Back to Journals » OncoTargets and Therapy » Volume 13
Long Non-Coding RNA NEAT1 Serves as Sponge for miR-365a-3p to Promote Gastric Cancer Progression via Regulating ABCC4
Authors Gao M, Liu L, Zhang D, Yang Y, Chang Z ![]()
Received 10 January 2020
Accepted for publication 14 April 2020
Published 11 May 2020 Volume 2020:13 Pages 3977—3985
DOI https://doi.org/10.2147/OTT.S245557
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
Ming Gao,1,* Liying Liu,2,* Dianbao Zhang,3 Yudan Yang,1 Zhiwei Chang1
1Department of Oncology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou 450052, People’s Republic of China; 2Department of Medical Record, The First People’s Hospital of Zhengzhou, Zhengzhou 450004, People’s Republic of China; 3Department of Oncology, The First Affiliated Hospital of Henan University of Science and Technology, Luoyang 471003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ming Gao
Department of Oncology, The First Affiliated Hospital of Zhengzhou University, No. 1 East Jianshe Road, Zhengzhou 450052, People’s Republic of China
Email [email protected]
Introduction: Long non-coding RNA (lncRNA) was reported to be a crucial regulator in cancer. In this work, our purpose is to explore the biological roles of nuclear paraspeckle assembly transcript 1 (NEAT1) in gastric cancer (GC).
Methods: Quantitative real-time polymerase chain reaction (qRT-PCR) was performed to detect NEAT1 expression in GC cells and normal cells. GC cell behaviors after NEAT1 overexpression or downregulation were analyzed by Cell Counting Kit-8 assay, colony formation assay, wound-healing assay, and flow cytometry assay. Bioinformatic tools were used to analyze the significance of NEAT1 in GC. The involvement of microRNA-365a-3p (miR-365a-3p) and ATP-binding cassette subfamily C member 4 (ABCC4) in the biological roles of NEAT1 in GC progression was validated by luciferase activity reporter assay and rescue experiments.
Results: We found NEAT1 increased expression in both GC tissues and cells and correlated with poorer overall survival of cancer patients. We found NEAT1 overexpression promotes, while its knockdown inhibits GC cell proliferation, colony formation, invasion, and cell cycle progression in vitro. Mechanism analyses showed that NEAT1 serves as a ceRNA to upregulate ABCC4 expression via sponging miR-365a-3p.
Conclusion: In this study, we revealed a NEAT1/miR-365a-3p/ABCC4 triplet in GC progression, which may provide novel targeted therapy markers for GC.
Keywords: long non-coding RNA, NEAT1, miR-365a-3p, ABCC4, gastric cancer
Introduction
More than half of gastric cancer (GC) cases are diagnosed in East Asia.1 Most of GC patients were diagnosed at late stages with malignant metastasis and thus results in poorer overall survival of cancer patients.2 Hence, identifying novel therapeutic targets for GC is needed to improve the survival quality of GC patients.3
The in-depth understanding of RNA biology demonstrated RNA is not only a bridge between DNA and protein but also played crucial roles in multiplies cellular processes.4 Long non-coding RNA (lncRNA) is a type RNA with limited coding capacity and at the length of over 200 nucleotides.5 The importance of lncRNAs in regulating cancer progression has been appreciated in recent years.6
Nuclear paraspeckle assembly transcript 1 (NEAT1) is a newly discovered lncRNA that is an essential component of nuclear paraspeckles.7 Moreover, NEAT1 was reported could function as oncogenic lncRNA in cancers. Shan et al showed NEAT1 was elevated expression in bladder cancer, and its overexpression was found to promote bladder cancer progression in vitro via regulating microRNA-410 and high-mobility group box 1.8 In prostate cancer, NEAT1 was found highly expressed in docetaxel-resistant samples, and its oncogenic role was exerted through sponging miR-34a-5p and miR-204-5p.9 Moreover, NEAT1 was also found elevated expression in GC, and its overexpression was found to promote cancer peogewaaion via upregulating miR-17 and activating PI3K/AKT and GSK3β pathways.10
lncRNA was found could exert its biological roles through competitively binding with miRNA.11 miR-365a-3p was reported to have a tumor-suppressive role in pancreatic cancer but an oncogenic role in laryngeal squamous cell carcinoma.12,13 For instance, miR-365a-3p was found to decrease expression in pancreatic cancer tissues and regulates c-Rel to inhibit cancer progression.12 On the contrary, miR-365a-3p was found elevated expression in laryngeal squamous cell carcinoma, and the knockdown of miR-365a-3p could suppress cell growth, metastasis through regulating PI3K/AKT signaling pathway.13 These results implied the importance of miR-365-3p in regulating cancer progression. However, the connection of NEAT1 and miR-365a-3p was not explored previously.
In this work, we aimed to explore the biological roles of NEAT1 and its associated mechanisms in GC, with the hope to develop novel therapeutic targets for GC treatment.
Materials and Methods
Cell Culture
GC cells MGC-803, HGC-27, and BGC-823, and normal gastric cell GES-1 were purchased from Cell Bank of Chinese Academy of Sciences (Shanghai, China). These cells were incubated in RPMI 1640 in supplement with 10% fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) at a 37°C moist incubator contains 5% CO2.
Cell Transfection
miR-365a-3p mimic and negative control (NC-miR) were purchased from GenePharm (Shanghai, China). NEAT1 sequence was cloned into pcDNA3.1 to generate pNEAT1 by GenScript (Nanjing, Jiangsu, China). Small interfering RNA against NEAT1 (si-NEAT1), ABCC4 (si-ABCC4) and negative control (NC-siR) were purchased from GenePharm. Cell transfection was conducted using Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific). After 48 h of transfection, cells were collected for further analyses.
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
RNA was isolated with Trizol reagent and then reverse-transcribed into first-strand complementary DNA using M-MLV Reverse Transcriptase kit (Invitrogen) according to the provided protocols. RT-qPCR was conducted using SYBR Green (Takara, Dalian, Liaoning, China) at ABI 7500 system (Applied Biosystems, Foster City, CA, USA). Primers used were as follows: miR-365a-3p forward, 5′-TAATGCCCCTAAAAATCCTTAT-3′ and reverse, 5′-CAGTGCGTGTCGTGGAGT-3′; U6 snRNA forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′, NEAT1 forward, 5′-TGGCTAGCTCAGGGCTTCAG-3′ and reverse, 5′- TCTCCTTGCCAAGCTTCCTTC-3′; ABCC4 forward, 5′- GGCAGTGACGCTGTATGG-3′ and reverse, 5′-CGCCAGGTCTGACAGTAAAG-3′; GAPDH forward, 5′-CTGGGCTACACTGAGCACCAG-3′ and reverse, 5′-CCAGCGTCAAAGGTGGAG-3′. Relative gene expression level was calculated by 2−ΔΔCt method with GAPDH or U6 snRNA as internal controls.
Cell Proliferation Assay
Cell Counting Kit-8 (CCK-8, Beyotime, Haimen, Jiangsu) was used to analyze cell proliferation. Cells were seeded in 96-well plate at the density of 3 × 103 and incubated for indicated time points. Then, CCK-8 reagent was added to each well and further incubated for 2 h. Absorbance of each well was measured at 450 nm using a microplate reader.
Cell Cycle Analysis
Cells were collected and then digested by trypsin and fixed by 70% cold ethanol. The treated cells were stained by PI (Beyotime) in the supplement of RNase A for 30 min. Cell cycle distribution was analyzed at FACSCalibur system (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) equipped with ModFit software (Verity Software House, Topsham, ME, USA).
Colony Formation Assay
A 5 × 103 cells were seeded in 6-well plates and cultured for 2 weeks at the above-described conditions. Then, cells were fixed by methanol and dyed by crystal violet. Finally, colony numbers were counted using a microscope.
Transwell Invasion Assay
Transwell chamber with 8 μm pore size was used to detect cell invasion ability. A 5 × 104 cells in serum-free medium were seeded in the upper chamber. Moreover, medium in supplement with FBS was filled into the lower chamber. After 48 h incubation, non-invasive cells were removed, while invaded cells were fixed by paraformaldehyde and stained with crystal violet. Invasive cell numbers were counted under a microscope.
Dual-Luciferase Reporter Assay
Bioinformatic analysis tools were utilized to predict the miRNA target of NEAT1 and we found miR-365a-3p was a putative target. In addition, the target of miR-365a-3p was predicted by bioinformatic tool and showed ABCC4 was a possible target. The wild-type sequence of NEAT1 and ABCC4 was cloned into pmirGLO (Promega, Madison, WI, USA) and designated as NEAT1-wt or ABCC4-wt. In addition, the sequences with mutations in binding regions were generated using site-direct mutagenesis kit (Takara) and named as NEAT1-mt or ABCC4-mt. Cells were co-transfected with luciferase vectors and miRNAs using Lipofectamine 2000. Relative luciferase activity was measured using Dual-Luciferase Reporter assay system (Promega) after 48 h transfection using Renilla luciferase activity as an internal control.
Detection of NEAT1, miR-365a-3p, and ABCC4 Expression in GC Tissues and Normal Tissues
ENCORI website (http://starbase.sysu.edu.cn/) was used to detect NEAT1, miR-365a-3p, and ABCC4 expression levels in GC tissues and normal tissues.
Detection of the Significance of NEAT1 and ABCC4 Expression in GC
Kaplan–Meier curve website (http://kmplot.com/analysis/index.php?p=service&start=1) was used to detect the clinical significance of NEAT1 and ABCC4 on GC patients.
Statistical Analysis
Data collected from three independent experiments were analyzed at GraphPad Prism 6.0 and presented as mean ± standard deviation. Two-tailed Student’s t-test and one-way analysis of variance were used to measure significant differences in groups. P < 0.05 was believed to indicate statistically significant.
Results
NEAT1 Was Upregulated Expression in GC and Correlated with Poor Overall Survival
RT-qPCR analysis showed NEAT1 expression in GC cells was significantly higher than that in normal cells (Figure 1A). Moreover, we found that NEAT1 also upregulated expression in GC tissues compared with normal tissues using ENCORI (Figure 1B). Importantly, we showed high NEAT1 expression was an indicator of poorer overall survival of GC patients (Figure 1C). These results implied the importance of NEAT1 in GC progression.
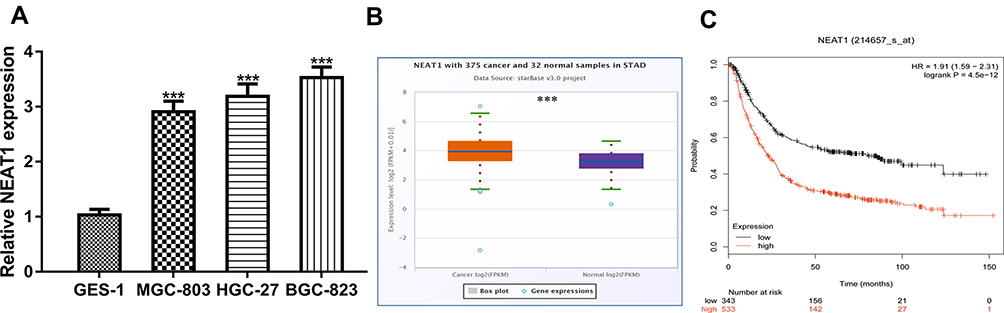 |
Figure 1 NEAT1 was upregulated in GC and correlated with poorer overall survival. Notes: (A) NEAT1 expression in GC cells and normal cells. (B) NEAT1 expression in GC tissues and normal tissues. (C) High NEAT1 expression was a predictor for poorer overall survival of cancer patients. ***P<0.001. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer. |
Knockdown and Overexpress Efficiency of NEAT1 in GC Cells
To explore the biological roles of NEAT1 in GC progression, we established NEAT1 knockdown and overexpression cells. RT-qPCR showed the introduction of si-NEAT1 significantly decreased NEAT1 expression in GC cells compared with NC-siR (Figure 2A). Moreover, the introduction of pNEAT1 increased the levels of NEAT1 in GC cells (Figure 2B).
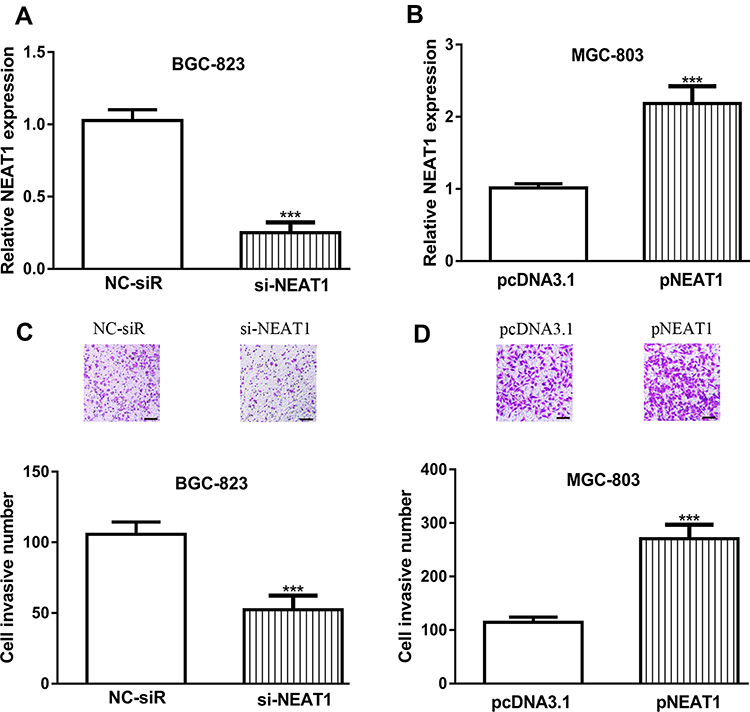 |
Figure 2 Influences of NEAT1 expression on GC cell invasion. Notes: (A) NEAT1 expression in GC cells with si-NEAT1 and NC-siR transfection. (B) NEAT1 expression in GC cells with pNEAT1 and pcNDA3.1 transfection. (C) Cell invasion ability in GC cells with si-NEAT1 and NC-siR transfection. (D) Cell invasion ability in GC cells with pNEAT1 and pcNDA3.1 transfection. ***P<0.001. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer; si-NEAT1, small interfering RNA against NEAT1; NC-siR, negative control small interfering RNA. |
Influence of NEAT1 on GC Cell Invasion
Transwell invasion assay was performed to detect the effects of NEAT1 on GC cell invasion. As shown in Figure 2C and D, we found NEAT1 knockdown could inhibit, while NEAT1 overexpress could stimulate GC cell invasion.
Influence of NEAT1 on GC Cell Growth
Then, the effects of NEAT1 on GC cell growth were detected in vitro. CCK-8 assay showed cell proliferation rate was higher in pNEAT1 group than that in pcDNA3.1 group, and lower in si-NEAT1 group than in NC-siR group (Figure 3A and B). Flow cytometry showed the percentage in G0/G1 phase in pNEAT1 group was lower than that in pcDNA3.1 group, while the introduction of si-NEAT1 caused cell cycle arrest (Figure 3C and D). Colony formation assay indicated colony numbers were higher in pNEAT1 group than that in pcDNA3.1 group (Figure 3E). Moreover, compared with the NC-siR group, colony numbers in si-NEAT1 group were significantly decreased (Figure 3F). We also transfect pNEAT1 into GES-1 cell and found NEAT1 overexpression could promote cell proliferation (Supplementary Figure 1A and 1B).
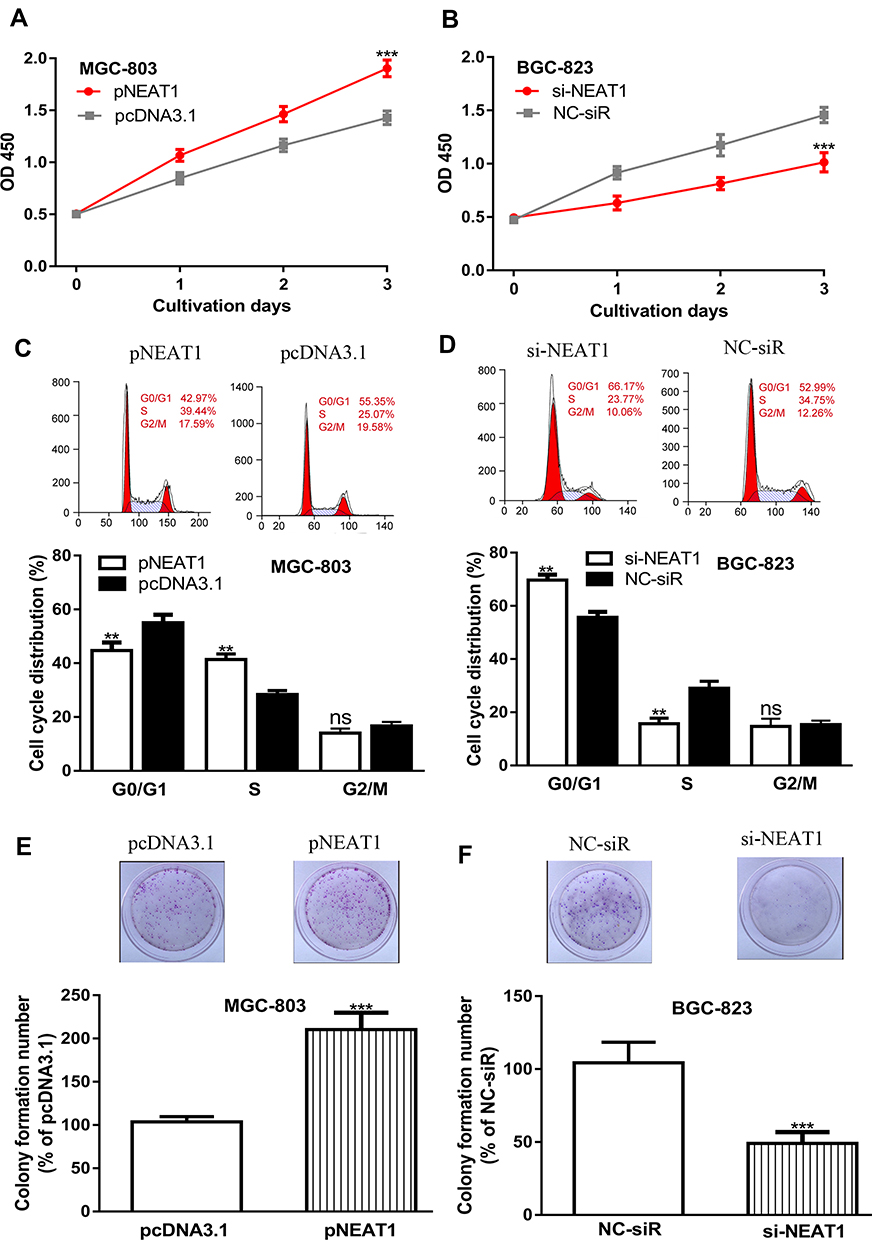 |
Figure 3 Influence of NEAT1 expression on GC cell growth. Notes: (A) Cell proliferation rate in GC cells with pNEAT1 and pcNDA3.1 transfection. (B) Cell proliferation rate in GC cells with si-NEAT1 and NC-siR transfection. (C) Cell cycle distribution in GC cells with pNEAT1 and pcNDA3.1 transfection. (D) Cell cycle distribution in GC cells with si-NEAT1 and NC-siR transfection. (E) Colony formation ability in GC cells with pNEAT1 and pcNDA3.1 transfection. (F) Colony formation ability in GC cells with si-NEAT1 and NC-siR transfection. ***P<0.001; **P<0.01; ns not significant. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer; si-NEAT1, small interfering RNA against NEAT1; NC-siR, negative control small interfering RNA. |
NEAT1 Could Interact with miR-365a-3p in GC
Bioinformatic analysis tool showed miR-365a-3p was a putative target of NEAT1 in GC (Figure 4A). RT-qPCR results indicated that miR-365a-3p expression level was upregulated by si-NEAT1 and downregulated by pNEAT1 in GC cells (Figure 4B and C). The results of luciferase activity reporter assay in Figure 4D indicated miR-365a-3p mimic decreased luciferase activity in GC cells transfected with NEAT1-wt. In addition, the analysis of miR-365a-3p in GC tissues at ENCORI showed it was decreased expression in GC tissues compared with normal tissues (Figure 4E).
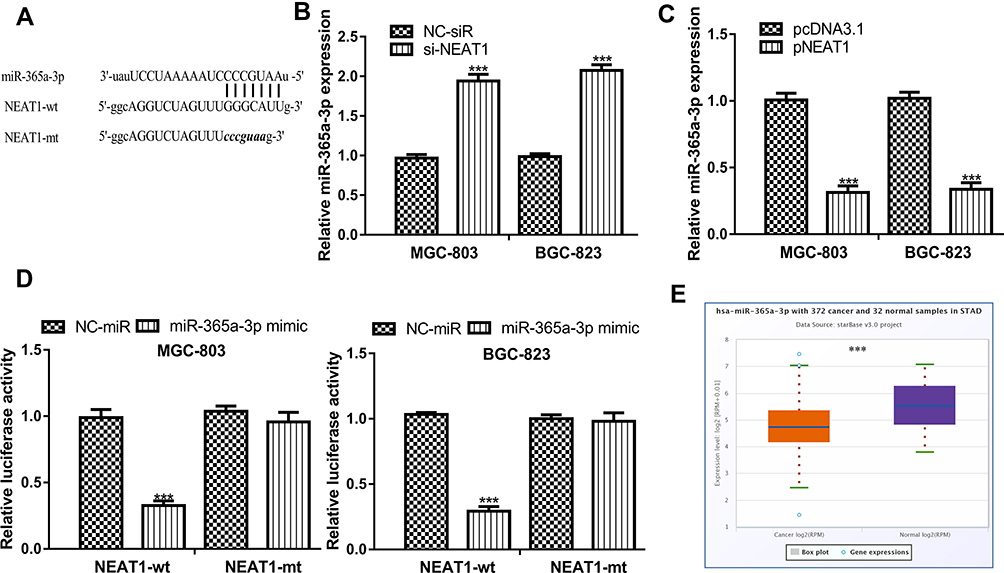 |
Figure 4 NEAT1 interact with miR-365a-3p in GC. Notes: (A) Binding model between NEAT1 and miR-365a-3p. (B) miR-365a-3p expression in GC cells with si-NEAT1 and NC-siR transfection. (C) miR-365a-3p expression in GC cells with pNEAT1 and pcNDA3.1 transfection. (D) Relative luciferase activity in GC cells with luciferase plasmids and miRNAs transfection. (E) miR-365a-3p expression in GC tissues and normal tissues. ***P<0.001. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer; si-NEAT1, small interfering RNA against NEAT1; NC-siR, negative control small interfering RNA; miR-365a-3p, microRNA-365a-3p; wt, wild-type; mt, mutant; NC-mimic, negative control mimic. |
miR-365a-3p Regulates ABCC4 Expression in GC
TargetScan showed miR-365a-3p has a binding site in 3ʹ-UTR of ABCC4 (Figure 5A). Results in Figure 5B revealed miR-365a-3p overexpression could decrease ABCC4 expression. We also found NEAT1 overexpression could increase, while NEAT1 knockdown could decrease ABCC4 expression in GC cells (Figure 5C and D). Luciferase activity assay showed luciferase activity in GC cells harboring ABCC4-wt was decreased by miR-365a-3p mimic (Figure 5E). Importantly, we showed ABCC4 was highly expressed in GC tissues and correlated with poorer overall survival of GC patients (Figure 5F and G).
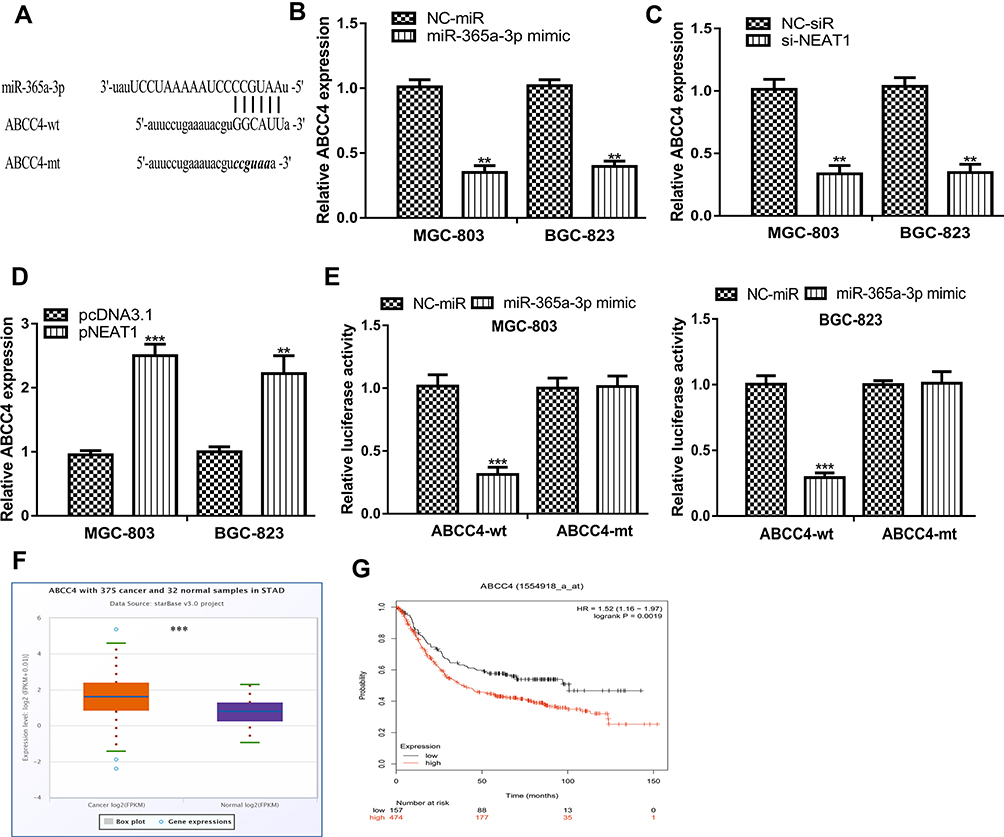 |
Figure 5 miR-365a-3p interact with ABCC4 in GC. Notes: (A) Binding model between ABCC4 and miR-365a-3p. (B) ABCC4 expression in GC cells with miR-365a-3p mimic and NC-mimic transfection. (C) ABCC4 expression in GC cells with pNEAT1 and pcNDA3.1 transfection. (D) ABCC4 expression in GC cells with si-NEAT1 and NC-siR transfection. (E) Relative luciferase activity in GC cells with luciferase plasmids and miRNAs transfection. (F) ABCC4 expression in GC tissues and normal tissues. (G) High ABCC4 expression was a predictor for poorer overall survival of cancer patients. ***P<0.001; **P<0.01. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer; si-NEAT1, small interfering RNA against NEAT1; NC-siR, negative control small interfering RNA; miR-365a-3p, microRNA-365a-3p; wt, wild-type; mt, mutant; NC-mimic, negative control mimic; ABCC4, ATP-binding cassette subfamily C member 4. |
NEAT1 Regulates GC Cell Behaviors via miR-365a-3p/ABCC4 Axis
At length, we explored whether NEAT1 regulates GC cell behaviors via miR-365a-3p/ABCC4 axis using rescue experiments. Results showed miR-365a-3p mimic or si-ABCC4 transfection decreased GC cell growth and invasion (Figure 6A–D). Rescue experiments showed miR-365a-3p mimic and si-ABCC4 could partially reverse the stimulation effects of pNEAT1 on GC cell growth and invasion to a different extent (Figure 6A–D).
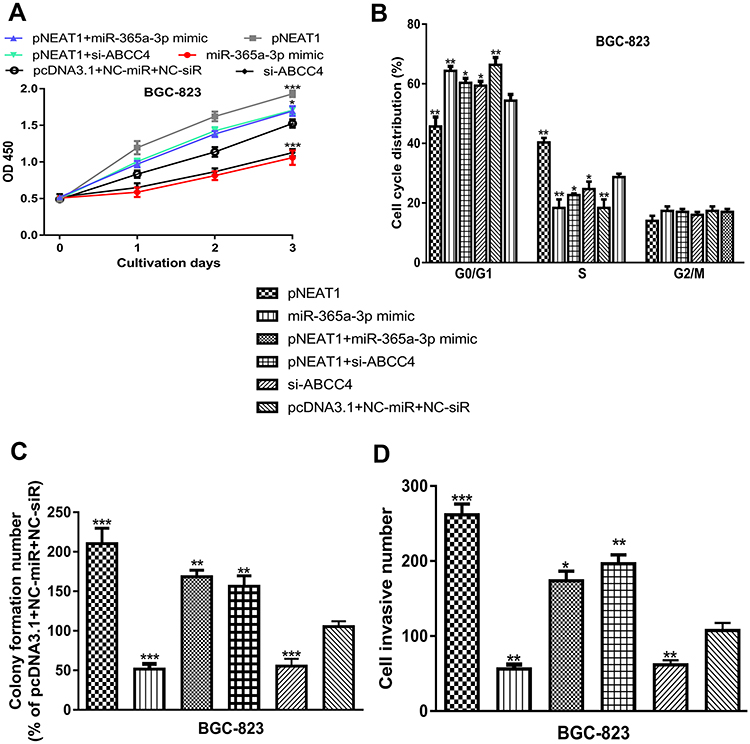 |
Figure 6 NEAT1 regulates GC cell behaviors via targeting miR-365a-3p and ABCC4 axis. Notes: (A) Effects of NEAT1 overexpression, miR-365a-3p overexpression, and ABCC4 knockdown on GC cell proliferation rate. (B) Effects of NEAT1 overexpression, miR-365a-3p overexpression, and ABCC4 knockdown on GC cell cycle progression. (C) Effects of NEAT1 overexpression, miR-365a-3p overexpression, and ABCC4 knockdown on GC cell colony formation. (D) Effects of NEAT1 overexpression, miR-365a-3p overexpression, and ABCC4 knockdown on GC cell invasion. ***P<0.001; **P<0.01; *P<0.05. Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; GC, gastric cancer; miR-365a-3p, microRNA-365a-3p; ABCC4, ATP-binding cassette subfamily C member 4; NC-mimic, negative control mimic; si-ABCC4, small interfering RNA against ABCC4; NC-siR, negative control small interfering RNA. |
Discussion
It has been widely recognized that lncRNAs play crucial roles in regulating carcinogenesis including GC.14,15 For instance, LINC00682 was found to decrease expression in GC cells, and its overexpression could upregulate LIM homeobox transcription factor 1α expression via serving as a sponge for miR-9 to thus result in the inhibition on GC cell growth, migration, and invasion.14 Another work indicated small nucleolar RNA host gene 7 (SNHG7) was highly expressed in GC and correlated with advanced tumor stages, poorer lymph-node metastasis, and overall survival.15 Moreover, SNHG7 was found could bind with miR-34a to stimulate GC migration and invasion via regulating Snail-EMT axis.15 However, more studies are needed to explore the biological roles of lncRNAs in GC.
Previous studies indicated NEAT1 functions as an oncogenic lncRNA in cancers.7–9 In line with these studies, we showed NEAT1 was upregulated expression in both GC tissues and cells. In addition, we showed high NEAT1 level was an indicator of poorer overall survival of GC patients. In addition, gain and loss-of-function experiments indicated NEAT1 overexpression could promote GC cell proliferation, colony formation, cell cycle progression, and invasion. In addition, the knockdown of NEAT1 caused opposite effects on GC cell behaviors. The results showed NEAT1 functions as an oncogenic lncRNA in GC. There are some studies to indicate the roles of NEAT1 in regulating cancer viability and migration.10,16 Here, we also showed NEAT1 could regulate GC cell growth by regulating cell cycle distribution.
Increasing evidence suggested lncRNA exerts its function mainly through functioning ceRNA for miRNA.17 Hence, we also supposed that NEAT1 acts as ceRNA to participate in GC development. Bioinformatic analysis tool and luciferase activity reporter assay showed NEAT1 could bind with miR-365a-3p. miR-365a-3p was previously revealed as a target for lncRNA ZEB1-AS1 and regulates the liver cancer cell proliferation.18 In this work, we found overexpression of miR-365a-3p could suppress GC cell growth and invasion, suggesting the tumor-suppressive role of miR-365a-3p.
Furthermore, we also showed ABCC4 was a possible target of miR-365a-3p. ABCC4 is a member of MRP family of transporters and able to transport organic anionic compounds (both endogenous and xenobiotic) out of the cell.19 Besides that, it was found to have a close association with extracellular signaling pathways and drug resistance.19 Moreover, ABCC4 was widely expressed in human tissues.20 ABCC4 was found highly expressed in non-small cell lung cancer cells and its knockdown could suppress cancer cell growth.21 Here, we also showed ABCC4 expression was elevated in GC and correlated with poorer overall survival of cancer patients.
Consider NEAT1 and ABCC4 can interact with miR-365a-3p, and hence, we speculated that NEAT1 may serve as sponge for miR-365a-3p to regulate ABCC4 expression in GC. Rescue experiment showed that overexpression of miR-365a-3p or knockdown of ABCC4 could partially reverse the effects of NEAT1 overexpression on GC cell events. However, we have to admit there is still a limitation in this work, which is we explored the expression of NEAT1, miR-365a-3p, and ABCC4 in GC tissues and normal tissues using the data from TCGA but we did not explore their expression levels in collected human tissues. Hence, in the future, we will recurit patients diagnosed with GC into our study to further investigate the roles of NEAT1 in GC.
Conclusion
In summary, we found NEAT1 was significantly upregulated in GC and correlated with poorer survival. Functional analyses indicated NEAT1 overexpression stimulates, and NEAT1 knockdown inhibits GC cell behaviors. Our study provided novel insights into the mechanisms underlying GC development.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Russo AE, Strong VE. Gastric cancer etiology and management in asia and the west. Annu Rev Med. 2019;70(1):353–367. doi:10.1146/annurev-med-081117-043436
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
3. Shah MA. Gastrointestinal cancer: targeted therapies in gastric cancer-the dawn of a new era. Nat Rev Clin Oncol. 2014;11(1):10–11. doi:10.1038/nrclinonc.2013.231
4. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
5. Zeng J, Gupta VK, Jiang Y, Yang B, Gong L, Zhu H. Cross‐kingdom small RNAs among animals, plants and microbes. Cells. 2019;8(4):371. doi:10.3390/cells8040371
6. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
7. Clemson CM, Hutchinson JN, Sara SA, et al. An architectural role for a nuclear noncoding RNA: NEAT1 RNA is essential for the structure of paraspeckles. Mol Cell. 2009;33(6):717–726. doi:10.1016/j.molcel.2009.01.026
8. Shan G, Tang T, Xia Y, Qian HJ. Long non-coding RNA NEAT1 promotes bladder progression through regulating miR-410 mediated HMGB1. Biomed Pharmacother. 2020;121:109248. doi:10.1016/j.biopha.2019.109248
9. Jiang X, Guo S, Zhang Y, et al. LncRNA NEAT1 promotes docetaxel resistance in prostate cancer by regulating ACSL4 via sponging miR-34a-5p and miR-204-5p. Cell Signal. 2020;65:109422. doi:10.1016/j.cellsig.2019.109422
10. Wang CL, Wang D, Yan BZ, Fu JW, Qin L. Long non-coding RNA NEAT1 promotes viability and migration of gastric cancer cell lines through up-regulation of microRNA-17. Eur Rev Med Pharmacol Sci. 2018;22(13):4128–4137. doi:10.26355/eurrev_201807_15405
11. Zhang G, Li S, Lu J, et al. LncRNA MT1JP functions as a ceRNA in regulating FBXW7 through competitively binding to miR-92a-3p in gastric cancer. Mol Cancer. 2018;17(1):87. doi:10.1186/s12943-018-0829-6
12. Yin L, Xiao X, Georgikou C, et al. MicroRNA-365a-3p inhibits c-Rel-mediated NF-κB signaling and the progression of pancreatic cancer. Cancer Lett. 2019;452:203–212. doi:10.1016/j.canlet.2019.03.025
13. Geng J, Liu Y, Jin Y, et al. MicroRNA-365a-3p promotes tumor growth and metastasis in laryngeal squamous cell carcinoma. Oncol Rep. 2016;35(4):2017–2026. doi:10.3892/or.2016.4617
14. Zhang X, Li J, Li F, Zhao Z, Feng L. LINC00682 inhibits gastric cancer cell progression via targeting microRNA-9-LMX1A signaling axis. Aging. 2019;11(23):11358–11368. doi:10.18632/aging.102533
15. Zhang Y, Yuan Y, Zhang Y, Cheng L, Zhou X, Chen K. SNHG7 accelerates cell migration and invasion through regulating miR-34a-Snail-EMT axis in gastric cancer. Cell Cycle. 2020;19(1):142–152. doi:10.1080/15384101.2019.1699753
16. Tan HY, Wang C, Liu G, Zhou X. Long noncoding RNA NEAT1-modulated miR-506 regulates gastric cancer development through targeting STAT3. J Cell Biochem. 2019;120(4):4827–4836. doi:10.1002/jcb.26691
17. Hu Y, Tian H, Xu J, Fang JY. Roles of competing endogenous RNAs in gastric cancer. Brief Funct Genomics. 2016;15(3):266–273. doi:10.1093/bfgp/elv036
18. Li M, Guan H, Liu Y, Gan X. LncRNA ZEB1-AS1 reduces liver cancer cell proliferation by targeting miR-365a-3p. Exp Ther Med. 2019;17(5):3539–3547. doi:10.3892/etm.2019.7358
19. Hardy D, Bill RM, Jawhari A, Rothnie AJ. Functional expression of multidrug resistance protein 4 MRP4/ABCC4. SLAS Discov. 2019;24(10):1000–1008. doi:10.1177/2472555219867070
20. Borst P, de Wolf C, van de Wetering K. Multidrug resistance-associated proteins 3, 4, and 5. Pflugers Arch. 2007;453(5):661–673. doi:10.1007/s00424-006-0054-9
21. Zhao X, Guo Y, Yue W, Zhang L, Gu M, Wang Y. ABCC4 is required for cell proliferation and tumorigenesis in non-small cell lung cancer. Onco Targets Ther. 2014;7:343–351. doi:10.2147/OTT.S56029
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.