Back to Journals » Cancer Management and Research » Volume 13
Long Non-Coding RNA TRIM52-AS1 Promotes Growth and Metastasis via miR-218-5p/ROBO1 in Hepatocellular Carcinoma
Authors Liu Y, Wu Y, Liu S, Dai Y
Received 13 October 2020
Accepted for publication 26 December 2020
Published 22 January 2021 Volume 2021:13 Pages 547—558
DOI https://doi.org/10.2147/CMAR.S286205
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xueqiong Zhu
Yuanjun Liu,* Yakun Wu,* Shuang Liu, Yi Dai
Department of Hepatobiliary Surgery, Suining Central Hospital, Suining 629000, Sichuan Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yi Dai
Department of Hepatobiliary Surgery, Suining Central Hospital, No. 127 Desheng West Road, Chuanshan District, Suining 629000, Sichuan Province, People’s Republic of China
Email [email protected]
Background: Hepatocellular carcinoma (HCC) is a malignant disease with a high mortality among primary HCC patients worldwide. Lots of studies have shown that lncRNAs are known as the biomarkers in diagnosis, treatment and prognosis of hepatocellular carcinoma. Therefore, clarifying the detailed function and mechanism of the lncRNA in the HCC progressing seems particularly important.
Methods: The TCGA and GEO database and RT-qPCR were used to analyse the expression of TRIM52-AS1 in HCC tissues and cell lines. Clinical data were collected to further analyze the correlation between indicators of clinical samples and the expression of TRIM52-AS1. CCK-8, plate clone and transwell assays were employed to evaluate the role of TRIM52-AS1 on cell proliferation, migration and invasion. Then, bioinformatics prediction, luciferase reporter, RNA immunoprecipitation (RIP), and RT-qPCR were employed to analyze the direct interaction among TRIM52-AS1, miR-218-5p and ROBO1. Additionally, the rescue function assays were used to verify that miR-218-5p/ROBO1 was the function downstream of TRIM52-AS1.
Results: TRIM52-AS1 was overexpressed in HCC according to the TCGA database and RT-qPCR assay. The expression of TRIM52-AS1 was higher in the metastatic foci compared with primary tumor according to the GEO database. Additionally, TRIM52-AS1 knockdown inhibited the proliferation and metastasis of HCC cells. TRIM52-AS1 could act as competitive endogenous RNA to regulate ROBO1 through miR-218-5p, then promoted the HCC cell progression.
Conclusion: TRIM52-AS1 is overexpressed in HCC and can promote the proliferation and metastasis of HCC cells through miR-218-5p/ROBO1 axis, then drives the HCC cell progression.
Keywords: TRIM52-AS1, ROBO1, miR-218-5p, hepatocellular carcinoma, growth, metastasis
Background
Hepatocellular carcinoma (HCC) is a malignant disease with a high mortality among primary HCC patients worldwide. It is one of the most common malignant tumors globally, and HCC is the sixth most common cancer in the world recently.1 The HCC mortality rate remains high mainly due to the lack of early diagnosis, high recurrence rate and high metastasis rate.2 The treatment of hepatocellular carcinoma has always been a difficult problem. Transplantation is the most effective method for the treatment of liver cancer. However, during the process of transplantation, the tumor recurrence rate and metastasis rate are relatively high.3 Therefore, we must accelerate the research on the molecular mechanism of HCC occurrence and progression, which can promote the effective treatment of HCC. In recent years, more and more studies have shown that lncRNAs are closely related to the diagnosis, treatment and prognosis of hepatocellular carcinoma.4–6
LncRNAs are a new class of transcriptional substances and important gene expression regulators.7 In addition, the biological functions of some lncRNAs in human cancers such as liver cancer, stomach cancer and lung cancer have increased exponentially.8–10 For example, the abnormal expression of MALAT1 is related to tumor metastasis and various cancers such as breast cancer, lung cancer, prostate cancer and liver cancer. Its overexpression can be regarded as an early prognostic indicator, indicating a low survival rate.11–14 Using lncRNA microarray, lncRNA HULC and HEIH were also found to be abnormally expressed in liver cancer tissues and to be involved in the occurrence of liver cancer.15 In addition, it was reported that several lncRNAs including lncRNA-p21 were involved in cell processes such as cell growth and apoptosis.16 Liu et al studies had shown that lncCSMD1-1 was upregulated in liver cancer and promoted the progression of liver cancer by activating the MYC signaling pathway. The underlying mechanism was that lncCSMD1-1 could directly bind to MYC protein in the nucleus of HCC cells, leading to an increase in MYC protein. At the same time, lncCSMD1-1 interacted with MYC protein to block its ubiquitin-proteasome degradation pathway, thereby activating its downstream target gene.17 Research by Xu et al found that the expression of lncRNA H19 was negatively correlated with sorafenib sensitivity in HCC cells. Inhibition of lncRNA H19 could increase the sensitivity of sorafenib by inhibiting the epithelial-mesenchymal transition (EMT) of HCC cells.18 Wu et al proved that the up-regulation of lncRNA FTX inhibited the conversion of non-alcoholic fatty liver disease (NAFLD) to hepatocellular carcinoma by promoting M1 polarization of Kupffer cells.19 Therefore, lncRNAs can be used as novel prognostic markers and potential therapeutic targets for HCC.
Studies have found that lncRNA TRIM52-AS1 is differentially expressed in cancer. Liu et al found that lncRNA TRIM52-AS1 was down-regulated and significantly inhibited cell proliferation and metastasis in renal cell carcinoma (RCC). Therefore, lncRNA TRIM52-AS1 played a role in the genesis and development of RCC.20 Currently, lncRNA TRIM52-AS1 related research is rarely reported in HCC, this study aimed to screen differentially expressed lncRNA through TCGA databases, and validated in the organization of patients with HCC. Additionally, cell function experiments were performed to verify the effects of TRIM52-AS1 on the proliferation, migration, invasion and invasion of HCC cells. To further explore the possible molecular mechanisms of TRIM52-AS1 promoting HCC progress, it might provide us more novel early diagnosis and specific therapeutic basis for HCC.
Methods
Patient and Clinical Samples
All the 42 pairs of HCC cancer samples and corresponding adjacent tissues were from patients at Suining Central Hospital (Suining, Suining, China). Specimens were frozen in liquid nitrogen immediately after resection and stored at Liquid nitrogen tank. These tissue samples were examined by pathological testing. All patients provided written informed consent and the Ethics Committee of Suining Central Hospital approved the current study. All procedures were aligned with the Declaration of Helsinki.
Cell Culture
The L02, Hep3B, Bel-7402, Huh7 cell lines were purchased from the American Type Culture Collection (USA) and Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). The cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Gibco, NY, USA) supplemented with 100 μg/mL streptomycin, 100 U/mL penicillin and 10% fetal bovine serum (FBS) at 37°C in a humidified atmosphere of 5% CO2.
RNA Extraction and PCR Assays
The HCC tumor and adjacent tissues were stored in liquid nitrogen immediately after collection and the cells were collected with TRIzol. Then the tissues and cell total RNA isolation were according to the TRIzol reagent (Invitrogen). Total RNA was reverse transcribed into cDNA using a RNA reverse transcription kit (Takara, Dalian, China). Then the RT-qPCR was performed using the Bio-Rad CFX96 Real-Time PCR with SYBR qPCR Mix kit (Takara, Dalian, China). The 2−ΔΔCt method was used to calculate the relative gene expression normalized by GAPDH and U6. The lncRNA and mRNA were normalized with GAPDH, and the miRNA was normalized with U6.
RNA Immunoprecipitation (RIP)
The RNA immunoprecipitation (RIP) used in the experiment was performed with the RNA-Binding Protein Immunoprecipitation Kit (Millipore, MA). The AGO2 antibody was used for RIP assay and the Co- immunoprecipitation was detected by real-time PCR. The IgG Co- immunoprecipitation was used as the negative control of AGO2 immunoprecipitation output.
Cell Proliferation Assay
CCK8 Assay
A cell proliferation assay was performed with the Cell Counting Kit-8 (CCK8) assay (DOJINDO, Japanese). The Huh7 and Hep3B cells were seed in the 96-well plate at the same concentration of 3000 cells per well transfected with si-TRIM52-AS1, miR-218-5p inhibitor and ROBO1 overexpression vector. Then the indicated cells were incubated at 37°C and then measured at 450 nm every 24h.
Colony Formation Assay
For the colony formation assay, Huh7 and Hep3B cells transfected with si-TRIM52-AS1, miR-218-5p inhibitor and ROBO1 overexpression vector were seeded in each well of a six-well plate and cultured in the appropriate medium containing 10% FBS. After 14 days, the colonies were fixed with paraformaldehyde and stained with 0.2% crystal violet. The clone number was determined by counting the stained colonies.
Transwell Migration and Invasion Assay
The transfected cells were collected and resuspended in serum-free medium. Then, 2×105 cells in 200ul serum-free medium were seeded in the upper chamber of the 24 well plate. After 24 h incubation, the migration cells adhered to the lower membrane surface. Then the lower membrane was fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. And for the invasion assay, the upper chamber was coated with a pre-packed Matrigel (BD Bioscience, USA). The Cells were then photographed and counted under an optical microscope.
Luciferase Reporter Assay
The luciferase reporter vector combined with wild type (WT) TRIM52-AS1-3ʹuntranslated region (UTR) and mutant type TRIM52-AS1 sequence with mutant (MUT) of miR-218-5p binding site were purchased from Guangzhou Ribobio Biology Company (Guangzhou, China). Renilla luciferase plasmid pRL-TK was used as the positive control. The luciferase plasmid and miRNA mimic were co-transfected into the HCC cells. The indicated cells were seeded into a 6-well plate after 24h. The cells were lysed and the luciferase activity was detected for firefly and renilla luciferases, the renilla luciferase activity was act as the control.
Statistical Analysis
In cell function experiments were repeated three times, giving reproducible results. Data are presented as mean values ± standard deviation (SD) of three independent experiments. The correlation between TRIM52-AS1 expression and clinical pathological features was assessed via Fisher’s exact test or the wilcoxon rank sum test. The correlation of the expressions of TRIM52-AS1 and miR-218-5p or miR-218-5p and ROBO1 was established by Pearson correlation coefficient. All statistical analyses were performed using SPSS software, version 20.0 (SPSS Inc., USA) and GraphPad Prism, version 8.00 (GraphPad Software, USA).
Results
TRIM52-AS1 is Overexpressed in Hepatocellular Carcinoma and Correlated with Lymph Node Metastasis
To explore the express level of TRIM52-AS1 in TCGA-LIHC database, the results illustrated that TRIM52-AS1 was overexpressed in the HCC tissue than the adjacent normal tissues (Figure 1A). Further, the HCC specimen was used to verify the expression of TRIM52-AS1, RT-qPCR assay indicated that TRIM52-AS1 was upregulated in the HCC tissues (Figure 1B). The RT-qPCR was used to determine the expression of TRIM52-AS1 in normal hepatocyte cell line (L02) and HCC cell lines (Hep3B, Bel-7402, Huh7), the result demonstrated TRIM52-AS1 was significantly elevated compared with that in L02 (Figure 1C). In addition, we have further compared the contents of corresponding components in metastasis and cancer tissues in the GEO database (GSE63124), the result indicated that the expression of TRIM52-AS1 was higher in the metastatic foci compared with primary tumor (Figure 1D). To further analyze the correlation between indicators of clinical samples and the expression of TRIM52-AS1, we collated the relevant clinical data and the results showed that high expression of TRIM52-AS1 was significantly correlated with lymph node metastasis in HCC. However, there were no significant differences between other indicators of clinical samples (such as TNM stage, distant metastasis, HBV virus infection or cirrhosis) and TRIM52-AS1 expression (Table 1).
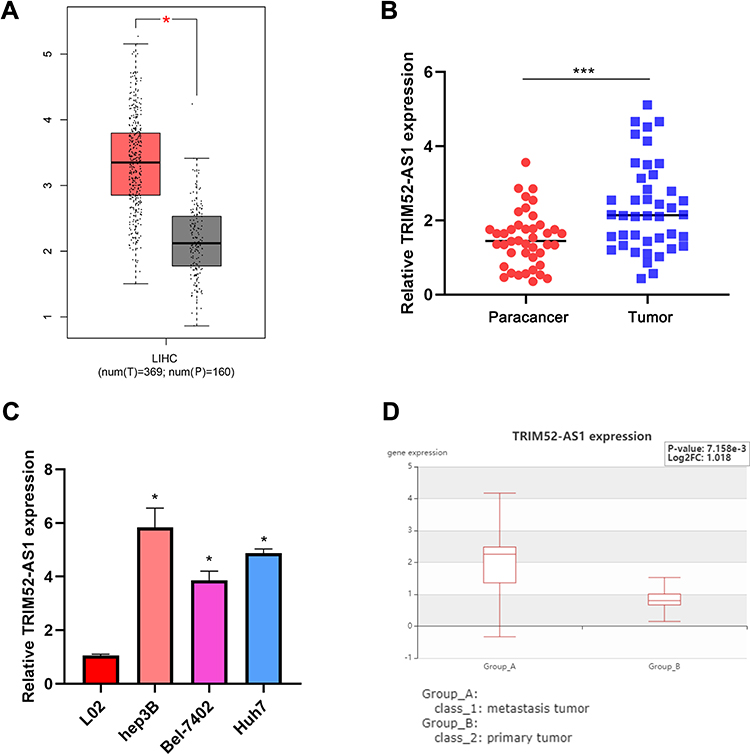 |
Figure 1 TRIM52-AS1 is overexpressed in hepatocellular carcinoma. (A) Bioinformation analysis of the expression level of TRIM52-AS1 in TCGA-LIHC database. (B) qRT-PCR analysis of relative expression level of TRIM52-AS1 in HCC tissue and adjacent normal tissues. (C) qRT-PCR analysis of relative expression levels of TRIM52-AS1 in HCC cell lines and normal liver cells. (D) Bioinformation analysis of the expression level of TRIM52-AS1 between metastasis and primary tumor in GEO database (GSE63124) (*P < 0.05, ***P < 0.001). |
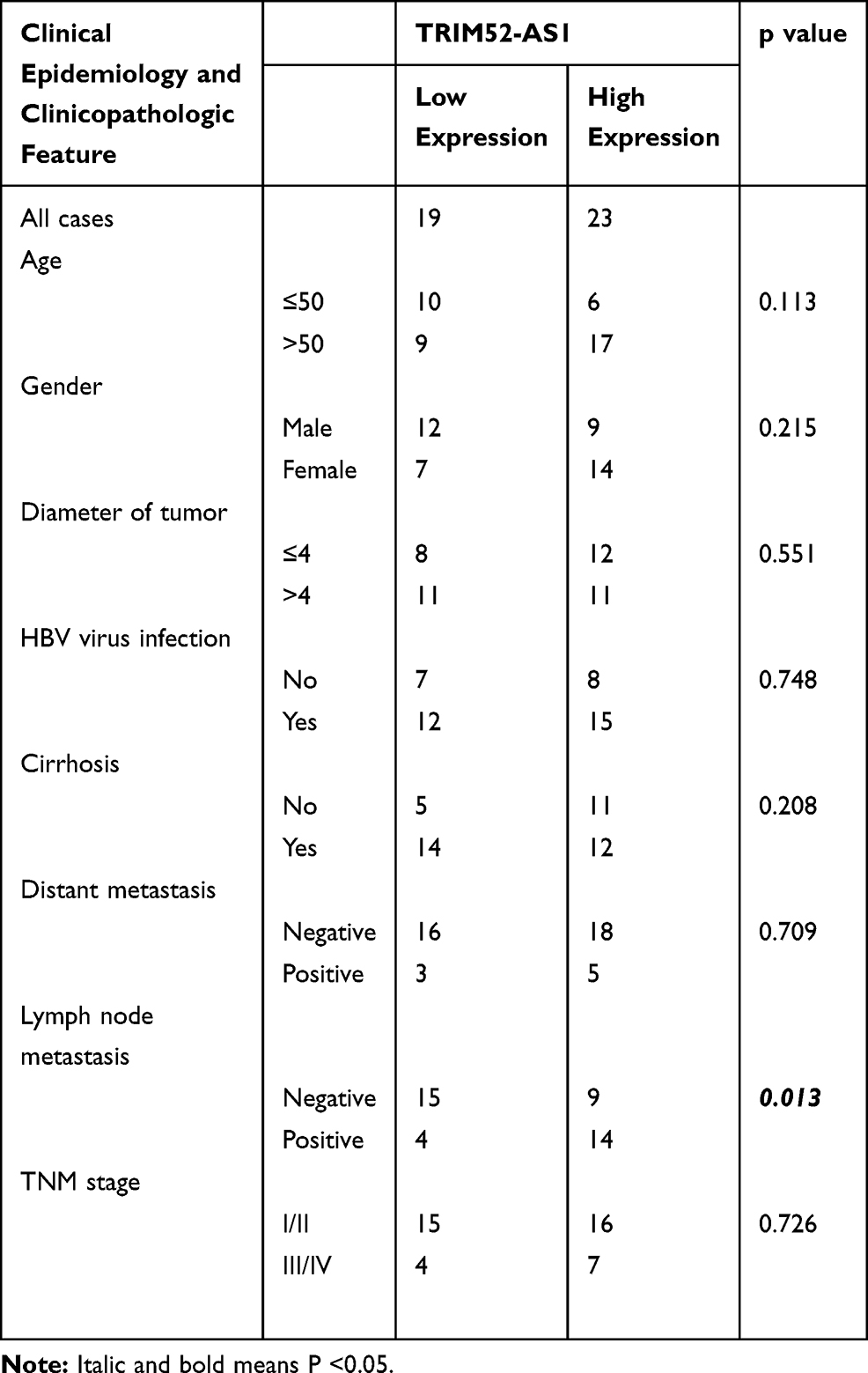 |
Table 1 General Clinicopathological Characteristics of Patients |
TRIM52-AS1 Knockdown Inhibits the Proliferation and Metastasis of HCC Cells
As the TRIM52-AS1 is upregulated in HCC tissue and cell lines, TRIM52-AS1 probably plays an essential role in proliferation and metastasis in HCC. Firstly, we transfected the target siRNA of TMEM52-AS1 in the Huh-7 and Hep3B cells. RT-qPCR verified the transfection efficiency of TRIM52-AS1 siRNA, the results demonstrated that TRIM52-AS1 was significantly downregulated in the TRIMM52-AS1 siRNA groups (si-TRIM52-AS1) (Figure 2A). CCK8 and plate clone assays were performed to evaluate the proliferation ability of Black control (BC), si-NC and si-TRIM52-AS1 groups, the results indicated that si-TRIM52-AS1 group exerted significantly decreased growth ability in Huh7 and He3B cells (Figure 2B–E). Further, the transwell experiment was used to detect the migration and invasion in the Huh7 and He3B cells, the result indicated that si-TRIM52-AS1 group showed lower migratory and invasive activities compared with NC group (Figure 2F–I).
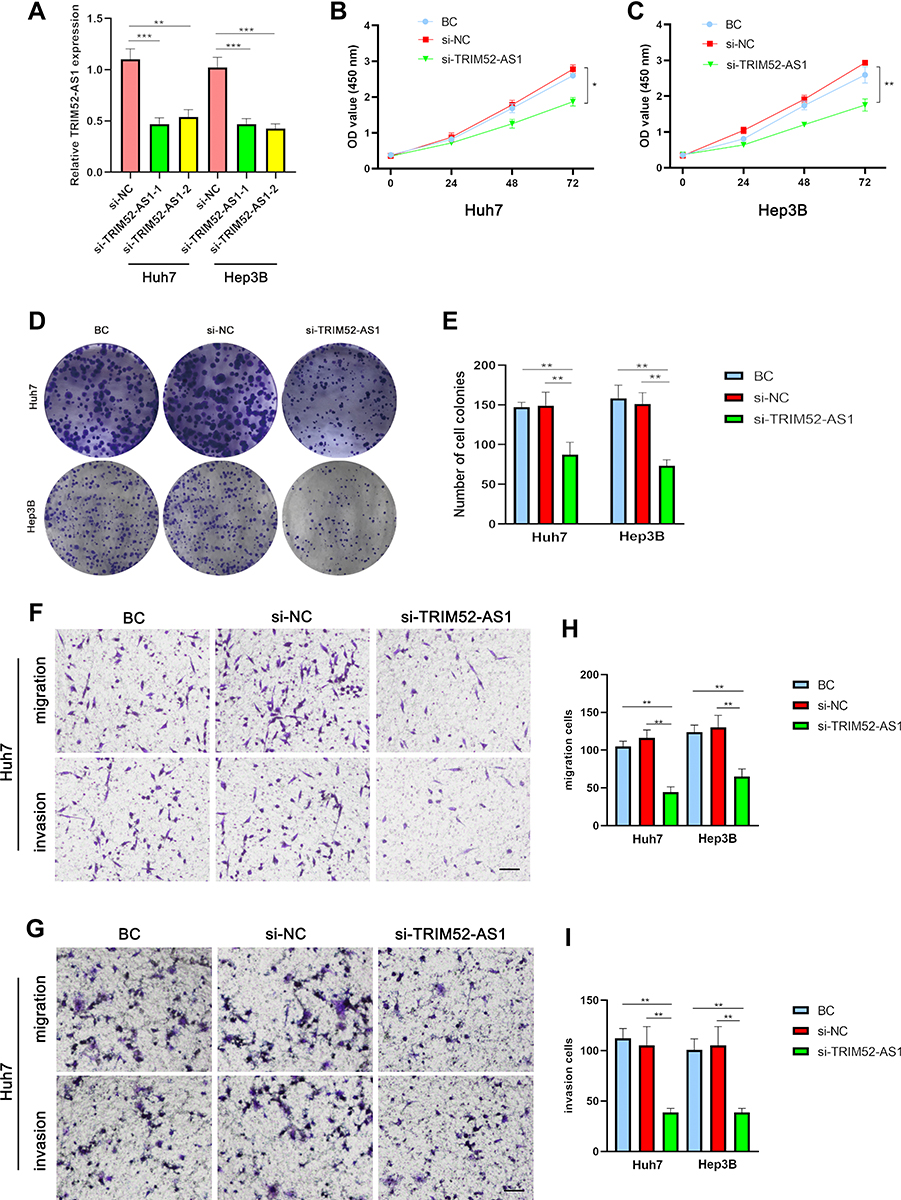 |
Figure 2 TRIM52-AS1 promotes cell proliferation of PC cells in vitro. (A) qRT-PCR analysis showing TRIM52-AS1 expression after transfection of si-RNA of TRIM52-AS1 (TRIM52-AS1-1/TRIM52-AS1-2) and negative control (NC) in Huh-7 and Hep3B cells. (B and C) CCK-8 assay analysis of the cell growth of TRIM52-AS1 knockdown, NC and BC group in Huh-7 and Hep3B cells. (D and E) Plate clone formation assays analysis of the cell growth of TRIM52-AS1 knockdown, NC and black control (BC) group in Huh-7 and Hep3B cells. (F–I) Transwell assays analysis the migration and invasion ability of TRIM52-AS1 knockdown, NC and BC group in Huh-7 and Hep3B cells (bar: 50 μm). The data are shown as means ± S.D. of three independent assays (*P < 0.05, **P < 0.01, ***P < 0.001). |
TRIM52-AS1 Activity is Directly and Partially Negatively Regulated by miR-218-5p
Further, we thus aimed to determine whether TRIM52-AS1 affect post-transcriptionally regulation with miRNA to regulate the HCC progress. According to the bioinformation prediction, the result indicated that miR-218 probably was the host miRNA of TRIM52-AS1 (Figure 3A). Further, the luciferase reporter assay demonstrated that miR-218-5p could be binding with TIMR52-AS1 (Figure 3B and C). And the RIP assay demonstrated that miR-218-5p and TRIM52-AS1 were combined in the protein of AGO2 (Figure 3D). Then, the RT-qPCR was used to evaluate the expression of miR-218-5p in the HCC normal and tumor tissues, the result indicated that miR-218-5p was downregulated in the HCC tissues (Figure 3E). RT-qPCR assay analysis results showed that miR-218-5p expression was negatively correlated with TRIM52-AS1 expression (Figure 3F). Further, the RT-qPCR assay indicated that miR-218-5p was significantly upregulated in the TRIM52-AS1 knockdown HCC cells (Figure 3G).
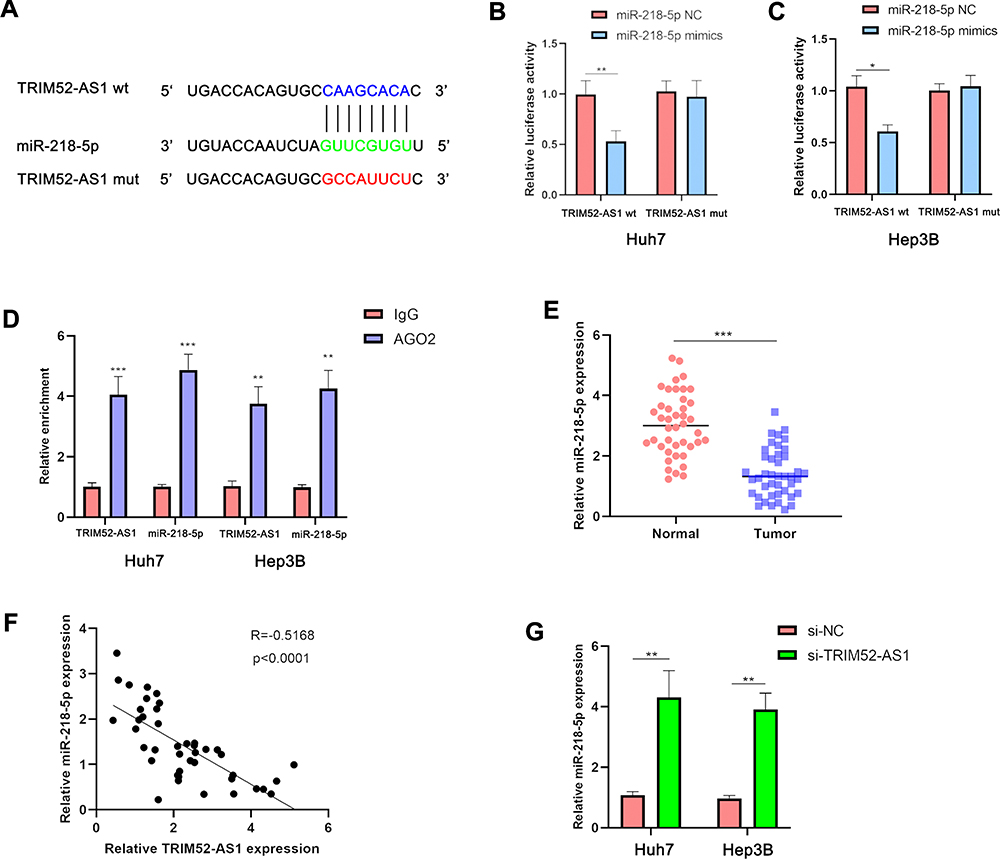 |
Figure 3 TRIM52-AS1 is directly and partially negatively regulated by miR-218-5p. (A) Bioinformation prediction of the potential binding site of miR-218-5p and TRIM52-AS1. (B and C) Luciferase reporter experiment analysis of the interaction with miR-218-5p and TRIM52-AS1. (D) RIP assay detected the combine of miR-218-5p and TRIM52-AS1 in the AGO protein. (E) qPCR assay analysis the expression of miR-218-5p in the HCC tissue and adjacent tissues. (F) The correlation of miR-218-5p and TRIM52-AS1 expression in the HCC tissues. (G) qPCR assay analysis the miR-218-5p expression of TRIM52-AS1 knockdown and NC group in Huh-7 and Hep3B cells (*P < 0.05, **P < 0.01, ***P < 0.001). |
ROBO1 is a Target Gene of miR-218-5p and Regulated by TRIM52-AS1
Base on the miRNA target gene prediction website (Taegetscan, Pic Tar, miRDB, miRTarbase), the result indicated that the ROBO1 and BMI1 were target genes of miR-218-5p in the four prediction softs (Figure 4A). The TCGA-LIHC database indicated that ROBO1 was upregulated in the liver cancer tissues, and BMI1 expression was not different in the database (Figure 4B). Then the survival analysis demonstrated that ROBO1 was negative correlated with the HCC patient prognosis (Figure 4C). According to the prediction, miR-218-5p probably binding with the 5ʹ UTR of the ROBO1 (Figure 4D). Further, the luciferase reporter assay eluted that miR-218-5p could directly be binding with the 5ʹ UTR of the ROBO1 (Figure 4E and F). The RT-qPCR assay was used to detect the ROBO1 expression in the HCC tissues, the result eluted that ROBO1 was overexpressed in the HCC tissues (Figure 4G). To clarify the miR-218-5p, ROBO1 and TRIM52RT-qPCR assay analysis results showed that miR-137 expression-AS1 expression regulation relationship, we used the RT-qPCR to evaluate the ROBO1 expression in the HCC cells, the results indicated that ROBO1 was downregulated in the miR-218-5p overexpressed and TRIM52-AS1 knockdown group (Figure 4H and K), was negatively correlated with miR-218-5p and positively correlated with TRIM52-AS1 expression (Figure 4I and J). Then, the Western blot assay demonstrated ROBO1 expression was downregulated in the TRIM52-AS1 knockdown group (Figure 4L).
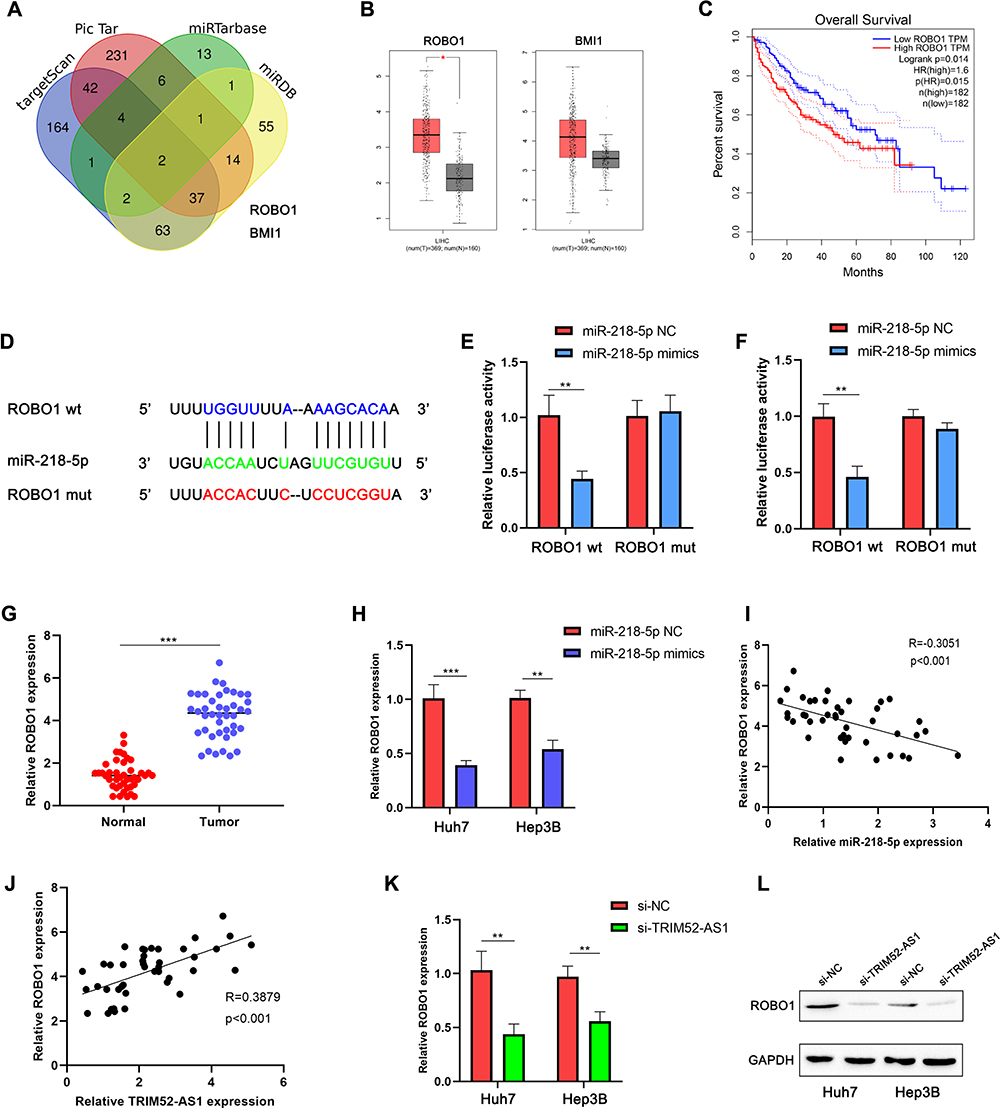 |
Figure 4 ROBO1 is a function target of miR-218-5p and regulated by TRIM52-AS1. (A) Bioinformation prediction of the potential target genes of miR-218-5p by venn analysis according to 4 miRNA databases (TargetScan, Pic Tar, miRTarbase, miRDB). (B) Bioinformation analysis of the expression of the potential target genes (ROBO1, BMI1) in the TCGA-LIHC database. (C) K-M Survival analysis the ROBO1 expression on HCC patients in TCGA-LIHC database. (D) Bioinformation prediction of the potential binding site target gene ROBO1 with the miR-218-5p. (E and F) Luciferase reporter assay analysis that the interaction with miR-218-5p and ROBO1. (G) qPCR assay analysis the expression of ROBO1 in the HCC tissues and adjacent tissues. (H) qPCR assay analysis the ROBO1 expression of miR-218-5p mimic and NC group in Huh-7 and Hep3B cells. (I) Pearson correlation coefficient show the correlation of miR-218-5p and ROBO1 expression in the HCC tissues. (J) Pearson correlation coefficient show the correlation of TRIM52-AS1 and ROBO1 expression in the HCC tissues. (K) qPCR assay analysis the ROBO1 expression of TRIM52-AS1 knockdown and NC group in Huh-7 and Hep3B cells. (L) Western-blot assay analysis the ROBO1 expression of TRIM52-AS1 knockdown and NC group in Huh-7 and Hep3B cells (*P < 0.05, **P < 0.01, ***P < 0.001). |
TRIM52-AS1 Promotes HCC Cancer Progression Through Regulating miR-218-5p/ROBO1
As the ROBO1 was found as a target of miR-218-5p and regulated by TRIM52-AS1, we further explore the regulation of TRIM52-AS1 dependent regulation of ROBO1 on the HCC cell proliferation and metastasis. Firstly, we divided the HCC cells as four groups, NC (negative group), TRIM52-AS1 knockdown, TRIM52-AS1 knockdown transfected with miR-218-5p inhibitors and TRIM52-AS1 knockdown transfected with ROBO1 overexpress vector. Then, the CCK8 and plate clone assay indicated that TRIM52-AS1 knockdown significantly inhibits the proliferation of HCC cells, and miR-218-5p inhibitor and ROBO1 overexpress vector could partly reverse the effect of TRIM52-AS1 knockdown (Figure 5A–C). Further, the transwell was used to evaluate the migration and invasion ability of the four group HCC cells, the results TRIM52-AS1 knockdown significantly inhibit the migration and invasion ability of HCC cells, and miR-218-5p inhibitor and ROBO1 overexpress vector could partly reverse the metastasis inhibited by TRIM52-AS1 knockdown (Figure 5D–G).
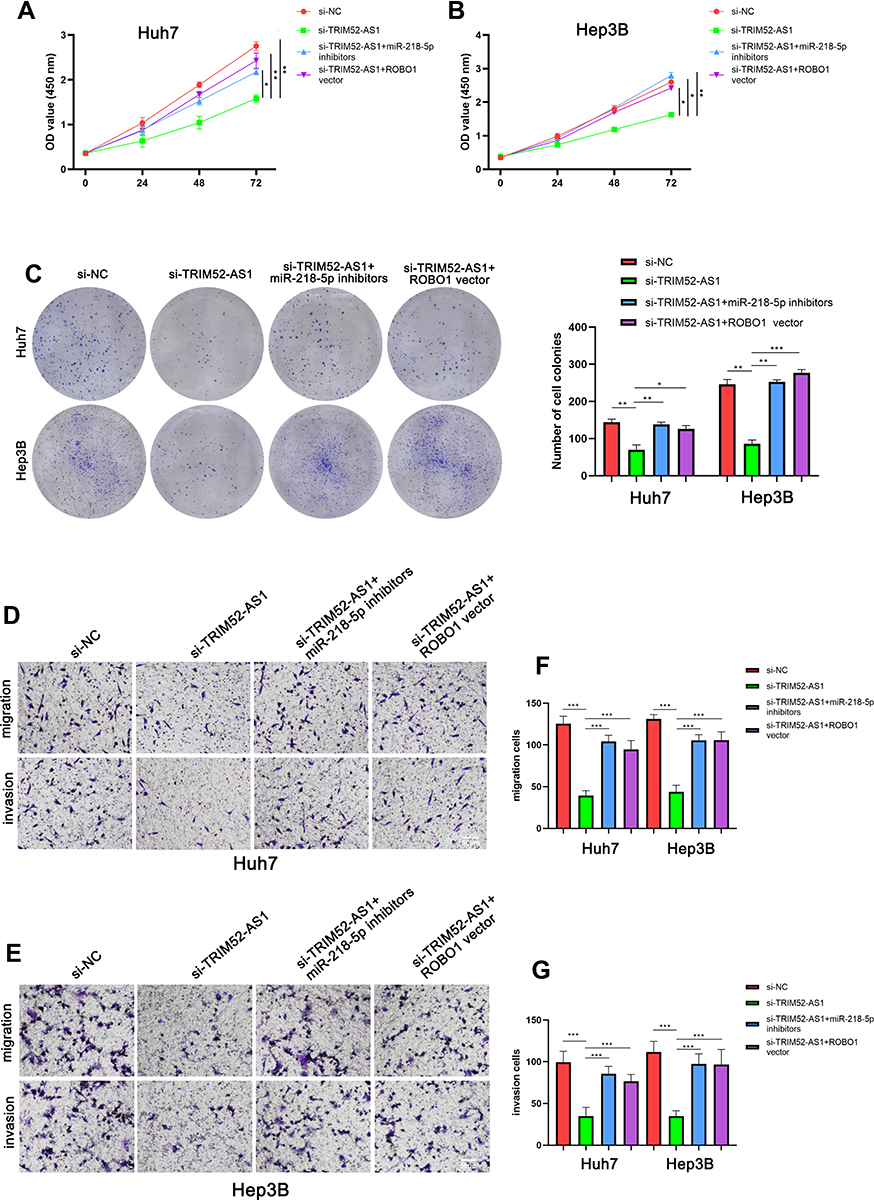 |
Figure 5 TRIM52-AS1 promotes HCC proliferation and metastasis through ROBO1 by sponging miR-218-5p (A and B) CCK8 assay analysis of the growth of TRIM52-AS1 knockdown, TRIM52-AS1 knockdown with miR-218-5p knockdown, TRIM52-AS1 knockdown with ROBO1 overexpressed and NC group. (C) Plate clone formation assays analysis of the growth of TRIM52-AS1 knockdown, TRIM52-AS1 knockdown with miR-218-5p knockdown, TRIM52-AS1 knockdown with ROBO1 overexpressed and NC group. (D–G) Transwell assays analysis of the migration and invasion of TRIM52-AS1 knockdown, TRIM52-AS1 knockdown with miR-218-5p knockdown, TRIM52-AS1 knockdown with ROBO1 overexpressed and NC group (bar: 50 μm) (*P < 0.05, **P < 0.01, ***P < 0.001). |
Discussion
Hepatocellular carcinoma (HCC) is the most common malignancy of the digestive tract and accounts for nearly 90% of primary liver cancer.21 The occurrence and development of HCC is an extremely complex biological process, which is known as the result of abnormal molecular expression or mutation.22 In the past few decades, many scientists have studied the molecular mechanism of the onset and progression of liver cancer, but the underlying mechanism remains unclear.
Previous research focuses on abnormally expressed protein involving in the tumor progress. New molecular targets including noncoding RNA may provide new perspectives for better exploration of hepatocellular carcinoma. Now, more and more evidences show that lncRNAs are involved in the process of gene expression regulation and play a very critical role in the HCC malignant progression.23–25 The expression of lncRNAs is abnormal in a variety of cancers, such as HOTAIR in breast cancer, BACE1-AS in gastric, MEG3 in liver cancer.26–28 There are also related lncRNA reports in hepatocellular carcinoma, such as H19, HEIH, MVIH, HULC and MEG3.29–33 The differential expression of these lncRNAs was also known as the tumor driver in other cancer diseases.34 Therefore, understanding the expression profile of lncRNA in liver cancer and adjacent tissues is a very critical step for revealing the pathogenesis of liver cancer.
The main work of this paper is to analyze the expression profiles of lncRNA using the TCGA-LIHC dataset. Based on the different lncRNA expression profile, we found that TRIM52-AS1 was significantly upregulated in the HCC tissues, and then the result was verified in the HCC cell lines. As TRIM52-AS1 was overexpressed in the HCC tissues and cell lines, we further detected the biological role in HCC cell proliferation and metastasis ability. The cell function assay illustrated that TRIM52-AS1 knockdown HCC cells exerted lower growth and metastasis ability. However, how TRIM52-AS1 regulates the proliferation and metastasis of hepatocellular carcinoma is still unclear.
Recent studies have shown that lncRNA has the function of regulating gene expression on multiple levels including pre-transcriptional and post-transcriptional regulation.35 In addition to the above-mentioned mechanism, TRIM52-AS1 can also regulate target gene expression through the “molecular sponge” mechanism when it functions. “Molecular sponge” is that lncRNA competitively binds to microRNA based on base complementary pairing, thereby inhibiting the binding of microRNA to its target gene, thereby up-regulating the expression of the microRNA target gene and making it function.36 In previous study, Zhou et al found that TRIM52-AS1 could sponge miR-514a-5p to promote HCC progression through increasing MRPS18A.37 And they firstly discussed the effect of lncRNA TRIM52-AS1 on proliferation and migration in HCC cells. The underlying mechanism was that TRIM52-AS1 upregulated MRPS18A expression by competitive binding miR-514a-5p. However, we tried to explore a new molecular mechanism to elucidate how TRIM52-AS1 affects the progression of HCC. Compared with Chunhui Zhou’s study, they did not discuss the correlation between indicators of clinical samples and TRIM52-AS1 expression in HCC, while we found that high expression of TRIM52-AS1 was significantly correlated with lymph node metastasis in HCC, which was our another novel point in our study. Through preliminary screening and comparison with previous reports, we finally selected miR-218-5p as the target of lncRNA TRIM52-AS1. In this study, it was found that the expression of miR-218-5p was significantly decreased and was negatively regulated by TRIM52-AS1 in the HCC cell lines Huh7 and Hep3B. Then, the luciferase reporter and RIP assay experiment verified that TIRM52-AS1 could competitively bind with the miR-218-5p, which met the conditions for its “molecular sponge” mechanism to regulate gene expression. Further, we explored to analyse the function target gene of miR-218-5p, ROBO1 was selected as the downstream function target regulated by TRIM52-AS1/miR-218-5p. Then, we found ROBO1 was upregulated in HCC tissues and as a tumor driver in the HCC cell growth and metastasis. And the ROBO1 and miR-218-5p could reverse the effect of TRIM52-AS1 knockdown in HCC cells. ROBO1 has been reported many times to play a key regulatory role in the occurrence and development of HCC, which was known as the target of miR-218-5p or miR-490-5p and triggered the Rho signal pathway.38–40 Therefore, the result indicated that TRIM52-AS1/miR-218-5p/ROBO1 was an effective pathway to regulate the HCC progression, and the ROBO1 probably was an oncogene which has not been studied clearly.
Ethics Approval
All patients provided written informed consent and the Ethics Committee of Suining Central Hospital approved the current study. All procedures were aligned with the Declaration of Helsinki.
Acknowledgment
This study was supported by Sichuan Health and Family Planning Commission project (NO.17ZD020)
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no financial or non-financial conflicts of interest for this work.
References
1. Tang A, Hallouch O, Chernyak V, Kamaya A, Sirlin CB. Epidemiology of hepatocellular carcinoma: target population for surveillance and diagnosis. Abdom Radiol. 2018;43(1):13–25. doi:10.1007/s00261-017-1209-1
2. Cao MQ, You AB, Zhu XD, et al. miR-182-5p promotes hepatocellular carcinoma progression by repressing FOXO3a. J Hematol Oncol. 2018;11(1):12. doi:10.1186/s13045-018-0555-y
3. Sarveazad A, Agah S, Babahajian A, Amini N, Bahardoust M. Predictors of 5 year survival rate in hepatocellular carcinoma patients. J Res Med Sci. 2019;24:86. doi:10.4103/jrms.JRMS_1017_18
4. Pan W, Li W, Zhao J, et al. lncRNA-PDPK2P promotes hepatocellular carcinoma progression through the PDK1/AKT/Caspase 3 pathway. Mol Oncol. 2019;13(10):2246–2258. doi:10.1002/1878-0261.12553
5. Ni W, Zhang Y, Zhan Z, et al. A novel lncRNA uc.134 represses hepatocellular carcinoma progression by inhibiting CUL4A-mediated ubiquitination of LATS1. J Hematol Oncol. 2019;10(1):91. doi:10.1186/s13045-017-0449-4
6. Wang Y, Yang L, Chen T, et al. A novel lncRNA MCM3AP-AS1 promotes the growth of hepatocellular carcinoma by targeting miR-194-5p/FOXA1 axis. Mol Cancer. 2019;18(1):28. doi:10.1186/s12943-019-0957-7
7. Su K, Zhao Q, Bian A, Wang C, Cai Y, Zhang Y. A novel positive feedback regulation between long noncoding RNA UICC and IL-6/STAT3 signaling promotes cervical cancer progression. Am J Cancer Res. 2018;8(7):1176–1189.
8. Wang H, Huo X, Yang XR, et al. STAT3-mediated upregulation of lncRNA HOXD-AS1 as a ceRNA facilitates liver cancer metastasis by regulating SOX4. Mol Cancer. 2017;16(1):136. doi:10.1186/s12943-017-0680-1
9. Zhao J, Du P, Cui P, et al. LncRNA PVT1 promotes angiogenesis via activating the STAT3/VEGFA axis in gastric cancer. Oncogene. 2018;37(30):4094–4109. doi:10.1038/s41388-018-0250-z
10. Loewen G, Jayawickramarajah J, Zhuo Y, Shan B. Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol. 2014;7:90. doi:10.1186/s13045-014-0090-4
11. Arun G, Spector DL. MALAT1 long non-coding RNA and breast cancer. RNA Biol. 2019;16(6):860–863. doi:10.1080/15476286.2019.1592072
12. Feng C, Zhao Y, Li Y, Zhang T, Ma Y, Liu Y. LncRNA MALAT1 promotes lung cancer proliferation and gefitinib resistance by acting as a miR-200a sponge. Arch Bronconeumol. 2019;55(12):627–633. doi:10.1016/j.arbres.2019.03.026
13. Chang J, Xu W, Du X, Hou J. MALAT1 silencing suppresses prostate cancer progression by upregulating miR-1 and downregulating KRAS. Onco Targets Ther. 2018;11:3461–3473. doi:10.2147/OTT.S164131
14. Malakar P, Stein I, Saragovi A, et al. Long noncoding RNA MALAT1 regulates cancer glucose metabolism by enhancing mTOR-mediated translation of TCF7L2. Cancer Res. 2018;79(10):2480–2493. doi:10.1158/0008-5472.CAN-18-1432
15. Xin X, Wu M, Meng Q, et al. Long noncoding RNA HULC accelerates liver cancer by inhibiting PTEN via autophagy cooperation to miR15a. Mol Cancer. 2018;17(1):94. doi:10.1186/s12943-018-0843-8
16. Wang T, Liu J, Li S, Yuan Z, Huang X. Effects of knockout of lincRNA-p21 on the proliferation, migration and invasion ability of HepG2 liver cancer cells. Oncol Lett. 2019;17(6):5103–5107. doi:10.3892/ol.2019.10201
17. Liu J, Xu R, Mai SJ, et al. LncRNA CSMD1-1 promotes the progression of hepatocellular carcinoma by activating MYC signaling. Theranostics. 2020;10(17):7527–7544. doi:10.7150/thno.45989
18. Xu Y, Liu Y, Li Z, et al. Long noncoding RNA H19 is involved in sorafenib resistance in hepatocellular carcinoma by upregulating miR675. Oncol Rep. 2020;44(1):165–173. doi:10.3892/or.2020.7608
19. Wu H, Zhong Z, Wang A, et al. LncRNA FTX represses the progression of non-alcoholic fatty liver disease to hepatocellular carcinoma via regulating the M1/M2 polarization of Kupffer cells. Cancer Cell Int. 2020;20:266. doi:10.1186/s12935-020-01354-0
20. Liu Z, Yan HY, Xia SY, Zhang C, Xiu YC. Downregulation of long non-coding RNA TRIM52-AS1 functions as a tumor suppressor in renal cell carcinoma. Mol Med Rep. 2016;13(4):3206–3212. doi:10.3892/mmr.2016.4908
21. Llovet JM, Zucman-Rossi J, Pikarsky E, et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2016;2:16018.
22. Brown AL, Li M, Goncearenco A, Panchenko AR. Finding driver mutations in cancer: elucidating the role of background mutational processes. PLoS Comput Biol. 2019;15(4):e1006981. doi:10.1371/journal.pcbi.1006981
23. Lin Z, Ni X, Dai S, et al. Screening and verification of long noncoding RNA promoter methylation sites in hepatocellular carcinoma. Cancer Cell Int. 2020;20:311. doi:10.1186/s12935-020-01407-4
24. Wang C, Chen Y, Chen K, Zhang L. Long noncoding RNA LINC01134 promotes hepatocellular carcinoma metastasis via activating AKT1S1 and NF-kappaB signaling. Front Cell Dev Biol. 2020;8:429. doi:10.3389/fcell.2020.00429
25. Li Y, Li G, Chen X, et al. A novel lncRNA NONHSAT053785 acts as an independent risk factor for intrahepatic metastasis of hepatocellular carcinoma. Onco Targets Ther. 2020;13:5455–5466. doi:10.2147/OTT.S254455
26. Wang Y, Gong G, Xu J, Zhang Y, Wu S, Wang S. Long noncoding RNA HOTAIR promotes breast cancer development by targeting ZEB1 via sponging miR-601. Cancer Cell Int. 2020;20:320. doi:10.1186/s12935-020-01410-9
27. Esfandi F, Ghafouri-Fard S, Oskooei VK, Taheri M. Beta-secretase 1 and its naturally occurring anti-sense RNA are down-regulated in gastric cancer. Pathol Oncol Res. 2019;25(4):1627–1633. doi:10.1007/s12253-019-00621-4
28. Liu Z, Chen JY, Zhong Y, Xie L, Li JS. lncRNA MEG3 inhibits the growth of hepatocellular carcinoma cells by sponging miR-9-5p to upregulate SOX11. Braz J Med Biol Res. 2019;52(10):e8631. doi:10.1590/1414-431x20198631
29. Li L, Han T, Liu K, Lei CG, Wang ZC, Shi GJ. LncRNA H19 promotes the development of hepatitis B related hepatocellular carcinoma through regulating microRNA-22 via EMT pathway. Eur Rev Med Pharmacol Sci. 2019;23(12):5392–5401. doi:10.26355/eurrev_201906_18208
30. Wu MM, Shen WD, Zou CW, Chen HJ, Guo HM. LncRNA-HEIH suppresses hepatocellular carcinoma cell growth and metastasis by up-regulating miR-199a-3p. Eur Rev Med Pharmacol Sci. 2020;24(11):6031–6038. doi:10.26355/eurrev_202006_21497
31. Zhang Y, Lin S, Yang X, Zhang X. Prognostic and clinicopathological significance of lncRNA MVIH in cancer patients. J Cancer. 2019;10(6):1503–1510. doi:10.7150/jca.28541
32. Ghafouri-Fard S, Esmaeili M, Taheri M, Samsami M. Highly upregulated in liver cancer (HULC): an update on its role in carcinogenesis. J Cell Physiol. 2019;10(6):1503–1510.
33. Sun Y, Cao FL, Qu LL, Wang ZM, Liu XY. MEG3 promotes liver cancer by activating PI3K/AKT pathway through regulating AP1G1. Eur Rev Med Pharmacol Sci. 2019;23(4):1459–1467. doi:10.26355/eurrev_201902_17103
34. Sanchez Calle A, Kawamura Y, Yamamoto Y, Takeshita F, Ochiya T. Emerging roles of long non-coding RNA in cancer. Cancer Sci. 2018;109(7):2093–2100. doi:10.1111/cas.13642
35. Parikshak NN, Swarup V, Belgard TG, et al. Genome-wide changes in lncRNA, splicing, and regional gene expression patterns in autism. Nature. 2016;540(7633):423–427. doi:10.1038/nature20612
36. Qi X, Zhang DH, Wu N, Xiao JH, Wang X, Ma W. ceRNA in cancer: possible functions and clinical implications. J Med Genet. 2015;52(10):710–718. doi:10.1136/jmedgenet-2015-103334
37. Zhou CH, Chen Z, Peng CL, et al. TRIM52-AS1 long noncoding RNA sponges to facilitate hepatocellular carcinoma progression through increasing. Cancer Biother Radiopharm. 2020. doi:10.1089/cbr.2019.3271
38. Wang J, Zhou Y, Fei X, et al. Integrative bioinformatics analysis identifies ROBO1 as a potential therapeutic target modified by miR-218 in hepatocellular carcinoma. Oncotarget. 2017;8(37):61327–61337. doi:10.18632/oncotarget.18099
39. Chen W, Ye L, Wen D, Chen F. MiR-490-5p inhibits hepatocellular carcinoma cell proliferation, migration and invasion by directly regulating ROBO1. Pathol Oncol Res. 2019;25(1):1–9. doi:10.1007/s12253-017-0305-4
40. Ao JY, Chai ZT, Zhang YY, et al. Robo1 promotes angiogenesis in hepatocellular carcinoma through the Rho family of guanosine triphosphatases’ signaling pathway. Tumour Biol. 2015;36(11):8413–8424. doi:10.1007/s13277-015-3601-1
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.