Back to Journals » Journal of Pain Research » Volume 18
Mechanotransduction Pathways in Massage Therapy: CB1/Piezo2 Receptor Crosstalk Modulates Myocellular Anti-Inflammatory Responses
Authors Liu Q, He W, Li M ![]() , Li Z, Wu C
, Li Z, Wu C
Received 18 June 2025
Accepted for publication 21 October 2025
Published 27 November 2025 Volume 2025:18 Pages 6359—6367
DOI https://doi.org/10.2147/JPR.S548026
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Houman Danesh
Qian Liu,1 Wen He,2 Man Li,3 Zeyun Li,4 Cai Wu4
1School of Sports and Health, Hubei University of Chinese Medicine, Wuhan, People’s Republic of China; 2Department of Rehabilitation Medicine, Wuhan Red Cross Hospital, Wuhan, People’s Republic of China; 3Department of Physiology, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China; 4School of Traditional Chinese Medicine, Hubei University of Chinese Medicine, Wuhan, People’s Republic of China
Correspondence: Cai Wu, School of Traditional Chinese Medicine, Hubei University of Chinese Medicine, No. 188 Tan Hualin Road, Wuchang District, Wuhan, 430061, People’s Republic of China, Tel +8618062600291, Email [email protected]
Background: Massage therapy is widely used to alleviate pain, however, its molecular mechanisms, particularly those integrating Traditional Chinese Medicine (TCM) with mechanobiology, remain inadequately elucidated. This study investigates the novel hypothesis that coordinated signaling between cannabinoid receptor-1 (CB1) and the Epac1-Piezo2 axis underlies massage-induced analgesia in inflammatory pain.
Methods: Female C57BL/6 mice (8– 10 weeks) were randomly allocated into three groups (n=6/group): Control group: Received a 25-μL intramuscular microinjection of saline into the right tibialis anterior muscle; CFA group: Inflammatory hyperalgesia was established via 25-μL intramuscular CFA (complete Freund’s adjuvant) microinjection. Model validation was performed 48 hours post-CFA administration by assessing ipsilateral hindlimb edema, erythema, exudation, and mechanical hyperalgesia using von Frey filament testing. Massage group: CFA mice received daily ipsilateral ST36/GB34 acupressure for 14 days starting 24h post-modeling. Thermal and mechanical hyperalgesia were assessed via hot plate and von Frey tests. CB1 (Western blot), Epac1 (IHC), and Piezo2 (IF) levels in L4-6 DRGs were analyzed.
Results: Acupoint massage (targeting ST36 and GB34) significantly elevated mechanical and thermal pain thresholds in CFA-induced inflammatory pain mice. Western blot analysis revealed upregulation of CB1 protein expression in L4– 6 DRGs. Notably, massage downregulated Epac1 and Piezo2 protein levels, suggesting that the analgesic mechanism involves modulation of the CB1, Epac1, and Piezo2 signaling pathways.
Conclusion: These findings demonstrate that massage therapy attenuates inflammatory pain, potentially through activation of cannabinoid type 1 (CB1) receptors and modulation of the Epac1/Piezo2 signaling axis. This study provides mechanistic insights into the analgesic properties of massage treatment.
Keywords: massage, analgesic, cannabinoid receptor-1, Epac1, Piezo2, inflammation
Introduction
Massage therapy is an established intervention in sports medicine and clinical rehabilitation, effectively aiding pain management and recovery.1,2 This manual technique employs controlled mechanical stimuli to specific anatomical regions, achieving therapeutic outcomes through three principal mechanisms: myofascial tension reduction, localized hemodynamic enhancement, and neural modulation of nociceptive pathways.3–6 Inflammatory pain involves neurochemical alterations leading to mechanical allodynia.7 Emerging research has identified Piezo2, a mechanosensitive ion channel integral to somatosensory transduction, as a critical mediator in both massage analgesia and mechanical hypersensitivity.8–12 Its involvement in pathological pain makes it a promising therapeutic target.12–15 Current mechanistic investigations delineate two synergistic pathways underlying manual therapy effects: The Epac1-Piezo2 axis mediates mechanical allodynia through injury-induced Epac1 mRNA elevation in dorsal root ganglia (DRGs).16–18 Central endocannabinoid signaling modulates descending pain inhibition via CB1 receptor activation.19–21 This study elucidates the novel interaction between cannabinoid signaling and mechanotransduction pathways, demonstrating that massage-induced analgesia requires CB1-mediated regulation of the Epac1-Piezo2 cascade. Our findings provide a molecular framework for understanding manual therapy mechanisms while identifying promising targets for precision pain management.
Materials and Methods
Animals
Female C57BL/6J mice (6–8 weeks old, specific pathogen-free [SPF]) were obtained from Collection Medicine Kang Biotechnology Co. Ltd. (SPF, China) and randomly assigned to experimental subgroups. All animals were maintained under controlled conditions (23°C, 12 h/12 h light/dark cycle) with ad libitum access to standard rodent chow and water. The experimental protocol was approved by the Institutional Animal Care and Use Committee of Hubei University of Chinese Medicine (Approval No. HUCMS00304833) and followed the Regulations on Laboratory Animal Management (revised 2018).
Piezo2 Ion Channels
Piezo2 is a mechanosensitive ion channel critical for transducing mechanical stimuli into electrochemical signals in sensory neurons.8–12 Its role in touch and pain sensation makes it a key target in mechanotherapy studies.
Group Design and Timing
The three experimental groups were designed to assess both immediate and sustained effects of massage: Control group: baseline saline injection; CFA group: inflammatory pain model Massage group: CFA + daily massage for 14 days. The 14-day intervention was chosen to evaluate cumulative neuroadaptive changes, while acute responses were assessed daily.
Complete Freund’s Adjuvant (CFA) Injection
Inflammatory injury was induced via intramuscular injection of 25 μL Complete Freund’s Adjuvant (CFA; Sigma, St. Louis, MO) into the right tibialis anterior muscle of anesthetized C57BL/6J mice (sodium pentobarbital, 50 mg/kg i.p). The CFA suspension (25 μL), containing inactivated dried mycobacteria, was slowly administered using a microsyringe to ensure precise delivery and minimize leakage. Contralateral tibialis anterior muscles received an equivalent volume (25 μL) of sterile saline as controls. This procedure was performed on 18 animals (180–220 g). Model validation included: (1) gross morphological assessment of inflammation (swelling, congestion, exudation) in ipsilateral hindlimbs 48 h post-injection, and (2) quantitative mechanical hyperalgesia testing with von Frey filaments following CFA challenge.
Massage Treatment
Mice were randomly divided into 3 groups: the control group, the CFA group, and the massage group, with 6 mice in each group. The solvent control group received 25 µL of physiological saline via intramuscular injection into the anterior tibialis muscle on the right side of the mouse. The CFA model group established a myositis pain model by administering a small amount of 25 µL complete Freund’s adjuvant (CFA) intramuscularly into the anterior tibialis muscle on the right side of the mouse. Two days after CFA injection, the mice were observed for signs such as swelling, congestion, and exudation in the right hind limb, and whether mechanical hyperalgesia occurred, to confirm the success of the model. In the massage group, starting from the second day after successful CFA modeling, the affected area’s Zusanli (ST36) and Yanglingquan (GB34) acupoints were massaged once daily for 5 minutes each time, for a total of 14 sessions.
Von Frey Filaments Test
After the model meets the standards, mechanical pain sensitivity is assessed using von Frey fibers. The experimental animals are placed in a transparent resin observation box with a metal mesh bottom for about 20 minutes to acclimate to the environment until their limb movements stabilize. According to the modified “up-down” method,7 eight different fiber sizes (0.4, 0.6, 1.0, 2.0, 4.0, 6.0, 8.0, and 15.0g) are selected, with an initial force intensity set at 2.0g. Precise mechanical stimulation is applied to the plantar region of the hind limbs of mice. During the application of force, ensure that the fibers form standard “C” or “S” bends and maintain pressure for 6–8 seconds. Behavioral responses are recorded according to international standards:6 no significant foot retraction or transient mild reflex is marked as “O”; persistent strong foot retraction or licking behavior is labeled as “X”.
Adjust the intensity of stimulation dynamically based on the type of response: increase the force gradient after a negative response, and decrease the stimulation level after a positive response. The interval between each application of force should be no less than 30 seconds. The mechanical pain threshold is ultimately calculated using the standard formula (50% contraction threshold (g) = 10 [Xf + kδ] / 10,000).22 To avoid pain sensitization caused by repeated stimulation, the experiment strictly adheres to a testing interval of over 10 minutes.23
Hot Plate Test
The thermal plate instrument must be pre-opened and calibrated to (52.0±0.2) °C. The experiment can only proceed after the device has reached temperature equilibrium. At this point, the acrylic protective cover should be removed, and the experimental mouse must be securely held by the operator to ensure that its hind feet remain in contact with the heating surface. The timing device should start strictly synchronized with the contact of the animal with the thermal plate. When a clear foot-holding-off response is observed in the animal, the timer should immediately pause to record the duration. The determination of the baseline pain threshold requires two independent measurements, with a minimum interval of 15 minutes between the two tests. The final pain threshold before administration is taken as the average of the two valid measurements.
Western Blotting Assay
Mice were anesthetized via intraperitoneal sodium pentobarbital injection (50 mg/kg, 1% in saline) and euthanized by cervical dislocation. The right lumbar DRGs (L4-L6) were immediately dissected post-euthanasia and flash-frozen in liquid nitrogen. Tissue lysates were prepared using RIPA buffer supplemented with 1% (v/v) phenylmethanesulfonyl fluoride (PMSF) as previously described. Total protein concentration was quantified in triplicate using a BCA assay kit per manufacturer’s protocol, with final values representing triplicate averages. Equal protein aliquots (20 μg) underwent electrophoretic separation on 12% SDS-PAGE gels and were subsequently electroblotted onto PVDF membranes (Millipore, USA). After 1 h blocking with 5% non-fat milk or 5% BSA, membranes were sectioned for target-specific incubation with primary antibodies against CNR1 (1:1000) and GAPDH (1:1000) overnight at 4°C, the latter serving as loading control. Following three 5-min TBS-T washes, membranes were incubated with secondary antibodies for 2 h at room temperature, then subjected to six 5-min TBS-T washes.
Immunohistochemistry
Right lumbar dorsal root ganglia (DRG; L4-L6) were immediately dissected from mice and flash-frozen in liquid nitrogen. Following fixation in 4% paraformaldehyde (PFA) for ≥24 h, samples were paraffin-embedded and sectioned at 5 μm. Sections underwent dewaxing followed by 10-min incubation with 3% H2O2 to quench endogenous peroxidase activity. Antigen retrieval was performed via high-pressure heating, then non-specific binding was blocked with 5% BSA (30 min, room temperature). Sections were probed with primary antibody against EPAC1 (90 min, 37°C), then sequentially incubated with: (1) biotinylated goat anti-rabbit IgG secondary antibody, (2) streptavidin-biotin complex (SABC), and (3) diaminobenzidine (DAB) chromogenic substrate. After hematoxylin counterstaining of nuclei, sections were dehydrated, mounted, and imaged at 40× magnification.
Immunofluorescence
Right lumbar dorsal root ganglia (DRG; L4-L6) were dissected from mice and cryopreserved in liquid nitrogen. Following ≥24 h fixation in 4% paraformaldehyde (PFA), tissues were paraffin-embedded and sectioned at 5 μm. After deparaffinization, high-pressure epitope retrieval was performed. Sections were blocked with 5% bovine serum albumin (BSA; 30 min, room temperature), then incubated with anti-Piezo2 primary antibody in a humidified chamber (15 h, 4°C). Sequential incubations included: (1) biotin-conjugated goat anti-rabbit IgG secondary antibody, and (2) 4’,6-diamidino-2-phenylindole (DAPI) for nuclear counterstaining under light-protected conditions. Finally, slides were mounted with anti-fade medium and imaged at 40× magnification.
Statistical Analysis
Data are expressed as mean ± standard error of the mean (SEM). Statistical analyses were performed using GraphPad Prism 9.5 and SPSS Statistics 21.0 (IBM Corp., Armonk, NY). Intergroup differences were assessed by one-way ANOVA with Bonferroni post-hoc tests for multiple comparisons. For longitudinal measurements, two-way ANOVA (factors: time and group) followed by Bonferroni correction was applied. Statistical significance was defined as P < 0.05 (two-tailed). A schematic overview of the experimental timeline and procedures is provided in Supplementary Figure 1.
Results
Massage Increased the Mechanical Threshold of Inflammatory Muscle Pain
Three days before the experiment, the baseline pain threshold of mice was measured. Three days later, a model was established to prepare a CFA-induced inflammatory pain mouse model. On the second day after successful modeling, massage was performed on the Zusanli (ST36) and Yanglingquan (GB34) acupoints of the mice once daily for 5 minutes each time, for a total of 14 consecutive days. After massage, the mechanical hyperalgesia of each group of mice was tested daily using a touch needle (Von Frey wire) according to the “up and down” method reported by Chaplan et al. The results showed that two days after CFA injection, swelling appeared in the tibialis anterior muscle, and some mice exhibited signs of congestion and exudation. All mice injected with CFA showed a decrease in pain threshold. After massage at the Zusanli and Yanglingquan acupoints, the mechanical pain threshold of the mice gradually increased. (See Figure 1).
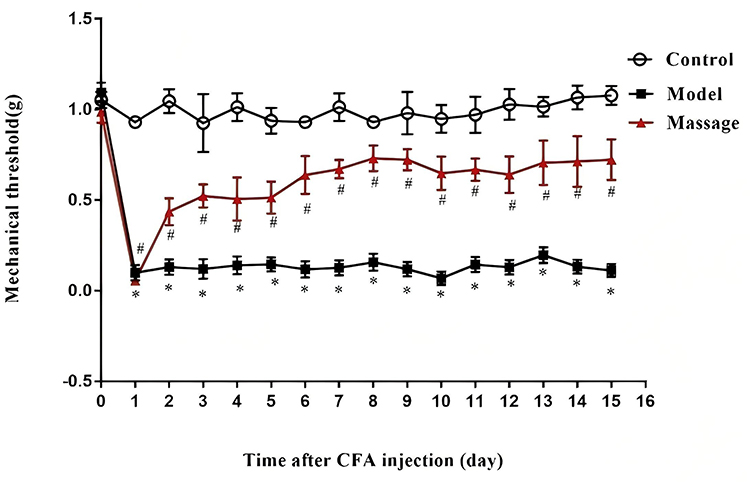 |
Figure 1 Effects of massage on mechanical pain threshold in mice. Pain behavior was measured after massage treatment on the second day after CFA modeling. Compared with the control group, *P<0.05; compared with the CFA group, #P<0.05. |
Massage Increased the Thermal Latency of Inflammatory Muscle Pain
Three days before the experiment, the baseline pain threshold of mice was measured. Three days later, a model was established to prepare a CFA-induced inflammatory pain mouse model. On the second day after successful modeling, massage was performed on the Zusanli and Yanglingquan acupoints of the mice (the treatment protocol was the same as above). The hot plate method was used to measure the thermal pain threshold. It was found that (as shown in Figure 2) all mice injected with CFA exhibited a decrease in their thermal pain threshold. After massage at the Zusanli (ST36) and Yanglingquan (GB34) acupoints, the thermal pain threshold of the mice gradually increased.
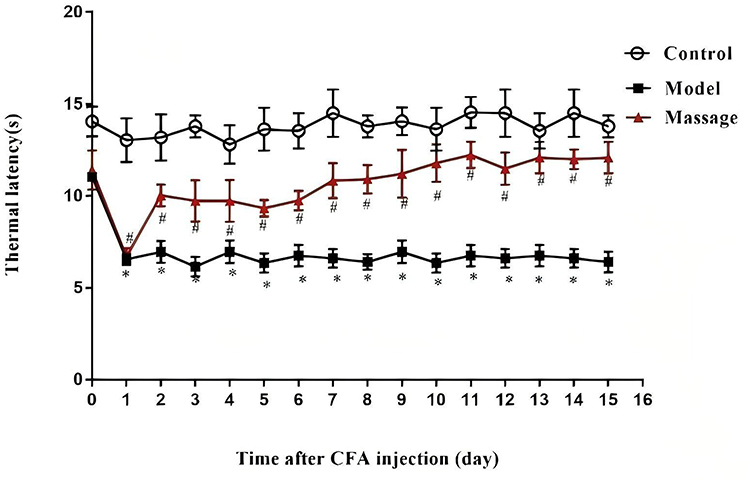 |
Figure 2 Effects of massage on thermal pain threshold in mice. Pain behavior was measured on the second day after CFA modeling with massage treatment. Compared with the control group, *P<0.05; compared with the CFA group, #P<0.05. |
Massage Could Upregulate the Protein Expression of CB1 in L4-6 DRGs
This study proposes that massage therapy based on the “massage point of pain” theory alleviates inflammatory pain by activating the endogenous cannabinoid system, specifically manifested as increased CB1 receptor expression in dorsal root ganglion (DRG) neurons. The CB1 receptor is widely distributed in both central and peripheral nervous systems and plays a core role in pain modulation: its activation can inhibit the release of presynaptic neurotransmitters (such as glutamate) and induce hyperpolarization of postsynaptic neurons, thereby reducing pain signal transmission. In the experiment, Western blot results showed that CB1 protein expression was significantly higher in the massage group compared to the CFA model group (P<0.01), indicating that mechanical stimulation of pain-sensitive acupoints can promote the release of endogenous cannabinoids. This mechanism aligns with the “gate control theory” of pain, which suggests that non-harmful stimuli (such as massage) can inhibit harmful signal transmission. Additionally, CB1 activation reduces neurogenic inflammation by inhibiting mast cell degranulation and pro-inflammatory factor release, echoing the traditional Chinese medical concept of unblocking meridians to relieve pain. This dual effect of neural regulation and anti-inflammatory action provides a biochemical basis for the analgesic effects of “massage point of pain” massage.
Endocannabinoid CB1 receptor activation in GABAergic neurons and inhibitory glutamatergic neurons of the vLPAG exerts analgesic effects, indicating that CB1 receptors on GABAergic neurons in the vLPAG are important targets for massage-induced analgesia. In this study, a CFA-induced inflammatory pain model in mice was used. On the second day after successful modeling, massage was performed on the Zusanli and Yanglingquan acupoints of the mice once daily for 5 minutes each time, for a total of 14 days. Samples were taken from L4-6DRG to detect CB1 levels. Western blot results showed that compared with the Control group and the CFA model group, the CB1 protein expression level in DRG neurons was significantly increased in the massage group (Figure 3, P<0.05).
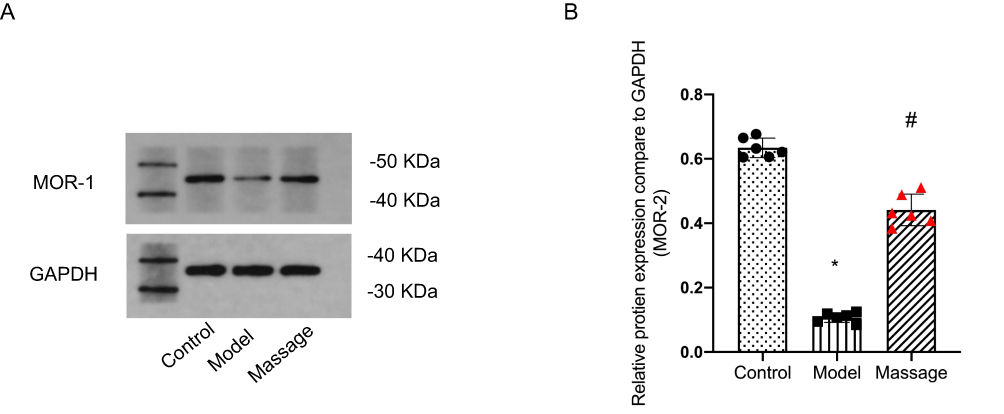 |
Figure 3 (A) Representative Western blots of CB1 and GAPDH in L4-L6 DRGs. The CB1 band intensity was increased in the Massage group compared to the Model group; (B) Effects of massage on CB1 receptor protein expression in DRG neurons of the dorsal root of mice with CFA-induced myalgia. Compared with Control group: *P<0.05; Compared with Model group: #P<0.05. |
Massage Could Downregulate the Protein Expression of Epac1 and Piezo2 in L4-6 DRGs
This study systematically investigated the therapeutic mechanisms of acupoint massage on inflammatory pain using a Complete Freund’s Adjuvant (CFA)-induced murine myositis model, which effectively recapitulates clinical inflammatory pain phenotypes. Within 24 hours post-CFA injection, mice developed sustained mechanical and thermal hyperalgesia, as validated by von Frey filament and hot plate tests (both P<0.05). Intervention with daily acupoint massage at Zusanli (ST36) and Yanglingquan (GB34)—selected based on meridian theory for their roles in regulating visceral function and alleviating musculoskeletal pain—was administered for 5 minutes per session over 14 consecutive days. Behavioral analyses demonstrated that massage significantly reversed CFA-induced reductions in pain thresholds (P<0.05), exhibiting rapid-onset and sustained analgesic effects. At the molecular level, downregulation of Epac1 and Piezo2 proteins in dorsal root ganglion (DRG) neurons was identified as a critical pathway mediating mechanical nociception. Epac1 potentiates TRPV4 channel activity to amplify pain signaling, while Piezo2 directly transduces mechanical stimuli. Acupoint massage concurrently suppressed both targets, providing mechanistic insights into its efficacy in restoring mechanical pain thresholds.
Interventions commenced on day 2 post-CFA modeling, with L4–6 DRG tissues systematically harvested after the 14-day regimen. Immunohistochemical quantification (cell diameter classification: 50 μm) revealed a significant reduction in Epac1-positive neurons in the massage group compared to CFA controls (P<0.05, Figure 4A). Parallel immunofluorescence analyses confirmed markedly decreased Piezo2 expression in DRG neurons (P<0.05, Figure 4B). This dual modulation of the Epac1-Piezo2 axis, validated through integrated behavioral and molecular approaches, substantiates the traditional “treating pain at acupoints” theory within a contemporary neurobiological framework. The findings highlight acupoint massage as a non-pharmacological strategy for inflammatory pain management, bridging classical meridian concepts with modern mechanotransduction pathways. Western blot analysis further corroborated the downregulation of both Epac1 and Piezo2 protein levels in DRG tissues (P<0.05, Figure 4C and D).
 |
Figure 4 (A) Representative IHC images of Epac1 (scale bar: 50 µm). Epac1-positive staining was reduced in the Massage group compared to the Model group; (B) Statistical analysis of Epac1-positive cell counts. *P < 0.05 vs Control group; #P < 0.05 vs Model group; (C) Representative IF images of Piezo2 (red) and DAPI (blue; scale bar: 50 µm). The Piezo2 fluorescence signal was attenuated in the Massage group compared to the Model group; (D) Effects of massage on the expression of Epac1 and Piezo2 proteins in dorsal root DRG neurons of CFA-induced muscle inflammatory pain mice. Compared with Control group: *P<0.05; Compared with Model group: #P<0.05. |
Discussion
This study demonstrates a mechanobiological mechanism by which acupoint massage alleviates inflammatory pain through coordinated regulation of the CB1 receptor and Epac1-Piezo2 pathways. Targeted mechanical stimulation at Zusanli (ST36) and Yanglingquan (GB34) acupoints significantly reduced mechanical and thermal hyperalgesia in CFA-treated mice. Behavioral improvements were accompanied by increased CB1 expression and decreased Epac1/Piezo2 levels in L4–6 dorsal root ganglia (DRGs). These findings align with the traditional meridian theory of “unblocking collaterals to relieve pain” and provide molecular evidence linking TCM practices to modern pain modulation pathways.
Mechanical stimulation at these acupoints activated CB1 receptors, which are known to inhibit nociceptive signaling through presynaptic neurotransmitter suppression and postsynaptic neuronal hyperpolarization.24 Elevated CB1 levels in DRGs after massage suggest enhanced endocannabinoid activity, potentially counteracting neurogenic inflammation by limiting mast cell activation and cytokine release.21 This observation is consistent with prior reports of CB1-mediated analgesia in spinal and supraspinal circuits. Simultaneously, reduced Epac1 and Piezo2 expression highlights the interaction between inflammatory signaling and mechanotransduction. Epac1 exacerbates mechanical hypersensitivity via TRPV4 channel activation,25 whereas Piezo2 directly mediates noxious mechanical stimuli.14,26 Acupoint massage disrupts both pathways, effectively reducing nociceptive signal amplification and mechanosensory transmission.
The progressive normalization of pain thresholds over 14 days of intervention indicates cumulative neuroadaptive changes. These results corroborate earlier work establishing Piezo2’s role in mechanical allodynia and Epac1’s contribution to neuropathic pain. Selective Piezo2 downregulation in DRGs supports its therapeutic relevance, as genetic deletion of Piezo2 reduces mechanical hypersensitivity without affecting non-noxious tactile perception.
Several limitations should be noted. First, the study focused on peripheral DRG mechanisms; future work should investigate central pathways, particularly spinal and supraspinal CB1-mediated circuits. Second, temporal changes in protein expression during the intervention period require further characterization. Third, clinical validation in patients with inflammatory pain is needed to confirm translational relevance.
In summary, this work clarifies the therapeutic potential of acupoint massage in inflammatory pain management. By integrating meridian theory with mechanobiological principles, it reveals molecular interactions between endocannabinoid signaling and mechanotransduction. These findings validate TCM practices scientifically and identify CB1 and Piezo2 as targets for mechanotherapy. Future research should investigate synergistic strategies combining acupoint stimulation with pharmacological agents targeting these pathways to enhance analgesic efficacy.
Conclusions
This study establishes that targeted acupoint massage alleviates inflammatory pain through synergistic modulation of cannabinoid and mechanotransduction pathways. Mechanistically, mechanical stimulation at ST36/GB34 acupoints upregulated CB1 receptor expression while downregulating Epac1 and Piezo2 proteins in lumbar dorsal root ganglia, thereby attenuating nociceptive signal transmission. These findings bridge traditional meridian theory with contemporary neurobiology by demonstrating that: (1) CB1 activation suppresses neurogenic inflammation via presynaptic inhibition,(2) Dual inhibition of the Epac1-Piezo2 axis disrupts mechanical hypersensitivity. Our integrated approach provides a molecular framework for massage-induced analgesia, positioning CB1/Piezo2 crosstalk as a therapeutic target for inflammatory pain management.
Data Sharing Statement
All data for this study are publicly available.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 82505788).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Dupuy O, Douzi W, Theurot D, et al. An evidence-based approach for choosing post-exercise recovery techniques to reduce markers of muscle damage, soreness, fatigue, and inflammation: a systematic review with meta-analysis. Front Physiol. 2018;9:403. doi:10.3389/fphys.2018.00403
2. Young C, Argáez C. Manual Therapy for Chronic Non-Cancer Back and Neck Pain: A Review of Clinical Effectiveness. Ottawa (ON): Canadian Agency for Drugs and Technologies in Health; 2020.
3. Deng G. Integrative medicine therapies for pain management in cancer patients. Cancer J. 2019;25(5):343–348. doi:10.1097/PPO.0000000000000399
4. Guo J, Li L, Gong Y, et al. Massage alleviates delayed onset muscle soreness after strenuous exercise: a systematic review and meta-analysis. Front Physiol. 2017;8:747. doi:10.3389/fphys.2017.00747
5. Ernst E. The safety of massage therapy. Rheumatology. 2003;42(9):1101–1106. doi:10.1093/rheumatology/keg306
6. Loew LM, Brosseau L, Tugwell P, et al. Deep transverse friction massage for treating lateral elbow or lateral knee tendinitis. Cochrane Database Syst Rev. 2014;2014(11):CD003528. doi:10.1002/14651858.CD003528.pub2
7. Wang L, Zhou H, Zhang M, et al. Structure and mechanogating of the mammalian tactile channel PIEZO2. Nature. 2019;573(7773):225–229. doi:10.1038/s41586-019-1505-8
8. Feng J, Luo J, Yang P, et al. Piezo2 channel-Merkel cell signaling modulates the conversion of touch to itch. Science. 2018;360(6388):530–533. doi:10.1126/science.aar5703
9. Coste B, Mathur J, Schmidt M, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330(6000):55–60. doi:10.1126/science.1193270
10. Woo SH, Ranade S, Weyer AD, et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature. 2014;509(7502):622–626. doi:10.1038/nature13251
11. Ranade SS, Woo SH, Dubin AE, et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature. 2014;516(7529):121–125. doi:10.1038/nature13980
12. Szczot M, Liljencrantz J, Ghitani N, et al. PIEZO2 mediates injury-induced tactile pain in mice and humans. Sci Transl Med. 2018;10(462):eaat9892. doi:10.1126/scitranslmed.aat9892
13. Chesler AT, Szczot M, Bharucha-Goebel D, et al. The role of PIEZO2 in human mechanosensation. New Engl J Med. 2016;375(14):1355–1364. doi:10.1056/NEJMoa1602812
14. Murthy ES, Loud CM, Daou I, et al. The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice. Sci Trans Med. 2018;10(462):eaat9897.
15. Liu M, Li Y, Zhong J, et al. The effect of IL-6/Piezo2 on the trigeminal neuropathic pain. Aging. 2021;13(10):13615–13625. doi:10.18632/aging.202887
16. Grandoch M, Roscioni SS, Schmidt M. The role of Epac proteins, novel cAMP mediators, in the regulation of immune, lung and neuronal function. Br J Pharmacol. 2010;159(2):265–284. doi:10.1111/j.1476-5381.2009.00458.x
17. Eijkelkamp N, Linley JE, Torres JM, et al. A role for Piezo2 in EPAC1-dependent mechanical allodynia. Nat Commun. 2013;4(1):1682. doi:10.1038/ncomms2673
18. Ni K, Zhang W, Ni Y, et al. Dorsal root ganglia NR2B-mediated Epac1-Piezo2 signaling pathway contributes to mechanical allodynia of bone cancer pain. Oncol Lett. 2021;21(4):338. doi:10.3892/ol.2021.12599
19. Lindgren L, Gouveia-Figueira S, Nording ML, et al. Endocannabinoids and related lipids in blood plasma following touch massage: a randomised, crossover study. BMC Res Notes. 2015;8(1):504. doi:10.1186/s13104-015-1450-z
20. Ho YC, Lee HJ, Tung LW, et al. Activation of orexin 1 receptors in the periaqueductal gray of male rats leads to antinociception via retrograde endocannabinoid (2-arachidonoylglycerol)-induced disinhibition. J Neurosci. 2011;31(41):14600–14610. doi:10.1523/JNEUROSCI.2671-11.2011
21. Meng ID, Manning BH, Martin WJ, et al. An analgesia circuit activated by cannabinoids. Nature. 1998;395(6700):381–383. doi:10.1038/26481
22. Chaplan SR, Bach FW, Pogrel JW, et al. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53(1):55–63. doi:10.1016/0165-0270(94)90144-9
23. Bonin RP, Bories C, De Koninck Y. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol Pain. 2014;10:26. doi:10.1186/1744-8069-10-26
24. Yang F, Xu Q, Shu B, et al. Activation of cannabinoid CB1 receptor contributes to suppression of spinal nociceptive transmission and inhibition of mechanical hypersensitivity by Aβ-fiber stimulation. Pain. 2016;157(11):2582–2593. doi:10.1097/j.pain.0000000000000680
25. Hu X, Du L, Liu S, et al. A TRPV4-dependent neuroimmune axis in the spinal cord promotes neuropathic pain. J Clin Invest. 2023;133(5):e161507. doi:10.1172/JCI161507
26. Ferrari FL, Bogen O, Green P, et al. Contribution of Piezo2 to endothelium-dependent pain. Molecular Pain. 2015;11(1):189–195. doi:10.1186/s12990-015-0068-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.