Back to Journals » OncoTargets and Therapy » Volume 13
Long Noncoding RNA LINC01485 Promotes Tumor Growth and Migration via Inhibiting EGFR Ubiquitination and Activating EGFR/Akt Signaling in Gastric Cancer
Authors Zhou J, Wu L, Li W, Xu X, Ju F, Yu S, Guo J, Li G, Shi J, Zhou S
Received 6 April 2020
Accepted for publication 10 July 2020
Published 21 August 2020 Volume 2020:13 Pages 8413—8425
DOI https://doi.org/10.2147/OTT.S257151
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
Jianping Zhou1 ,* Lulu Wu1 ,* Weiling Li2 ,* Xiao Xu,1 Feng Ju,1 Shao Yu,1 Jianfeng Guo,3 Gang Li,3 Jun Shi,1 Sujun Zhou1
1Department of Gastrointestinal Surgery, The Affiliated Yixing Hospital of Jiangsu University, Yixing, Jiangsu, People’s Republic of China; 2Department of Obstetrics and Gynecology, The Affiliated Yixing Hospital of Jiangsu University, Yixing, Jiangsu, People’s Republic of China; 3Department of B-Ultrasound Room, The Affiliated Yixing Hospital of Jiangsu University, Yixing, Jiangsu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Sujun Zhou; Jianping Zhou
Department of Gastrointestinal Surgery, The Affiliated Yixing Hospital of Jiangsu University, No. 75 Tongzhenguan Road, Yixing 214200, Jiangsu, People’s Republic of China
Tel/ Fax +86-510-87921098
Email [email protected]; [email protected]
Background: Although several long non-coding RNAs (lncRNAs) have been found to be involved in gastric cancer tumorigenesis, the more comprehensive contributions of lncRNAs to gastric cancer require further investigation. Here, we identify a cytoplasmic lncRNA, LINC01485, which promotes tumor growth and migration in gastric cancer.
Materials and Methods: Microarray and computational analysis were utilized to identify differential expression in LINC01485 and EGFR. Real-time PCR and Western blotting assays were used to confirm the expression of LINC01485 and EGFR in gastric cancer cells. Cell proliferation, wound-healing and transwell assays were performed to measure cell growth, migration and invasion. Immunoprecipitation, RNA pull-down, and RNA fluorescence in situ hybridization (RNA-FISH) assays were used to test the interaction of c-Cbl with LINC01485 and EGFR. Furthermore, tumor xenograft in nude mice was performed to test tumor growth in vivo.
Results: LINC01485 was upregulated and associated with tumor size, lymphatic metastasis and advanced pathological stage in gastric cancer. LINC01485 promoted gastric cancer cell proliferation, migration and invasion in vitro and in vivo. Furthermore, LINC01485 levels were positively correlated with EGFR expression in gastric cancer tissues and significantly increased the expression and phosphorylation (Tyr1045) of EGFR in gastric cancer cells. Mechanistically, LINC01485 competes with c-Cbl for binding to phosphorylated Tyr1045 site of EGFR, thus interfering with c-Cbl-mediated ubiquitination and subsequent degradation of EGFR.
Conclusion: LINC01485 promoted EGFR stabilization and activation of EGFR/Akt signaling in gastric cancer. Our findings illustrate the diversity of cytoplasmic lncRNAs in signal transduction and highlight the important roles of lncRNAs in gastric cancer.
Keywords: gastric cancer, long noncoding RNA, LINC01485, EGFR, ubiquitination
Introduction
Gastric cancer is the 4th most common incident cancer and the 2nd most common cause of cancer death worldwide, each year about 990,000 people are diagnosed with gastric cancer, approximately 738,000 people die of this disease.1 Strikingly, gastric cancer cases in China alone account for 42% of the total number of cases worldwide.2 Nevertheless, surgery is the only means of cure for patients with gastric cancer and is the treatment of choice for early-stage disease.3 Although oncogene activation and tumor suppressor gene loss are leading causes of gastric carcinogenesis,4 the underlying molecular mechanisms of gastric carcinogenesis are largely unknown. Thus, the detection of new prognostic markers and a better comprehension of molecular mechanisms for gastric tumorigenesis are still an urgent problem.
Epidermal growth factor receptor (EGFR), a member of receptor tyrosine kinase family, is a transmembrane glycoprotein. Moreover, EGFR is involved in the development of malignancies, including gastric cancer.5 Mechanistically, EGFR contributes to oncogenesis primarily through activation of Akt pathway, thus stimulating cancer cell proliferation, survival, invasion, and drug resistance.6 Significantly, EGFR activation has been implicated in events leading to gastric cancer and the phosphorylation of EGFR was upregulated in gastric cancer.7,8 Furthermore, EGFR ubiquitination is required for routing the receptor to the lysosomal degradative compartment.9 The ubiquitin ligase Cbl (Casitas B-lineage lymphoma) can direct binding to phosphorylated EGFR (Tyr1045) and target activated EGFR for ubiquitination and subsequent endosomal sorting.10–12 EGFR inhibitor Gefitinib has been used in the treatment of various cancers, including proximal gastric cancer.13
The long noncoding RNAs (lncRNAs) are uniquely expressed in differentiated tissues or specific cancer types and functionally defined as transcripts >200 nucleotides in length with no protein-coding potential.14,15 Recently, significant attention has been paid to define the complex mechanisms underlying tumor development and progression of lncRNAs.16,17 Mechanistically, lncRNAs modulate many important cancer phenotypes by associating with other cellular macromolecules including DNA, protein, and RNA.15 For example, LncRNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer.16 LncRNA GClnc1 promotes gastric carcinogenesis and may act as a modular scaffold of WDR5 and KAT2A complexes to specify the histone modification pattern.18 However, the more comprehensive contributions of lncRNAs to gastric cancer require further investigation.
We hypothesized that there is still a significant amount of previously unexplored transcriptomic differences of lncRNAs in gastric cancer. To establish a more in-depth understanding of the interaction between protein-coding genes and lncRNAs in gastric cancer, we performed the lncRNA and mRNA microarray of three paired gastric cancer tissues and their matched normal counterparts. We discovered LINC01485 (Gene ID: 101,928,154; NR_108028.1), an EGFR–associated lncRNA, that is overexpressed in gastric cancer. Its functional association in gastric cancer progression may partly explain how EGFR exerts its oncogenic effects in gastric cancer.
Materials and Methods
Tissue Samples and Clinical Ethics
Tumors and the adjacent gastric tissues were obtained from patients with gastric cancer who underwent surgery at the Affiliated Yixing Hospital of Jiangsu University. All methods were achieved according to the guideline approved by the Ethics Committee of the Affiliated Yixing Hospital of Jiangsu University. All patients gave informed consent and written informed consent was obtained.
Microarray and Computational Analysis
For microarray detection, total RNA from each sample was quantified by the NanoDrop ND-1000. The lncRNA/mRNA crosslinked microarray was employed (Capitalbio, Beijing, China). The sample preparation and microarray hybridization were performed based on the manufacturer’s standard protocols with minor modifications. Briefly, mRNA was purified from total RNA after removal of rRNA (mRNA-ONLY™ Eukaryotic mRNA Isolation Kit, Epicentre, WI, USA). Then, each sample was amplified and transcribed into fluorescent cRNA along the entire length of the transcripts without 3ʹ bias utilizing a random priming method. The labeled RNAs were hybridized onto the Human LncRNA Array v3.0 (8 x 60K, Arraystar). After having washed the slides, the arrays were scanned by the Agilent Scanner (Agilent, CA, USA).
For the co-expression network, the system was built according to the normalized signal intensity of specific expression in gene and lncRNA. For each pair of mRNA-lncRNA, gene-gene or mRNA-mRNA, we calculate the Pearson correlation and choose the significant correlation pairs with which to construct the network. In a network analysis, degree centrality is the simplest and most important measures of a gene or lncRNA centrality within a network that determining the relative importance. Degree centrality is defined as the link numbers one node has to the other. The Purpose of Network Structure Analysis is to locate core regulatory factors (mRNA). In one network, core regulatory factors connect most adjacent mRNAs and lncRNA, and have the biggest degrees. While considering different networks, Core regulatory factors were determined by the degree differences between two class samples. Cytoscape (Gladstone Institutes, CA, USA) was applied for the presentation of predicted network.
Cell Culture and Lentiviral Infection
Human gastric cell lines were purchased from the American Type Culture Collection (ATCC, USA) and the Cell Bank of Chinese Academy of Sciences (Shanghai, China). ShRNA and overexpression lentiviruses were produced in HEK293T cells with the GenePharma Lentiviral Expression System (GenePharma, Shanghai, China). Cell lines were cultured in DMEM medium (Gibco) supplemented with 10% FBS at 37°C in an atmosphere of 5% CO2.
Real-Time PCR Assay
Total RNA was extracted by TRIzol reagent (Invitrogen), and 1 μg of total RNA was reverse-transcribed using the PrimeScript RT Reagent Kit (Perfect Real Time; Takara). The amplified transcript level of each specific gene was normalized to that of GAPDH. RT-PCR was performed using ABI Prism 7900HT (Applied Biosystems, CA, USA) according to the direction of the reagents. 2−ΔΔCT method was used to calculate the genes expression.
Western Blotting and Immunoprecipitation
Equal amounts of cell lysates were subjected to Western blotting or immunoprecipitation assays. Immunoprecipitation was performed using Pierce Co-Immunoprecipitation Kit (Thermo, 26,149) according to the manufacturer’s instructions. Cells were lysed and incubated with Protein-G coupled sepharose beads that had been incubated with antibody to EGFR at room temperature for 1 h. Immunoprecipitation was for 1 h at 4°C before washing four times with immunoprecipitation buffer. The immunoprecipitated was eluted in sample buffer, incubated at 95°C for 5 min and subjected to Western blotting.
For analysis of EGFR ubiquitination, 1 mM Methylmaleimide (NEM) was added in the immunoprecipitation buffer. Immunoprecipitated EGFR was analyzed by Western blotting with antibody to ubiquitin.
Cells protein lysates were separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE), transferred to 0.22 μm PVDF membranes (Millipore) and visualized using an ECL kit (Millipore, MA, USA). Antibodies: Cell Signaling Technology: anti-EGFR (D38B1) rabbit monoclonal antibody (4267), anti-Phospho-EGFR (Tyr1045) rabbit antibody (2237), anti-Akt (11E7) rabbit monoclonal antibody (4685), anti-phospho-Akt (Ser473) (D9E) rabbit monoclonal antibody (4060), anti-Ubiquitin (P4D1) mouse monoclonal antibody (3936), anti-c-Cbl (C49H8) rabbit monoclonal antibody (2179), anti-GAPDH (D16H11) rabbit monoclonal antibody (5174). The antibodies were used as 1:1000 dilutions for Western blotting experiments and 1:200 for immunoprecipitation.
Cell Proliferation Assay
The ability of cells proliferation was assayed using CCK-8 (Dojin Laboratories, Kumamoto, Japan) according to the manufacturer’s instructions. The cells were seeded at a density of 104 cells/well in 96-well flat-bottom and, respectively, cultured for CCK-8 assay according to the protocol provided by the manufacturer. The OD 450 nm was detected at 0, 24, 48 and 72 h.
Wound-Healing and Transwell Assay
When reaching 80~90% confluence, the cell layer was scratched. Cells were then continued cultured and the average extent of wound closure was quantified. Transwell assays were performed using 24-well transwell (8-μm pore size; Millipore) precoated with Matrigel (BD Biosciences). Cells were then harvested, and 105 cells were seeded in serum-free medium into the upper chamber, whereas medium supplemented with 20% FBS was applied to the lower chamber as a chemoattractant. After 24 hours of incubation, the migrated cells at the bottom surface of the filter were fixed with pre-cooled methanol, stained with 0.2% crystal violet solution, and counted with a microscope.
RNA Pull-Down Assay
Synthesis of LINC01485 lncRNA (wild type and deletion mutant LINC01485 (nucleotides 340–391)) sequences were purchased from Genewiz (Suzhou, China). The biotin-labeled lncRNA was transcribed with a Biotin RNA Labeling Mix (Roche, CA, USA) and the T7 RNA polymerase (Roche, CA, USA), treated with RNase-free DNase I (Roche) and purified with an RNeasy Mini Kit (Qiagen, Germany). Protein extracted from cells was mixed with biotinylated RNA. Sixty μL washed streptavidin agarose beads (Invitrogen, USA) was then added to each binding reaction and washed. The associated proteins were resolved by SDS-PAGE and analyzed by Western blotting.
RNA Fluorescence in situ Hybridization (RNA-FISH)
The cells were rinsed briefly in PBS and then fixed in 4% formaldehyde in PBS for 20 min at 25°C. Further, the cells were permeabilized in PBS containing 0.5% Triton X-100 on ice for 10 min; washed with PBS and rinsed once in 2× SSC (300 mM NaCl, 30 mM sodium citrate, pH 7.0) prior to hybridization. Hybridization was carried out using a nick-translated cDNA probe (Takara, Japan) in a moist chamber at 37°C for 16 h.19 For DAPI staining, cells were incubated in 30 nM DAPI in PBS at room temperature for 15 min.
Probes and Primers
RT-PCR primers for gene expression: LINC01485 (741bp) (5′- GAG AGA GCA CTT TCG GGC TT −3′ and 5′- CTT GCC ACG CTC ACA ATC TG −3′), EGFR (5′- AGG CAC GAG TAA CAA GCT CAC −3′ and 5′- ATG AGG ACA TAA CCA GCC ACC −3′). shRNA targeted sequences: LINC01485 shRNA1 (5′- GAG TAC TGC CAA CAA GAA G −3′), LINC01485 shRNA1 (5′- AGA GAG ATT GGG CAC AGA C −3′). RNA FISH probe: LINC01485 (/FITC/5′- GGC AGT ACT CTT TTT TCT GA −3′), U6 (/FITC/5′- TGA TCA TCT CTG TAT TGT TTC −3′).
The Subcutaneous Xenograft Models
Animal care and euthanasia were approved by the Jiangsu University animal studies committee and is based on the National Institutes of Health’s Guide to the Care and Use of Laboratory Animals. Indicated cells (105) were subcutaneously implanted into the bilateral axilla of nude mice. Tumors were measured every week after implantation, and the volume of each tumor was calculated (length × width2 × 0.5). All mice were sacrificed 7 weeks afterwards, and the xenografts were peeled off subcutaneously.
Statistics
The experiment was independently repeated three times. Results are reported as mean ± standard error of the mean (SEM) of at least three independent experiments. Comparisons were performed using two-tailed Student’s t-test or one-way ANOVA (*P < 0.05), as indicated in the individual figures. F-test was employed for statistical analyses of linear regression correlation analysis. Chi-squared test was implemented for statistical analyses of the correlation between clinical parameters. All statistical analyses were presented with GraphPad prism 5 software.
Results
Microarray Analysis of Differentially Expressed lncRNAs and mRNAs in Gastric Cancer
Firstly, we performed the lncRNA and mRNA microarray in three paired gastric cancer tissues and their matched normal counterparts. The result of hierarchical (Figure 1A and B) clustering exhibited the significantly differentially expressed lncRNAs and mRNAs, which were defined as the absolute fold change of >2 between gastric cancer tissues and the normal counterparts. The top 6 aberrant expressed mRNA has been validated in 20 paired gastric cancer tumor tissues and paratumor counterparts (Figure 1C). To confirm the microarray results, we selected eight upregulated novel candidate lncRNAs in the microarray for authentication in about 20 paired gastric cancer tumor tissues and paratumor counterparts. As shown in Figure 1D, six novel candidate lncRNAs upregulated in gastric cancer tumor tissues than the paratumor counterparts. Using WGCNA (weighted correlation network analysis), we generated correlations among these upregulated lncRNAs and mRNAs and uncovered a positive correlation between EGFR and lncRNAs MATN1-AS1, LINC01356, LINC01485, and TCEB3-AS1 (Figure 1E). In order to conform the co-expressed lncRNA, expression and correlation analysis of EGFR and 4 lncRNAs were performed in 20 paired gastric cancer specimens. Figure 1F displays that only LINC01485 expression was positively correlated with EGFR levels.
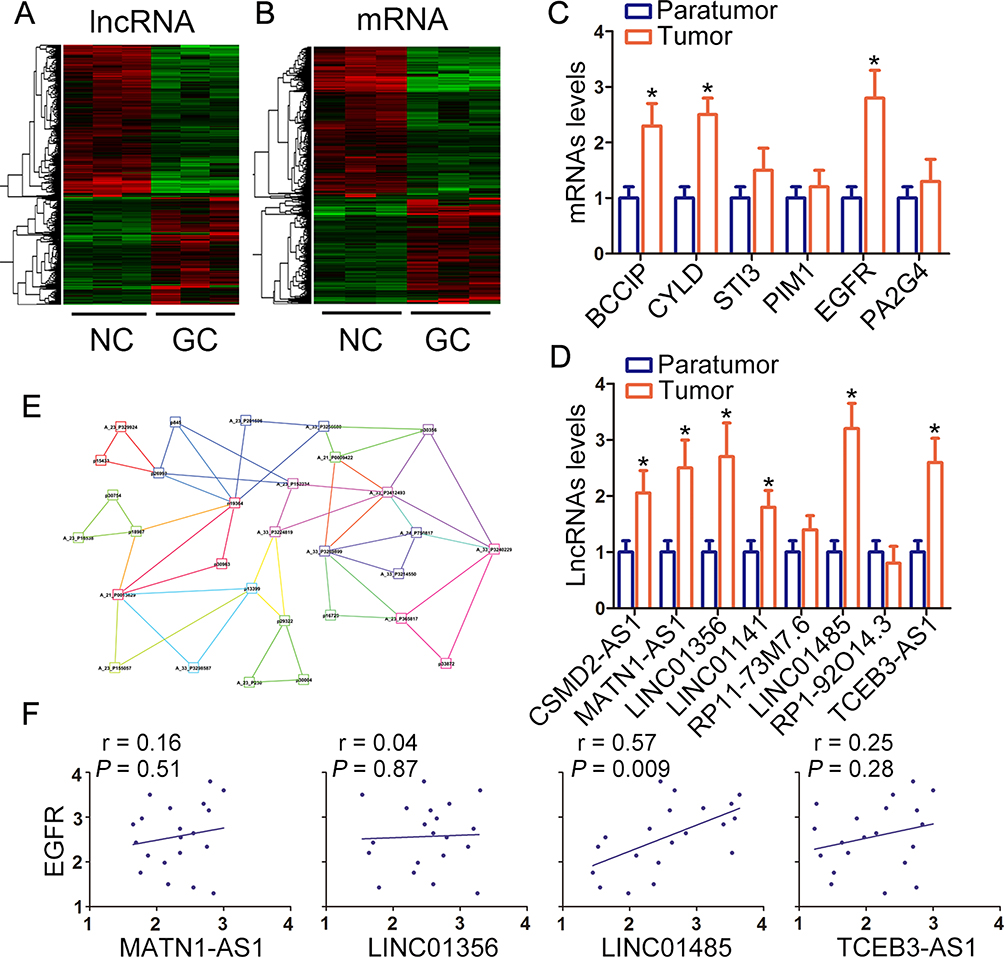 |
Figure 1 Microarray analysis of differentially expressed lncRNAs and mRNAs in gastric cancer. (A) Heat map and hierarchical clustering analyzed the distinguishable lncRNA (A) and mRNA (B) profiling in gastric cancer tumor tissues and paratumor counterparts. The relative expression from high to low level was indicated with red and blue color. (C and D) The expression levels of 6 upregulated candidate mRNAs and 8 upregulated novel candidate lncRNAs were validated in 20 paired gastric cancer tumor tissues and paratumor counterparts with RT-PCR. The data were presented as the mean ± SEM, *P < 0.05, Student’s t-test. (E) The specific gene-expression patterns were determined by weighted gene co-expression network analysis (WGCNA). (F) The correlation analysis of EGFR and 4 lncRNAs were examined in 20 gastric cancer tumor tissues with RT-PCR. Abbreviations: lncRNA, long non-coding RNA; RT-PCR, real-time polymerase chain reaction; SEM, standard error of the mean; EGFR, epithelial growth factor receptor. |
LINC01485 Level is Upregulated and Positively Correlated to EGFR Expression in Gastric Cancer
Considering that EGFR was previously implicated with stemness in gastric epithelial cells,20 we analyzed the expression patterns of LINC01485 and EGFR based on the single-cell data released by the previous study (GSE134520).21 Our analyses showed that both LINC01485 and EGFR were upregulated in cancer cell and metaplastic stem-like cell, but not in endothelial cell and smooth muscle cell (Figure 2A and B). Additionally, we proceeded to validate the expression and correlation of LINC01485 and EGFR in 61 paired gastric cancer tumor tissues and the paratumor counterparts. As shown in Figure 2C and D, both LINC01485 and EGFR expression were higher in gastric cancer tumor tissues than the paratumor counterparts. The LINC01485 levels were positively correlated with EGFR expression by correlation analysis (Figure 2E). Moreover, the upregulation of LINC01485 positively correlated with the larger tumor size, lymphatic metastasis, and TNM stage III&IV as shown in Table 1.
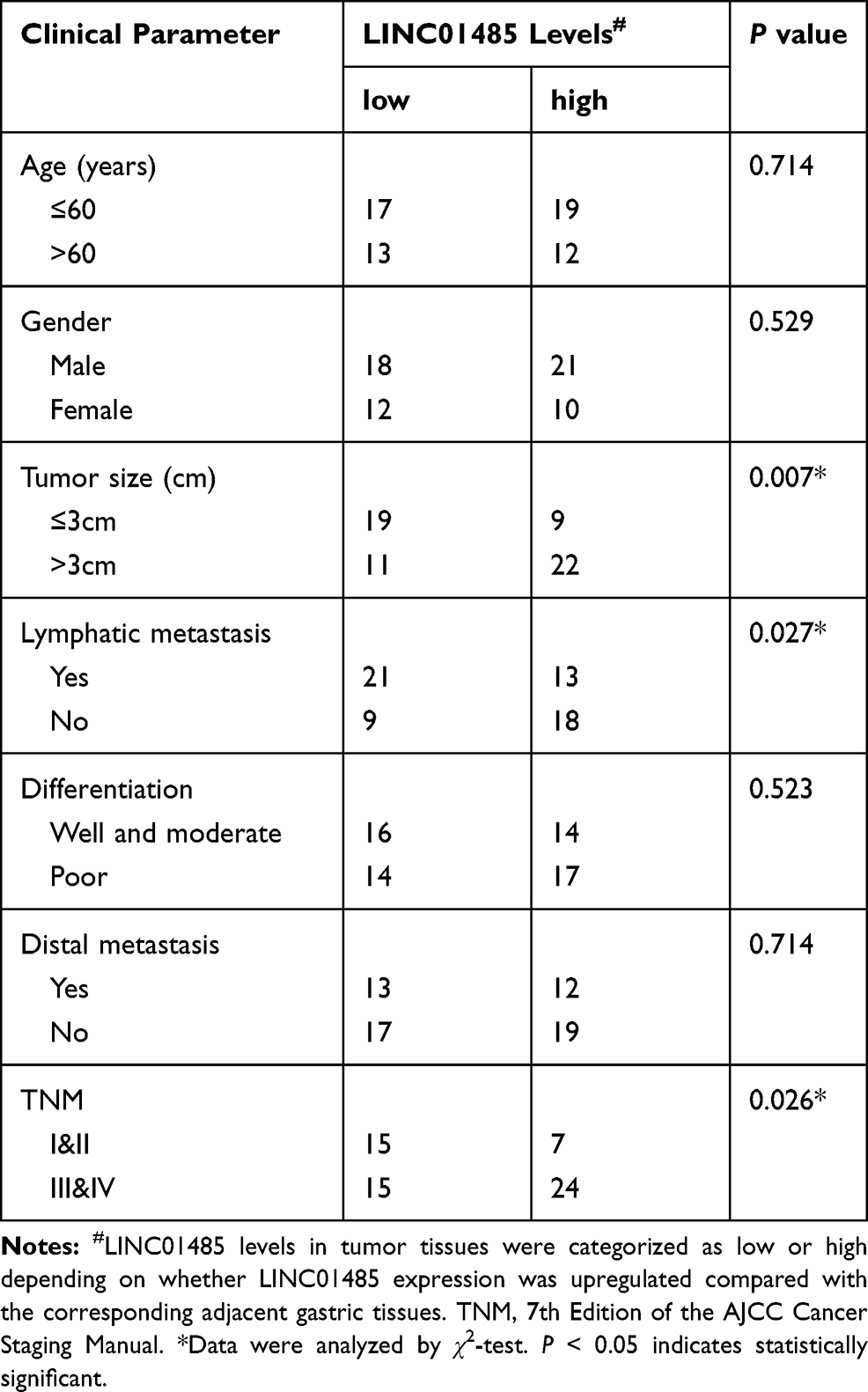 |
Table 1 Clinicopathological Characteristics and LINC01485 Expression in 61 Patient Samples of Gastric Cancer |
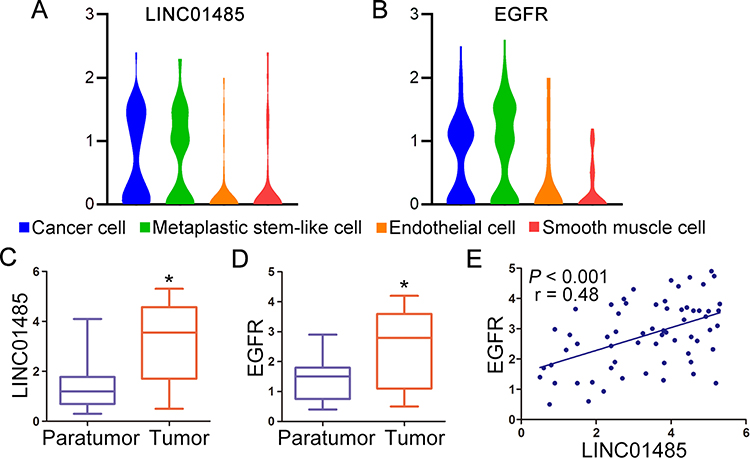 |
Figure 2 LINC01485 level is upregulated and positively correlated to EGFR expression in gastric cancer. Violin plots display the distribution of expression of LINC01485 (A) and EGFR (B) across cancer cell, metaplastic stem-like cell, endothelial cell, and smooth muscle cell. (C) The expression levels of LINC01485 and EGFR (D) were validated in 61 paired gastric cancer tumor tissues and paratumor counterparts with RT-PCR. The data were presented as the box and whisker plots, *P < 0.05, Student’s t-test. (E) The correlation analysis of EGFR and LINC01485 were observed in 61 gastric cancer tumor tissues with RT-PCR. *P < 0.05, linear regression correlation analysis (F-test). Abbreviations: RT-PCR, real-time polymerase chain reaction; EGFR, epithelial growth factor receptor. |
Knockdown of LINC01485 Expression Reduces Cell Proliferation, Migration, and Invasion in Gastric Cancer Cells in vitro
In five gastric cancer cell lines, LINC01485 expression was the highest in gastric cancer cell line NCI-N87 compared with other cells (Figure 3A). The significant upregulation of LINC01485 in gastric cancer tumor tissues was related to larger tumor size and lymphatic metastasis. Subsequently, we silenced LINC01485 in the gastric cancer cell line NCI-N87 by employing two shRNA lentivirus (Figure 3B). The influence of LINC01485 knockdown on gastric cancer cell proliferation was tested by CCK-8 assay. LINC01485 shRNA infection-mediated LINC01485 knockdown significantly reduced cell proliferation compared with negative control scrambled shRNA infection in NCI-N87 cells (Figure 3C). Furthermore, wound-healing assays revealed that LINC01485 shRNA-treated cells showed remarkedly declined cell migration rate compared with the scrambled shRNA in NCI-N87 cells (Figure 3D). The effect of LINC01485 knockdown on gastric cancer cell invasion was examined by Transwell assay. Findings showed that knockdown of LINC01485 also repressed the invasion activity of gastric cancer cell NCI-N87 (Figure 3E). These results demonstrated that LINC01485 was involved in the promotion of gastric cancer cell proliferation, migration, and invasion.
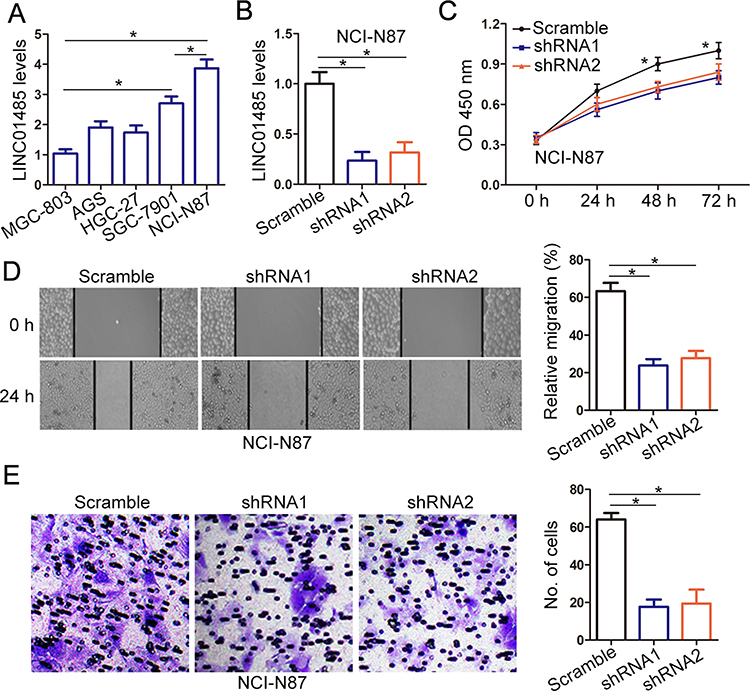 |
Figure 3 Knockdown of LINC01485 expression reduces cell proliferation, migration, and invasion in gastric cancer cells in vitro. (A) The expression levels of LINC01485 were studied in 5 gastric cancer cell lines with RT-PCR. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (B) The NCI-N87 cells were infected with LINC01485 shRNA1 and shRNA2 lentivirus, and the expression levels of LINC01485 were examined with RT-PCR. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (C) The NCI-N87 cells were infected with LINC01485 shRNA1 and shRNA2 lentivirus, and cell growth rates were determined by CCK-8 viability assay. The absorbance at 450 nm was measured at indicated times. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (D) The NCI-N87 cells were infected with LINC01485 shRNA1 and shRNA2 lentivirus, and wound-healing assay was performed to determine the migration of gastric cancer cells. The relative migration rate was quantified after scratch 24 h. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (E) The NCI-N87 cells were infected with LINC01485 shRNA1 and shRNA2 lentivirus, and transwell assay was used to determine the invasion of gastric cancer cells. The images show the number of cells that penetrated the porous membrane. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. Abbreviations: RT-PCR, real-time polymerase chain reaction; SEM, standard error of the mean; shRNA, short hairpin RNA; CCK-8, Cell Counting Kit-8; ANOVA, analysis of variance. |
LINC01485 Enhances EGFR/AKT Signaling via Inhibiting EGFR Ubiquitination
Considering that ubiquitination of the EGFR is an important intracellular signal that occurs upon EGF stimulation,22 we examined EGFR protein levels in four paired gastric cancer tissues. In these cases, EGFR protein levels were found to be higher in tumor tissue than they were in matched paratumor counterparts (Figure 4A and B). Consistent with these findings, we proposed that LINC01485 might interact with EGFR protein and regulated the ubiquitination of EGFR. Therefore, we queried the LINC01485 and EGFR sequences through catRAPID algorithm to estimate their binding propensity. This algorithm predicted that lncRNA LINC01485 (nucleotides 340–391) could bind to intracellular domain (amino acids 1001–1052) of EGFR as depicted in the schematic representation (Figure 4C). To test whether LINC01485 indeed binds EGFR, we firstly performed RNA fluorescence in situ hybridization (RNA-FISH) to evaluate the location of LINC01485 in the gastric cancer cell NCI-N87. LINC01485 transcript was mainly located to the cytoplasm in the gastric cancer cell NCI-N87 (Figure 4D). We then performed deletion mutation and in vitro RNA pulldown experiments to determine whether EGFR would associate within the predicted region (nucleotides 340–391) of LINC01485. We found that 340–391 nt region of LINC01485 is required for the association with EGFR (Figure 4E and F). Meanwhile, lentivirus-mediated ectopic expression of wild type LINC01485 significantly increased the expression and phosphorylation (Tyr1045) of EGFR in gastric cancer cell line MGC-803. However, the expression and phosphorylation of EGFR remained unaltered in the deletion mutant LINC01485 (nucleotides 340-391)-overexpressed gastric cancer cell MGC-803 compared with the vector controls (Figure 4G and H). Moreover, ectopic expression of wild type LINC01485 substantially increased phosphorylation of Akt, whereas ectopic expression of deletion mutant LINC01485 had no effect on Akt phosphorylation (Figure 4G and H). The phosphorylation of EGFR Tyr1045 site is a major docking site for c-Cbl, we next compared the EGFR ubiquitination and association between EGFR and c-Cbl in wild type and deletion mutant LINC01485-overexpressed gastric cancer cell MGC-803. The level of EGFR ubiquitination was higher in Lv-WT MGC-803 cells than in vector control cells. Consistent with the reduction in EGFR ubiquitination, the association of EGFR with c-Cbl was obviously decreased in Lv-WT MGC-803 cells than in vector control cells (Figure 4I and J). Conversely, ectopic expression of deletion mutant LINC01485 in MGC-803 cells did not decrease the EGFR ubiquitination and EGFR/c-Cbl association compared with that of vector control cells (Figure 4I and J). These findings illustrated that LINC01485 could increase the phosphorylation of EGFR and inhibit EGFR/c-Cbl association and subsequent c-Cbl-mediated EGFR ubiquitination.
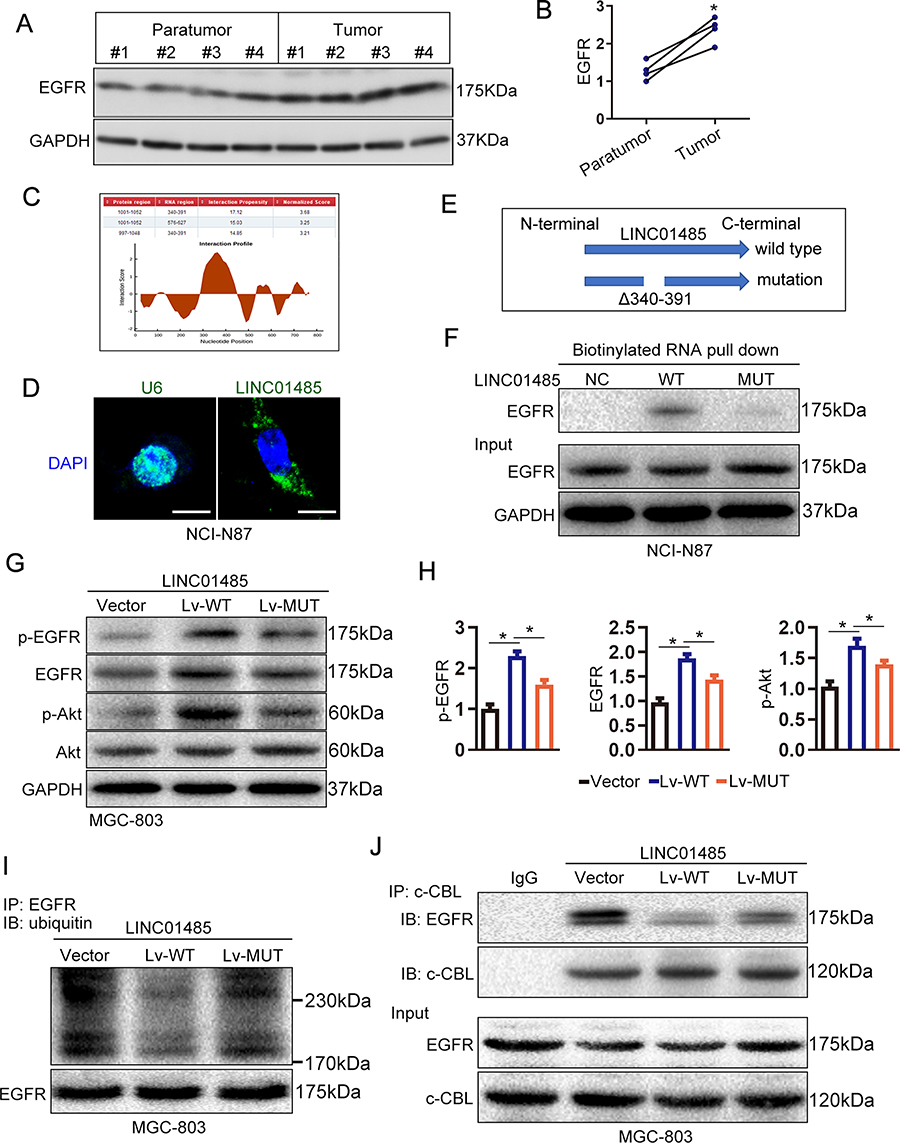 |
Figure 4 LINC01485 enhances EGFR/AKT signaling via inhibiting EGFR ubiquitination. (A) The expression of EGFR was examined by Western blotting in 4 paired gastric cancer tumor tissues and matched paratumor counterparts. (B) The quantification of blots in figure (A). The data were presented as the mean ± SEM, n = 4, *P < 0.05, Student’s t-test. (C) The catRAPID method was used to predict EGFR-LINC01485 interactions and their binding sites. (D) RNA fluorescence in situ hybridization (RNA-FISH) with LINC01485 probe (green) was employed to evaluate the location of LINC01485 in the gastric cancer cell NCI-N87. U6 snRNA probe (green) was used as normalization control. DAPI is shown as blue. Scale bar, 10 μm. (E) Diagram of wild type and deletion mutant LINC01485 (nucleotides 340–391). (F) In vitro transcribed wild type and deletion mutant (nucleotides 340–391) LINC01485 were incubated with cell lysates extracted from gastric cancer cell NCI-N87 for an in vitro RNA pull-down assay. The EGFR levels were examined by Western blotting. (G) The wild type (Lv-WT) or deletion mutant (Lv-MUT) LINC01485 lentivirus were used to construct wild type or deletion mutant LINC01485-overexpressed gastric cancer cell MGC-803. The expression and phosphorylation of EGFR and Akt were examined by Western blotting in Lv-WT MGC-803 and Lv-MUT MGC-803 cells. (H) The quantification of blots in figure (G). The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (I and J) LINC01485 inhibited EGFR ubiquitination and EGFR–c-Cbl association. EGFR immunoprecipitates (IP EGFR) or whole-cell lysates (Input) of Lv-WT MGC-803 and Lv-MUT MGC-803 cells were subjected to Western blotting with the indicated antibodies. GAPDH served as loading control. Representative results of three replicates are shown.Abbreviations: RNA-FISH, RNA fluorescence in situ hybridization; DAPI, 4ʹ,6-diamidino-2-phenylindole; EGFR, epithelial growth factor receptor; Lv, lentivirus; IP, immunoprecipitates. |
LINC01485 Exert Oncogenesis Effects is Dependent on Its Association with EGFR
To evaluate the biological impact of LINC01485/EGFR association in gastric cancer, we explored the effect of lentivirus-mediated ectopic expression of wild type (Lv-WT) or deletion mutant (Lv-MUT) LINC01485 on cell proliferation, migration, and invasion. The CCK-8 assays showed that cell growth was remarkedly improved in Lv-WT infected MGC-803 cells (Figure 3A and B), while the proliferation of MGC-803 cells did not increase in Lv-MUT infected cells compared with vector controls (Figure 5A and B). Similarly, the findings of wound-healing assays displayed that cell migration and invasion ability was increased following upregulation of wild type LINC01485 in Lv-WT MGC-803 cells, while no change in deletion mutant (Lv-MUT) LINC01485 infected MGC803 cells (Figure 5C and D). Taken together, these findings illustrated that the upregulation of LINC01485 promoted gastric cancer cell proliferation, migration, and invasion and was dependent on its association with EGFR.
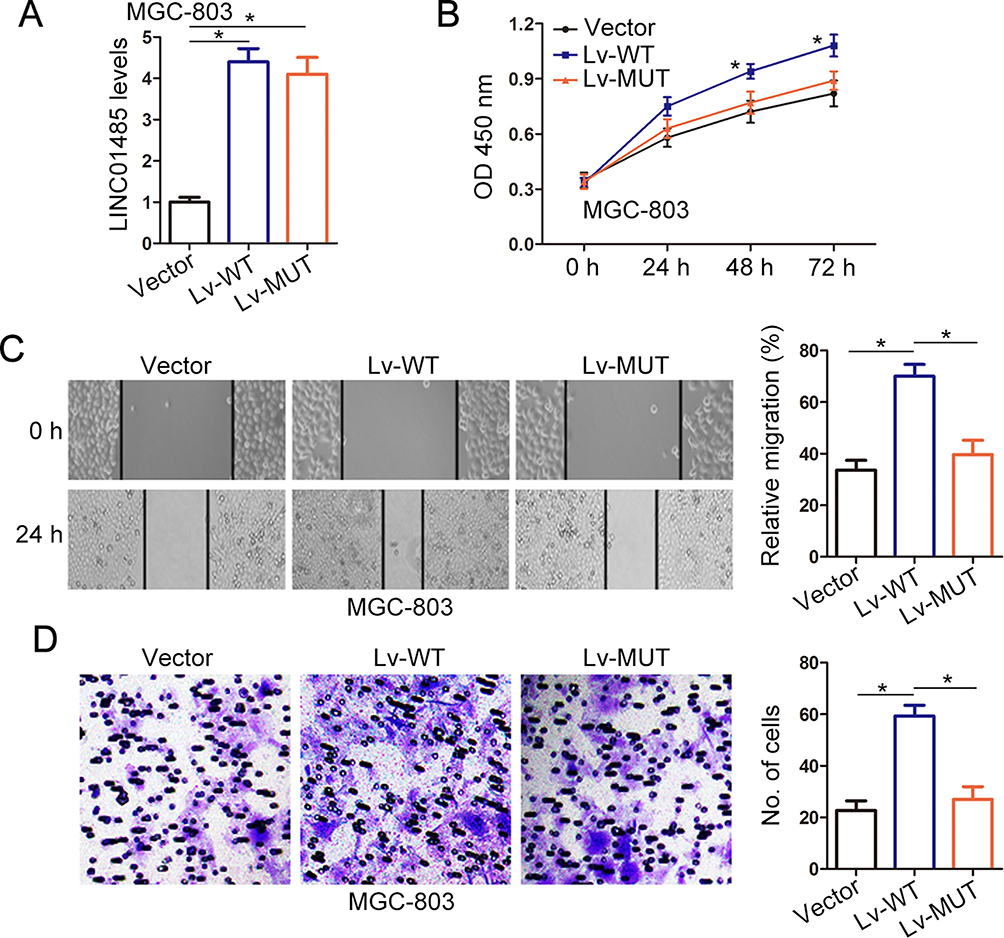 |
Figure 5 LINC01485 exert oncogenesis effects is dependent on its association with EGFR. (A) The wild type (Lv-WT) or deletion mutant (Lv-MUT) LINC01485 lentivirus were used to construct wild type or deletion mutant LINC01485-overexpressed gastric cancer cell MGC-803. The expression levels of LINC01485 were examined with RT-PCR. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (B) The cell growth rates of Lv-WT and Lv-MUT MGC-803 cells were determined by CCK-8 viability assay. The absorbance at 450 nm was measured at indicated times. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (C) The wound-healing assay was performed to determine the migration of Lv-WT and Lv-MUT MGC-803 gastric cancer cells. The relative migration rate was quantified after scratch 24 h. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. (D) The transwell assay was used to determine the invasion of Lv-WT and Lv-MUT MGC-803 gastric cancer cells. The images show the number of cells that penetrated the porous membrane. The data were presented as the mean ± SEM, n = 3, *P < 0.05, ANOVA. Abbreviations: RT-PCR, real-time polymerase chain reaction; Lv, lentivirus; EGFR, epithelial growth factor receptor; SEM, standard error of the mean; ANOVA, analysis of variance. |
LINC01485 Promotes Gastric Cancer Cells Tumor Growth in vivo
To study the effect of LINC01485 on gastric cancer cells tumor growth in vivo, we generated xenograft models by implanting LINC01485 shRNA or scramble shRNA-treated NCI-N87 cells into nude mice. Seven weeks after injection, the tumors formed in shRNA-treated NCI-N87 group were greatly smaller than those in the scramble group (Figure 6A and B). Additionally, tumor weight was also obviously reduced in the shRNA-treated NCI-N87 group compared to the scramble group (Figure 6C). Therefore, suppression of LINC01485 produced a marked decrease in the rate of xenograft subcutaneous tumor growth. To determine whether LINC01485 expression and LINC01485/EGFR association could affect tumor proliferation and invasion in vivo, wild type or deletion mutant LINC01485 ectopic expressed-MGC-803 cells were inoculated into nude mice. The tumors formed in Lv-WT MGC-803 group were markedly larger than those in the vector control group (Figure 6D and E). Additionally, tumor weight was also obviously increased in the Lv-WT MGC-803 group compared to the vector control group (Figure 6F). However, ectopic expression of deletion mutant LINC01485 failed to significantly tumor formation and weight in the Lv-MUT MGC-803 group compared to the vector control group (Figure 6D–F). These data confirmed the effect of LINC01485 on EGFR degradation and strongly support our notion that LINC01485 contributes to the malignant phenotype by interacting with EGFR.
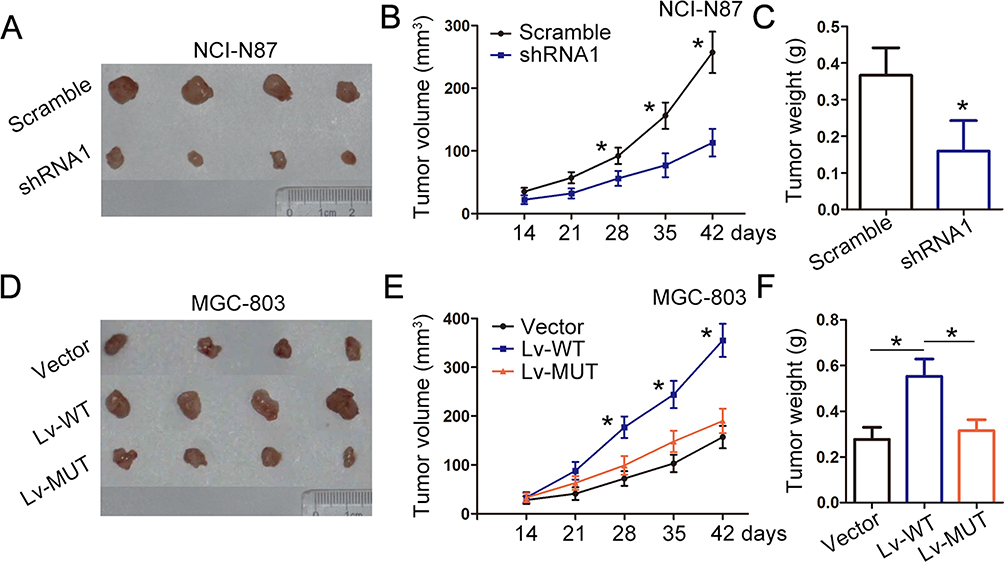 |
Figure 6 LINC01485 promotes gastric cancer cells tumor growth in vivo. (A and B) The tumor volume was calculated every week after injection of scramble or shRNA1 NCI-N87 cells (stably knockdown of LINC01485). The data were presented as the mean ± SEM, n = 6, *P < 0.05, Student’s t-test. (C) Tumor weights were represented as means of tumor weights ± SEM, n = 6, *P < 0.05, Student’s t-test. (D and E) The tumor volume was calculated every week after injection of Lv-WT, Lv- MUT, or vector control MGC-803 cells. The data were presented as the mean ± SEM, n = 6, *P < 0.05, ANOVA. (F) Tumor weights were represented as means of tumor weights ± SEM, n = 6, *P < 0.05, ANOVA. Abbreviations: Lv, lentivirus; SEM, standard error of the mean; ANOVA, analysis of variance. |
Discussion
Recent studies have discovered that lncRNAs play an important role in cancer progression. The lncRNAs HOTAIR, MEG3, MALAT1, H19, and GAPLINC have been found to be implicated in various malignancies.18 Importantly, the lncRNA PCA3 has become the first approved diagnostic biomarker in prostate cancer by FDA.23 Nevertheless, dysregulated lncRNAs and their functional mechanisms have not yet been extensively studied in gastric cancer.
In this study, we performed the transcriptomic mRNAs/lncRNAs association analysis on gastric cancer. We report here that one of the lncRNAs, LINC01485, is shown to be interacted with EGFR and has dramatic oncogenic effects on gastric cancer cells. Our data revealed that the expression of LINC01485 was upregulated and positively correlated to EGFR expression in gastric cancer. Moreover, LINC01485 upregulation correlated with larger tumor size, lymphatic metastasis and advanced pathological stage. Furthermore, LINC01485 overexpression promoted the gastric cancer cell proliferation, migration and invasion, while LINC01485 knockdown repressed cell viability and cell invasion, and induced growth arrest both in vitro and in vivo. These results illustrate that LINC01485 has an important effect on the pathophysiological phenotypes and progression of gastric cancer.
LncRNAs exert their functions through cis- and trans-regulatory mechanisms and formation of RNA–protein interactions.24 Critically, the cellular localization of lncRNA partly determines its biological functions. Cytoplasmic lncRNAs can modulate mRNA stability or translation and influence cellular signaling cascades, while nuclear lncRNAs are enriched for functionality involving chromatin interactions, transcriptional regulation, and RNA processing.15 LncRNA LINK-A enables the recruitment of BRK to the EGFR/GPNMB complex and BRK kinase activation, thus activating normoxic HIF1α signaling in triple-negative breast cancer.25 Subsequently, we observed LINC01485 subcellular localization and found that LINC01485 mainly express in both the cytoplasm and close to the cellular membrane. Therefore, we speculated that LINC01485 may exert its functions in the cytosol and cellular signaling cascades. Then, we predicted the association of LINC01485 with EGFR by using bioinformatics-based tool (catRAPID algorithm) and found the highest interaction sore (normalized) was 3.68 (location: lncRNA LINC01485 region: 340–391, HOXA1 region: 1001–1052). The LINC01485 deletion mutation and RNA pull‐down assay revealed that LINC01485 could combine with the protein EGFR, further promote the expression of EGFR, and activating its downstream Akt signaling. Akt (protein kinase B) has been found to be involved in cancer cell survival, metastasis, and chemotherapy resistance and dysregulated Akt activation is one of the most common molecular findings in human malignancies including gastric cancer.26 Based on the single-cell data released by the previous study (GSE134520),21 we found that both LINC01485 and EGFR were upregulated in cancer cell and metaplastic stem-like cell, but not in endothelial cell and smooth muscle cell. The results indicated that the regulation between LINC01485 and EGFR might occur in the cellular context, particularly in gastric epithelial stem cells.
It is well known that direct association between EGFR (pTyr1045) and c-Cbl is required to sort the EGFR to lysosomes for degradation,11,27 leading us to examine the ubiquitination of EGFR to investigate whether LINC01485/EGFR association modulates c-Cbl-mediated EGFR ubiquitination. Moreover, both EGFR ubiquitination and interaction of EGFR with c-Cbl were markedly reduced in LINC01485-overexpressed gastric cancer cells. However, the deletion mutation of LINC01485 (nucleotides 340–391) partially restored EGFR ubiquitination and the interaction between EGFR and c-Cbl. Thus, LINC01485 competes with c-Cbl for binding to phosphorylated Tyr1045 site of EGFR, thus interfering with c-Cbl-mediated ubiquitination and subsequent degradation of EGFR. Considering that lncRNAs act as ceRNAs via lncRNA-miRNA-mRNA Networks, several microRNAs were predicted to be bound to LINC01485 (Table 2). Among them, miR-30b inhibits non-small cell lung cancer cell growth by targeting the EGFR.28 Furthermore, the deletion mutation of LINC01485 also did not alter the gastric cancer cell proliferation, tumor growth, migration and invasion both in vitro and in vivo. Although our findings suggest a molecular mechanism by which LINC01485 contributes to EGFR function, further investigation of whether and how LINC01485 regulate de novo recruitment of other interacting proteins to EGFR are required to address.
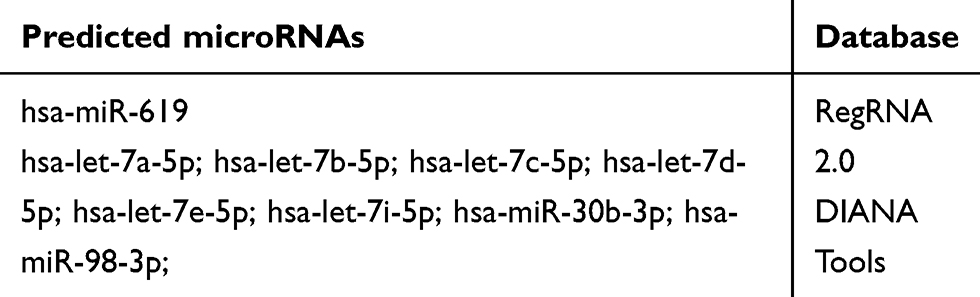 |
Table 2 The Predicted microRNAs Interacted with LINC01485 |
However, there were some limitations associated with our findings. For instance, we found that lncRNA LINC01485 was mostly overexpressed in EGFR-upregulated cancer patient samples. Since EGFR-targeted therapies eventually develop resistance in several cancers,29 it would be of great interest to study if LINC01485 is involved in EGFR resistance mechanisms.
Conclusions
Our study describes lncRNA-protein kinase module that regulates ubiquitination and degradation of EGFR regarding the functional consequences in Akt signaling activation and gastric cancer cell proliferation, migration and invasion. Our data provide new insights and an underlying mechanism for the function of lncRNA LINC01485 in gastric cancer.
Ethical Approval
All methods were achieved according to the guideline approved by the Ethics Committee of the Affiliated Yixing Hospital of Jiangsu University. All patients gave informed consent and written informed consent was obtained. All animal experiments were approved by the Committee on the Ethics of Animal Experiments of Jiangsu University and based on the National Institutes of Health’s Guide to the Care and Use of Laboratory Animals.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Karimi P, Islami F, Anandasabapathy S, et al. Gastric cancer: descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol Biomarkers Prev. 2014;23(5):700–713. doi:10.1158/1055-9965.EPI-13-1057
2. Zhou Z, Ji Z, Wang Y, et al. TRIM59 is up-regulated in gastric tumors, promoting ubiquitination and degradation of p53. Gastroenterology. 2014;147(5):1043–1054. doi:10.1053/j.gastro.2014.07.021
3. Lordick F, Allum W, Carneiro F, et al. Unmet needs and challenges in gastric cancer: the way forward. Cancer Treat Rev. 2014;40(6):692–700. doi:10.1016/j.ctrv.2014.03.002
4. Choi Y, Park J, Ko YS, et al. c-Jun N-terminal kinase activation has a prognostic implication and is negatively associated with FOXO1 activation in gastric cancer. BMC Gastroenterol. 2016;16(1):59. doi:10.1186/s12876-016-0473-9
5. Sierra JC, Asim M, Verriere TG, et al. Epidermal growth factor receptor inhibition downregulates Helicobacter pylori-induced epithelial inflammatory responses, DNA damage and gastric carcinogenesis. Gut. 2018;67(7):1247–1260. doi:10.1136/gutjnl-2016-312888
6. Feng H, Lopez GY, Kim CK, et al. EGFR phosphorylation of DCBLD2 recruits TRAF6 and stimulates AKT-promoted tumorigenesis. J Clin Invest. 2014;124(9):3741–3756. doi:10.1172/JCI73093
7. Stahl P, Seeschaaf C, Lebok P, et al. Heterogeneity of amplification of HER2, EGFR, CCND1 and MYC in gastric cancer. BMC Gastroenterol. 2015;15:7. doi:10.1186/s12876-015-0231-4
8. Chaturvedi R, Asim M, Piazuelo MB, et al. Activation of EGFR and ERBB2 by Helicobacter pylori results in survival of gastric epithelial cells with DNA damage. Gastroenterology. 2014;146(7):1739–1751.e1714. doi:10.1053/j.gastro.2014.02.005
9. Tomas A, Futter CE, Eden ER. EGF receptor trafficking: consequences for signaling and cancer. Trends Cell Biol. 2014;24(1):26–34. doi:10.1016/j.tcb.2013.11.002
10. Grovdal LM, Stang E, Sorkin A, et al. Direct interaction of Cbl with pTyr 1045 of the EGF receptor (EGFR) is required to sort the EGFR to lysosomes for degradation. Exp Cell Res. 2004;300(2):388–395. doi:10.1016/j.yexcr.2004.07.003
11. Yao S, Zheng P, Wu H, et al. Erbin interacts with c-Cbl and promotes tumourigenesis and tumour growth in colorectal cancer by preventing c-Cbl-mediated ubiquitination and down-regulation of EGFR. J Pathol. 2015;236(1):65–77. doi:10.1002/path.4502
12. Li X, Lu Y, Liang K, et al. Brk/PTK6 sustains activated EGFR signaling through inhibiting EGFR degradation and transactivating EGFR. Oncogene. 2012;31(40):4372–4383. doi:10.1038/onc.2011.608
13. Soria JC, Wu YL, Nakagawa K, et al. Gefitinib plus chemotherapy versus placebo plus chemotherapy in EGFR-mutation-positive non-small-cell lung cancer after progression on first-line gefitinib (IMPRESS): a Phase 3 randomised trial. Lancet Oncol. 2015;16(8):990–998. doi:10.1016/S1470-2045(15)00121-7
14. Iyer MK, Niknafs YS, Malik R, et al. The landscape of long noncoding RNAs in the human transcriptome. Nat Genet. 2015;47(3):199–208. doi:10.1038/ng.3192
15. Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways. Cancer Cell. 2016;29(4):452–463. doi:10.1016/j.ccell.2016.03.010
16. Liu XH, Sun M, Nie FQ, et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer. Mol Cancer. 2014;13:92. doi:10.1186/1476-4598-13-92
17. Hu Y, Wang J, Qian J, et al. Long noncoding RNA GAPLINC regulates CD44-dependent cell invasiveness and associates with poor prognosis of gastric cancer. Cancer Res. 2014;74(23):6890–6902. doi:10.1158/0008-5472.CAN-14-0686
18. Sun TT, He J, Liang Q, et al. LncRNA GClnc1 promotes gastric carcinogenesis and may act as a modular scaffold of WDR5 and KAT2A complexes to specify the histone modification pattern. Cancer Discov. 2016;6(7):784–801. doi:10.1158/2159-8290.CD-15-0921
19. Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. 2010;39(6):925–938. doi:10.1016/j.molcel.2010.08.011
20. Wang C, Guo X, Xi R. EGFR and Notch signaling respectively regulate proliferative activity and multiple cell lineage differentiation of Drosophila gastric stem cells. Cell Res. 2014;24(5):610–627. doi:10.1038/cr.2014.27
21. Zhang P, Yang M, Zhang Y, et al. Dissecting the single-cell transcriptome network underlying gastric premalignant lesions and early gastric cancer. Cell Rep. 2019;27(6):1934–1947.e1935. doi:10.1016/j.celrep.2019.04.052
22. Pinilla-Macua I, Grassart A, Duvvuri U, et al. EGF receptor signaling, phosphorylation, ubiquitylation and endocytosis in tumors in vivo. Elife. 2017;6. doi:10.7554/eLife.31993
23. Zhao J, Liu Y, Zhang W, et al. Long non-coding RNA Linc00152 is involved in cell cycle arrest, apoptosis, epithelial to mesenchymal transition, cell migration and invasion in gastric cancer. Cell Cycle. 2015;14(19):3112–3123. doi:10.1080/15384101.2015.1078034
24. Guttman M, Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature. 2012;482(7385):339–346. doi:10.1038/nature10887
25. Lin A, Li C, Xing Z, et al. The LINK-A lncRNA activates normoxic HIF1alpha signalling in triple-negative breast cancer. Nat Cell Biol. 2016;18(2):213–224. doi:10.1038/ncb3295
26. Almhanna K, Strosberg J, Malafa M. Targeting AKT protein kinase in gastric cancer. Anticancer Res. 2011;31(12):4387–4392.
27. Shrestha N, Shrestha H, Ryu T, et al. delta-Catenin increases the stability of EGFR by decreasing c-Cbl interaction and enhances EGFR/Erk1/2 signaling in prostate cancer. Mol Cells. 2018;41(4):320–330. doi:10.14348/molcells.2018.2292
28. Qi Z, Zhang B, Zhang J, et al. MicroRNA-30b inhibits non-small cell lung cancer cell growth by targeting the epidermal growth factor receptor. Neoplasma. 2018;65(2):192–200. doi:10.4149/neo_2018_170217N118
29. Chong CR, Janne PA. The quest to overcome resistance to EGFR-targeted therapies in cancer. Nat Med. 2013;19(11):1389–1400. doi:10.1038/nm.3388
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.