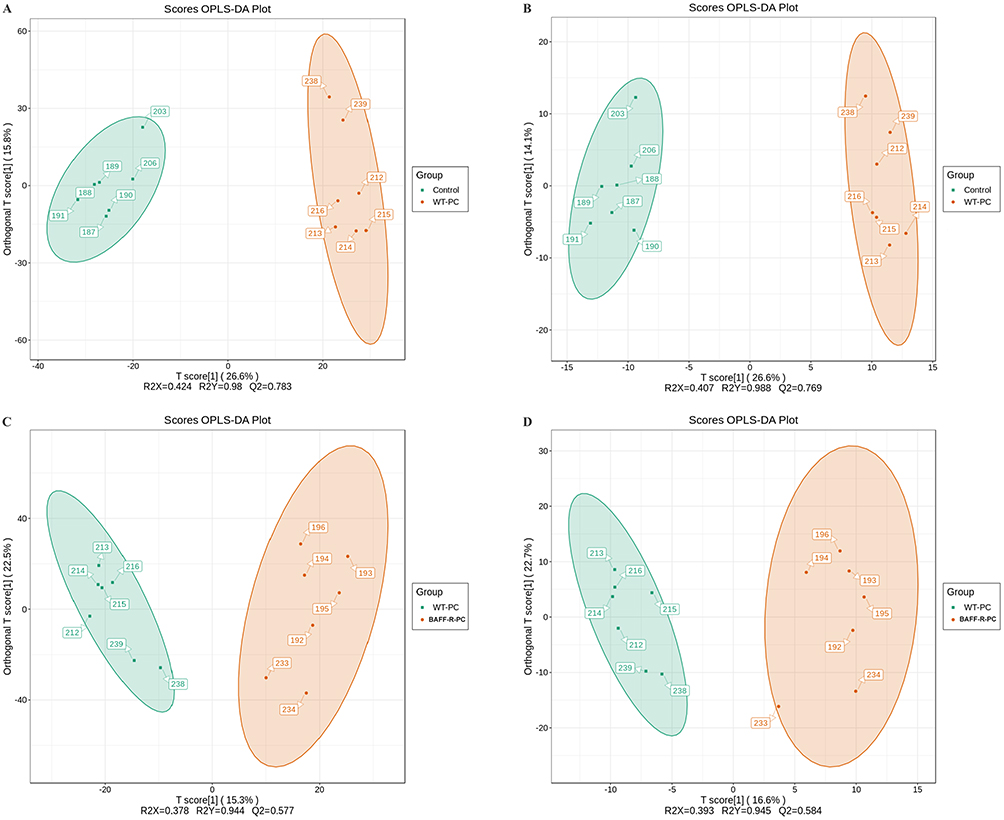 |
Figure 2 Scatter plot of scores based on the OPLS-DA model. (A) ESI+ scan between the control group and Pneumocystis-infected WT mice; (B) ESI− scan between the control group and Pneumocystis-infected WT mice; (C) ESI+ scan between the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice; (D) ESI− scan between the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice. |
A total of 302 differential metabolites in the lungs were identified in the positive ion mode between control mice and WT Pneumocystis-infected mice. The levels of 73 metabolites were higher, whereas the levels of 229 metabolites were lower in the lungs of WT Pneumocystis-infected mice compared with control mice. In the negative ion mode, we detected 66 differential metabolites between the control group and the WT Pneumocystis-infected group, of which 21 metabolites were upregulated and 45 metabolites were downregulated. The details are shown in Supplemental Tables 2, 3, and Figure 3. Heatmaps presented a different pattern of metabolites between the control and WT Pneumocystis-infected mice in both the positive and negative ion modes (Supplemental Figures 1 and 2).
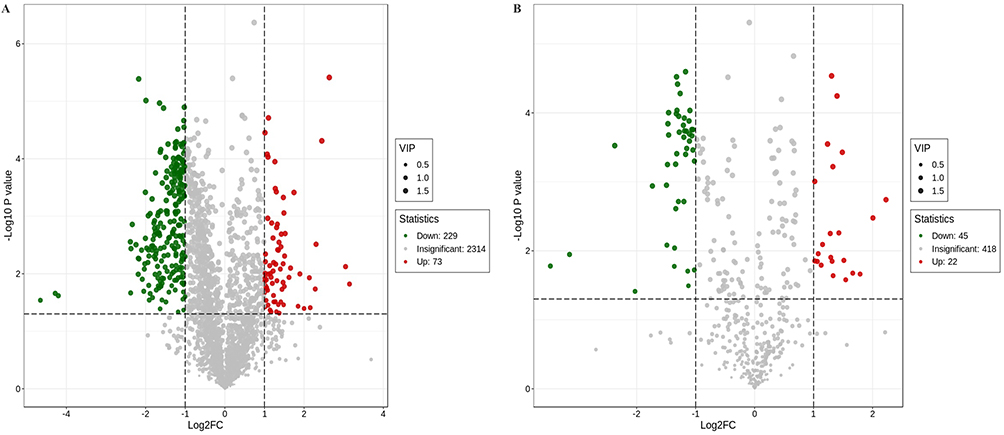 |
Figure 3 Volcano plot showing the number and distribution of lung metabolites in the control group and Pneumocystis-infected WT mice. (A) Positive and (B) negative ion modes. |
In the comparison between WT Pneumocystis-infected mice and BAFF-R–/– Pneumocystis-infected mice, 105 differential metabolites were identified in the lung samples, of which 85 were found in the positive ion mode and 23 in the negative ion mode. The results are summarized in Supplemental Tables 4 and 5. In the positive ion mode, 44 metabolites were upregulated and 41 metabolites were downregulated in the lungs of BAFF-R–/– Pneumocystis-infected mice compared with WT Pneumocystis-infected mice. In the negative ion mode, seven metabolites were upregulated and 16 metabolites were downregulated. A volcano plot is shown in Figure 4. Heatmaps showed differential metabolic profiles of lung samples between WT Pneumocystis-infected mice and BAFF-R–/– Pneumocystis-infected mice (Supplemental Figures 3 and 4).
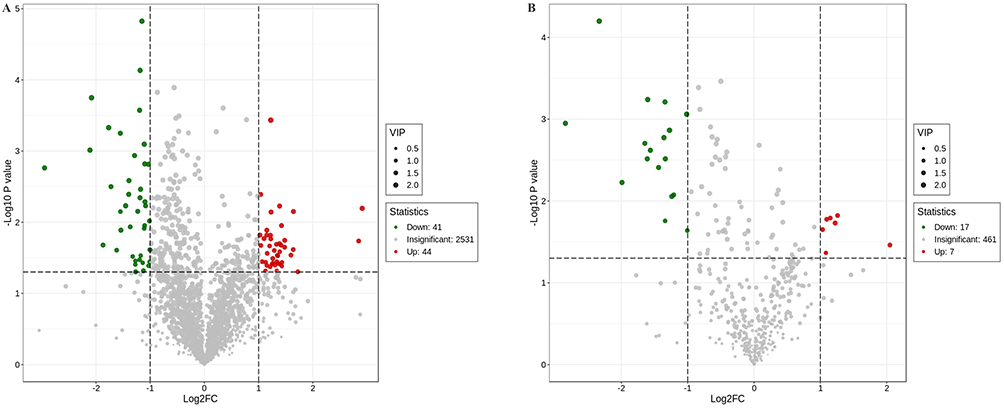 |
Figure 4 Volcano plot showing the number and distribution of lung metabolites in the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice. (A) Positive and (B) negative ion modes. |
Dysregulated Lipids and Lipid-Like Molecules and Significant Changes in Products from Tryptophan Metabolism in WT Pneumocystis-Infected Mice
The significantly dysregulated metabolites included lipids and lipid-like molecules, amino acid and its metabolomics, and organic acids and derivatives (Table 1). The findings are consistent with those of other studies on patients with ventilator-associated pneumonia (VAP) or community-acquired pneumonia (CAP).27,28 Most of the differential metabolites exhibited downregulation in WT Pneumocystis-infected mice, such as PA (21:0/i-15:0), TG (18:2 (9Z,12Z)/20:5 (5Z,8Z,11Z,14Z,17Z)/18:4 (6Z,9Z,12Z,15Z)), PA (i-21:0/a-15:0), medroxyprogesterone 17-acetate, methylprednisolone succinate, PE-NMe2 (20:5 (5Z,8Z,11Z,14Z,17Z)/22:6 (4Z,7Z,10Z,13Z,16Z,19Z)), P-aminohippuric acid, 2-amino-3-(3-methylbut-2-enylthio) propionic acid, and raltitrexed (Supplemental Figure 5). However, some lipids and lipid-like molecules, such as PC (14:0/22:2 (13Z,16Z)), DG (22:1 (13Z)/20:5 (5Z,8Z,11Z,14Z,17Z)/0:0), and cis-7,10,13,16,19-docosapentaenoic acid, were upregulated (Supplemental Figure 5) in WT Pneumocystis-infected mice. The top 20 differential metabolites between the control group and the Pneumocystis-infected WT group with the highest VIP values under ESI- and ESI+ modes are presented in Table 1.
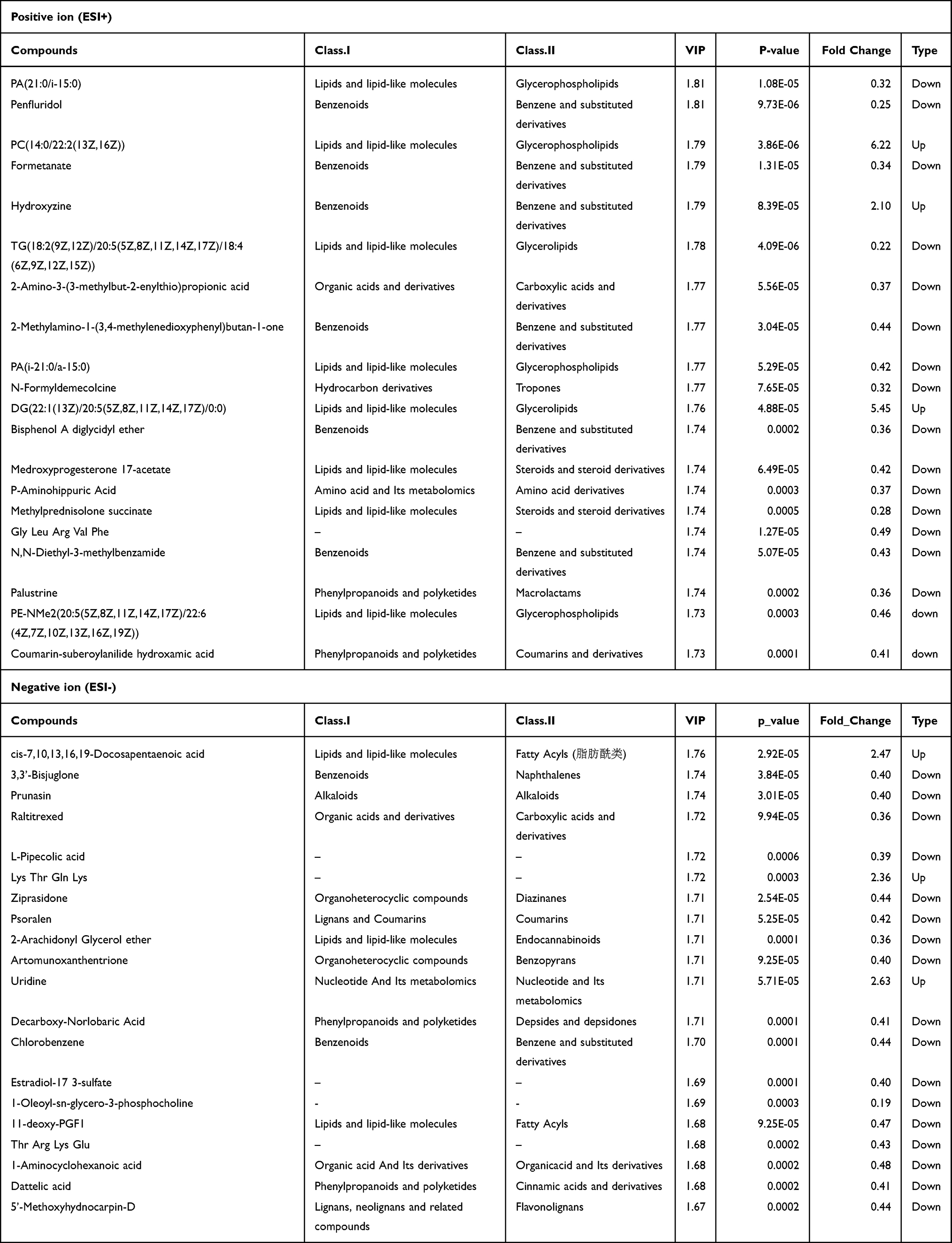 |
Table 1 Top 20 Differential Metabolites Between the Control Group and Pneumocystis-Infected WT Group with the Highest VIP Values Under ESI- and ESI+ Modes |
Based on the differential metabolites, the Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis was conducted. The path analysis results are shown in Figure 5 and Table 2, where we have selected the top 20 pathways in the positive ion mode and the negative ion mode. Previous studies have suggested that the changes in tryptophan metabolism play an important regulatory role in the host response during infection.29 Our pathway enrichment analysis also showed that the tryptophan metabolism pathway was significantly enriched. We observed that some products of tryptophan metabolism were significantly changed in the lung samples of WT Pneumocystis-infected mice, including kynurenine, 4-(2-aminophenyl)-2,4-dioxobutanoic acid, 4,6-dihydroxyquinoline, N-acetyl-5-hydroxytryptamine. The pathway map of tryptophan metabolism is shown in Supplemental Figure 6.
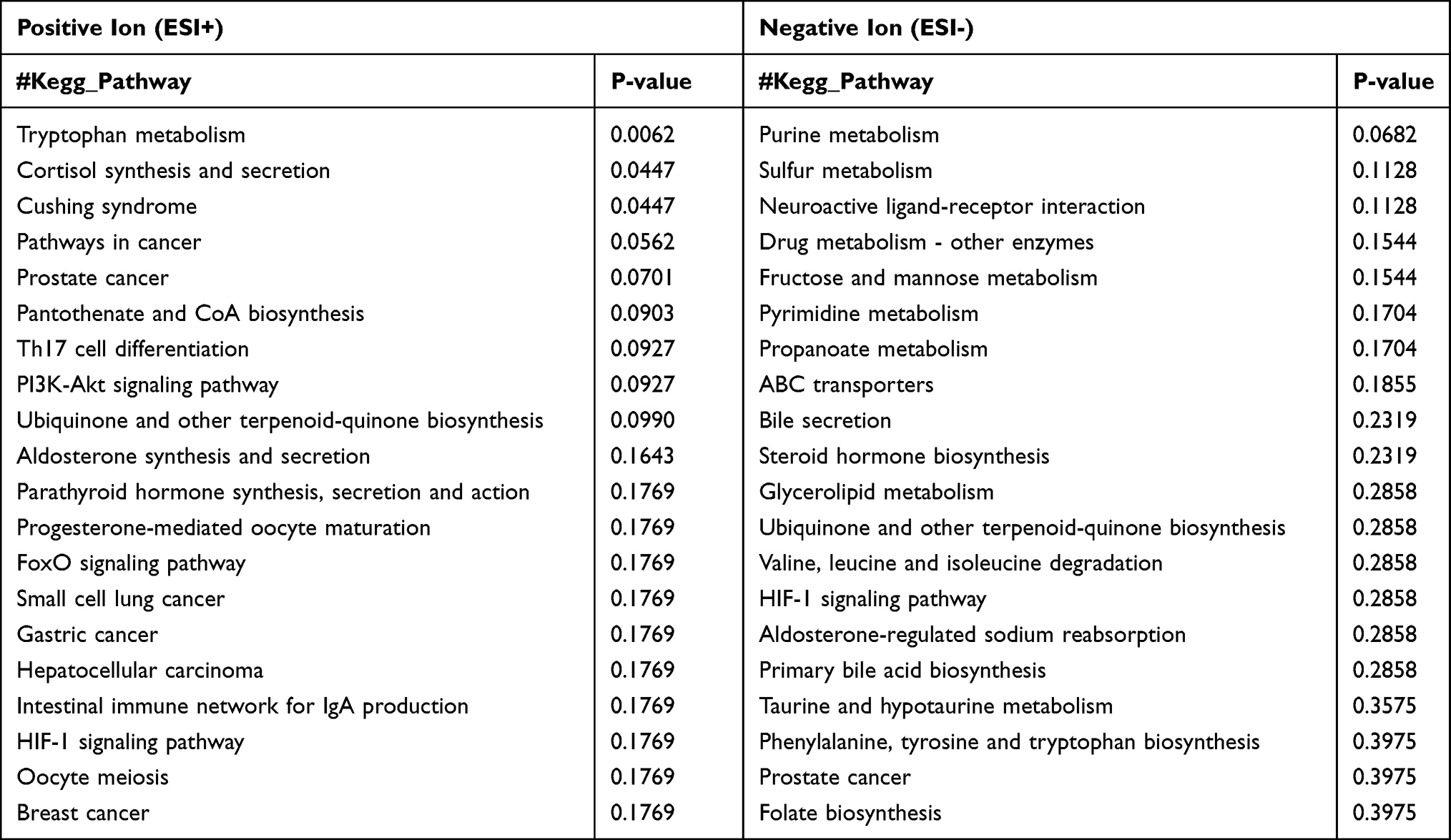 |
Table 2 Pathway Enrichment Analysis of the Identified Metabolites Showing a Difference Between the Control Group and Pneumocystis-Infected WT Group Under ESI- and ESI+ Modes |
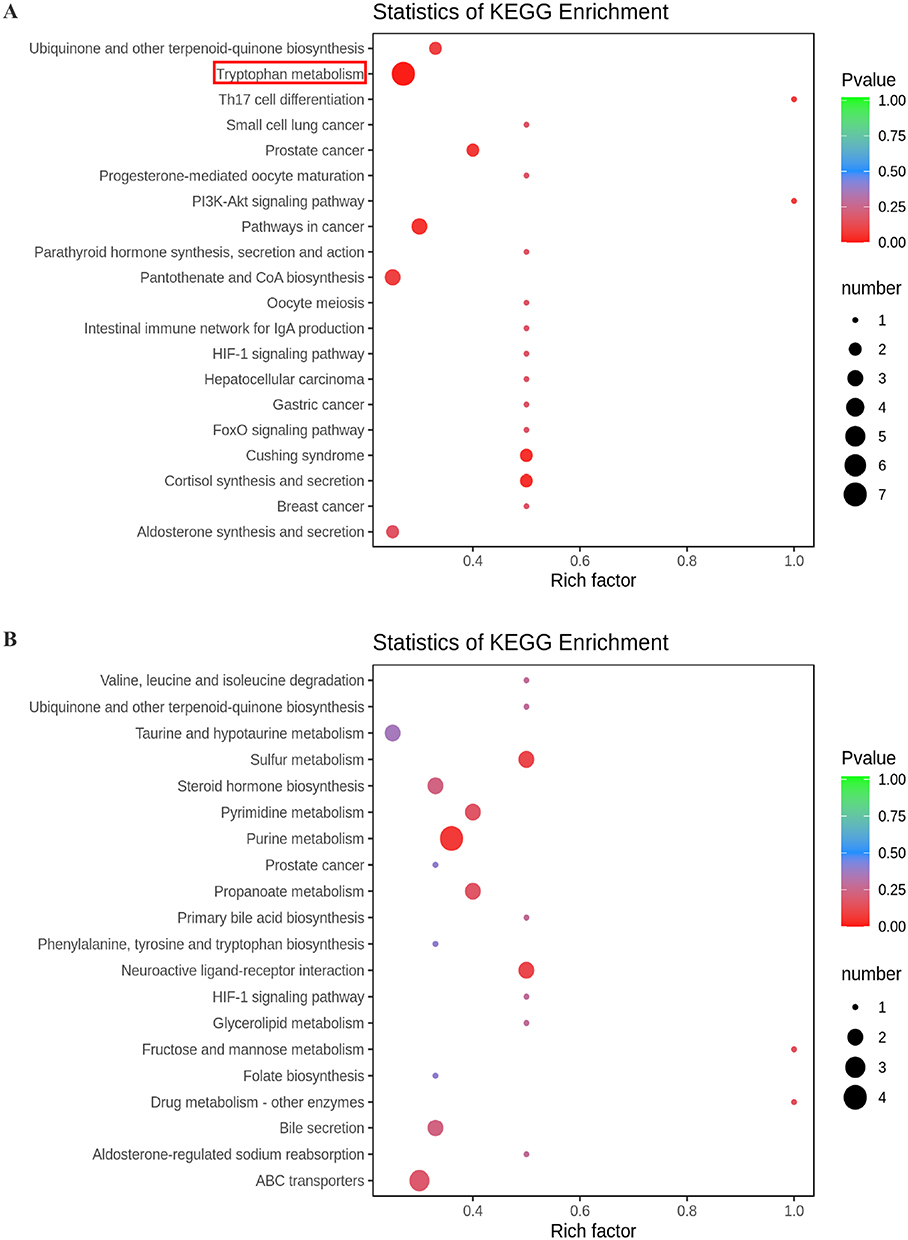 |
Figure 5 KEGG pathway enrichment analysis of differential metabolites between the control group and Pneumocystis-infected WT mice. (A) Positive and (B) negative ion modes. |
To investigate why some products’ levels of tryptophan metabolism were changed in the lung of WT Pneumocystis-infected mice, we measured the mRNA expression levels of key enzymes associated with the tryptophan metabolism in the lungs of control mice and Pneumocystis-infected WT mice. RT-PCR analysis showed that the mRNA level of indoleamine 2,3-dioxygenase 1(IDO1) and tryptophan hydroxylase (TPH) in the lungs was actually upregulated in Pneumocystis-infected WT mice (Figure 6). The results suggested that the tryptophan metabolism was activated and may play a potential role during Pneumocystis infection.
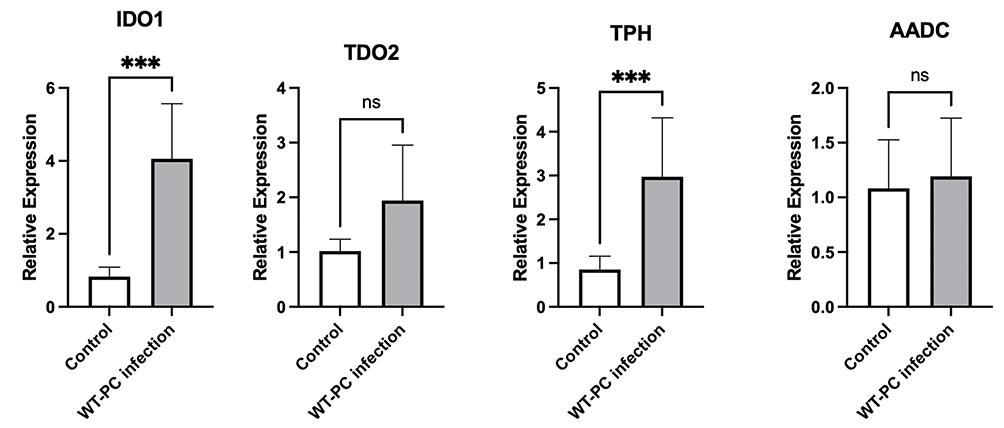 |
Figure 6 mRNA expression levels of enzymes associated with tryptophan metabolism in the lungs in the control group and Pneumocystis-infected WT mice (***P < 0.001). |
Lower Level of Alitretinoin and Abnormal Fatty Acid Metabolism in BAFF-R Gene Knockout Pneumocystis-Infected Mice
One hundred and five metabolites, mainly lipids and lipid-like molecules, were dysregulated (Supplemental Tables 4 and 5). Most of the differential metabolites of lipids and lipid-like molecules, such as farnesylthiosalicylic acid, PA (21:0/12:0), TG (14:0/18:1 (11Z)/18:1 (11Z)), PA (22:1 (13Z)/19:2 (10Z,13Z)), eicosapentaenoyl PAF C-16, PE-NMe (14:0/18:1 (11Z)), PC (18:1 (9Z)/22:2 (13Z,16Z)), alitretinoin, TG (16:0/14:1 (9Z)/20:3 (5Z,8Z,11Z)), and PE-Nme2 (15:0/16:0), were downregulation in BAFF-R–/– Pneumocystis-infected mice compared with WT Pneumocystis-infected mice (Supplemental Figure 7). The top 20 differential metabolites between the Pneumocystis-infected WT group and the Pneumocystis-infected BAFF-R–/– group with the highest VIP values under ESI- and ESI+ modes are shown in Table 3.
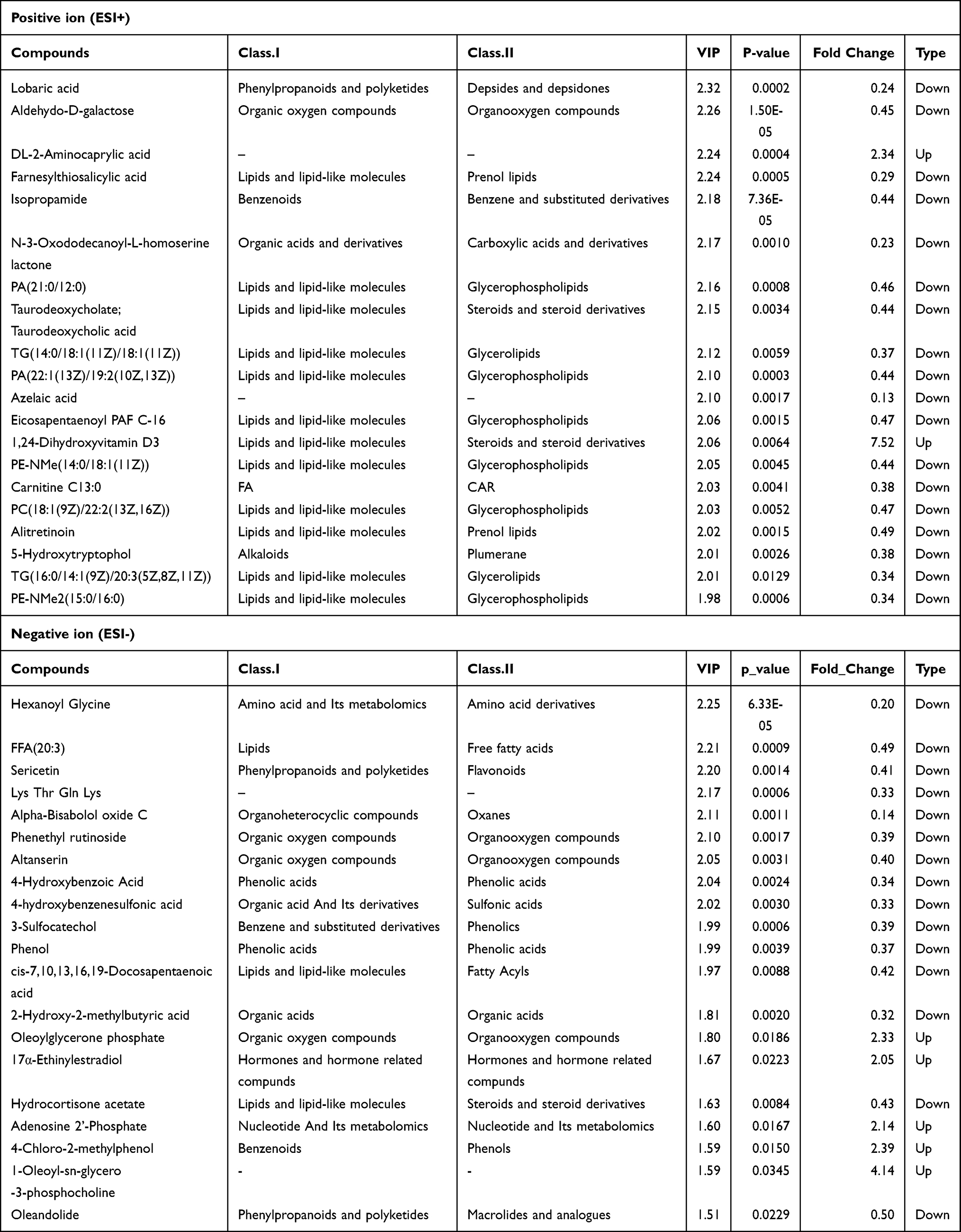 |
Table 3 Top 20 Differential Metabolites Between the Pneumocystis-Infected WT Group and Pneumocystis-Infected BAFF-R–/– Group with the Highest VIP Values Under ESI- and ESI+ Modes |
We next concentrated on metabolites and pathways involved in infection immunity and inflammation. Notably, the level of vitamin A’s metabolite, alitretinoin, was lower in the lung samples of BAFF-R–/– Pneumocystis-infected mice than in WT Pneumocystis-infected mice. The KEGG enrichment analysis results are shown in Figure 7 and Table 4. The analysis highlighted pathways related to fatty acid metabolism, such as the adipocytokine signaling pathway and fatty acid elongation. The pathway maps of the two pathways are presented in Supplemental Figures 8 and 9. Other studies have shown that fatty acid metabolism is related to infection immunity.30
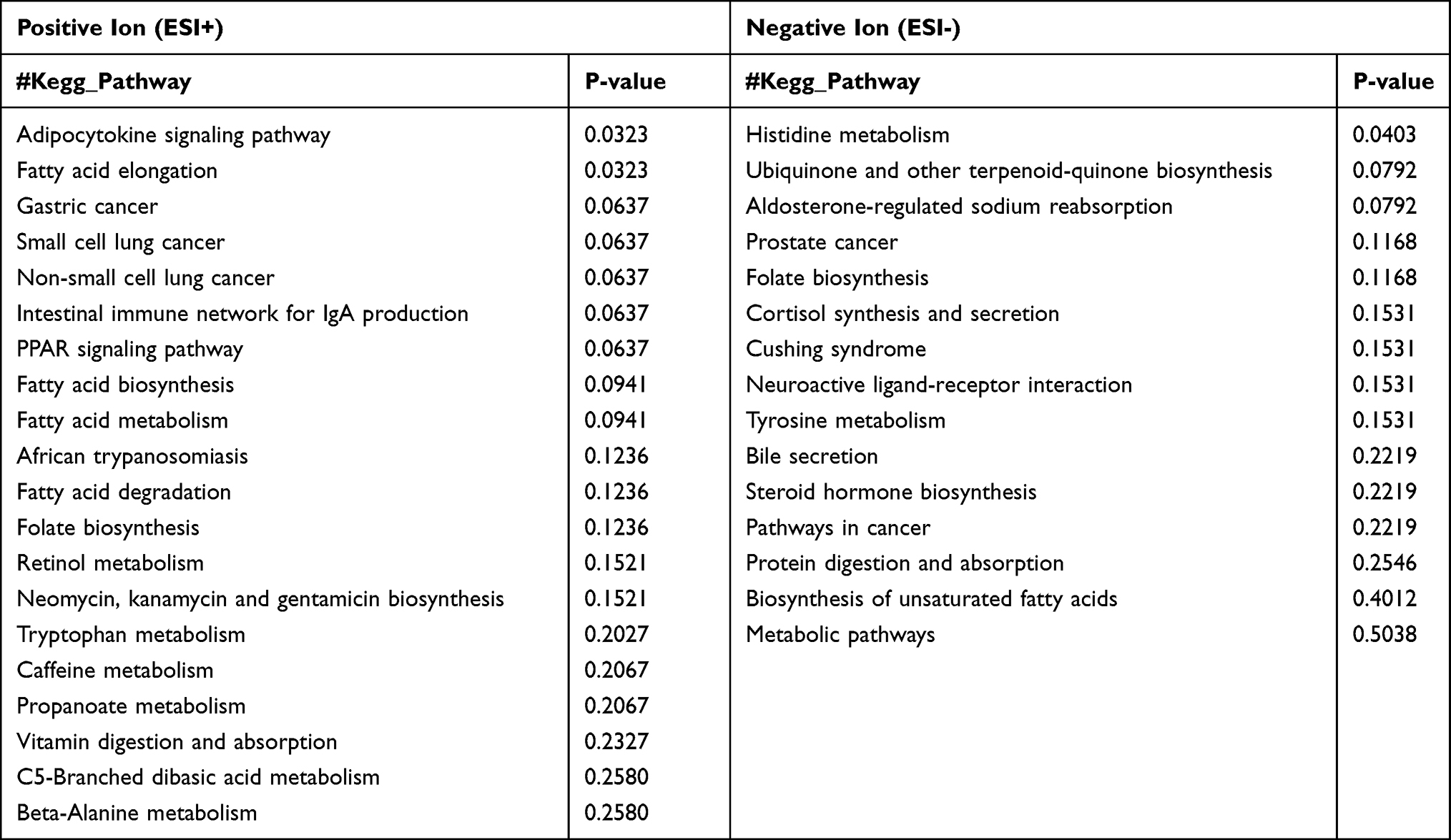 |
Table 4 Pathway Enrichment Analysis of the Identified Metabolites Showing a Difference Between the Pneumocystis-Infected WT Group and Pneumocystis-Infected BAFF-R–/– Group Under ESI- and ESI+ Modes |
 |
Figure 7 KEGG pathway enrichment analysis of differential metabolites between the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice. (A) Positive and (B) negative ion modes. |
We also measured the mRNA expression levels of enzymes associated with fatty acid metabolism in the lungs of the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice. The results of RT-PCR showed that the mRNA levels of stearoyl-CoA desaturase 1 (SCD1) and fatty acid synthase (FASN) in the lungs were significantly upregulated in BAFF-R–/– Pneumocystis-infected mice, and they both were positively correlated with the mRNA level of IL17A (Figure 8). The results revealed the metabolic dysregulation of fatty acid metabolism in BAFF-R–/– Pneumocystis-infected mice, possibly related to greater inflammatory cell infiltration in the lungs.
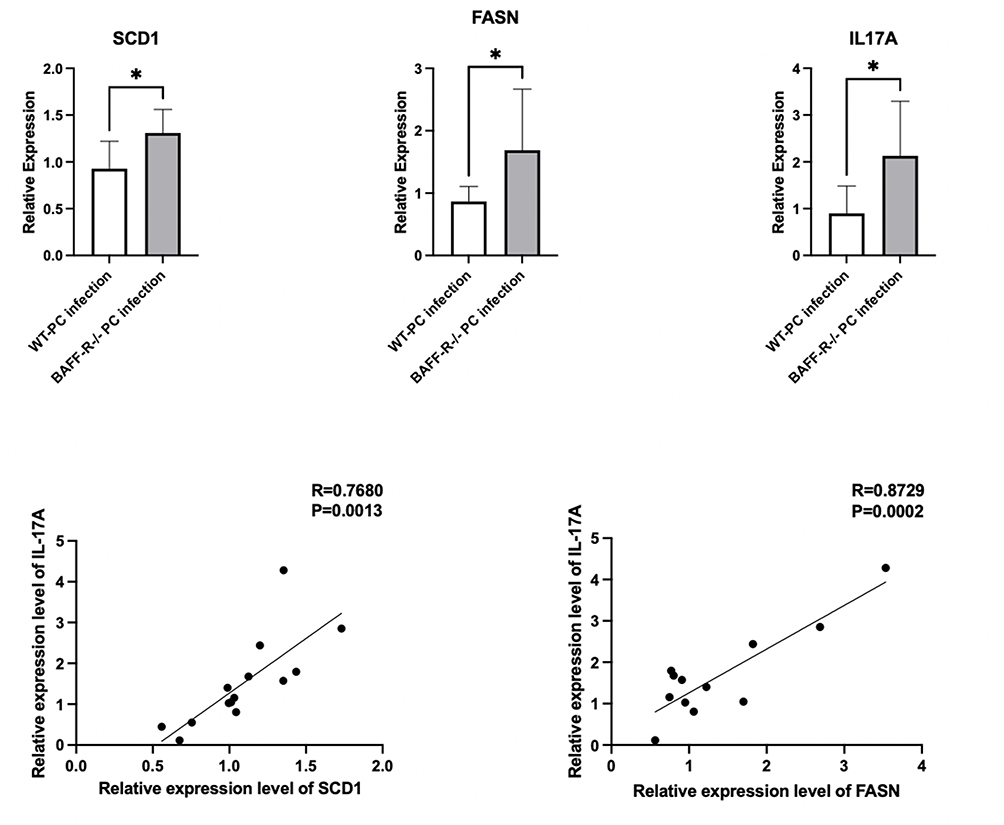 |
Figure 8 mRNA expression levels of enzymes associated with fatty acid metabolism in the lungs in the Pneumocystis-infected WT mice and Pneumocystis-infected BAFF-R–/– mice, and the correlation analysis of the mRNA expression levels of IL17A and SCD1 or FASN (*P < 0.05). |
Discussion
Changes in the metabolic profile are the body’s final response to stimuli. Untargeted metabolomics is an advanced way to screen unknown metabolite alterations. In recent years, metabolomics has been widely applied in infectious and immune diseases, and some metabolic changes have been found.31–36 However, there is a lack of metabolomic information on Pneumocystis infection.
We found that many metabolites in WT Pneumocystis-infected mice were changed compared with non-infected mice. Lipids and lipid-like molecules were the primary dysregulated metabolites, indicating that lipid metabolism may be abnormal in Pneumocystis infection. Lipids have three main functions, namely, storing energy, forming an important component of cell membranes, and acting as important signaling molecules within cells. Many recent studies have explored lipid metabolism in diseases. As for infectious diseases, some studies have found lipid metabolism dysregulation in patients with COVID-19 or Mycoplasma pneumoniae pneumonia.32,37 Our previous studies have suggested that B-cell counts are reduced after Pneumocystis infection,18 and corticosteroid treatment might suppress the B-cell immunity in the PCP hosts.15 Recent studies have shown that B-cell development and function are related to lipid metabolism.38,39 Thus, correcting lipid metabolism disorders is a possible therapeutic target that may help promote B-cell function against Pneumocystis infection.
The current study focused on enriched metabolic pathways associated with anti-infection immunity. We noted significant changes in some metabolites associated with the tryptophan metabolism pathway during Pneumocystis infection. We also found that the expression levels of key enzymes of tryptophan metabolism, such as IDO1, was significantly upregulated. Many studies have revealed the role of tryptophan metabolism in regulating immune response.40–42 In antitumor immunity, the activation of IDO1 may restrain antitumor immune responses by inhibiting T cell function.43 There is also evidence suggesting that tryptophan metabolism plays a vital role in infectious diseases.43 Activation of IDO1 in our study may lead to impaired T-cell immunity against Pneumocystis infection and possibly lead to immune tolerance. Similar to our findings, in fungal infections, a recent study showed that P. brasiliensis infection induces a vigorous IDO expression.44 In addition, the main pathway of tryptophan catabolism metabolism is the kynurenine pathway mediated by the key enzyme IDO1.40,45,46 Kynurenine is the main downstream product of the IDO1. However, our study showed that compared with control mice, the product of tryptophan metabolism, kynurenine, was downregulated in the lung of WT Pneumocystis-infected mice, although the expression level of IDO1 was upregulated. This was probably due to an increase in kynurenine’s incoming but more pronounced outgoing route. Hence, the results suggested a potential role of tryptophan metabolism in Pneumocystis infection. Given the complexity of the body’s anti-Pneumocystis immunity, the current results are speculative, and further mechanistic studies are needed to explore the role of tryptophan metabolism in Pneumocystis infection.
CD4+ T cells are crucial in controlling Pneumocystis infection.47 Increasing evidence suggests that B cells also play an essential role in Pneumocystis infection.48,49 Our previous study18 found that the counts of B cells in mouse peripheral blood and lung tissue decreased after Pneumocystis infection, suggesting the significance of B cells. We further found that the clearance of Pneumocystis was delayed in BAFF-R–/– mice. However, current studies focus on the cellular level. Although we have done some molecular mechanism studies, metabolism-related studies on PCP are still lacking. Metabolism plays a crucial role in regulating the body’s response to infection. Our study showed that many metabolites in the lung samples were altered in Pneumocystis-infected mice. In addition, there were differences in metabolites involved in the fatty acid metabolism in the lungs between WT Pneumocystis-infected mice and BAFF-R–/– Pneumocystis-infected mice.
Similar to our previous study,18 as for the uninfected mice, BAFF-R–/– mice exhibited mainly the absence of mature B cells and a reduced but not complete lack of total B cells. We also observed no statistically significant differences in appearance, body weight, growth, and development between uninfected BAFF-R–/– mice and WT mice, so we did not set the group of uninfected BAFF-R–/– mice in the metabolomic analysis. As for the infected mice, we found that BAFF-R deficiency resulted in delayed clearance of Pneumocystis and more serious inflammatory cell infiltration in the lungs compared with Pneumocystis-infected WT mice in our previous study.18 In comparing WT Pneumocystis-infected mice and BAFF-R–/– Pneumocystis-infected mice, we found some differential metabolites, mainly lipids and lipid-like molecules. For example, the level of vitamin A’s metabolite, alitretinoin, was lower in the lung samples of BAFF-R–/– Pneumocystis-infected mice than in WT Pneumocystis-infected mice. Alitretinoin, also known as 9-cis-retinoic acid, is an isomer of all-trans retinoic acid.50 As the active metabolites of vitamin A, much evidence has suggested that all-trans-retinoic acid and 9-cis-retinoic acid could increase the frequency of Treg cells and downregulate the frequency of Th17 cells.51–53 It has been suggested that retinoic acid promotes the production of Tregs but restrains the differentiation of Th17 cells; it enhances the generation of Th2 but suppresses the Th1 cell differentiation of naïve T cells.50 Furthermore, retinoic acid signaling is required for Th1 and Th17 immune responses to infection.54 The lack of retinoic acid may enhance proinflammatory responses and diminish anti-inflammatory responses. It may be related to the recruitment of more inflammatory cells in the lung tissue of BAFF-R–/– mice. Although the recruitment of more inflammatory cells is intended to promote Pneumocystis clearance, excessive recruitment may lead to inflammatory damage, so more obvious alveolar hemorrhage was observed in the lung tissue of BAFF-R–/– mice.
We found some enriched metabolic pathways in BAFF-R–/– Pneumocystis-infected mice compared to WT Pneumocystis-infected mice, such as adipocytokine signaling pathway and fatty acid elongation. We also found significantly increased expression of key enzymes of fatty acid metabolism, such as SCD1, FASN, in the lungs of BAFF-R–/– Pneumocystis-infected mice. Many studies have suggested that the significant expression increase of SCD1 and other key enzymes of fatty acid metabolism is associated with the dysregulation of proinflammatory genes.55–57 This indicated a key role for fatty acid metabolism in regulating inflammation responses.
Our previous study demonstrated a significant role of B cells in defense against Pneumocystis infection.15 Another study explored the role of B cells in regulating helper T lymphocytes’ response to Pneumocystis infection18 and suggested that BAFF-R deficiency prompted Th17-cell immune responses during Pneumocystis infection. This study found that elevated expression of key enzymes of fatty acid metabolism, such as SCD1, was positively correlated with IL17A levels. The results indicated that the abnormal fatty acid metabolism might be associated with the increased Th17-cell immune responses. We hypothesize that the over-recruitment of Th17 cells, although intended to promote Pneumocystis clearance, also caused more obvious lung damage during Pneumocystis infection in BAFF-R gene knockout Pneumocystis-infected mice.
Our results suggested that the metabolites, alitretinoin, as well as the products of fatty acid metabolism, may influence the inflammatory responses in Pneumocystis infection. In BAFF-R–/– Pneumocystis-infected mice, the balance between anti- and pro-inflammatory responses was disrupted, resulting in increased inflammatory damage. In short, these findings indicated that an appropriate Th1/Th17 response may contribute to the clearance of Pneumocystis, but an excessive response may result in inflammatory damage, where the balance between anti- and pro-inflammatory immunity is critical.
Overall, our study revealed the metabolic changes in the lungs of mice with Pneumocystis infection. However, there are some limitations to the study. First, the results of this study were based on the mouse model with limited sample sizes and lacked validation in PCP patients. Second, we only chose to observe the change of metabolites at a time point when the Pneumocystis burden was relatively high instead of observing the dynamic change of metabolites during the whole progression of Pneumocystis infection. Last but not least, future studies should be conducted to further explore the mechanism of metabolic changes.
Conclusions
In conclusion, many lipids and lipid-like molecules are dysregulated in Pneumocystis infection, which may influence B-cell development and function. The results also show noticeable changes in tryptophan metabolism in mice with Pneumocystis infection. In addition, the metabolomics analysis of the lung revealed a lower level of alitretinoin and the abnormality of fatty acid metabolism in BAFF-R gene knockout Pneumocystis-infected mice as compared with the WT Pneumocystis-infected mice, which may be related to the greater inflammatory cell infiltration in the lungs of BAFF-R–/– Pneumocystis-infected mice. The findings of metabolomic profiling of the lungs provide insights into developing novel therapeutic targets for PCP.
Abbreviations
PCP, Pneumocystis pneumonia; HIV, human immunodeficiency virus; BAFF-R, B cell-activating factor receptor; WT, wild-type; BAFF-R–/–, BAFF-R deficient; ICU, Intensive care unit; TNF, tumor necrosis factor; SCID, severe combined immunodeficient; PCA, principal component analysis; OPLS-DA, orthogonal partial least squares discriminant analysis; VIP, variable importance in projection; FC, fold change; ESI+, electrospray ionization positive; ESI-, electrospray ionization negative; VAP, ventilator-associated pneumonia; CAP, community-acquired pneumonia; IDO1, indoleamine 2,3-dioxygenase 1; TPH, tryptophan hydroxylase; SCD1, stearoyl-CoA desaturase 1; FASN, fatty acid synthase.
Ethics Statement
The animal study was approved by Beijing Chao Yang Hospital, Capital Medical University. No. AEEI-2020-121.
Funding
This work was funded by the National Natural Science Foundation of China (No. 82270009) and the Natural Science Foundation of Capital Medical University (PYZ21084).
Disclosure
These authors report no conflicts of interest in this work.
References
1. Grønseth S, Rogne T, Hannula R, Åsvold BO, Afset JE, Damås JK. Epidemiological and clinical characteristics of immunocompromised patients infected with Pneumocystis jirovecii in a twelve-year retrospective study from Norway. BMC Infect Dis. 2021;21(1):659. doi:10.1186/s12879-021-06144-1
2. Cillóniz C, Dominedò C, Álvarez-Martínez MJ, et al. Pneumocystis pneumonia in the twenty-first century: HIV-infected versus HIV-uninfected patients. Expert Rev Anti Infect Ther. 2019;17(10):787–801. doi:10.1080/14787210.2019.1671823
3. Roux A, Canet E, Valade S, et al. Pneumocystis jirovecii pneumonia in patients with or without AIDS, France. Emerg Infect Dis. 2014;20(9):1490–1497. doi:10.3201/eid2009.131668
4. Enomoto T, Azuma A, Kohno A, et al. Differences in the clinical characteristics of Pneumocystis jirovecii pneumonia in immunocompromized patients with and without HIV infection. Respirology. 2010;15(1):126–131. doi:10.1111/j.1440-1843.2009.01660.x
5. Gingerich AD, Norris KA, Mousa JJ. Pneumocystis pneumonia: immunity, vaccines, and treatments. Pathogens. 2021;10(2). doi:10.3390/pathogens10020236
6. Shellito J, Suzara VV, Blumenfeld W, Beck JM, Steger HJ, Ermak TH. A new model of Pneumocystis carinii infection in mice selectively depleted of helper T lymphocytes. J Clin Invest. 1990;85(5):1686–1693. doi:10.1172/jci114621
7. Beck JM, Warnock ML, Curtis JL, et al. Inflammatory responses to Pneumocystis carinii in mice selectively depleted of helper T lymphocytes. Am J Respir Cell Mol Biol. 1991;5(2):186–197. doi:10.1165/ajrcmb/5.2.186
8. Fishman JA. Pneumocystis jiroveci. Semin Respir Crit Care Med. 2020;41(1):141–157. doi:10.1055/s-0039-3399559
9. Garvy BA, Wiley JA, Gigliotti F, Harmsen AG. Protection against Pneumocystis carinii pneumonia by antibodies generated from either T helper 1 or T helper 2 responses. Infect Immun. 1997;65(12):5052–5056. doi:10.1128/iai.65.12.5052-5056.1997
10. Hu T, Takamoto M, Hida S, Tagawa Y, Sugane K. IFN-γ deficiency worsen Pneumocystis pneumonia with Th17 development in nude mice. Immunol Lett. 2009;127(1):55–59. doi:10.1016/j.imlet.2009.08.013
11. Otieno-Odhiambo P, Wasserman S, Hoving JC. The contribution of host cells to pneumocystis immunity: an update. Pathogens. 2019;8(2):52. doi:10.3390/pathogens8020052
12. Kolls JK. An emerging role of B cell immunity in susceptibility to pneumocystis pneumonia. Am J Respir Cell Mol Biol. 2017;56(3):279–280. doi:10.1165/rcmb.2016-0360ED
13. Elsegeiny W, Eddens T, Chen K, Kolls JK, Deepe GS. Anti-CD20 antibody therapy and susceptibility to Pneumocystis pneumonia. Infect Immun. 2015;83(5):2043–2052. doi:10.1128/iai.03099-14
14. Opata MM, Hollifield ML, Lund FE, et al. B lymphocytes are required during the early priming of CD4+ T cells for clearance of pneumocystis infection in mice. J Immunol. 2015;195(2):611–620. doi:10.4049/jimmunol.1500112
15. Hu Y, Wang D, Zhai K, Tong Z. Transcriptomic analysis reveals significant B lymphocyte suppression in corticosteroid-treated hosts with pneumocystis pneumonia. Am J Respir Cell Mol Biol. 2017;56(3):322–331. doi:10.1165/rcmb.2015-0356OC
16. Mackay F, Schneider P, Rennert P, Browning J. BAFF AND April: a tutorial on B cell survival. Annu Rev Immunol. 2003;21(1):231–264. doi:10.1146/annurev.immunol.21.120601.141152
17. Thompson JS, Bixler SA, Qian F, et al. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science. 2001;293(5537):2108–2111. doi:10.1126/science.1061965
18. Rong H-M, Li T, Zhang C, et al. IL-10-producing B cells regulate Th1/Th17-cell immune responses in Pneumocystis pneumonia. Am J Physiol Lung Cell Mol Physiol. 2019;316(1):L291–l301. doi:10.1152/ajplung.00210.2018
19. Jung J, Zeng H, Horng T. Metabolism as a guiding force for immunity. Nat Cell Biol. 2019;21(1):85–93. doi:10.1038/s41556-018-0217-x
20. Loftus RM, Finlay DK. Immunometabolism: cellular metabolism turns immune regulator. J Biol Chem. 2016;291(1):1–10. doi:10.1074/jbc.R115.693903
21. Rahman AN, Liu J, Mujib S, et al. Elevated glycolysis imparts functional ability to CD8(+) T cells in HIV infection. Life Sci Alliance. 2021;4(11). doi:10.26508/lsa.202101081
22. Jing Y, Luo L, Chen Y, et al. SARS-CoV-2 infection causes immunodeficiency in recovered patients by downregulating CD19 expression in B cells via enhancing B-cell metabolism. Signal Transduct Target Ther. 2021;6(1):345. doi:10.1038/s41392-021-00749-3
23. Gertsman I, Barshop BA. Promises and pitfalls of untargeted metabolomics. J Inherit Metab Dis. 2018;41(3):355–366. doi:10.1007/s10545-017-0130-7
24. Thomas T, Stefanoni D, Reisz JA, et al. COVID-19 infection alters kynurenine and fatty acid metabolism, correlating with IL-6 levels and renal status. JCI Insight. 2020;5(14). doi:10.1172/jci.insight.140327
25. Ceballos FC, Virseda-Berdices A, Resino S, et al. Metabolic profiling at COVID-19 onset shows disease severity and sex-specific dysregulation. Front Immunol. 2022;13:925558. doi:10.3389/fimmu.2022.925558
26. Xu J, Zhou M, Luo P, et al. Plasma metabolomic profiling of patients recovered from coronavirus disease 2019 (COVID-19) with pulmonary sequelae 3 months after discharge. Clin Infect Dis. 2021;73(12):2228–2239. doi:10.1093/cid/ciab147
27. Antcliffe D, Jiménez B, Veselkov K, Holmes E, Gordon AC. Metabolic profiling in patients with pneumonia on intensive care. EBioMedicine. 2017;18:244–253. doi:10.1016/j.ebiom.2017.03.034
28. den Hartog I, Zwep LB, Vestjens SMT, et al. Metabolomic profiling of microbial disease etiology in community-acquired pneumonia. PLoS One. 2021;16(6):e0252378. doi:10.1371/journal.pone.0252378
29. Schmidt SV, Schultze JL. New insights into IDO biology in bacterial and viral infections. Front Immunol. 2014;5:384. doi:10.3389/fimmu.2014.00384
30. Zhou X, Zhu X, Zeng H. Fatty acid metabolism in adaptive immunity. Febs J. 2021. doi:10.1111/febs.16296
31. Hussain H, Vutipongsatorn K, Jiménez B, Antcliffe DB. Patient stratification in sepsis: using metabolomics to detect clinical phenotypes, sub-phenotypes and therapeutic response. Metabolites. 2022;12(5). doi:10.3390/metabo12050376
32. Li J, Luu LDW, Wang X, et al. Metabolomic analysis reveals potential biomarkers and the underlying pathogenesis involved in Mycoplasma pneumoniae pneumonia. Emerg Microbes Infect. 2022;11(1):593–605. doi:10.1080/22221751.2022.2036582
33. Adu-Gyamfi CG, Snyman T, Hoffmann CJ, et al. Plasma indoleamine 2, 3-dioxygenase, a biomarker for tuberculosis in human immunodeficiency virus-infected patients. Clin Infect Dis. 2017;65(8):1356–1358. doi:10.1093/cid/cix550
34. Dewulf JP, Martin M, Marie S, et al. Urine metabolomics links dysregulation of the tryptophan-kynurenine pathway to inflammation and severity of COVID-19. Sci Rep. 2022;12(1):9959. doi:10.1038/s41598-022-14292-w
35. Zardini Buzatto A, Sarkar I, van Drunen Littel-van den Hurk S, Li L. Comprehensive Lipidomic and metabolomic analysis for studying metabolic changes in lung tissue induced by a vaccine against respiratory syncytial virus. ACS Infect Dis. 2020;6(8):2130–2142. doi:10.1021/acsinfecdis.0c00210
36. Zahoor I, Suhail H, Datta I, et al. Blood-based untargeted metabolomics in relapsing-remitting multiple sclerosis revealed the testable therapeutic target. Proc Natl Acad Sci U S A. 2022;119(25):e2123265119. doi:10.1073/pnas.2123265119
37. Hao Y, Zhang Z, Feng G, et al. Distinct lipid metabolic dysregulation in asymptomatic COVID-19. iScience. 2021;24(9):102974. doi:10.1016/j.isci.2021.102974
38. Weisel FJ, Mullett SJ, Elsner RA, et al. Germinal center B cells selectively oxidize fatty acids for energy while conducting minimal glycolysis. Nat Immunol. 2020;21(3):331–342. doi:10.1038/s41590-020-0598-4
39. Zhou X, Zhu X, Li C, et al. Stearoyl-CoA desaturase-mediated monounsaturated fatty acid availability supports humoral immunity. Cell Rep. 2021;34(1):108601. doi:10.1016/j.celrep.2020.108601
40. Dagenais-Lussier X, Loucif H, Beji C, Telittchenko R, Routy JP, van Grevenynghe J. Latest developments in tryptophan metabolism: understanding its role in B cell immunity. Cytokine Growth Factor Rev. 2021;59:111–117. doi:10.1016/j.cytogfr.2021.02.003
41. Grohmann U, Bronte V. Control of immune response by amino acid metabolism. Immunol Rev. 2010;236:243–264. doi:10.1111/j.1600-065X.2010.00915.x
42. Munn DH, Mellor AL. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013;34(3):137–143. doi:10.1016/j.it.2012.10.001
43. Platten M, Nollen EAA, Röhrig UF, Fallarino F, Opitz CA. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov. 2019;18(5):379–401. doi:10.1038/s41573-019-0016-5
44. de Araújo EF, Loures FV, Preite NW, et al. AhR ligands modulate the differentiation of innate lymphoid cells and T Helper cell subsets that control the severity of a pulmonary fungal infection. Front Immunol. 2021;12:630938. doi:10.3389/fimmu.2021.630938
45. Peyraud F, Guegan JP, Bodet D, Cousin S, Bessede A, Italiano A. Targeting tryptophan catabolism in cancer immunotherapy era: challenges and perspectives. Front Immunol. 2022;13:807271. doi:10.3389/fimmu.2022.807271
46. Moffett JR, Namboodiri MA. Tryptophan and the immune response. Immunol Cell Biol. 2003;81(4):247–265. doi:10.1046/j.1440-1711.2003.t01-1-01177.x
47. Morris A, Norris KA. Colonization by Pneumocystis jirovecii and its role in disease. Clin Microbiol Rev. 2012;25(2):297–317. doi:10.1128/cmr.00013-12
48. Lund FE, Schuer K, Hollifield M, Randall TD, Garvy BA. Clearance of Pneumocystis carinii in mice is dependent on B cells but not on P carinii-specific antibody. J Immunol. 2003;171(3):1423–1430. doi:10.4049/jimmunol.171.3.1423
49. Ruan S, Cai Y, Ramsay AJ, Welsh DA, Norris K, Shellito JE. B cell and antibody responses in mice induced by a putative cell surface peptidase of Pneumocystis murina protect against experimental infection. Vaccine. 2017;35(4):672–679. doi:10.1016/j.vaccine.2016.11.073
50. Larange A, Cheroutre H. Retinoic acid and retinoic acid receptors as pleiotropic modulators of the immune system. Annu Rev Immunol. 2016;34:369–394. doi:10.1146/annurev-immunol-041015-055427
51. Xiao S, Jin H, Korn T, et al. Retinoic acid increases Foxp3+ regulatory T cells and inhibits development of Th17 cells by enhancing TGF-beta-driven Smad3 signaling and inhibiting IL-6 and IL-23 receptor expression. J Immunol. 2008;181(4):2277–2284. doi:10.4049/jimmunol.181.4.2277
52. Abdolahi M, Yavari P, Honarvar NM, Bitarafan S, Mahmoudi M, Saboor-Yaraghi AA. Molecular mechanisms of the action of vitamin A in Th17/Treg axis in multiple sclerosis. J Mol Neurosci. 2015;57(4):605–613. doi:10.1007/s12031-015-0643-1
53. Mucida D, Park Y, Kim G, et al. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science. 2007;317(5835):256–260. doi:10.1126/science.1145697
54. Hall JA, Cannons JL, Grainger JR, et al. Essential role for retinoic acid in the promotion of CD4(+) T cell effector responses via retinoic acid receptor alpha. Immunity. 2011;34(3):435–447. doi:10.1016/j.immuni.2011.03.003
55. Guo R, Xu X, Babcock SA, Zhang Y, Ren J. Aldehyde dedydrogenase-2 plays a beneficial role in ameliorating chronic alcohol-induced hepatic steatosis and inflammation through regulation of autophagy. J Hepatol. 2015;62(3):647–656. doi:10.1016/j.jhep.2014.10.009
56. Fang R, Yang S, Gu X, Li C, Bi N, Wang HL. Early-life exposure to bisphenol A induces dysregulation of lipid homeostasis by the upregulation of SCD1 in male mice. Environ Pollut. 2022;304:119201. doi:10.1016/j.envpol.2022.119201
57. Zhou J, Zhang N, Aldhahrani A, Soliman MM, Zhang L, Zhou F. Puerarin ameliorates nonalcoholic fatty liver in rats by regulating hepatic lipid accumulation, oxidative stress, and inflammation. Front Immunol. 2022;13:956688. doi:10.3389/fimmu.2022.956688
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.