Back to Journals » Journal of Inflammation Research » Volume 13
Mesenchymal Stem Cells Anti-Inflammatory Activity in Rats: Proinflammatory Cytokines
Authors Petryk N ![]() , Shevchenko O
, Shevchenko O
Received 14 April 2020
Accepted for publication 27 June 2020
Published 7 July 2020 Volume 2020:13 Pages 293—301
DOI https://doi.org/10.2147/JIR.S256932
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Video abstract presented by Nataliia Petryk.
Views: 1228
Nataliia Petryk, Oleksandr Shevchenko
Department of Pathology, Kharkiv National Medical University, Kharkiv, Ukraine
Correspondence: Nataliia Petryk
Department of Pathology, Kharkiv National Medical University, Kharkiv, Ukraine
Email [email protected]
Introduction: Many widespread intractable diseases are caused or supported by chronic inflammation. Such conditions include the 2nd type of diabetes mellitus, atherosclerosis, neurodegenerative diseases, chronic inflammatory diseases of the connective tissue - ankylosing spondylitis, rheumatoid arthritis, autoimmune myositis, etc. Therefore, the search for targeted treatment of these illnesses is extremely in high demand. Immunomodulatory activity of mesenchymal stem cells is one of their remarkable properties. Several biomarkers (cytokines and nonspecific proteins) are known to be associated with chronic inflammation.
Methods: Our study aimed to investigate the serum levels of tumor necrosis factor-alpha, interleukin 6 and C-reactive protein in carrageenan myositis in rats, because there is more and more evidence of the significance of these markers in the course and resolution of the diseases mentioned above. For the first time in our experiment, it was shown and evaluated using analysis of variance, how MSCs influence the indicators of proinflammatory cytokines on the model of carrageenan myositis. The levels of α-TNF, IL-6 and CRP in the plasma of rats were studied in groups with chronic carrageenan inflammation and chronic inflammation with local injection of MSCs into the affected area.
Results: Our study proved the effectiveness of MSCs by showing a significant decrease in the levels of inflammatory mediators in the plasma of the studied animals.
Discussion and Conclusions: Thus, the administration of MSCs is a promising tool in the pathogenic treatment of chronic inflammation and concomitant conditions.
Keywords: chronic inflammation, mesenchymal stem cells, carrageenan, tumor necrosis factor-alpha, interleukin-6, C-reactive protein
Introduction
Mesenchymal stem cells (MSCs) are non-hematopoietic stromal cells. MSCs can differentiate into mesenchymal tissues (bones, cartilage, muscles, ligaments and fat) and provide their regeneration.1 MSCs represent in the bone marrow in an amount of 1 to 10,000 cells containing nuclei. Although these cells are not immortal, they are capable of multiple divisions in culture, while maintaining the potential for differentiation in different lines. MSCs are identified by expression of a variety of molecules, including CD 105, CD 73, and hematopoietic negativity markers CD 34, CD 45 so on. The properties of mesenchymal stem cells show them potentially ideal candidates for tissue engineering. It was concluded that with systemic use, MSCs can get into places of damage in animals, so MSCs can migrate. The mechanisms, responsible for stem cell migration are not known reliably. It is suggested that chemokine receptors and stem cell adhesion molecules provide such potential for MSCs.
It was well documented in the literature that MSCs come from perivascular cells called pericytes.2,3 This explains how MSCs can be isolated from almost all vascularized tissues in the body.4,5 When blood vessels are damaged or inflamed, pericytes separate from the basement of the vessel and become MSCs. These newly formed MSCs respond to the local environment by secretion of cytokines.
The presence of non-hematopoietic stem cells in the bone marrow was first suggested by Kongheim about 130 years ago. He had thought that the bone marrow could be the source of fibroblasts involved in wound healing in numerous peripheral tissues.6 Evidence that the bone marrow contains cells that can differentiate into other mesenchymal cells is now publicly available, starting with the work of Friedenstein and colleagues.7 They showed that the bone marrow of rodents had fibroblast-like cells with a clonogenic potential in vitro. In the early 70s, Friedenstein placed the bone marrow in plastic Petri dishes and, after several hours of incubation, washed off cells that did not adhere. Cells adhered to plastic had a fusiform shape and were various, but capable of forming colonies. These cells could also grow bone and restore the hematopoietic microenvironment in subcutaneous grafts. Moreover, the ability of these cells to regenerate heterotopic bone tissue in a series of transplants has been demonstrated, thus providing evidence in support of MSCs self-renewing potential. Over the years, many laboratories have approved and expanded knowledge about the regenerative abilities of MSCs. It was shown that cells, isolated with the Friedenstein protocol, were present in the human bone marrow as well. It was also described that these cells could be differentiated into many cells of mesenchymal lines in vitro, such as osteoblasts, chondrocytes, adipocytes, and myoblasts.8–10
The immunomodulating and anti-inflammatory properties of MSCs are remaining partially unexplained. MSCs express a moderate level of proteins of the main histocompatibility complex of the first class (MHC I) but do not express MHC II proteins.11 This phenotype is considered non-immunogenic, which suggests that such a transplant to the allogenic host will not require the use of immunosuppressants. Moreover, several studies show that MSCs have immunosuppressive properties by modulating the functioning of specific T cells in vitro.12 Some of these observations were made in vivo. It was shown an experimental model of graft-versus-host disease.13
MSCs, existing almost in all vascular tissues, have an important role in regulation mechanisms of chronic inflammation.14 Chronic inflammation, unsolved problems in our modern society, accompanies different conditions, such as II type diabetes, atherosclerosis, obesity, cancer, neurodegenerative diseases so on.15–19 A variety of mediators are believed to be responsible for the immunomodulatory ability of MSCs, such as transforming growth factor (TGF) -β, indolamine 2, 3-dioxygenase, inducible nitric oxide synthase (iNOS) and prostaglandin E2.20 These possibilities suggest great therapeutic potential for MSCs. Recently, several studies have reported that MSCs can alleviate liver fibrosis and promote liver regeneration.21 Whether MSCs can effectively reduce the level of pro-inflammatory cytokines and have an immunoregulatory effect in chronic inflammation on the model of carrageenan myositis remains unclear.
Tumor Necrosis Factor-Alpha (α-TNF)
α-TNF is an inflammatory protein that is secreted by monocytes/macrophages, T-cells, smooth muscle cells, adipocytes, and fibroblasts and is responsible for signaling events leading to either apoptosis or necrosis. It also has an important role in developing resistance to cancer and infections. α-TNF provides a wide spectrum of cellular responses including regulation of inflammation, immune response, cell survival, proliferation and apoptosis. In 1975, it was shown in experiments that after stimulation of the reticuloendothelial system in animals, a protein called the tumor necrosis factor was released into the circulation. This protein could cause necrotic regression of certain tumors.22,23
All known effects of α-TNF are mediated by two types of receptors. They are designated as TNFR1 and TNFR2. Binding with different receptors can either trigger similar or different responses. TNFR1 and TNFR2 have extracellular and intracellular ligand-binding.24
Extracellular domains are members of the nerve growth factor/TNF receptor gene family. Intracellular domains do not have similar sequences and therefore activate different signal transmission paths. This may explain how activation of TNFR1 or TNFR2 causes similar or different effects. The key role in α-TNF signal transduction is believed to belong to the transcription factor NF-κB. The stimuli that induce α-TNF production include bacterial toxins, infections, injury, antigen-antibody complexes and diverse environmental factors. According to numerous scientific studies, serum α-TNF levels are elevated in both acute and chronic inflammation.25
Interleukin-6 (IL-6)
IL-6 is a polypeptide with various regulatory functions concerning inflammation, the immune system and blood formation. Until 1989, various studies were conducted that described several molecules with different functions, such as the ability to stimulate B cells to differentiate into cells producing antibodies or to stimulate the production of acute-phase protein in the liver. After the identity of these molecules was established, a new name was proposed IL-6.
IL-6 is produced in response to tissue damage or infections primarily by monocytes and lymphocytes, but can also be produced by other cells.
In the case of an infectious process, the stimulation of the production of IL-6 occurs by binding to macrophage and monocyte pathogen-recognition receptors (PRRs). This leads to the activation of many pathways, including NF-kb and consequently to increasing of mRNA transcription which leads to releasing of inflammatory cytokines.
In the case of tissue injury damage-associated molecular patterns (DAMPs) are released.DAMPs from dying or damaged cells contain several molecules that stimulate IL-6 production.23
After the production in the initial site of inflammation IL-6 it moves to the bloodstream and reaches the liver.26
It causes rapid induction of C-reactive protein (CRP), serum amyloid A (SAA), fibrinogen, haptoglobin, and α1-antichymotrypsin. Simultaneously, IL-6 diminishes the production of transferrin, fibronectin, and albumin. Also, IL-6 causes a decrease in iron and zinc in the blood, stimulates megakaryocytes in the bone marrow. With the help of this cytokine, immunocompetent cells are differentiated.
C-Reactive Protein (CRP)
CRP is one of the acute phase proteins. It was first isolated in 1930 when studying the serum of patients suffering from pneumococcal infection. Its name comes from the ability to react with the capsular lipopolysaccharide of pneumococcus.
CRP is synthesized mainly in hepatocytes in response to increased levels of proinflammatory cytokines, mainly IL-6 and to a lesser extent interleukin−1(IL-1) and TNFα. In addition to hepatocytes, CRP can be produced by smooth muscle cells, endothelial cells, macrophages, lymphocytes and adipocytes.
It is synthesized in the form of a homopentamer and in the foci of inflammation irreversibly dissociates into monomers.
The level of CRP rises with rheumatic diseases, bacterial infections, cardiovascular diseases and other inflammatory processes.
CRP is mainly regarded as a marker of inflammation. However, it plays an important role in the regulation of the immune response. The main role of CRP in inflammation is to activate the C1q component of the complement system, followed by the opsonization of pathogens. Therefore, CRP demonstrates protective action against bacterial infections.27
Given the correlation between the levels of the aforementioned markers and the intensity of inflammation, it was decided to use them to evaluate the effect of mesenchymal stem cells on chronic inflammation in the experiment.
Materials and Methods
Our experiment was obtained with 132 adult male laboratory rats (200–220g of weight, 4–5-month in age) in the vivarium of Kharkiv National Medical University. The study was carried out following the national “General Ethical Principles for Animal Research” (Ukraine, 2001), the “European Convention for the Protection of Vertebrate Animals Used for Experimental and Other Scientific Purposes” (Strasbourg, 18.03.1986p.). Also, the Declaration of Helsinki, adopted by the General Assembly of the World Medical Association (1964–2000), the charter of the Ukrainian Association for Bioethics and GLP (1992). Animals were randomly divided into 22 groups with 6 animals in every group - the minimum, acceptable for statistical processing and obtaining reliable results conventional number of animals, as well as the minimum sufficient to achieve the goal and the objectives of the study, the number of experimental groups. Animals were sacrificed by inhalation of high concentrations of carbon dioxide (CO2) followed by decapitation. Laboratory animals were kept in separate cages, 6 rats in every cage at room temperature 21±2ºC, 45–50% humidity, 12h/12h light/dark day cycle, with 24 hours access to food and water. Kharkiv National Medical University Ethics Committee approved our study (Order № 370 dated “06” September 2017 Form B-5). The study was performed with minimal psychological loss from experimental animals.
Obtaining MSCs
MSCs were isolated from the bone marrow of the femur of rats by the standard procedure.28–31 Under sterile conditions, the bone marrow was removed from the epiphyses of the rat’s femur, then it was washed with a solution of Hanks (Biowest, France). The cells were centrifuged to precipitate them (1000 rpms for 10 min). Mononuclear cells were isolated by centrifugation in a Ficoll-Hypaque gradient (density of 1.077 g/mL) (Sigma, USA) at 400g for 25 min and washed twice with Hanks solution (Biowest, France). After that, the cells were resuspended in saline at a concentration of 1.0 x 106 in 1 mL.32,33 We counted the total number of live cells in a hemocytometer by staining with a 0.2% trypan blue solution (Janssen Chemica, Belgium). The structure of MSCs was investigated with a phase-contrast microscope; we studied cell cycles by flow cytometry. The phenotype of MSCs was studied by using immunocytochemical methods.
Primary cultured MSC were oval, spindle-shaped or polygonal, and adhered to a plastic surface within 24 h and reached 90% confluence within eight days. After purification and proliferation, they were uniformly long spindle-shaped forms and passed every five days. The adhesion rate within 24 hours was all. The flow cytometry showed 80% cells of the fourth generation of MSC were at the G0 phase. Immunocytochemistry showed MSC were positive for CD29, CD105, CD166, VLA-4, and P-selectin, while negative for CD34 and CD45.
Induction of Carrageenan Inflammation
Chronic inflammation was caused by an intramuscular injection into the right thigh of a rat with 0.1 mL of a 10% solution of λ-carrageenan (Sigma-Aldrich GmbH), equivalent to 10 mg of the active substance.34,35 For each animal was used a sterile 1cc syringe with a 27-G needle attached with 100 μL of carrageenan.36 The substance for injections was a gel and it required not rapid but slow injection during 10–11 seconds.
To avoid pained stress for animals, all injections were carried out under sufficient isoflurane anesthesia.37
Experiment and Plasma Collection
There were 60 rats with edema in the right hip due to injection of λ-carrageenan intramuscularly, 60 rats were simultaneously injected not only with carrageenan but, also, with the suspension of MSCs in the experiment. Control group consisted of 6 intact rats without any interventions, and 6 rats, administered MSCs without causing inflammation.
The concentration of MSCs was 2 million cells in 0.4 mL for every animal. There were 10 terms in the experiment. For every term, 6 rats with carrageenan inflammation and 6 rats with inflammation plus MSCs were analyzed. Anesthetized animals were sacrificed after 6 hours, on the 1st day, the 2nd day, the 3rd day, the 5th day, the 7th day, the 10th day, the 14th day, the 21st day, the 28th day. Blood samples were obtained by cardiac puncture and collected into sterile tubes. The blood clot appeared in 25–30 min after that tubes were placed into a centrifuge and processed at 3000 rpm for 10 min. Plasma was obtained and sent to a deep freezer (−20 ºC) until analysis was performed - after the 28th day of the experiment.
Determination of α-TNF, IL-6, and CRP
Plasma levels of α-TNF, IL-6 and CRP were measured with Enzyme-Linked Immunosorbent Assay kit (Sigma-Aldrich GmbH), for the quantitative measurement of target markers in biological liquids. We used The Rat TNF-α ELISA, The Rat IL-6 ELISA, The Rat CRP (C-Reactive Protein) ELISA kits.
Statistical Analysis
All calculations were performed with Statistica (data analysis software system), version 13. We use a one-way ANOVA test for comparison, Turkey post hoc test for α-TNF. IL-6 and CRP profiles for all terms and groups, where p < 0.05 was accepted as statistically significant.
Results
Increased α-TNF in the inflammation groups and decreased level of α-TNF in inflammation plus MSCs groups.
There was a significant difference in the level of α-TNF in all inflammation groups compared to control (P=0,0 - P=0,037). The bar chart (Figure 1) indicates that α-TNF in the inflammation groups steadily grow from 6 hours to 3rd day, then dropped gradually by the 21st day with a slight increase in the 28th day. Besides, the differences were significant in the level of α-TNF in all inflammation plus MSCs groups compared to control (P=0,0 - P=0,037) and compared to control plus MSCs group (P=0,0). Furthermore, α-TNF was significantly lower in groups myositis with MSCs between 14th day and 28 days in comparison with the control and the control plus MSCs. Moreover, the curve for inflammation with MSCs groups is flatter than the inflammation group curve. The level of α-TNF was significantly lower for the group of inflammation plus MSCs in comparison with the usual course of inflammation in the same term.
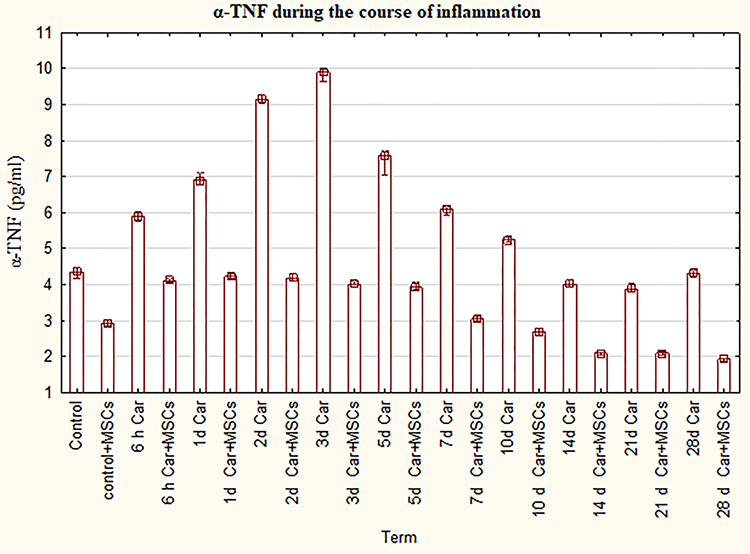 |
Figure 1 The levels of α-TNF for groups with the ordinary course of inflammation and inflammation on the background of MSCs. Data are represented as means ± SEM of 6 animals for each group. (One-way ANOVA and Tukey-Kramer multiple comparisons test, p<0.05). |
According to our statistical analysis, we can see that a statistically significant increase of IL-6 compared to intact control was between 6 hours and 5th day in the inflammation groups (P=0,0 - P=0,038) (Figure 2). On the 7th day and 28th day where the level of IL-6 was the same as in the control group (P=1,0), there was a slight decrease of IL-6 in the 10th day, 14 and 21 days (P=0,0 - P=0,031) in contrast to control. We got a wave-like curve for the course of inflammation in groups of myositis plus MSCs with a downward trend. But the main feature of the Figure 2 is dramatically dropping of IL-6 for the group of inflammation plus MSCs in comparison with the usual course of inflammation in the same term (P=0,0).
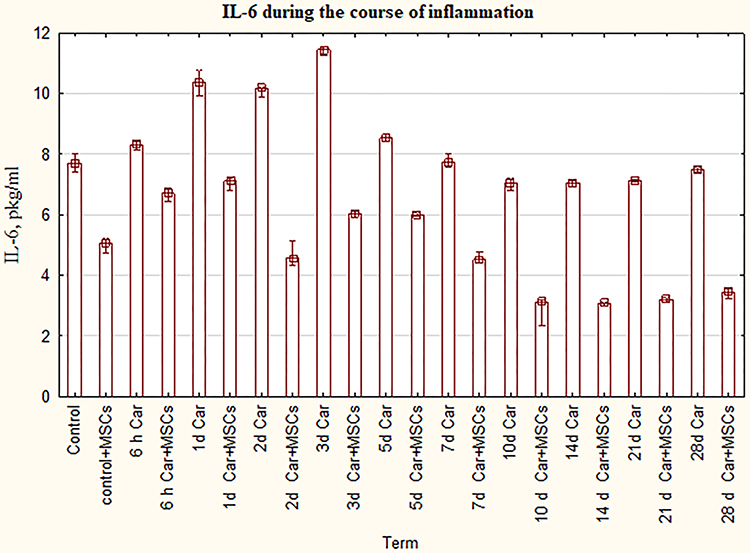 |
Figure 2 The levels of IL-6 for groups with the ordinary course of inflammation and inflammation on the background of MSCs. Data are represented as means ± SEM of 6 animals for each group. (One-way ANOVA and Tukey-Kramer multiple comparisons test, p<0.05). |
Measurement of CRP showed that inflammation plus MSCs group has drastically lower plasma levels of this protein in contrast to the carrageenan myositis group from the 1st day till the end of the experiment (P=0,0) (Figure 3). Furthermore, there was a slight decrease in CRP level in carrageenan plus MSCs groups between 6hours and 3rd day with dropping in the 5th day compared to control. Since 7th day, a more pronounced decline of the curve was supervised.
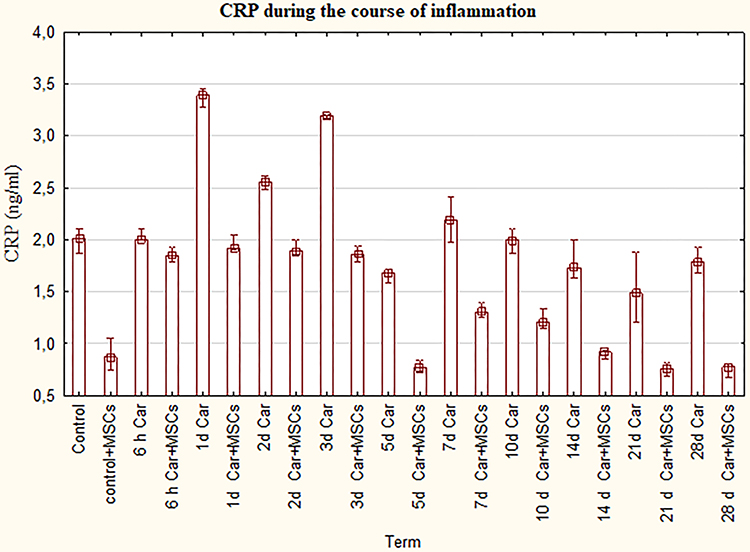 |
Figure 3 The levels of CRP for groups with the ordinary course of inflammation and inflammation on the background of MSCs. Data are represented as means ± SEM of 6 animals for each group. (One-way ANOVA and Tukey-Kramer multiple comparisons test, p<0.05). |
Also, we have noticed the same trends for all 3 figures. The levels of α-TNF, IL-6, CRP were significantly lower for the group of inflammation plus MSCs in comparison with the usual course of inflammation in the same term (P=0,0). Moreover, the level of inflammatory markers was noticeably higher in the intact control group compared to group MSCs without inflammation.
Therapeutic Benefits of MSCs in Clinic
Мany chronic intractable inflammatory diseases (rheumatoid arthritis, psoriasis, ankylosing spondylitis, juvenile arthritis, inflammatory bowel diseases) are pathogenetically associated with elevated levels of proinflammatory cytokines, in particular alpha TNF and interleukin 6. Adalimumab (monoclonal anti-TNF antibodies) and tocilizumab (antibodies to IL 6 receptor) are used to treat these diseases. The ability of mesenchymal stem cells to lower the levels of these pro-inflammatory cytokines may be used in the treatment of these diseases as an adjunct to monoclonal antibody therapy or as monotherapy.
Discussion
It is well known that many chronic inflammatory conditions are accompanied by increased levels of pro-inflammatory cytokines. There are several therapeutic options to lower their levels. Among them are monoclonal antibodies and cytokine receptor blockers, immunosuppressants and non-steroidal anti-inflammatory drugs. None of these medicines are completely safe and effective. Therefore, there is still a need to develop new approaches that can target other pathogenetic mechanisms.
One such approach may be to use MSCs to influence the production of pro-inflammatory cytokines in areas of inflammation or tissue damage. It is well known that MSCs can interact with immune cells of both innate and adaptive immune systems. This interaction is mediated by the secretion of numerous biologically active compounds such as cytokines, chemokines and growth factors.
The main reason for our study was to evaluate changes in proinflammatory cytokines profile throughout the carrageenan-induced inflammation. According to our study, the use of bone marrow-derived MSCs in the focus of chronic inflammation led to a significant decrease in the pro-inflammatory cytokines such as IL-6, TNFα, and CRP in animal plasma of the inflammation plus MSCs group. This decrease was statistically significant in comparison to the inflammation group. The results could be explained taking into account the immunosuppressive properties of MSCs. MSCs can interact with many types of immune cells, including B cells, T cells, dendritic cells (DC), natural killer cells (NK), neutrophils and macrophages.38 It was shown that the interaction mechanisms are based on intercellular contact, working in collaboration with the secretion of soluble immune factors to induce MSC-regulated immunosuppression.39 These specific modulators, including many immunomodulatory factors, cytokines, and growth factors, modulate inflammatory responses and balance immune profiles. MSCs also can regulate the inflammatory process and repair damaged cells and tissues by attaching to sites of inflammation.40 The integration of MSC with inflammatory processes can both strengthen and inhibit the immune response and depends on the general state of the immune system.41 Interestingly, MSCs modulate immunosuppression only when they are initially stimulated by inflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin- (IL-) 1.42 MSCs not only respond to inflammatory cytokines but also produce immunoregulatory secretors that mediate the inflammation process.43,44
It is important to note that despite the usage of allogeneic bone marrow-derived MSCs there was a significant decrease in IL-6, TNF-α and CRP in plasma of animals of the control plus MSCs group compared to the control group. It can be explained by the immunomodulatory capability of MSCs.45 Such a substantial decrease in proinflammatory cytokines could be the evidence of non- immunogenic properties of allogeneic MSCs. This fact may be important in such cases where harvesting autologous MSCs is impossible.
Even though an understanding of the mechanisms of MSC-based immunomodulation remains incomplete, the growing number of evidences are encouraging for further investigations of MSCs properties and putting them into practice.46 This can help develop pathogenetic treatments for chronic inflammatory and autoimmune diseases that will not have side effects, such as those associated with non-steroidal anti-inflammatory drugs, immunosuppressants, and monoclonal antibodies against inflammatory cytokines.
Conclusions
In the experiment in 132 rats, the immunomodulating and anti-inflammatory abilities of MSCs were reliably shown by significantly reducing the plasma levels of α-TNF, IL-6 and CRP in groups with local administration of MSCs to the affected area. Also, no hypersensitivity reactions or rejection were observed due to the use of allogeneic MSCs. Further additional researchers should be conducted to study not only the local effects of MSCs on the course of inflammation but also the systemic administration of MSCs. Further studies of MSCs and implementing into clinical practice will allow to treat a variety of diseases associated with chronic inflammation at a pathogenetic level and to provide their reliable prevention to reduce disability and premature mortality.
Abbreviations
MSCs, mesenchymal stem cells; Car, λ-carrageenan; CRP, C-reactive protein; α-TNF, tumor necrosis factor-alpha; IL 6, interleukin 6.
Data Sharing Statement
Research data are available from the corresponding author on a reasonable request.
Acknowledgments
This project was supported by Kharkiv National Medical University, Kharkiv, Ukraine.
Disclosure
The authors reports no conflicts of interest in this work.
References
1. Giselle C, James F, Brian A, Jim M. Concise review: mesenchymal stem cells: their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells. 2007;25(11):2739–2749.
2. Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human origins. Cell Stem Cell. 2008;3(3):301–313. doi:10.1016/j.stem.2008.07.003
3. Caplan AI. All MSCs are pericytes? Cell Stem Cell. 2008;3(3):229–230. doi:10.1016/j.stem.2008.08.008
4. da Silva Meirelles L, Chagastelles PC, Nardi NB. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J Cell Sci. 2006;119(11):2204–2213. doi:10.1242/jcs.02932
5. Campagnoli C, Roberts IA, Kumar S, Bennett PR, Bellantuono I, Fisk NM. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood. 2001;98(8):2396–2402. doi:10.1182/blood.V98.8.2396
6. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276:71–74. doi:10.1126/science.276.5309.71
7. Friedenstein AJ. Stromal mechanisms of bone marrow: cloning in vitro and retransplantation in vivo. HaematolBloodTransfus. 1980;25:19–29.
8. Bianco P, Robey PG, Simmons PJ. Mesenchymal stem cells: revisiting history, concepts, and assays. Cell Stem Cell. 2008;2:313–319. doi:10.1016/j.stem.2008.03.002
9. Caplan AI. Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J Cell Physiol. 2007;213:341–347. doi:10.1002/jcp.21200
10. Kolf C, Cho E, Tuan R. Biology of adult mesenchymal stem cells: regulation of niche, self-renewal and differentiation. Arthritis Res Ther. 2007;9(1):204. doi:10.1186/ar2116
11. Le Blanc K, Ringden O. Immunobiology of human mesenchymal stem cells and future use in hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2005;11:321–334. doi:10.1016/j.bbmt.2005.01.005
12. Beyth S, Borovsky Z, Mevorach D, et al. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T-cell unresponsiveness. Blood. 2005;105:2214–2219. doi:10.1182/blood-2004-07-2921
13. Dazzi F, Marelli-Berg FM. Mesenchymal stem cells for graft-versus-host disease: close encounters with T cells. Eur J Immunol. 2008;38:1479–1482. doi:10.1002/eji.200838433
14. Shi Y, Hu G, Su J, et al. Mesenchymal stem cells: a new strategy for immunosuppression and tissue repair. Cell Res. 2010;20:510–518. doi:10.1038/cr.2010.44
15. Leitner DR, Frühbeck G, Yumuk V, et al. Obesity and type 2 diabetes: two diseases with a need for combined treatment strategies - EASO can lead the way. Obes Facts. 2017;10(5):483–492. doi:10.1159/000480525
16. Boloker G, Wang C, Zhang J, et al. Updated statistics of lung and bronchus cancer in United States (2018). J Thorac Dis. 2018;10(3):1158–1161. doi:10.21037/jtd.2018.03.15.
17. Sitarz R, Skierucha M, Mielko J, et al. Gastric cancer: epidemiology, prevention, classification, and treatment. Cancer Manag Res. 2018;10(7):239–248. doi:10.2147/CMAR.S149619.
18. Chen -W-W, Zhang X, Huang W-J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol Med Rep. 2016;13(4):3391–3396. doi:10.3892/mmr.2016.4948
19. Nathan C, Ding A. Nonresolving Inflammation. Cell. 2010;140(6):871–882. doi:10.1016/j.cell.2010.02.029
20. Li W, Ren G, Huang Y, et al. Mesenchymal stem cells: hepatology 26 a double-edged sword in regulating immune responses. Cell Death Differ. 2012;19:1505–1513. doi:10.1038/cdd.2012.26
21. Zhang Y, Cai W, Huang Q, et al. Mesenchymal stem cells alleviate bacteria-induced liver injury in mice by inducing regulatory dendritic cells. Hepatology. 2013;59(2):671–682. doi:10.1002/hep.26670
22. Idriss HT, Naismith JH. TNFα and the TNF receptor superfamily: structure‐function relationship(s). Microsc Res Tech. 2000;50(3):184–195. doi:10.1002/1097-0029(20000801)50:3<84::aid-jemt2><84::aid-jemt2>3.0.CO;2-H
23. Swaminathan S, Alirio J M. New developments on the TNFα-mediated signalling pathways. Biosci Rep. 2011;1–31(1):63–76. doi:10.1042/BSR20100040
24. Al-Lamki RS, Wang J, Skepper JN, Thiru S, Pober JS, Bradley JR. Expression of tumor necrosis factor receptors in normal kidney and rejecting renal transplants. Lab Invest. 2001;81(11):1503–1515. doi:10.1038/labinvest.3780364
25. Melania Manco MD, Matilde Marcellini MD, Germana Giannone G, Valerio Nobili V. Correlation of serum TNF-α levels and histologic liver injury scores in pediatric nonalcoholic fatty liver disease. Am J Clin Pathol. 2007;127(6):954–960. doi:10.1309/6VJ4DWGYDU0XYJ8Q
26. Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014;6(10):a016295. doi:10.1101/cshperspect.a016295
27. Heinrich PC, Castell JV, Andus T. Interleukin-6 and the acute phase response. Biochem J. 1990;265(3):621–636. doi:10.1042/bj2650621
28. Sproston NR, Ashworth JJ. Role of C-reactive protein at sites of inflammation and infection. Front Immunol. 2018;9:754. doi:10.3389/fimmu.2018.00754
29. Morrison SJ, Scadden DT. The bone marrow niche for haematopoietic stem cells. Nature. 2014;505(7483):327–334. doi:10.1038/nature12984.
30. Susanne K, Hermann E, Johannes S, Harald K, Karen B. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2012;24(5):1294–1301.
31. Gulati BR, Kumar R, Mohanty N, Kumar P, Somasundaram K, Yadav PS. Bone morphogenetic protein-12 induces tenogenic differentiation of mesenchymal stem cells derived from equine amniotic fluid. Cells Tissues Organs. 2013;198(5):377–389. doi:10.1159/000358231.
32. Kassis I, Zangi L, Rivkin R, et al. Isolation of mesenchymal stem cells from G-CSF-mobilized human peripheral blood using fibrin microbeads. Bone Marrow Transplant. 2006;37(10):967–976. doi:10.1038/sj.bmt.1705358.
33. Uchida K, Urabe K, Naruse K, Ujihira M, Mabuchi K, Itoman M. Comparison of the cytokine-induced migratory response between primary and subcultured populations of rat mesenchymal bone marrow cells. J Orthop Sci. 2007;12:484–492. doi:10.1007/s00776-007-1159-5.
34. Sotiropoulou PA, Perez SA, Salagianni M, Baxevanis CN, Papamichail M. Characterization of the optimal culture conditions for clinical scale production of human mesenchymal stem cells. Stem Cells. 2006;24(2):462–471. doi:10.1634/stemcells.2004-0331.
35. Klimenko NA, Tatarko SV, Shevchenko AN, Gubina-Vakulik GI. Justification of the model of chronic (secondary chronic) inflammation. Eksperim I Clinich Medicine No. 2007;2:24–28.
36. Necas J, Bartosikova L. Carrageenan: a review. Vet Med (Praha). 2013;58(4):187–205. doi:10.17221/6758-VETMED
37. Fehrenbacher JC, Vasko MR, Duarte DB. Models of inflammation: carrageenan- or complete freund’s adjuvant-induced edema and hypersensitivity in the rat. Curr Protoc Pharmacol. 2012;2012:4. doi:10.1002/0471141755.ph0504s56
38. Wang Y, Chen X, Cao W, Shi Y. “Plasticity of mesenchymal stem cells in immunomodulation: pathological and therapeutic implications,”. Nat Immunol. 2014;15(11):1009–1016. doi:10.1038/ni.3002
39. Kean TJ, Lin P, Caplan AI, Dennis JE. “MSCs: delivery routes and engraftment, cell-targeting strategies, and immune modulation,”. Stem Cells Int. 2013;2013:13. doi:10.1155/2013/732742
40. Ren G, Zhang L, Zhao X, et al. “Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide,”. Cell Stem Cell. 2008;2(2):141–150. doi:10.1016/j.stem.2007.11.014
41. Anzalone R, Iacono ML, Corrao S, et al. “New emerging potentials for human Wharton’s jelly mesenchymal stem cells: immunological features and hepatocyte-like differentiative capacity,”. Stem Cells Dev. 2010;19(4):423–438. doi:10.1089/scd.2009.0299
42. Abomaray FM, Al Jumah MA, Alsaad KO, et al. “Phenotypic and functional characterization of mesenchymal stem/multipotent stromal cells from decidua basalis of human term placenta,”. Stem Cells Int. 2016;2016:18. doi:10.1155/2016/5184601
43. Nejati M, Azami Tameh A, Vahidinia Z, Atlasi MA. Mesenchymal stem cells improve ischemic stroke injury by anti-inflammatory properties in rat model of middle cerebral artery occlusion. Iran Red Crescent Med J. 2018;20(1):e55085. doi:10.5812/ircmj.55085.
44. Nejati M, Tameh AA, Atlasi MA. Role of toll‐like receptors 2 and 4 in the neuroprotective effects of bone marrow–derived mesenchymal stem cells in an experimental model of ischemic stroke. J Cell Biochem. 2019;120:
45. Gao F, Chiu SM, Motan DA, et al. Mesenchymal stem cells and immunomodulation: current status and future prospects. Cell Death Dis. 2016;7(1):e2062. doi:10.1038/cddis.2015.327
46. Petryk N, Shevchenko O. Anti-inflammatory activity of mesenchymal stem cells in λ-carrageenan-induced chronic inflammation in rats: reactions of the blood system, leukocyte-monocyte ratio. Inflammation. 2020. doi:10.1007/s10753-020-01262-5
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.