Back to Journals » Infection and Drug Resistance » Volume 17
Mechanisms of Antibiotic Resistance and Developments in Therapeutic Strategies to Combat Klebsiella pneumoniae Infection
Authors Li Y ![]() , Kumar S
, Kumar S ![]() , Zhang L
, Zhang L
Received 11 December 2023
Accepted for publication 12 March 2024
Published 19 March 2024 Volume 2024:17 Pages 1107—1119
DOI https://doi.org/10.2147/IDR.S453025
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Yanping Li,1,2 Suresh Kumar,3 Lihu Zhang1
1Pharmacy Department, Jiangsu Vocational College of Medicine, Yancheng, Jiangsu, People’s Republic of China; 2Post Graduate Centre, Management and Science University, Shah Alam, Malaysia; 3Department of Diagnostic and Allied Health Science, Faculty of Health and Life Sciences, Management and Science University, Shah Alam, Malaysia
Correspondence: Lihu Zhang; Suresh Kumar, Email [email protected]; [email protected]
Abstract: Infections with drug-resistant bacteria have become one of the greatest public health challenges, and K. pneumoniae is among the top six drug-resistant bacteria. K. pneumoniae often causes nosocomial infections, leading to illnesses such as pneumonia, liver abscesses, soft tissue infections, urinary tract infections, bacteremia, and in some cases death. As the pathogen continues to evolve and its multidrug resistance increases, K. pneumoniae poses a direct threat to humans. Drug resistance in K. pneumoniae may occur due to the formation of biofilms, efflux pumps, and the production of β-lactamases. In many cases, resistance is further enhanced by enzymatic modification and loss of porins. Drug resistance to K. pneumoniae has led to a decline in the effectiveness of conventional therapies against this pathogen. Therefore, there is an urgent need to accelerate the development of new antibiotics and explore new therapeutic approaches such as antimicrobial peptides, phages, traditional Chinese medicine, immunotherapy, Antimicrobial nanoparticle technology, antisense oligonucleotides and gene editing technologies. In this review, we discuss the mechanisms of drug resistance in K. pneumoniae and compare several new potential therapeutic strategies to overcome drug resistance in the treatment of K. pneumoniae infections.
Keywords: Klebsiella pneumoniae, antibiotic resistance mechanisms, therapeutic strategies
Introduction
Klebsiella pneumoniae (K. pneumoniae) is a gram-negative bacterium found primarily in the respiratory tract and intestines of humans, often causes nosocomial infections, leading to illnesses such as pneumonia, liver abscesses, soft tissue infections, urinary tract infections, bacteremia, and in some cases death.1–3 Due to the irrational use of some antibiotics and the evolution of bacteria, drug-resistant bacterial infections have become one of the greatest public health challenges. In 2019, more than 1.2 million people died from drug-resistant bacterial infections, exceeding the number of deaths caused by HIV/AIDS or malaria. K. pneumoniae is also one of the six major drug-resistant bacteria.2,3 The World Health Organization recognizes that extended-spectrum β-lactam (ESBL) and carbapenem-resistant K. pneumoniae (CRKP) pose significant public health threats. Currently, CRKP has been reported in Greece, China, Eastern Europe, Argentina and the Philippines.4 In Europe alone, this strain type is reported to cause over 90,000 infections and over 7000 deaths annually, accounting for 25% of deaths caused by multidrug-resistant (MDR) bacterial infections.5 The global resistance rate of MDR bacteria is increasing. The detection rate of CRKP is increasing from less than 0.1% to 24.6% in the United States. The resistance rate of K. pneumoniae isolates in Greek hospital wards reaches 52.4%. The China Antibiotic Surveillance Network (CHINET) shows that the resistance rate of K. pneumoniae to carbapenems increased from 2.4% to 32.8%.6
This increased resistance has severely limited therapeutic options and makes combating infections with multidrug-resistant (MDR) and extremely drug-resistant (XDR) K. pneumoniae a global challenge.7–9 The search for innovative therapeutic strategies has become particularly urgent. K. pneumoniae poses an immediate threat to humans and is becoming increasingly multi-resistant as the pathogen continues to develop. However, there are no approved treatments or vaccines other than traditional antibiotics, and there is an urgent need to accelerate the development of new antibiotics and explore new therapeutic approaches. In recent years, the number of published manuscripts on novel therapeutic strategies for K. pneumoniae has increased dramatically. In this review, we present the mechanisms of antibiotic resistance in K. pneumoniae and compare several new potential therapeutic strategies to overcome resistance in the treatment of K. pneumoniae infections.
Mechanisms of Antibiotic Resistance
Antibiotic resistance in K. pneumoniae is achieved through multiple mechanisms that enhance antimicrobial resistance through biofilm formation while also undergoing frequent horizontal transfer of antibiotic resistance genes;7 Production of a variety of β-lactamases, including broad-spectrum β-lactamases (eg TEM-30, TEM-31, TEM-50) and AmpC β-lactamases, capable of antibiotics, including clavulanate, vincristine and tazobactam, neutralize and sulbactam. Mutations in KPC enzymes, AmpC, efflux pumps, and outer membrane permeability result in resistance to β-lactam/β-lactamase inhibitor; Furthermore, mutations in efflux pumps (eg, AcrAB, KdeA, CepA, etc.) result in resistance to a variety of drugs, including mucins, tigecycline, and ceftazidime-avibactam.10 Aminoglycoside acetyltransferases (eg aac(3)-II and aac(6′)-Ib) and plasmid-mediated quinolone resistance are key factors in the resistance of K. pneumoniae to these drugs; Reduced membrane permeability, evidenced by the deletion or mutation of the pore proteins OmpK35 and OmpK36, as well as modification of the lipopolysaccharide structure (via phosphoethanolamine, galactosamine, etc.) have also been associated with resistance to peptide antibiotics. The various mechanisms by which K. pneumoniae develops resistance to multiple antibiotics and the associated resistance genes are summarized in Table 1.
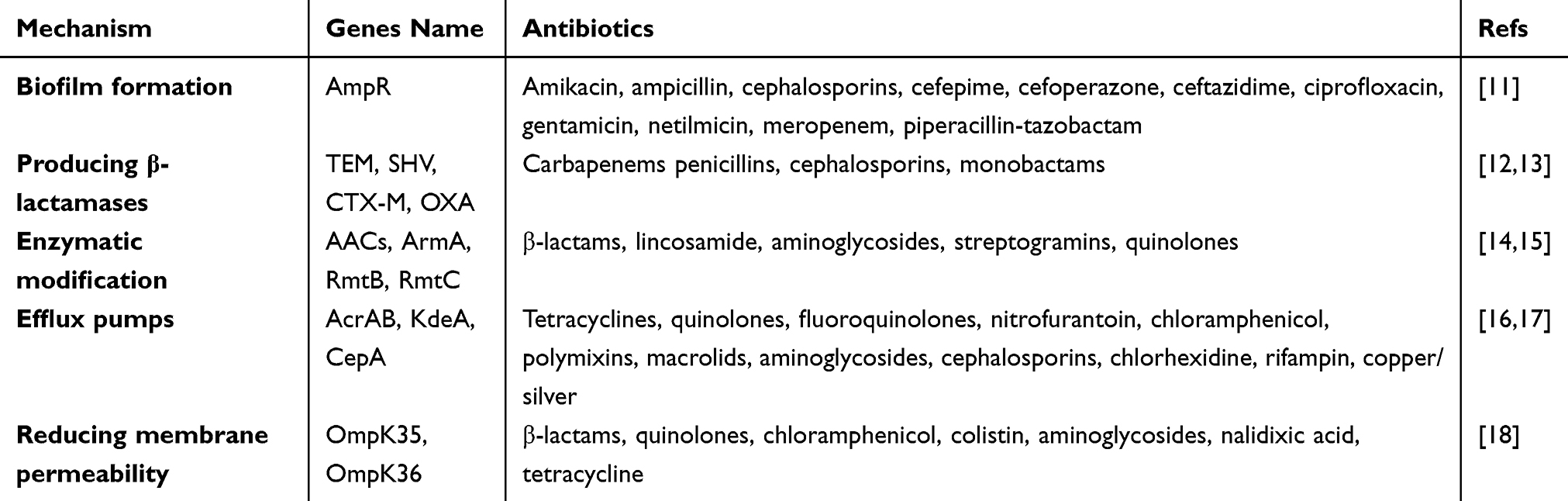 |
Table 1 K. pneumoniae Resistance to Multiple Antibiotics and the Associated Resistance Genes |
Biofilm Formation
Biofilms are highly resistant to antimicrobial drugs and immune responses and help organisms to attach to microbial communities, including proteins, extracellular DNA and extracellular polysaccharides, on living or non-living surfaces, providing a protective lifestyle for bacteria.19
K. pneumoniae usually forms biofilms on the surfaces of medical devices and objects. K. pneumoniae can protect bacteria from adverse conditions in the host body (eg, hypoxia and nutrient deficiency) as well as antimicrobial drugs through the formation of biofilms,20 which increase their persistence on the organism’s tissue surface and on medical devices. Biofilms increase bacterial resistance and are closely associated with high rates of resistance in clinical isolates of K. pneumoniae, with resistance to antimicrobial drugs being 10 to 1000 times higher in the biofilm state of K. pneumoniae than in the planktonic state.21 This increases the pressure on diagnosis and treatment.22 K. pneumoniae biofilm-forming strains isolated from medical devices were found to be 10–25 times more resistant to ceftazidime, ciprofloxacin, and gentamicin than free-living strains.23 Biofilm formation in K. pneumoniae is closely related to the luxS gene, and luxS, together with genes such as micF, ompK36 and ramA, can regulate the biofilm formation.24 Some researchers have also found that the acrA gene is upregulated to some extent during biofilm production25 and that a certain concentration of efflux pump inhibitor can promote the formation of K. pneumoniae biofilm.
Producing β-Lactamases
β-lactams, including carbapenems, cephalosporins, and penicillins, are widely used due to their therapeutic efficacy and wide range of options. In recent decades, the number of β-lactam-resistant bacteria (eg, cephalosporin-resistant Enterobacteriaceae) has increased rapidly worldwide. The production of β-lactamases is the main cause of resistance to this class of antimicrobials.26
K. pneumoniae produces extended-spectrum beta-lactamases (ESBLs) that are capable of hydrolyzing a variety of antibiotics such as penicillin and cephalosporins. Since the first report of penicillin resistance in K. pneumoniae in the 1960s, there are now more than 200 ESBLs, most of which can hydrolyze via serine residues and a small number of which use zinc ions to destroy the β-lactam ring.2 Carbapenemase is a type of ESBLs, and carbapenemase production is the most common mechanism resistance of K. pneumoniae to carbapenem antibiotics. Currently, carbapenemases are mainly divided into three categories: A, B and D. Class A includes KPC, SPM and SME; Class B includes metallo-β-lactamases such as NDM, VIM and IMP; and class D consists primarily of OXA, with classes A and D being serine carbapenemases. The genes encoding carbapenemases are mainly located on plasmids and spread through horizontal transmission, easily leading to nosocomial infection outbreaks.27 The AmpC enzyme is also a type of β-lactamase that evolved through the translocation of the AmpC gene from the chromosome to the plasmid.28 It not only hydrolyzes broad-spectrum β-lactam antibiotics, but also resists the inhibitory effect of β-lactamase inhibitors such as clavulanic acid, but the enzyme cannot hydrolyze carbapenems. However, if K. pneumoniae produces ESBLs or AmpC together with altered membrane pore proteins, then this is also an important mechanism for its resistance to carbapenem antibiotics.
Enzymatic Modification
Bacteria use a number of molecular strategies such as DNA mutations and protein translation modifications to resist antibiotics, so that they cannot interfere with bacterial metabolism and can be degraded intracellularly.29 Among the mechanisms of the main resistance of K. pneumoniae bacteria to aminoglycoside antibiotics, the most widespread is resistance mediated by aminoglycoside acetyltransferases enzymes which effectively prevents the binding of aminoglycoside antibiotics to the ribosome and deprives them of antibacterial activity. Aminoglycoside-acetyltransferase enzymes are present in the bacterial cytoplasm, which can be classified into (AACs), nucleotidyltransferases (ANTs), and phosphotransferases (APHs), etc. According to their modifying activity.14 The genes of aminoglycoside acetyltransferases enzymes are encoded by plasmids and transposons that can be easily transferred between strains, resulting in increased bacterial resistance to aminoglycoside antimicrobials.30 The most common aminoglycoside acetyltransferase enzymes in K. pneumoniae isolates are the enzymes aac(3)-II and aac(6′)-Ib.31 Furthermore, modification of 16SrRNA methyltransferases is a high-level aminoglycoside resistance mechanism in Gram-negative pathogens,32 and the prevalence of 16SrRNA methylase in clinical isolates of K. pneumoniae has been reported to be quite high. Genes encoding aminoglycoside acetyltransferase enzymes and 16S rRNA methylases (ArmA, RmtB and RmtC) are transferred to other organisms via mobile genetic elements linked to other resistance determinants such as ESBLs and carbapenemases, leading to the emergence of multidrug-resistant tribes leads.33
Efflux Pumps
Drug efflux pumps are active transport proteins that can actively excrete various antibiotics and other compounds from bacteria after substrate-induced activation of the efflux pump gene,1,34 thereby reducing the concentration of drugs in bacteria and thus generating drug resistance, and they include five families (SMR, MFS, RND, ABC and MATE families).
Currently, the efflux system of K. pneumoniae is dominated by the AcrAB-TolC system and the OqxAB system in the RND family, while the AcrAB-TolC system is dominant and β-lactam antibiotics, macrolide antibiotics, fluoroquinolone antibiotics, tetracycline antibiotics, amido-alcohol antibiotics etc. and disinfectants. The repressive regulatory genes of this system, acrR and ramR, carry deletion mutations that lead to overexpression of the AcrAB-TolC efflux pump, which in turn plays a key role in pan-resistance of K. pneumoniae.35,36 Autosomal transfer of the OqxAB gene to the plasmid can result in a ten-fold increase in its efflux pump expression level,37 and over expression of OqxAB reduces the susceptibility of K. pneumoniae to a variety of antimicrobial drugs by four-fold or more, resulting in the development of drug resistance.38
Reducing Membrane Permeability
Altered membrane permeability affects drug influx and hinders the interaction between the drug and its intracellular target, leading to resistance. Outer membrane proteins (OMP) or porins are trimeric protein channels on the outer membrane of gram-negative bacteria through which antimicrobial drugs can enter the bacterium and bind the corresponding proteins to act.39 When a mutation occurs in the bacterium that causes the outer membrane pore proteins to be absent or reduced, this results in a reduction in the number of antimicrobial drug molecules that can enter the bacterium, which can lead to bacterial drug resistance.40
K. pneumoniae primarily expresses two nonspecific porins, OmpK35 and OmpK36, which allow passive diffusion of hydrophilic small molecules and antimicrobial drugs (eg, β-lactams and fluoroquinolones) into the bacterium to exert antimicrobial effects. Since porin expression can be influenced by factors such as coding sequence or promoter site mutations or insertional deletions, resulting in deletion or alteration of porins, antimicrobial resistance can occur by preventing entry of antimicrobial agents into the bacterium. The expression of OmpK35 and OmpK36 porins in K. pneumoniae is inversely associated with ESBL production and confers resistance to cephalosporins, meropenem, ertapenem, ciprofloxacin and chloramphenicol.41 Strains lacking OmpK35 and OmpK36 also show high levels of resistance to antimicrobial drugs.42 Experiments in animal models of peritonitis have shown that deficiency of Ompk35/36 proteins reduces pathogenicity and increases resistance to various antibiotics.43
Therapeutic Strategies
Treatment of carbapenem-resistant K. pneumoniae (CRKP) infections is a major challenge, with polymyxins, tigecycline, fosfomycin and aminoglycosides being the main treatment agents.44 However, increasing antibiotic resistance and difficulties in developing new antibiotics have made the search for new therapeutic strategies an urgent task for the medical community.45 CRKP exhibits a high level of resistance to aminoglycosides and carbapenems, although carbapenems are still an effective option for the treatment of CRKP that does not produce carbapenemases.45 Antimicrobial combination therapies, particularly polymyxin-based combinations, have shown significant synergistic effects.46,47 Compared with traditional treatment regimens, new antibiotics and their synergistic combinations that have been marketed or approved in the later stages of the procedure have shown superior efficacy and safety.
Referring to the 2022 Infectious Diseases Society of America guidelines for the treatment of ultra-broad-spectrum β-lactamase-producing Enterobacteriaceae bacteria, carbapenem-resistant Enterobacteriaceae bacteria, and refractory drug-resistant Pseudomonas aeruginosa infections, the Conventional therapeutic regimens for K. pneumoniae summarized in Table 2. Given the increasing multidrug resistance in K. pneumoniae, it is essential to take new measures to combat drug-resistant bacteria, including the development of novel drugs and non-antibiotic alternatives that are both effective against infectious diseases and safe. The different treatment alternatives are described in Table 3 and Figure 1.
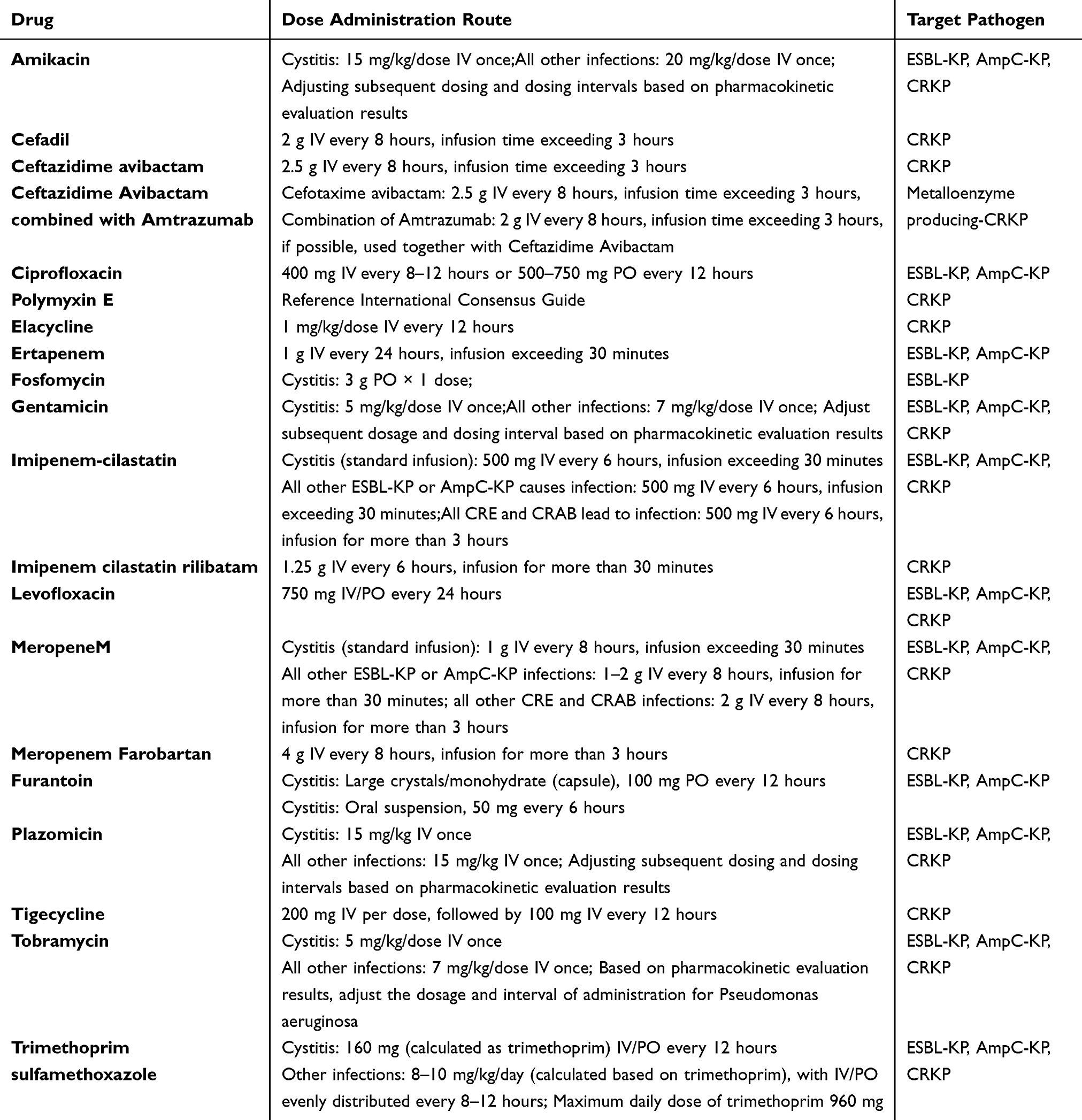 |
Table 2 Recommended Dosage of Antibiotics for Treating Infections Caused by K. Pneumoniae |
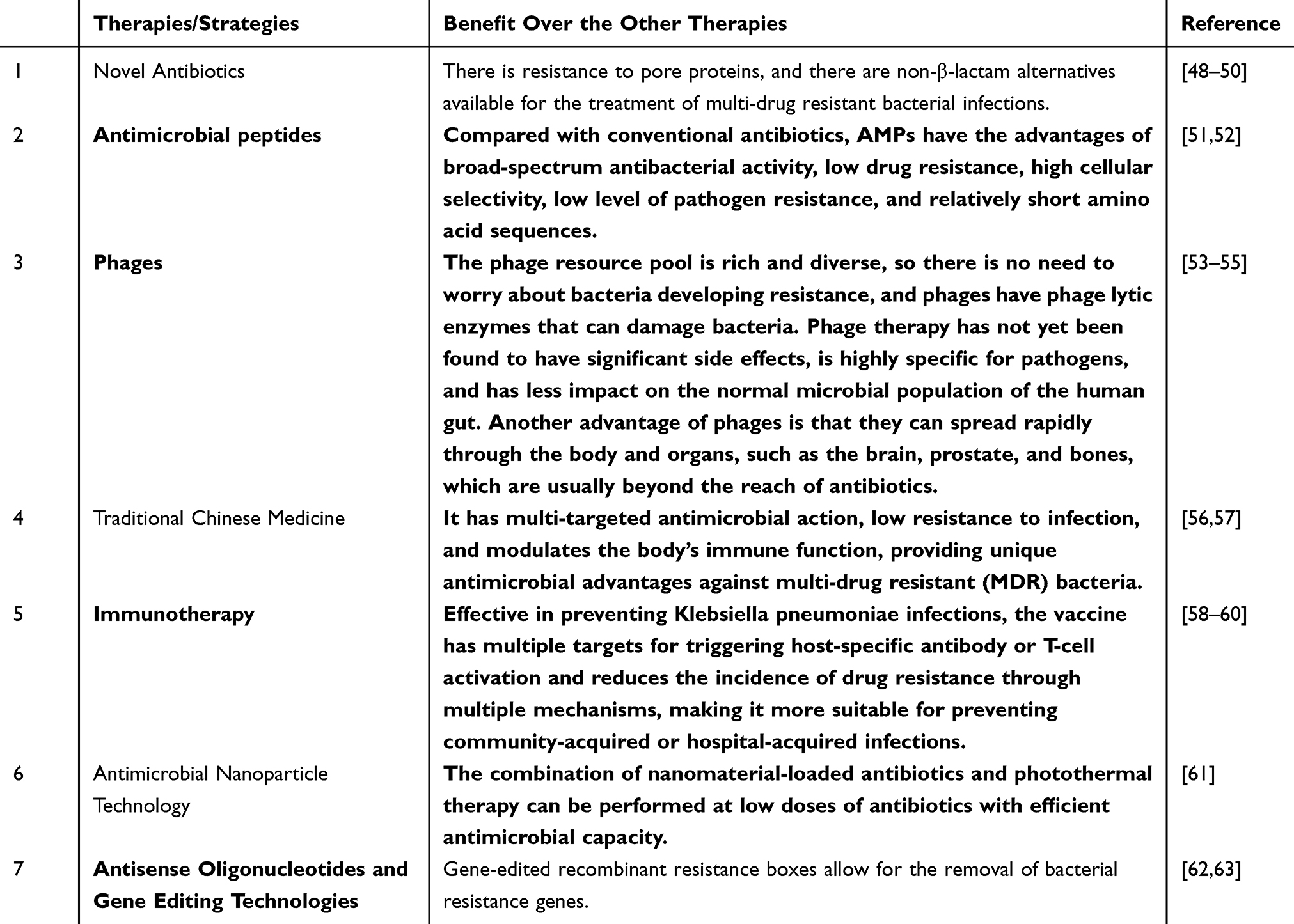 |
Table 3 A Account of Alternative Treatments for K. pneumoniae Infections Available |
 |
Figure 1 Novel Therapies/Strategies for the treatment of K. pneumoniae infections. |
Novel Antibiotics
Several studies have focused on finding drugs that act synergistically with polymyxins, such as sertraline, levopromazine, the LpxC inhibitor Chir-090, and selective estrogen receptor modulators.64–66 Meanwhile, the design and development of novel polymyxin analogues such as CB-182804 and Queensland aim to improve the activity and safety of polymyxins.67,68 Furthermore, the synthesis of novel antibiotics such as avibactam, plazomicin, eravacycline and cefiderocol as well as the development of synthetic peptides have provided new ideas for the treatment of CRKP infections.
Prazosin is a new semi-synthetic aminoglycoside antibiotic with better antibacterial activity against Klebsiella pneumoniae, which produces carbapenemases or ESBLs, which improves efficacy and reduces toxicity compared to mucomycin.69 Cefdinir is a new iron-carrying cephalosporin that enters the bacterial cell via an iron transport protein. Compared to ceftazidime or cefepime, it binds to free iron and increases its concentration in the bacterial periplasm, is extremely stable against carbapenemases and avoids resistance caused by mutations in the pore protein channel and overproduction of the efflux pump.70
The advent of bacterial genomics has opened the door to the discovery of new antibiotics. A recent study used a subtractive genomics approach to identify the bacterial DNA adenine methyltransferase of K. pneumoniae as a new drug target from hypothetical proteins. This suggests that it is theoretically possible to find important target proteins in K. pneumoniae using computational analysis methods.48 Although inhibiting these protein targets in vitro is challenging and requires overcoming obstacles in penetrating cellular constraints, this study succeeded in identifying potential new antibiotics, konembin, through a combination of computational drug design and bioinformatics approaches. In addition, microbial secondary metabolites found by the traditional route from ecosystems have shown potential for the discovery of new antibiotics, such as the antibiotic ludunin produced by Staphylococcus lugdunensis, which has been shown to have an inhibitory effect against gram-positive pathogens, including S. aureus.49 The application of microbiomics has also led to the discovery of new molecules with antimicrobial activity, such as tetracycline against methicillin-resistant Staphylococcus aureus soil microbiota and humin, which inhibits the enzyme lipid II flippase and acts synergistically with certain β-lactam antibiotics.50
Antimicrobial Peptides
Antimicrobial peptides (AMPs) are broad-spectrum, positively charged antimicrobial host defense peptides produced by all organisms in nature and represent a potential source of new drugs for the treatment of K. pneumoniae infections. Recent studies have shown that AMPs have made significant progress in combating pneumonia caused by this bacterium. As a class of immune effector molecules, AMPs act through a variety of mechanisms, including membrane permeability, inhibition of protein, DNA and RNA synthesis, and degradation of genetic material.51 This has led to AMPs having a broad spectrum of activity against pathogenic microorganisms. As a class of small-molecule peptides or small-molecule proteins, the antimicrobial mechanisms of AMPs include causing cell wall damage and cell membrane damage, influencing the activity of key enzymes for the synthesis and metabolism of biomolecules, and triggering immunoregulatory inhibitory mechanisms. As a result, antimicrobial peptides not only kill bacteria directly, but also mobilize the body’s own immune cells to participate in the regulation of inflammation.52 This unique antimicrobial mechanism makes AMPs a promising class of antimicrobial agents against Gram-positive and negative multidrug-resistant bacteria.
It has been found that AMPs can act synergistically when combined with difficult-to-penetrate bacteria or antibiotics in the context of membrane modifications, thereby improving therapeutic efficacy. For example, combination with mucomycin reduced the minimum inhibitory concentration (MIC) of mucomycin against MDR-K. pneumoniae strains by up to 11-fold without increasing the hemolytic effect of mucomycin on sheep erythrocytes, suggesting that the combination could improve the safety of mucomycin and the possible use of mucomycin could improve mucomycin therapy by using lower levels Expand mucomycin concentrations and/or shorter treatment regimens.71 AMPs also induced synergistic antimicrobial susceptibility of ST258 K. pneumoniae strains to rifampicin and azithromycin, suggesting that AMPs may act as antimicrobial enhancers.72
Although AMPs have shown remarkable activity in antimicrobial therapy, their application still faces several challenges, including lack of stability, susceptibility to hydrolysis by proteases, low activity under physiological conditions, and high production cost. Therefore, optimizing the structure of AMPs to improve their biological activity, extend half-life, and reduce cytotoxicity has become a current research focus in the field of AMPs. Although less than 50 AMPs have been enrolled in clinical trials worldwide, the listed AMP drugs such as anidulafungin, baclofungin, caspofungin, daptomycin, etc. provide useful experience and inspiration for the future application of AMPs.
Phages
Phages are viruses that infect and lyse bacteria and have the potential to treat severe drug-resistant bacterial infections. Phage therapy for K. pneumoniae infections is still in the early stages of research, with sepsis and pneumonia being the most studied in vivo models for phage therapy of K. pneumoniae infections.73
One of the major virulence factors that make K. pneumoniae immune to immune responses is podoplanin polysaccharides. Phages can be designed to specifically target podoplanin polysaccharides in K. pneumoniae. Phages provide a targeted approach to weaken K. pneumoniae defenses such as biofilms and reduce their virulence. The use of phages holds promise for the treatment of CRKP infections. Certain phages, such as phage vB_kpnM_17-11, have demonstrated promising efficacy against K. pneumoniae infections in vitro and in vivo, with in vitro assays demonstrating a 105-fold reduction in the number of KPs by vB_kpnM_17-11. In a mouse model of infection, phage treatment improved survival;53 it was demonstrated that phage pKp11 and pKp383 lysed 51 strains of multidrug-resistant K. pneumoniae (including strains ST11 and ST383), providing an effective treatment for the early stages of pneumonia in a mouse model of infection without significant side effects.54 Scientists at the National Institutes of Health (NIH) successfully treated mice infected with multidrug-resistant K. pneumoniae ST258 using two different phage viruses, alone and in combination. These findings are encouraging and show successful therapeutic effects in animal models.55
Phage depolymerases are proteins that specifically degrade bacterial surface polysaccharides, specifically recognize bacterial pod types, remove bacterial biofilms, and effectively reduce bacterial virulence.53 In recent years, phage and phage depolymerization enzymes have shown promise in the treatment of CRKP infections. However, phage therapy faces limitations in clinical application due to the lack of standardized guidelines and the variable nature of phage-bacteria interactions. Despite these challenges, more and more researchers are realizing the therapeutic value of phages and it is believed that phage therapy will play an important role in clinical settings in the future as research continues to progress.
Traditional Chinese Medicine
The antimicrobial effects of Traditional Chinese medicines (TCM) against K. pneumoniae have been confirmed by several studies, such as the inhibitory effect of baicalein from Scutellaria baicalensis on the growth and biofilm formation of highly virulent K. pneumoniae,74 and the synergistic effect of baicalein combined with imipenem on CRKP, as well as the ability to reduce the resistance of CRKP to imipenem.75 Studies on baicalein, the active ingredient in the root bark of Paeonia lactiflora, showed that it had significant antibacterial and antibiofilm activity against K. pneumoniae. Paeoniflorin effectively inhibited the growth of K. pneumoniae with a minimum inhibitory concentration (MIC) of 64 μg/mL. it was found to disrupt the integrity of bacterial cell membranes and alter cell morphology. In addition, paeoniflorin showed potent effects in inhibiting bacterial adhesion and biofilm formation, which are key factors in the process of bacterial infection. It also disperses mature biofilms; therefore, the current study suggests that bryonoxin is a promising new antibacterial and anti-biofilm agent for the treatment of infections caused by K. pneumoniae.56
Huanglianin is an important alkaloid extracted from the plants such as Cortex Eucommiae, Phellodendron Bark, and Rhizoma Coptidis, and it is recognised as an antibacterial drug in traditional Chinese medicine. Huanglianin can inhibit the formation of K. pneumoniae biofilm by down-regulating the expression of the genes for type I and type III bacterial hairs, reducing the surface adhesion ability of K. pneumoniae, and thus effectively inhibiting the formation of K. pneumoniae biofilm. Recent studies have explored the mechanisms of Chinese herbal medicines against K. pneumoniae resistance, revealing their multiple modes of action. Herbal medicines are able to inhibit β-lactamase (an enzyme used by bacteria to resist β-lactam antibiotics), counteract biofilm formation (which protects the structure of the bacterial community), counteract the effects of endotoxin (a component of the outer membrane of Gram-negative bacteria such as K. pneumoniae), and inhibit the exocytosis pump (a mechanism used by bacteria to excrete antibiotics and other harmful substances). By understanding these mechanisms, the effects of herbal medicines can be accurately assessed to inform the selection of appropriate antibiotic therapy in the future57
K. pneumoniae, especially those pan-resistant strains, pose a major challenge in clinical treatment due to their high resistance rates. TCM has attracted widespread attention as a novel approach against multi-drug resistant bacteria, either alone or in combination with western drugs, due to its low resistance and broad-spectrum antimicrobial effect against multiple targets of action.
Immunotherapy
In response to the problem of drug resistance in K. pneumoniae, especially in the context of the epidemic of drug-resistant bacterial infections and drug-resistant strains faced globally, the production of vaccines and antibodies based on active and passive immunization has provided new ideas for the prevention and treatment of infections with multidrug-resistant bacteria. Immunotherapy has emerged as a promising means to break the dead-end cycle of antibiotic resistance-new antibiotic development-new antibiotic resistance.
Vaccine studies for K. pneumoniae infections are currently in the research and development phase, using strategies involving whole cell vaccines, outer membrane vesicles (OMVs), ribosomes, polysaccharides, lipopolysaccharides (LPS) and protein vaccine formulations.58 The goal of these approaches is to induce antibodies and cell-mediated immune responses to effectively combat K. pneumoniae as a pathogen.58 Although overcoming challenges such as the diversity of K. pneumoniae strains remains a key issue, preclinical studies have shown that vaccine-induced antimicrobial antibodies are protective against K. pneumoniae infection.59 In addition, monoclonal antibodies have been explored as one of the strategies against drug-resistant bacteria. About 10 monoclonal antibodies are in clinical trials, of which three have been approved for marketing.60 Despite the encouraging progress in the development of vaccines, the diversity of K. pneumoniae strains presents significant challenges that need to be overcome to create effective vaccines. Ongoing research is essential to find effective vaccines that offer protection, especially against this increasingly resistant pathogen.
Antimicrobial Nanoparticle Technology
Studies have shown that metal nanoparticles (eg, silver, zinc oxide, magnesium, nitrogen oxide, etc.) possess antimicrobial activity, and their mechanisms of action include induction of oxidative stress, non-oxidative mechanisms, and interactions of released metal ions with functional groups of proteins and nucleic acids. Factors such as the size, zeta potential, charge, surface morphology and crystal structure of the nanoparticles determine their antimicrobial activity.76
Common types of antimicrobial nanoparticles include silver nanoparticles, metal oxide nanoparticles, photothermal conversion nanoparticles, and antibiotic conjugated nanoparticles. These nanoparticles achieve their antimicrobial effects in a variety of ways, including disrupting bacterial cell membranes, interacting with bacterial DNA or proteins, and indirectly initiating ROS production.61
Researchers explored the antibacterial and antibiofilm efficiency of zinc ferrate nanoparticles (ZnFeO NPs) against K. pneumoniae. The results showed that ZnFeO nanoparticles exhibited excellent antimicrobial efficiency, inducing reactive oxygen species (ROS)-induced bacterial damage, as well as significant results in anti-biofilm.77 The therapeutic possibilities of ZnFeO NPs in antimicrobial applications, especially their overcoming of challenges related to biofilm infectivity. This offers promising avenues for finding more effective and sustainable antimicrobial practices.
Overall, antimicrobial nanoparticle technology shows potential efficacy in the treatment of K. pneumoniae infections, offering promising avenues for overcoming drug resistance and providing new therapeutic options.
Antisense Oligonucleotides and Gene Editing Technologies
Research on antisense oligonucleotides and gene editing technologies in the treatment of K. pneumoniae infections has shown impressive progress. By classifying and applying different types of antisense oligonucleotides, researchers have successfully achieved the inhibition of gene expression in K. pneumoniae, specifically including the inhibition of gene expression by RNA silencing, especially the generation of resistance to antibiotics.78 In order to overcome the problem of antisense oligonucleotides reaching sufficiently high concentrations in bacteria, cell-penetrating peptide (CPP) was introduced, a strategy that is expected to improve the effective delivery of oligonucleotides in cells and provide a new direction for the development of antimicrobial agents.78
On the other hand, gene editing technologies, especially the CRISPR/Cas system, have been applied to programmable modification of the K. pneumoniae genome. The CRISPR/Cas system is capable of re-sensitizing bacteria to antibiotics by selectively eliminating drug-resistant genes, and even by eliminating plasmids containing drug-resistant genes.62 This provides a promising solution for the treatment of drug-resistant K. pneumoniae infections. However, despite the remarkable progress, the CRISPR/Cas system still faces a series of challenges for in vivo application, including the problems of efficient delivery to bacteria and the side effects of potential off-target modifications, which need to be solved by future intensive research.62,63 Therefore, antisense oligonucleotide and gene editing technologies provide new ideas and possibilities for the treatment of K. pneumoniae infections, offering promising solutions to tackle the drug resistance problem.
Other Emerging Strategies
In terms of new strategies f to treat K. pneumoniae infections, researchers have employed a variety of innovations to address the serious challenge of antimicrobial drug resistance. These new strategies include host-directed therapy (HDT), virulence factor inhibitors, QS inhibitors, and nanoenzymes.79 HDT, a promising alternative therapy to antibiotics, has the potential to eradicate pathogenic bacteria by targeting host factors and eliminating intracellular replication of pathogenic bacteria while stimulating the host immune response to the pathogen.79 Virulence factor inhibitors, particularly molecules targeting pathogen virulence factors, are expected to disarm bacteria, slow down the development of resistance, and reduce the pathogenicity of the strain.
On the other hand, QS inhibitors achieve antimicrobial effects by inhibiting signaling molecular communication between bacteria and realizing the regulation of physiological processes, such as bacterial metabolism, virulence, and biofilm formation.80 Nanoenzymes, as a new generation of artificial mimetic enzymes, combine the unique physicochemical properties of nanomaterials with enzyme-like catalytic activity, and have the advantages of structural stability, broad antimicrobial spectrum, and low production cost. The introduction of these new strategies offers diverse options for the treatment of K. pneumoniae infections and is expected to address the problem of antibiotic resistance.
In addition, there are intervention strategies for the treatment of K. pneumoniae infections that target specific virulence factors, including determinants involved in host cell attachment, host immune modulation, and biofilm regulation.81–83 Research and development of these strategies is expected to provide more effective means of treating drug-resistant K. pneumoniae infections.
Conclusion
With the widespread use of antibiotics in recent decades, the incidence of multidrug-resistant K. pneumoniae isolates has increased frequently worldwide, posing a major threat to public health. K. pneumoniae is a priority pathogen for research and development of new therapeutic approaches, but its innovation remains a challenge. In this paper, the mechanisms and effects of emerging therapeutic strategies for K. pneumoniae in recent years are summarized, Potential therapies were also be characterized in Figure 1. The application of these new technologies and their synergistic application with various antimicrobial compounds may be a new direction for future research. In addition, preventive measures are more important than therapeutic measures. Early detection, early diagnosis and early treatment, reduction and avoidance of high-risk factors for CRKP infection, as well as increased management of antimicrobials and drug resistance monitoring, can play a multiplying role in controlling the spread of K. pneumoniae infection.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bengoechea JA, Pessoa JS. Klebsiella pneumoniae infection biology: living to counteract host defences. FEMS Microbiol Rev. 2019;2019:2.
2. Collaborators AR, Darboe S, Carvalheiro CG, Haller S, Roca A. Articles global burden of bacterial antimicrobial resistance in 2019: a systematic analysis Antimicrobial Resistance Collaborators*. Lancet. 2022;2022:1.
3. Murray CJL, Ikuta KS, Sharara F, et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;10325:399.
4. Diallo OO, Baron SA, Abat C, et al. Antibiotic resistance surveillance systems: a review. J Glob Antimicrob Resist. 2020;23:430–438.
5. B ACA, A LDH, A DP, et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: a population-level modelling analysis. Lancet Infect Dis. 2018;2018:1.
6. Hu F, Guo Y, Zhu D, et al. CHINET surveillance of antimicrobial resistance among the bacterial isolates in 2021. Chin J Infect Chemother. 2021;25(5):20.
7. Li Y, Kumar S, Zhang L, Wu H, Wu H. Characteristics of antibiotic resistance mechanisms and genes of Klebsiella pneumoniae. Open Med. 2023;18:1. doi:10.1515/med-2023-0707
8. Pei N, Sun W, He J, et al. Genome-wide association study of Klebsiella pneumoniae identifies variations linked to carbapenems resistance. Front Microbiol. 2022;13:997769. doi:10.3389/fmicb.2022.997769
9. Wang L, Yuan XD, Pang T, Duan SH. The risk factors of carbapenem-resistant Klebsiella pneumoniae infection: a Single-Center Chinese Retrospective Study. Infect Drug Resist. 2022;15:1477–1485. doi:10.2147/IDR.S352070
10. Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol Rev. 2017;41(3):252–275. doi:10.1093/femsre/fux013
11. Hennequin C, Robin F, Cabrolier N, Bonnet R, Forestier C. Characterization of a DHA-1-producing Klebsiella pneumoniae strain involved in an outbreak and role of the AmpR regulator in virulence. Antimicrob Agents Chemother. 2012;56(1):288–294. doi:10.1128/AAC.00164-11
12. Garcia-Fernandez S, Bala Y, Armstrong T, et al. Multicenter evaluation of the new etest gradient diffusion method for piperacillin-tazobactam susceptibility testing of enterobacterales, Pseudomonas aeruginosa, and Acinetobacter baumannii complex. J Clin Microbiol. 2020;58:2. doi:10.1128/JCM.01042-19
13. Abdelraouf K, Chavda KD, Satlin MJ, Jenkins SG, Kreiswirth BN, Nicolau DP. Piperacillin-tazobactam-resistant/third-generation cephalosporin-susceptible Escherichia coli and Klebsiella pneumoniae isolates: resistance mechanisms and in vitro-in vivo discordance. Int J Antimicrob Agents. 2020;55(3):105885. doi:10.1016/j.ijantimicag.2020.105885
14. Ramirez MS, Tolmasky ME. Aminoglycoside modifying enzymes. Drug Resist Updates. 2010;13(6):151–171. doi:10.1016/j.drup.2010.08.003
15. Liao W, Liu Y, Zhang W. Virulence evolution, molecular mechanisms of resistance and prevalence of ST11 carbapenem-resistant Klebsiella pneumoniae in China: a review over the last 10 years. J Glob Antimicrob Resist. 2020;23:174–180. doi:10.1016/j.jgar.2020.09.004
16. Rodriguez-Martinez JM, Diaz de Alba P, Briales A, et al. Contribution of OqxAB efflux pumps to quinolone resistance in extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae. J Antimicrob Chemother. 2013;68(1):68–73. doi:10.1093/jac/dks377
17. Zhong X, Xu H, Chen D, Zhou H, Hu X, Cheng G. First emergence of acrAB and oqxAB mediated tigecycline resistance in clinical isolates of Klebsiella pneumoniae pre-dating the use of tigecycline in a Chinese hospital. PLoS One. 2014;9(12):e115185. doi:10.1371/journal.pone.0115185
18. Chen JH, Lin JC, Chang JL, Tsai YK, Siu LK. Different culture medium formulations induce variant protein expression patterns of outer membrane porins in Klebsiella pneumoniae. J Chemother. 2011;23(1):9–12. doi:10.1179/joc.2011.23.1.9
19. Al-Mogbel MS, Khan M, Shadeed M, Meqdam M, Hossain A. Influence of growth in biofilm in the formation of new biofilm by clinical isolates of Acinetobacter baumannii. J Pure Appl Microbiol. 2015;Spec.1:9.
20. Perez L. Equal, but different: fluctuant biofilm formation and its impact on polymyxin B susceptibility among a clonal spreading of KPC-2-producing Klebsiella pneumoniae isolates. Infect Control Hosp Epidemiol. 2019;40(8):954–955. doi:10.1017/ice.2019.106
21. Mohamed SH, Mohamed MSM, Khalil MS, Azmy M, Mabrouk MI. Combination of essential oil and ciprofloxacin to inhibit/eradicate biofilms in multidrug-resistant Klebsiella pneumoniae. J Appl Microbiol. 2018;125(1):84–95. doi:10.1111/jam.13755
22. Flemming H-C, Wingender J, Szewzyk U, Steinberg P, Rice SA, Kjelleberg S. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol. 2016;14(9):563–575. doi:10.1038/nrmicro.2016.94.
23. Vuotto C, Longo F, Balice M, Donelli G, Varaldo P. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens. 2014;3(3):743–758. doi:10.3390/pathogens3030743
24. Vuotto CL, F.Pascolini CD, G.Balice MPL, F.Tiracchia M, V.Salvia AV, E P. Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. In Vitro Cell Dev Biol Plant. 2017;53:3.
25. Wei X. Antibiotic Resistance Profile of Klebsiella Pneumoniae and the Relationship with Biofilm Formation and Efflux Pump Genes. Southwest Medical University; 2018.
26. Bevan ER, Jones AM, Hawkey PM, et al. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–2155. doi:10.1093/jac/dkx146
27. Savov E, Todorova I, Politi L, Trifonova A, Borisova M. Colistin resistance in KPC-2- and SHV-5-producing Klebsiella pneumoniae clinical isolates in Bulgaria. Chemotherapy. 2017;62(6):339–342. doi:10.1159/000464275
28. Sidjabat HE, Silveira FP, Potoski BA, et al. Interspecies spread of Klebsiella pneumoniae carbapenemase gene in a single patient. Clinl Infect Dis. 2009;49(11):1736–1738. doi:10.1086/648077
29. Schaenzer AJ, Wright GD. Antibiotic resistance by enzymatic modification of antibiotic targets. Trends Mol Med. 2020;26:768–782.
30. Radlinski LC, Rowe SE, Brzozowski R, Wilkinson AD, Conlon BP. Chemical induction of aminoglycoside uptake overcomes antibiotic tolerance and resistance in Staphylococcus aureus. Cell Chem. Biol. 2019;26:10. doi:10.1016/j.chembiol.2019.07.009
31. Butler DA, Rana AP, Krapp F, et al. Optimizing Aminoglycoside Selection for KPC-Producing Klebsiella pneumoniae with the Aminoglycoside-Modifying Enzyme (AME) Gene aac(6’)-Ib. Oxford Academic. 2021:3.
32. Shanshan SU, Xue G, Jisheng Z, et al. Antibiotic resistance mechanism and homology analysis of carbapenem-resistant Klebsiella pneumoniae outbreak in ICU. Chin J Infect Chemother. 2018. doi:10.1016/j.jiac.2018.05.003
33. Haldorsen BRC, Simonsen GS, Sundsfjord A, Samuelsen R. Increased prevalence of aminoglycoside resistance in clinical isolates of Escherichia coli and Klebsiella spp. in Norway is associated with the acquisition of AAC(3)-II and AAC(6’)-Ib. Diagn Microbiol Infect Dis. 2014;78(1):66–69. doi:10.1016/j.diagmicrobio.2013.10.001
34. Chiu SK, Chan MC, Huang LY, et al. Tigecycline resistance among carbapenem-resistant Klebsiella Pneumoniae: clinical. PloS One. 2017;12:e0175140.
35. Lv F, Cai J, He Q, Wang WQ, Luo W. Overexpression of efflux pumps mediate pan resistance of Klebsiella pneumoniae sequence type 11. Microb Drug Resist. 2021;27(10):1405–1411. doi:10.1089/mdr.2020.0395
36. Q Z, L L, Y P, J C. Characterization of tigecycline-heteroresistant Klebsiella pneumoniae clinical. Front Microbiol. 2021;12:671153.
37. Wong MHY, Chan EWC, Chen S. Evolution and dissemination of OqxAB-like efflux pumps, an emerging quinolone resistance determinant among members of Enterobacteriaceae. Antimicrob Agents Chemother. 2015;59(6):3290–3297. doi:10.1128/AAC.00310-15
38. Li J, Zhang H, Ning J, et al. The nature and epidemiology of OqxAB, a multidrug efflux pump. Antimicrob Resist Infect Control. 2019;8(1). doi:10.1186/s13756-019-0489-3
39. Vergalli J, Bodrenko IV, Masi M, Moynié L, Pagès J-M. Porins and small-molecule translocation across the outer membrane of gram-negative bacteria. Nat Rev Microbiol. 2019;18:3.
40. Choi U, Lee CR, Zhou H. Distinct roles of outer membrane porins in antibiotic resistance and membrane integrity in Escherichia coliData_Sheet_1.docx. Front Microbiol. 2019;10:10. doi:10.3389/fmicb.2019.00010
41. Hamzaoui Z, Ocampo-Sosa A, Maamar E, et al. An outbreak of NDM-1-producing Klebsiella pneumoniae, associated with OmpK35 and OmpK36 Porin Loss in Tunisia. Microb Drug Resist. 2018;24:1137.
42. B AKA, b AFLA, C LM, d RCYLAB, E JRIAB. Characterizing the role of porin mutations in susceptibility of beta lactamase producing Klebsiella pneumoniae isolates to ceftaroline and ceftaroline-avibactam - ScienceDirect. Inter J Infect Dis. 2020;93:252–257. doi:10.1007/s00284-016-1034-8
43. Chung HS, Yong D, Lee M. Mechanisms of ertapenem resistance in Enterobacteriaceae isolates in a tertiary university hospital. J Investig Med. 2016;64(5):1042. doi:10.1136/jim-2016-000117
44. Chang-Ro L, Jung H, Lee K, Seung P, Young B. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front Microbiol. 2016;7. doi:10.3389/fmicb.2016.00895
45. Sommer LM, Johansen HK, Molin S. Antibiotic resistance in Pseudomonas aeruginosa and adaptation to complex dynamic environments. Microb Genom. 2020;6(5):1.
46. Liu E, Jia P, Li X, Zhou M, Yang Q. In vitro and in vivo effect of antimicrobial agent combinations against carbapenem-resistant Klebsiella pneumoniae with different resistance mechanisms in China. Infect Drug Resist. 2021;14:917–928. doi:10.2147/IDR.S292431
47. Sheu CC, Chang YT, Lin SY, Chen YH, Hsueh PR. Infections caused by carbapenem-resistant Enterobacteriaceae: an update on therapeutic options. Front Microbiol. 2019;10:80. doi:10.3389/fmicb.2019.00080
48. Omeershffudin UNM, Kumar S. Antimicrobial resistance in Klebsiella pneumoniae: identification of bacterial DNA adenine methyltransferase as a novel drug target from hypothetical proteins using subtractive genomics. Genomics Inform. 2022;20(4):e47. doi:10.5808/gi.22067
49. Janek D, Brotz-Oesterhelt H, Weidenmaier C, Peschel A, Schilling N. Human commensals producing a novel antibiotic impair pathogen colonization. Nature. 2016;539(7628):314. doi:10.1038/nature19781
50. Kallifidas D, Kang HS, Brady SF. Tetarimycin A, an MRSA-active antibiotic identified through induced expression of environmental DNA gene clusters. J Am Chem Soc. 2012;134(48):19552. doi:10.1021/ja3093828
51. Molchanova N, Hansen PR, Franzyk H. Advances in development of antimicrobial peptidomimetics as potential drugs. Molecules. 2017;22(9):1430. doi:10.3390/molecules22091430
52. Olekson MA, You T, Savage PB, Leung KP. Antimicrobial ceragenins inhibit biofilms and affect mammalian cell viability and migration in vitro. Febs Open Bio. 2017;7(7):953–967. doi:10.1002/2211-5463.12235
53. Bai J, Zhang F, Liang S, et al. Isolation and characterization of vB_kpnM_17-11, a novel phage efficient against carbapenem-resistant Klebsiella pneumoniae. Front Cell Infect Microbiol. 2022;12:897531. doi:10.3389/fcimb.2022.897531
54. Gan L, Fu H, Tian Z, et al. Bacteriophage effectively rescues pneumonia caused by prevalent multidrug-resistant Klebsiella pneumoniae in the early stage. Microbiol Spectr. 2022;10(5):e0235822. doi:10.1128/spectrum.02358-22
55. Ichikawa M, Nakamoto N, Kredo-Russo S, et al. Bacteriophage therapy against pathologicalKlebsiella pneumoniae ameliorates the course of primary sclerosing cholangitis. Nat Commun. 2023;14:1. doi:10.1038/s41467-023-39029-9
56. Qian W, Zhang J, Wang W, et al. Antimicrobial and antibiofilm activities of paeoniflorin against carbapenem-resistant Klebsiella pneumoniae. J Appl Microbiol. 2020;128(2):401–413. doi:10.1111/jam.14480
57. Pei-feng CZ. Screening of the traditional Chinese medicine with antibacterial activity against pan-drug resistant Klebsiella pneumoniae in vitro. J Bengbu Med Coll. 2020;45(4):515–519.
58. Assoni L, Girardello R, Converso TR, Darrieux M. Current stage in the development of Klebsiella pneumoniae vaccines. Infect Dis Ther. 2021;10(4):2157–2175. doi:10.1007/s40121-021-00533-4
59. Frost I, Sati H, Garcia-Vello P, et al. The role of bacterial vaccines in the fight against antimicrobial resistance: an analysis of the preclinical and clinical development pipeline. Lancet Microbe. 2023;4(2):e113–e125. doi:10.1016/S2666-5247(22)00303-2
60. Wantuch PL, Rosen DA. Klebsiella pneumoniae: adaptive immune landscapes and vaccine horizons. Trends Immunol. 2023;44(10):826–844. doi:10.1016/j.it.2023.08.005
61. Kedziora A, Korzekwa K, Strek W, Pawlak A, Doroszkiewicz W, Bugla-Ploskonska G. Silver nanoforms as a therapeutic agent for killing Escherichia coli and certain ESKAPE pathogens. Curr Microbiol. 2016;73(1):139–147.
62. Kim JS, Cho DH, Park M, Chung WJ, Kweon DH. CRISPR/Cas9-mediated re-sensitization of antibiotic-resistant Escherichia coli harboring extended-spectrum β-lactamases. J Microbiol Biotechnol. 2016;26(2):1.
63. Gholizadeh P, Kse Ü, Dao S, Ganbarov K, Kafil HS. How CRISPR-Cas system could be used to combat antimicrobial resistance. Infect Drug Resist. 2020;13:1111–1121. doi:10.2147/IDR.S247271
64. Hussein MH, Schneider EK, Elliott AG, Han M, Velkov T. From breast cancer to antimicrobial: combating extremely resistant gram-negative ”superbugs” using novel combinations of polymyxin B with selective estrogen receptor modulators. Microb Drug Resist. 2016;23(5):640–650. doi:10.1089/mdr.2016.0196
65. Jun H, Tan C, Vidaillac J, Kuok H, Yam S. In vitro and in vivo efficacy of an LpxC Inhibitor, CHIR-090, alone or combined with colistin against Pseudomonas aeruginosa biofilm. Antimicrob. Agents Chemother. 2017;2017:1.
66. Otto RG, Van Gorp E, Kloezen W, Meletiadis J, Sanne VDB, Mouton JW. An alternative strategy for combination therapy: interactions between polymyxin B and non-antibiotics. Int J Antimicrob Agents. 2019;53:34.
67. Velkov T, Roberts KD. Discovery of novel polymyxin-like antibiotics. Adv Exp Med Biol. 1145;2019:343–362.
68. Francesc R, Cajal Y. Recent advances and perspectives in the design and development of polymyxins. Nat Prod Rep. 2017;34(7):886–908. doi:10.1039/c7np00023e
69. Eljaaly K, Alharbi A, Alshehri S, Ortwine JK, Pogue JM. Plazomicin: a novel aminoglycoside for the treatment of resistant gram-negative bacterial infections. Drugs. 2019;79:243.
70. Karaiskos I, Galani I, Papoutsaki V, et alCarbapenemase producing Klebsiella pneumoniae: implication on future therapeutic. Expert Rev Anti Infect Ther. 2022;1744-8336:53–69.
71. Witherell KS, Price J, Bandaranayake AD, Olson J, Call DR. In vitro activity of antimicrobial peptide CDP-B11 alone and in combination with colistin against colistin-resistant and multidrug-resistant Escherichia coli. Sci Rep. 2021;11(1):2151. doi:10.1038/s41598-021-81140-8
72. Baker KR, Jana B, Hansen AM, Nielsen HM, Franzyk H, Guardabassi L. Repurposing azithromycin and rifampicin against gram-negative pathogens by combination with peptidomimeticsData_Sheet_1.PDF. Front Cell Infect Microbiol. 2019;9. doi:10.3389/fcimb.2019.00236
73. Iszatt JJ, Larcombe AN, Chan HK, et al. Phage therapy for multi-drug resistant respiratory tract infections. Viruses. 2021;13(9):1809.
74. Yi-qiang HJ. A preliminary study on the effects of different concentrations of baicalin on the growth and biofilm formation of hypervirulent Klebsiella pneumoniae. J Clin Pulm Med. 2022;27(3):342–345.
75. Tang JR, Zhang D, Li S. Antibacterial activity of four traditional Chinese medicine momonomers combined with imipenem against carbapenem-resistant Klebsiella Pneumoniae in vitro. J Mod Lab Med. 2022;37(6):162–166.
76. Slomberg DL, Lu Y, Broadnax AD, Hunter RA, Carpenter AW, Schoenfisch MH. Role of size and shape on biofilm eradication for nitric oxide-releasing silica nanoparticles. ACS Appl Mater Interfaces. 2013;5(19):9322–9329. doi:10.1021/am402618w
77. Sharma RP, Raut SD, Jadhav VV, Mulani RM, Kadam AS, Mane RS. Assessment of antibacterial and anti-biofilm effects of zinc ferrite nanoparticles against Klebsiella pneumoniae. Folia Microbiol. 2022;67(5):747–755.
78. Abushahba MFN, Mohammad H, Thangamani S, Hussein AAA, Seleem MN. Impact of different cell penetrating peptides on the efficacy of antisense therapeutics for targeting intracellular pathogens. Rep. 2016;6(1):20832.
79. Mohan M, Bhattacharya D. Host-directed therapy: a new arsenal to come. Comb Chem High Throughput Screen. 2021;24(1):59–70. doi:10.2174/1386207323999200728115857
80. Sikdar R, Elias M. Quorum quenching enzymes and their effects on virulence, biofilm, and microbiomes: a review of recent advances. Exp Rev Anti-Infective Ther. 2020;1:1–13.
81. Vila J, Moreno-Morales J, Ballesté-Delpierre C. Current landscape in the discovery of novel antibacterial agents. Clin Microbiol Infect. 2020;26(5):5. doi:10.1016/j.cmi.2019.09.015
82. Aggarwal C, Jimenez JC, Lee H, Chlipala GE, Ratia K, Federle MJ. Identification of quorum-sensing inhibitors disrupting signaling between rgg and short hydrophobic peptides in Streptococci. Mbio. 2015;6(3). doi:10.1128/mBio.00393-15
83. Ursula T, Laura J, Piddock V. Non-traditional ANTIBACTERIAL THERAPEUTIC OPTIONS AND CHALLENGes - ScienceDirect. Cell Host Microbe. 2019;26(1):61–72. doi:10.1016/j.chom.2019.06.004
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.