Back to Journals » Biologics: Targets and Therapy » Volume 19
Mesenchymal Stem Cell Therapy in Osteoarthritis and Rheumatoid Arthritis: A Systematic Review of Exosomal microRNAs
Authors Hejazian SS, Hejazian SM, Mostafavi Montazeri SS, Abediazar S, Zununi Vahed S ![]() , Barzegari A
, Barzegari A
Received 30 September 2025
Accepted for publication 23 December 2025
Published 30 December 2025 Volume 2025:19 Pages 747—785
DOI https://doi.org/10.2147/BTT.S571417
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shein-Chung Chow
Seyyed Sina Hejazian,1,2 Seyyedeh Mina Hejazian,3 Seyede Saba Mostafavi Montazeri,4 Sima Abediazar,3 Sepideh Zununi Vahed,3 Abolfazl Barzegari5– 7
1Neurosciences Research Center, Tabriz University of Medical Sciences, Tabriz, Iran; 2Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran; 3Kidney Research Center, Tabriz University of Medical Sciences, Tabriz, Iran; 4Student Research Committee, Alborz University of Medical Sciences, Karaj, Iran; 5Department of Medical Biotechnology, Faculty of Advanced Medical Sciences, Tabriz University of Medical Sciences, Tabriz, Iran; 6Research Center for Pharmaceutical Nanotechnology (RCPN), Tabriz University of Medical Sciences, Tabriz, Iran; 7Laboratory for Vascular Translational Science, Nanotechnologies for Vascular Medicine and Imaging Team, Université Sorbonne Paris Nord, Villetaneuse, 93430, France
Correspondence: Sepideh Zununi Vahed, Kidney Research Center, Tabriz University of Medical Sciences, Tabriz, Iran, Email [email protected] Abolfazl Barzegari, Department of Medical Biotechnology, Faculty of Advanced Medical Sciences, Tabriz University of Medical Sciences, Tabriz, Iran, Email [email protected]
Summary: Osteoarthritis (OA) and rheumatoid arthritis (RA) are chronic diseases characterized by persistent inflammation and autoimmune responses that affect the joints and other organs. Scientific evidence indicates that the therapeutic effects of mesenchymal stem cells (MSCs) are mediated through the release of soluble factors and extracellular vesicles (EVs), particularly exosomes. The release of microRNAs from MSCs holds substantial potential for cell-free treatment in OA and RA.
Methods: A comprehensive search was conducted on Web of Science, PubMed, Scopus, and Google Scholar to identify relevant publications until 24 March 2024. The systematic review aimed at illuminating the current understanding of MSC-derived exosomal microRNAs, the origin of MSCs, potential mechanisms of action, and their therapeutic implications in managing OA and RA.
Results: A total of fifty-five articles (OA, n= 41 and RA, n= 14) were deemed eligible for inclusion in this study. Regardless of MSC origin, exosomal miRNAs could induce anti-inflammatory, protective, and chondroregenerative potential in in vitro and in vivo models of OA by targeting different signaling pathways. Fourteen studies have highlighted the role of MSC-derived exosomal miRNAs in modulating immune responses, reducing pro-inflammatory cytokine production, and potentially ameliorating synovial inflammation and joint symptoms associated with RA. To suppress joint inflammation and preserve cartilage, miR-140, miR-92a-3p, and miR-136-5p emerged as leading candidates for OA because they help restore the anabolic/catabolic balance and modulate key signaling pathways. For RA, the most effective candidates were miR-146a, miR-150-5p and miR-205-5p that target innate and adaptive immune signaling and synoviocyte activation, with NF-κB modulators such as miR-361-5p offering overlap across both diseases.
Conclusion: The mounting body of preclinical evidence supports that MSC-exosomal-miRNAs present a promising solution for OA and RA as a novel therapeutic strategy. However, human studies and more clinical trials are warranted.
Keywords: arthritis, extracellular vesicles, miRNAs, stem cells, MSCs, osteoarticular disease
Introduction
Osteoarthritis (OA) and rheumatoid arthritis (RA), the most common types of arthritis, are chronic inflammatory, autoimmune, and age-related osteoarticular diseases. They are characterized by the degeneration of cartilage and the inflammation of the synovial membrane, respectively. Both OA and RA involve inflammation and joint degeneration, although through different mechanisms.
Osteoarthritis is the most prevalent type of arthritis, involving over 250 million individuals worldwide.1,2 Currently, the prevalence of knee OA among the population aged over 40 is as high as 23%.3 At present, there are limited treatment optiones for knee OA, none of which can reverse cartilage degradation. Autologous chondrocyte implantation (ACI) is a technique used for larger cartilage defects.4 In patients with more severe knee OA, total knee arthroplasty (TKA) is a good choice with remarkable outcomes; however, the artificial joints require revision 15 to 25 years after surgery.5
Rheumatoid arthritis is considered a systemic autoimmune disorder due to leukocyte influx into joints that produce inflammatory mediators and destroy bone and cartilage.6 The incidence of RA worldwide is 40 per 100,000 individuals per year, the prevalence is ~0.24%7 and men are significantly more exposed.8 Over the last decade, therapeutic options for RA have been improved; however, their serious complications and low efficacy in some patients remain unsolved. Some RA patients remain untreated because of their resistance or intolerant reactions.9 Thus, there is a necessity to develop other therapeutic strategies for RA.
Mesenchymal stem cell (MSC) therapy is a much newer therapy for patients with OA/RA, which practically fills the gap between conventional pharmaceutical and surgical therapies. This treatment is available in an intra-articular manner with a relatively low cost, but the risk of non-optimal transformation of the transplanted cells and hyper-inflammatory reactions is always there.10 This is why scientists have recently proposed the use of cell-free therapeutic strategies such as MSC-derived extracellular vesicles (EVs), especially exosomes (MSC-Exos), to enhance the ideal therapeutic impact against cartilage and bone diseases via selectively administering the specified factors.11
It is worth mentioning that a part of the therapeutic effects of MSCs is attributable to EVs /exosomes, which carry selected cargo.12 Given recent advancements, these exosomes are currently exploited as effective drug delivery strategies that can be modified to carry particular agents like small non-coding microRNAs (miRNAs; miR) as regulators of gene expression and various biochemical pathways.
Although research on EVs and MSC-Exos in joint diseases is increasing rapidly, existing reviews either address EV cargoes in general or focus on a single condition, such as OA or RA. However, miRNAs packaged within MSC-Exos represent a distinct and biologically active subset of EV cargo with demonstrated roles in cartilage repair, immunomodulation, and bone remodeling in preclinical models of both OA and RA.13,14 This specific area has not yet been systematically reviewed, creating a clear gap in the current literature. Numerous experimental studies have shown that transferring defined miRNA cargos via MSC-derived exosomes can alter disease progression, highlighting their potential as diagnostic biomarkers and therapeutic candidates.15 However, published studies vary substantially in EV isolation, characterization, and miRNA analytic methods. Synthesizing evidence across OA and RA, therefore, offers a unique opportunity to identify miRNAs that function as shared mediators of joint pathology, as well as those specific to each disease. Such a cross-disease approach can reveal converging molecular mechanisms underlying cartilage degeneration and inflammation, which are not captured in single-disease reviews. In this review, the recent concepts of MSC derived exosomal-miRNAs are presented that can be applied to the treatment of joint disorders such as OA and RA.
microRNAs Carried by MSC-Derived Exosomes
The discovery of mRNAs and miRNAs being packed inside exosomes and shuttled between cells is a breakthrough, making these cargo-carrying components promising tools in various conditions. Since miRNAs participate in the regulation of inflammatory factors and chondrocyte homeostasis,16 they are important controllers in preserving a healthy joint, whereas Dicer knock-out mice are proof of this claim.17 Dicer is a crucial enzyme in the biosynthesis of miRNAs.
In both animal models and humans, dysregulated levels of miRNAs have been connected with osteoarticular diseases. Murata et al found that miRNA profiles of synovial fluid and plasma successfully discriminate RA from OA and can be used as diagnostic biomarkers. In synovial fluid of patients with RA, concentrations of miR-16, miR-146a, miR-223, and miR-155 were higher than those with OA significantly.18
Certainly, exosomal miRNAs have drawn greater attention than other molecules, given their functions in the regulation of gene expression. Besides, it is verified that a greater proportion of miRNAs is present in exosomes compared to their parent cells.19 More miRNA-profiling investigations have shown that the packaging of miRNAs into exosomes is done selectively. Accordingly, Guduric-Fuchs et al reported that subgroups of miRNAs, including miR-451, miR-142-3p, and miR-150, are preferentially wrapped inside exosomes.20 Thus, miRNAs are thought to be the key functional components in target cells.
Exosomal miRNAs are preserved against endogenous RNase-based degradation by the exosomal membrane. Their capacity for survival in body fluids and cellular culture medium makes them suitable for intra-articular administration. Further investigations have demonstrated that exosomal miRNAs are related to cartilage tissue development and homeostasis. A comparative microarray analysis between exosomes produced by undifferentiated human bone marrow-derived MSCs (hBM-MSCs) and cartilage-differentiating hBM-MSCs revealed that 35 miRNAs, consisting of miR-1290, miR-1246, miR-320c, miR-193a-5p, and miR-92a are upregulated in exosomes secreted from cartilage-differentiating hBM-MSCs.21 The same study also demonstrated that hBM-MSCs overexpressing exosomal miR-320c enhance the chondrocyte level of SOX9 (SRY-box transcription factor 9) and decrease MMP-13, leading to the expanded chondrogenesis of hBM-MSCs.21
Regarding the well-known cartilage regeneration role of miRNAs, miRNA (ie miR-155, miR-140, miR-136, miR-92a miR-26a, and miR-9) -overexpressing exosomes indicate greater therapeutic perspective in the recipient cells in comparison to the naive exosomes.13,22–25
Methods
Search Strategy and Inclusion and Exclusion Criteria
The systematic review was carried out to examine the effects of exosomal miRNAs derived from MSCs on arthritis models, following the PRISMA guidelines. Various databases including Web of Sciences, PubMed, Scopus, and Google Scholar were searched with no time restriction, up to 24 March 2024 by a qualified librarian. The search strategy utilized terms and MeSH-based keywords, are presented in the Supplementary Table S1. In this review, original research articles in English focusing on preclinical in vitro and animal models of arthritis, osteoarthritis, and rheumatoid arthritis utilizing MSCs secretome (exosomal miRNAs) were considered. Abstracts, review articles, non-English articles, retracted articles, and those with low quality were excluded. Two researchers independently reviewed records from the primary search (SMH and SSMM) with a third researcher resolving any discrepancies. Data extraction was performed by two researchers. The extracted information can be found in Tables 1–4.
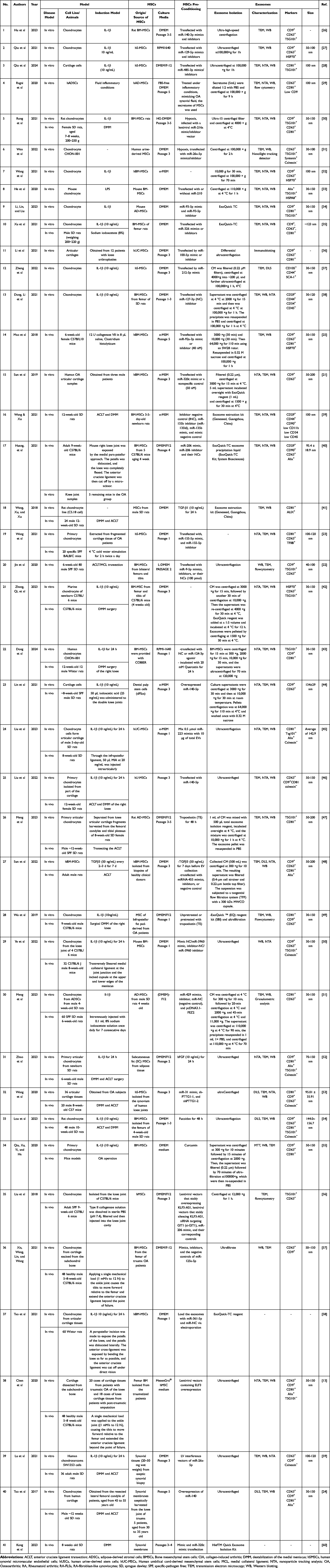 |
Table 1 In vitro/in vivo Osteoarthritis Models Applied MSC-Derived Exosomal microRNAs |
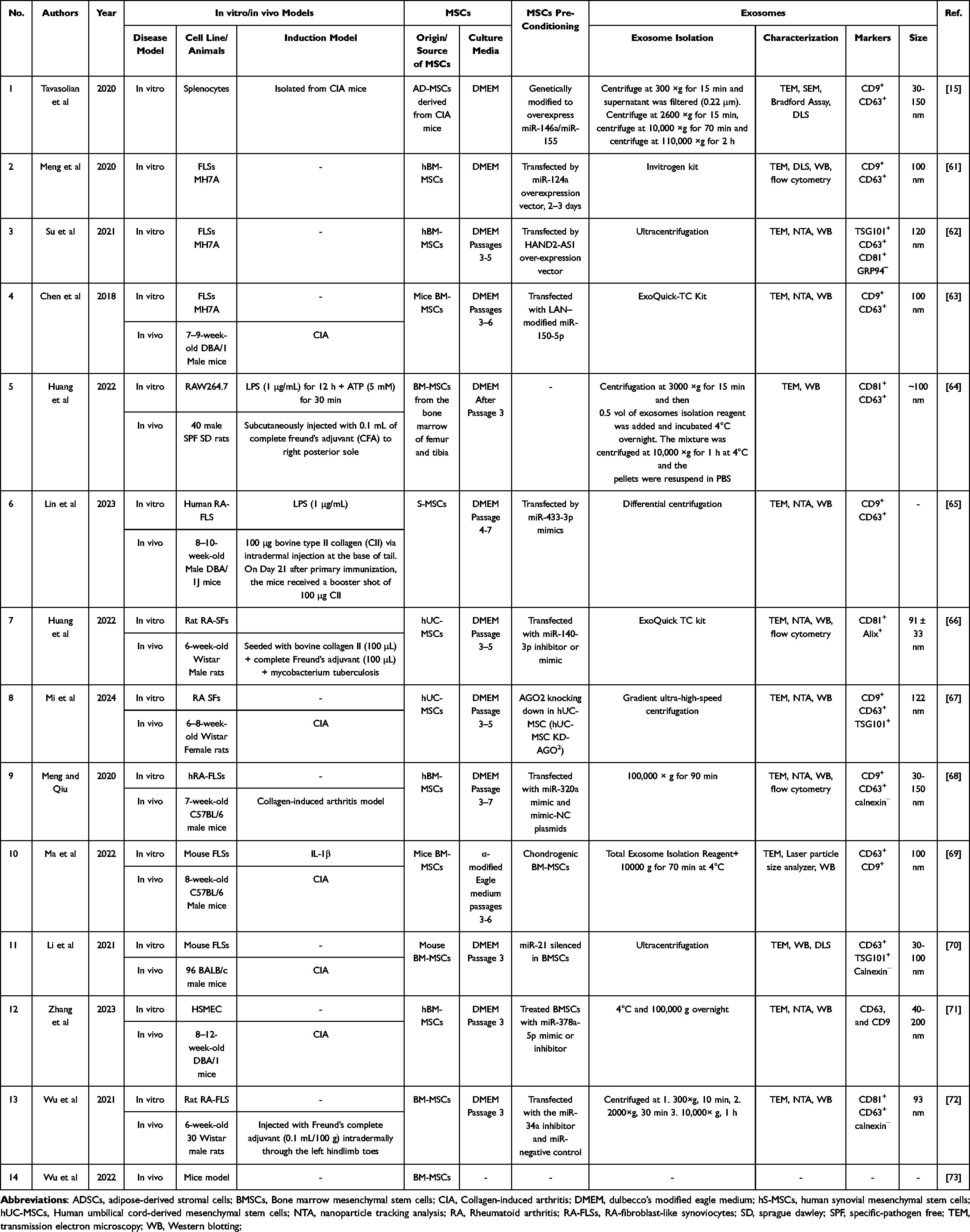 |
Table 2 In Vitro/in Vivo Rheumatoid Arthritis Models Applied MSC-Derived Exosomal microRNAs |
 |
Table 3 Preclinical Studies Applied MSC-Derived Exosomal microRNAs for Osteoarthritis-Additional Information |
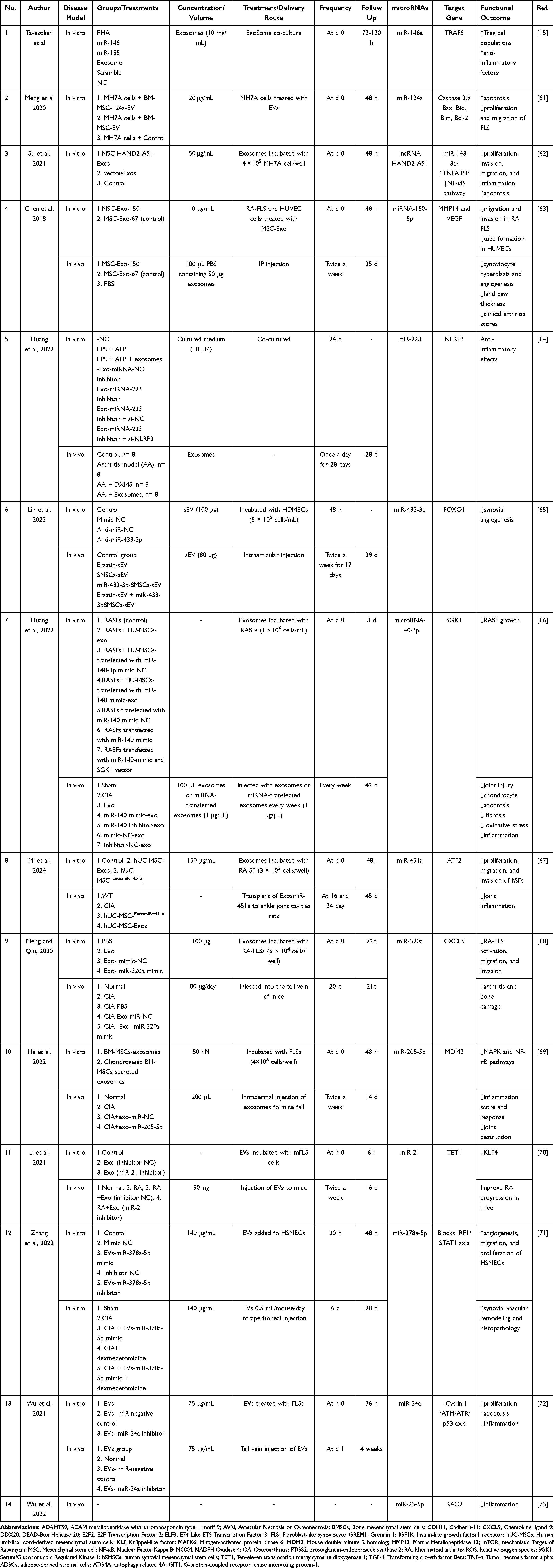 |
Table 4 Preclinical Studies Applied MSC-Derived Exosomal microRNAs for Rheumatoid Arthritis-Additional Information |
Assessment of the Quality of Studies
The quality of the studies included in the review was assessed based on the ARRIVE guidelines 2.0 and the risk of bias was evaluated using SYRCLE’s risk of bias tool.
Results
Included Studies
The initial search yielded 447 articles, with 191 duplicates removed. One hundred and seven reports were screened through abstracts. Stem cell-based therapies, review articles, and irrelevant and retracted studies were excluded. Fifty-five articles were included in the review based on our criteria.13,15,21–49,51–73 The PRISMA flow diagram in Figure 1 outlines the literature search process.
 |
Figure 1 PRISMA flow diagram of the literature search and selection process. *Web of Sciences, PubMed, Scopus, and Google Scholar. |
MSC Sources and Pre-Conditioning Methods to Obtain Engineered Exosomes in OA
Included OA articles were twelve in vitro,21,26–29,31–34,36–38 four in vivo,22,25,39,60 and twenty-five mixed in vivo+ in vitro studies, Table 1. Different types of chondrocyte cell lines, including C5.18 and CHON-001 or isolated chondrocytes from cartilage of OA patients, were used in cell culture studies. IL-1β or sodium iodoacetate was used to induce OA in most of these studies. The anterior cruciate ligament transection (ACLT) and destabilization of the medial meniscus (DMM) surgeries were commonly used surgical models to induce OA in animal studies. Additionally, other methods such as monosodium iodoacetate (MIA) injection45 and collagenase VII25,56 administration were employed to create OA animal models.
Among OA studies, BM-MSCs were the most utilized cells.13,21,22,25,26,30,32,33,35,38–40,42,43,48,50,54,55,57,58 Beyond BM-MSC, S-MSCs,23,24,27,28,37,53,59,60 AD-MSCs,29,34,47,51,52 urine-derived MSCs,31,46 umbilical-cord MSCs (UC-MSCs),36,45 infrapatellar fat pad derived MSCs (IPFP-MSCs),49 dental pulp-derived MSCs,44 and undefined MSCs41,56 were utilized in OA studies. In most of the OA studies, MSCs were pre-conditioned with hypoxia,30,31 TGF-β,41,48 curcumin,55 fucoidan,54 bFGF,52 quercetin,43 or transfected with mimic/inhibitory miRNAs before EV/exosome isolation, Table 1.
MSC Sources and Pre-Conditioning Methods to Obtain Engineered Exosomes in RA
Fourteen studies in total were conducted to assess the impact of MSC-derived EVs on the treatment of RA, with a detailed description of their characteristics found in Table 2. Among these studies, three were in vitro studies,15,61,62 ten utilized a combination of methodologies (in vitro+ in vivo,63–72 and one was in vivo study.73 The MH7A cell line was selected as a dependable in vitro model for replicating Fibroblast-like synoviocytes from RA patients. Various animal models, including mice (n=6)63,65,68–71,73 and rats (n=4)64,66,67,72 were employed. Collagen-induced arthritis (CIA) was the most widely used model for studying RA.
The included RA studies showcased a range of MSC types. The most commonly utilized MSCs were BM-MSCs (n=10),61–64,68–73 followed by umbilical cord MSCs (UC-MSCs),66,67 AD-MSCs,15 and synovial MSCs (S-MSCs).65 In most studies, MSCs were transfected with specific miRNA mimics or inhibitors to generate engineered exosomes, Table 2.
Isolation and Characterization of Exosomes
The ultracentrifugation technique was predominantly used to extract exosomes from MSCs, although some studies opted for different exosome extraction kits like Invitrogen,61 ExoQuick-TC,21,34,35,40,42,49,58,63,66 or HiefTM60 kits. Transmission electron microscopy (TEM), Western blotting, and nanoparticle tracking analysis were utilized to examine the structure, presence of exosome-related markers, and size distribution, respectively. The extracted MSC exosomes typically exhibited a bilayer lipid membrane structure under the TEM, showcased a size distribution ranging from around 30 to 200 nm, and harbored exosome-related markers, including endosomal proteins ALIX and TSG101, tetraspanins CD81, CD63, and CD9 (Table 1 and Table 2).
Delivery Routes and Treatment Frequency for miRNAs Derived from MSC-Exosomes
A wide range of exosome concentrations, ranging from 20–200 μg/μL, were utilized across the studies. The intraperitoneal, intraarticular, intradermal, and tail vein routes were employed for the administration of exosomes. Additionally, in one investigation, exosomes were transplanted into ankle joint cavities.67 Furthermore, in certain instances, the concentration of exosomes was unspecified, with only the total quantity of injected exosomes being referenced. The exact dosage and route of administration were not elucidated in some studies. In animal experiments, the duration of observation varied between 14 and 45 days after treatment (Table 3 and Table 4).
Outcomes/Improvement
Generally, the results indicated that the administration of MSC-exosomal miRNAs had a positive impact on cartilage regeneration and cartilage repair in the defect area. Moreover, studies illustrated that MSC-secretome therapy of OA patients could reduce apoptosis, pyroptosis of cartilage, ROS production, pathological severity, inflammation, and degrading of the extracellular matrix in the chondrocytes. This therapy could also increase the production of cartilage, homeostasis, chondrocyte proliferation, differentiation, migration, autophagy of chondrocytes (Table 3). Based on the obtained results of experimental RA studies, it was observed that MSC-secretome decreased proliferation, invasion, migration, inflammation, hyperplasia, angiogenesis, and arthritis scores of synoviocyte cells (Table 4).
Quality and Risk of Bias Assessment
Considerable heterogeneity was observed among the included studies in several key methodological aspects. Various sources of MSCs, including bone marrow, adipose tissue, umbilical cord, and synovium, potentially influence the composition and biological potency of secreted exosomes. Exosome isolation and characterization techniques were not standardized; studies employed differential ultracentrifugation, precipitation kits, size-exclusion chromatography, or microfiltration, leading to variations in particle purity and yield. Animal models ranged from chemically induced to surgically created OA and RA models, with differences in disease severity and duration that may affect treatment response. Likewise, the dosage and administration routes of exosomes (intra-articular injection, intravenous infusion, or local tissue delivery) varied substantially, limiting direct comparability across studies. These methodological inconsistencies contribute to variability in reported outcomes and complicate the identification of dose-response relationships or optimal treatment regimens. As a result, while consistent beneficial effects of MSC-derived exosomal miRNAs were reported, caution is warranted when generalizing these findings across experimental systems or translating them to clinical settings.
In the included studies, the main risk of bias was the lack of clear information about the blinding and randomization process in the performance and detection phase (Figure 2). Blinding of investigators and outcome assessors was rarely implemented, particularly for histological scoring or behavioral analyses, which represent a risk of detection bias. Only a few studies provided sample size justification, technical replicates, or independent repeat experiments, which limit reproducibility.
 |
Figure 2 Risk of bias chart. |
Discussion
MSC-Derived EVs in Osteoarthritis
The regenerative and immunomodulatory role of MSC-EVs in OA has attracted the attention of many researchers. Accordingly, several studies have revealed the significant therapeutic role of MSC-EVs in OA by suppressing degenerative pathways and specific pro-inflammatory elements and improving the migration and proliferation of chondrocytes.74,75 By inhibiting apoptosis, pyroptosis, and ferroptosis, they can inhibit chondrocyte death. These cargo-carrying components are also capable of improving chondrogenesis and cellular differentiation and stimulating autophagy to preserve chondrocyte integrity, which is directly correlated with chondrocyte regeneration and efficient metabolism (reviewed by Wu et al).76 MSC-derived exosomes have also been demonstrated to reduce the anabolic capacity of chondrocytes.24 It is also reported that overexpressing miR-140 in urine-derived MSCs could ease knee OA in a rat model by downregulation of endothelial growth factor A.46
Vonk et al revealed that MSC-EVs are capable of blocking NF-κB signaling by suppressing phosphorylation of IκBα, leading to downregulation of TNF-α (tumor necrosis factor-alpha)-derived COX2 activity, cytokine-mediated responses, and collagenase function.74 Additionally, MSC-EVs are involved in the upregulation of SOX9 and WNT7A expression. In vitro synthesis of proteoglycan and type II collagen is enhanced by MSC-EVs.74 Induction of OA-like chondrocytes with MSC-EVs is associated with an elevated level of chondrogenesis (type II collagen and aggrecan) indicators, and reduced level of catabolic (MMP-13), and inflammatory (iNOS) factors.75 In another study, the MSC-EV treatment of OA in mouse models was associated with decreased cartilage and bone degradation.77 Ruiz et al also demonstrated that the therapeutic impact of MSC-EVs in a mouse model of OA was partially regulated by TGF-βI.78 Further studies have shown that miR-206, miR-212-5p, and miR-136-5p present in the MSCs-derived exosomes enhance chondrocyte migration, prevent chondrocyte degeneration, and decline inflammatory responses in OA models by targeting E74-like factor 3 (ELF3).13,37,40 ELF3 is an inflammatory mediator activated by pro-inflammatory cytokines in chondrocytes, which elevates the expression level of inflammatory and catabolic pathways, involved in cartilage degradation in OR. EVs obtained from MSCs with overexpressed circHIPK3 (EVs-circHIPK3) are associated with promotion of chondrocyte migration and proliferation and suppression of chondrocyte apoptosis induced by the miR-124/MYH9 pathway.79
Exosomal-miRNAs Derived from BM-MSCs in Osteoarthritis
Various studies have demonstrated that BM-MSC-exosomal miRs have therapeutic effects on OA by attenuating inflammatory-mediated cartilage injury, promoting proliferation and migration and suppressing apoptosis of chondrocytes in OA joints. BM-MSC-exsosomal miRNAs, such as miR-136-5p,13 miR-6515-5p32 prevent the apoptosis and senescence of chondrocytes. Surprisingly, one study found that in HIF-1α-induced BM-MSC-EVs, hypoxia results in overexpression of miR-216a, leading to migration and proliferation, and a drop in apoptosis of chondrocytes in OA (through suppression of the JAK2 and STAT3 signaling pathway).30 Hu et al indicated that under hypoxia, the overexpression of miR-140-3p in exosomes derived from BM-MSCs could significantly increase migration, survival rate, and chondrogenic differentiation in inflammatory chondrocytes and consequently delay OA pathogenesis and protect joints.26 On the whole, BM-derived exosomal miR-9-5p, miR-92a-3p, miR-127-3p, miR-210, miR-135b, miR-326, miR-206, miR-361-5p, miR-347-3b, miR-124-3p, miR-146-5p, miR-124, miR-143, miR-455, miR-125a-5p, and miR-3960 by decreasing inflammation, cell death, and inducing the proliferation and migration of chondrocytes represent therapeutic effects on OA.21,22,25,26,30,33,35,38,40,42,43,48,50,54,55,57,58,72
Exosomal-miRNAs Derived from AD-MSC in Osteoarthritis
AD-MSCs-exosomes are uptaken by chondrocytes80 and reduce the catabolic state in these cells,81 making them potential therapeutic candidates for OA. Experiments on animal models of OA have demonstrated that AD-MSCs-EVs are capable of suppressing inflammation by inhibiting M1 macrophage infiltration to joint synovium and protecting against cartilage degeneration in both subacute and chronic animal models of OA.81 Early stages of OA are associated with inflammatory-mediated catabolism in joint cartilage by proteases that can be reversed by IR injection of AD-MSCs-EVs. They repress the progression of OA, which is mediated by enhancing collagen II synthesis and downregulation of ADAMTS-5, MMP-13, MMP-3, and MMP-1 in the presence of IL-1β.81 AD-MSCs-EVs are also capable of neutralizing the IL-1β-mediated inflammatory responses by inhibiting p65 in the NF-κB signaling pathway.82 Exosomes derived from pretreated AD-MSCs with Tropoelastin could promote cartilage repair in animal models of OA and preserve the chondrocyte phenotype in cell culture through miR-451-5p.47
Comparing the effect of AD-MSCs-EVs on chondrocytes and synovial cells showed that the uptake of exosomes by synovial cells happens quicker with greater intensity, which leads to a better modulation of secretion of interleukins, catabolic, angiogenesis, and pain factors in synovial cells. Moreover, the absorption of exosomes by synovial cells is associated with the release of secondary vesicles from these cells that improve the biological function of exosomes via positive feedback.82
IPFP-MSCs are a type of AD-MSCs that have already been demonstrated to contribute to chondrogenesis in OA.83 It is now being proposed that a part of this impact may be due to IPFP-MSCs-derived exosomes. In vitro studies have shown that these miR-100-5p-enriched exosomes contribute to the suppression of apoptosis and improvement of matrix synthesis by inhibiting the mTOR autophagy pathway, leading to reduced cartilage degradation and gait abnormalities in DMM-induced OA models.49 Other in vitro investigations have proposed that IPFP-MSC-EVs improve chondrogenesis in periosteal cells by enhancing the miR-145 and miR-221 expression and downregulating the synthesis of pro-inflammatory cytokines.84 Overexpressed exosomal miR-199a-3p in chondrocytes is shown to target the 3ʹ UTR region of mTOR mRNA and reduce protein levels of mTOR and its downstream factors along with increasing chondrocyte autophagy, enhance anabolism and suppressing catabolism for the OA treatment.52
Exosomal-miRNAs Derived from Umbilical Cord-MSC in Osteoarthritis
UC-MSCs have the advantages of easy access, minor immune rejection, and significant immunity-regulating effects, which make them a promising treatment for OA. Researchers have found that UC-MSCs-exosomes can prevent apoptosis and ROS production. This may be due to inhibition of NOX4 expression by exosomal miR-100-5p.36 miR-223, which directly interacts with the 3’ untranslated region of NLRP3 mRNA, has been identified as a pivotal miR that facilitates the beneficial effects of hUC-EVs in the realms of inflammation suppression and cartilage protection. The dual-engineered EVs demonstrated a maximal capacity for inhibiting chondrocyte pyroptosis and NLRP3 inflammasome activation, thereby yielding remarkable outcomes in the therapeutic management of osteoarthritis.45
Exosomal-miRNAs Derived from Synovial-MSC in Osteoarthritis
Liu et al reviewed the potential roles of EVs and their combination with biomaterials in OA pathogenesis, diagnosis, and therapy.85 Tao et al indicated that miR-140-5p overexpression could block Wnt-5a and −5b signaling pathway that activates YAP and decreases ECM secretion.24 In rat OA models induced by cold-water stimulation, OA chondrocytes were treated with overexpressed miR-155-5p S-MSCs-exosomes. This therapy could enhance the release of ECM, cellular proliferation and migration and diminish cell apoptosis, effectively prohibiting OA from happening.23 Moreover, it is indicated that overexpressed miR-155-5p SMSCs-exosomes could target Runx2 and increase ECM secretion, causing increased cartilage regeneration and decreased OA.23 Wang et al found that overexpressed miR-31 S-MSCs-EVs could mitigate OA by regulating the KDM2A (lysine demethylase 2A)/E2F1/PTTG1 (pituitary tumor-transforming gene 1) pathway.53 It is also reported that S-MSC-exosomal miR-129 could inhibit the IL-1β-mediated OA by preventing the release of high mobility group protein −1 (HMGB1).27 Kong et al found that S-MSC-exosomal miR-320c prevents chondrocyte apoptosis and ECM degradation by targeting ADAM19-dependent Wnt signaling to assist cartilage injury repair in OA rats.60 Modified exosomes derived from miR-155-overexpressing S-MSCs by reducing apoptosis and modulating ECM in chondrocytes could prevent OA.23
MSC-Derived EVs in Rheumatoid Arthritis
BM-MSC-EVs, like their parental cells, have promising therapeutic effects against RA.86 The BM-MSC-Exosomal RNAs are absorbed by fibroblast-like synovial cells (FLSs) and prevent inflammation during RA treatment. It is found that RA-FLS inflammation and proliferation were decreased by miR-34 in BM-MSC-EVs through the prevention of cyclin I/ATM/ATR/p53 signaling pathway.72 Exosomal miR-320 derived from BM-MSC significantly downregulates the chemokine ligand CXCL9 and preventes the invasion, migration, and activation of RA-FLSs.68 FLS regulation and angiogenesis inhibition occur by exosomal miR-150-5p, resulting in downregulated levels of MMP14 and VEGF.63 It is also reported that the combination of HAND2-AS1 (a long non-coding RNA) with BM-MSC-EVs could suppress RA progression by inhibiting the RA-FLSs tumor-like behavior through the miR-143/TNFAIP3/NF-κB pathway.62 MiRNA-223 in BM-MSCs-exosomes regulates the activation of inflammasomes by targeting NLRP3 in macrophages of RA models.66 Generally, BM-MSC-EVs are shown to protect against RA but more research is needed to clarify their anti-inflammatory mechanisms and immune regulatory properties in RA.61–64,66,68–73
Yin et al indicated that miR-99 b-3p in exosomes of subcutaneous AD-MSCs could regulate the cartilage ECM degradation via targeting ADAMTS4 during OA treatment.87 Li et al revealed that human AD-MSC-exosomal miR-376c-3p could inhibit the Wnt-β-catenin pathway and then alleviate synovial fibrosis and chondrocyte degradation in an OA animal model.65 Altogether, the results showed that AD-MSCs-EVs could be a novel proper alternative RA treatment.
Upregulated exosomal miR-140 in UC-MSCs reduced joint damage of RA rats through targeting serum- and glucocorticoid-inducible kinase 1.66
Across MSC sources, a convergent set of intracellular signaling pathways emerges as the principal targets of exosomal miRNAs in OA and RA. Many of the most frequently reported miRNAs (such as miR-140-5p, miR-146a, miR-21, miR-92a-3p, and miR-100-5p) modulate key regulatory cascades that collectively govern inflammation, cartilage matrix turnover, and cell survival. NF-κB signaling is the most consistently inhibited pathway, underpinning the anti-inflammatory and immunosuppressive effects observed across studies. Several miRNAs (eg, miR-146a, miR-21, miR-320a) attenuate NF-κB activity by targeting upstream mediators such as TRAF6 and IRAK1, thereby reducing cytokine production and synovial inflammation. Wnt/β-catenin and PI3K/AKT/mTOR pathways are recurrently modulated to promote anabolic and anti-apoptotic effects in chondrocytes: miR-92a-3p and miR-100-5p restore cartilage matrix synthesis via activation of SOX9 and COL2A1, while inhibition of mTOR signaling by miR-100-5p enhances autophagy and cell survival. In addition, suppression of MAPK/ERK and TGF-β/Smad cascades contributes to reduced catabolic enzyme expression (MMP-13, ADAMTS-5) and fibrosis. Together, these pathways form a core regulatory network through which MSC-derived exosomal miRNAs exert anti-inflammatory, pro-regenerative, and cytoprotective effects, regardless of MSC tissue origin.
Prospects and Challenges in the Clinical Application of MSCs-Exosomal miRNAs in Arthritis
Based on previous preclinical studies, although cell-free exosomes have protective effects in OA/RA, their efficacy is only at early stages and several problems should be addressed. For example, the types and content of molecules in exosomes/EVs can be different depending on the tissue source, donor characteristics, and culture conditions of the parent MSCs. These points can alter exosomal miRNA profiles and consequently influence therapeutic potency and reproducibility. This point can impact MSC-exosomal function at recipient cells and cause alterations in physiological processes.
The efficiency of exosomal content delivery to chondrocytes is influenced by the exosome diameter (30–150 nm) because the average pore size of the articular cartilage ECM is about 6.0 nm88 and only small cationic nanocarriers (<15 nm in diameter) can cross over this biological barrier.89 During in vivo studies, the thickness of cartilage in small animal models is lower than humans90 and cultured chondrocytes are investigated instead of full-thickness cartilage explants in most time during in vitro studies. These inconsistencies should be examined before the application of exosomes in the clinic.
The pleiotropic nature of miRNAs means that each can regulate multiple target genes; thus, off-target effects may occur if exosomal miRNAs inadvertently modulate pathways unrelated to joint repair, altering immune or metabolic functions in distant tissues. This concern is further heightened by evidence that repeated or high-dose administrations of MSC-derived exosomes could elicit unexpected immune activation or antibody formation, particularly if exosomes are derived from allogeneic sources. Moreover, key safety parameters such as dose-dependent toxicity and biodistribution remain poorly defined. Most preclinical studies use variable exosome concentrations and delivery routes without standardized pharmacokinetic evaluation. These issues make it difficult to predict or control these off-target and immunological risks.
The most important problem regarding the use of miRNAs is the delivery of appropriate amounts of selective miRNAs to save and repair cartilage tissue. The engineering of exosomes and their cargo, such as MSC pre-conditioning, exosome surface modification, and miRNA enrichment would be an effective therapy for RA and OA. Surface or genetic modification can enable targeted delivery of exosomes to inflamed synovial tissue or degenerative cartilage, thereby improving local uptake and minimizing off-target effects. Encapsulating MSC-derived exosomes within alginate hydrogels, collagen scaffolds, or chitosan-based matrices can allow sustained release and prolonged residence time at the target joint site. This point can address the rapid clearance typically observed after intra-articular injection.91 Future studies should therefore investigate optimal hydrogel formulations and release kinetics, as well as evaluate biocompatibility and functional outcomes in large-animal and clinical models. Comparative studies assessing exosomes derived from different MSC sources are warranted to determine which cell type yields the most functionally potent and clinically scalable exosomes. It is also important to investigate whether the complex regulatory network of miRNAs and their targets can initiate other diseases necessities further study.92
Because of the local injection of exosomes mostly into the articular cavity, it is much safer than systematic administration and less toxic as cell-released products. Most in vivo MSC-exosome therapeutic studies were performed in small animals with less focus on their safety in arthritis treatment. Presently, insufficient evidence from clinical trials and preclinical studies delays the translation of MSC-exosomal therapies to clinical applications.
From a manufacturing and translational point, large-scale production of clinical-grade exosomes presents significant challenges (reviewed in Ref.93). Scaling up MSC culture while maintaining cell viability, consistent phenotype, and exosome yield requires bioreactor-based culture systems and stringent Good Manufacturing Practice (GMP) compliance. Additionally, the lack of standardized protocols for exosome isolation and purification leads to variability in purity, particle concentration, and cargo integrity. Developing robust quality control assays to characterize exosomal size distribution, miRNA content, and bioactivity is essential for ensuring batch-to-batch consistency.
Exosome stability and storage also remain unresolved issues. Freeze–thaw cycles, lyophilization, and prolonged storage can alter exosomal membrane integrity and reduce biological activity.93 Establishing validated methods for long-term preservation without functional loss will be critical for clinical manufacturing and distribution. Comparative profiling of MSC-derived exosomal miRNAs from healthy donors and arthritis patients, using sequencing or microarray approaches, may also identify disease-specific cargo signatures suitable for precision therapies. Addressing these challenges through standardized manufacturing protocols, rigorous dose optimization, and in vivo safety studies will be essential before clinical translation.
Conclusion
MSC-derived exosomal miRNAs represent a promising, low-risk, cell-free therapeutic strategy for OA and RA, combining the biological potency of stem cells with the safety and scalability required for clinical translation. In preclinical OA models, MSC-exosomal miR-92a-3p, miR-140-5p, miR-199a-3p, miR-95-5p, miR-320c and miR-100-5p are the most consistently reported therapeutic miRNAs. They protect cartilage and inflammation by targeting WNT5A/Wnt, ADAMTS/MMP, MAPK/NF-κB, HDAC2/8→SOX9, ADAM-dependent Wnt and NOX4/mTOR (oxidative-stress/autophagy) pathogenic axes, respectively. In RA preclinical models, MSC-derived exosomes carrying miR-150-5p, miR-146a-5p, and miR-205-5p have demonstrated potent anti-inflammatory, anti-angiogenic, and joint-protective effects by downregulating key pathogenic mediators (MMP14, VEGF, MDM2) and reprogramming immune responses (eg, enhancing Treg, suppressing NF-κB/MAPK).
Their intrinsic ability to cross biological barriers and their low immunogenicity make MSC-Exos an ideal platform for cell-free therapeutic development. Translationally, the identified miRNAs offer two major opportunities. First, as therapeutic targets, they can be enriched within exosomes through pre-conditioning of MSCs or encapsulated into advanced delivery systems to enhance joint repair and immune regulation. Second, as biomarkers, specific exosomal miRNA signatures may be used to monitor disease activity, predict treatment response, or stratify in future clinical trials. Future work should prioritize validating the most consistent miRNA candidates in human samples and developing standardized production and quantification methods to accelerate their path toward therapeutic and diagnostic application.
Abbreviations
ADAMTS9, ADAM metallopeptidase with thrombospondin type 1 motif 9; AVN, Avascular Necrosis or Osteonecrosis; BMSCs, Bone mesenchymal stem cells; CDH11, Cadherin-11; CXCL9, Chemokine ligand 9; DDX20, DEAD-Box Helicase 20; E2F2, E2F Transcription Factor 2; ELF3, E74 Like ETS Transcription Factor 3; FLS, Fibroblast-like synoviocyte; GREM1, Gremlin 1; IGF1R, Insulin-like growth factor1 receptor; hUC-MSCs, Human umbilical cord-derived mesenchymal stem cells; KLF, Krüppel-like factor; MAPK6, Mitogen-activated protein kinase 6; MDM2, Mouse double minute 2 homolog; MMP13, Matrix Metallopeptidase 13; mTOR, mechanistic Target of Rapamycin; MSC, Mesenchymal stem cell; NF-κB, Nuclear Factor Kappa B; NOX4, NADPH Oxidase 4; OA, Osteoarthritis; PTGS2, prostaglandin-endoperoxide synthase 2; RA, Rheumatoid arthritis; ROS, Reactive oxygen species; SGK1, Serum/Glucocorticoid Regulated Kinase 1; hSMSCs, human synovial mesenchymal stem cells; TET1, Ten-eleven translocation methylcytosine dioxygenase 1; TGF-β, Transforming growth factor Beta; TNF-α, Tumor necrosis factor alpha; ADSCs: adipose-derived stromal cells, ATG4A: autophagy related 4A, GIT1: G-protein-coupled receptor kinase interacting protein-1.
Data Sharing Statement
Data will be made available upon a reasonable request.
Acknowledgments
The authors declare that they have not used AI-generated work in this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Tabriz University of Medical Sciences, Tabriz, Iran (Grant # 72684).
Disclosure
The authors confirm that there is no conflict of interest related to the manuscript.
References
1. Nelson AE, Allen KD, Golightly YM, Goode AP, Jordan JM. A systematic review of recommendations and guidelines for the management of osteoarthritis: the chronic osteoarthritis management initiative of the U.S. bone and joint initiative. Semin Arthritis Rheum. 2014;43(6):701–712. doi:10.1016/j.semarthrit.2013.11.012
2. Bannuru RR, Schmid CH, Kent DM, Vaysbrot EE, Wong JB, McAlindon TE. Comparative effectiveness of pharmacologic interventions for knee osteoarthritis: a systematic review and network meta-analysis. Ann Intern Med. 2015;162(1):46–54. doi:10.7326/m14-1231
3. Cui A, Li H, Wang D, Zhong J, Chen Y, Lu H. Global, regional prevalence, incidence and risk factors of knee osteoarthritis in population-based studies. EClinicalMedicine. 2020;29-30:100587. doi:10.1016/j.eclinm.2020.100587
4. Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl J Med. 1994;331(14):889–895. doi:10.1056/nejm199410063311401
5. Evans JT, Walker RW, Evans JP, Blom AW, Sayers A, Whitehouse MR. How long does a knee replacement last? A systematic review and meta-analysis of case series and national registry reports with more than 15 years of follow-up. Lancet. 2019;393(10172):655–663. doi:10.1016/s0140-6736(18)32531-5
6. Scott DL, Wolfe F, Huizinga TW. Rheumatoid arthritis. Lancet. 2010;376(9746):1094–1108. doi:10.1016/s0140-6736(10)60826-4
7. Cross M, Smith E, Hoy D, et al. The global burden of rheumatoid arthritis: estimates from the global burden of disease 2010 study. Ann Rheum Dis. 2014;73(7):1316–1322. doi:10.1136/annrheumdis-2013-204627
8. Krasselt M, Baerwald BC. Sex, Symptom Severity, and Quality of Life in Rheumatology. Clin Rev Allergy Immunol. 2019;56(3):346–361. doi:10.1007/s12016-017-8631-6
9. Singh JA, Saag KG, Bridges SL Jr, et al. 2015 American College of Rheumatology Guideline for the Treatment of Rheumatoid Arthritis. Arthritis Rheumatol. 2016;68(1):1–26. doi:10.1002/art.39480
10. Lalu MM, McIntyre L, Pugliese C, et al. Safety of cell therapy with mesenchymal stromal cells (SafeCell): a systematic review and meta-analysis of clinical trials. PLoS One. 2012;7(10):e47559. doi:10.1371/journal.pone.0047559
11. Pourakbari R, Khodadadi M, Aghebati-Maleki A, Aghebati-Maleki L, Yousefi M. The potential of exosomes in the therapy of the cartilage and bone complications; emphasis on osteoarthritis. Life Sci. 2019;236:116861. doi:10.1016/j.lfs.2019.116861
12. Witwer KW, Van Balkom BWM, Bruno S, et al. Defining mesenchymal stromal cell (MSC)-derived small extracellular vesicles for therapeutic applications. J Extracell Vesicles. 2019;8(1):1609206. doi:10.1080/20013078.2019.1609206
13. Chen X, Shi Y, Xue P, Ma X, Li J, Zhang J. Mesenchymal stem cell-derived exosomal microRNA-136-5p inhibits chondrocyte degeneration in traumatic osteoarthritis by targeting ELF3. Arthritis Res Ther. 2020;22(1):256. doi:10.1186/s13075-020-02325-6
14. Wu Y, Li J, Zeng Y, et al. Exosomes rewire the cartilage microenvironment in osteoarthritis: from intercellular communication to therapeutic strategies. Int J Oral Sci. 2022;14(1):40. doi:10.1038/s41368-022-00187-z
15. Tavasolian F, Hosseini AZ, Soudi S, Naderi M. miRNA-146a improves immunomodulatory effects of MSC-derived exosomes in rheumatoid arthritis. Current Gene Ther. 2020;20(4):297–312. doi:10.2174/1566523220666200916120708
16. Mirzamohammadi F, Papaioannou G, Kobayashi T. MicroRNAs in cartilage development, homeostasis, and disease. Curr Osteoporos Rep. 2014;12(4):410–419. doi:10.1007/s11914-014-0229-9
17. Kobayashi T, Lu J, Cobb BS, et al. Dicer-dependent pathways regulate chondrocyte proliferation and differentiation. Proc Natl Acad Sci U S A. 2008;105(6):1949–1954. doi:10.1073/pnas.0707900105
18. Murata K, Yoshitomi H, Tanida S, et al. Plasma and synovial fluid microRNAs as potential biomarkers of rheumatoid arthritis and osteoarthritis. Arthritis Res Ther. 2010;12(3):R86. doi:10.1186/ar3013
19. Goldie BJ, Dun MD, Lin M, et al. Activity-associated miRNA are packaged in Map1b-enriched exosomes released from depolarized neurons. Nucleic Acids Res. 2014;42(14):9195–9208. doi:10.1093/nar/gku594
20. Guduric-Fuchs J, O’Connor A, Camp B, c l O, Medina RJ, Simpson DA. Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genomics. 2012;13(1):357. doi:10.1186/1471-2164-13-357
21. Sun H, Hu S, Zhang Z, Lun J, Liao W, Zhang Z. Expression of exosomal microRNAs during chondrogenic differentiation of human bone mesenchymal stem cells. J Cell Biochem. 2019;120(1):171–181. doi:10.1002/jcb.27289
22. Jin Z, Ren J, Qi S. Exosomal miR-9-5p secreted by bone marrow–derived mesenchymal stem cells alleviates osteoarthritis by inhibiting syndecan-1. Cell Tissue Res. 2020;381(1):99–114. doi:10.1007/s00441-020-03193-x
23. Wang Z, Yan K, Ge G, et al. Exosomes derived from miR-155-5p–overexpressing synovial mesenchymal stem cells prevent osteoarthritis via enhancing proliferation and migration, attenuating apoptosis, and modulating extracellular matrix secretion in chondrocytes. Cell Biol Toxicol. 2021;37(1):85–96. doi:10.1007/s10565-020-09559-9
24. Tao S-C, Yuan T, Zhang Y-L, Yin W-J, Guo S-C, Zhang C-Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics. 2017;7(1):180. doi:10.7150/thno.17133
25. Mao G, Zhang Z, Hu S, et al. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res Ther. 2018;9(1):247. doi:10.1186/s13287-018-1004-0
26. Hu Y, Liu HX, Xu D, Xue X, Xu X. The Anti-Inflammatory Effect of miR-140-3p in BMSCs-Exosomes on Osteoarthritis. Acta Chirurgiae Orthopaedicae Et Traumatologiae Cechoslovaca. 2023;90(4):267–276. doi:10.55095/achot2023/032
27. Qiu M, Liu D, Fu Q. MiR-129-5p shuttled by human synovial mesenchymal stem cell-derived exosomes relieves IL-1β induced osteoarthritis via targeting HMGB1. Life Sci. 2021;269:118987. doi:10.1016/j.lfs.2020.118987
28. Qiu M, Xie Y, Tan G, Wang X, Huang P, Hong L. Synovial mesenchymal stem cell-derived exosomal miR-485-3p relieves cartilage damage in osteoarthritis by targeting the NRP1-mediated PI3K/Akt pathway: exosomal miR-485-3p relieves cartilage damage. Heliyon. 2024;10(2):e24042 doi:10.1016/j.heliyon.2024.e24042..
29. Ragni E, Perucca Orfei C, De Luca P, Colombini A, Viganò M, de Girolamo L. Secreted factors and EV-miRNAs orchestrate the healing capacity of adipose mesenchymal stem cells for the treatment of knee osteoarthritis. Int J Mol Sci. 2020;21(5):1582. doi:10.3390/ijms21051582
30. Rong Y, Zhang J, Jiang D, et al. Hypoxic pretreatment of small extracellular vesicles mediates cartilage repair in osteoarthritis by delivering miR-216a-5p. Acta Biomater. 2021;122:325–342. doi:10.1016/j.actbio.2020.12.034
31. Wan S, Bao D, Li J, et al. Extracellular vesicles from hypoxic pretreated urine-derived stem cells enhance the proliferation and migration of chondrocytes by delivering miR-26a-5p. Cartilage. 2022;13(2):19476035221077401. doi:10.1177/19476035221077401
32. Wang X, Li Z, Cui Y, Cui X, Chen C, Wang Z. Exosomes Isolated From Bone Marrow Mesenchymal Stem Cells Exert a Protective Effect on Osteoarthritis via lncRNA LYRM4-AS1-GRPR-miR-6515-5p. Front Cell Dev Biol. 2021;9:644380. doi:10.3389/fcell.2021.644380
33. He L, Chen Y, Ke Z, et al. Exosomes derived from miRNA-210 overexpressing bone marrow mesenchymal stem cells protect lipopolysaccharide induced chondrocytes injury via the NF-κB pathway. Gene. 2020;751:144764. doi:10.1016/j.gene.2020.144764
34. Li Y, Duan J, Lin W, Liu J. Exosomal miR-93-5p regulated the progression of osteoarthritis by targeting ADAMTS9. Open Med. 2023;18(1):20230668. doi:10.1515/med-2023-0668
35. Xu H, Xu B. BMSC-Derived Exosomes Ameliorate Osteoarthritis by Inhibiting Pyroptosis of Cartilage via Delivering miR-326 Targeting HDAC3 and STAT1//NF-κB p65 to Chondrocytes. Mediators Inflamm. 2021;2021:9972805. doi:10.1155/2021/9972805
36. Li X, Wang Y, Cai Z, Zhou Q, Li L, Fu P. Exosomes from human umbilical cord mesenchymal stem cells inhibit ROS production and cell apoptosis in human articular chondrocytes via the miR-100-5p/NOX4 axis. Cell Biol Int. 2021;45(10):2096–2106. doi:10.1002/cbin.11657
37. Zheng T, Li Y, Zhang X, Xu J, Luo M. Exosomes derived from miR-212-5p overexpressed human synovial mesenchymal stem cells suppress chondrocyte degeneration and inflammation by targeting ELF3. Front Bioeng Biotechnol. 2022;10:816209. doi:10.3389/fbioe.2022.816209
38. Dong J, Li L, Fang X, Zang M. Exosome-encapsulated microRNA-127-3p Released from bone marrow-derived mesenchymal stem cells alleviates osteoarthritis through regulating CDH11-mediated Wnt/β-catenin pathway. J Pain Res. 2021;14:297–310. doi:10.2147/jpr.S291472
39. Wang R, Xu B. TGF-β1-modified MSC-derived exosomal miR-135b attenuates cartilage injury via promoting M2 synovial macrophage polarization by targeting MAPK6. Cell Tissue Res. 2021;384(1):113–127. doi:10.1007/s00441-020-03319-1
40. Huang Y, Zhang X, Zhan J, et al. Bone marrow mesenchymal stem cell–derived exosomal miR-206 promotes osteoblast proliferation and differentiation in osteoarthritis by reducing Elf3. J Cell Mol Med. 2021;25(16):7734–7745. doi:10.1111/jcmm.16654
41. Wang R, Xu B, Xu H. TGF-β1 promoted chondrocyte proliferation by regulating Sp1 through MSC-exosomes derived miR-135b. Cell Cycle. 2018;17(24):2756–2765. doi:10.1080/15384101.2018.1556063
42. Zhang Y, Qi G, Yan Y, et al. Exosomes derived from bone marrow mesenchymal stem cells pretreated with decellularized extracellular matrix enhance the alleviation of osteoarthritis through miR-3473b/phosphatase and tensin homolog axis. J Gene Med. 2023;25(8):e3510. doi:10.1002/jgm.3510
43. Dong S, Xu G, Li X, et al. Exosomes derived from quercetin-treated bone marrow derived mesenchymal stem cells inhibit the progression of osteoarthritis through delivering miR-124-3p to chondrocytes. DNA Cell Biol. 2024;43(2):85–94. doi:10.1089/dna.2023.0341
44. Lin T, Wu N, Wang L, Zhang R, Pan R, Chen Y-F. Inhibition of chondrocyte apoptosis in a rat model of osteoarthritis by exosomes derived from miR-140-5p-overexpressing human dental pulp stem cells. IntJ Mol Med. 2021;47(3):7 doi:10.3892/ijmm.2020.4840.
45. Liu W, Liu A, Li X, et al. Dual-engineered cartilage-targeting extracellular vesicles derived from mesenchymal stem cells enhance osteoarthritis treatment via miR-223/NLRP3/pyroptosis axis: toward a precision therapy. Bioact Mater. 2023;30:169–183. doi:10.1016/j.bioactmat.2023.06.012
46. Liu Y, Zeng Y, Si H-B, Tang L, Xie H-Q, Shen B. Exosomes derived from human urine–derived stem cells overexpressing miR-140-5p alleviate knee osteoarthritis through downregulation of VEGFA in a rat model. Am J Sports Med. 2022;50(4):1088–1105. doi:10.1177/03635465221073991
47. Meng S, Tang C, Deng M, et al. Tropoelastin-pretreated exosomes from adipose-derived stem cells improve the synthesis of cartilage matrix and alleviate osteoarthritis. J Funct Biomat. 2023;14(4):203. doi:10.3390/jfb14040203
48. Sun Y, Zhao J, Wu Q, et al. Chondrogenic primed extracellular vesicles activate miR-455/SOX11/FOXO axis for cartilage regeneration and osteoarthritis treatment. npj Regen Med. 2022;7(1):53. doi:10.1038/s41536-022-00250-7
49. Wu J, Kuang L, Chen C, et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials. 2019;206:87–100. doi:10.1016/j.biomaterials.2019.03.022
50. Ye P, Mi Z, Wei D, Gao P, Ma M, Yang H. miR-3960 from mesenchymal stem cell-derived extracellular vesicles inactivates SDC1/Wnt/β-Catenin axis to relieve chondrocyte injury in osteoarthritis by targeting PHLDA2. Stem Cells Int. 2022;2022:9455152. doi:10.1155/2022/9455152
51. Meng C, Na Y, Han C, et al. Exosomal miR-429 derived from adipose-derived stem cells ameliorated chondral injury in osteoarthritis via autophagy by targeting FEZ2. Int Immunopharmacol. 2023;120:110315. doi:10.1016/j.intimp.2023.110315
52. Zhao S, Xiu G, Wang J, et al. Engineering exosomes derived from subcutaneous fat MSCs specially promote cartilage repair as miR-199a-3p delivery vehicles in Osteoarthritis. J Nanobiotechnol. 2023;21(1):341. doi:10.1186/s12951-023-02086-9
53. Wang K, Li F, Yuan Y, et al. Synovial mesenchymal stem cell-derived EV-packaged miR-31 downregulates histone demethylase KDM2A to prevent knee osteoarthritis. Mol Ther Nucleic Acids. 2020;22:1078–1091. doi:10.1016/j.omtn.2020.09.014
54. Lou C, Jiang H, Lin Z, et al. MiR-146b-5p enriched bioinspired exosomes derived from fucoidan-directed induction mesenchymal stem cells protect chondrocytes in osteoarthritis by targeting TRAF6. J Nanobiotechnol. 2023;21(1):486. doi:10.1186/s12951-023-02264-9
55. Qiu B, Xu X, Yi P, Hao Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J Cell Mol Med. 2020;24(18):10855–10865. doi:10.1111/jcmm.15714
56. Liu Y, Lin L, Zou R, Wen C, Wang Z, Lin F. MSC-derived exosomes promote proliferation and inhibit apoptosis of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in osteoarthritis. Cell Cycle. 2018;17(21–22):2411–2422. doi:10.1080/15384101.2018.1526603
57. Xia Q, Wang Q, Lin F, Wang J. miR-125a-5p-abundant exosomes derived from mesenchymal stem cells suppress chondrocyte degeneration via targeting E2F2 in traumatic osteoarthritis. Bioengineered. 2021;12(2):11225–11238. doi:10.1080/21655979.2021.1995580
58. Tao Y, Zhou J, Wang Z, et al. Human bone mesenchymal stem cells-derived exosomal miRNA-361-5p alleviates osteoarthritis by downregulating DDX20 and inactivating the NF-κB signaling pathway. Bioorg Chem. 2021;113:104978. doi:10.1016/j.bioorg.2021.104978
59. Lu L, Wang J, Fan A, et al. Synovial mesenchymal stem cell-derived extracellular vesicles containing microRN555A-26a-5p ameliorate cartilage damage of osteoarthritis. J Gene Med. 2021;23(11):e3379. doi:10.1002/jgm.3379
60. Kong R, Zhang J, Ji L, Yu Y, Gao J, Zhao D. Synovial mesenchymal stem cell-derived exosomal microRNA-320c facilitates cartilage damage repair by targeting ADAM19-dependent Wnt signalling in osteoarthritis rats. Inflammopharmacology. 2023;31(2):915–926. doi:10.1007/s10787-023-01142-y
61. Meng H-Y, Chen L-Q, Chen L-H. The inhibition by human MSCs-derived miRNA-124a overexpression exosomes in the proliferation and migration of rheumatoid arthritis-related fibroblast-like synoviocyte cell. BMC Musculoskel Disorders. 2020;21(1):150. doi:10.1186/s12891-020-3159-y
62. Su Y, Liu Y, Ma C, Guan C, Ma X, Meng S. Mesenchymal stem cell-originated exosomal lncRNA HAND2-AS1 impairs rheumatoid arthritis fibroblast-like synoviocyte activation through miR-143-3p/TNFAIP3/NF-κB pathway. J Orthop Surg Res. 2021;16(1):116. doi:10.1186/s13018-021-02248-1
63. Chen Z, Wang H, Xia Y, Yan F, Lu Y. Therapeutic potential of mesenchymal cell–derived miRNA-150-5p–expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J Immunol. 2018;201(8):2472–2482. doi:10.4049/jimmunol.1800304
64. Huang Y, Lu D, Ma W, et al. miR-223 in exosomes from bone marrow mesenchymal stem cells ameliorates rheumatoid arthritis via downregulation of NLRP3 expression in macrophages. Mol Immunol. 2022;143:68–76. doi:10.1016/j.molimm.2022.01.002
65. Lin Z, Li W, Wang Y, et al. SMSCs-derived sEV overexpressing miR-433-3p inhibits angiogenesis induced by sEV released from synoviocytes under triggering of ferroptosis. Int Immunopharmacol. 2023;116:109875. doi:10.1016/j.intimp.2023.109875
66. Huang Y, Chen L, Chen D, Fan P, Yu H. Exosomal microRNA-140-3p from human umbilical cord mesenchymal stem cells attenuates joint injury of rats with rheumatoid arthritis by silencing SGK1. Mol Med. 2022;28(1):36. doi:10.1186/s10020-022-00451-2
67. Mi L, Gao J, Li N, et al. Human umbilical cord mesenchymal stem cell-derived exosomes loaded miR-451a targets ATF2 to improve rheumatoid arthritis. Int Immunopharmacol. 2024;127:111365. doi:10.1016/j.intimp.2023.111365
68. Meng Q, Qiu B. Exosomal MicroRNA-320a derived from mesenchymal stem cells regulates rheumatoid arthritis fibroblast-like synoviocyte activation by suppressing CXCL9 expression. Front Physiol. 2020;11:441. doi:10.3389/fphys.2020.00441
69. Ma W, Tang F, Xiao L, et al. miR-205-5p in exosomes divided from chondrogenic mesenchymal stem cells alleviated rheumatoid arthritis via regulating MDM2 in fibroblast-like synoviocytes. J Musculoskelet Neuronal Interact. 2022;22(1):132–141.
70. Li G-Q, Fang Y-X, Liu Y, et al. MicroRNA-21 from bone marrow mesenchymal stem cell-derived extracellular vesicles targets TET1 to suppress KLF4 and alleviate rheumatoid arthritis. Ther Adv Chronic Dis. 2021;12:20406223211007369. doi:10.1177/20406223211007369
71. Zhang Y, Jiao Z, Wang S. Bone marrow mesenchymal stem cells release miR-378a-5p-carried extracellular vesicles to alleviate rheumatoid arthritis. J Innate Immun. 2023;15(1):893–910. doi:10.1159/000534830
72. Wu H, Zhou X, Wang X, et al. miR-34a in extracellular vesicles from bone marrow mesenchymal stem cells reduces rheumatoid arthritis inflammation via the cyclin I/ATM/ATR/p53 axis. J Cell Mol Med. 2021;25(4):1896–1910. doi:10.1111/jcmm.15857
73. Wu L, Wang Z, Hou Z, Zheng L, Gu Z. Exosomal MicroRNA-23-5p Derived from Bone Marrow Mesenchymal Stem Cells Relieves Inflammatory Response in Rheumatoid Arthritis. J Biomater Tissue Eng. 2022;12(5):939–946. doi:10.1166/jbt.2022.2997
74. Vonk LA, van Dooremalen SFJ, Liv N, et al. Mesenchymal stromal/stem cell-derived extracellular vesicles promote human cartilage regeneration in vitro. Theranostics. 2018;8(4):906–920. doi:10.7150/thno.20746
75. Li S, Stöckl S, Lukas C, et al. hBMSC-derived extracellular vesicles attenuate IL-1β-induced catabolic effects on OA-chondrocytes by regulating pro-inflammatory signaling pathways. Front Bioeng Biotechnol. 2020;8:603598. doi:10.3389/fbioe.2020.603598
76. Wu C, He Y, Yao Y, Yang H, Lu F. Exosomes treating osteoarthritis: hope with challenge. Heliyon. 2023;9(1):e13152. doi:10.1016/j.heliyon.2023.e13152
77. Cosenza S, Ruiz M, Toupet K, Jorgensen C, Noël D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep. 2017;7(1):16214. doi:10.1038/s41598-017-15376-8
78. Ruiz M, Toupet K, Maumus M, Rozier P, Jorgensen C, Noël D. TGFBI secreted by mesenchymal stromal cells ameliorates osteoarthritis and is detected in extracellular vesicles. Biomaterials. 2020;226:119544. doi:10.1016/j.biomaterials.2019.119544
79. Li S, Liu J, Liu S, Jiao W, Wang X. Mesenchymal stem cell-derived extracellular vesicles prevent the development of osteoarthritis via the circHIPK3/miR-124-3p/MYH9 axis. J Nanobiotechnol. 2021;19(1):194. doi:10.1186/s12951-021-00940-2
80. Mortati L, de Girolamo L, Perucca Orfei C, et al. In vitro study of extracellular vesicles migration in cartilage-derived osteoarthritis samples using real-time quantitative multimodal nonlinear optics imaging. Pharmaceutics. 2020;12(8):734. doi:10.3390/pharmaceutics12080734
81. Woo CH, Kim HK, Jung GY, et al. Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J Extracell Vesicles. 2020;9(1):1735249. doi:10.1080/20013078.2020.1735249
82. Cavallo C, Merli G, Borzì RM, et al. Small extracellular vesicles from adipose derived stromal cells significantly attenuate in vitro the NF-κB dependent inflammatory/catabolic environment of osteoarthritis. Sci Rep. 2021;11(1):1053. doi:10.1038/s41598-020-80032-7
83. Huang S, Song X, Li T, et al. Pellet coculture of osteoarthritic chondrocytes and infrapatellar fat pad-derived mesenchymal stem cells with chitosan/hyaluronic acid nanoparticles promotes chondrogenic differentiation. Stem Cell Res Ther. 2017;8(1):264. doi:10.1186/s13287-017-0719-7
84. Zhao C, Chen J-Y, Peng W-M, Yuan B, Bi Q, Xu Y-J. Exosomes from adipose‑derived stem cells promote chondrogenesis and suppress inflammation by upregulating miR‑145 and miR‑221. Mol Med Rep. 2020;21(4):1881–1889. doi:10.3892/mmr.2020.10982
85. Liu Z, Zhuang Y, Fang L, Yuan C, Wang X, Lin K. Breakthrough of extracellular vesicles in pathogenesis, diagnosis and treatment of osteoarthritis. Bioact Mater. 2023;22:423–452. doi:10.1016/j.bioactmat.2022.10.012
86. Cosenza S, Toupet K, Maumus M, et al. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics. 2018;8(5):1399–1410. doi:10.7150/thno.21072
87. Yin Z, Qin C, Pan S, et al. Injectable hyperbranched PEG crosslinked hyaluronan hydrogel microparticles containing mir-99a-3p modified subcutaneous ADSCs-derived exosomes was beneficial for long-term treatment of osteoarthritis. Mater Today Bio. 2023;23:100813. doi:10.1016/j.mtbio.2023.100813
88. Sophia Fox AJ, Bedi A, Rodeo SA. The basic science of articular cartilage: structure, composition, and function. Sports Health. 2009;1(6):461–468. doi:10.1177/1941738109350438
89. Geiger BC, Wang S, Padera RF Jr, Grodzinsky AJ, Hammond PT. Cartilage-penetrating nanocarriers improve delivery and efficacy of growth factor treatment of osteoarthritis. Sci Transl Med. 2018;10(469):eaat8800. doi:10.1126/scitranslmed.aat8800
90. Bajpayee AG, Scheu M, Grodzinsky AJ, Porter RM. A rabbit model demonstrates the influence of cartilage thickness on intra-articular drug delivery and retention within cartilage. J Orthop Res. 2015;33(5):660–667. doi:10.1002/jor.22841
91. Venkatesan J, Liu W, Madry H, Cucchiarini M. Alginate hydrogel-guided rAAV-mediated FGF-2 and TGF-β delivery and overexpression stimulates the biological activities of human meniscal fibrochondrocytes for meniscus repair. Eur Cells Mat. 2024;47:1–14. doi:10.22203/eCM.v047a01
92. Eirin A, Zhu X-Y, Puranik AS, et al. Integrated transcriptomic and proteomic analysis of the molecular cargo of extracellular vesicles derived from porcine adipose tissue-derived mesenchymal stem cells. PLoS One. 2017;12(3):e0174303. doi:10.1371/journal.pone.0174303
93. Zununi Vahed S, Hejazian SM, Bakari WN, et al. Milking mesenchymal stem cells: updated protocols for cell lysate, secretome, and exosome extraction, and comparative analysis of their therapeutic potential. Methods. 2025;238:40–60. doi:10.1016/j.ymeth.2025.03.004
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.